
Nectoneanthes oxypoda ( Marenzeller, 1879 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.743609 |
|
publication LSID |
lsid:zoobank.org:pub:8868B0C6-E0A5-42BC-B7F6-9FF4F0960D27 |
|
DOI |
https://doi.org/10.5281/zenodo.4742631 |
|
persistent identifier |
https://treatment.plazi.org/id/03A8316D-FFC8-C813-F142-CACEF4DEFC0F |
|
treatment provided by |
Carolina |
|
scientific name |
Nectoneanthes oxypoda ( Marenzeller, 1879 ) |
| status |
|
Nectoneanthes oxypoda ( Marenzeller, 1879) View in CoL
(Japanese name: Ougi-gokai) (Korean name: Gyeong-nam-neob-jeok-bal-charm-gaet-ji-reong-i)
( Figures 1 View Figure 1 , 2 View Figure 2 , 3 View Figure 3 A–C, 4A, 5–9)
Nereis (Alitta) oxypoda Marenzeller, 1879: 120–122 View in CoL , pl. 2, fig. 3.
Nereis (Neanthes) oxypoda: Fauvel, 1932: 22 View in CoL ; Treadwell, 1936: 268; Uschakov, 1965: 187–188, fig. 63E.
Nereis oxypoda: Izuka, 1912: 171–173 View in CoL , pl. 18, figs. 8–11 (in part); Mori et al., 1932: 3, pl. 1, fig. 2, pl. 3, figs. 1–6 (in part); Monro, 1934: 362–363 (in part, specimen collected by Chen); 1938: 614–617, figs. 1–5.
Neanthes oxypoda: Hutchings and Murray, 1984: 37–39 View in CoL , fig. 12.
Nectoneanthes oxypoda: Imajima, 1972: 116–117 View in CoL , figs. 36 a–d (in part, epitokes); Imajima, 1996: 145, fig. 116’ (in part, epitokes).
Nereis alatopalpis Wesenberg-Lund, 1949: 281–283 View in CoL , figs. 15–17. syn. nov.
Nectoneanthes alatopalpis: Wu et al., 1985: 168–169 View in CoL , fig. 94; Khlebovich, 1996: 115.
Nectoneanthes donghaiensis He, 1987: 346–349 View in CoL , figs. 1–7. syn. nov.
Nereis ijimai Izuka, 1912: 174–176 View in CoL , pl. 2, fig.1, pl. 19, figs. 1–9; Horst, 1918: 258. syn. nov.
Neanthes ijimai: Imajima and Hartman, 1964: 144–145 View in CoL .
Nectoneanthes ijimai: Imajima, 1972: 117–119 View in CoL ; Imajima, 1996: 143, fig. 115; Wu et al., 1985: 161–163, fig. 90.
Nectoneanthes latipoda Paik, 1973: 81–84 View in CoL , figs. 1a–j, 2; Imajima, 1996: 146–147, figs. 117–117’; Khlebovich, 1996: 116, pl. XXIII; Nishi et al., 1998: 199; Lee et al., 2003: 191; Choi et al., 2005: 387; Yamanishi and Sato, 2007: appendix 20. syn. nov.
Neanthes latipoda: Wilson, 1988: 9 View in CoL .
Nereis (Nereis) legeri Gravier and Dantan, 1934: 57–61 View in CoL , figs. 17–21. syn. nov.
Nectoneanthes multignatha Wu, Sun and Yang, 1981: 163–164 View in CoL , fig. 91. syn. nov.
Neanthes multignatha: Wilson, 1988: 9 View in CoL .
Nereis singularis Wesenberg-Lund, 1949: 278–281 View in CoL , figs. 13–14. syn. nov.
Nectoneanthes singularis: Khlebovich, 1996: 115 View in CoL .
Neanthes succinea: Wilson, 1984: 218–221 View in CoL (in part, 9 specimens from NSW and 5 specimens from WA, Australia); 1988: 5–7 (in part).
Not Nectoneanthes oxypoda View in CoL ; Imajima, 1972: 113–116, figs. 35a–k (in part, atokes, = N. uchiwa View in CoL sp. nov.); Ibáñez, 1972: 24–26, fig. 1 (= Neanthes succinea View in CoL ?); Wu et al., 1985: 164–167, figs. 92–93 (= N. uchiwa View in CoL sp. nov.); Srinivasa Rao and Rama Sarma, 1982: 447, figs. 6–7; Yang and Sun, 1988: 39, figs. 8A–D (= N. uchiwa View in CoL sp. nov.); Imajima, 1996: 144, fig. 116 (= N. uchiwa View in CoL sp. nov.); Khlebovich, 1996: 115–116, pl. XXII (= N. uchiwa View in CoL sp. nov.); Lee et al., 2003: 189, 190 (= N. uchiwa View in CoL sp. nov.); Yamanishi and Sato, 2007: 183 (= N. uchiwa View in CoL sp. nov.).
Type material
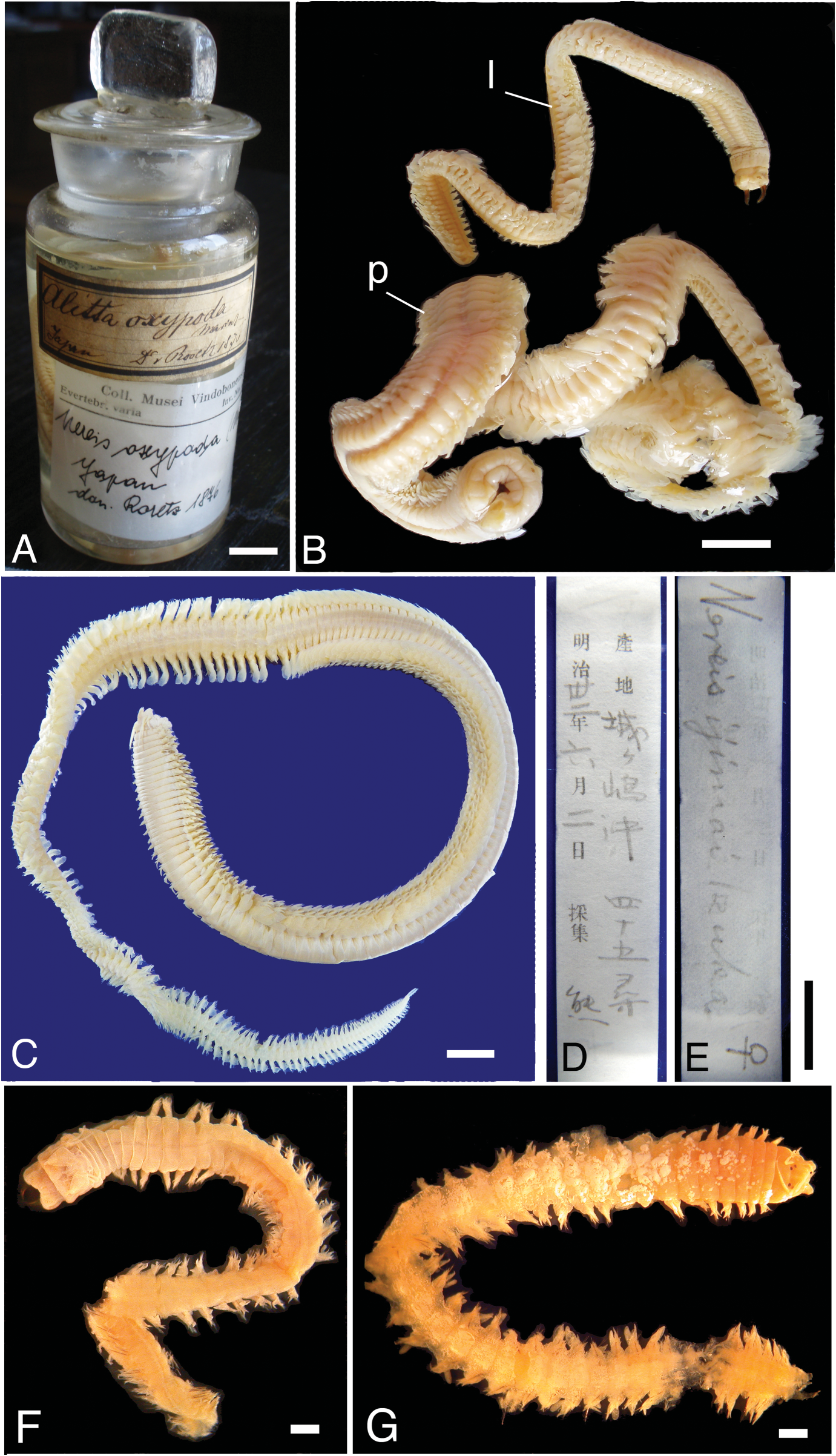
Syntypes of Nereis (Alitta) oxypoda (two atokous individuals), collected from Japan (“wahrscheinlich bei Yokohama”, Marenzeller, 1879: 122) by Dr A. v. Roretz, and donated to NHMW in 1876, stored in a glass jar ( Figure 1A, B View Figure 1 ). The original label attached on the outer surface of the jar read: “ Alitta oxypoda Marenz. Japan D v. Roretz 1876”. Smaller individual of syntypes designated as lectotype ( NHMW ZooEV 20.125); larger individual designated as paralectotype ( NHMW ZooEV 2140) .
Holotype of Nereis ijimai ( UMUTZ Ann-Pc-54, epitokous ovigerous female), collected off Johgashima (35 ◦ 07 ′ N, 139 ◦ 07 ′ E, about 82 m in depth) in Sagami Bay, central Japan on 2 June 1898 by Kuma ( Mr Kumakichi Aoki , a technician of Misaki Marine Biological Station of the University of Tokyo) ( Figure 1 View Figure 1 C–E). In this specimen, right parapodia of chaetigers 5, 30, 80, 99, 125, 177 had been detached from the body. Ventral surface of the anteriormost portion had been dissected to observe paragnaths on retracted proboscis.
Holotype of Nereis alatopalpis (ZMUC POL 1481) collected from St. 72C (25 ◦ 07 ′ N, 57 ◦ 07 ′ E, 12 m deep), Gulf of Oman on 20 April 1937, by G. Thorson ( Figure 1F View Figure 1 ). Several parapodia had been detached from the body.
Holotype of Nereis singularis (ZMUC POL 1488) collected from St. 24A (29 ◦ 07 ′ N, 49 ◦ 07 ′ E, 41 m deep), northern part of Persian Gulf on 13 March 1937, by G. Thorson ( Figure 1G View Figure 1 ). Ventral surface of the anteriormost portion had been dissected to observe paragnaths on retracted proboscis. Several parapodia had been detached from the body.
Other material examined
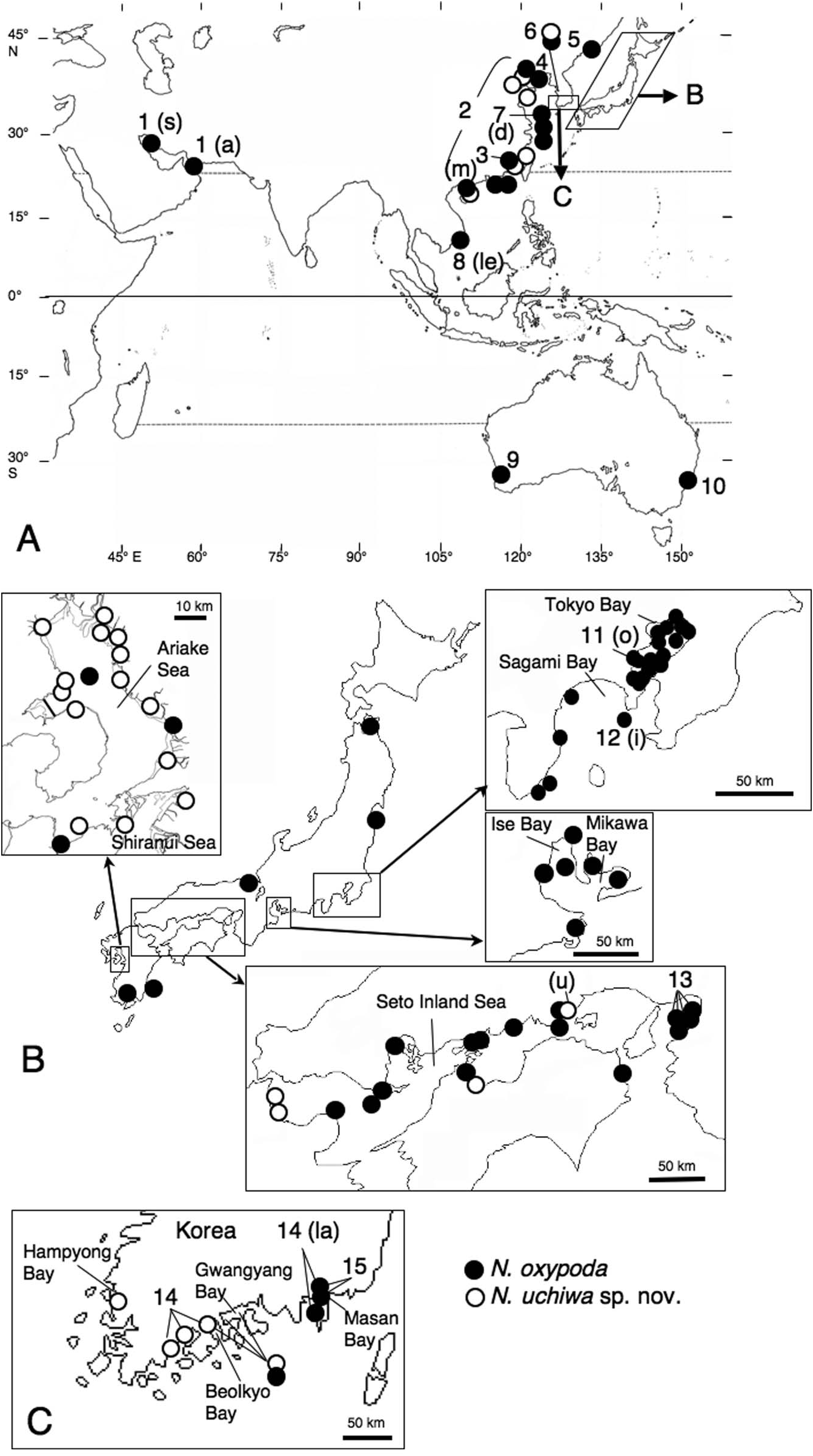
Sexually immature specimens (atokes) collected from Japan. Tohoku Region: Mouth of Tanabu-gawa River (41 ◦ 16 ′ 40 ′′ N, 141 ◦ 10 ′ 30 ′′ E, 7 m deep) in Mutsu Bay , Aomori Prefecture, January 1973, 2 specimens (BW, 0.8–1.5 mm; NSMT Pol 38864–38865) . Intertidal flats at Matsukawa-ura (37 ◦ 49 ′ 17.1 ′′ –11.2 ′′ N, 140 ◦ 59 ′ 04.7 ′′ –53.8 ′′ E), Soma , Fukushima Prefecture, 30 May 2002, coll. T. Suzuki et al., 2 ( OMNH) . Tokyo Bay: Detailed locality and date unknown, coll. Döderlein, 1 (BW, 10.0 mm; NHMW 781 View Materials ) . Subtidal 19 sites (35 ◦ 17.8 ′ –38.0 ′′ N, 139 ◦ 40.0 ′ –140 ◦ 04.0 ′′ E, 7–49 m deep), May and August 1957, March 1958, coll. Fishery Experiment Station of Kanagawa Prefecture, 56 (BW, 0.7–5.2 mm; NSMT Pol 58167–58223) . Off Kisarazu Port (35 ◦ 23 ′ N, 139 ◦ 53 ′ 50 ′′ E), Chiba Prefecture, 17 October 1996, coll. K. Ueda, 3 (BW, 3.2–4.0 mm; NSMT Pol 11395, ZMH P-26034) . Intertidal flats at Banzu (35 ◦ 23 ′ 02 ′′ N, 139 ◦ 54 ′ 11.2 ′′ E), Chiba Prefecture, 31 July 2004, coll. T. Furota et al., 4 (BW, 2.0– 2.7 mm; OMNH) . Eight sites off Banzu , June 1974, 30 (BW, 0.5–2.8 mm; NSMT Pol 32784–32851) . Six sites (8.6–34 m deep) in Nagaura Bay (35 ◦ 17 ′ 48 ′′ –19 ′ 15 ′′ N, 139 ◦ 38 ′ 06 ′′ –39 ′ 49 ′′ E), Yokosuka , Kanagawa Prefecture, 12 and 13 October 1977, coll. M. Imajima, 25 (BW, 0.5–3.0 mm; NSMT Pol 69152–69175) . Intertidal flats at Nojima (35 ◦ 19 ′ 43.6 ′′ –45.8 ′′ N, 139 ◦ 38 ′ 13.9 ′′ –16.4 ′′ E), Kanagawa Prefecture, 13 July 2002, coll. T. Furota et al., 3 ( OMNH) . Sagami Bay: Subtidal bottom, 19 July 1979, 1 (BW, 0.7 mm; NSMT Pol 66301) . Off Morito, Sakawa (35 ◦ 16 ′ N, 139 ◦ 12 ′′ E, 68–80 m deep), Kanagawa Prefecture, 7 May 1966, 10 (BW, 0.5–1.0 mm; NSMT Pol 15023–15065) . Two sites off Shimoda (34 ◦ 39.7 ′ –44.9 ′ N, 138 ◦ 57.0 ′ –139 ◦ 02.1 ′ E, 17–85 m deep), Shizuoka Prefecture, 19 October 1981, coll. M. Imajima, 4 (BW, 0.8–1.5 mm; NSMT Pol 55590–55594) . Mikawa Bay: Subtidal bottom in Atsumi Bay , April 1973, 2 (BW, 2.5 mm; NSMT Pol 64970–71) . Mouth of Yahagi-gawa River (34 ◦ 49 ′ N, 136 ◦ 58 ′ E, 4 m deep, silt), Aichi Prefecture, Chita Bay in Mikawa Bay , 7 November 1995, coll. K. Suganuma, 4 (BW, 1.7–4.0 mm; NSMT Pol 11394, MS) . Ise Bay: Detailed locality and date unknown, 4 (BW, 1.0– 3.2 mm; NSMT Pol 64955–64958) . Matoya Bay (34 ◦ 20 ′ – 23 ′ N, 136 ◦ 49 ′ –55 ′ E), Mie Prefecture, March 1970, 1 (BW, 5.0 mm; NSMT Pol 15066) . Intertidal flats at Shitomo-gawa River (34 ◦ 44 ′ 06.7 ′′ N, 136 ◦ 31 ′ 42.5 ′′ E), Mie Prefecture, 26 September 2003, coll. T. Kimura et al., 1 ( OMNH) . Intertidal flats at Fujimaehigata (35 ◦ 04 ′ 54.4 ′′ N, 136 ◦ 50 ′ 05.5 ′′ E), Nagoya , Aichi Prefecture, 14 August 2004, coll. T. Kimura et al., 1 ( OMNH) . Seto Inland Sea: Intertidal flats at Komatsu-kaigan (34 ◦ 05 ′ 43 ′′ N, 134 ◦ 36 ′ 24 ′′ E), Tokushima, Tokushima Prefecture, 14 March 2009, coll. M. Nii, 2 (BW, 5.5 mm; ZMUC-POL-2217, MS) . Off Hachihama (34 ◦ 33 ′ 20 ′′ N, 133 ◦ 56 ′ 20 ′′ E) Kojima Bay , Okayama Prefecture, 17 December 1906, coll. A. Izuka. 1 (BW, 5.2 mm; NSMT Pol 5290) . Intertidal flats in Kasaoka (34 ◦ 27 ′ –29 ′ N, 133 ◦ 31 ′ – 32 ′ E), Okayama Prefecture, 4 March 2009, coll. K. Okada, 3 (BW, 3.0–5.0 mm; MS) . Around Mukaishima Island (34 ◦ 21 ′ –25 ′ N, 133 ◦ 10 ′ –15 ′ E), Onomichi, Hiroshima Prefecture, October 1963, 1 (BW, 1.6 mm; NSMT Pol 16637); October 1966, 1 (BW, 2.7 mm; NSMT Pol 16636) . Shijushima Island (34 ◦ 21 ′ 30 ′′ –43 ′′ N, 133 ◦ 9 ′ 40 ′′ – 50 ′ E), Onomichi , Hiroshima Prefecture, 27 June 1934, 1 (BW, 3.0 mm; NSMT Pol 14993) . Intertidal flats at Jigozen (34 ◦ 20 ′ 10 ′′ N, 132 ◦ 19 ′ 13 ′′ E), Hatsukaichi , Hiroshima Prefecture, 30 March 2009, coll. K. Okada, 1 (BW, 9.0 mm; MS) . Ariake Sea: Two sites (32 ◦ 58.2 ′ –58.8 ′′ N, 130 ◦ 16.3 ′ –18.7 ′ E, 5–12 m deep), September 1958, coll. Seikai National Fisheries Research Institute, 2 (BW, 1.0– 1.5 mm; NSMT Pol 5285–5286) . Intertidal flats at Shioya (32 ◦ 49 ′ 51.0 ′′ N, 130 ◦ 35 ′ 13.6 ′′ E), Kumamoto Prefecture, 8 May 2004, coll. Y. Henmi et al., 2 (BW, 4.0 mm; OMNH) . Intertidal flats at Hondo (32 ◦ 28 ′ 14.5 ′′ N, 130 ◦ 12 ′ 30.7 ′′ E), Amakusa , Kumamoto Prefecture, 27 September 2003, coll. Y. Henmi et al., 1 (BW, 1.5 mm; OMNH) . Japan Sea: Muddy bottom in Maizuru Bay (35 ◦ 27.29 ′ N, 135 ◦ 19.09 ′ E, 11.5 m deep), Kyoto Prefecture, 27 January, 1975, coll. I. Hayashi, 3 (BW, 0.8–1.1 mm; OMNH) .
Atokes collected from Korea. Beolkyo Bay : Sta. 18 (34 ◦ 47 ′ 37 ′′ N, 127 ◦ 28 ′ 45 ′′ E, 5 m deep), February 2009, coll. J.-W. Choi, 1 (BW, 1.0 mm; KIOST) . Gwangyang Bay : Sta. 5 (34 ◦ 54 ′ 55 ′′ N, 127 ◦ 47 ′ 29 ′′ E, 10 m deep), 7 August 2008, coll. J.-W. Choi, 5 (BW, 1.0– 1.5 mm; KIOST) . Sta. 6 (34 ◦ 50 ′ 58 ′′ N, 127 ◦ 39 ′ 53 ′′ E, 10 m deep), 7 August 2008, coll. J.-W. Choi, 1 (BW, 1.4 mm; KIOST) . Masan Bay : Sta. 1 (35 ◦ 11 ′ 59 ′′ N, 128 ◦ 35 ′ 07 ′′ E, 12 m deep), February 2004, coll. J.-W. Choi, 15 (BW, 1.2–4.3 mm; KIOST) ; April 2004, coll. J.-W. Choi, 6 (BW, 3.0– 4.7 mm; KIOST) ; March 2005, coll. J.-W. Choi, 8 (BW, 2.3–3.2 mm; KIOST) . Sta. 5 (35 ◦ 11 ′ 01 ′′ N, 128 ◦ 34 ′ 16 ′′ E, 15 m deep), March 2004, coll. J.-W. Choi, 2 (BW, 3.0– 4.2 mm; KIOST) . Sta. 8 (35 ◦ 08 ′ 59 ′′ N, 128 ◦ 36 ′ 05 ′′ E, 15 m deep), February 2004, coll. J.-W. Choi, 5 (BW, 1.8–3.8 mm; KIOST) ; March 2004, coll. J.-W. Choi, 6 (BW, 1.7–4.0 mm; KIOST) . Sta. 12 (35 ◦ 05 ′ 50 ′′ N, 128 ◦ 36 ′ 56 ′′ E, 15 m deep), March 2004, coll. J.-W. Choi, 2 (BW, 3.5–4.0 mm; KOIST) .
Atokes collected from Australia. Port Hacking , NSW Sta.631, artificial reef, mud, 17 December 1984, coll. NSW Fisheries, 1 (BW, 1.2 mm; AM W-195222). Lake Macquarie, NSW, Zostera beds, January 1978, coll. K. Robinson, 1 (BW, 1.8 mm; AM W-17830). Hawkesbury R., NSW D4-3, 0.5 km coast of Dongar Is., 21 August 1980, coll. A. Jones et al., 1 (BW, 0.7 mm; AM W-196463). Hawkesbury R., NSW D2-2, Eastern end of Brooklyn Boat Channel, 21 August 1980, coll. A. Jones et al., 1 (BW, 1.3 mm; AM W-196461) .
Sexually mature specimens (epitokes) collected during reproductive swarming in Japan. Tokyo Bay: Honmaki (35 ◦ 24 ′ 10 ′′ –46 ′′ N, 139 ◦ 39 ′ –40 ′ E), Naka-ku, Yokohama , Kanagawa Prefecture, April 1999, coll. T. Kudo, 3 females (BW, 6.0–10.0 mm; NSMT Pol 11398) and 7 males (BW, 4.0–6.0 mm; NSMT Pol 11396, 11397; ZMH P- 26035; ZMUC-POL-2219). Sagami Bay : Ito Port (34 ◦ 58 ′ 15 ′′ N, 139 ◦ 06 ′ 22 ′′ E), Izu Peninsula , Shizuoka Prefecture, 16 May 1970, 1 female (BW, 4.0 mm; NSMT Pol 16635). Seto Inland Sea: Uno Port (34 ◦ 29 ′ 24 ′′ N, 133 ◦ 57 ′ 18 ′′ E) , Okayama Prefecture, 26 October 2009 (18:55–21:00), coll. M. Sato et al., 2 females (BW, 3.0 mm, NSMT Pol 11400; BW, 6.0 mm, NSMT Pol 11403) and 3 males (BW, 3.0 mm; NSMT Pol 11401, 11402), 10–11 November 2010 (18:00, 18:17, 1:00), coll. Y. Urata et al., 2 females (BW, 5.0– 5.5 mm; MS) and 1 male (BW, 2.7 mm; MS). Imabari Port (34 ◦ 03 ′ 45 ′′ N, 133 ◦ 01 ′ 15 ′′ E) , Ehime Prefecture, 27 October 2009 (18:20–18:45), coll. M. Sato et al., 2 females (BW, 6.5 mm, NSMT Pol 11404; BW, 5.5 mm, ZMUC- POL-2218). Hime-shima Port (33 ◦ 43 ′ 17 ′′ N, 131 ◦ 38 ′ 50 ′′ E), Hime-shima Island , Oita Prefecture, 28 October 2009 (20:08), coll. M. Sato et al., 1 male (BW, 3.5 mm; NSMT Pol 11405). Yanai Port (33 ◦ 57 ′ 23 ′′ N, 132 ◦ 08 ′ 11 ′′ E) , Yamaguchi Prefecture, 29 October 2009 (17:50–18:30), coll. M. Sato et al., 4 males (BW, 2.5–3.5 mm; NSMT Pol 11406). Iwai-jima Port (33 ◦ 47 ′ 14 ′′ N, 131 ◦ 59 ′ 20 ′′ E), Iwai-jima Island , Kaminoseki , Yamaguchi Prefecture, 8 September, 2010 (19:00), coll. M. Sato, 2 females (BW, 7.0 mm, NSMT Pol 11407; BW, 9.0 mm, MS). Kyushu: Around Miyazaki, Miyazaki Prefecture, 20 April 1970, 1 female (BW, 2.5 mm; NSMT Pol 14994). Hakamagoshi (31 ◦ 35 ′ 34 ′′ N, 130 ◦ 35 ′ 47 ′′ E), Sakura-jima Island , Kagoshima Prefecture, 17 March 2010 (20:00–20:25), coll. Y. Urata, 1 female (BW, 3.7 mm; MS); 5 March 2011 (18:40–19:50), coll. Y. Urata, 19 females (BW, 2.7–5.0 mm; MS) and 7 males (BW, 2.8–4.5 mm; MS); 21 March 2011 (18:55–19:55), coll. Y. Urata, 4 females (BW, 4.0– 4.5 mm; MS) and 2 males (BW, 3.5–4.0 mm; MS) .
Epitokes collected from sediment samples in Japan. Intertidal flats at the mouth of Edogawa-hosuiro River (35 ◦ 40 ′ 56.5 ′′ N, 139 ◦ 56 ′ 31.0 ′′ E), Tokyo Bay , Chiba Prefecture, 21 April 2003, coll. A. Iijima, 1 female (BW, 11.0 mm; OMNH). Intertidal pebble beach at Hakamagoshi (31 ◦ 35 ′ 05 ′′ N, 130 ◦ 35 ′ 32 ′′ E), Sakura-jima Island , Kagoshima Bay, Kagoshima Prefecture, 8 April 2008, coll. S. Kusumure, 1 female (BW, 3.5 mm; NSMT Pol 11399) .
Diagnosis
Paragnaths in group VII–VIII arranged in distal (on everted proboscis) single transverse row of large paragnaths and proximal irregular one or two rows of large and small paragnaths, extending to lateral and often dorsal surface (near group VI) in oral ring of proboscis, not reaching group V. Relatively small number of paragnaths in group II (10–24 on each side), III (0–11) and IV (10–22 on each side) in maxillary ring of proboscis.
Description
Lectotype, paralectotype and non-type atokes. Lectotype, incomplete individual 105 mm BL, 5.0 mm BW, with 104 chaetigers, with proboscis fully everted ( Figure 1B View Figure 1 ). Paralectotype, almost complete ovigerous female without epitokous metamorphosis, 260 mm BL, 9.0 mm BW, with 170 chaetigers, with proboscis partially everted. Non-types (n = 148) 0.7–9.0 mm BW.
Body stout anteriorly, posteriorly gradually tapering toward pygidium. Dorsum convex, venter relatively flat with longitudinal midventral groove. Colour in preserved specimens whitish cream with brownish pigmentation in anterior dorsum.
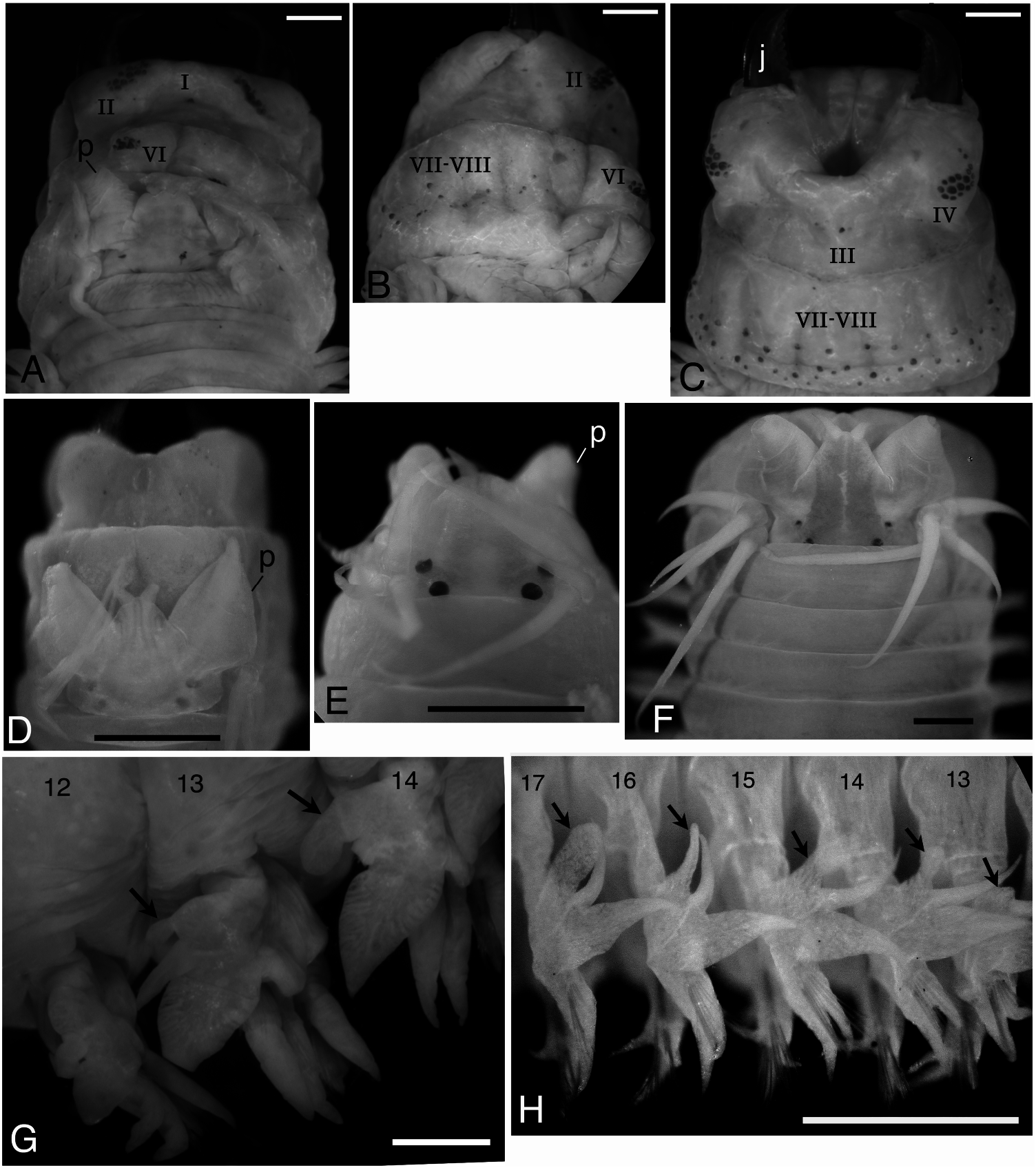
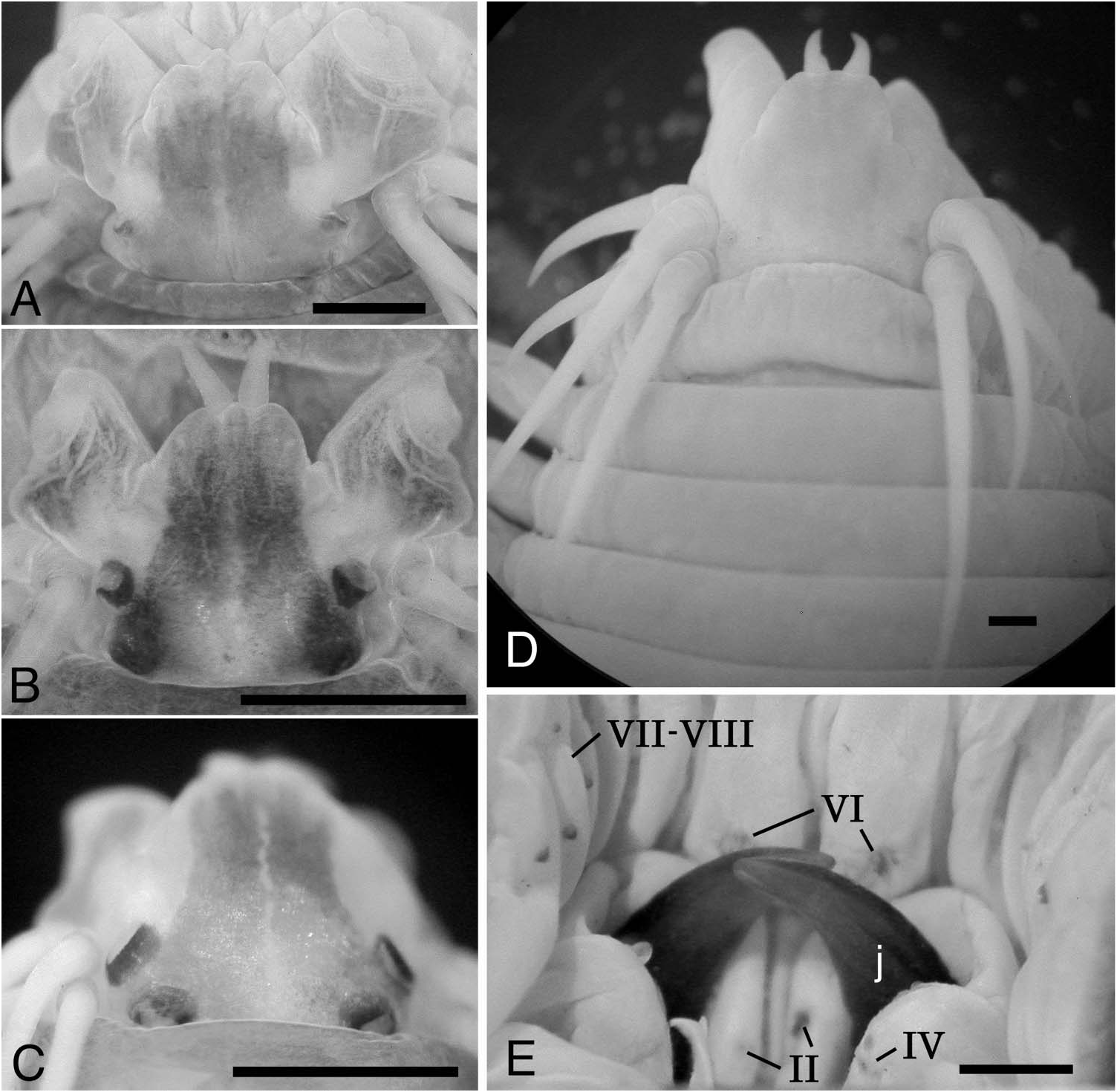
Prostomium with trapezoidal shape, broader than long, with pair of smooth tapered antennae situated at anterior end ( Figure 2A View Figure 2 , D–F). Pair of palps with massive palpophores and short round palpostyles. Two pairs of eyes arranged trapezoidally (anterior pair with space 1.2–1.4 times as wide as that of posterior pair); anterior pair reniform and smaller; posterior pair round and larger (subdermal eyes unclear in some large specimens). Trapezoidal dark pigmentation with mid-longitudinal white slit present on dorsal anterior surface of prostomium; dark pigmentation with white slits often present also on dorsal surface of palpophore ( Figure 2F View Figure 2 ).
Peristomium subequal to (or slightly longer than) subsequent chaetigers, with four pairs of tentacular cirri of unequal length; posterior dorsal tentacular cirri longest, reaching back to chaetigers 2 to 8 (chaetiger 4 in lectotype, chaetiger 3 in paralectotype, usually chaetigers 3 to 5); anterior dorsal tentacular cirri next longest ( Figure 2F View Figure 2 ).
Proboscis with pair of brown jaws, each with 10 or11 teeth in lectotype; teeth indistinct in paralectotype and non-type large individuals. Black paragnaths with usually pointed tip present on both maxillary and oral rings ( Figure 2 View Figure 2 A–D). Paragnath numbers in lectotype (range for all examined specimens in parentheses) as follows, group I: 1 (0–4, n = 42) in longitudinal arrangement, II: 14 on right and 16 on left in three or four arched rows, total 30 (21–44, n = 13), III: 4 (0–9, n = 62) in triangular or rhomboid patch, IV: 15 on right and 17 on left in three or four arched rows, total 32 (21–41, n = 15), V: 1 (0–5, n = 108) in longitudinal arrangement, VI: 11 on right and 15 on left in circular cluster, total 26 (13–28, n = 58), VII–VIII: 63 (59–96, n = 6), arranged in distal (on everted proboscis) single transverse row of large paragnaths and proximal irregular single row (1–2 rows for all examined specimens) of large and small paragnaths, extending to lateral and often dorsal surface (near group VI), not reaching group V ( Figure 2B View Figure 2 ). Some paragnaths obviously lost in paralectotype, resulting in low numbers of paragnaths (e.g. 0 on right and 11 on left in group VI).
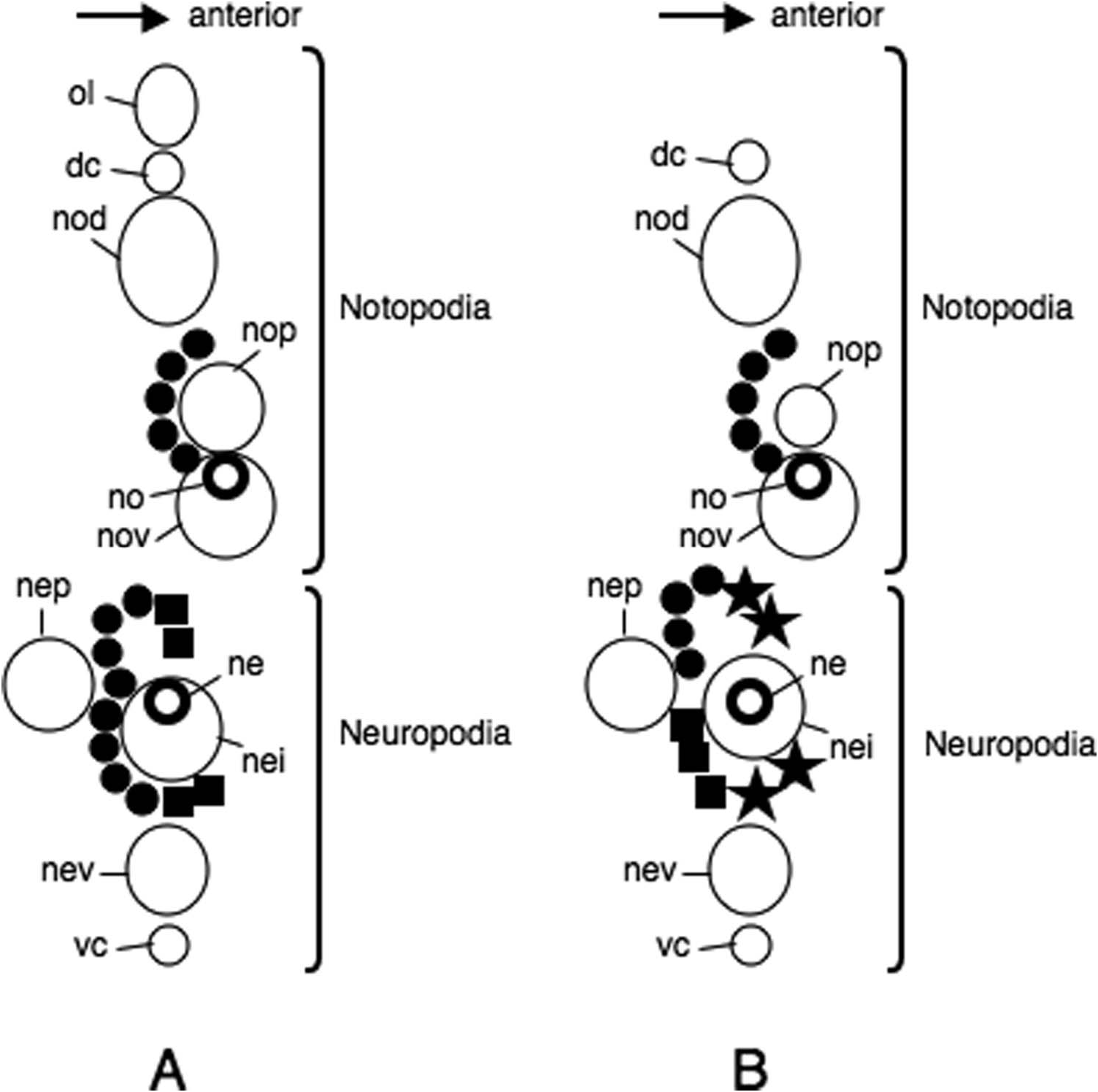
Parapodia of first two chaetigers sub-biramous, all following parapodia biramous. Sub-biramous parapodia with thin notoacicula and reduced notopodia consisting of dorsal cirrus and dorsal ligule, and with ordinary neuroacicula and ordinary neuropodia.
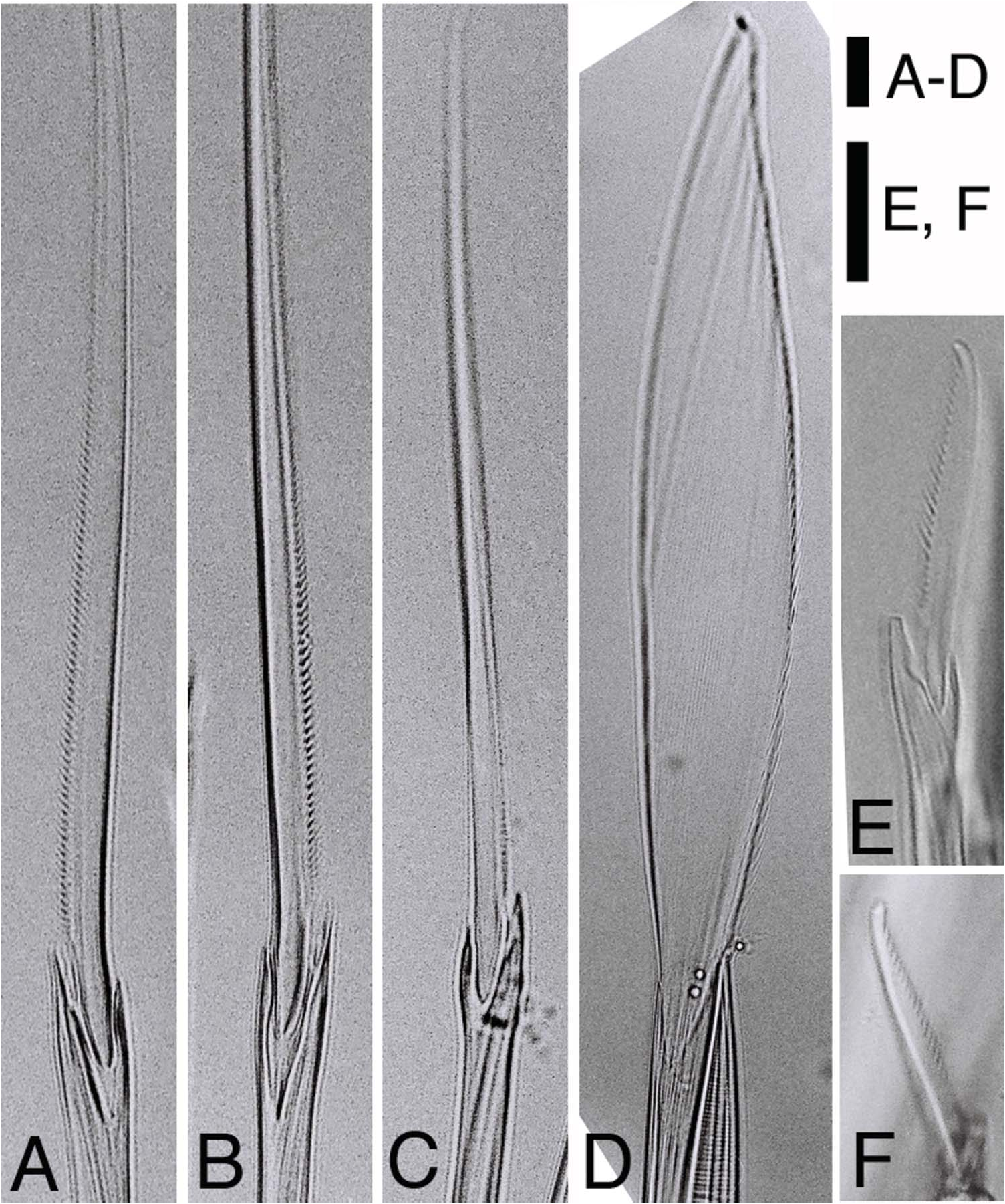
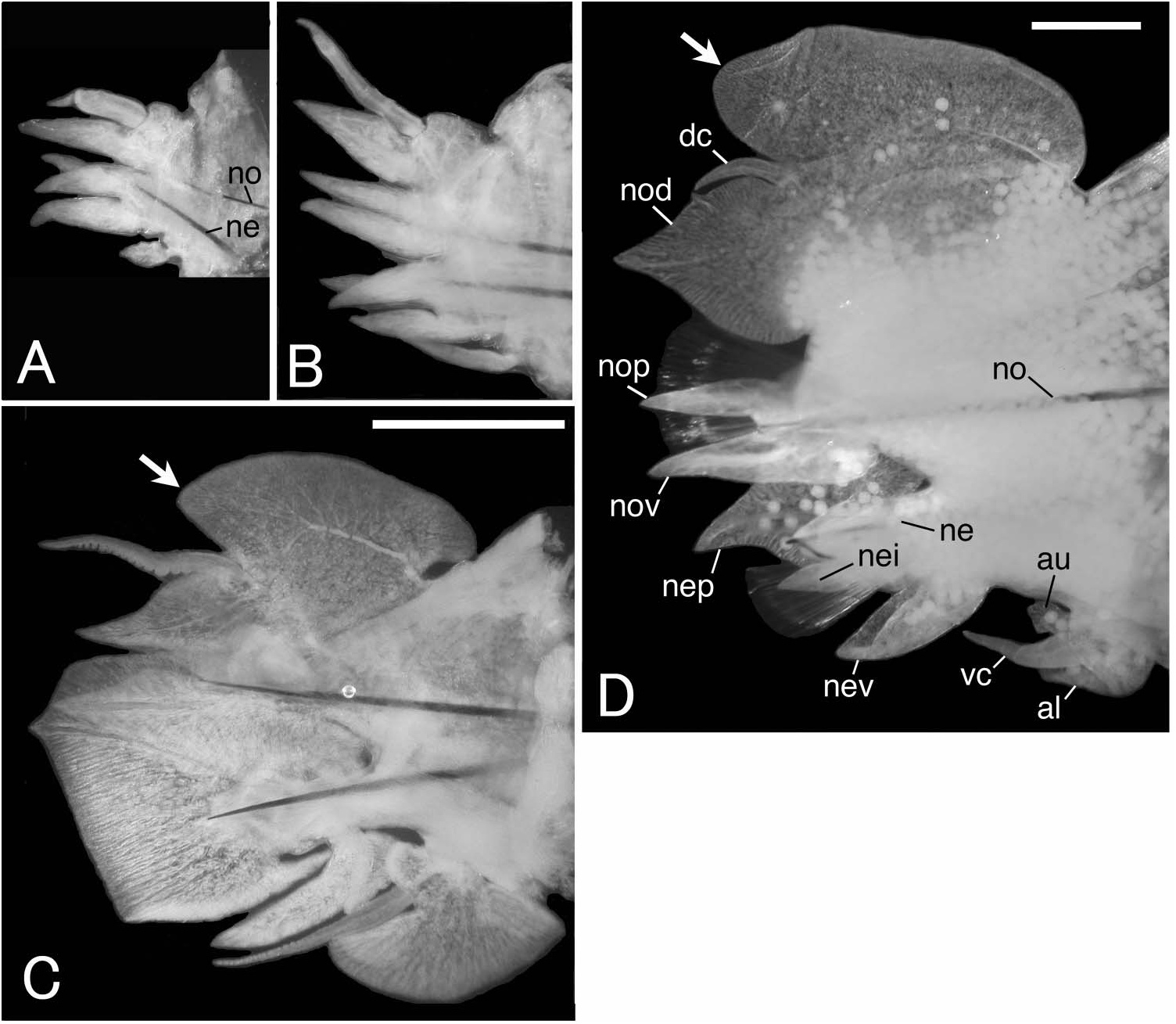
Notopodia consisting of dorsal cirrus, dorsal ligule, prechaetal lobe and ventral ligule in biramous parapodia. Notopodial dorsal ligule slender with tapering tip in anterior parapodia, expanded to large triangular with prominent ovoid lobe developing above dorsal cirrus in middle and posterior parapodia; first ovoid lobe above dorsal cirrus appearing at chaetigers 12–16 (chaetiger 13 in both lectotype and paralectotype) ( Figure 2G, H View Figure 2 ), except at around chaetiger 20 in some juveniles; notopodial dorsal ligule most enlarged in middle parapodia, gradually diminishing in size in posteriormost parapodia ( Figure 3 View Figure 3 A–C). Notopodial prechaetal lobe and notopodial ventral ligule slender with tapering tip throughout; notopodial prechaetal lobe subequal to notopodial ventral ligule in anterior and middle parapodia, diminishing in size in posteriormost parapodia ( Figure 3 View Figure 3 A–C). Dorsal cirri slender, tapering.
Neuropodia consisting of inferior lobe, postchaetal lobe, ventral ligule and ventral cirrus throughout; all lobes and ligule slender with tapering tip throughout, of similar length; ventral cirrus slender with tapering tip ( Figure 3 View Figure 3 A–C).
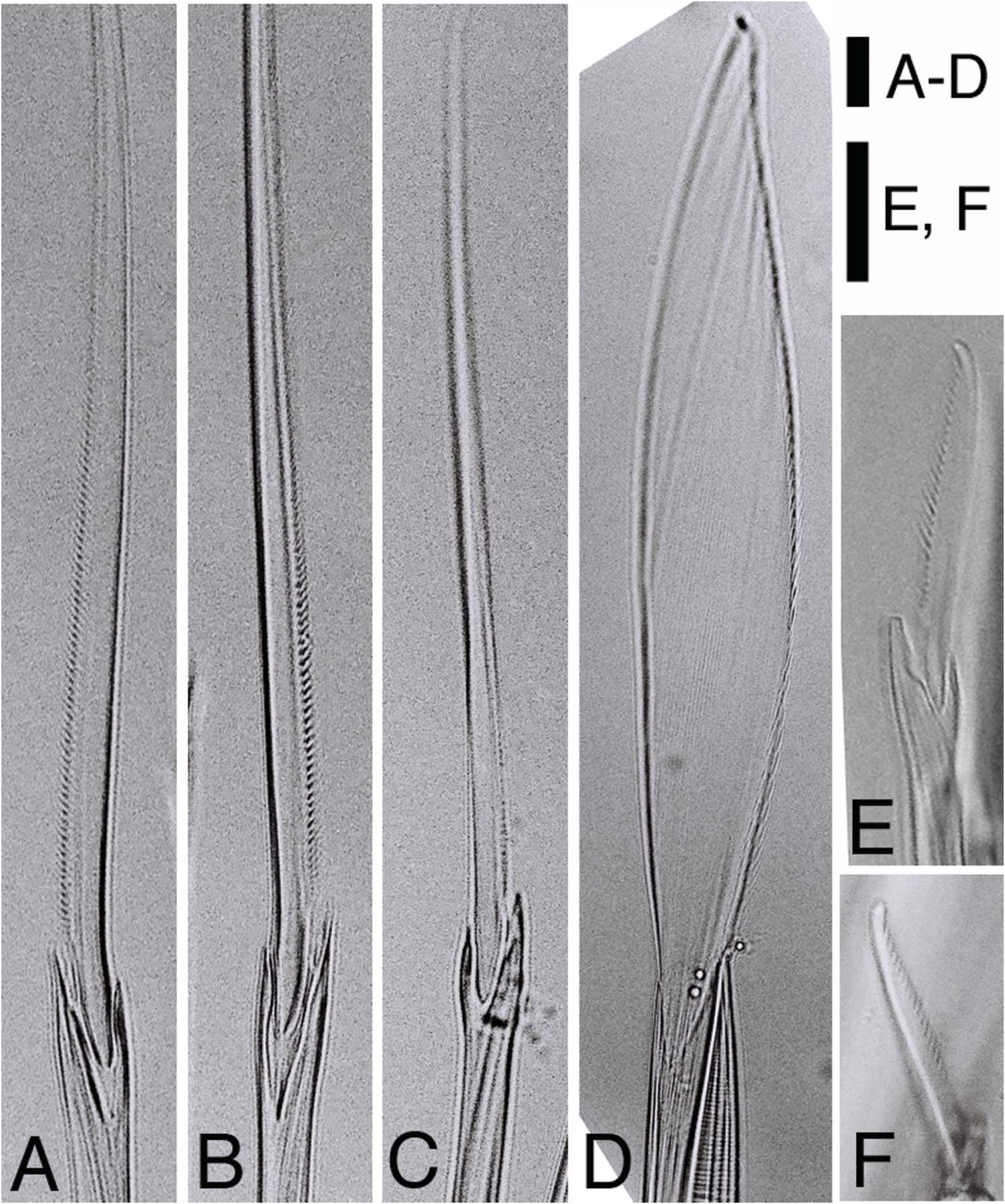
Most chaetae lost or broken in both lectotype and paralectotype. The following description based on only non-type specimens. Only spinigers observed except for juveniles. Notochaetae all homogomph or sesquigomph spinigers; blades long with finely serrated edge ( Figures 4A View Figure 4 , 5A View Figure 5 ). Upper neurochaetae consisting of heterogomph spinigers with short serrated blades ( Figure 5C View Figure 5 ) at superior / anterior position, and homogomph or sesquigomph spinigers with long serrated blades ( Figure 5B View Figure 5 ) at inferior / posterior position ( Figure 4A View Figure 4 ). Lower neurochaetae consisting of homogomph or sesquigomph spinigers with long serrated blades at superior / posterior position, and heterogomph spinigers with short serrated blades at inferior / anterior position. One or two heterogomph falcigers with slender serrated blades present at lowermost portion in lower neurochaetae throughout in small juveniles (less than 2 mm in BW) ( Figure 5E, F View Figure 5 ).
Pygidium with anus on dorsal side, with pair of cylindrical slender anal cirri.
Holotype of Nereis alatopalpis syn. nov. Incomplete atokous specimen composed of anterior fragment (about 20 mm BL, 2.0 mm BW, with 45 chaetigers) and posterior three small fragments (7, 1.5, 4 mm BL, with 15, 5, 11 chaetigers, respectively), with proboscis well everted ( Figures 1F View Figure 1 , 2D View Figure 2 ). Colour in preserved specimens whitish cream.
Prostomium with pair of palps with broad massive palpophores and digitate palpostyles.
Paragnath numbers and arrangement as follows, group I: 1, II: 12 on right and 11 on left, total 23, III: 0, IV: 10 on both sides, total 20 (instead of total 19 in original description by Wesenberg-Lund 1949), V: 0, VI: 11 on right and 9 on left, total 20, VII–VIII: two transverse rows of paragnaths, extending to lateral surface; distal (on everted proboscis) single transverse row of large paragnaths and proximal irregular single row of large and small paragnaths.
Notopodial dorsal ligule slender with tapering tip in anterior parapodia, expanded to triangular with prominent ovoid lobe developing above dorsal cirrus in posterior parapodia; first ovoid lobe above dorsal cirrus appearing at chaetiger 14 (both right and left parapodia of chaetigers 13 had been lost; a “cushion-like” bulge above dorsal cirrus at chaetigers 13 was drawn in original description by Wesenberg-Lund 1949).
Other characteristics well described and drawn in Wesenberg-Lund (1949).
Holotype of Nereis singularis syn. nov. Incomplete atokous specimen of anterior fragment, about 30 mm BL, 2.1 mm BW, with 41 chaetigers, with proboscis retracted ( Figures 1G View Figure 1 , 2E View Figure 2 ). Colour in preserved specimens whitish cream with slightly brownish pigmentation in anterior dorsum.
Prostomium with pair of palps with small palpophores and short round palpostyles.
Paragnath numbers and arrangement as follows, group I: 1, II: 12 on right and 11 on left, total 23, III: 3, IV: 13 on both sides, total 26, V: 0, VI: 9 on right and 9 on left, total 18, VII–VIII: two transverse rows of paragnaths; proximal (on retracted proboscis) single transverse row of large paragnaths and distal irregular single row of large and small paragnaths.
Notopodial dorsal ligule slender with tapering tip in anterior parapodia, expanded to triangular with prominent ovoid lobe developing above dorsal cirrus in posterior parapodia; first ovoid lobe above dorsal cirrus appearing at chaetiger 14.
Other characteristics well described and drawn in Wesenberg-Lund (1949).
Non-type epitokes. Largest complete specimen (female) 250 mm BL, 9.0 mm BW, with 192 chaetigers. Smallest complete specimen (male) 33 mm BL, 2.5 mm BW, with 92 chaetigers. Body widths of females (range: 2.7–11.0 mm, average ± SD: 4.9 ± 1.9 mm, n = 38) significantly larger than those of males (2.5–6 mm, 3.6 ± 0.8 mm, n = 25) (Wilcoxon–Mann–Whitney test, P = 0.0002).
Eyes slightly enlarged in both males and females ( Figure 6 View Figure 6 A–C); border of subdermal eyes unclear in large females. Other characteristics of prostomium and peristomium similar to those of atokes.
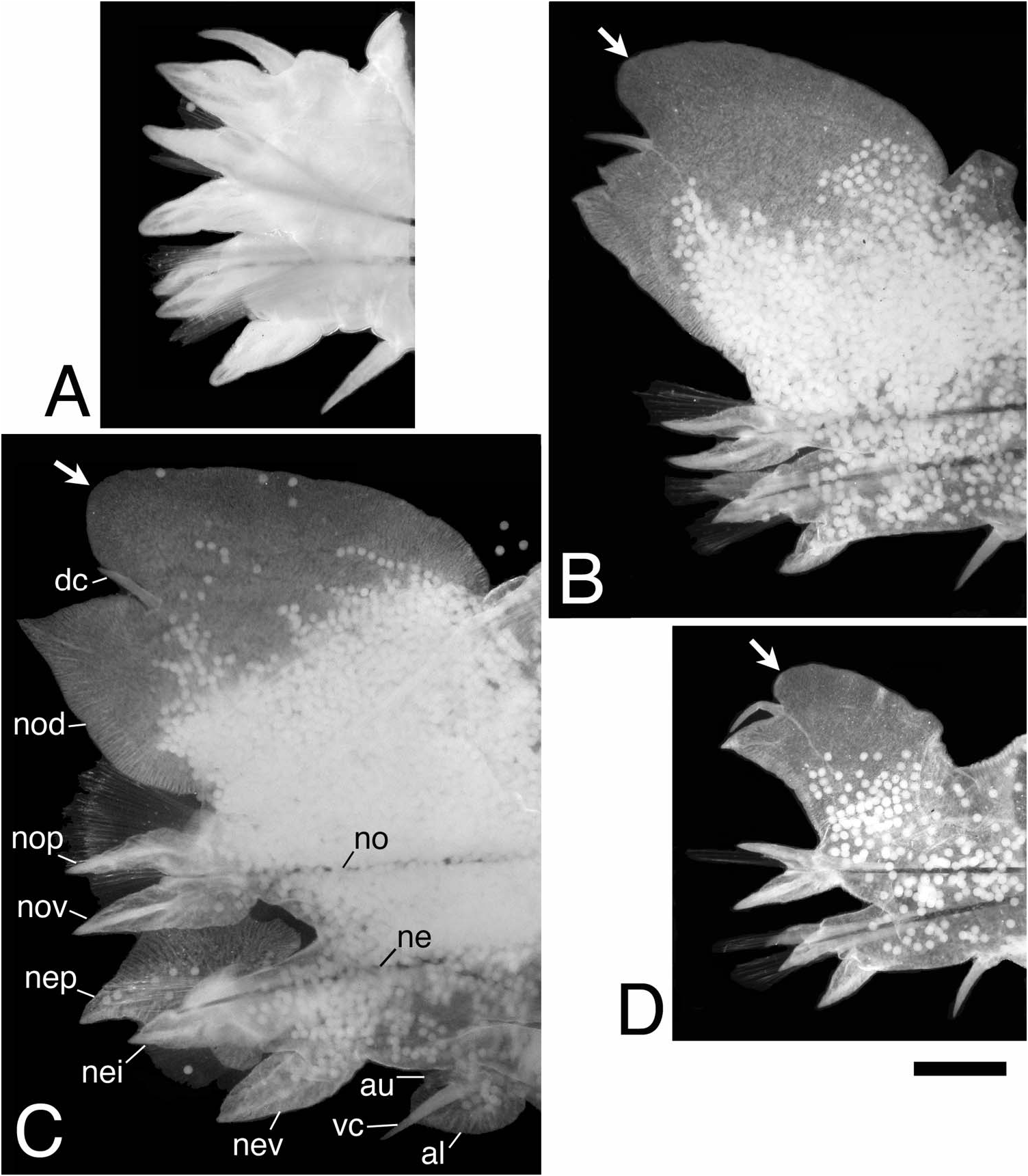
Parapodia and chaetae in anterior body (first 16–19 chaetigers in males, 27–30 chaetigers in females) similar to those of atokes, with no marked morphological change in females, with slight changes in males; dorsal cirri of first seven chaetigers and ventral cirri of first five chaetigers thickened in males ( Figure 7A, B View Figure 7 ).
Parapodia and chaetae in middle body modified ( Figure 7C, D View Figure 7 ). Additional lobes present at upper and lower portions on base of ventral cirri; upper lobe smaller, lower lobe larger, appearing from chaetigers 17–20 to around chaetiger 80 in males, from chaetigers 28–31 to chaetigers 50–100 in females, developing much larger in males than in females. Neuropodial postchaetal lobe developing into large square-shaped flat lamella with small triangular protrusion on lateral edge, and with digitate protrusion on inner side of lower edge in males ( Figure 7C View Figure 7 ); neuropodial postchaetal lobe expanded to triangular lamella with tapering tip in females ( Figure 7D View Figure 7 ). Notopodial ventral ligule expanded remarkably to triangular lamella in males, enlarged slightly in females. Dorsal cirri with serration on lower edge, slightly elongated in males; dorsal cirri with neither serration nor elongation in females. Ventral cirri with minute serration on upper edge, slightly elongated in males; ventral cirri with neither serration nor elongation in females. Atokous chaetae substituted by epitokous natatory ones ( Figure 5D View Figure 5 ) completely in males, and incompletely (several atokous spinigers remaining) in females; blade of epitokous natatory chaetae flat and wide, with minutely serrated edge on one side, and tapering tip.
Parapodia and chaetae in posterior body (behind around chaetiger 80 in males, behind chaetigers 50–100 in females) similar to those of atokes.
Coelom of females filled with eggs (full-grown oocytes); eggs spherical, 125–135 µm in diameter in fixed specimen collected from Iwai-jima Island , Yamaguchi Prefecture , 130–135 µm in diameter in fixed specimen collected from Uno Port, Okayama Prefecture, 130–140 µm in diameter in fixed specimen collected from Sakura-jima Island , Kagoshima Prefecture .
Holotype of Nereis ijimai syn. nov. Complete ovigerous female with epitokous metamorphosis, about 370 mm BL, 9.0 mm BW, with 212 chaetigers, with proboscis retracted ( Figure 1 View Figure 1 C–E).
Border of subdermal eyes unclear ( Figure 6D View Figure 6 ). Pigmentation on dorsal surface of prostomium and palpophore unclear probably because of discoloration from longterm preservation.
Peristomium with four pairs of tentacular cirri; posterior dorsal tentacular cirri longest, reaching back to chaetiger 4.
Proboscis with pair of dark brown jaws; teeth of jaws indistinct ( Figure 6E View Figure 6 ). Black paragnaths with round tip; some paragnaths obviously lost, leaving traces of their bases. Paragnath numbers and arrangement as follows, group I: 0, II: 4 on right and 8 on left, total 12, III: 0, IV: 0 on right and 3 on left, total 3, V: 0, VI: 12 on right and 10 on left, total 22, VII–VIII: proximal (on retracted proboscis) single transverse row of large paragnaths and distal irregular single row of small paragnaths, extending to lateral surface ( Figure 6E View Figure 6 ).
Parapodia and chaetae in anterior body (up to first 30 chaetigers, Figure 8A, B View Figure 8 ) similar to those of atokes described above; ovoid lobe above dorsal cirrus in notopodia first appearing at chaetiger 14.
Parapodia and chaetae in middle body modified ( Figure 8C View Figure 8 ), similar to those of non-type epitokes described above. Additional lobes present at upper and lower portions on base of ventral cirri, appearing from chaetiger 31 to chaetiger 104. Neuropodial postchaetal lobe expanded to triangular lamella with tapering tip. Notopodial ventral ligule enlarged slightly. Most of atokous chaetae substituted by epitokous natatory ones; several atokous spinigers remaining in both notopodia and neuropodia.
Parapodia and chaetae in posterior body (behind chaetiger 104) similar to those of atokes ( Figure 8D View Figure 8 ).
Coelom filled with eggs; eggs spherical, 110–120 µm in diameter, with shrivelled surface (probably because of shrinkage of eggs due to long-term preservation).
Variation of paragnath number
Paragnath numbers in groups I–VI in Nectoneanthes oxypoda in 11 localities are summarized in Table 1 View Table 1 .
Reproduction
Epitokous mature males and females swarm at the surface in a long period from March to November in central and western Japan. Most of them swam within 3 hours after sunset, with no relation to high tide or phase of the moon.
Habitat
Sandy or muddy bottom in mainly subtidal (sometimes intertidal) coastal areas (up to 100 m deep).
Geographical distribution
Asian Pacific coasts north to Far East Russia and south to Vietnam, Persian Gulf, Southern Australia ( Figure 9 View Figure 9 ).
Remarks
The present results of the examination of the syntype and non-type specimens of Nectoneanthes oxypoda agreed well with Marenzeller’s (1879) original description and Wilson’s (1988) redescription of the syntypes in general. The presence of neuropodial falcigers in an early stage of juveniles is first recorded as a characteristic of this species by the present study. The other morphological characteristics were adequately drawn in Marenzeller (1879), Monro (1938), Wesenberg-Lund (1949), Paik (1973), Wu et al. (1985), He (1987) and Khlebovich (1996) for the atokous form, and in Gravier and Dantan (1934) and Imajima (1996) for the epitokous form.
Wilson (1988) regarded this species to be synonymous with Neanthes succinea ( Leuckart, 1847) , which was later transferred to the genus Alitta by Bakken and Wilson (2005), because the key characteristics of Nectoneanthes oxypoda (the absence of neuropodial falcigers in adults and the presence of an ovoid lobe above the dorsal cirri in notopodia) were judged to be characteristics associated with the development of epitoky in Neanthes succinea . However, the present study confirmed that the key characteristics are found consistently in both epitokous and atokous specimens of Nectoneanthes oxypoda , showing that N. oxypoda is not synonymous with Neanthes succinea . Moreover, Nectoneanthes oxypoda differs from Neanthes succinea (see details below) also in the following eight additional diagnostic characteristics: (1) larger maximum body size (BL up to 370 mm, with 212 chaetigers), in contrast to that of Neanthes succinea (BL up to 190 mm, with 160 chaetigers); (2) the morphology of the expanded notopodial dorsal ligule in posterior parapodia without conspicuous elongation, in contrast to that with conspicuous elongation in Neanthes succinea ( Figure 3C, F View Figure 3 ); (3) the presence of notopodial prechaetal lobes throughout, in contrast to the absence of the same in the posterior parapodia in Neanthes succinea ( Figure 3C, F View Figure 3 ); (4) the presence of the digitiform neuropodial inferior lobe with a tapering tip throughout as in the postchaetal lobe, in contrast to that in Neanthes succinea , which is not digitiform and has a round tip ( Figure 3 View Figure 3 ); (5) the small number of paragnaths (less than 15, usually less than 10) in group III in the proboscis, in contrast to the presence of 20–60 in Neanthes succinea ; (6) the presence of heterogomph spinigers in the upper neurochaetae, in contrast to the absence of the same in Neanthes succinea ( Figure 4 View Figure 4 ); (7) epitokous modification of the middle parapodia first appearing in chaetigers 17–20 in males and chaetigers 28–31 in females, in contrast to the first appearance of this modification in chaetigers 14–16 in males and chaetigers 14–19 in females in Neanthes succinea ; and (8) neuropodial postchaetal lobe developing into a square-shaped lamella with a digitate protrusion on the inner side of the lower edge in male epitokes, in contrast to that developing into a round lamella without digitate protrusion on the inner side of the lower edge in Neanthes succinea .
Wilson (1984) reported that among Australian specimens identified as Neanthes succinea , nine specimens collected from NSW (materials examined by Hutchings and Murray 1984) and five specimens from Swan River in WA (materials examined by Monro 1938) have fewer paragnaths (two to four) than most specimens collected from Victoria (20–30). I judged the former as Nectoneanthes oxypoda , and the latter as Neanthes succinea .
Nectoneanthes latipoda Paik, 1973 syn. nov., which was originally described from Korea and was later recorded from Japan ( Imajima 1996) and Far East Russia ( Khlebovich 1996), was in this study found to be synonymous with N. oxypoda , because its description agreed well with results of the present examination of N. oxypoda . The type of N. latipoda syn. nov. seems to be deposited in the laboratory of the late Professor E-I. Paik at the Department of Science Education, Hyosung Women’s College, Korea, but I have not examined it.
Nectoneanthes multignatha Wu, Sun and Yang, 1981 syn. nov. which was originally described from southern China and later reduced to a synonym of N. latipoda syn. nov. by Khlebovich (1996), was also in this study synonymized with N. oxypoda . Its description agreed well with results of the present examination of N. oxypoda . The holotype (No. A-62) and paratype (No. A-63) of N. multignatha syn. nov. seem to be deposited in the First Institute of Oceanography, Qingdao, China, but I have not examined them.
Nectoneanthes donghaiensis He, 1987 syn. nov. was described from the East China Sea. He (1987) distinguished it from N. oxypoda and N. multignatha syn. nov. by the characteristics of the shape of the eyes of the anterior pair (crescent), paragnath numbers, and the chaetiger number of the first occurrence of an ovoid lobe above dorsal cirrus (chaetiger 17). These characteristics, however, are within the range of the intraspecific variation of N. oxypoda , which was shown by the present examination (e.g. Table 1 View Table 1 ). Therefore N. donghaiensis syn. nov. is here synonymized with N. oxypoda . The holotype (No. 848E-50-1) and paratype (No. 848E-50-2) of N. donghaiensis syn. nov. were deposited in the Third Institute of Oceanography, SOA, Xiamen, China ( He, 1987), but I have not examined them.
Nereis ijimai Izuka, 1912 syn. nov. which was transferred to the genus Nectoneanthes by Imajima (1972), was originally described from the coast of central Japan based on a single extremely large specimen of an epitokous female, and later found in China ( Wu et al. 1985). This species was also judged to be synonymous with N. oxypoda . The holotype of Nereis ijimai syn. nov. agreed well in morphological characteristics with the epitokous females of N. oxypoda examined in the present study. Small numbers of paragnaths in the proboscis of the holotype of Nereis ijimai syn. nov., which have been regarded as a diagnostic characteristic for this species ( Imajima 1972, 1996; Wu et al. 1985; Khlebovich 1996), seem to be caused by the deletion of some paragnaths during the process of growing to a large body size. Izuka (1912) described the number of paragnaths of group VI to be six or seven in the holotype of N. ijimai syn. nov. However, I counted 12 on the right and 10 on the left, including several traces of paragnath bases remaining, indicating missing paragnaths. Srinivasa Rao and Rama Sarma (1982) recorded “ Nectoneanthes ijimai ” on the east coast of India. However, this seems to be a misidentification, judging from their drawing of a posterior parapodium without an ovoid lobe above the dorsal cirri.
Nereis alatopalpis Wesenberg-Lund, 1949 syn. nov. and Nereis singularis Wesenberg-Lund, 1949 syn. nov., which were transferred to the genus Nectoneanthes by Wu et al. (1985) and Khlebovich (1996), respectively, were originally described from the Iranian Gulf based on a single small specimen of each species; the former was later found in China ( Wu et al. 1985). They were also judged here to be synonymous with N. oxypoda . Characteristics of holotypes of the two species, described by Wesenberg- Lund (1949) and observed by myself, agreed well with the characteristics of N. oxypoda shown in the present study, with comparable paragnath numbers ( Table 1 View Table 1 ). The broad palps in the prostomium ( Figure 2D View Figure 2 ), which was regarded as a diagnostic characteristic for Nereis alatopalpis syn. nov. by Wesenberg-Lund (1949), seem to be within the range of the intraspecific variation of Nectoneanthes oxypoda observed in the present study.
Nereis (Nereis) legeri Gravier and Dantan, 1934 syn. nov., which was described from the coast of Vietnam based on a total of 81 epitokes collected in June and November, was also in this study synonymized with Nectoneanthes oxypoda , because its description agreed well with results of the present examination of N. oxypoda , except for the adult body sizes. All specimens of Nereis (Nereis) legeri syn. nov. 30–40 mm in BL and less than 3 mm in BW ( Gravier and Dantan 1934) are comparable with the smallest epitoke (33 mm BL, 2.5 mm BW) in the Japanese population of N. oxypoda found in the present study, and much smaller than the largest one (370 mm BL, 11.0 mm BW). The syntypes of Nereis (Nereis) legeri syn. nov. were deposited in the Muséum national d’Histoire naturelle, Paris (MNHN POLY TYPE 107, 108) ( Solís-Weiss et al. 2004), but they were not examined in the present study.
A specimen that was collected on the Spanish coast and described as Nereis (Neanthes) oxypoda by Ibáñez (1972) is similar to N. oxypoda in terms of the presence of an ovoid lobe above the dorsal cirri in the notopodia of chaetigers 20–70. However, the other characteristics of this specimen, such as the presence of neuropodial falcigers in the adult stage and the marked elongation of the notopodial dorsal ligule in posterior parapodia, agree with the diagnostic characteristics of Neanthes succinea (mentioned below). It is most probable that this specimen belongs to Neanthes succinea in the process of epitokous metamorphosis, just as in the case that led Wilson (1988) to question the validity of N. oxypoda .
| NHMW |
Naturhistorisches Museum, Wien |
| UMUTZ |
Department of Zoology, University Museum |
| NSMT |
National Science Museum (Natural History) |
| T |
Tavera, Department of Geology and Geophysics |
| OMNH |
Osaka Museum of Natural History |
| ZMH |
Zoologisches Museum Hamburg |
| MS |
Herbarium Messanaensis, Università di Messina |
| AM |
Australian Museum |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Nectoneanthes oxypoda ( Marenzeller, 1879 )
| Sato, Masanori 2013 |
Nectoneanthes singularis:
| Khlebovich VV 1996: 115 |
Neanthes latipoda: Wilson, 1988: 9
| Wilson RS 1988: 9 |
Neanthes multignatha: Wilson, 1988: 9
| Wilson RS 1988: 9 |
Nectoneanthes donghaiensis
| He M-H 1987: 349 |
Nectoneanthes alatopalpis:
| Khlebovich VV 1996: 115 |
| Wu BL & Sun R & Yang D 1985: 169 |
Neanthes oxypoda:
| Hutchings P & Murray A 1984: 39 |
Neanthes succinea: Wilson, 1984: 218–221
| Wilson RS 1984: 221 |
Nectoneanthes latipoda
| Choi J-W & Seo J-Y & Lee C-H & Ryu T-K & Sung C-G & Han G-M & Hyun S 2005: 387 |
| Lee J-H & Park J-Y & Lee H-G & Park H-G & Kim D 2003: 191 |
| Nishi K & Hanaoka M & Yamanishi R 1998: 199 |
| Imajima M 1996: 146 |
| Khlebovich VV 1996: 116 |
| Paik E-I 1973: 84 |
Nectoneanthes oxypoda: Imajima, 1972: 116–117
| Imajima M 1996: 145 |
| Imajima M 1972: 117 |
Nectoneanthes ijimai: Imajima, 1972: 117–119
| Imajima M 1996: 143 |
| Wu BL & Sun R & Yang D 1985: 161 |
| Imajima M 1972: 119 |
Nectoneanthes oxypoda
| Yamanishi R & Sato M 2007: 183 |
| Lee J-H & Park J-Y & Lee H-G & Park H-G & Kim D 2003: 189 |
| Imajima M 1996: 144 |
| Khlebovich VV 1996: 115 |
| Yang D & Sun R 1988: 39 |
| Wu BL & Sun R & Yang D 1985: 164 |
| Srinivasa Rao D & Rama Sarma DV 1982: 447 |
| Imajima M 1972: 113 |
| Ibanez M 1972: 24 |
Neanthes ijimai:
| Imajima M & Hartman, O 1964: 145 |
Nereis alatopalpis
| Wesenberg-Lund E 1949: 283 |
Nereis singularis
| Wesenberg-Lund E 1949: 281 |
Nereis (Nereis) legeri
| Gravier C & Dantan JL 1934: 61 |
Nereis (Neanthes) oxypoda:
| Uschakov PV 1965: 187 |
| Treadwell AL 1936: 268 |
| Fauvel P 1932: 22 |
Nereis oxypoda: Izuka, 1912: 171–173
| Monro CCA 1934: 362 |
| Mori T & Tamura M & Makino K 1932: 3 |
| Izuka A 1912: 173 |
Nereis ijimai
| Horst R 1918: 258 |
| Izuka A 1912: 176 |
Nereis (Alitta) oxypoda
| Marenzeller E 1879: 122 |