
Alpheus longipalma, Komai & Ohtomi, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4434.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:152955BF-B12D-43DD-8E3D-30689FC710E3 |
|
DOI |
https://doi.org/10.5281/zenodo.5965997 |
|
persistent identifier |
https://treatment.plazi.org/id/AF99BD81-CFD8-4685-B2C7-84ED83465F20 |
|
taxon LSID |
lsid:zoobank.org:act:AF99BD81-CFD8-4685-B2C7-84ED83465F20 |
|
treatment provided by |
Plazi |
|
scientific name |
Alpheus longipalma |
| status |
sp. nov. |
Alpheus longipalma View in CoL n. sp.
[New Japanese name: Satsuma-teppou-ebi]
Figs 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6
Material examined. Holotype: CBM-ZC 14482, male (cl 13.0 mm), T/V “Nansei-maru”, stn 5, center of Kagoshima Bay, 31°26.99’N, 130°37.78’E to 31°26.69’N, 130°37.96’E, trawl, depth 226.6–226.7 m, coll. J. Ohtomi, 13 October 2015.
Paratypes: CBM-ZC 14483, 3 males (cl 8.8–11.0 mm), 1 ovigerous female (cl 14.0 mm), same collection data as holotype; CBM-ZC 14484, 21 males (cl 9.5–12.8 mm), 4 females (cl 8.1–13.4 mm), T/V “Nansei-maru”, stn 5, similar locality, 31°26.93’N, 130°37.72’E to 31°26.79’N, 130°37.91’E, trawl, depth 226.3–227.2 m, coll. J. Ohtomi, 17 July 2016.
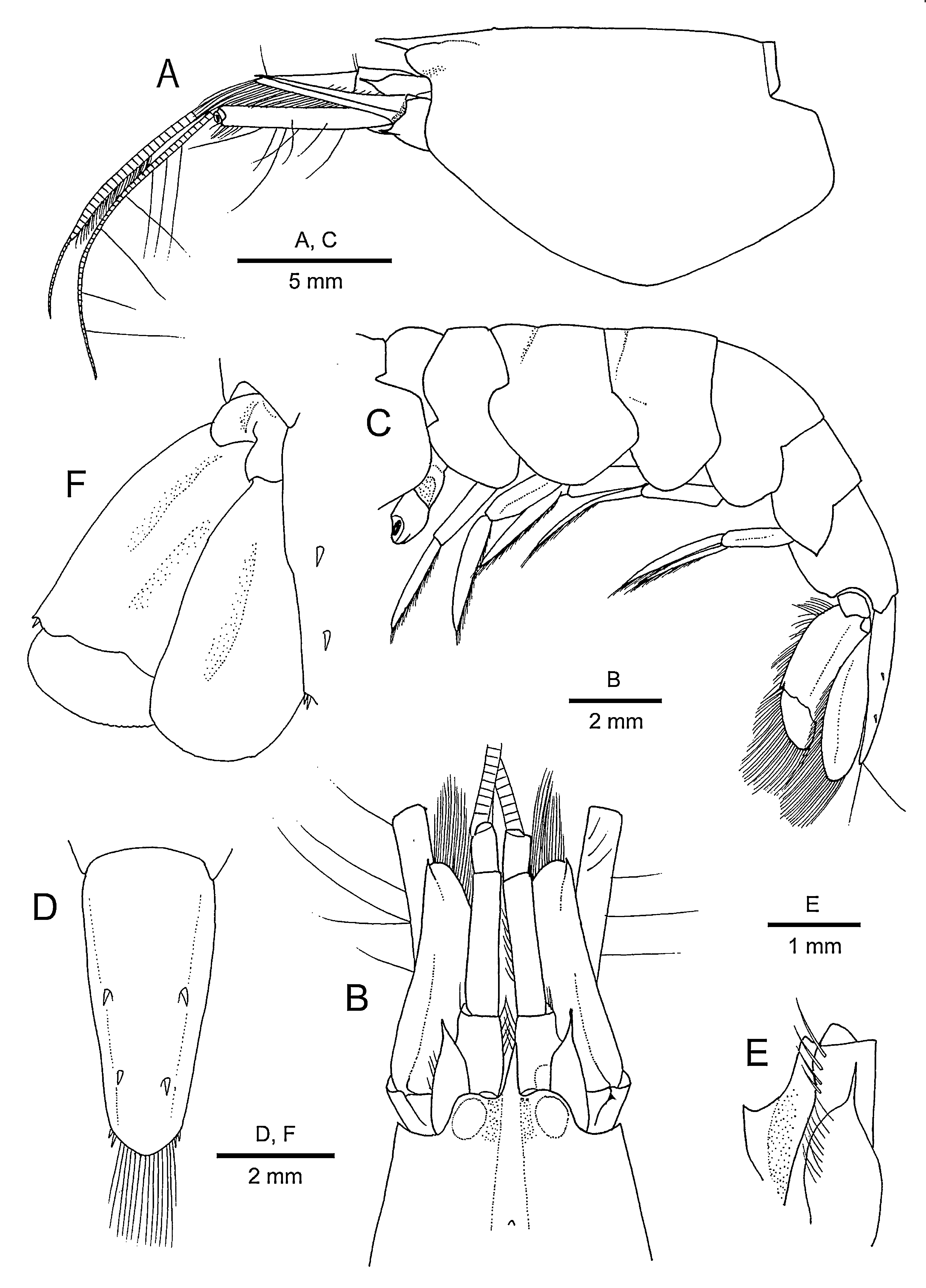
Description. Males. Body ( Figs. 1A, B View FIGURE 1 , 2A, C View FIGURE 2 ) moderately slender. Rostrum ( Fig. 2A, B View FIGURE 2 ) slender, slightly compressed laterally, overreaching mid-length to reaching distal margin of article 1 of antennular peduncle, directed forward or slightly ascending; dorsal carina blunt, not extending to gastric region of carapace. Carapace ( Fig. 2A, B View FIGURE 2 ) smooth, glabrous, moderately compressed laterally; adrostral grooves shallow; gastric region with minute median tubercle; dorsum nearly straight in lateral view, slightly sloping in anterior part of gastric region to rostral apex; orbital hoods not particularly inflated, anteriorly unarmed; anterior margin between rostrum and orbital hoods slightly concave; pterygostomial angle broadly rounded; cardiac notch deep, V-shaped.
Pleon ( Fig. 2C View FIGURE 2 ) subcylindrical, not particularly compressed laterally; pleura 1–4 all generally rounded, pleuron 2 subtriangular; pleuron 5 armed with small posteroventral spine; pleomere 6 without articulated flap, preanal area not protruding as spine, posterolateral process obliquely truncate. Telson ( Fig. 2D View FIGURE 2 ) gradually narrowing posteriorly, about 2.3 times as long as proximal width; dorsal surface with 2 pairs of small dorsolateral spiniform setae, situated at some distance from lateral margins, first pair located at 0.4–0.5 of telson length, second pair at 0.7; posterior margin convex, with 2 pairs of unequal spiniform setae at posterolateral angles (mesial spiniform setae much longer than lateral spiniform setae) and with row of long plumose setae between them.
Eyes completely concealed by orbital hoods in both dorsal and lateral views ( Fig. 2A, B View FIGURE 2 ), anteromesial margin slightly produced, with minute blunt tubercle; cornea darkly pigmented ( Fig. 1 View FIGURE 1 ). Ocellar beak not conspicuously protruding between eyes.
Antennular peduncle ( Fig. 2A, B View FIGURE 2 ) relatively slender, overreaching distal margin of scaphocerite (antennal scale) by half-length of article 3. Article 1 bearing large triangular lamina with subacute apex on ventromesial carina ( Fig. 2E View FIGURE 2 ); stylocerite somewhat inflated proximally, terminating in slender spine reaching nearly to distal margin of article 1 ( Fig. 2E View FIGURE 2 ). Article 2 about 4.0 times or more as long as broad. Lateral flagellum with numerous tufts of aesthetascs on about 12 articles; secondary ramus rudimentary.
Antenna ( Fig. 2A, B View FIGURE 2 ) with basicerite bearing slender spine at ventrolateral distal angle. Carpocerite relatively slender, distinctly overreaching lamella of scaphocerite, with row of sparse long setae on ventromesial margin. Scaphocerite 0.4 times as long as carapace and about 2.8 times as long as wide, greatest width at proximal 0.2 of length; lateral margin slightly concave, terminating in small distolateral spine slightly overreaching distal margin of lamella; cleft between lamella and distolateral spine slit-like, deep; lamella rounded distally.
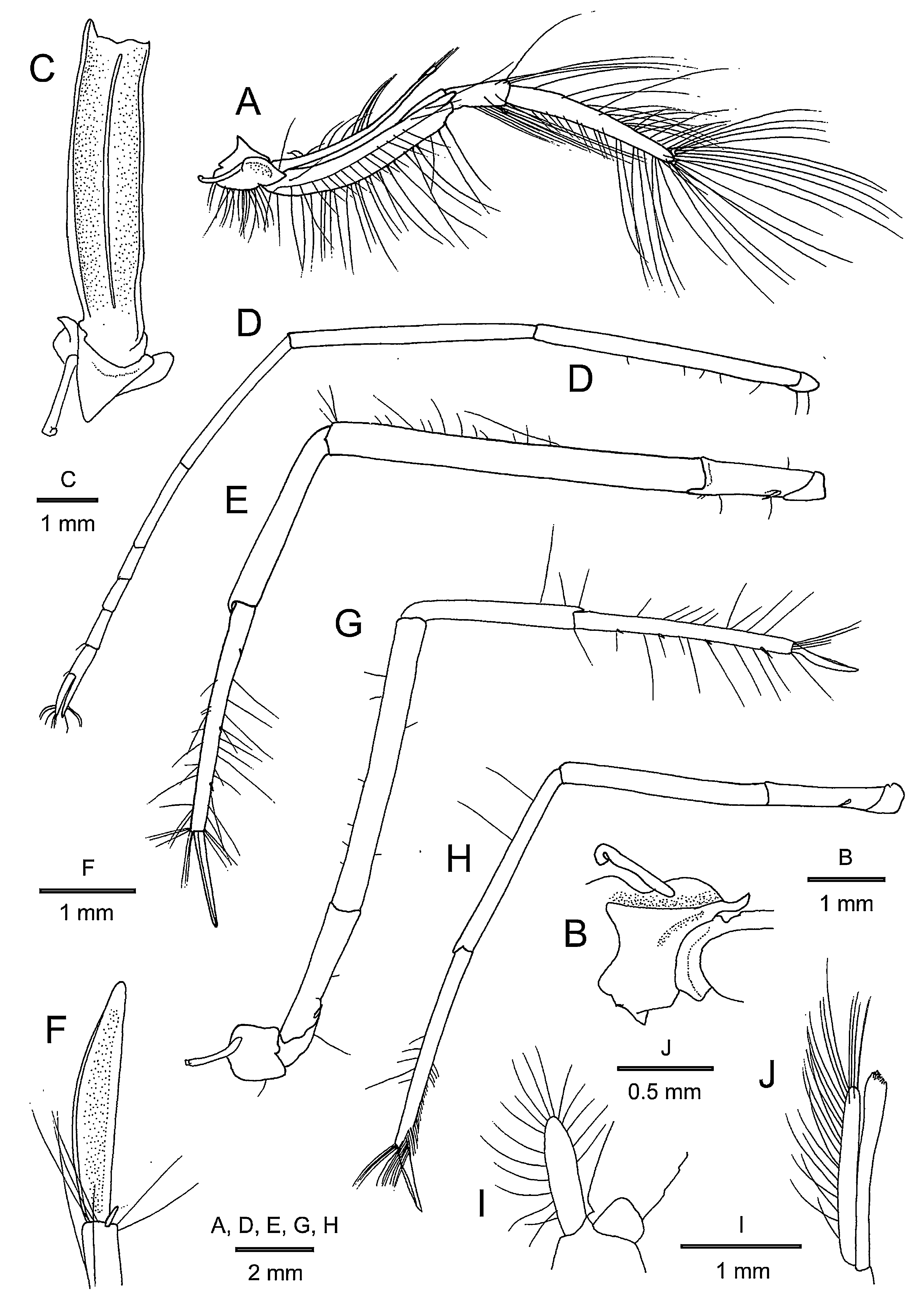
Mouthparts typical for Alpheus in external observation. Maxilliped 3 ( Fig. 5A–C View FIGURE 5 ) moderately slender, reaching level of distal end of antennal scaphocerite. Coxa with lateral plate terminating distally in subacute, laterally curving, triangular tooth ( Fig. 5B View FIGURE 5 ); ventral surface of coxa crested laterally, continuous with lateral plate. Antepenultimate article strongly flattened dorsoventrally in proximal half; dorsolateral margin forming distinct keel with sharply edged dorsal and ventral margins, produced distally into blunt process, latter with tuft of very long setae; dorsomesial margin sharply carinate over entire length; ventral surface with sharp, crest-like carina mesial to mid-line ( Fig. 5C View FIGURE 5 ), with row of short to long setae. Penultimate article slightly flattened dorsoventrally, slightly widened distally, with very long setae on distal margin. Ultimate article 2.8 times as long as penultimate article, gradually tapering, trigonal in cross-section, with scattered tufts of elongated setae on dorsolateral surface and on blunt tip ( Fig. 5A View FIGURE 5 ).
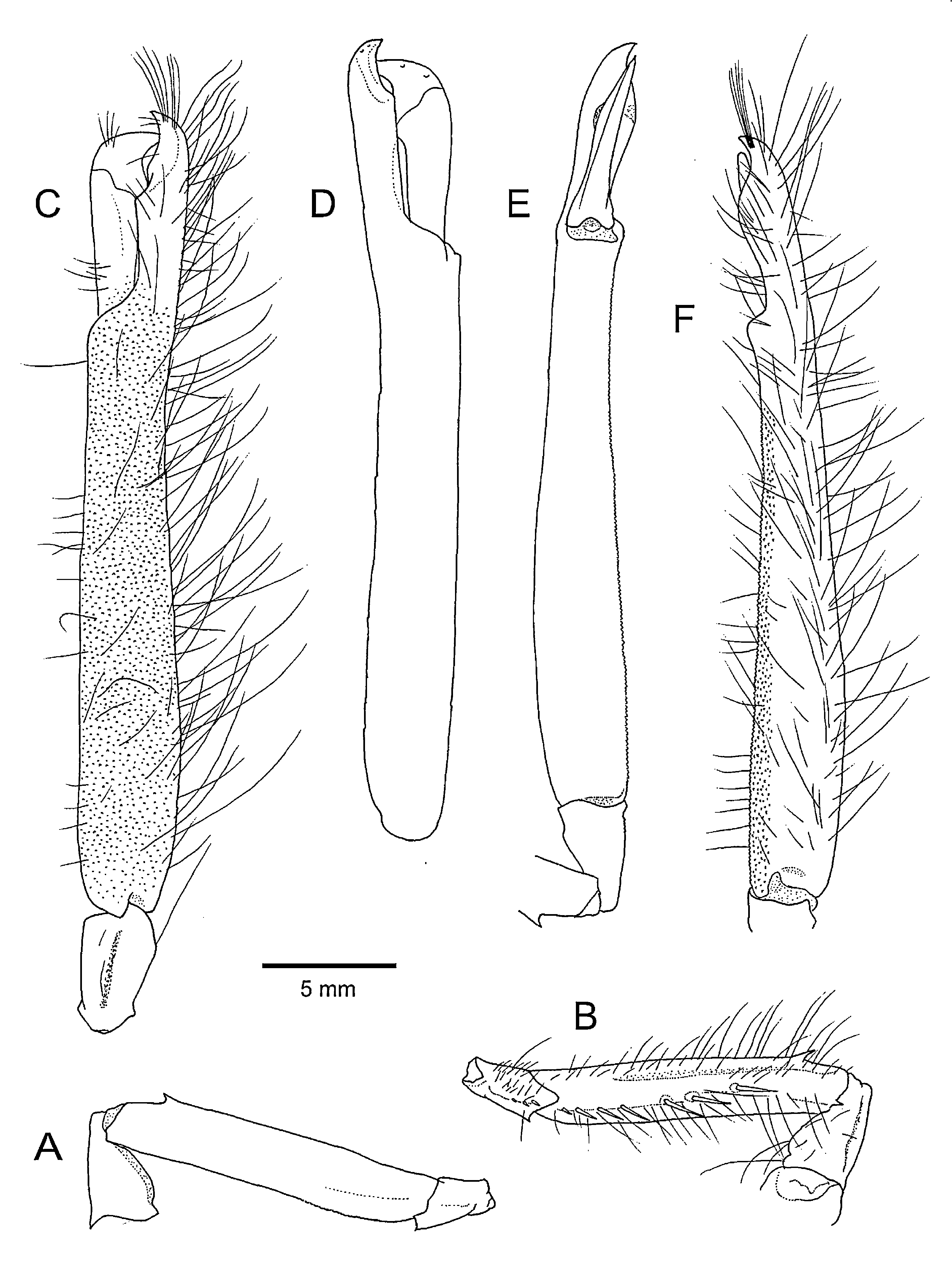
Major cheliped ( Figs 1A, B View FIGURE 1 , 3 View FIGURE 3 ) elongate, slender, longer than body in large individuals. Ischium short; ventral surface bicarinate, ventrolateral margin terminating in acute or subacute tooth, ventromesial margin armed with row of 3–5 minute spiniform setae. Merus slightly broadened distally, trigonal in cross-section; dorsal margin forming sharp crest-like carina bearing sharp subdistal tooth; ventral surface distinctly bicarinate, ventrolateral margin minutely granulate, ventromesial margin armed with small sharp distal tooth and 4–8 widely spaced, elongate spiniform setae in proximal 0.5–0.7 of its length, their bases typically formed by small protuberances. Carpus very short, cup-shaped; dorsal (extensor) surface with shallow longitudinal groove on mid-line; ventral (flexor) surface slightly inflated mesially. Chela somewhat compressed, suboval in cross section, strongly elongate, about 8 times as long as greatest width, sometimes reaching triple carapace length ( Fig. 1B View FIGURE 1 ), slightly upcurved; lateral margin nearly straight, mesial margin slightly sinuous, with shallow concavity proximal to base of fixed finger. Palm about 6 times as long as wide, with greatest width at about proximal 0.2 length, without conspicuous grooves or depressions on minutely granulate surfaces; distal part proximal to base of dactylus somewhat inflated. Pollex about 0.3 times as long as palm, terminating in curved, subacute tip; dorsal side of socket forming low convexity. Dactylus strongly flattened, nearly straight in longitudinal plane, with blunt tip not reaching tip of pollex; lateral (extensor) margin sharply keeled; plunger reduced, located distal to mid-length of dactylus, obliquely truncate, defined only by proximal angle. Margins of each article of chela bearing sparse but numerous short and long setae, particularly on mesial margin of chela.
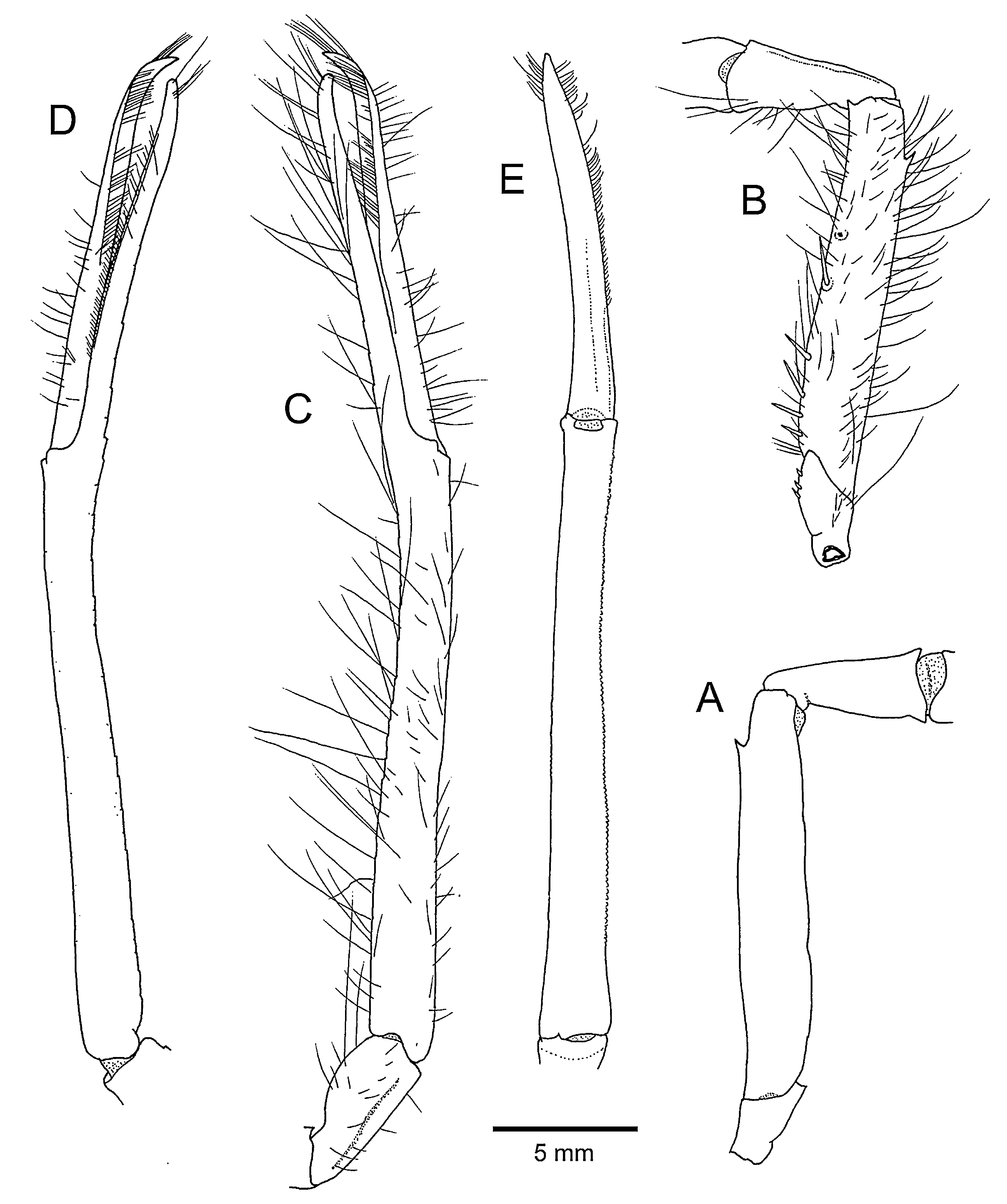
Minor cheliped ( Figs. 1A, B View FIGURE 1 , 4 View FIGURE 4 ) balaeniceps in males, longer than major cheliped. Ischium and merus generally similar to those of major cheliped, merus, slightly more slender. Carpus cup-shaped, longer and more slender than that of major cheliped. Chela extremely slender, subcylindrical, about 13 times as long as wide, without marked depressions or grooves, slightly upcurved. Palm about 8 times as long as wide, surfaces minutely granulate, mesial face with minute protuberances furnished with setae. Fingers slender, noticeably curved near tip, crossing; dorsal and ventral sides each with oblique balaeniceps ridges in distal half; occlusal margins each with thin corneous blade; dactylus 0.66–0.84 (mean 0.74, n=15) times as long as palm.
Pereopod 2 ( Fig. 5D View FIGURE 5 ) slender, overreaching distal end of antennal carpocerite by length of chela and 0.8–0.9 length of carpus. Ischium slightly longer than merus; carpus divided into 5 articles, with approximate ratio 1: 0.6: 0.2: 0.2: 0.3; chela about 0.2 times as long as carpus, fingers distinctly longer than palm.
Pereopod 3 ( Fig. 5E View FIGURE 5 ) slender, overreaching distal end of antennal carpocerite by full length of propodus, moderately setose. Ischium with 1 small spiniform seta on ventrolateral surface. Merus unarmed, about 10 times as long as wide. Carpus unarmed. Propodus about 1.4 times as long as carpus; ventral (flexor) margin with 1 or 2 spiniform setae and row of long simple setae; ventrodistal margin with 1 minute spiniform seta obscured by tuft of long setae. Dactylus ( Fig. 5F View FIGURE 5 ) subspatulate, terminating in subacute or blunt tip, about half as long as propodus.
Pereopod 4 ( Fig. 5G View FIGURE 5 ) similar to pereopod 3, slightly shorter, overreaching distal end of antennal scaphocerite by 0.3–0.4 length of propodus.
Pereopod 5 ( Fig. 5H View FIGURE 5 ) overreaching scaphocerite by 0.2 length of propodus. Ischium unarmed or armed with 1 spiniform seta on ventrolateral surface. Propodus with grooming brush consisting of several transverse rows of stiff setulose setae on distal half; one small spiniform seta present at ventrodistal angle (usually concealed by grooming setae, not visible in lateral view); dactylus narrower than that of third or fourth pereopods.
Male pleopod 1 with endopod about 0.2 length of exopod ( Fig. 5I View FIGURE 5 ); pleopod 2 with appendix masculina ( Fig. 5J View FIGURE 5 ) subequal in length to appendix interna, furnished with numerous slender stiff setae on lateral surface and apex.
Uropod ( Fig. 2F View FIGURE 2 ) with protopod armed with 2 distal processes, lateral process rounded, mesial process terminating in subacute tooth. Exopod with nearly straight lateral margin, terminating in small posterolateral tooth, adjacent spiniform seta very small; diaeresis faintly trilobed; distal margin without spiniform setae dorsal to main setal fringe.
Gill/exopod formula typical of Alpheus .
Females. Pleuron of pleomere 2 wider and more rounded than in males. Chelipeds ( Fig. 6A, B View FIGURE 6 ) proportionally shorter than in males; major cheliped with chela 1.0–1.5 times as long as carapace; minor cheliped slightly shorter to subequal in length to major cheliped, chela 1.2–1.5 times as long as carapace, dactylus 0.90–1.04 (mean 0.95, n=4) times as long as palm, with obliquely longitudinal carina on dorsal and ventral surfaces, without balaeniceps setae.
Colouration in life. Body generally dominated by red chromatophores ( Fig. 1 View FIGURE 1 ). Carapace reddish orange dorsally, paler on flanks. Pleon with broad transverse red bands on lighter semitransparent-reddish background, bands concentrated near posterior margin of pleomeres. Antennules, antennae, maxilliped 3 and tail fan red-orange generally, flagella paler. Chelipeds almost entirely orange-red, except for paler ischium. Pereopod 2 reddish orange generally, except for transparent chela; pereopods 3 and 4 with ischia and meri reddish and distal three articles transparent; pereopod 5 and pleopods transparent.
Distribution and habitat. Presently known only from Kagoshima Bay, Kyushu, Japan, at depths of 226–227 m, on muddy bottoms, probably burrowing as most other species of the A. brevirostris group (e.g., Anker 2000).
Remarks. The present new species is referred to the Alpheus brevirostris species group mainly because of the compressed palm of the major cheliped and the subspatulate dactylus of the pereopods 3 and 4. One of the most important features of A. longipalma n. sp. is the possession of extremely elongate, slender chelipeds. Amongst about 50 species presently assigned to the A. brevirostris species group ( Bruce 1994; Hayashi & Nagata 2000, 2002; Anker & Dworschak 2004; Anker et al. 2007; Bracken-Grissom & Felder, 2014; Komai 2015; Ramos-Tafur 2018), similarly elongate chelipeds are also seen in A. kagoshimanus (also found in Kagoshima Bay, see above), A. acutocarinatus de Man, 1909 , A. explorator Boone, 1935 , A. macroskeles Alcock & Anderson, 1894 , A. migrans Lewinsohn & Holthuis, 1958 , A. nonalter Kensley, 1969 , A. notabilis Stebbing, 1915 and A. talismani Coutière, 1898 . The new species is morphologically most similar to A. macroskeles , known with certainty only from the northeastern Indian Ocean (Bay of Bengal and Andaman Sea), and A. talismani from the eastern Atlantic ( Portugal to Angola), in the lack of a distinct postrostral carina on the carapace and possession of a subdistal tooth on the meri of both chelipeds. In A. acutocarinatus , A. explorator , A. kagoshimanus , A. migrans and A. notabilis , the carapace has a distinct postrostral carina extending at least to the gastric region. The cheliped meri are unarmed on the dorsal or distodorsal margin in A. acutocarinatus , A. explorator , A. kagoshimanus , A. migrans and A. notabilis (cf. de Man 1909, 1911; Stebbing 1915; Boone 1935; Lewinsohn & Holthuis 1958; Hayashi & Nagata 2000), whereas they are armed with a distodorsal, i.e. clearly distal (not subdistal) subacute or blunt tooth in A. nonalter ( Kensley 1969; Chace 1988; Komai 2011).
Alpheus macroskeles View in CoL was originally described on the basis of material from two stations in the Bay of Bengal [off Godévari Delta, 270 fms (= 486 m); off Plicat, 145–250 fms (= 261–450 m)], “Swatch” (193 fms = 347 m) and one station from the Andaman Sea (193 fms = 347 m) ( Alcock & Anderson 1894). In the original description, no name bearing type was designated. Subsequently, Alcock (1901: 141) designated one specimen from the Bay of Bengal (off Godévari Delta, 486 m) as “type” and Banner & Banner (1978: 224; 1981: 232) interpreted this specimen as holotype. Coutière (1898) provided a figure of the anterior portion of the carapace and cephalic appendages of a specimen of A. macroskeles View in CoL sent from A. Alcock for comparison with his A. talismani View in CoL . Alcock & Anderson (1895) published a figure of A. macroskeles View in CoL , showing the habitus of the species in lateral view. Since then, there have been several regional records from various parts in the Indo-West Pacific under this name, for example, from the Red Sea ( Balss 1915), Gulf of Aden ( Calman 1939), Indonesia (de Man 1911; Chace 1988), South China Sea ( Banner & Banner 1978), the Philippines ( Banner & Banner 1981; Chace 1988) and Taiwan ( Wang et al. 2013), with de Man (1911), Banner & Banner (1978) and Chace (1988) also providing some morphological details, including illustrations, on their specimens. De Man (1911) expressed some doubts about his own identification, because his specimen (one ovigerous female from the Bali Sea) was lacking both first pereopods and exhibited some slight differences when compared to the descriptions by Alcock & Anderson (1894) and Alcock (1901), and the figure of Coutière (1898). Banner & Banner (1978) compared the type material (holotype and two paratypes) of A. macroskeles View in CoL deposited in the Indian Museum, Calcutta, with four specimens from the South China Sea, noting that “These are excellent agreement with the specimens in the present collections.” However, Chace (1988) stated that the only authentic records of A. macroskeles View in CoL were those mentioned by Alcock & Anderson (1894), i.e. from the Bay of Bengal and Andaman Sea.
Considering the possibility that some records of A. macroskeles View in CoL in previous taxonomic accounts may refer to other species, we refer herein only to the original description of that species ( Alcock & Anderson 1894) and the subsequent accounts by Coutière (1898) and Alcock (1901), along with the illustration of Alcock & Anderson (1895). We believe that the following differences between A. macroskeles View in CoL and the specimens from Kagoshima Bay warrant the establishment of a new species, A. longipalma View in CoL n. sp.:
1 The minor cheliped is subequal to or longer than the major cheliped in males of A. longipalma n. sp. ( Fig. 1A, B View FIGURE 1 ). In contrast, the figure of A. macroskeles in Alcock & Anderson (1895: pl. IX, fig. 5) clearly shows the minor cheliped being distinctly shorter than the major cheliped.
2 The dactylus of the minor cheliped is distinctly shorter than the palm and has balaeniceps setae in males of A. longipalma n. sp. ( Fig. 4 View FIGURE 4 ). In contrast, both Alcock & Anderson (1894: 153) and Alcock (1901: 141) specifically noted that the dactylus was equal in length to the palm in their male specimens of A. macroskeles . Neither Alcock & Anderson (1894) nor Alcock (1901) mentioned the presence of balaeniceps setae on the dactylus of the minor cheliped in their descriptions of A. macroskeles .
3 Alcock & Anderson (1894) specifically noted that setae on the chelipeds are few and fine as to be invisible to the naked eye, in A. macroskeles . However, in A. longipalma n. sp., the chelipeds can be considered as fairly setose (cf. Figs 3 View FIGURE 3 , 4 View FIGURE 4 , 6 View FIGURE 6 ).
4 The rostrum appears to be relatively more slender and longer in A. longipalma n. sp. (cf. Fig. 2B View FIGURE 2 ) than in A. macroskeles (more than 3 times longer than the basal width versus approximately 2 times, cf. Coutière 1898: fig. 3’).
5 The pigmentation of the eye was said to be markedly deficient in the original description of A. macroskeles ( Alcock & Anderson 1894) , whilst the cornea is darkly pigmented in A. longipalma n. sp. ( Fig. 1 View FIGURE 1 ). Noteworthy, most specimens of A. macroskeles were dredged from depths greater than 400 m, whereas the Japanese material of A. longipalmus sp. n. was trawled from less than 230 m.
The new species is readily distinguished from A. talismani in the relatively shorter article 2 of the antennular peduncle and shorter antennal scale, with a better developed, rounded distal lamella ( Fig. 2A View FIGURE 2 ). Alpheus talismani has a very slender article 2 of the antennular peduncle and more elongate antennal scaphocerite, with a narrow, obliquely truncate distal lamella (cf. Coutière 1898: fig. 3; Holthuis 1951: fig. 14a).
The occurrence of Alpheus nonalter in Japanese waters has been recently confirmed by Komai (2011). This species is generally very similar to A. longipalma n. sp., but in addition to the different position of the dorsal tooth on the cheliped meri (see above), A. nonalter differs from the new species in the shorter antennular peduncle (not reaching the distal margin of the antennal scaphocerite in A. nonalter vs. distinctly overreaching it in the new species) and in the more developed distolateral tooth of the scaphocerite, which is reaching far beyond the blade in A. nonalter ( Chace 1988; Komai 2011) and barely overreaching it in A. longipalma n. sp. ( Fig. 2A View FIGURE 2 ). As pointed out by Komai (2011), the original illustrations of A. nonalter by Kensley (1969) are not very accurate; therefore, illustrations provided by Chace (1988) and Komai (2011) should be used for interspecific comparisons.
Banner & Banner (1978: fig. 1) provided figures of selected parts from two specimens identified as A. macroskeles from the South China Sea off Hong Kong. However, these specimens are substantially different from the descriptions and/or figures of A. macroskeles provided by Alcock & Anderson (1894, 1899), Coutière (1898) and Alcock (1901), in several aspects. Their pleura 2 and 3 are broadly rounded, even in males; the second article of the antennular peduncle is more robust; the antennal scaphocerite has a gently concave lateral margin, an elongate distolateral tooth and a narrow, distally obliquely truncate blade (vs. having a nearly straight lateral margin, moderately short distolateral tooth and moderately broad, distally rounded blade). Alpheus longipalma n. sp. also differs from Banners’ Hong Kong material, for instance, in the longer article 2 of the antennular peduncle; the non-elongate distolateral tooth and rounded blade of the antennal scaphocerite; the much more slender palm of the minor cheliped; the absence of spiniform setae on the dorsal margin of the cheliped meri; the presence of a sharp distoventral tooth on the cheliped ischia; the absence of long spiniform setae on the flexor margin of the propodus and of spiniform seta on the ischium of the third pereopod. We therefore believe that the Hong Kong specimens represent a different deep-water species of the A. brevirostris group, closely related to A. macroskeles and Alpheus longipalma n. sp.
Chace (1988: fig. 8) illustrated two male specimens from Indonesia that he identified as A. macroskeles . They differ from the present specimens of A. longipalma n. sp. in the shorter rostrum, more strongly produced distolateral tooth of the antennal scaphocerite, and the dactylus of the minor cheliped being subequal in the length to the palm. Thus Chace’s specimens seem to agree with A. macroskeles in all diagnostic characters, except for the development of the distolateral tooth of the antennal scaphocerite. Their taxonomic identity remains to be confirmed by a more careful morphological comparison and perhaps molecular analyses.
Etymology. The specific epithet is a combination of the Latin, longus (= long) and palma (= palm), in reference to the greatly elongate palms of the chelipeds in the present new species; used as a noun in apposition.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Alpheus longipalma
| Komai, Tomoyuki & Ohtomi, Jun 2018 |
Alpheus longipalma
| Komai & Ohtomi 2018 |
A. longipalma
| Komai & Ohtomi 2018 |
A. talismani
| Coutiere 1898 |
Alpheus macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |
A. macroskeles
| Alcock & Anderson 1894 |