
Kroeyerina sudamericana, Irigoitia, Manuel M., Cantatore, Delfina M. P., Incorvaia, Inés S. & Timi, Juan T., 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4174.1.10 |
|
publication LSID |
lsid:zoobank.org:pub:E2DFA756-B6B2-44F3-BC03-40A08CB89646 |
|
DOI |
https://doi.org/10.5281/zenodo.5662704 |
|
persistent identifier |
https://treatment.plazi.org/id/B1116A44-190E-AF48-E996-FD5CFB91FE6C |
|
treatment provided by |
Plazi |
|
scientific name |
Kroeyerina sudamericana |
| status |
sp. nov. |
Kroeyerina sudamericana sp. nov.
( Figs. 1–47 View FIGURES 1 – 11 View FIGURES 12 – 15 View FIGURES 16 – 27 View FIGURES 28 – 31 View FIGURES 32 – 39 View FIGURES 40 – 47 )
Type host. Spotback skate Atlantoraja castelnaui ( Rajiformes : Arhynchobatidae ).
Other hosts. Smallnose fanskate Sympterygia bonapartii , bignose fanskate Sympterygia acuta and zipper sand skate Psammobatis extenta ( Rajiformes : Arhynchobatidae ).
Attachment site. Olfactory sacs.
Type locality. Waters off Buenos Aires province, Argentina (34°–41°S; 53°–62°W).
Material examined. 17 females and six males from A. castelnaui (collected November 2011 and November 2013), 14 females and two males from S. bonapartii (collected from 2010 to 2013), 12 females and eight males from S. acuta (Collected November 2014) and one female from P. ex t en t a (collected November 2014).
Type material. Holotype MLP-Cr 26.980 (female), host: A. castelnaui . Allotype MLP-Cr 26.981 (male), host: A. castelnaui . Paratypes MLP-Cr 26.982 (five females and two males from A. castelnaui ). Voucher specimens (two females and two males from S. bonapartii MLP-Cr 26.983; two females and two males from S. acuta MLP-Cr 26.984). All specimens are deposited in the Carcinological Collection of the Museo de La Plata ( CCMLP), La Plata , Argentina.
Prevalence. Atlantoraja castelnaui = 28.6% (CL = 13.3–50.6), Sympterygia bonapartii = 7.5% (CL = 3.9– 12.8), S. acuta 18.2% (CL = 3.3–50).
Mean intensity (range). Atlantoraja castelnaui = 4.8 ± 5.6 (1–15), Sympterygia bonapartii = 1.7 ± 1.2 (1–5), S. acuta = 10 ± 11.3 (2–18).
Etymology. The specific name refers to the subcontinent where the parasites were found, alluding to the first record of the genus Kroeyerina for South America.
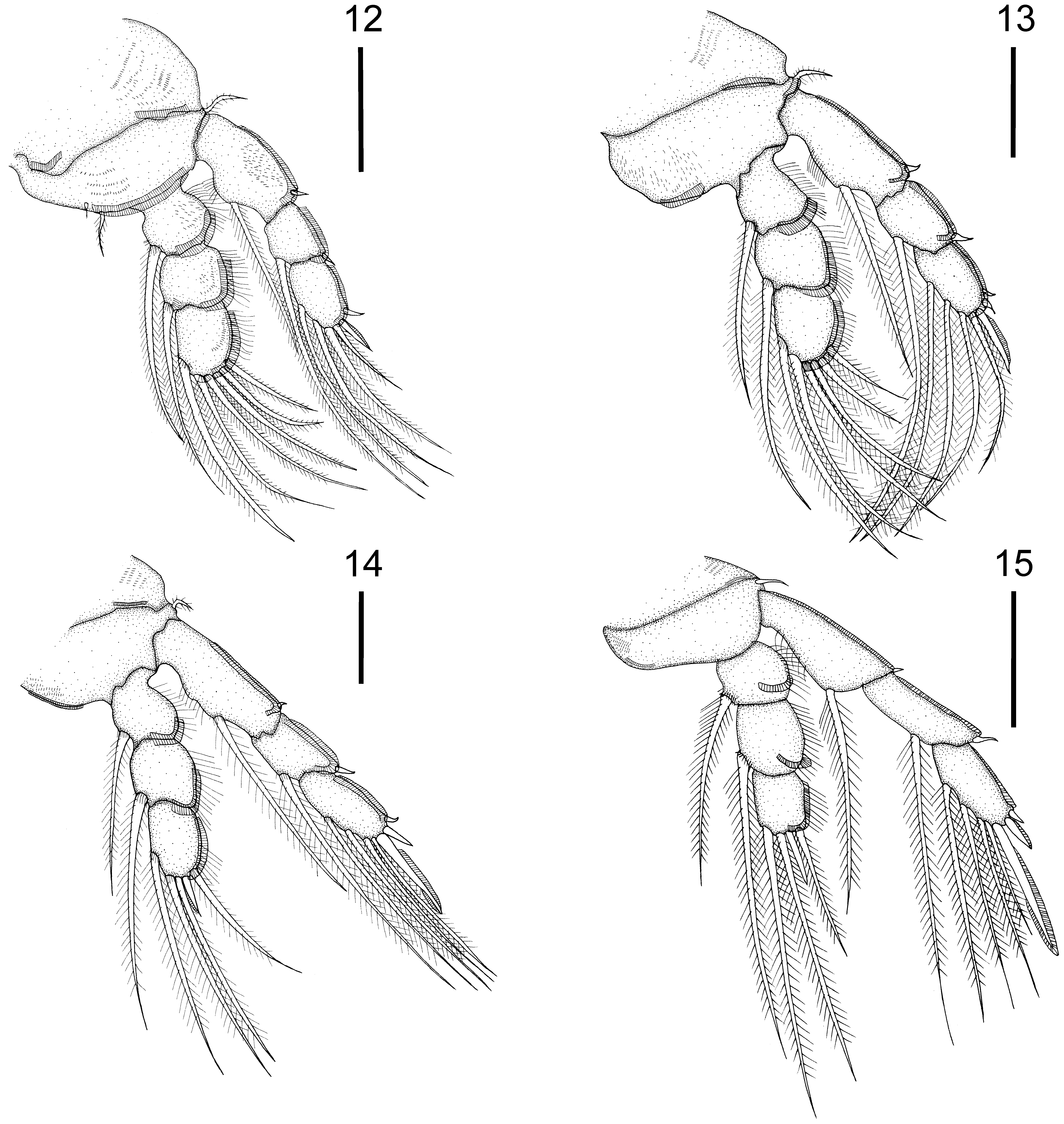
Description of female. ( Figs. 1–15 View FIGURES 1 – 11 View FIGURES 12 – 15 , 32–39 View FIGURES 32 – 39 ; measurements based on 15 specimens). Overall length in dorsal view 3797 ± 568 µm (2500–4,420 µm). Cephalothorax 583 ± 50 µm (490–650 µm) long and 612 ± 85 µm (420– 700 µm) wide. Cephalothoracic sutures ( Fig. 1 View FIGURES 1 – 11 ) arising anterolaterally, uniting posteromedially. Three free thoracic somites with overlapping terga, decreasing in width posteriorly. Genital complex cylindrical ( Figs. 1, 2 View FIGURES 1 – 11 ), 1915 ± 391 µm (1020–2420 µm) long, 531 ± 120 µm (380–750 µm) wide, comprising approximately 50% of total body length, with lateral rows of spinules along entire length. Fifth leg ( Figs. 4 View FIGURES 1 – 11 , 35 View FIGURES 32 – 39 ) located near posterior third of genital complex. Abdomen ( Figs. 1, 2 View FIGURES 1 – 11 ) 477 ± 61 µm (320–540 µm) long, indistinctly 3-segmented, first somite with pair of ventral setules, third somite with membranes on ventral distal margin, all somites with patches of spinules on ventral surface. Caudal ramus ( Figs. 3 View FIGURES 1 – 11 , 33, 34 View FIGURES 32 – 39 ) about three times as long as wide, 195 ± 55 µm (90–250 µm) long, fringed with setules on medial margin and bearing membranes on distal margin of ventral side and six setae as follows: three distal setae, lateral one naked, short and curved, others long and pinnate; two ventral pinnate distolateral setae and one dorsal pinnate distomedial seta.
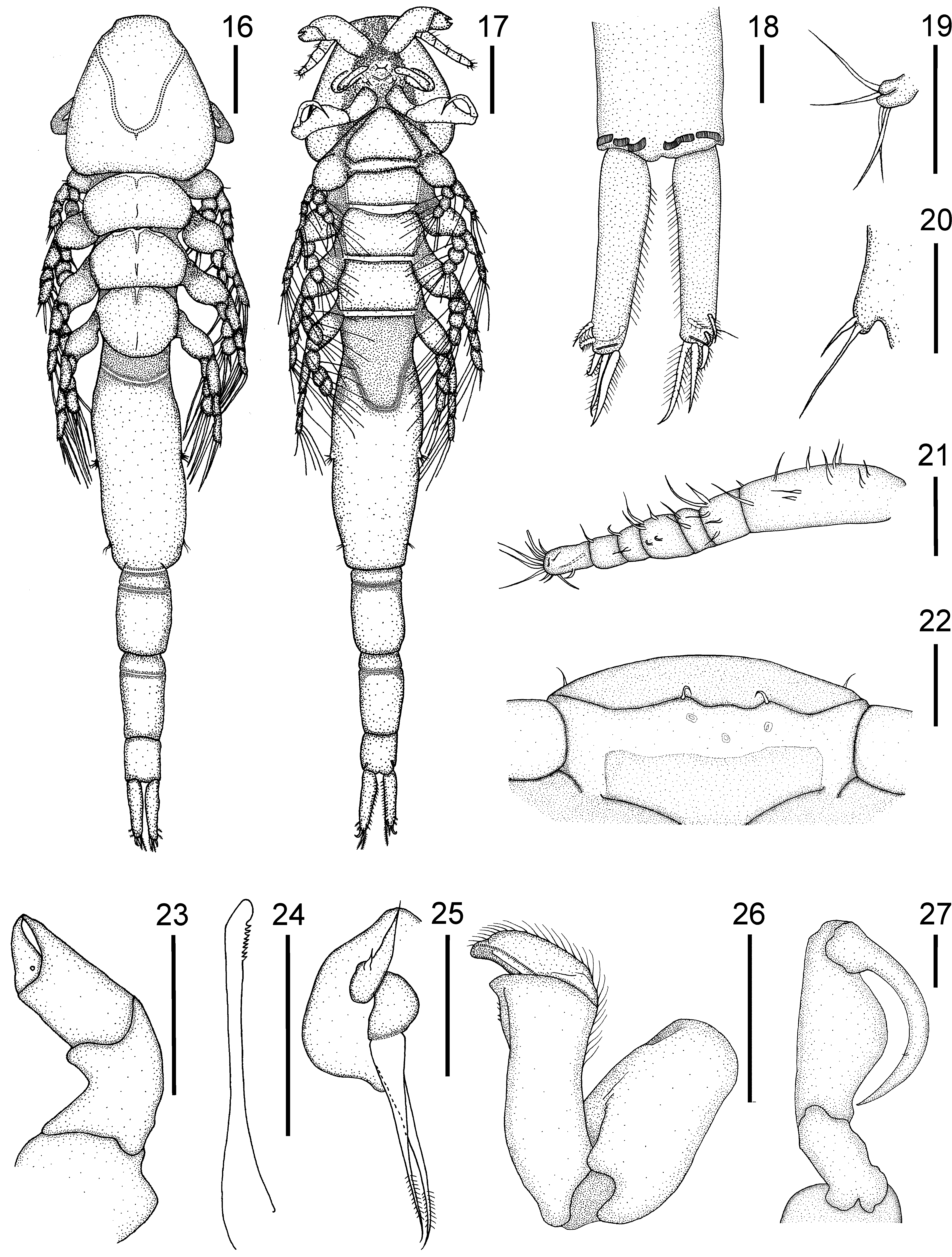
Antennule ( Fig. 5 View FIGURES 1 – 11 ) 7-segmented, armature (base to apex) as follows: 11, 5, 2, 3, 1, 1, 12 + 1 aesthetasc. Ventral surface between bases of antennules bearing two small rostral lobes ( Figs. 6 View FIGURES 1 – 11 , 36 View FIGURES 32 – 39 ) with lateral setule; an additional pair of setules located on dorsal shield near bases of the antennules.
Antenna ( Fig. 7 View FIGURES 1 – 11 ) robust, chelate, 4-segmented; third segment expanded distally into narrow arm with terminal receptacle to receive tip of fourth segment; fourth segment forming thickset claw.
Postantennary process papilliform, lateral to bases of mouthparts. Mouth tube ( Fig. 37 View FIGURES 32 – 39 ) siphonostomatoid; labrum with two setules, slightly anterior to curved transverse fringe of spinules; two lateral patches of spinules separated by scattered smaller spinules and patch of setules; apical border fringed; labium with two lateral rows of setules and fringing apical border.
Mandible ( Fig. 8 View FIGURES 1 – 11 ) siphonostomatoid, with eight equal teeth on blade.
Maxillule ( Figs. 9 View FIGURES 1 – 11 , 37 View FIGURES 32 – 39 ) biramous, composed of broad base and two lobes; outer lobe with two short setae, inner lobe ending in two longer pinnate setae.
Maxilla ( Figs. 10 View FIGURES 1 – 11 , 38 View FIGURES 32 – 39 ) brachiform, lacertus robust, distal half of brachium with depression on medial border, distomedial patch of spinules and two distodorsal rows of setules (not easily visible on ventral view), distal process bearing recurved spine. Calamus scoop-like, bearing two medial and two dorsal serrated membranes.
Maxilliped ( Figs. 11 View FIGURES 1 – 11 , 39 View FIGURES 32 – 39 ) subchelate, four-segmented, corpus unarmed and subchela with small spine.
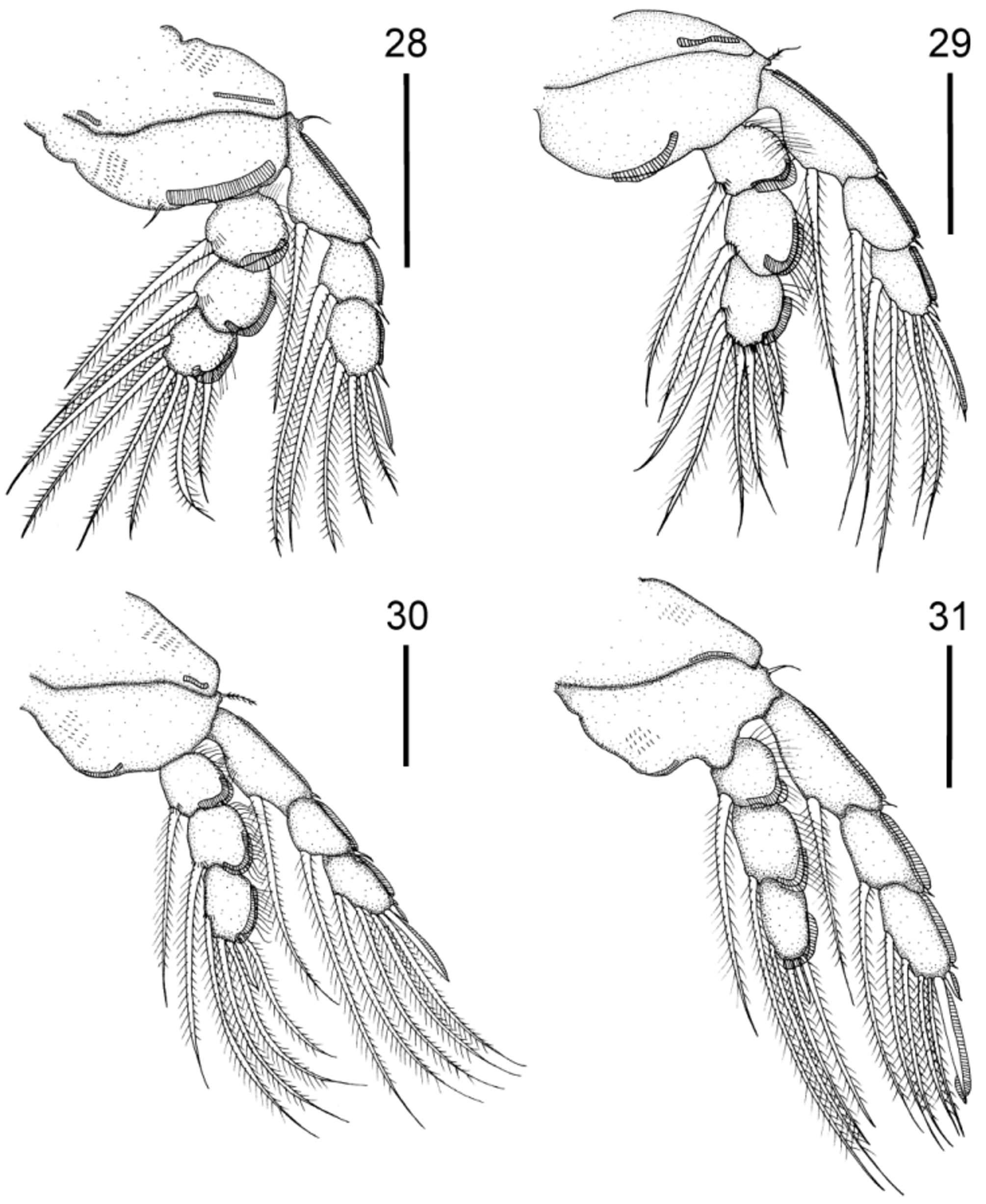
Legs 1–4 ( Figs. 12–15 View FIGURES 12 – 15 ), coxa 1 fringed with two distal membranes on ventral side, coxae 2–3 fringed with distolateral membrane. Basis 1 with lateral pinnate setae, two medial setae, larger pinnate and smaller naked and two distal membranes. Basis 2 with lateral pinnate setae and two distal membranes. Basis 3 with lateral pinnate setae and medial membrane. Basis 4 with lateral naked seta and medial membrane. All exopodal segments of legs 1–4 with lateral membrane. All endopodal segments of legs 1–4 with lateral membrane and dorsolateral row of setules. Armature formula of legs 1–4 as follows (spines in Roman numerals; setae in Arabic numerals):
exopod endopod Larger spines on third exopodal segment with lateral membranes in all legs, those of legs 2–3 also hemipinnate. Leg 5 vestigial, each ramus with pair of setae.
Description of male. ( Figs. 16–31 View FIGURES 16 – 27 View FIGURES 28 – 31 , 40–47 View FIGURES 40 – 47 ; measurements based on six specimens). Overall length in dorsal view 2467 ± 178 µm (2160–2660 µm). Cephalothorax 447 ± 28 µm (400–480 µm) long and 447 ± 43 µm (380–500 µm) wide. Cephalothoracic sutures ( Fig. 16 View FIGURES 16 – 27 ) arising anterolaterally, uniting posteromedially. Three free thoracic somites with overlapping terga, decreasing in width posteriorly. Genital complex ( Figs. 16, 17 View FIGURES 16 – 27 ) cylindrical, 670 ± 113 µm (500–800 µm) long, 258 ± 16 µm (240–280 µm) wide, comprising approximately 27% of total body length, with lateral rows of spinules ( Fig. 43 View FIGURES 40 – 47 ) along entire length. Fifth leg ( Fig. 19 View FIGURES 16 – 27 ) located laterally at middle of genital complex, with four spiniform setae. Sixth leg ( Fig. 20 View FIGURES 16 – 27 ) vestigial, consisting of protuberance with two spiniform setae, located laterally near posterior margin of genital complex. Abdomen ( Figs. 16, 17 View FIGURES 16 – 27 , 40 View FIGURES 40 – 47 ) 588 ± 73 µm (460– 660 µm) long, 3-segmented, first two somites with slight constriction at anterior third, third somite with membranes near junction with caudal rami with patches of spinules on ventral surface. Caudal ramus ( Figs. 18 View FIGURES 16 – 27 , 41, 42 View FIGURES 40 – 47 ) as in female, but four times as long as wide, 217 ± 48 µm (140–260 µm) long and with all setae pinnate.
Antennule ( Fig. 21 View FIGURES 16 – 27 ) 7-segmented, armature (base to apex) as follows: 11, 5, 2, 5, 2, 1, 12 + 1 aesthetasc. Rostral lobes ( Figs. 22 View FIGURES 16 – 27 , 44 View FIGURES 40 – 47 ), rest of cephalic appendages ( Figs. 23–26 View FIGURES 16 – 27 , 45, 46 View FIGURES 40 – 47 ), maxilliped ( Figs. 27 View FIGURES 16 – 27 , 47 View FIGURES 40 – 47 ) and general morphology of legs 1–4 ( Figs. 28–31 View FIGURES 28 – 31 ) as in female.
Remarks. In his phylogenetic analysis of Kroeyerina, Deets (1987) identified two clades presenting differences in the morphology of the rostral lobes ( Benz et al. 2001). One clade, with upturned rostral processes, infects sharks and the other composed of only three species, with reduced lobes, is parasitic on rays. More recently Benz et al. (2001) described a new species, Kroeyerina deetsorum Benz, Smith & Bullard, 2001 , parasitic on the shark Rhizoprionodon terraenovae ( Richardson, 1837) from the Mississippi Delta (Texas, USA). This parasite of sharks, however, does not possess rostral lobes or upturned projections. This characteristic was considered as a possible secondary loss due to the diminutive size (body length 1.2 mm) of K. deetsorum ( Benz et al. 2001) ; these both features clearly differentiate it from the new species.
In the possession of reduced rostral lobes, the new species is similar to congeners forming the second clade proposed by Deets (1987), namely Kroeyerina nasuta Wilson, 1932 , a parasite of Dasybatus marinus Garman, 1913 (= Dasyatis centroura (Mitchill, 1815)) from the western North Atlantic (Woods Hole, USA) ( Wilson 1932), Kroeyerina deborahae Deets, 1987 parasitic on Rhinobatos productus Ayres, 1854 from the eastern North Pacific (Palos Verdes, California) and Kroeyerina mobulae Deets, 1987 , a parasite of Mobula lucasana Beebe & Tee-Van, 1938 (= Mobula thurstoni (Lloyd, 1908)) and Mobula japanica ( Müller & Henle, 1841) from the eastern North Pacific (Sea of Cortez) ( Deets 1987).
Both males and females of the new species are readily distinguished from K. mobulae , which possesses a maxilliped with a more robust subchela and has extremely reduced setae on the caudal rami. Kroeyerina deborahae differs from the new species in having a caudal ramus with all setae naked; an almost straight subchela on the maxilliped, a smaller number of elements on the antennule and a proportionally shorter genital complex (45% of total body length). Kroeyerina nasuta has a caudal ramus similar to the new species, but can be distinguished from it by possessing a different armature of the antennule and a proportionally larger genital complex (57% of total body length). Additionally, in both K. deborahae and K. nasuta , the chela of the antenna which, when closed, leaves no gap between the corpus and the claw. The claw bears stout spines ( Deets 1987). The males of K. deborahae and K. nasuta are unknown.
Based on these differences a new species, Kroeyerina sudamericana sp. nov., is proposed. This new species represents the first record of Kroeyerina parasitizing members of the Arhynchobatidae (Rajiformes) . It is noteworthy that the hosts of K. nasuta and K. mobulae belong to the order Myliobatiformes , and although the host of K. deborahae belongs to the Rajiformes , it is a guitarfish of the family Rhynobatidae.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.