
Cyrtodactylus tahuna, Riyanto & Arida & Koch, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4399.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:3F55047D-F272-4B5E-AACA-85379B535A2D |
|
DOI |
https://doi.org/10.5281/zenodo.5986471 |
|
persistent identifier |
https://treatment.plazi.org/id/03EE87EB-1F73-FFFB-FF01-F933FD717CA1 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus tahuna |
| status |
sp. nov. |
Cyrtodactylus tahuna sp. nov.
Common name: Tahuna bent-toed gecko Indonesian name: Cicak Jari-lengkung Tahuna Figs. 2–5 View FIGURE2 View FIGURE 3 View FIGURE 4 View FIGURE 5
Holotype ( Fig. 2A View FIGURE2 , 3A View FIGURE 3 ). MZB.Lace.5123 (field number AK028), an adult male collected on 8 July 2005 in Kecamatan (district) Tahuna , Kabupaten (regency) Kepulauan Sangihe, Sangihe Island, Sulawesi Utara, Indonesia (3o36’47” N; 125o30’11” E; 30 m) by E. Arida and A. Koch. GoogleMaps
Paratypes ( Fig. 2B–C View FIGURE2 , 3 View FIGURE 3 ). MZB. Lace.5097 (field number AK032), an adult female, same data as for holotype except for collection date, i.e., 10 July 2005. MZB . Lace.5133 (field number AK029), an unsexed juvenile, same data as for holotype.
Diagnosis. Cyrtodactylus tahuna sp. nov. can be readily distinguished from the congeners on Sulawesi, the Moluccas, and the Lesser Sunda Islands by the following unique combination of characters: (1) medium size, with a SVL reaching 79.2 mm, (2) brachium and antebrachium tuberculated, (3) raised conical tubercles along ventrolateral body folds, (4) 49–50 ventral scales, (5) precloacal depression in shape of a pit, (6) 14 precloacal pores arranged in the shape of a ‘ ˄’, (7) enlarged precloacal and femoral scales separated by smaller scales, (8) enlarged femoral scales in two rows, (9) precloacal and femoral pores separated by poreless scales, (10) five femoral pores on both sides, (11) 20–24 lamellae under fourth toes, (12) no transversely enlarged median subcaudals, (13) tail not prehensile, and (14) tubercles extend along about 60% of tail length.
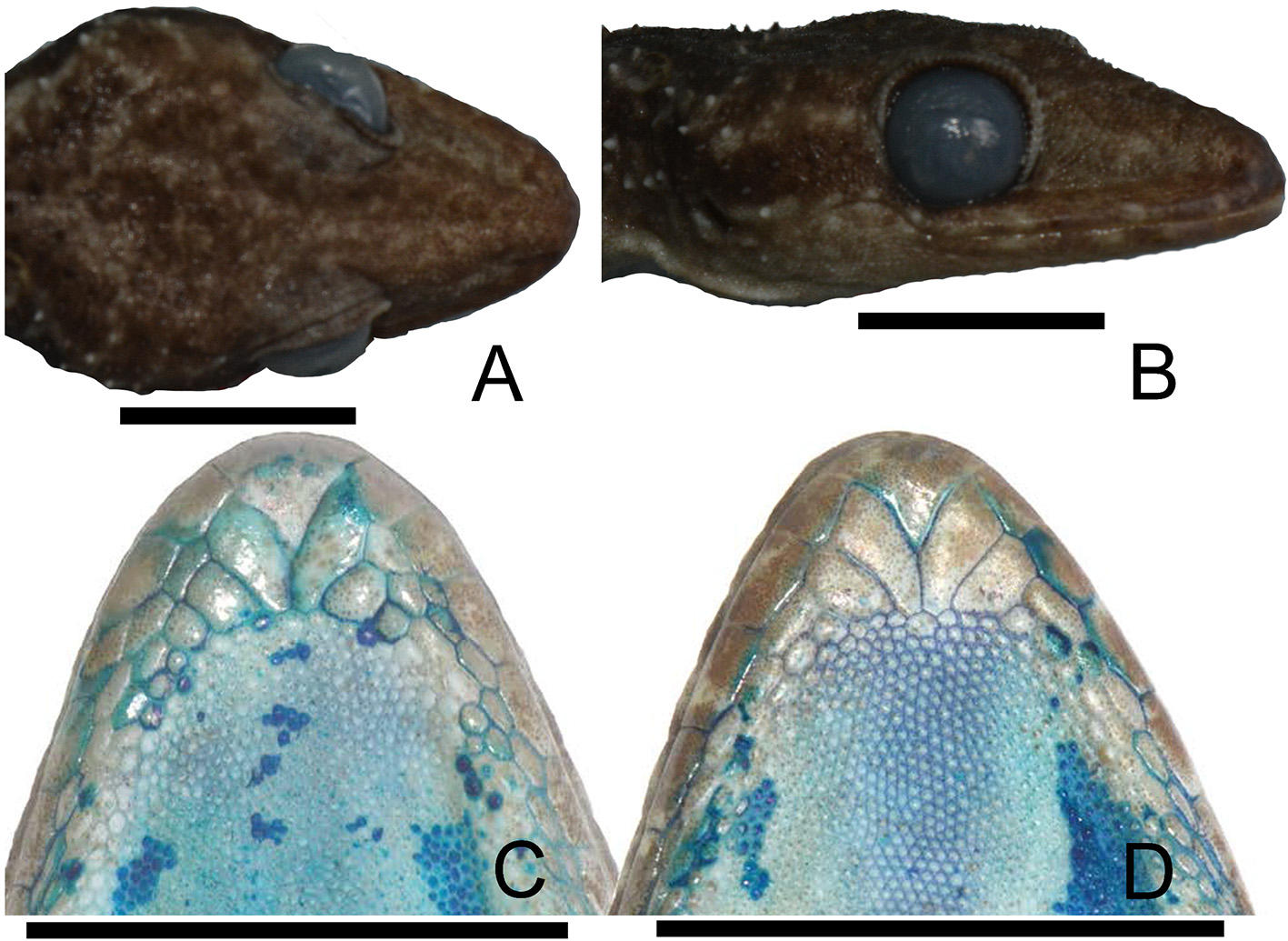
Description of holotype. An adult male, SVL 78.5 mm; head triangular in dorsal view ( Fig. 4A View FIGURE 4 ), distinct from neck; HeadL 31.1% of SVL, HeadW 61.6% of HeadL, and HeadH 11.2% of SVL; prefrontal region concave, canthus rostralis rounded ( Fig. 4B View FIGURE 4 ); SnoutL 39.6% of HeadL, OD 27.7% of HeadL, EarL 7.7% of HeadL and OD 98.0% of EyeEar.
Rostral rectangular, approximate height about 60% of width; bordered posterolaterally by first supralabials and naris, and dorsally by three postrostral scales; naris oval, bordered anteriorly by rostral, anterodorsally by one postrostral, posteriorly by five scales on right side and by six scales on left side, and ventrally by first supralabials; orbit separated from supralabials by two rows of small lorilabial scales; 13 SuL on right side; 10 InfL on right.
Mental triangular, length 73.7% of width; bordered laterally by first infralabials, posteriorly by one pair of enlarged first postmentals, which are in contact medially over about 40.2% of their length; second postmentals smaller, divided, and separated from each other by three granular scales ( Fig. 4C View FIGURE 4 ); gular scales small, granular, slightly grading in size posteriorly.
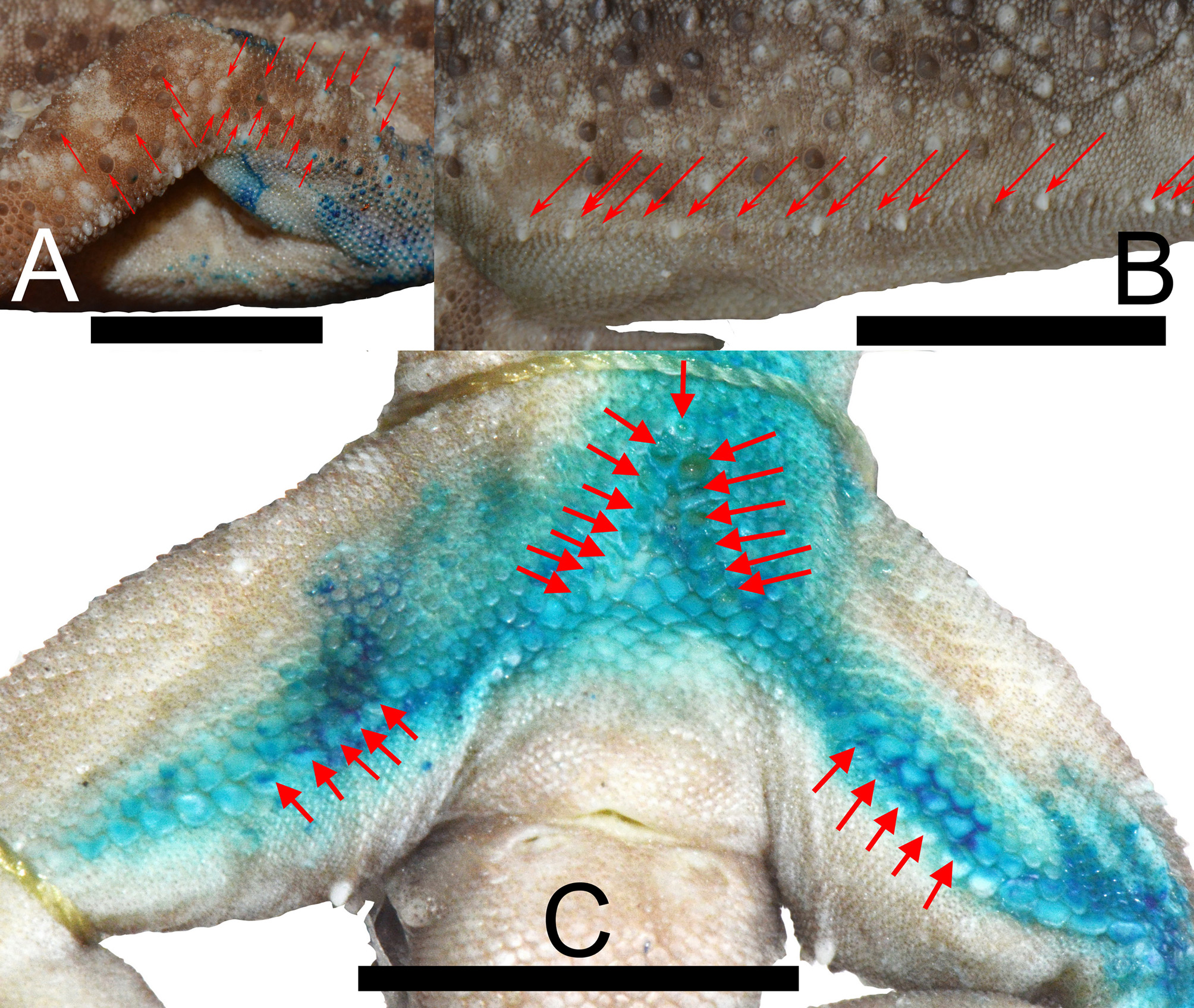
Forelimbs relatively short, ArmL 15.5% of SVL; dorsal scales on forelimbs slightly larger than those of body, weakly keeled; dorsal surface of antebrachium with keeled tubercles, dorsal surface of brachium with few tubercles, close to elbow very few, but multiple close to the axilla ( Fig. 5A View FIGURE 5 ); palmar scales flat, smooth, subimbricate; digits well-developed, inflected at basal interphalangeal joints, digits slightly narrower distal to inflection; subdigital scales transversely expanded along the entire length of each digit, but slightly compressed in both length and width immediately distal to interphalangeal inflection; 12 LamF1, 17 LamF2, 17 LamF3, 19 LamF4, and 16 LamF5; claws well developed, sheathed by two dorsal scales and one ventral scale.
Hindlimbs longer than forelimbs, TibL 19.2% of SVL; hindlimbs covered dorsally by granular scales interspersed with larger, conical tubercles; EPS and EFS present, discontinues, interrupted by five smaller scales on each side; 14 PP arranged in ˄ - shape; precloacal depression in shape of a pit; EFS arranged in two rows, 13 right and 11 left side of EFS; 5 FP on both side, separated from PP by eight poreless scales on right and seven poreless scales on left side, respectively ( Fig. 5C View FIGURE 5 ); scales on ventral surface of thigh bordering EFS and InfraS from below very small; ventral scales on tibia smooth, flat, subimbricate; plantar scales slightly raised; digits well-developed, inflected at basal metapodial-phalangeal joints, digits slightly narrower distal to inflection; subdigital scales transversely expanded along the entire length of each digit, but slightly compressed in both length and width immediately distal to interphalangeal inflection; 14 LamT1, 17 LamT2, 18 LamT3, 20 LamT4, and 19 LamT5; claws well-developed, sheathed by two dorsal scales and one ventral scale.
Body elongate, AGL 38.9% of SVL; ventrolateral body folds with strongly raised conical tubercles, which are various from not separated to separate each other until by three granules ( Fig. 5B View FIGURE 5 ); dorsal scales small, granular, interspersed with large, conical, semi-regularly arranged, keeled tubercles; 19 DorsT; 28 PVT. Ventral scales larger than dorsals, smooth, flat, imbricate, 50 VentS between ventrolateral body folds.
Tail regenerated, TailL 77.1 mm with original part 7.2 mm and regenerated part 69.9 mm in length, subcylindrical, somewhat flattened; dorsal and lateral side of tail with keeled tubercles, which extend along original part of tail only; distribution of tubercles at base of tail similar to dorsum, tubercles on tail not extending to ventral side; subcaudals under original tail small, irregular, no enlarged transverse plates; on the regenerated tail scales form transversely enlarged subcaudal plates; tip of tail pointed; three postcloacal tubercles on right and four on left side.
Coloration in life. Dorsal ground color of head, body and tail light brownish grey. Nuchal region light brown with a dark brown V-shaped blotch, which is bordered by a yellowish white line; three darkish brown transverse blotches between axilla and groin. Some tubercles on sides are yellowish white. A darkish brown longitudinal line is running from posterior orbit to above of the ear and joins dorsolaterally with the first of the darkish brown transverse blotches. The regenerated tail is dorsally light brown, scattered with small yellowish white dots distally. The yellowish color faded in preservative. We missed to record the ventral color in life; it is grey in preserved vouchers. Iris grey to dark during the day with brick-red pupil ( Fig. 2A–C View FIGURE2 ).
Variation. The paratypes resemble the holotype in coloration and there seems to be no sexual dimorphism in coloration. Both paratypes with nine, wide dark brown bands (including tail tip) separated by narrower light greyish to off-white bands. Male have pores and a precloacal depression, whereas female do not. Paratype MZB.Lace.5133 is an unsexed juvenile, in which pores and a precloacal depression are absent. Detailed variation of mensural and meristic characters are presented in Table 1.
Etymology. The species epithet tahuna is a noun used in apposition. Tahuna is the capital town of Kabupaten Sangihe, where the specimens were collected.
Distribution. Cyrtodactylus tahuna sp. nov. is currently only known from its type locality, Tahuna, Kepulauan Sangihe Regency, Sulawesi Utara Province, Sangihe Island, Indonesia ( Fig. 4 View FIGURE 4 ). Although the species range is probably not restricted to Tahuna, it may exhibit a relatively limited distribution in Sangihe Island and should be regarded as endemic to the region, until evidence to the contrary becomes available.
Natural history. Specimen MZB.Lace.5123 (AK028), the holotype, was found on a steep concrete wall of a bridge crossing a small river at around 08:30 pm after a long rainfall. The anthropogenically influenced and mountainous surrounding near Tahuna was characterized by a street and a house in a densely forested area. In direct vicinity specimen MZB.Lace.5133 (AK029) was found at the base of a tree that night. Two days later, specimen MZB.Lace.5097 (AK032) was collected at a bridge across the small river around 08:00 pm. At this occasion, the sky was clear, but humidity was still high. The accompanying nocturnal herpetofauna comprised of several specimens of Hylarana celebensis ( Peters, 1872) . An artificial water pool close by was inhabited by unidentified tadpoles. During daytime Limnonectes sp., Eutropis multifasciata ( Kuhl, 1820) , Lamprolepis smaragdina (Lesson, 1829), Sphenomorphus sp., Varanus salvator ( Laurenti, 1768) , and Ahaetulla prasina ( Boie, 1827) were encountered on Sangihe Island ( Koch 2012).
Comparisons. The new species can be readily distinguished from all recognized congeners occurring on Sulawesi, the Moluccas, and the Lesser Sunda Islands (detailed diagnostic characters in Table 2) except Cyrtodactylus fumosus , C. halmahericus ( Mertens, 1929) , C. papuensis Brongersma, 1934 and C. tambora Riyanto, Mulayadi, McGuire, Kusrini, Febylasmia, Basyir & Kaiser, 2017 , by the following combination of characters: the presence of precloacal pores, femoral pores, enlarged precloacal and femoral scales that are disconnected, and the lack of enlarged median subcaudal scales in original tails. In the following comparisons, the characters for C. tahuna are provided in parentheses. Cyrtodactylus fumosus (data from Mecke et al. 2016a, b) can be easily distinguished from C. tahuna by the absence of tubercles on ventrolateral folds (versus tubercles on ventrolateral skin fold present), the presence of 4–7 unkeeled DorsT (versus 19 keeled DorsT), the presence of only, this is indicated either by ̒Ƌ̕ or ̒ ♀ ̓), 6 = enlarged femoral scales (the number of pores is given in parentheses and includes primordia; if pores are present in one sex only,
is indicated either by ̒Ƌ̕ or ̒ ♀ ̓), 7 = enlarged precloacofemoral scales (the number of pores is given in parentheses and includes primordia; if pores are present in males only, this
indicated by ̒Ƌ̕), 8 = precloacal and femoral pores in a continuous series, 9 = expression of precloacal depression (N = no depression, G = groove, P = pit; if a depression is present
males only, this is indicated by ̒Ƌ̕), 10 = pattern of dorsum (bd = banded; bl = blotched; mo = mottled; pl = patternless; st = striped), 11 = data derived from literature sources are abbreviated by letters as follows: A = this publication, B = Riyanto et al. (2017), C = Mecke et al. (2016a), D = Mecke et al. (2016b), E = Riyanto et al. (2016), D = Rösler & Kaiser 2016), E = Hartmann et al. (2016), F = Iskandar et al. (2011), G = Oliver et al. (2009), H = Hayden et al. (2008), I = Linkem et al. (2008) and J = Rösler et al. (2007). If data for a character are not available, this is indicated by a question mark (?).
EPFS in both sexes (versus EFS and EPS), and the presence of a precloacal groove in males (versus males with a pit-like precloacal depression). Cyrtodactylus halmahericus (data obtained from specimens listed in appendix) can be distinguished from C. tahuna by the presence of EPFS in both sexes (versus EFS and EPS in both sexes), the presence of a continuous row of PFP in males (versus PP and FP in males), and the presence of a precloacal groove in males (versus males with a pit-like precloacal depression). Cyrtodactylus papuensis (data from Rösler et al. 2007) differs from C. tahuna by possessing a deep groove-like precloacal depression (versus pit-like). Cyrtodactylus tambora (data from Riyanto et al. 2017) can be distinguished from C. tahuna by smaller body size with an adult SVL of 39.4–47.4 mm (versus 78.5–79.2 mm), the presence of EPFS in both sexes (versus EFS and EPS in both sexes), the absence of tubercles on the dorsal surface of brachium (versus tubercles on brachium present), and the absence of FP in both sexes (versus FP present in males).
Cyrtodactylus tahuna sp. nov. differs from the following species from central Indonesia by possessing tubercles on the dorsal surface of brachium, with tubercles being absent in C. celatus Kathriner, Bauer, O’Shea, Sanchez & Kaiser, 2014 , C. fumosus , C. gordongekkoi (Das, 1994) , C. hitchi Riyanto, Kurniati & Engilis, 2016 , C. laevigatus Darevsky, 1964 , C. tambora , and C. wetariensis ( Dunn, 1927) . In having tubercles on ventrolateral body folds, the new species differs from Cyrtodactylus celatus , C. fumosus , C. laevigatus , and C. tambora . By having enlarged posterior precloacal scales, Cyrtodactylus tahuna sp. nov. can be distinguished from Cyrtodactylus batik Iskandar, Rachmansah & Umilaela, 2011 , C. hitchi , C. nuaulu Oliver, Edgar, Mumpuni, Iskandar & Lilley, 2009 , C. spinosus Linkem, McGuire, Hayden, Setiadi, Bickford & Brown, 2008 , and C. wallacei Hayden, Brown, Gillespie, Setiadi, Linkem, Iskandar, Umilaela, Bickford, Riyanto, Mumpuni & McGuire, 2008 .
In addition, the new species possesses EFS, which distinguishes it from Cyrtodactylus batik , C. celatus , C. hitchi , C. jellesmae (Boulenger, 1897) , C. laevigatus , C. spinosus and C. wallacei .
Finally, C. tahuna sp. nov. lacks enlarged transversely subcaudal scales in original tails as opposed to the presence of transversely enlarged, median subcaudal scales as seen in C. batik , C. darmandvillei (Weber, 1890) , C. deveti ( Brongersma, 1948) , C. hitchi , and C. wallacei .


| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cyrtodactylus tahuna
| Riyanto, Awal, Arida, Evy & Koch, André 2018 |
Cyrtodactylus tahuna
| Riyanto & Arida & Koch 2018 |
Cyrtodactylus tahuna
| Riyanto & Arida & Koch 2018 |
C. tambora
| Riyanto, Mulayadi, McGuire, Kusrini, Febylasmia, Basyir & Kaiser 2017 |
C. tambora
| Riyanto, Mulayadi, McGuire, Kusrini, Febylasmia, Basyir & Kaiser 2017 |
C. hitchi
| Riyanto, Kurniati & Engilis 2016 |
C. hitchi
| Riyanto, Kurniati & Engilis 2016 |
C. hitchi
| Riyanto, Kurniati & Engilis 2016 |
C. celatus Kathriner, Bauer, O’Shea, Sanchez & Kaiser, 2014
| Kathriner, Bauer, O'Shea, Sanchez & Kaiser 2014 |
Cyrtodactylus celatus
| Kathriner, Bauer, O'Shea, Sanchez & Kaiser 2014 |
C. celatus
| Kathriner, Bauer, O'Shea, Sanchez & Kaiser 2014 |
Cyrtodactylus batik
| Iskandar, Rachmansah & Umilaela 2011 |
Cyrtodactylus batik
| Iskandar, Rachmansah & Umilaela 2011 |
C. nuaulu
| Oliver, Edgar, Mumpuni, Iskandar & Lilley 2009 |
C. spinosus
| Linkem, McGuire, Hayden, Setiadi, Bickford & Brown 2008 |
C. wallacei
| Hayden, Brown, Gillespie, Setiadi, Linkem, Iskandar, Umilaela, Bickford, Riyanto, Mumpuni & McGuire 2008 |
C. spinosus
| Linkem, McGuire, Hayden, Setiadi, Bickford & Brown 2008 |
C. wallacei
| Hayden, Brown, Gillespie, Setiadi, Linkem, Iskandar, Umilaela, Bickford, Riyanto, Mumpuni & McGuire 2008 |
C. gordongekkoi
| Das 1994 |
C. laevigatus
| Darevsky 1964 |
C. laevigatus
| Darevsky 1964 |
C. laevigatus
| Darevsky 1964 |
C. wetariensis (
| Dunn 1927 |
C. jellesmae
| Boulenger 1897 |