
Caspia makarovi ( Golikov and Starobogatov, 1966 )
|
publication ID |
https://doi.org/ 10.1080/00222933.2012.742934 |
|
persistent identifier |
https://treatment.plazi.org/id/5D4987D3-CF45-FFCD-FEB7-FF5BFEA1F6F9 |
|
treatment provided by |
Carolina |
|
scientific name |
Caspia makarovi ( Golikov and Starobogatov, 1966 ) |
| status |
|
Caspia makarovi ( Golikov and Starobogatov, 1966) View in CoL
( Figures 2 View Figure 2 A–E, 3E)
Caspia gmelini W. Dybowski, 1887 View in CoL – Makarov 1938: 1058, figure only [not Clessin and W. Dybowski in W. Dybowski, 1887]
Pyrgula (Caspia) makarovi Golikov and Starobogatov, 1966: 353 View in CoL , fig. 1, 5;
Pyrgula (Caspia) gmelini aluschtensis Golikov and Starobogatov, 1966: 354 View in CoL , fig. 1, 8;
Caspia gmelini aluschtensis ( Golikov and Starobogatov, 1966) – Golikov and Starobogatov 1972: 100, pl. 2, fig. 19;
Caspia (Clathrocaspia) makarovi makarovi ( Golikov and Starobogatov, 1966) View in CoL – Alexenko and Starobogatov 1987: 36;
Caspia (Clathrocaspia) makarovi ( Golikov and Starobogatov, 1966) View in CoL – Anistratenko 2007b: 796–797, fig. 2, 20.
The teleoconch is ovate, thick-walled, minute. The shell consists of 4.0–4.5 moderately inflated and regularly increasing whorls separated by an incised suture; it is about 2.2–2.4 mm in height and up to 1.3 mm in width. The apical angle is about 40–45 ◦, spire profile is almost straight. The whorls of the teleoconch are ornamented with strongly expressed spiral ribs (up to 20 on body whorl), crossed by axial ribs, creating a reticulate sculpture ( Figure 2 View Figure 2 A–C). Body whorl moderately expanded, occupying about 0.75 of total shell height. Aperture medium-sized, oval, with a thickened peristome. Height of shell exceeds 2.5 times the height of aperture. The umbilical chink is narrow or completely covered by the columellar margin.
The protoconch ( Figure 2 View Figure 2 C–E) is dome-like, and comparatively large; it consists of 1.25 whorls and has a maximum diameter 0.28–0.30 mm; diameter of the first embryonic whorl is 0.25–0.26 mm. The initial cap-like onset of the embryonic shell is 0.11–0.12 mm wide. The ornamentation of the embryonic shell is the same as in C. knipowitschii , consisting of irregular honeycomb-like pits covering the entire surface; some fine spiral rows of grains can be recognized on it in some specimens ( Figure 2 View Figure 2 ,E,F). The transition from the protoconch to the teleoconch is quite sharp and is marked by a thickening, followed by a constriction. The onset of the teleoconch is indicated by the formation of axial and spiral striae, which grade into the adult sculptural pattern.
In fact the fully grown shells of C. makarovi and C. knipowitchii are similar in size, general shape and ornamentation patterns (see Figures 1 View Figure 1 , 2 View Figure 2 ). They differ in the dimensions of the protoconch and in the proportions and degree of inflation of the whorls. The shell of C. makarovi is more slender and has slightly more inflated whorls than C. knipowitschii : the apical angle in C. makarovi is 40–45 ◦ compared with 45–50 ◦ in C. knipowitschii .
Analysis of the SEM images shows a uniform protoconch sculpture in both species ( C. makarovi and C. knipowitchii ) ( Figure 1 View Figure 1 , 2 View Figure 2 ). At the same time the protoconch measurements can help to differentiate between these species. The parameters summarized in Table 1, show that the protoconch of C. makarovi is slightly smaller than that of C. knipowitchii . Compared with Caspiohydrobia johanseni Frolova, 1984 , the protoconch shape and its ornamentation are similar in the Caspia species, though some details are specific: the surface of the embryonic shell of Caspiohydrobia johanseni is covered by a more delicate and less dense net (see Figure 4C View Figure 4 ). At the same time the teleoconch characters enable the two genera to be clearly distinguished since the adult shell of Caspiohydrobia (like most hydrobiid species) lacks any ornamentation, except growth lines.
Radula morphology
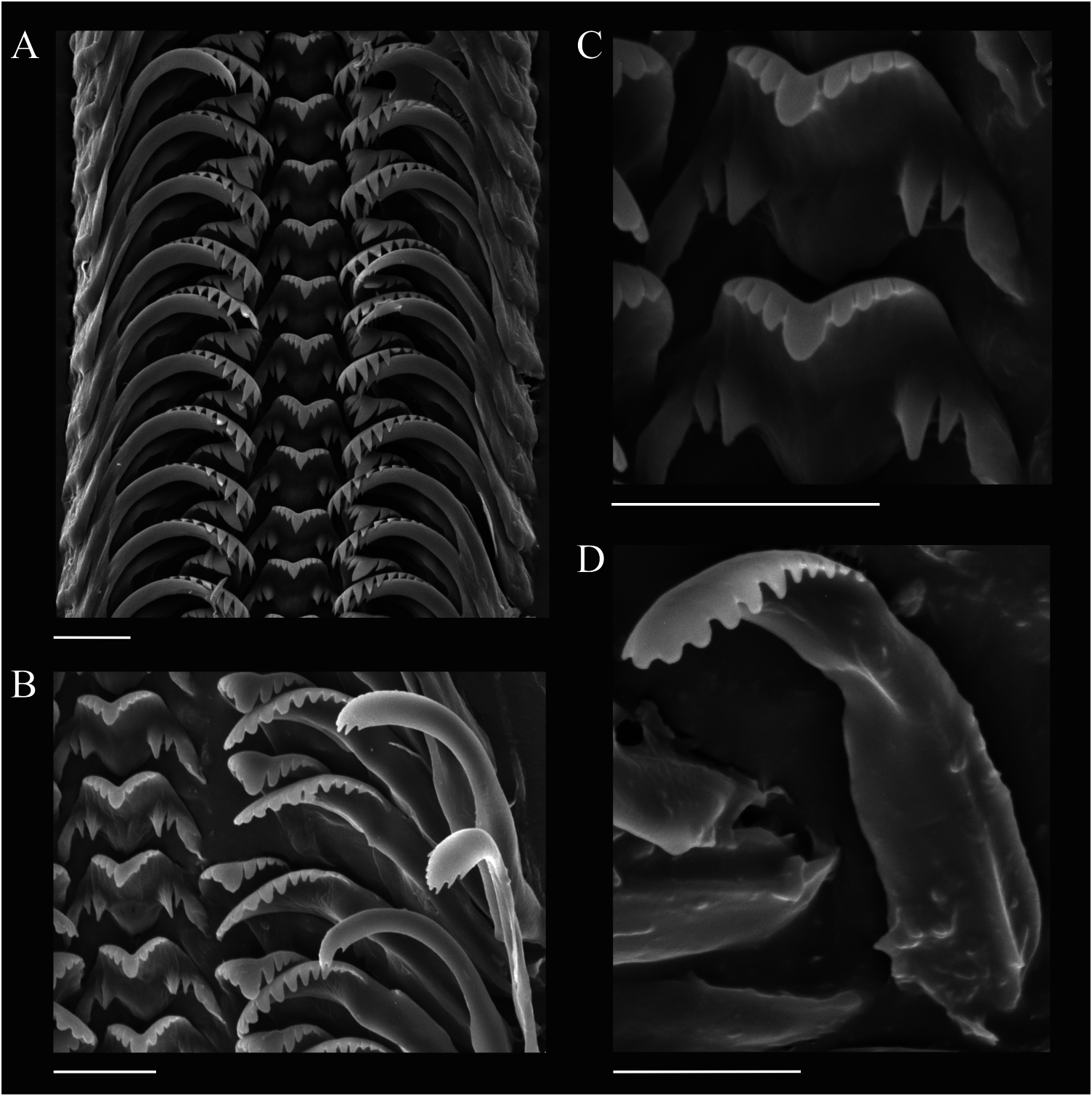
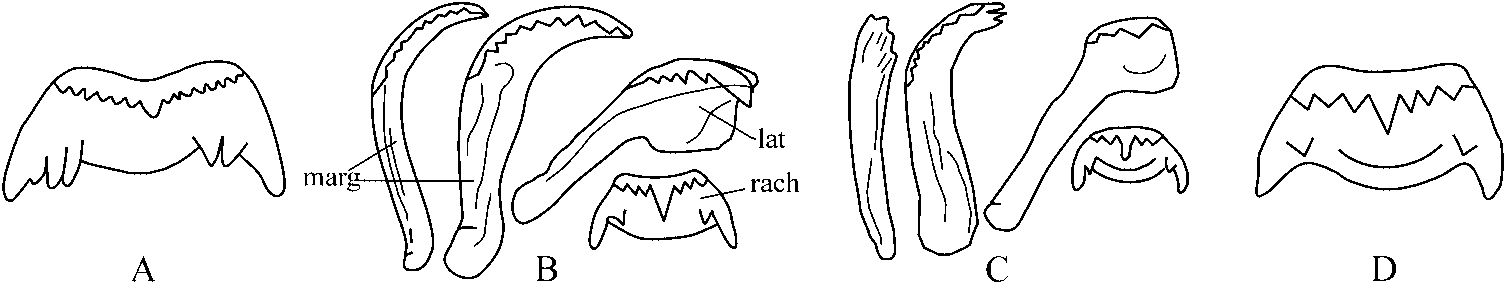
The radula of C. knipowitchii is of the taenioglossate type, about 0.06–0.07 mm wide and 0.23–0.25 mm long, bearing around 32 rows of teeth ( Figure 5A View Figure 5 ). The general shape of the rachidian tooth is trapeziform with tapered latero-basal corners. The maximum width of the rachidian tooth is about 0.015 mm, the height does not exceed 0.008 mm. On the cutting edge of the rachidian (central) tooth there are four lateral cusps on each side of the median denticle (4-1-4: Figure 5B,C View Figure 5 ), which is much larger than the lateral cusps and triangular in shape. The basal plate of the rachidian tooth bears two pairs of well-developed sub-basal cusps (compare with Sitnikova and Starobogatov 1998).
The lateral teeth have the formula 3-1-4 ( Figure 5B View Figure 5 ), they have a broad and massive plate with broad cutting edge bearing cusps, the biggest of them is moderately prominent. The inner marginal tooth bears 10–11 prominent, moderately long knife-shaped cusps. The cutting edges of the outer marginal teeth are multicusped and terminally rounded; both the inner and outer marginal teeth are attached by a narrow base and have a length of 0.03–0.04 mm ( Figure 5 View Figure 5 B–D).
The radula of C. knipowitchii from the Azov–Black Sea Basin (Lower Dnieper) studied here is almost identical to that of Caspia gmelinii Clessin and W. Dybowski in W. Dybowski, 1887 from the Caspian Sea ( Sitnikova and Starobogatov 1998) (see Figure 6A View Figure 6 ). A difference appears to be the larger number (up to six) of lateral cusps on each side of the median denticle on the cutting edge of the rachidian tooth of C. gmelinii , as drawn by the authors ( Sitnikova and Starobogatov 1998: 1361, fig. 3, 12). However, their text mentions four or five lateral cusps and their SEM image shows exactly four cusps as in C. knipowitchii ( Sitnikova and Starobogatov 1998: 1363, fig. 5, 4). Another difference appears to be that, according to the authors, on the base of the rachidian tooth of C. gmelinii there are two (as is here shown for C. knipowitchii ), or rarely, three pairs of sub-basal cusps ( Sitnikova and Starobogatov 1998: 1363). Most likely the data quoted demonstrate a range of variability of the rachidian tooth, as the authors studied more specimens than I did. Anyway, the radulae of both Caspia species discussed here show extremely close resemblance to each other in most of their quantitative as well as qualitative features.
The comparison of the radulae of Caspia and those of Caspiohydrobia studied by Sitnikova et al. (1992) shows their remarkable similarity not only in composition and general view of radulae but also in many tiny details. The main difference is that the basal plate of the rachidian tooth of all Caspia bears two pairs of sub-basal cusps, whereas only a single pair is present in Caspiohydrobia ( Figure 6 View Figure 6 B–D). This supports the separate generic rank of the two genera.
The radulae of Pyrgula annulata (Linnaeus, 1758) from Lake Garda in Italy (type species of Pyrgula Cristofori and Jan, 1832 ), Dianella thiesseana (Kobelt, 1878) from Greece as well as Turricaspia spp. from the Caspian Sea and Euxinipyrgula spp. from the Azov–Black Sea Basin are characterized by a simple rachidian tooth with no basal cusps ( Radoman 1983; Sitnikova and Starobogatov 1998, 1999; Riedel et al. 2001; Szarowska et al. 2005). The absence of basal cusps on the rachidian tooth can be considered a diagnostic feature of all “true” pyrgulid taxa listed here in contrast to the remaining Ponto-Caspian hydrobiids (namely Caspia and Caspiohydrobia ). Further, the median denticle on the cutting edge of the rachidian tooth of Turricaspia and Euxinipyrgula is usually smaller than in Caspia and Caspiohydrobia ( Figure 6 View Figure 6 ).
A combination of these features and the presence of a pair of small lateral protrusions on the rachidian tooth of Pyrgula , Dianella and Falsipyrgula (the latter from south-western Turkish lakes) could be considered as evidence for independent evolutionary changes in this branch of hydrobioid gastropods (see Anistratenko 2008). With regard to the radular morphology the representatives of Caspia and Caspiohydrobia appear to be more closely related to Hydrobia than to Pyrgula . These observations corroborate the viewpoint of Riedel et al. (2001) that Caspiinae B. Dybowski, 1913 (sensu Sitnikova and Starobogatov 1998) might represent a lineage independent from “true” pyrgulids (see taxonomic implications).
Taxonomic implications
According to Starobogatov (1970) the family Pyrgulidae Brusina, 1882 (= Micromelaniidae B. Dybowski and Grochmalicki, 1913 of subsequent authors) included all the taxa of Ponto-Caspian hydrobioids inhabiting the brackish waters of the Caspian Sea, the entire Azov–Black Sea Basin and some Balkan lakes. Many authors mention that classification of the biogeographical group discussed here (i.e. all Ponto-Caspian hydrobioids) remains uncertain (e.g. Radoman 1983; Sitnikova and Starobogatov 1998, 1999; Riedel et al. 2001). The range and taxonomic rank (families or subfamilies) of hydrobiid and pyrgulid gastropods is still not widely agreed upon. Some systematists consider them as clearly distinct families ( Thiele 1929; Radoman 1983; Alexenko and Starobogatov 1987; Ponder and Warén 1988; Alexenko and Anistratenko 1998; Riedel et al. 2001), while others regard pyrgulids as a subfamily within Hydrobiidae ( Kabat and Hershler 1993; Wilke et al. 2001, 2007; Bouchet and Rocroi 2005).
Based on conchological traits the genera Caspia and Caspiohydrobia were originally attributed to the family Pyrgulidae ( Starobogatov 1970) . Later, the reproductive system and radula of some Caspiohydrobia species were described and illustrated ( Sitnikova et al. 1992). Anatomical investigations suggested that Caspiohydrobia species are systematically closer to Hydrobiidae s. str. than to Pyrgulidae . It was shown that Caspiohydrobia differs strongly from all other Ponto-Caspian pyrgulids by having a single pair of basal cusps on the rachidian tooth of radula and male and female reproductive organs very similar to those of Hydrobia ( Sitnikova et al. 1992: fig.1, 9, 10, 17–21), and it was therefore transferred to the family Hydrobiidae .
The protoconch examinations of Caspia and Caspiohydrobia reveal a similar kind of pitted ornamentation, though the protoconchs of Caspia bear more complicated and specific sculpture. This type of ornamentation occurs with variations in most Hydrobiidae s. lat. and is here interpreted as a plesiomorphic character. The radula morphology of Caspia and Caspiohydrobia reveals that these two genera are distinguished from all other pyrgulids sensu Starobogatov ( Starobogatov 1970) by the presence of basal denticles on the rachidian tooth.
Based on anatomical (female reproductive system) and radular characteristics, Sitnikova and Starobogatov (1998) have already suggested that the genus Caspia should be placed in a separate subfamily Caspiinae within Pyrgulidae . Caspia and Caspiohydrobia differ from other “pyrgulid” taxa (namely, Turricaspia , Euxinipyrgula and Falsipyrgula ) by the presence of basal denticles on the rachidian tooth and should therefore be assigned to the family Hydrobiidae . Caspiohydrobia is closely related to, and possibly synonymous with, Ecrobia Stimpson, 1865 ( Bank et al. 2001; Filippov and Riedel 2009; Haase et al. 2010) and should be placed in Hydrobiinae . Caspia is sufficiently different to be placed in a separate subfamily Caspiinae , which should be transferred from the family Pyrgulidae to the family Hydrobiidae .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Caspia makarovi ( Golikov and Starobogatov, 1966 )
| Anistratenko, Vitaliy V. 2013 |
Caspia (Clathrocaspia) makarovi ( Golikov and Starobogatov, 1966 )
| Anistratenko VV 2007: 796 |
Caspia (Clathrocaspia) makarovi makarovi ( Golikov and Starobogatov, 1966 )
| Alexenko TL & Starobogatov YI 1987: 36 |
Caspia gmelini aluschtensis ( Golikov and Starobogatov, 1966 )
| Golikov AN & Starobogatov YI 1972: 100 |
Pyrgula (Caspia) makarovi
| Golikov AN & Starobogatov YI 1966: 353 |
Pyrgula (Caspia) gmelini aluschtensis
| Golikov AN & Starobogatov YI 1966: 354 |
Caspia gmelini W. Dybowski, 1887
| Makarov AC 1938: 1058 |