
Hungarosoma bokori Verhoeff, 1928
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4178.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:466B4B88-F8C2-4321-80F8-59E38F8EBAE9 |
|
DOI |
https://doi.org/10.5281/zenodo.5631232 |
|
persistent identifier |
https://treatment.plazi.org/id/03C087BB-473D-FFAF-FF6D-2090FAFDFBF2 |
|
treatment provided by |
Plazi |
|
scientific name |
Hungarosoma bokori Verhoeff, 1928 |
| status |
|
Hungarosoma bokori Verhoeff, 1928
( Table 2 View TABLE 2 , Figs 1–17 View FIGURE 1 View FIGURES 2 – 5 View FIGURES 6 – 9 View FIGURES 10 – 11 View FIGURES 12 – 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 )
Hungarosoma bokori: Verhoeff, 1928: 192 –195 (description from a female); Verhoeff 1932: 1485 –6, 1893 (diagnosis, notes on systematics and distribution); Gebhardt 1933; 1963; 1967; (all faunistic notes from type locality); Gebhardt 1934: 203 –207 (reprint of the original description); Moritz & Fischer 1978 (data on type material); Loksa 1961 (first record outside the type locality); Korsós 2000 (new record on the type locality); Papáč et al. 2014 (first record in the Carpathians).
Ochogona moravica nomen nudum: Skoumalová, 2010: 229 –230, 324 (preliminary diagnosis of a presupposed new species, drawings orig. P. Kocourek, faunistic data from the Moravian Karst).
Material examined. Holotype: Hungary: Abaliget Höhle (=cave), ZSM-A- 20032282: 1♀ , 12.viii.1924, leg. E. Bokor , det. K. W. Verhoeff, mounted on permanent microscopic slide, dissected. Deposited in The Bavarian State Collection of Zoology in Munich, Germany .
Topotypoid material. Hungary: Abaliget (cave?), ZMB 13596: 1♀, without date and collector, det. K. W. Verhoeff, complete body mounted on a permanent slide, deposited in the Museum für Naturkunde in Berlin; Abaliget Cave, material determined by K. W. Verhoeff: ZSM-A-200322811: 1 juvenile, permanent slide, without date and collector; ZSM-A-20034336: 1 juvenile ; ZSM-A-20060246: 1 subadult male and 3 juveniles in a vial with ethanol, all deposited in The Bavarian State Collection of Zoology in Munich, Germany.
New material from type locality. The cave at the end of main corridor (at siphon), sampled on the old cavetimber from oak wood, 18.iv.2013, 1♂, 3♀, 5 juveniles (stages III, VI, VII), leg. A. Mock.
Other examined material. Austria, 2 ♀: Lower Austria: Waldviertel ( Southern Bohemian Massif ), Albrechtsberg an der Großen Krems , Sommerbachtal N of church, montane beech forest on metamorphites as bedrock, on right hand slope (NW exposition) between a footpath higher up and the creek, collected from valley floor a few meters upwards, 28.x.1999, hand collecting, 1 ♀, leg. J. Gruber; NMW 8831 View Materials ; 15.xi.2000, leaf litter sifting, 1 ♀, leg. J. Gruber, NMW 8832.
Czech Republic, 47 ♀: Moravian Karst : Hostěnice near Ochoz u Brna , litter of Melampyro nemorosi- Carpinetum (mesophilous oak-hornbeam forest “Mokerský les” and forest/meadow ecotone), 13.x.2005, 1 ♀, leg. P. Kocourek ; 28.x.2005, 7 ♀, leg. P. Kocourek; 15.iv.2006, 1 ♀, leg. P. Kocourek; 23.iii.–19.x.2007, pitfall traps, 2 ♀, leg. P. Kocourek; 19.x.2007, 20 ♀, leg. P. Kocourek; 19.x.2008, 2 ♀, leg. P. Kocourek; 3.x.2009, 4 ♀, leg. P. Kocourek; 15.x.2011, 7 ♀, leg. K. Tajovský & P. Kocourek; 15.x.2011 – 7.v.2012, pitfall traps, 1 ♀, leg. K. Tajovský; 5.xi.2012 – 31.v.2013, pitfall traps, 2 ♀, leg. K. Tajovský; 16.ix.2013 – 28.iv.2014, pitfall traps, 2 ♀, leg.
K. Tajovský. Note: the samples from the years 2005–2008 are presented in the diploma thesis of Skoumalová (2010) under the name (nomen nudum) “ Ochogona moravica ”.
Hungary, 2 ♀, 1 juv.: Aggtelek Karst : Aggtelek , litter of Corneto-Quercetum at the foothills of reefs at the main entrance of Baradla, 21.iii.2013, 2 ♀, leg. A. Mock ; Vass Imre Cave , leaf litter of hornbeam-oak forest concentrated at the lower entrance to the corridor of discoverers, extracted organic material, 5.ix.2013, 1 juv. (IV st.), leg. A. Mock .
Slovakia, 36 ♀: Slovak Karst : Čertova diera Cave, litter of Corneto-Quercetum in the entrance hole, 23.x.1997, 1 ♀, leg. A. Mock ; 22.x.1999, 1 ♀, leg. A. Mock; 1.x.2000, 2 ♀, leg. A. Mock; 8.iii.2001, 1 ♀, leg. A. Mock; 24.ix.2003, 3 ♀, leg. K. Tajovský; 28.xi.2013, 2 ♀, leg. K. Tajovský & A. Mock; Líščia diera Cave, leaf litter of Corneto-Quercetum at the entrance, 1.xii.2000, 1 ♀, leg. A. Mock ; 13.iii.2012, 3 ♀, leg. A. Mock; 28.xi.2013, 4 ♀, leg. K. Tajovský & A. Mock; Domica Cave (artificial corridor to the Styx river), 10 m from the entrance, on wood, 9.x.2003, 1 ♀, leg. A. Mock ; Domica Cave , 30 m from the entrance of an artificial corridor, remainder of mine-timber on the ceiling, 3.x.2001, 1 ♀, leg. A. Mock ; Silická Plateau , sifting of leaf litter under bushes at the edge of Jašteričie jazierko sinkhole, 1.x.2010, 1 ♀, leg. A. Mock ; Gombasek Cave , Čierna vyvieračka Spring, litter and humus layers at the spring, 24.ix.2003, 2 ♀, leg. K. Tajovský ; New Brzotínska Cave , litter of oakhornbeam forest nearby the cave entrance, 3.x.2005, 2 ♀, leg. K. Tajovský ; Brzotínska Spring , litter of oakhornbeam forest nearby the spring, 3.x.2005, 1 ♀, leg. K. Tajovský ; forested scree slope at the Ardovská Cave , subterranean traps with water solution of ethylene glycol, 5 cm and 45 cm under the surface, 27.x.2014 – 29.iv.2015, 2 ♀, leg. P. Ľuptáčik; Revúcka vrchovina ( Drienčanský Karst ): Blh River valley, alluvium and foot of the slopes with Malá Drienčanská Cave, forest, sifted litter and soil, 18.x.2013, 1 ♀, leg. A. Jászayová ; Malé Karpaty : Driny Cave, 2001–2003, pitfall traps, 1 ♀, leg. O. Majzlan, P. Zvonár ; small corridor at the site “ Chodba spolupracovníkov”, sampled on wood, 29.iv.2005, 2 ♀, leg. P. Ľuptáčik ; same place, 29.iv.–13.ix.2005, pitfall traps, 3 ♀; litter at the entrance, 11.xi.2009, 3 ♀, leg. A. Mock.
Re-diagnosis. Body length 6.0– 6.6 mm, width 0.5–0.6 mm. The same habitus in both sexes, but females overgrow males moderately. Number of ocelli in the adults (11) differs from that in H. inexpectatum (9). Pale brownish colouration. Rounded paranota with notch prior to the front line of them ( Fig. 3 View FIGURES 2 – 5 ). Macrosetae long. In males two pairs of legs are modified as diversified copulatory organs, the 8th and 9th. Compared to its congener, H. inexpectatum , the cheirites of gonopods in H. bokori are elongated, with a flattened and rewound central part and the tail in a shape like “two fingers” ( Figs 13–16 View FIGURES 12 – 14 View FIGURE 15 View FIGURE 16 ), while in H. inexpectatum the gonopod cheirites are drawn short with a simple tail.
General morphology. Selected morphological characteristics of Hungarosoma bokori are summarised in Table 2 View TABLE 2 . Coloration: The small body of the earlier postembryonic stadia is without colouration. The subadults (stadium VII) have a poorly pigmented head; only the frons is light brown in colour. The adult stadium (VIII) is light brown, with a darker head, antennae and dorsal part of trunk segments; the metazonites are darker than the prozonites; without sexual dimorphism ( Figs 1 View FIGURE 1 , 2 View FIGURES 2 – 5 , 6, 7 View FIGURES 6 – 9 ).
Setation and paranota: Typical rounded paranota (pleurotergite processes) with triplets of long setae are present in both sexes during all postembryonic development ( Figs 3 View FIGURES 2 – 5 , 7–9 View FIGURES 6 – 9 ). There is a characteristic notch prior to the front line of the paranota. Setae are quite long (up to 0.18 mm) and slightly curved, sticking up from the body segments. Two setae are on the paranota, one after another. There is almost a right angle between the connecting line of both setae and the third, the short median seta (96–99º).
Surface: The surface of the body is smooth, but the edges of the paranota are toothed due to outgoing scales of the cuticle ( Figs 3 View FIGURES 2 – 5 , 10, 11 View FIGURES 10 – 11 ).
Eyes: Juveniles up to the stadium III are eyeless; then the number of ocelli increases from 5 (stadium IV) to 11 (stadium VIII, adults), forming a triangular field on both sides of the head. The first ocellus is separated from the triangular field of ocelli more behind and often smaller (“semi-oculus”), apparently rudimentary.
Extremities: No other specific characteristics on the antennae ( Figs 4 View FIGURES 2 – 5 , 12 View FIGURES 12 – 14 ) or legs, except the ones found in the male mating apparatus, were observed.
Male characters. The male is less robust than the female, with a body length of 6.0 mm, a body width of 0.5 mm.
Anterior legs 1–7 are slightly shorter and more robust than the legs following the gonopods, but without other species-specific characteristics. The tarsus carries a chain of short, erect setae, ordered in a moderate spiral (in both sexes).
Legs 8 modified to anterior or 'proper' gonopods ( Figs 13–16 View FIGURES 12 – 14 View FIGURE 15 View FIGURE 16 ). This apparatus consists of a short frontal median process followed by long and thick cheirites growing from the robust proximal parts. The cheirites are curved twice toward the posterior gonopods, similar to the shape of the neck of the common heron or duck, with long beak-like apices that are better visible laterally ( Figs 14 View FIGURES 12 – 14 , 16 View FIGURE 16 ). The ends of the cheirites are flattened with divided tails like fingers and/or the pedipalps of scorpions or nippers of crayfish ( Figs 15 View FIGURE 15 a, 16a). Medially, behind the cheirites, are paired delicate brush-like arms with dense fimbricated setae on their slightly extended flat top ( Fig. 16 View FIGURE 16 b), followed by another pair of stronger arms. Laterally the stronger arms seem to be completed in a simple projection, but the anterior view shows they are curved toward the median plane and divided into two “teeth” like a repair spanner ( Fig. 15 View FIGURE 15 c). These arms extend to half the length of the cheirites. In the median zone between the spanner-like arms and close to the posterior gonopods ( Fig. 15 View FIGURE 15 -e) there are two hyaline processes with rounded tips equipped apically with short spines ( Fig. 15 View FIGURE 15 d).
Leg 9 or posterior gonopods ( Figs 15 View FIGURE 15 e, 16e) are strongly reduced, formed by a uniformly wide basal (coxal?) segment, with long setae, the length of which increases towards the top of the gonopods. The tops of the posterior gonopods are armed by a prominent claw-shaped process, with the tip pointed medially and with long seta on a lateral base ( Figs 15 View FIGURE 15 g, 16g). This claw-shaped process seems to be a rudiment of the following podomeres of the original walking leg. Next-to this structure, a short hyaline prominence occurs in front of the ventral edge ( Figs 15 View FIGURE 15 f, 16f). The surface of the posterior gonopods is nodulated, especially on the anterior and posterior sides ( Fig. 16 View FIGURE 16 ).
Legs 10 and 11 are of full length and differ from the other legs only by the presence of coxal sacs for sperm material.
Female characters. The females are more robust; their body length reaches up to 6.6 mm, and body width is about 0.6 mm. No modifications on the legs were observed.
Vulvae fused to form one complex and are free of any postvulval structures ( Figs 5 View FIGURES 2 – 5 , 17 View FIGURE 17 ). Each bursa has an oval shape, with a roof-like structure on the top. Setation is present on the inner side of the bursae: 4 long setae consecutively one after another, with the longest one on the top. A short setation (5 or 6 in number) is located on the lateral edge of each bursa. At the lateral base of the bursae are 2 long setae. The receptaculum is a branch-free, pipe-shaped tube. The opercula are coupled together. Their distal edge is curved toward the bursae. Four very short setae are present on the external (anterior) surface of the opercula. They are ordered diagonally.
Molecular analysis. Analysis in BOLD (Barcoding of Life Database) via the “species identification” module revealed that H. bokori was thus far not registered; therefore, the three top matches are reported below. In H. bokori , two haplotypes were found among the 8 sequences, one in the samples from the Slovak/Aggtelek Karst and the Moravian Karst, while the second was found both in south-western Hungary (Abaliget) and the Slovak/ Aggtelek Karst, thus, not in clear dependence on geography ( Table 3).
The intraspecific distance (Kimura 2 parameter) of H. bokori was rather low, 0–0.4%, with an overall average of 0.2% (SE =0.1%). The optimal neighbor-joining tree, with the sum of the branch length =1.67893986, is presented ( Fig. 18 View FIGURE 18. A ). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) is shown next to the branches (values>45% are shown). The tree is drawn to scale, with branch lengths in the same units (the number of base substitutions per site) as those of the genetic distances (Kimura 2 parameter) used to infer the dendrogram. The analysis involved 64 nucleotide sequences, and the tree was rooted with Polyxenus lagurus .
The presence of the two haplotypes is also visible on the dendrogram ( Fig. 18 View FIGURE 18. A ), where all the samples with the same haplotype clustered together, although the cluster of haplotype 2 is not strongly supported. Within the dendrogram Hungarosoma represents a separate cluster. The proximity of H. bokori to some representatives of the families Attemsiidae ( Dendromonomeron oribates ( Latzel, 1884) , Syngonopodium aceris Verhoeff, 1913 , S. cornutum (Verhoeff, 1929)) and Neoatractosomatidae ( Pseudocraspedosoma grypischium (Rothenbühler, 1900)) is suggested rather than to the other Craspedosomatidae genera (species) or even to the Brachychaeteuma -species ( Brachychaeteumatidae ). However, the boostrap support is too low to draw more strict conclusions and more thorough analysis is needed.
Order Family Genus Species Similarity (%) Hungarosoma bokori - haplotype 1
Chordeumatida Attemsiidae Syngonopodium cornutum 85.74 Chordeumatida Chordeumatidae Chordeuma sylvestre 84.46 Chordeumatida Chordeumatidae Chordeuma sylvestre 84.19 Chordeumatida Craspedosomatidae not identified not identified 84.19 Chordeumatida Attemsiidae Schubartia lohmanderi 84.19 Chordeumatida Neoatractosomatidae Pseudocraspedosoma brentanum 84.19 Chordeumatida Attemsiidae not identified not identified 84.02 Chordeumatida Chordeumatidae Chordeuma sylvestre 84.02 Chordeumatida Craspedosomatidae Craspedosoma rawlinsii 83.85 transsilvanicum
Chordeumatida Craspedosomatidae Listrocheiritium noricum 83.85

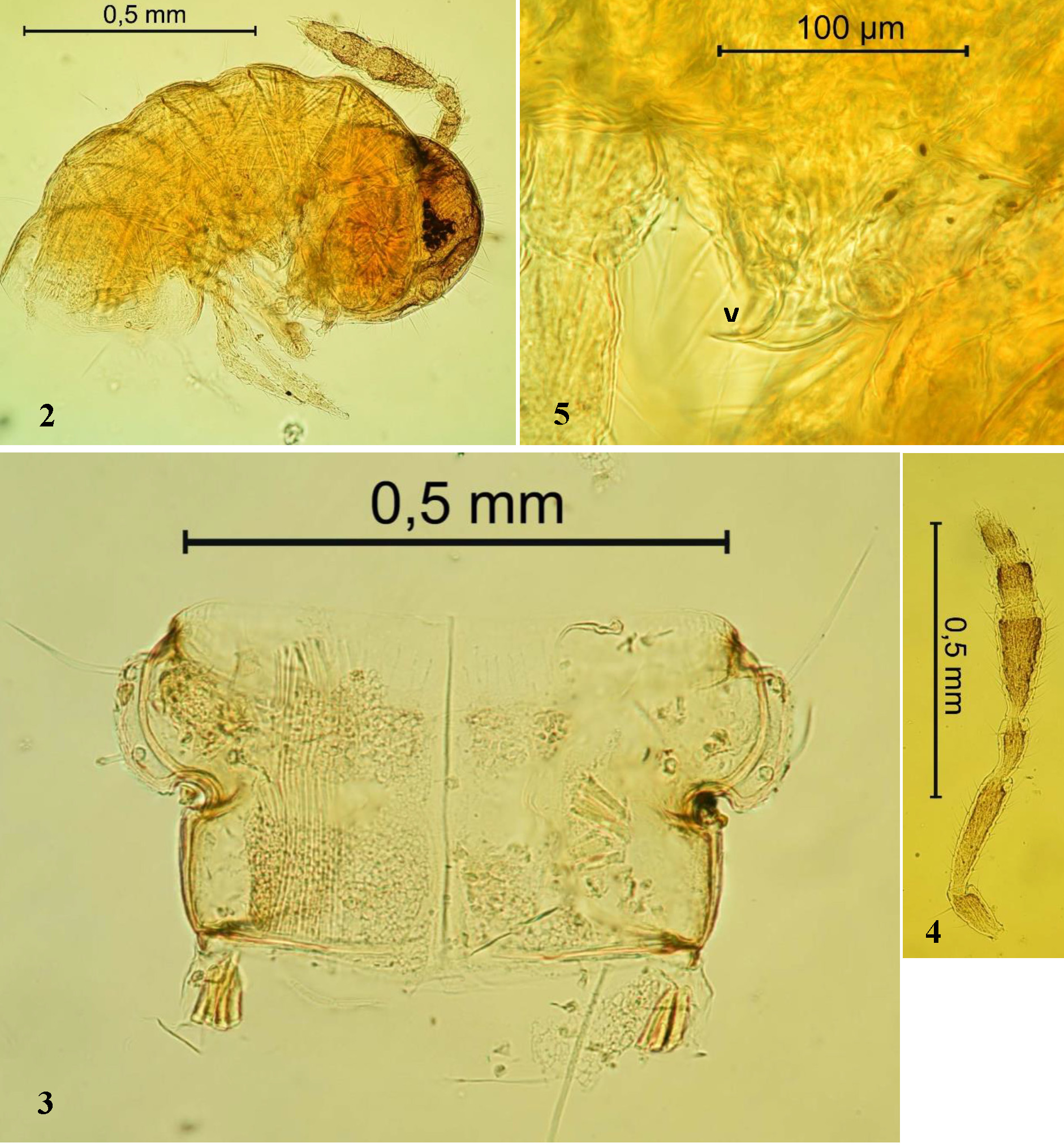

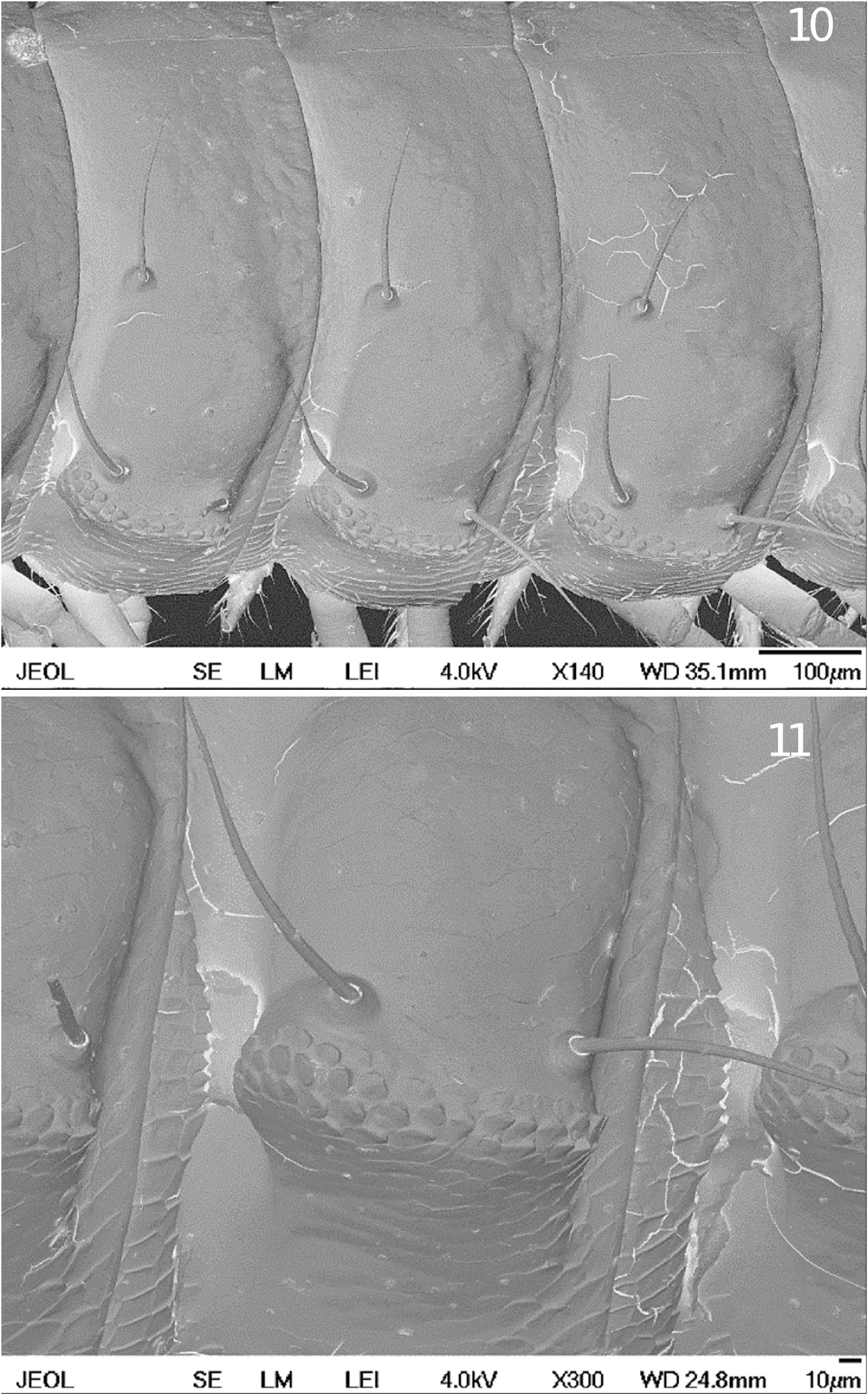
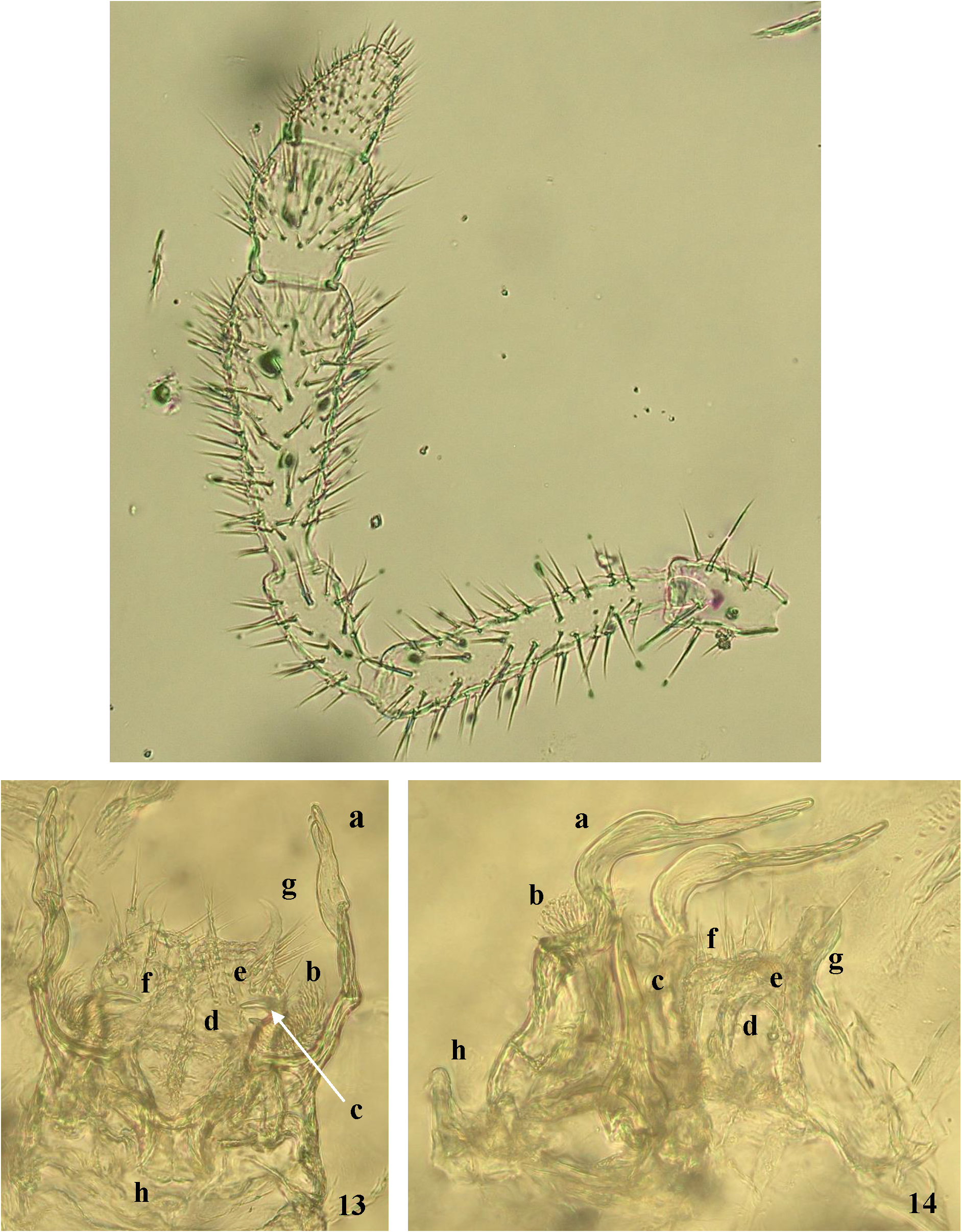
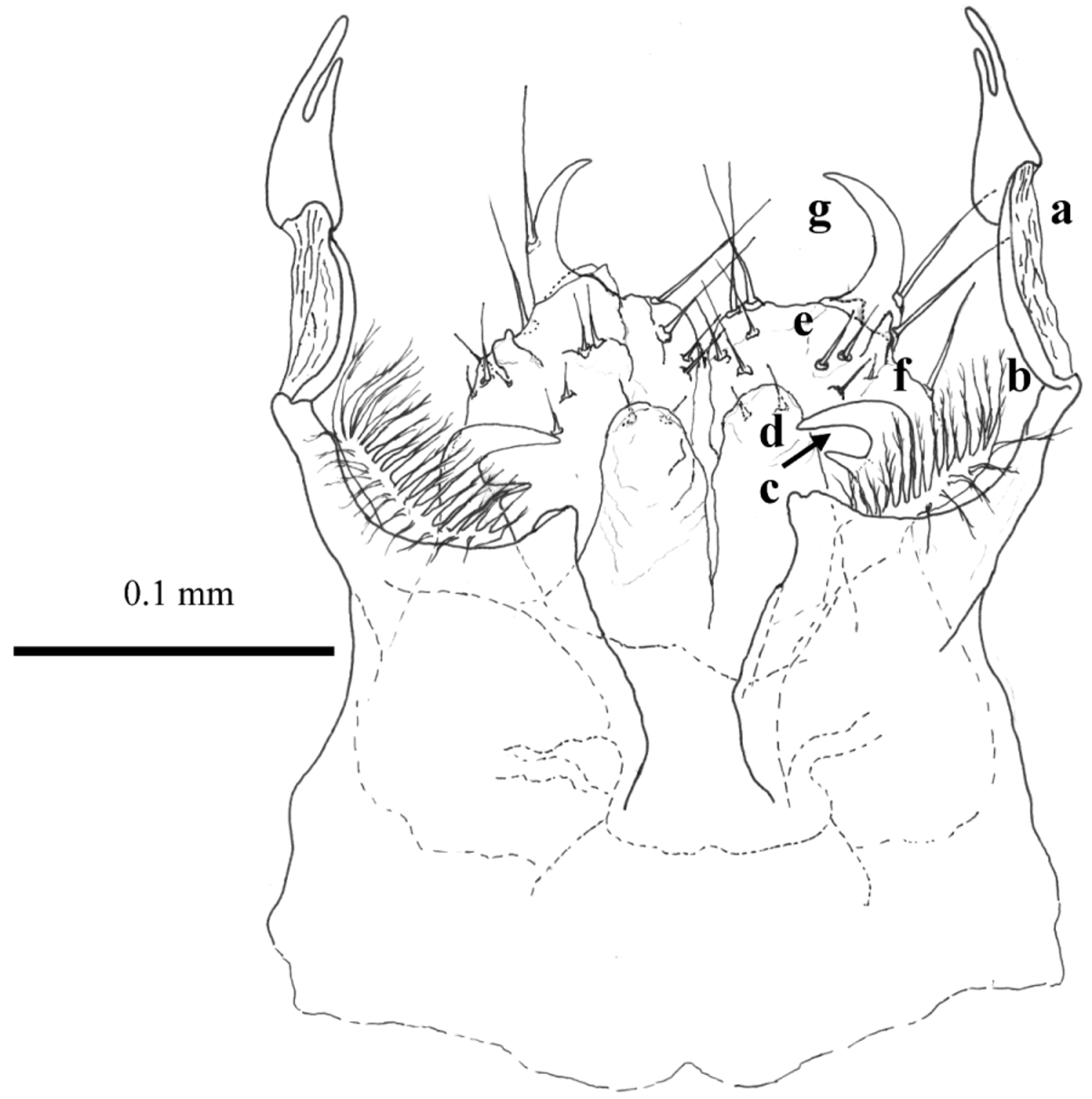
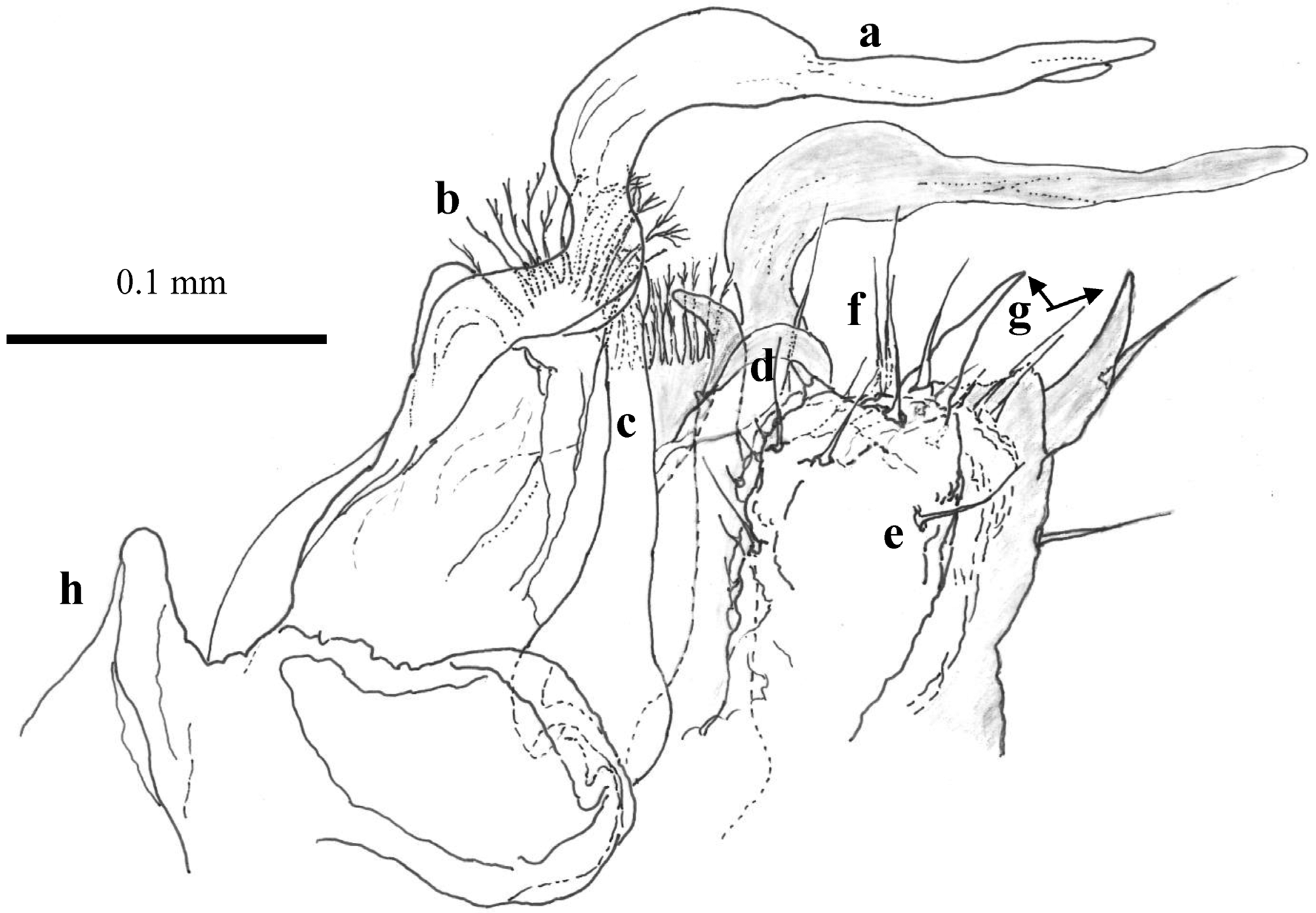
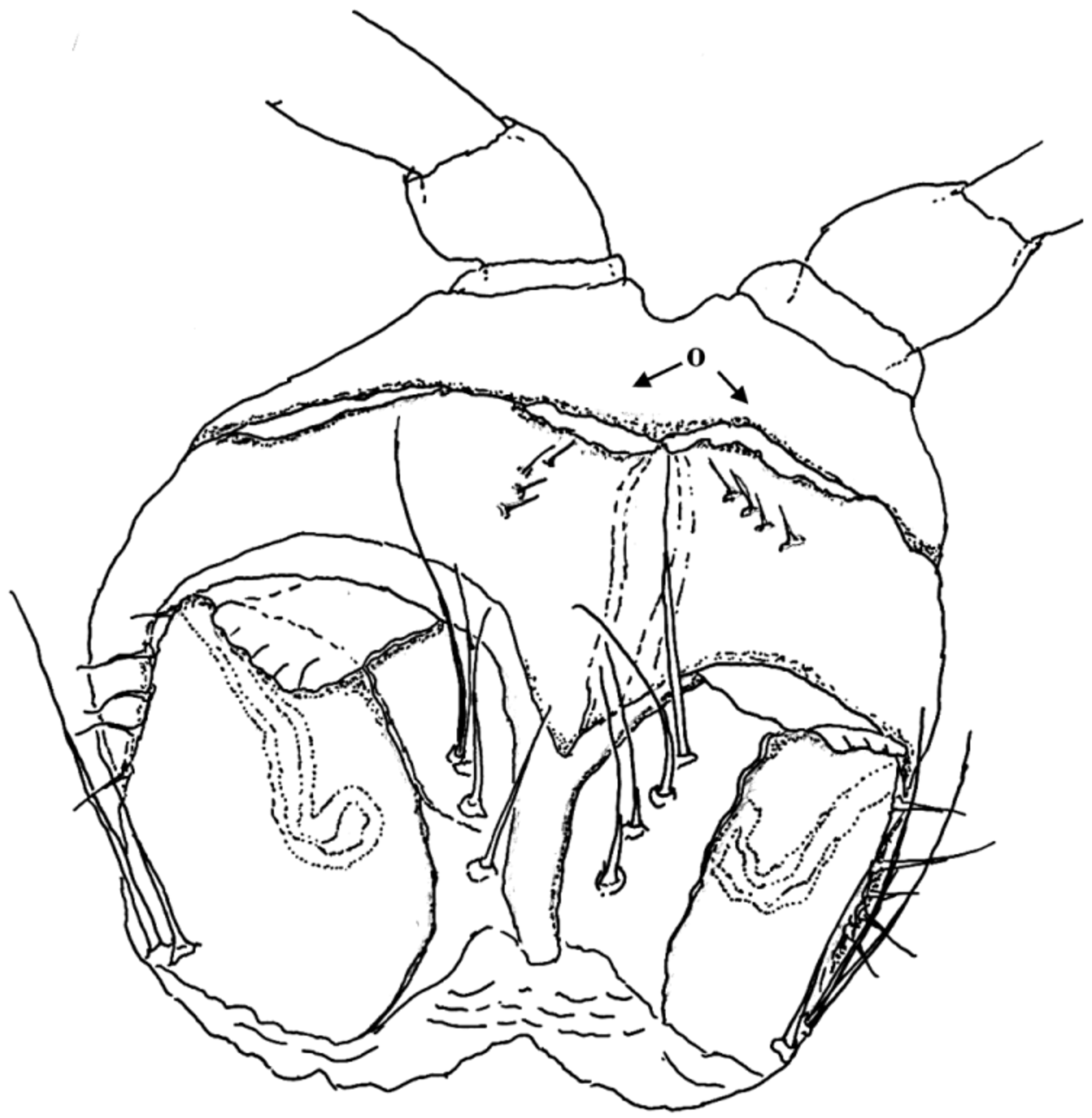
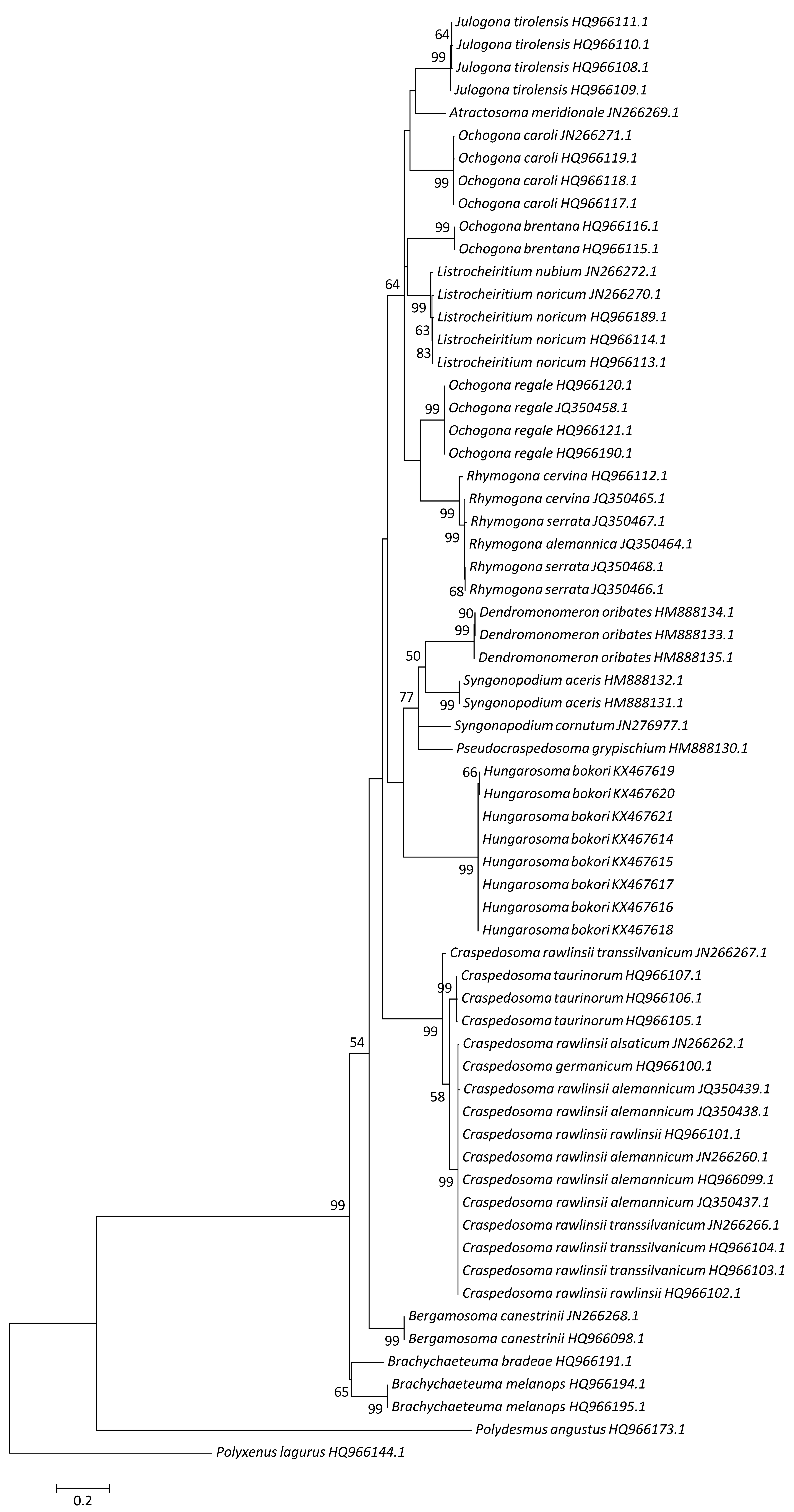
TABLE 2. Some characteristics of the anamorphosis of H. bokori from the type locality. The counting of pleurotergites was done according to Blower (1985). * — the first smaller (semi) ocellus separated from triangular field of ocelli to more posterior location on the head. Stadia I – II and IV – V were not found. Colour of cuticle: ○○○ — without colour; ● ○○ — light-coloured frons of the head, trunk white; ●●● — finely coloured head and trunk. Body length, height and width in mm.
| Character | Stadium III | VI | VII | VIII (male) | VIII (female) |
|---|---|---|---|---|---|
| Pleurotergites | 11 | 23 | 26 | 28 | 28 |
| Leg pairs | 10 | 32 | 40 | 44 | 46 |
| Ocelli | 0 | 4+1* | 6+1* | 10+1* | 10+1* |
| Body length | 1.9 | 4.2 | 5.2 | 6.0 | 6.2–6.6 |
| Body height | 0.2 | 0.3 | 0.4 | 0.4 | 0.5 |
| Body width | 0.3 | 0.4 | 0.4 | 0.6 | 0.6–0.7 |
| Colour | ○○○ | ○○○ | ●○○ | ●●● | ●●● |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hungarosoma bokori Verhoeff, 1928
| Mock, Andrej, Tajovský, Karel, Žurovcová, Martina, Jarošová, Andrea, Kocourek, Pavel, Gruber, Jürgen, Angyal, Dorottya & Spelda, Jörg 2016 |
Ochogona moravica
| Skoumalova 2010: 229 |
Hungarosoma bokori:
| Gebhardt 1934: 203 |
| Verhoeff 1932: 1485 |
| Verhoeff 1928: 192 |