
Microthalestris Sars, 1905
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5051.1.13 |
|
publication LSID |
lsid:zoobank.org:pub:F94203E7-FCD1-4975-BAD3-0DF534806712 |
|
DOI |
https://doi.org/10.5281/zenodo.5572440 |
|
persistent identifier |
https://treatment.plazi.org/id/951887EA-FFD7-FFBC-FF51-D13AE09BFBFD |
|
treatment provided by |
Plazi |
|
scientific name |
Microthalestris Sars, 1905 |
| status |
|
Various authors have reported on what they individually regard as Parastenhelia spinosa or varieties/species sharing some of its morphological characteristics, but recognise a wide range of variation in features such as segmentation of the female antennule, modification of the caudal ramus setae (particularly V), setation of the antennary exopod and endopod, relative proportions of P1 rami and form of their terminal elements, armature formulae of P2–P4, shape and armature of ♀ P5 exopod, and segmentation of ♂ P5 exopod. The magnitude of this variability, or the identity of differences which might represent true genetic discontinuity (and thus distinct species), is in many cases impossible to determine due to the inaccurate and/or incomplete descriptions and illustrations [especially the original description by Fischer (1860)] which have accompanied many published reports. The currently accepted concept of P. spinosa being a highly variable, cosmopolitan taxon has grown over time since 1860 by the gradual accumulation of differences observed in what were wrongly assumed to be geographically isolated populations of the same species. The historical review presented below aims at (a) pointing out some important misconceptions in the taxonomy of the species, (b) assigning distinct specific status to some of its radically divergent “populations”, and (c) formulating a working hypothesis and baseline for future comparisons and revisionary work .
Parastenhelia spinosa was first described as Harpacticus spinosus by Fischer [1860: 665–666, Table XXI (figures 51–53), XXII (figure 66)] from the island of Madeira [ Lang (1948: 589) stated incorrectly that Fischer did not disclose the type locality] but the description is severely lacking in any real detail (8-segmented antennule, ovate female P5 with four spines on exopod and baseoendopod, caudal ramus seta V swollen at base) and only his illustration of the P1 provides the barest minimum to identify it as a member of the Parastenheliidae .The inaccuracies expressed in the few poorly rendered drawings (maxilliped, P1, female P5, caudal rami) and the lack of the male make the description virtually useless for comparative purposes. Harpacticus spinosus is placed in Microthalestris solely on account of its elongate P1 exp-2 but cannot be treated as anything more than a species inquirenda in this genus. Had Lang’s (1948) fixation of H. spinosus as the type of Parastenhelia been valid, this would have threatened stability and universality in the application of the name of the type species since its taxonomic identity cannot be determined from its name-bearing type. Although Lang (1936a: 21) considered the possibility that Fischer’s (1860) species was conspecific with Parastenhelia forficula var. littoralis (sensu Sars 1911) he recommended to set it aside on the grounds that it cannot be identified with any degree of confidence. Unfortunately, he changed his opinion by not only reinstating it as a valid species but also by illegitimately fixing it as the type of the genus ( Lang 1944: 13; 1948: 586–588).
Claus (1863: 131–132, Plate XVII, figs 7–11) described Thalestris forficula Claus, 1863 as one of ten species assigned to his new genus Thalestris Claus, 1862 (type species by subsequent designation: Thalestris longimana Claus, 1863 ). The author reluctantly included H. spinosus in the genus as well as two other Harpacticus species previously described by Fischer (1860). His description of T. forficula , based on material from Messina (Sicily), included illustrations of the antennule of both sexes, P1, caudal rami and the female genital field.The most significant information that can be extracted from Claus’s (1863) concise description is the 8-segmented condition of the female antennule, the presence of an apophysis on the male P3 endopod, the modified caudal seta (V) displaying a basal swelling, and the length of the relatively short inner seta on P1 enp-1 which extends only to about 40% of the segment length. No information was given about the armature of P2–P 5 in both sexes, or the segmentation of the male P5 exopod.
......continued on the next page
17 Based on Lang’s (1936a: Fig. 37) supplementary description (as Pa. gracilis ).
18 Pallares (1982: 10) erroneously stated that P3–P4 exp-3 display a [333] pattern in the male; probably not having consulted Pallares’s drawings of the male, Mielke (1990: 166) accepted this as an additional difference between the type population and his material collected from Bahía Lapataia near Ushuaia ( Argentina).
19 Armature pattern of enp-3 021 in ♂.
20 0.1.12 + apo in ♂.
21 The single ♀ observed by Mielke (1974: 20) showed an aberrant setal pattern on one side of P2 enp-2 (with two inner setae) and P4 exp-3 (with inner seta).
22 There is confusion over the exact number of elements on the exopod. Pallares (1963) states that there are three outer, one inner and two apical elements but in a subsequent account ( Pallares 1968: 65) explicitly states that there are seven without going into detail about their position. We have adopted Pallares’s original observation pending reexamination of new material.
23 Lang’s (1936b: 24) only female displayed the atypical 121 pattern on enp-3.
24 Mielke (1990) recorded a high degree of variability in the armature of P2–P4. Some specimens displayed (a) an inner seta on P2–P3 exp-2, (b) only one (instead of two) inner seta on P3 exp-3, (c) two (instead of one) inner setae on P4 exp-3, (d) two (instead of one) inner setae on P3–P4 enp-3, and/or (e) only two (instead of three) outer spines on P4 exp-3. One male lacked the inner seta on P4 enp-2.
25 According to Wells (2007: 596) the original report of three inner setae on P2 exp-3 is wrong.
26 The endopodal armature pattern differs from Mielke’s (1994a: 257) in two aspects; the apical spiniform element on enp-3 is here considered as the homologue of the outer spine in other genera, and the two minute and fine setae originating from the inner corner of enp-3 are included in the formulae.
27 Inner seta minuscule ( Mielke 1994a: Fig. 4C View FIGURE 4 ).
28 2-segmented in ♂ (1.321).
29 1.1. 321 in ♂; Kunz (1975) did not illustrate the ♀ condition but listed the armature formula of P3 endopod as 1.1. 221 in his Table 5; Gee (2006) re-examined the type material and noted that the armature formula is as in F. anglica .
30 2-segmented in ♂ (1.121).
31 1.1.021 in ♂.
32 Willey (1935) reported some specimens with a 3-segmented endopod in the ♀ (1.1.221).
33 P3 endopod ♂ and P 5 ♂ based on Vervoort’s (1964) description of Pa. spinosa [partim]. According to Sewell (1940: Text-fig. 27H) P2 exp-1 lacks an inner seta but Vervoort shows a small inner element which is included in the armature formula here.
34 Scott (1894a: Plate 12, Fig. 39) most likely overlooked the short inner seta on exp-3.
35 Sewell (1940: 196) claims the proximal inner seta was overlooked in Scott’s (1894a) description, bringing the total to five.
36 1.221 in ♂.
In a paper written in old Danish and completely lacking in illustrations, Boeck (1865: 266–267) described a new species, Thalestris karmensis Boeck, 1865 , based on specimens collected from Karmøy in Norway; this species fell into oblivion until it was discussed by Sars (1905 – see below).
In a similar contribution, Boeck (1873: 56) provided a short unillustrated text description of Dactylopus longipes Boeck, 1873 , obtained from 29 m depth in the Oslofjord in Norway. His female specimens were characterized by a 9-segmented antennule, a long and slender P1 exopod which is shorter than enp-1, a 2-segmented P1 endopod with a very long enp-1 which bears a seta in the proximal quarter of the inner margin, and a P5 exopod with eight elements. Based on this short description Lang (1936a: 31) would later synonymize D. longipes with Thalestris forficula which he had previously placed in Parastenhelia ( Lang 1934: 24) .
Thomson’s (1883: 104–105; Plate X, Figs 16–21) material of Thalestris forficula from Otago Harbour in New Zealand agrees with the description of Claus (1863) in the 8-segmented condition of the female antennule and the basally inflated setae V on the caudal ramus but differs in size (1,000 vs 800 μm) and the morphology of P1. The female P5 bears six and five setae on the exopod and endopodal lobe, respectively. Lang (1934: 25) lists Thomson’s record but does not appear to question its identity. In later accounts he accepts it as a record of either Parastenhelia forficula var. littoralis ( Lang 1936a: 52) or P. spinosa ( Lang 1948: 588) .
Car (1884: 248–249, Plate XVIII, Figs 1 View FIGURE 1 –8) provided an illustrated description of a new species, Thalestris pectinimana Car, 1884 , based on a single ovigerous female collected in the vicinity of Trieste, northeastern Italy. The author did not compare T. pectinimana with other congeners nor did he present a justification for its recognition as a distinct species. Illustrations were provided for the female in lateral aspect, antennule, antenna, maxilliped, distal segments of the P1 rami and P5. The species is characterized by the 8-segmented antennule, the large P5 with seven setae on the foliaceous exopod and five elements on the endopodal lobe, and the presence of distinct pectinate spines on the terminal segments of P1 exopod and endopod. No information was given on the armature of P2–P4. The species has not been recorded again since its original description. Pesta (1920: 590) listed it as a junior subjective synonym of Phyllothalestris mysis ( Claus, 1863) but this was rejected by Lang (1936a: 49) who considered it a valid species of Thalestris . Lang (1948: 497, 588) revised his previous opinion and considered T. pectinimana conspecific with Parastenhelia spinosa , more specifically (based on P5 morphology) with the form described as Microthalestris littoralis f. penicillata by Willey (1935: 82). This claim is dubious since Willey did not describe the female P5 but only stated that it displays “… the fifth foot of littoralis ”. According to Sars’s (1911: Suppl. Plate 11) description of M. littoralis the female P5 has only six setae on the exopod while T. pectinimana exhibits seven.
Scott (1894a: 100–101, Plate XII, Figures 33–41) recorded a single ovigerous female of T. forficula from a shore gathering in Accra ( Ghana). The specimen is similar to Claus’s (1863) material in the 8-segmented antennule and the basally swollen caudal ramus setae (V), however, is distinctly smaller (500 vs 800 μm). The most conspicuous feature is the presence of two strong pectinate claws on both P1 exp-3 and P1 enp-2. His illustration of P4 shows that it lacks the inner seta on exp-1 and has two inner setae on enp-3 (221). The female P5 has a reduced armature (if Scott’s observations are correct), displaying six setae on the exopod and only four on the endopodal lobe. The latter observation was disputed by Sewell (1940: 196) who believed that the proximal inner seta of the endopodal lobe was overlooked.
Scott & Scott (1894: 142–144, Plate IX, Figs 4 View FIGURE 4 –9) described Thalestris forficuloides from mud near the lowwater mark at Seafield in the vicinity of Leith, Firth of Forth, Scotland. An expanded description of the female and additional illustrations of the male were provided by Scott (1894b: 255–256, Plate X, Figures 13–25) based on specimens from the type locality (note that the mandible and maxillule were transposed in his Plate X). As the name suggests this species was considered close to T. forficula , displaying differences in the segmentation of the female antennule (9-segmented vs 8-segmented in T. forficula ), the length of the P1 endopod, and some unspecified discrepancies in the ”… proportional lengths of the other thoracic feet” ( Scott & Scott 1894: 144).
Sars (1905: 122) proposed a new genus Microthalestris in the Thalestridae to accommodate T. forficula as its type and only species and stated that the forms recorded by Boeck (1865) as Thalestris karmensis and by Scott & Scott (1894) as T. forficuloides , both belong to M. forficula ( Claus, 1863) . His redescription of the latter ( Sars 1905: 123–124, Plate LXXVI) corresponds exactly to the illustrations of Scott (1894b) in the segmentation of the antennule, morphology of P1, and the basally inflated caudal ramus seta (V), however, also shows a number of important differences with T. forficuloides including (a) P3 exp-1 with inner seta instead of without, (b) P3 exp-3 with two inner setae instead of three, (c) P5 exopod ♀ comparatively longer, and (d) P5 exopod ♂ 3-segmented instead of 1-segmented. Although Sars was not specific about the total number of setae on the male P5 exopod it appears from his illustration that there are only six as opposed to the seven recorded in later descriptions. There is a hint of a short setal element among the spinular cluster on the outer margin of exp-1, however, the seta usually found in that position is typically very long (e.g. Chislenko 1967: Fig. 45; Mielke 1974: Fig. 9D). Lang’s (1936b: 23, Fig. 52) observation of a similar condition in a male from the Øresund indicates that it is genuinely lost (or extremely reduced) rather than broken off during dissection. Although Lang (1934: 24) expressed doubts, primarily based on differences in antennulary segmentation, about the conspecificity of Sars’s (1905) Norwegian specimens of M. forficula and Claus’s (1863) type material of T. forficula , he cited the former as a synonym of the new combination Parastenhelia forficula ( Claus, 1863) .
Sars (1911: 369–370, Supplement plate 11-1) added the new species M. littoralis Sars, 1911 to the genus which showed more or less the same distribution in Norway as M. forficula , including various localities along the south and west coasts and further northwards to the Trondhjem Fjord (Bejan). Sars (1911) only reported the female which differs from that of the type species in the more compact antennule (the number of segments was not disclosed), the longer and more slender P1, the number of exopodal setae on the P5 (six vs eight) and the normally developed seta V on the caudal ramus. Unfortunately, no information on the armature formula of P2–P4 was given which inevitably led to the incorrect assignment of various non-conspecific populations to this species.
Farran (1913) observed considerable variation in body size (500–850 μm) in his material of M. littoralis from Clare Island and Blacksod Bay, Co. Mayo ( Ireland) (see also Farran et al. 1915; Southern 1915) which he viewed as a possible indication of the presence of a second species. All specimens agreed with Sars’s (1905) description in the morphology of the female P5.
Pesta (1920: 591–593) summarized records of Microthalestris in the Adriatic, including M. forficula in the Venice Lagoon ( Grandori 1912, 1914) and M. littoralis from the Palagruža (Pelagosa) archipelago ( Steuer 1912).
Brian (1921: 77–80) reported M. littoralis from the Gulf of Genoa, illustrated naupliar stages I and III–V (Text figures 23–25; Plate VI, Fig. 3 View FIGURE 3 ) and described copepodids III–V (Plate VIII, Figs 10–22). Dahms & Hicks (1996) pointed out that Brian’s naupliar illustrations in reality refer to NIV–NVI. Brian’s (1921) description reveals little information about the female except for the lateral aspect view of an ovigerous specimen (Plate III, Fig. 7), showing the presence of a single egg sac, and the ventral view of the abdomen (Plate V, Fig. 15) which shows that the caudal ramus seta V is not swollen at the base. His illustrations of the male (Plate IV, Fig. 9; Plate V, Fig. 5 View FIGURE 5 ; Plate IX, Figs 9–14) document three characters of particular significance, (a) the presence of two penicillate spines on the antenna (already expressed in copepodid IV: his Plate VIII, Fig. 17), (b) the 3-segmented P3 endopod in the male bearing an inner seta on enp-1 and -2, in addition to the apophysis and two apical setae on enp-3, and (c) the 1-segmented condition of the male P5 exopod bearing seven setae. No information was provided on the segmentation of the female antennule or armature of the swimming legs and P5.
Monard (1927) retained Microthalestris in the Thalestridae as an “insufficiently described genus” (p. 157) in his Synopsis universalis generum harpacticoidarum but placed the closely related Parastenhelia in the Diosaccidae (= Miraciidae ). He admitted that the latter genus may belong to the same family as Microthalestris on the basis of P1 morphology but was unable to make a firm recommendation for its exclusion from the Miraciidae because the number of egg sacs was apparently unknown [this being untrue since both Car (1884) and Brian (1921) had made cursory observations and illustrated a single egg sac].
Gurney (1927: 545) confirmed the presence of a single egg sac in a damaged female of an undescribed Parastenhelia species from Port Taufiq (Suez Port), located at the southern boundary of the Suez Canal in Egypt. He classified the genus in the Canthocamptidae .
Monard (1928: 348–349, Fig. XVIII–2) reported numerous specimens of M. forficula from Port-Vendres and Banyuls-sur-Mer on the French mediterranean coast and considered the species as extremely variable with M. littoralis definitely falling within its limits. Contrary to Sars (1905), Monard also appeared to support the concept that M. forficula assumes a cosmopolitan distribution by accepting doubtful records from New Zealand ( Thomson 1883) and the Gulf of Guinea ( Scott 1894a) as valid. The French material differs from the Norwegian specimens used in Sars’s (1905) description in a number of significant aspects such as (a) the segmentation of the female antennule (eight vs nine), (b) antennary endopod with two penicillate elements, (c) caudal ramus setae IV–V with an inflated base (this also occurs in Claus’s description of T. forficula ), (d) claws on both rami of P1 distinctly shorter, (e) P3 exp-3 with three inner setae vs two, (f) P4 enp-3 with two inner setae vs one, (g) P5 exopod ♀ with six elements vs eight, and (h) P5 exopod ♂ 1-segmented vs 3-segmented. Monard (1928) also reported variability in the armature of the female P5 exopod with some (rare) specimens displaying seven elements, and variation in body length (650–800 μm). Sewell (1940: 191) stated that Monard misidentified his specimens and was in reality dealing with M. littoralis sensu Sars (1911) but there is no justification for this claim.
Stephensen (1929) recorded M. littoralis from two localities in the Faroe Islands, i.e. off Trangisvaag (Suðuroy = Suderø), and between Streymoy (Strømø) and Eysturoy (Østerø). Lang (1948) re-examined Stephensen’s material and confirmed the identity of the specimens from the latter site but confirmed that the male from Trangisvaag belongs to Amonardia normani ( Brady, 1872) (Thalestridae) .
Wilson (1932a) added the first North American record of M. littoralis from the plankton in Chesapeake Bay, Maryland, stating that he found both sexes. His record was not accompanied by any illustrations but the author seems to indicate that he accepts Brian’s (1921) identification of the male of this species as correct. Wilson (1932b) subsequently recorded both M. forficula (Penzance Pond) and M. littoralis (Cuttyhunk Island between Buzzards Bay and Vineyard Sound) from the Woods Hole region, Massachusetts. The report of the first species included illustrations of P1, P5 of both sexes and male P3 endopod, all of which agree well with Sars’s (1905) description except for the latter which shows an inner seta on the distal segment (only P. bulbosa shares this character; Table 1 View TABLE ), and the male P5 which displays a long outer seta on the proximal segment. No information was given about the segmentation of the female antennule, swimming leg armature or the form of caudal ramus seta V. The North American specimens are distinctly smaller (♀♀: 400–500 μm; ♂♂: 350 μm) than their European counterparts. The authenticity of Wilson’s (1932b) material of M. littoralis is indeterminable since the few illustrations provided are reproduced from Sars (1911). His text reveals that females (no males were found) correspond in the setation of the female and measure 500–600 μm. Interestingly, Wilson (1932b: 572) maintained Parastenhelia and Microthalestris as separate genera.
Jakubisiak (1933: 115) recorded M. littoralis from the Isle of Šolta along the Dalmatian coast of Croatia and opposed Monard’s (1928) view that this species must be subsumed under M. forficula , stating that it not only differs in the morphology of the female P5 but also in body size and the segmentation of the antennule.
Lang (1934) formally synonymized Microthalestris with Parastenhelia and transferred the latter to the Thalestridae based primarily on the similar position of the inner seta on P1 enp-2 and the reported presence of a single egg sac in members of the genus (cfr. Gurney 1927: 545). Morphological comparisons between single species of these genera were also used as additional evidence in support of their synonymy, e.g. similarities were pointed out in the morphology of ♀ P5 ( P. hornelli and M. littoralis ), ♂ P5 ( P. hornelli and M. forficula ), P1 ( P. antarctica and M. forficula / littoralis ) and caudal ramus seta V ( P. similis and M. forficula ). Lang (1934: 25) relegated M. littoralis to a mere variety of Parastenhelia forficula ( Claus, 1863) , dismissing the most clearcut character that consistently separates these species (number of exopodal setae on ♀ P5) along most of the Norwegian coastline ( Sars 1911) as of no significance. This course of action was based on the incongruous argument that differences had been reported between the widely disjunct “populations” of T. forficula (and M. littoralis ) from New Zealand ( Thomson 1883), Gulf of Guinea ( Scott 1894a) and Ireland ( Farran 1913). The first two sources contain incomplete and partly inaccurate descriptions (based on a single specimen in Scott’s case) while the third is completely lacking in illustrations and by Farran’s own account is probably based on an amalgam of two species. Lang’s (1934) paper set the scene for his authoritarian stance in shaping a very broad, encompassing taxonomic concept of Parastenhelia spinosa , with the result that it has unjustly come to be regarded as a highly variable, cosmopolitan species. He further fuelled the controversy by assigning a pair in amplexus from Perseverance Harbour on Campbell Island in the southern Pacific to P. forficula var. littoralis . Differences in P1, P5 and caudal rami indicate that his subantarctic specimens belong to a different, as yet unnamed species (see below). Lang (1934) listed T. karmensis and T. forficuloides as synonyms of the typical P. forficula .
Monard (1935a: 43) recorded both M. forficula and M. littoralis from the Roscoff region (Brittany). The former displayed a 3-segmented P5 exopod in the male while females of the latter were excessively large (900 μm) compared to the Norwegian populations (590 μm; cfr. Sars 1905). He continued to place Parastenhelia in the Miraciidae while maintaining Microthalestris and his newly established genus Thalestrella in the Thalestridae . During the same year, Monard (1935b: 47), adopting a different nomenclature, stated that he had found both the “typical” form and the littoralis variety of Parastenhelia forficula in Cartaghe (Salammbô), Tunisia and admitted having difficulties separating them. He reported variability in the number of segments in the female antennule (one specimen having seven!) and occasional left-right asymmetry in the setation of the female P5 exopod in some individuals. Without illustrations it is impossible to decide on the identity of his material.
Willey (1935: 82–83) distinguished two forms in his material of M. littoralis from Harrington Sound in Bermuda. One ovigerous female (500 μm) conformed to Sars’s (1911) description in displaying 9-segmented antennules and unmodified caudal ramus setae. The endopods of P2–P4 of this littoralis -form are distinctly 3-segmented, have a constricted mobile articulation between enp-2 and enp-3 and the inner distal corner of enp-2 produced into a sharp spur beyond the insertion of the inner seta (his Fig. 121); the formula of P4 enp-3 is 121. Furthermore, the proximal inner seta of P4 exp-3 is longer than the second inner seta, which is comparatively short and only slightly thickened. All other specimens (♀♀: 500–700 μm; ♂♂: 400–500 μm) had 8-segmented antennules and forcipately curved caudal setae IV–V with swollen bases. Willey (1935: Figs 119–120, 122–126) assigned them to a separate penicillata -form, based on the presence of two penicillate spines amongst the apical armature elements of the antenna similar to those figured by Brian (1921) for M. littoralis from the Gulf of Genoa. In addition, both claws at the distal margin of P1 enp-2 are coarsely penicillate, and P1 exp-3 bears a slender inner seta, a geniculate seta, and two penicillate spines. In the penicillata -form P2–P3 enp-2 and -3 are more often indistinctly separated, without a functional articulation between them. Specimens with distinctly 3-segmented endopods of P2–P3 lack a spur at the inner distal corner of enp-2. The armature formula of P4 enp-3 is sexually dimorphic, being 221 in the female, and 121 in the male. On P3–P4 exp-3, the distal inner seta is excessively fine and curving outwards, and the proximal inner seta of P4 exp-3 is distinctly shorter than the middle one. Females of both forms share the typical P5 setation pattern of M. littoralis (six elements on the exopod, five on the endopodal lobe). Males of the penicillata -form have a 3-segmented P3 endopod with two apical setae and a spinous apophysis on enp-3, and a 1-segmented P5 exopod with seven elements.
Lang (1936b) recorded both P. forficula (one ♀, one ♂) and P. forficula var. littoralis (one ♀) from the Øresund between Zealand ( Denmark) and Scania ( Sweden) with all three specimens having been collected from different localities. His P. forficula specimens agree with Sars’s (1911) description in the general facies of the P1 and P5 of both sexes but differs in the presence of only one inner seta on P3 enp-3 (instead of two) in the female; however, it must be noted that the latter observation was based on a single female and not accompanied by an illustration. Lang (1936b) assigned, except for his own record from Campbell Island ( Lang 1934), several older records to P. forficula var. littoralis without giving any explanation. These include the reports of Microthalestris littoralis from Norway ( Sars 1911), western Ireland ( Farran 1913), the Gulf of Genoa ( Brian 1917, 1921), Palagruža in Croatia ( Steuer 1912), Chesapeake Bay ( Wilson 1932a) and Cuttyhunk Island, Massachusetts ( Wilson 1932b), of Microthalestris forficula from Banyuls-sur-Mer ( Monard 1928), and of Thalestris forficula from New Zealand ( Thomson 1883) and Ghana ( Scott 1894a).
Lang (1936a) proposed a new subfamily Parastenheliinae within the Thalestridae and fixed Parastenhelia as its type and only genus. Contrary to his earlier opinion ( Lang 1934: 24) he accepted the conspecificity of Claus’s (1863) T. forficula and Sars’s (1905) M. forficula , and assigned the previous records of T. forficula from Lille-Karajak fjord, western Greenland ( Vanhöffen 1897) and of M. forficula from Woods Hole ( Wilson 1932b) to Parastenhelia forficula . He maintained Sars’s (1911) M. littoralis as a variety of P. forficula , made Dactylopus longipes a synonym of this species, but surprisingly made no mention of Fischer’s (1860) Harpacticus spinosus . The two females of P. gracilis he reported from the Falkand Islands are here assigned to P. antarctica (see below).
Monard (1937: 59, 62, Fig. 3–E View FIGURE 3 ) collected material from Algiers and Bou Ismaïl (formerly Castiglione) in Algeria and assigned it to P. forficula littoralis . He figured the male P5 exopod as 1-segmented bearing a total of seven elements and based on his previous observations ( Monard 1928, 1935a) prudently postulated that this character may be dimorphic among males.
In his review of the genus Parastenhelia Sewell (1940: 189–196)) reassessed the great degree of reported variability in antennule segmentation, armature of P2–P4, segmentation of male P5 exopod, and shape of terminal caudal ramus setae. He forcefully opposed the views of Lang (1934, 1936a, 1936b) and Monard (1935b, 1937) for uncritically accepting the synonymy of T. forficula , T. forficuloides and M. littoralis , claiming that these authors and others before them had been dealing with a mixture of species. Sewell (1940) reinstated the latter two as valid species of Parastenhelia and maintained Willey’s (1935) penicillate variety as a distinct form of P. littoralis . His material from seaweed washings in Addu Atoll, the southernmost atoll of the Maldives, revealed three different forms which he attributed to P. littoralis , P. littoralis f. penicillata , and a new form, P. littoralis f. scotti. Sewell’s P. littoralis material (his Text-Fig. 26) is characterized by the following features: (a) body length 440–560 μm (♀), unknown (♂); (b) antennule ♀ 9-segmented; (c) antennary endopod with two penicillate spines; (d) P1 enp-1 very long, about 1.45 times length of exopod; enp-2 with two penicillate claws; exp-3 with three penicillate elements and one long seta; (e) P3 endopod ♂ 3-segmented, with armature formula [1.1.02 + apo]; (f) P4 exp-1 without inner seta and exp-3 with seven elements; endopod with inner seta on exp-1 and -2, and four elements on enp-3; (g) P5 exopod ♀ elongate, about 3.7 times as long as maximum width, with six setae; endopodal lobe with five setae; (h) P5 exopod ♂ 1-segmented, with seven setae; and (i) caudal ramus seta V unmodified or only slightly swollen at the base. Despite the absence of information about female antennulary segmentation, antennary morphology and swimming leg armature in M. littoralis , it is still obvious that Sewell’s (1940) specimens do not belong to this species; this is best illustrated by the morphology of the P1 and the length:width ratio of the ♀ P5 exopod. The absence of the inner seta on P1 exp-2 is most likely based on an observational error. It is puzzling why Sewell listed T. forficula sensu Thomson (1883) and M. forficula sensu Monard (1928) as misidentified records of P. littoralis since both sets of specimens are significantly divergent from Sars’s (1905) description.
Sewell’s (1940: Text-Fig. 27) second form, based on females only and assigned to P. littoralis f. penicillata , is characterized by the following features: (a) body length 600 μm (♀), unknown (♂); (b) antennule ♀ 8-segmented; (c) antennary endopod with two penicillate spines; (d) P1 enp-1 long, about 1.15 times length of exopod; enp-2 with two penicillate claws; exp-3 with two pinnate claws and two (?) setae; (e) P2–P3 exp-3 with six and seven setae/ spines, respectively, P2 exp-1 without inner seta; (f) P2–P4 enp-3 with four, five and five elements, respectively; (g) P5 exopod ♀ elongate, about three times as long as maximum width, with six setae; endopodal lobe with five setae; and (h) caudal ramus setae IV–V markedly swollen at the base (occasionally less pronounced in seta IV). According to Sewell (1940) there is no doubt that his material is identical to Willey’s (1935) specimens from Bermuda but a comprehensive comparison is hampered by the lack of information on (a) relative segment lengths of P1, armature of P2, and setation and length:width ratio of the female P5 exopod in the Bermudian material, and (b) armature of P4 exopod, sexual dimorphism in P4 enp-3 setal pattern, and both segmentation and armature of the male P5 exopod in the Maldivian specimens. It is unclear whether the absence of the thin, outwardly curved, distal inner seta on P3 (and possibly P4) exp- 3 in Sewell’s (1940: Text-Fig. 27–I) specimens is genuine or due to an observational error; a minute notch along the distal protion of the inner margin appears to hint at its presence.
The final form, based on a single female (Text-Fig. 28), was described as a new variety, forma scotti, of P. littoralis . It is characterized by the following features: (a) body length 560 μm; (b) antennule 8-segmented; (c) antennary endopod without penicillate spines; (d) P1 enp-1 very long, about 1.33 times length of exopod and twice as long as exp-2; enp-2 with two pinnate claws; exp-3 with three claws and one seta; (e) P5 exopod large (reaching to middle of third abdominal somite), elongate-oval, about three times as long as maximum width (measured in proximal 1/3), with six setae all inserting in distal one-third; endopodal lobe with five setae; and (f) caudal ramus very broad (about 4.5 times as wide as long), seta V forming pyriform swelling at the base. Sewell (1940) considered this form identical with Scott’s (1894a) ovigerous female of T. forficula from Ghana, primarily on account of the general shape of the P5 exopod and the modification of caudal seta V. Although both females are strikingly similar in these two characters and Sewell dismissed the absence of the proximal inner seta on the P5 endopodal lobe in Scott’s specimen as an observational error, others demonstrate that they are clearly different. Such features are particularly related to the P1 (relative lengths of rami, armature of exp-3) and P5 (length:width ratio, size relative to urosome, position and length of setae). A comparison of the swimming legs is impossible since only the P4 was figured by Scott (1894a: Plate XII, Fig. 39) and no information was given by Sewell (1940) other than that they “… appear to resemble those of P. littoralis ”.
Lang (1944: 13) invalidly fixed Harpacticus spinosus as the type species of Parastenhelia .
Nicholls (1945: 3–4, Text fig. 2A–B) illustrated two forms of P. forficula from the reef fringing Leander Point at Port Denison near Dongarra in Western Australia. Both forms differ considerably in the shape of caudal ramus setae IV–V, P1 and female P5. Adopting the view of both Monard (1928) and Lang (1934, 1936a, 1936b) he considered M. littoralis a variety of P. forficula . Three females (720 μm), which were assigned to the “typical” form of P. forficula , display (a) distincty basally swollen caudal setae IV–V, (b) a P 1 in which exp-2 and enp-1 are extremely elongate, being much longer than in Sars’s (1905) material from Norway, and (c) a P5 which is ovate in shape and shows the 6-element arrangement typical of P. littoralis instead of the eight setae/spines in typical P. forficula . Nicholls (1945: 4) himself regarded the identification of his material as provisional, suggesting it may represent a new species. The second form, described as P. forficula var. littoralis (♀♀: 630–670 μm.; ♂: 540 μm), exhibits unmodified caudal setae IV–V. Females differ slightly from Sars’s (1911) description of M. littoralis in (a) the proportionally longer P1 exp-2 and enp-1, and (b) the P5 endopodal lobe which is narrower and less truncate. Nicholls (1945) claimed that the male P5 (1-segmented with seven elements) did not conform with the descriptions of Sars and Lang but this statement cannot be substantiated since Sars (1911) did not describe the male and Lang’s (1934: Fig. 56) illustration of the P5 refers to a different species (see below).
Having previously claimed that Harpacticus spinosus was indeterminable ( Lang 1936a: 21), Lang (1948: 311) stated that “Fischer’s illustrations of this species clearly show that it is the same species that Claus described as Thalestris forficula and which Lang (1934) subsequently placed in the genus Parastenhelia . That I make this identification without any reservation, is based on the fact that there is no other harpacticoid whose P1 and P5 are built in the way Fischer depicts” [our translation]. As a result of Lang’s groundless reinstatement of H. spinosus as a valid species it not only became the senior synonym of a whole string of species and forms previously considered as conspecific with M. forficula and M. littoralis but was also unjustly designated as the type species ( P. spinosa ) of Parastenhelia . Lang (1948) assembled arguments in favour of his very broad concept of P. spinosa , including all the species, forms and varieties that in the course of time had been described and he considered conspecific – mostly on dubious grounds. These include Thalestris forficula , T. forficuloides , T. karmensis , T. pectinimana , Dactylopus longipes , Microthalestris littoralis and M. littoralis f. penicillata . With regard to the latter Lang (1948: 586) stated emphatically that comparison with Fischer’s (1860) description of Harpacticus spinosus shows beyond doubt that both forms are identical. Since Fischer only illustrated, and quite inadequately, the maxilliped, P1, female P5 and the caudal ramus, it is inexplicable how Lang could have arrived at this conclusion because none of these appendages (except for the caudal ramus) was figured or described by Willey (1935). Given that Fischer’s (1860) illustrations do not inspire confidence in their accuracy it would not be extraordinary to state that there is no tangible evidence for synonymizing any of the other species/forms mentioned above with H. spinosus . Lang (1948: 586) suggests in his review that M. forficula , M. littoralis and M. littoralis f. penicillata should be regarded as three forms (forma typica , forma littoralis and forma penicillata , respectively) of P. spinosa , which however are not geographically or ecologically distinct. Separation of these forms was based on the shape and/or modification of caudal ramus setae IV–V, number of segments in the female antennule, presence/absence of penicillate spines on the antennary endopod, shape and ornamention of the distal claws on P1 enp-2, number of setae on P3–P4 exp-3, P4 enp-3 (in both sexes where known) and P5 exopod ♀, and segmentation of P5 exopod ♂. Rather than exploring the possibility that P. spinosa may represent a species complex Lang claimed that it is impossible to keep the three forms distinct because a number of intermediate forms are known to exist and the combination of characters varies in every conceivable manner. Lang (1948: 587) underpinned his claim by shoehorning into P. spinosa every form and variety from geographically disparate locations such as Campbell Island ( Lang 1934), the Woods Hole region ( Wilson 1932b), Gulf of Guinea ( Scott 1894a), Brittany ( Monard 1935a), the Banyuls area ( Monard 1928) and the Gullmar Fjord in Sweden ( Lang 1948), irrespective of the morphological variation reported. Since Lang failed to recognize sibling species boundaries, which negatively affects studies on biodiversity, speciation and ecology, his attempt to present a unifying, deliberately broad, taxonomic concept of P. spinosa should be seen as an exercise in futility. Unfortunately, most subsequent workers have uncritically adopted this concept and even attempted to expand it (e.g. Vervoort 1964) rather than subjecting it to scrutiny [ Gee (2006) and Kuru & Karaytuğ (2014) are notable exceptions]. The species is now widely viewed as assuming an allegedly cosmopolitan distribution, covering all continents and including records from the Arctic ( Vanhöffen 1897; Scott 1899; Scott & Scott 1901; Sars 1909; Brotskaya 1962; Mielke 1974; Chislenko 1967, 1977; Chertoprud et al. 2005; Kornev & Chertoprud 2008; Dvoretsky & Dvoretsky 2010) and remote islands such as Tristan da Cunha ( Wiborg 1964) and Moce Island, Fiji ( Wells 1978).
Lang’s (1948) view was not generally accepted by some Irish workers who continued to separate “populations” of P. spinosa into forms or even species. Williams (1954) recorded both Microthalestris forficula and M. littoralis from pools at Ardglass (the latter also from similar habitats at Kilclief) in Co. Down, Northen Ireland. Roe (1958) reported both P. spinosa var. littoralis and P. spinosa var. penicillata in intertidal pools around Dalkey Island and The Muglins (lighthouse), Co. Dublin, Ireland. Both forms were subsequently obtained from littoral stone and seaweed washings and net sweepings in Lough Hyne (Ine), West Cork, Ireland ( Roe 1960).
Noodt (1955b: 204) suggested that Sewell’s (1940) Parastenhelia littoralis f. scotti probably falls within the boundaries of Lang’s concept of P. spinosa .
Božić (1955: 5, 11, Plate 4, Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ) described P. spinosa from Kernic, west of Roscoff, Brittany and illustrated the female caudal ramus, antennary exopod and mandibular palp. Females display a 9-segmented antennule and a P5 with six setae on the exopod and five on the endopodal lobe. Males have a 3-segmented P5 exopod bearing seven elements. Božić (1955) reported sexual dimorphism in the shape of caudal ramus seta IV, being distinctly swollen and bulb-shaped in the female and unmodified in the male. In both sexes the inner terminal seta V was more or less normal at the base. The material was accorded distinct specific rank by Gee (2006 – see below).
Şerban (1959) reported P. spinosa from Agigea along the Romanian Black Sea coast, stating that his specimens approached Willey’s (1935) penicillata form because of the 1-segmented exopod in the male P5, a condition he attributed to neotenic development rather than variability. It is not clear whether his material displayed the penicillate spines on the antenna and P1 endopod. Pór (1960) subsequently recorded P. littoralis from Eforie and Agigea and advocated that P. spinosa (which he regarded as conspecific with Microthalestris forficula ) and P. littoralis are separate species.
Pesta (1959: 128–129; Figs 66–69) collected both sexes of P. spinosa from algal washings at Sorrento in the Bay of Naples, Italy. Illustrations were provided for the P1, the female P5 and abdomen, and the male P3. Pesta accepted Lang’s (1948) concept of P. spinosa , being a variable species containing three ill-defined and overlapping forms, and stated that the females in his material displayed the P5 armature of f. littoralis and the basally swollen caudal ramus seta V of f. typica , and that one of the males resembled f. penicillata in the structure of the P3 endopod. The body size of the male was erroneously given as 4.4 mm (it is more likely 440 μm). Among all the records that have been attributed to P. spinosa , Pesta’s is probably the one that is most similar to Claus’s (1863) original from Messina. Apart from being the one that is geographically closest to the type locality in Sicily, the Napolitanean female agrees with the original description in the relative proportions of the P1 rami, the degree of modification of caudal ramus seta V and the ovate shape of the P5. Except for the male P3 Pesta (1959) does not provide any information about the armature of the remaining swimming legs or the segmentation and setation of the male P5.
In his first paper on the harpacticoids from the Isles of Scilly Wells (1961) reported finding populations of P. spinosa containing a mixture of f. typica , f. littoralis and many females which were identical with Božić’s (1955) unnamed form characterized by the bulbous base of caudal ramus seta IV. The species was found on all four principal islands (St Mary’s, Tresco, St Martin’s and St Agnes). Wells (1963a: 14) subsequently found the bulbous form in clean sandy substrates at two sites in the estuary of the River Exe and formally proposed to name it P. spinosa forma bulbosa . A second form, P. spinosa forma littoralis was also recorded from one of the stations in the estuary. Gee (2006: 2634–2643, Figs 14–19) re-examined Wells’s (1963a) forma bulbosa based on new material from the Isles of Scilly and accorded it full specific rank (as P. bulbosa ). Both Bodin & Boucher (1983) and Bodin (1988) had previously recorded the taxon from Nord-Finistère (Brittany) as P. spinosa bulbosa . Since neither satisfied the criteria of availability (ICZN Arts 11–18) they cannot claim authorship for the subspecific name (ICZN Art. 45.5.1). Wells’s (1970: 261) record of P. spinosa from Port Hellick on St Mary’s was also confirmed to be conspecific with P. bulbosa ( Gee 2006) .
Vervoort (1964: 182) found that most of his Ifaluk material, although showing great variability in body length, agreed with Lang’s (1948) forma typica of P. spinosa (formerly M. forficula ). No illustrations were given but Vervoort stated that the females were characterized by a 9-segmented antennule, normally developed spines on the antennary endopod, P1 exp-2 (Vervoort erroneously referred to as the “2 nd endopodal segment”) 4–5 times as long as wide with an inner seta near the apex, P4 enp-3 with one inner seta, P5 exopod with eight elements, and a slightly basally swollen caudal ramus seta V. Males displayed a 3-segmented P5 exopod bearing six setae. One female and one male of a different form (from his locality 592), which he claimed would formerly have been assigned to Willey’s (1935) forma penicillata , were described in considerable detail ( Vervoort 1964: 182–184, Figs 64c, 66c, 67, 68d, 70a–g). The female (520 μm) exhibits a fairly long 8-segmented antennule, two penicillate spines on the antennary endopod, P1 exopod with two penicillate spines on exp-3 (note that Vervoort had misinterpreted this ramus as the endopod), basally swollen caudal ramus setae IV–V, and a P5 exopod with six elements. No information was provided about the female armature of P2–P4 or P5 endopodal lobe but Vervoort did illustrate the exopods of P2–P3 and the endopod of P3 of the male (420 μm). His material is assigned distinct species status below.
Bodin (1964: 128, Plate III, Fig. 15) recorded specimens of P. spinosa f. penicillata in high abundance in the Marseille region, southern France. Females display eight segments in the antennule but the proportional segment lengths are different from those reported by Monard (1928) and Sewell (1940). The only illustration given is that of the male P3 endopod which differs from Willey’s (1935) account in the absence of the inner seta on enp-1.
Chislenko (1967: 140–144, Figs 45–46) provided illustrations of both sexes and an aberrant specimen of P. spinosa from the White Sea. His material (♀: 550–800 μm, ♂: 400–420 μm) shows some similarities with Sars’s (1905) illustrations of M. forficula (e.g. 9-segmented antennule and eight elements on P5 exopod ♀) but differs in the length:width ratio of the female P5. The P1 has a similar exopod:endopod length ratio but the rami are significantly longer in the White Sea specimens. The males among Chislenko’s material display the full complement of seven elements on the P5 exopod while the outer seta is absent on the proximal exopodal segment in the Scandinavian populations of M. forficula ( Sars 1905; Lang 1936b). Considerable variability was observed in the degree of the basal swelling of caudal ramus seta V, including left-right asymmetry in the same specimen. In one female individual the P5 exopod on one side only had seven instead of the typical eight setae but the taxonomic significance of this aberration was dismissed. Chislenko (1967) drew particular attention to the structure of the female genital field, pointing out that the condition in the White Sea material differed significantly from that of Lang’s (1934: Fig. 51) illustration of the structure in his forma littoralis but was more similar to Claus’s (1863: Plate XVII, Fig. 11) approximate representation of that in the typica form. He expressed a sceptical attitude towards the recognition of forms or varieties in P. spinosa , claiming instead that a proper comparative analysis of the female genital field of these infrasubspecific taxa would confirm their specific status.
Apostolov (1968: 398, Plate 2, Figs 1 View FIGURE 1 –10) reported both sexes of P. spinosa from a sandy beach in Tsarevo (formerly Michurin), Bulgaria, provided a partial description, and assigned them to a new form, f. bulgarica. The Black Sea specimens are characterized by unmodified caudal ramus setae IV–V, the absence of the inner seta on P2 exp-1 and enp-1, and the presence of seven setae on the ♀ P5 exopod. The male displays a 1-segmented P5 exopod with seven elements and a bisetose endopodal lobe which is distinctly larger than in any other reported population. Apostolov (1968) illustrated sexual dimorphism in the setation of the antennary exopod but did not comment on it in the text. In a later paper Apostolov (1973: 270, 272–273, Fig. 8) recorded additional specimens from Tsarevo and supplemented his original descriptions of the female with illustrations of the genital field, P1, P2, P4, P5 and caudal rami. The latter two differ in some aspects (e.g. P5 exopod with only six setae) from his 1968 drawings, rendering future morphological comparison difficult.
Mielke (1974: 20–22, Fig. 9) reported on Parastenhelia specimens he collected in the intertidal zone at Longyearbyen in Svalbard. He assigned the material to P. spinosa , stating that it could not be attributed to any of the forms recognized so far. Illustrations were presented of the antennary exopod and P1 of the female, and the P3 endopod and P5 of the male. The female (590 μm) is characterized by a 9-segmented antennule, normally developed caudal ramus setae, and eight and five setae on the exopod and endopodal lobe of the P5, respectively. Males (420– 510 μm) possess a characteristic, acutely recurved, spinous apophysis on the P3 endopod. Some aberrations were recorded in the armature of P2 enp-2 (one or two inner setae) and P4 exp-1 (with or without inner seta).
Marinov (1974: 80–82, Figs 1–2 View FIGURE 1 View FIGURE 2 ) provided a description of P. spinosa specimens collected from algal washings at Sozopol on the southern Bulgarian Black Sea coast. Females display an 8-segmented antennule, six and five elements on the P5 exopod and endopodal lobe, respectively, and what appear to be unmodified caudal ramus setae IV–V. The distal endopod segment of P1 bears two pinnate (or penicillate) claws while enp-1 is excessively long, measuring 2.2 times the length of exp-2. The inner seta on P4 enp-1 is exceptionally long, extending beyond the distal tip of the endopod. The male shows the typical modification on the P3 endopod [1.1.02 + apo] and a 1-segmented P5 exopod with seven elements. Marinov (1974) reported some aberrant specimens that had a 1-segmented exopod in P3–P4, each carrying two apical elements, and observed a malformation in one male exopod which carried only six setae. Due to its dubious taxonomic status Apostolov’s (1968) forma bulgarica was dismissed from further consideration.
Chislenko (1977: 246, Fig. 6 View FIGURE 6 –1,2) collected two females (675–700 μm) of P. spinosa from washings of the demosponge, Semisuberites cribrosa ( Miklucho-Maclay, 1870) , in Franz Josef Land. He maintained that his specimens were morphologically similar to the material he described from the White Sea ( Chislenko 1967) which is confirmed by the two illustrations presenting the genital field and the P5 (with eight elements on the exopod).
According to Wells & Rao (1987) specimens (two ♀♀, one ♂) of P. spinosa from the Andaman Islands appear to be morphologically most similar to Sewell’s (1940) forma scotti, with eight segments in the female antennule, non-pectinate spines on the antennary endopod and a relatively short P1 exopod. The exopod of the male P5 is 3- segmented with six setae.
Gee (2006) stated that the Langian concept of P. spinosa as a very variable, cosmopolitan species is probably erroneous and a thorough reappraisal of this species is required. He convincingly demonstrated that P. spinosa var. bulbosa must be accorded distinct specific status as P. bulbosa and argued that Willey’s (1935) forma penicillata displays several characteristics warranting its elevation to species level.
Kornev & Chertoprud (2008: 196–197) provided an illustrated description of a White Sea population which they identified as P. spinosa . It appears that most of the text and at least some of the illustrations are a verbatim account of Chislenko’s (1967) description, including the substantial variation recorded in female body length. The authors do not specify whether the observed variability in P3–P4 armature is based on their White Sea material or on previous literature reports.
Kuru & Karaytuğ (2015) added a new species, P. aydini , from the eastern Mediterranean and Aegean coasts of Turkey and considered it morphologically closest to P. spinosa . The species is unique in the family by displaying a 2-segmented P5 exopod with a total of five setae in the male (a 2-segmented condition with different armature has been observed in Johnwellsia bipartita gen. et sp. nov. and Parastenhelia willemvervoorti sp. nov.). According to Kuru & Karaytuğ (2015), females of P. aydini differ from those of P. spinosa by the 9-segmented antennule, the reduction of the spinular rows around the outer margins of the bases of P1–P4, the presence of only four elements on P4 enp-3, ornamentation of the P5 baseoendopod and exopodal setae different, and the normally developed caudal ramus seta V. Males differ in the absence of a modified element on the seventh segment of the antennule (apparently present in P. spinosa ), the notched apophysis on P3 enp-3 and the presence of only four setae on P4 enp-3. It is not clear which material of P. spinosa the authors used as their reference for comparison.
Zaleha et al. (2018) provided a short text description of a single ovigerous female of Parastenhelia sp. from sediment in seagrass bed in Sungai Pulai, Johor, Malaysia . The antennule was described as 8-segmented and the 2- segmented P1 endopod as being much longer than the exopod. However, the armature of P2–P4 and the endopodal lobe of P5 is radically divergent from any other species in the genus, casting serious doubts on the validity of their identification. Zaleha et al. (2010) had previously identified Parastenhelia littoralis from rockpool seaweeds and sediment in Pulau Besar, Melaka.
Cordell (2018: 117–118, Fig. 5 View FIGURE 5 ) found several Parastenhelia specimens on debris, generated from the earthquake and tsunami that struck Japan on 11 March 2011, that landed in North America. The species was recorded on floating structures (docks, vessels, buoys) in several localities along the coasts of Washington and Oregon. While Cordell assigned the material to P. spinosa he acknowledged that they may be a different species within a larger species complex. Photographs were provided of the P1 and female urosome, showing that the caudal ramus seta V is somewhat swollen near the base.Although this character was observed in every female, Cordell (2018) mentioned that specimens he had previously examined from the northeast Pacific had almost no swelling at the base of this seta.
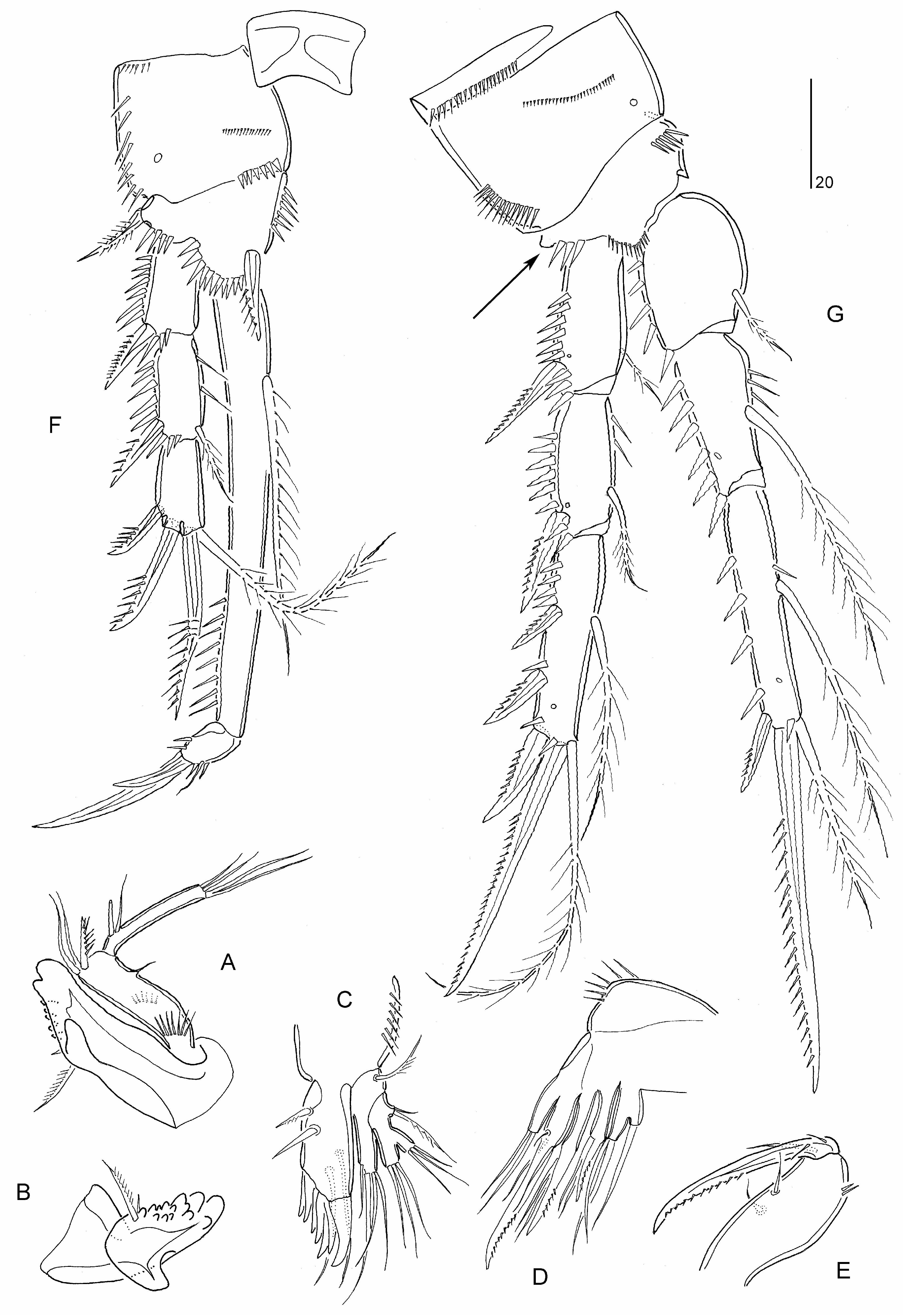
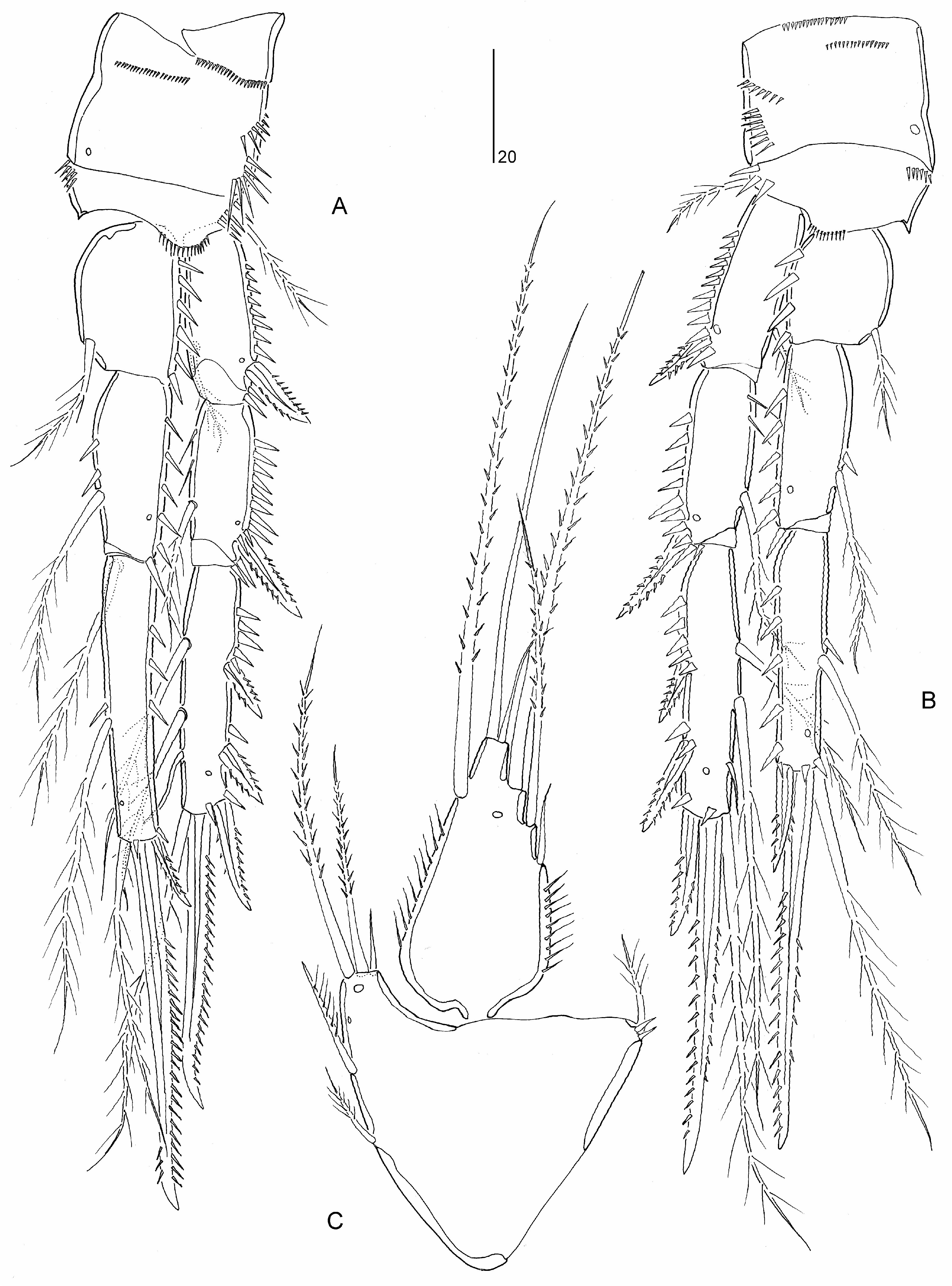
Diagnosis. Parastenheliidae . Sexual dimorphism in antennule, P3 endopod, P5–P6 and urosomal segmentation; occasionally endopods of P2 and P4, and caudal ramus setae IV– V. Body subcylindrical, short; posterior margin of urosomites (except anal somite) typically with denticulodigitate hyaline frills. Rostrum defined at base, of moderate size, reaching at most to distal margin of second antennulary segment; linguiform. Anal operculum semicircular, typically bordered with fine spinules. Caudal ramus wider than long, without spinular row on dorsal surface; with at least seven setae (seta I possibly absent); setae IV and/or V slightly swollen at base in ♀; seta IV ♀ with bulbous base in some species ( M. bulbosa comb. nov.) .
Antennule ♀ elongate and 9-segmented in ♀, segment 1 not elongate, segments 7–8 (when 9-segmented) shortest, with aesthetascs on segments 4 and 8 or 9. Antennule ♂ haplocer with four segments distal to geniculation but segmentation and segmental homologies unconfirmed for most species; in M. bulbosa comb. nov. segment 5 swollen and with aesthetasc, segmental homologies as follows: I, II– VIII, IX – XII, XIII, XIV – XVII, XVIII, XIX – XX, XXI – XXII, XXIII, XXIV – XXV, XXVI – XXVIII. Antenna not sexually dimorphic; proximal endopodal segment largely separated from basis or forming allobasis, with pinnate seta on abexopodal margin; exopod 2- segmented, proximal segment with 1–2 seta(e), distal segment with 1–2 lateral and three apical elements; distal endopodal segment without penicillate elements. Mandible with 3–4 elements on basis; endopod with eight setae; exopod 1-segmented with 2–5 setae. Maxillulary coxal epipodite represented by one seta. Maxilla with three endites on syncoxa; endopod discrete, with 2–3 setae. Maxilliped with 2–3 setae on syncoxa; basis with two setae on palmar margin; endopod represented by curved claw, accompanied by up to three accessory seta(e) .
P1 inner basal spine not sexually dimorphic. P1 exopod 3-segmented; exp-2 elongate, about 2–2.5 times length of exp-1 and between two-thirds and 80% length of endopod, with inner seta; exp-3 very small, typically with two pinnate spines, one geniculate seta or claw, and one non-geniculate seta. P1 endopod 2-segmented; enp-1 elongate, longer than exopod, with well developed inner seta originating either from near end of proximal quarter of segment or from middle third of segment, segment margins without area of reduced chitinization; enp-2 very small, with one naked minute seta and two dentate claws (or one of claws replaced by geniculate seta/spine). P2–P4 rami 3- segmented. P2 endopod ♂ typically 3-segmented and not modified; occasionally 2-segmented but with same total number of spines/setae as in ♀ ( M. costata comb. nov., M. variabilis sp. nov.). P3 endopod ♂ 3-segmented; without inner setae on enp-3 and outer spine modified into spinous apophysis. P4 endopod ♂ occasionally with one ( M. bulbosa comb. nov.) or two ( M. minuta comb. nov.) less(er) inner seta(e) on enp-3. Armature formula of P2–P4 as follows:
P 5 ♀ endopodal lobe with five setae; inner margin without transverse striae. P 5 ♀ exopod elongate, with six or eight elements. P 5 ♂ endopodal lobe with two elements, outer one shortest; exopod 1- or 3-segmented, with 6–7 elements in total. Vestigial P 6 ♀ represented by three minute setae. P 6 ♂ with three setae.
Type species. Thalestris forficula Claus, 1863 = Microthalestris forficula ( Claus, 1863) comb. nov. (by monotypy; Sars 1905: 123) [species inquirenda].
Other species. Thalestris forficuloides Scott & Scott, 1894 = M. forficuloides ( Scott & Scott, 1894) comb. nov.; Parastenhelia gracilis Brady, 1910 = M. gracilis ( Brady, 1910) comb. nov.; M. littoralis Sars, 1911 ; Parastenhelia antarctica Scott, 1912 = M. antarctica ( Scott, 1912) comb. nov.; Parastenhelia costata Pallares, 1982 = M. costata ( Pallares, 1982) comb. nov.; Parastenhelia minuta Pallares, 1982 = M. minuta ( Pallares, 1982) comb. nov.; Parastenhelia bulbosa Gee, 2006 = M. bulbosa ( Gee, 2006) comb. nov.; M. campbelliensis sp. nov.; M. polaris sp. nov.; M. santacruzensis sp. nov.; M. sarsi sp. nov.; M. variabilis sp. nov.
Additional species inquirendae. Harpacticus spinosus Fischer, 1860 ; Parastenhelia spinosa ( Fischer, 1860) sensu Pesta (1959) , sensu Marinov (1974), sensu Wells & Rao (1987) and sensu Kornev & Chertoprud (2008); Parastenhelia spinosa f. bulgarica Apostolov, 1968; Microthalestris forficula ( Claus, 1863) sensu Wilson (1932b) ; Microthalestris littoralis Sars, 1911 sensu Willey (1935) and sensu Nicholls (1945).
Species incertae sedis. Thalestris karmensis Boeck, 1865 ; Dactylopus longipes Boeck, 1873 .


| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.