CN113966401A - 用于治疗amd的htra1调节 - Google Patents
用于治疗amd的htra1调节 Download PDFInfo
- Publication number
- CN113966401A CN113966401A CN202080042929.4A CN202080042929A CN113966401A CN 113966401 A CN113966401 A CN 113966401A CN 202080042929 A CN202080042929 A CN 202080042929A CN 113966401 A CN113966401 A CN 113966401A
- Authority
- CN
- China
- Prior art keywords
- htra1
- sequence
- risk
- dna
- protein
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- 102100021119 Serine protease HTRA1 Human genes 0.000 title claims abstract description 345
- 101001041393 Homo sapiens Serine protease HTRA1 Proteins 0.000 title claims abstract description 306
- 206010064930 age-related macular degeneration Diseases 0.000 title claims abstract description 159
- 238000011282 treatment Methods 0.000 title claims description 27
- 230000014509 gene expression Effects 0.000 claims abstract description 191
- 238000000034 method Methods 0.000 claims abstract description 180
- 208000002780 macular degeneration Diseases 0.000 claims abstract description 156
- 210000000349 chromosome Anatomy 0.000 claims abstract description 35
- 108700028369 Alleles Proteins 0.000 claims description 208
- 210000004027 cell Anatomy 0.000 claims description 169
- 108020004414 DNA Proteins 0.000 claims description 140
- 108090000623 proteins and genes Proteins 0.000 claims description 137
- 108020005004 Guide RNA Proteins 0.000 claims description 109
- 102000004169 proteins and genes Human genes 0.000 claims description 101
- 108091033409 CRISPR Proteins 0.000 claims description 83
- 230000001965 increasing effect Effects 0.000 claims description 68
- 239000003795 chemical substances by application Substances 0.000 claims description 63
- 101150007028 HTRA1 gene Proteins 0.000 claims description 61
- 239000013598 vector Substances 0.000 claims description 58
- 108091006106 transcriptional activators Proteins 0.000 claims description 53
- 125000003729 nucleotide group Chemical group 0.000 claims description 52
- 239000002773 nucleotide Substances 0.000 claims description 51
- 230000027455 binding Effects 0.000 claims description 50
- 230000001105 regulatory effect Effects 0.000 claims description 49
- 101001020544 Homo sapiens LIM/homeobox protein Lhx2 Proteins 0.000 claims description 41
- 102100036132 LIM/homeobox protein Lhx2 Human genes 0.000 claims description 41
- 238000004519 manufacturing process Methods 0.000 claims description 39
- 238000013518 transcription Methods 0.000 claims description 39
- 230000035897 transcription Effects 0.000 claims description 39
- 238000010354 CRISPR gene editing Methods 0.000 claims description 38
- 150000007523 nucleic acids Chemical class 0.000 claims description 35
- 230000008685 targeting Effects 0.000 claims description 35
- 230000000694 effects Effects 0.000 claims description 33
- 239000003623 enhancer Substances 0.000 claims description 30
- 102000040430 polynucleotide Human genes 0.000 claims description 30
- 108091033319 polynucleotide Proteins 0.000 claims description 30
- 239000002157 polynucleotide Substances 0.000 claims description 30
- 108090000765 processed proteins & peptides Proteins 0.000 claims description 30
- 108091028043 Nucleic acid sequence Proteins 0.000 claims description 28
- 102000037865 fusion proteins Human genes 0.000 claims description 28
- 108020001507 fusion proteins Proteins 0.000 claims description 28
- 102000039446 nucleic acids Human genes 0.000 claims description 28
- 108020004707 nucleic acids Proteins 0.000 claims description 28
- 239000013603 viral vector Substances 0.000 claims description 27
- 101710163270 Nuclease Proteins 0.000 claims description 25
- 230000004913 activation Effects 0.000 claims description 22
- 102000004196 processed proteins & peptides Human genes 0.000 claims description 22
- 238000011161 development Methods 0.000 claims description 21
- 102200139266 rs10490924 Human genes 0.000 claims description 21
- 230000008439 repair process Effects 0.000 claims description 19
- 230000000295 complement effect Effects 0.000 claims description 18
- 229920001184 polypeptide Polymers 0.000 claims description 18
- 239000003814 drug Substances 0.000 claims description 15
- 241000702421 Dependoparvovirus Species 0.000 claims description 14
- 241000713666 Lentivirus Species 0.000 claims description 12
- 208000032023 Signs and Symptoms Diseases 0.000 claims description 12
- 210000002287 horizontal cell Anatomy 0.000 claims description 12
- 108700039691 Genetic Promoter Regions Proteins 0.000 claims description 11
- 108020001580 protein domains Proteins 0.000 claims description 8
- 108700004991 Cas12a Proteins 0.000 claims description 7
- 102220050813 rs3750847 Human genes 0.000 claims description 7
- 108010068250 Herpes Simplex Virus Protein Vmw65 Proteins 0.000 claims description 6
- 125000006850 spacer group Chemical group 0.000 claims description 6
- 102100032606 Heat shock factor protein 1 Human genes 0.000 claims description 5
- 101000867525 Homo sapiens Heat shock factor protein 1 Proteins 0.000 claims description 5
- 238000002360 preparation method Methods 0.000 claims description 4
- 229920002477 rna polymer Polymers 0.000 claims description 4
- 241001430294 unidentified retrovirus Species 0.000 claims description 4
- 102000015280 CCAAT-Enhancer-Binding Protein-beta Human genes 0.000 claims description 3
- 108010064535 CCAAT-Enhancer-Binding Protein-beta Proteins 0.000 claims description 3
- 108010040467 CRISPR-Associated Proteins Proteins 0.000 claims description 3
- 101000666382 Homo sapiens Transcription factor E2-alpha Proteins 0.000 claims description 3
- 108010057466 NF-kappa B Proteins 0.000 claims description 3
- 102000003945 NF-kappa B Human genes 0.000 claims description 3
- 108010018525 NFATC Transcription Factors Proteins 0.000 claims description 3
- 102000002673 NFATC Transcription Factors Human genes 0.000 claims description 3
- 102100038313 Transcription factor E2-alpha Human genes 0.000 claims description 3
- 241001529453 unidentified herpesvirus Species 0.000 claims description 3
- 102100022103 Histone-lysine N-methyltransferase 2A Human genes 0.000 claims description 2
- 101001045846 Homo sapiens Histone-lysine N-methyltransferase 2A Proteins 0.000 claims description 2
- 102000004389 Ribonucleoproteins Human genes 0.000 claims 18
- 108010081734 Ribonucleoproteins Proteins 0.000 claims 18
- 238000001415 gene therapy Methods 0.000 abstract description 37
- 239000000203 mixture Substances 0.000 abstract description 18
- 210000000844 retinal pigment epithelial cell Anatomy 0.000 abstract description 3
- 235000018102 proteins Nutrition 0.000 description 86
- 239000013612 plasmid Substances 0.000 description 67
- 108020004999 messenger RNA Proteins 0.000 description 43
- 108091034117 Oligonucleotide Proteins 0.000 description 36
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 36
- 230000015572 biosynthetic process Effects 0.000 description 36
- 108091027544 Subgenomic mRNA Proteins 0.000 description 34
- 210000001519 tissue Anatomy 0.000 description 34
- 102000040945 Transcription factor Human genes 0.000 description 32
- 108091023040 Transcription factor Proteins 0.000 description 32
- 238000003786 synthesis reaction Methods 0.000 description 31
- 230000000875 corresponding effect Effects 0.000 description 24
- 230000002829 reductive effect Effects 0.000 description 22
- 210000001525 retina Anatomy 0.000 description 22
- 238000013459 approach Methods 0.000 description 20
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 20
- 108700019146 Transgenes Proteins 0.000 description 18
- 230000009467 reduction Effects 0.000 description 17
- 102100038568 Age-related maculopathy susceptibility protein 2 Human genes 0.000 description 16
- 101000808726 Homo sapiens Age-related maculopathy susceptibility protein 2 Proteins 0.000 description 16
- 230000002068 genetic effect Effects 0.000 description 16
- 230000002207 retinal effect Effects 0.000 description 16
- 239000000523 sample Substances 0.000 description 16
- 201000010099 disease Diseases 0.000 description 15
- 238000002347 injection Methods 0.000 description 15
- 239000007924 injection Substances 0.000 description 15
- 108010008532 Deoxyribonuclease I Proteins 0.000 description 14
- 102000007260 Deoxyribonuclease I Human genes 0.000 description 14
- 238000004458 analytical method Methods 0.000 description 14
- 210000003161 choroid Anatomy 0.000 description 14
- 238000002560 therapeutic procedure Methods 0.000 description 14
- 230000002103 transcriptional effect Effects 0.000 description 14
- 108091026890 Coding region Proteins 0.000 description 13
- 102000010834 Extracellular Matrix Proteins Human genes 0.000 description 13
- 108010037362 Extracellular Matrix Proteins Proteins 0.000 description 13
- 101100517196 Arabidopsis thaliana NRPE1 gene Proteins 0.000 description 12
- 101100190825 Bos taurus PMEL gene Proteins 0.000 description 12
- 101100073341 Oryza sativa subsp. japonica KAO gene Proteins 0.000 description 12
- 125000003275 alpha amino acid group Chemical group 0.000 description 12
- 210000001775 bruch membrane Anatomy 0.000 description 12
- 102000054766 genetic haplotypes Human genes 0.000 description 12
- 101150005492 rpe1 gene Proteins 0.000 description 12
- 238000012360 testing method Methods 0.000 description 12
- 238000003776 cleavage reaction Methods 0.000 description 11
- 238000012217 deletion Methods 0.000 description 11
- 239000002245 particle Substances 0.000 description 11
- 230000007017 scission Effects 0.000 description 11
- 230000001225 therapeutic effect Effects 0.000 description 11
- 238000010453 CRISPR/Cas method Methods 0.000 description 10
- ISAKRJDGNUQOIC-UHFFFAOYSA-N Uracil Chemical compound O=C1C=CNC(=O)N1 ISAKRJDGNUQOIC-UHFFFAOYSA-N 0.000 description 10
- 235000001014 amino acid Nutrition 0.000 description 10
- 238000003556 assay Methods 0.000 description 10
- 230000037430 deletion Effects 0.000 description 10
- 239000000047 product Substances 0.000 description 10
- 230000004568 DNA-binding Effects 0.000 description 9
- 108700028146 Genetic Enhancer Elements Proteins 0.000 description 9
- 102000012479 Serine Proteases Human genes 0.000 description 9
- 108010022999 Serine Proteases Proteins 0.000 description 9
- 238000010459 TALEN Methods 0.000 description 9
- 241000700605 Viruses Species 0.000 description 9
- 150000001413 amino acids Chemical class 0.000 description 9
- 150000001875 compounds Chemical class 0.000 description 9
- 238000013461 design Methods 0.000 description 9
- 230000007246 mechanism Effects 0.000 description 9
- 229940124597 therapeutic agent Drugs 0.000 description 9
- 241000702423 Adeno-associated virus - 2 Species 0.000 description 8
- 102000004533 Endonucleases Human genes 0.000 description 8
- 108010042407 Endonucleases Proteins 0.000 description 8
- OPTASPLRGRRNAP-UHFFFAOYSA-N cytosine Chemical compound NC=1C=CNC(=O)N=1 OPTASPLRGRRNAP-UHFFFAOYSA-N 0.000 description 8
- 210000002744 extracellular matrix Anatomy 0.000 description 8
- 239000000284 extract Substances 0.000 description 8
- 239000012634 fragment Substances 0.000 description 8
- 230000006798 recombination Effects 0.000 description 8
- 238000005215 recombination Methods 0.000 description 8
- 230000002441 reversible effect Effects 0.000 description 8
- -1 small molecule compound Chemical class 0.000 description 8
- 208000024891 symptom Diseases 0.000 description 8
- 238000010361 transduction Methods 0.000 description 8
- 238000011144 upstream manufacturing Methods 0.000 description 8
- 208000005590 Choroidal Neovascularization Diseases 0.000 description 7
- 206010060823 Choroidal neovascularisation Diseases 0.000 description 7
- 102000053602 DNA Human genes 0.000 description 7
- 101000729271 Homo sapiens Retinoid isomerohydrolase Proteins 0.000 description 7
- 102100031176 Retinoid isomerohydrolase Human genes 0.000 description 7
- 238000004113 cell culture Methods 0.000 description 7
- 239000012228 culture supernatant Substances 0.000 description 7
- 230000007423 decrease Effects 0.000 description 7
- 230000002950 deficient Effects 0.000 description 7
- 238000010362 genome editing Methods 0.000 description 7
- 230000004048 modification Effects 0.000 description 7
- 238000012986 modification Methods 0.000 description 7
- 230000035772 mutation Effects 0.000 description 7
- 210000000608 photoreceptor cell Anatomy 0.000 description 7
- 230000029279 positive regulation of transcription, DNA-dependent Effects 0.000 description 7
- KDCGOANMDULRCW-UHFFFAOYSA-N 7H-purine Chemical compound N1=CNC2=NC=NC2=C1 KDCGOANMDULRCW-UHFFFAOYSA-N 0.000 description 6
- 241000701022 Cytomegalovirus Species 0.000 description 6
- 108010017070 Zinc Finger Nucleases Proteins 0.000 description 6
- 230000009286 beneficial effect Effects 0.000 description 6
- 239000002299 complementary DNA Substances 0.000 description 6
- 230000003247 decreasing effect Effects 0.000 description 6
- 238000004520 electroporation Methods 0.000 description 6
- 230000001973 epigenetic effect Effects 0.000 description 6
- 230000004438 eyesight Effects 0.000 description 6
- 239000012530 fluid Substances 0.000 description 6
- 230000006870 function Effects 0.000 description 6
- 230000006872 improvement Effects 0.000 description 6
- 230000001404 mediated effect Effects 0.000 description 6
- 230000002018 overexpression Effects 0.000 description 6
- 238000006467 substitution reaction Methods 0.000 description 6
- 230000026683 transduction Effects 0.000 description 6
- 238000001890 transfection Methods 0.000 description 6
- 241000701161 unidentified adenovirus Species 0.000 description 6
- 230000003827 upregulation Effects 0.000 description 6
- 230000003612 virological effect Effects 0.000 description 6
- 102100022794 Bestrophin-1 Human genes 0.000 description 5
- 101000903449 Homo sapiens Bestrophin-1 Proteins 0.000 description 5
- 108020005198 Long Noncoding RNA Proteins 0.000 description 5
- 108010029485 Protein Isoforms Proteins 0.000 description 5
- 102000001708 Protein Isoforms Human genes 0.000 description 5
- 241000193996 Streptococcus pyogenes Species 0.000 description 5
- 101000910035 Streptococcus pyogenes serotype M1 CRISPR-associated endonuclease Cas9/Csn1 Proteins 0.000 description 5
- 108091046869 Telomeric non-coding RNA Proteins 0.000 description 5
- 108010043645 Transcription Activator-Like Effector Nucleases Proteins 0.000 description 5
- HCHKCACWOHOZIP-UHFFFAOYSA-N Zinc Chemical compound [Zn] HCHKCACWOHOZIP-UHFFFAOYSA-N 0.000 description 5
- 101150063416 add gene Proteins 0.000 description 5
- 230000008901 benefit Effects 0.000 description 5
- 239000000872 buffer Substances 0.000 description 5
- 239000013611 chromosomal DNA Substances 0.000 description 5
- 230000001186 cumulative effect Effects 0.000 description 5
- 208000031513 cyst Diseases 0.000 description 5
- 230000001419 dependent effect Effects 0.000 description 5
- 239000012636 effector Substances 0.000 description 5
- 238000002474 experimental method Methods 0.000 description 5
- VPZXBVLAVMBEQI-UHFFFAOYSA-N glycyl-DL-alpha-alanine Natural products OC(=O)C(C)NC(=O)CN VPZXBVLAVMBEQI-UHFFFAOYSA-N 0.000 description 5
- 230000012010 growth Effects 0.000 description 5
- 238000001727 in vivo Methods 0.000 description 5
- 239000002502 liposome Substances 0.000 description 5
- 239000008194 pharmaceutical composition Substances 0.000 description 5
- 230000008488 polyadenylation Effects 0.000 description 5
- 230000007115 recruitment Effects 0.000 description 5
- 229940035893 uracil Drugs 0.000 description 5
- 230000000007 visual effect Effects 0.000 description 5
- 239000011701 zinc Substances 0.000 description 5
- 229910052725 zinc Inorganic materials 0.000 description 5
- KRHRBKYBJXMYBB-WHFBIAKZSA-N Ala-Cys-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CS)C(=O)NCC(O)=O KRHRBKYBJXMYBB-WHFBIAKZSA-N 0.000 description 4
- 108010077544 Chromatin Proteins 0.000 description 4
- 206010016654 Fibrosis Diseases 0.000 description 4
- PGPJSRSLQNXBDT-YUMQZZPRSA-N Gln-Arg-Gly Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)NCC(O)=O PGPJSRSLQNXBDT-YUMQZZPRSA-N 0.000 description 4
- IVGJYOOGJLFKQE-AVGNSLFASA-N Glu-Leu-Lys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)O)N IVGJYOOGJLFKQE-AVGNSLFASA-N 0.000 description 4
- 108010033040 Histones Proteins 0.000 description 4
- 101001033249 Homo sapiens Interleukin-1 beta Proteins 0.000 description 4
- 101000979347 Homo sapiens Nuclear factor 1 X-type Proteins 0.000 description 4
- 102100039065 Interleukin-1 beta Human genes 0.000 description 4
- 108010018650 MEF2 Transcription Factors Proteins 0.000 description 4
- 102100021148 Myocyte-specific enhancer factor 2A Human genes 0.000 description 4
- 108010079364 N-glycylalanine Proteins 0.000 description 4
- 102100023049 Nuclear factor 1 X-type Human genes 0.000 description 4
- 241000192142 Proteobacteria Species 0.000 description 4
- 238000003559 RNA-seq method Methods 0.000 description 4
- 238000011529 RT qPCR Methods 0.000 description 4
- 108091028113 Trans-activating crRNA Proteins 0.000 description 4
- 102100031142 Transcriptional repressor protein YY1 Human genes 0.000 description 4
- 238000009825 accumulation Methods 0.000 description 4
- 230000021736 acetylation Effects 0.000 description 4
- 238000006640 acetylation reaction Methods 0.000 description 4
- 230000000692 anti-sense effect Effects 0.000 description 4
- 230000033228 biological regulation Effects 0.000 description 4
- 239000000090 biomarker Substances 0.000 description 4
- 238000010804 cDNA synthesis Methods 0.000 description 4
- 101150038500 cas9 gene Proteins 0.000 description 4
- 238000006243 chemical reaction Methods 0.000 description 4
- 210000003483 chromatin Anatomy 0.000 description 4
- 230000002759 chromosomal effect Effects 0.000 description 4
- 230000002596 correlated effect Effects 0.000 description 4
- 229940104302 cytosine Drugs 0.000 description 4
- 230000002255 enzymatic effect Effects 0.000 description 4
- 230000004761 fibrosis Effects 0.000 description 4
- 238000003780 insertion Methods 0.000 description 4
- 230000037431 insertion Effects 0.000 description 4
- 230000017730 intein-mediated protein splicing Effects 0.000 description 4
- 108010034529 leucyl-lysine Proteins 0.000 description 4
- 239000003550 marker Substances 0.000 description 4
- 239000000463 material Substances 0.000 description 4
- 230000009437 off-target effect Effects 0.000 description 4
- 238000003757 reverse transcription PCR Methods 0.000 description 4
- 239000000243 solution Substances 0.000 description 4
- 239000013607 AAV vector Substances 0.000 description 3
- 108010079649 APOBEC-1 Deaminase Proteins 0.000 description 3
- UXRVDHVARNBOIO-QSFUFRPTSA-N Asp-Val-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CC(=O)O)N UXRVDHVARNBOIO-QSFUFRPTSA-N 0.000 description 3
- 206010003694 Atrophy Diseases 0.000 description 3
- 201000004569 Blindness Diseases 0.000 description 3
- 102100040397 C->U-editing enzyme APOBEC-1 Human genes 0.000 description 3
- 101710172824 CRISPR-associated endonuclease Cas9 Proteins 0.000 description 3
- 108020004705 Codon Proteins 0.000 description 3
- 108020004635 Complementary DNA Proteins 0.000 description 3
- 108010031325 Cytidine deaminase Proteins 0.000 description 3
- 238000012286 ELISA Assay Methods 0.000 description 3
- RIMMMMYKGIBOSN-DCAQKATOSA-N Leu-Asn-Met Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CCSC)C(O)=O RIMMMMYKGIBOSN-DCAQKATOSA-N 0.000 description 3
- QOJDBRUCOXQSSK-AJNGGQMLSA-N Lys-Ile-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCCN)C(O)=O QOJDBRUCOXQSSK-AJNGGQMLSA-N 0.000 description 3
- 102100026261 Metalloproteinase inhibitor 3 Human genes 0.000 description 3
- SITLTJHOQZFJGG-UHFFFAOYSA-N N-L-alpha-glutamyl-L-valine Natural products CC(C)C(C(O)=O)NC(=O)C(N)CCC(O)=O SITLTJHOQZFJGG-UHFFFAOYSA-N 0.000 description 3
- 101150114527 Nkx2-5 gene Proteins 0.000 description 3
- 102000011931 Nucleoproteins Human genes 0.000 description 3
- 108010061100 Nucleoproteins Proteins 0.000 description 3
- CZPWVGJYEJSRLH-UHFFFAOYSA-N Pyrimidine Chemical compound C1=CN=CN=C1 CZPWVGJYEJSRLH-UHFFFAOYSA-N 0.000 description 3
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 3
- 108010031429 Tissue Inhibitor of Metalloproteinase-3 Proteins 0.000 description 3
- 239000012190 activator Substances 0.000 description 3
- 238000000246 agarose gel electrophoresis Methods 0.000 description 3
- 108010087924 alanylproline Proteins 0.000 description 3
- 108010068380 arginylarginine Proteins 0.000 description 3
- 230000037444 atrophy Effects 0.000 description 3
- 210000002469 basement membrane Anatomy 0.000 description 3
- 230000001364 causal effect Effects 0.000 description 3
- 108010016616 cysteinylglycine Proteins 0.000 description 3
- 230000008021 deposition Effects 0.000 description 3
- 208000035475 disorder Diseases 0.000 description 3
- 230000005782 double-strand break Effects 0.000 description 3
- 230000001605 fetal effect Effects 0.000 description 3
- 238000001476 gene delivery Methods 0.000 description 3
- 238000000338 in vitro Methods 0.000 description 3
- 238000001802 infusion Methods 0.000 description 3
- 239000003112 inhibitor Substances 0.000 description 3
- 230000003902 lesion Effects 0.000 description 3
- 239000013642 negative control Substances 0.000 description 3
- 230000001537 neural effect Effects 0.000 description 3
- 238000010606 normalization Methods 0.000 description 3
- 238000005457 optimization Methods 0.000 description 3
- 108010051242 phenylalanylserine Proteins 0.000 description 3
- 239000013641 positive control Substances 0.000 description 3
- 238000012545 processing Methods 0.000 description 3
- 239000013646 rAAV2 vector Substances 0.000 description 3
- 238000011160 research Methods 0.000 description 3
- 230000004044 response Effects 0.000 description 3
- 238000012552 review Methods 0.000 description 3
- 230000035945 sensitivity Effects 0.000 description 3
- 239000000126 substance Substances 0.000 description 3
- 238000001356 surgical procedure Methods 0.000 description 3
- 238000013519 translation Methods 0.000 description 3
- AXFMEGAFCUULFV-BLFANLJRSA-N (2s)-2-[[(2s)-1-[(2s,3r)-2-amino-3-methylpentanoyl]pyrrolidine-2-carbonyl]amino]pentanedioic acid Chemical compound CC[C@@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O AXFMEGAFCUULFV-BLFANLJRSA-N 0.000 description 2
- 108091032973 (ribonucleotides)n+m Proteins 0.000 description 2
- 102000040650 (ribonucleotides)n+m Human genes 0.000 description 2
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 2
- 101150036609 ARMS2 gene Proteins 0.000 description 2
- 108010052875 Adenine deaminase Proteins 0.000 description 2
- UWQJHXKARZWDIJ-ZLUOBGJFSA-N Ala-Ala-Cys Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CS)C(O)=O UWQJHXKARZWDIJ-ZLUOBGJFSA-N 0.000 description 2
- SSSROGPPPVTHLX-FXQIFTODSA-N Ala-Arg-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(O)=O SSSROGPPPVTHLX-FXQIFTODSA-N 0.000 description 2
- GFBLJMHGHAXGNY-ZLUOBGJFSA-N Ala-Asn-Asp Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CC(O)=O)C(O)=O GFBLJMHGHAXGNY-ZLUOBGJFSA-N 0.000 description 2
- IKKVASZHTMKJIR-ZKWXMUAHSA-N Ala-Asp-Val Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O IKKVASZHTMKJIR-ZKWXMUAHSA-N 0.000 description 2
- FUSPCLTUKXQREV-ACZMJKKPSA-N Ala-Glu-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(O)=O FUSPCLTUKXQREV-ACZMJKKPSA-N 0.000 description 2
- LMFXXZPPZDCPTA-ZKWXMUAHSA-N Ala-Gly-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N LMFXXZPPZDCPTA-ZKWXMUAHSA-N 0.000 description 2
- PCIFXPRIFWKWLK-YUMQZZPRSA-N Ala-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@H](C)N PCIFXPRIFWKWLK-YUMQZZPRSA-N 0.000 description 2
- DVJSJDDYCYSMFR-ZKWXMUAHSA-N Ala-Ile-Gly Chemical compound [H]N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)NCC(O)=O DVJSJDDYCYSMFR-ZKWXMUAHSA-N 0.000 description 2
- DCVYRWFAMZFSDA-ZLUOBGJFSA-N Ala-Ser-Ala Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O DCVYRWFAMZFSDA-ZLUOBGJFSA-N 0.000 description 2
- RTZCUEHYUQZIDE-WHFBIAKZSA-N Ala-Ser-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O RTZCUEHYUQZIDE-WHFBIAKZSA-N 0.000 description 2
- NCQMBSJGJMYKCK-ZLUOBGJFSA-N Ala-Ser-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O NCQMBSJGJMYKCK-ZLUOBGJFSA-N 0.000 description 2
- GXCSUJQOECMKPV-CIUDSAMLSA-N Arg-Ala-Gln Chemical compound C[C@H](NC(=O)[C@@H](N)CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O GXCSUJQOECMKPV-CIUDSAMLSA-N 0.000 description 2
- KWKQGHSSNHPGOW-BQBZGAKWSA-N Arg-Ala-Gly Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)NCC(O)=O KWKQGHSSNHPGOW-BQBZGAKWSA-N 0.000 description 2
- KXOPYFNQLVUOAQ-FXQIFTODSA-N Arg-Ser-Ala Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O KXOPYFNQLVUOAQ-FXQIFTODSA-N 0.000 description 2
- FRBAHXABMQXSJQ-FXQIFTODSA-N Arg-Ser-Ser Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O FRBAHXABMQXSJQ-FXQIFTODSA-N 0.000 description 2
- NXVGBGZQQFDUTM-XVYDVKMFSA-N Asn-Ala-His Chemical compound C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC(=O)N)N NXVGBGZQQFDUTM-XVYDVKMFSA-N 0.000 description 2
- CTQIOCMSIJATNX-WHFBIAKZSA-N Asn-Gly-Ala Chemical compound [H]N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](C)C(O)=O CTQIOCMSIJATNX-WHFBIAKZSA-N 0.000 description 2
- MVXJBVVLACEGCG-PCBIJLKTSA-N Asn-Phe-Ile Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O MVXJBVVLACEGCG-PCBIJLKTSA-N 0.000 description 2
- VWADICJNCPFKJS-ZLUOBGJFSA-N Asn-Ser-Asp Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O VWADICJNCPFKJS-ZLUOBGJFSA-N 0.000 description 2
- HPBNLFLSSQDFQW-WHFBIAKZSA-N Asn-Ser-Gly Chemical compound NC(=O)C[C@H](N)C(=O)N[C@@H](CO)C(=O)NCC(O)=O HPBNLFLSSQDFQW-WHFBIAKZSA-N 0.000 description 2
- JBDLMLZNDRLDIX-HJGDQZAQSA-N Asn-Thr-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(O)=O JBDLMLZNDRLDIX-HJGDQZAQSA-N 0.000 description 2
- KDFQZBWWPYQBEN-ZLUOBGJFSA-N Asp-Ala-Asn Chemical compound C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)O)N KDFQZBWWPYQBEN-ZLUOBGJFSA-N 0.000 description 2
- KTTCQQNRRLCIBC-GHCJXIJMSA-N Asp-Ile-Ala Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O KTTCQQNRRLCIBC-GHCJXIJMSA-N 0.000 description 2
- 241000894006 Bacteria Species 0.000 description 2
- 241000701822 Bovine papillomavirus Species 0.000 description 2
- 238000010356 CRISPR-Cas9 genome editing Methods 0.000 description 2
- 102100035432 Complement factor H Human genes 0.000 description 2
- 108010051219 Cre recombinase Proteins 0.000 description 2
- 241000192700 Cyanobacteria Species 0.000 description 2
- XTHUKRLJRUVVBF-WHFBIAKZSA-N Cys-Gly-Ser Chemical compound SC[C@H](N)C(=O)NCC(=O)N[C@@H](CO)C(O)=O XTHUKRLJRUVVBF-WHFBIAKZSA-N 0.000 description 2
- MQQLYEHXSBJTRK-FXQIFTODSA-N Cys-Val-Cys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CS)N MQQLYEHXSBJTRK-FXQIFTODSA-N 0.000 description 2
- 102100026846 Cytidine deaminase Human genes 0.000 description 2
- 230000007018 DNA scission Effects 0.000 description 2
- 238000002965 ELISA Methods 0.000 description 2
- 102100021977 Ectonucleotide pyrophosphatase/phosphodiesterase family member 2 Human genes 0.000 description 2
- 208000002197 Ehlers-Danlos syndrome Diseases 0.000 description 2
- 102000004190 Enzymes Human genes 0.000 description 2
- 108090000790 Enzymes Proteins 0.000 description 2
- 241000192125 Firmicutes Species 0.000 description 2
- CLPQUWHBWXFJOX-BQBZGAKWSA-N Gln-Gly-Gln Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(O)=O CLPQUWHBWXFJOX-BQBZGAKWSA-N 0.000 description 2
- YPMDZWPZFOZYFG-GUBZILKMSA-N Gln-Leu-Ser Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CO)C(O)=O YPMDZWPZFOZYFG-GUBZILKMSA-N 0.000 description 2
- LGYCLOCORAEQSZ-PEFMBERDSA-N Glu-Ile-Asp Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=O LGYCLOCORAEQSZ-PEFMBERDSA-N 0.000 description 2
- OQXDUSZKISQQSS-GUBZILKMSA-N Glu-Lys-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(O)=O OQXDUSZKISQQSS-GUBZILKMSA-N 0.000 description 2
- NNQDRRUXFJYCCJ-NHCYSSNCSA-N Glu-Pro-Val Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O NNQDRRUXFJYCCJ-NHCYSSNCSA-N 0.000 description 2
- JWNZHMSRZXXGTM-XKBZYTNZSA-N Glu-Ser-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(O)=O JWNZHMSRZXXGTM-XKBZYTNZSA-N 0.000 description 2
- GNPVTZJUUBPZKW-WDSKDSINSA-N Gly-Gln-Ser Chemical compound [H]NCC(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CO)C(O)=O GNPVTZJUUBPZKW-WDSKDSINSA-N 0.000 description 2
- HQRHFUYMGCHHJS-LURJTMIESA-N Gly-Gly-Arg Chemical compound NCC(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)N HQRHFUYMGCHHJS-LURJTMIESA-N 0.000 description 2
- XPJBQTCXPJNIFE-ZETCQYMHSA-N Gly-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)CN XPJBQTCXPJNIFE-ZETCQYMHSA-N 0.000 description 2
- COVXELOAORHTND-LSJOCFKGSA-N Gly-Ile-Val Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C(C)C)C(O)=O COVXELOAORHTND-LSJOCFKGSA-N 0.000 description 2
- TWTPDFFBLQEBOE-IUCAKERBSA-N Gly-Leu-Gln Chemical compound [H]NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCC(N)=O)C(O)=O TWTPDFFBLQEBOE-IUCAKERBSA-N 0.000 description 2
- HFPVRZWORNJRRC-UWVGGRQHSA-N Gly-Pro-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)CN HFPVRZWORNJRRC-UWVGGRQHSA-N 0.000 description 2
- 102100031181 Glyceraldehyde-3-phosphate dehydrogenase Human genes 0.000 description 2
- NYHBQMYGNKIUIF-UUOKFMHZSA-N Guanosine Chemical compound C1=NC=2C(=O)NC(N)=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O NYHBQMYGNKIUIF-UUOKFMHZSA-N 0.000 description 2
- BCZFOHDMCDXPDA-BZSNNMDCSA-N His-Lys-Tyr Chemical compound C1=CC(=CC=C1C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC2=CN=CN2)N)O BCZFOHDMCDXPDA-BZSNNMDCSA-N 0.000 description 2
- 102000003964 Histone deacetylase Human genes 0.000 description 2
- 108090000353 Histone deacetylase Proteins 0.000 description 2
- 102000006947 Histones Human genes 0.000 description 2
- 102100026342 Homeobox protein BarH-like 2 Human genes 0.000 description 2
- 241000282412 Homo Species 0.000 description 2
- 101000897035 Homo sapiens Ectonucleotide pyrophosphatase/phosphodiesterase family member 2 Proteins 0.000 description 2
- 101000766187 Homo sapiens Homeobox protein BarH-like 2 Proteins 0.000 description 2
- 101000595904 Homo sapiens Procollagen-lysine,2-oxoglutarate 5-dioxygenase 1 Proteins 0.000 description 2
- 101000946167 Homo sapiens Transcription factor LBX1 Proteins 0.000 description 2
- 101000940144 Homo sapiens Transcriptional repressor protein YY1 Proteins 0.000 description 2
- XQFRJNBWHJMXHO-RRKCRQDMSA-N IDUR Chemical compound C1[C@H](O)[C@@H](CO)O[C@H]1N1C(=O)NC(=O)C(I)=C1 XQFRJNBWHJMXHO-RRKCRQDMSA-N 0.000 description 2
- JRYQSFOFUFXPTB-RWRJDSDZSA-N Ile-Gln-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N JRYQSFOFUFXPTB-RWRJDSDZSA-N 0.000 description 2
- LPXHYGGZJOCAFR-MNXVOIDGSA-N Ile-Glu-Leu Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CC(C)C)C(=O)O)N LPXHYGGZJOCAFR-MNXVOIDGSA-N 0.000 description 2
- CAHCWMVNBZJVAW-NAKRPEOUSA-N Ile-Pro-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)O)N CAHCWMVNBZJVAW-NAKRPEOUSA-N 0.000 description 2
- QHUREMVLLMNUAX-OSUNSFLBSA-N Ile-Thr-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)O)N QHUREMVLLMNUAX-OSUNSFLBSA-N 0.000 description 2
- APQYGMBHIVXFML-OSUNSFLBSA-N Ile-Val-Thr Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)O)C(=O)O)N APQYGMBHIVXFML-OSUNSFLBSA-N 0.000 description 2
- 208000032578 Inherited retinal disease Diseases 0.000 description 2
- 102100021244 Integral membrane protein GPR180 Human genes 0.000 description 2
- 241000880493 Leptailurus serval Species 0.000 description 2
- DXYBNWJZJVSZAE-GUBZILKMSA-N Leu-Gln-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)N[C@@H](CS)C(=O)O)N DXYBNWJZJVSZAE-GUBZILKMSA-N 0.000 description 2
- HNDWYLYAYNBWMP-AJNGGQMLSA-N Leu-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CC(C)C)N HNDWYLYAYNBWMP-AJNGGQMLSA-N 0.000 description 2
- YOKVEHGYYQEQOP-QWRGUYRKSA-N Leu-Leu-Gly Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)NCC(O)=O YOKVEHGYYQEQOP-QWRGUYRKSA-N 0.000 description 2
- NFLFJGGKOHYZJF-BJDJZHNGSA-N Lys-Ala-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCCN NFLFJGGKOHYZJF-BJDJZHNGSA-N 0.000 description 2
- WSXTWLJHTLRFLW-SRVKXCTJSA-N Lys-Ala-Lys Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCCN)C(O)=O WSXTWLJHTLRFLW-SRVKXCTJSA-N 0.000 description 2
- ZXEUFAVXODIPHC-GUBZILKMSA-N Lys-Glu-Asn Chemical compound NCCCC[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(N)=O)C(O)=O ZXEUFAVXODIPHC-GUBZILKMSA-N 0.000 description 2
- LNMKRJJLEFASGA-BZSNNMDCSA-N Lys-Phe-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC(C)C)C(O)=O LNMKRJJLEFASGA-BZSNNMDCSA-N 0.000 description 2
- IMDJSVBFQKDDEQ-MGHWNKPDSA-N Lys-Tyr-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)NC(=O)[C@H](CCCCN)N IMDJSVBFQKDDEQ-MGHWNKPDSA-N 0.000 description 2
- RIPJMCFGQHGHNP-RHYQMDGZSA-N Lys-Val-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](CCCCN)N)O RIPJMCFGQHGHNP-RHYQMDGZSA-N 0.000 description 2
- KDXKERNSBIXSRK-UHFFFAOYSA-N Lysine Natural products NCCCCC(N)C(O)=O KDXKERNSBIXSRK-UHFFFAOYSA-N 0.000 description 2
- 239000004472 Lysine Substances 0.000 description 2
- 241000407429 Maja Species 0.000 description 2
- PNDCUTDWYVKBHX-IHRRRGAJSA-N Met-Asp-Tyr Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 PNDCUTDWYVKBHX-IHRRRGAJSA-N 0.000 description 2
- 241001465754 Metazoa Species 0.000 description 2
- 108091092878 Microsatellite Proteins 0.000 description 2
- 108091061960 Naked DNA Proteins 0.000 description 2
- 241001274216 Naso Species 0.000 description 2
- 102000007999 Nuclear Proteins Human genes 0.000 description 2
- 108010089610 Nuclear Proteins Proteins 0.000 description 2
- 108091005461 Nucleic proteins Proteins 0.000 description 2
- 238000012408 PCR amplification Methods 0.000 description 2
- YYRCPTVAPLQRNC-ULQDDVLXSA-N Phe-Arg-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CC1=CC=CC=C1 YYRCPTVAPLQRNC-ULQDDVLXSA-N 0.000 description 2
- HNFUGJUZJRYUHN-JSGCOSHPSA-N Phe-Gly-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC1=CC=CC=C1 HNFUGJUZJRYUHN-JSGCOSHPSA-N 0.000 description 2
- NMELOOXSGDRBRU-YUMQZZPRSA-N Pro-Glu-Gly Chemical compound OC(=O)CNC(=O)[C@H](CCC(=O)O)NC(=O)[C@@H]1CCCN1 NMELOOXSGDRBRU-YUMQZZPRSA-N 0.000 description 2
- KHRLUIPIMIQFGT-AVGNSLFASA-N Pro-Val-Leu Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(C)C)C(O)=O KHRLUIPIMIQFGT-AVGNSLFASA-N 0.000 description 2
- 102100035202 Procollagen-lysine,2-oxoglutarate 5-dioxygenase 1 Human genes 0.000 description 2
- 108700008625 Reporter Genes Proteins 0.000 description 2
- 208000032430 Retinal dystrophy Diseases 0.000 description 2
- 108060007030 Ribulose-phosphate 3-epimerase Proteins 0.000 description 2
- 102100039270 Ribulose-phosphate 3-epimerase Human genes 0.000 description 2
- UQFYNFTYDHUIMI-WHFBIAKZSA-N Ser-Gly-Ala Chemical compound OC(=O)[C@H](C)NC(=O)CNC(=O)[C@@H](N)CO UQFYNFTYDHUIMI-WHFBIAKZSA-N 0.000 description 2
- JFWDJFULOLKQFY-QWRGUYRKSA-N Ser-Gly-Phe Chemical compound [H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O JFWDJFULOLKQFY-QWRGUYRKSA-N 0.000 description 2
- QUGRFWPMPVIAPW-IHRRRGAJSA-N Ser-Pro-Phe Chemical compound OC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 QUGRFWPMPVIAPW-IHRRRGAJSA-N 0.000 description 2
- VLMIUSLQONKLDV-HEIBUPTGSA-N Ser-Thr-Thr Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O VLMIUSLQONKLDV-HEIBUPTGSA-N 0.000 description 2
- 208000022758 Sorsby fundus dystrophy Diseases 0.000 description 2
- 241000191967 Staphylococcus aureus Species 0.000 description 2
- 101710172711 Structural protein Proteins 0.000 description 2
- ONNSECRQFSTMCC-XKBZYTNZSA-N Thr-Glu-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=O ONNSECRQFSTMCC-XKBZYTNZSA-N 0.000 description 2
- MGJLBZFUXUGMML-VOAKCMCISA-N Thr-Lys-Lys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCCN)C(=O)O)N)O MGJLBZFUXUGMML-VOAKCMCISA-N 0.000 description 2
- WPSKTVVMQCXPRO-BWBBJGPYSA-N Thr-Ser-Ser Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CO)C(O)=O WPSKTVVMQCXPRO-BWBBJGPYSA-N 0.000 description 2
- LXXCHJKHJYRMIY-FQPOAREZSA-N Thr-Tyr-Ala Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](C)C(O)=O LXXCHJKHJYRMIY-FQPOAREZSA-N 0.000 description 2
- REJRKTOJTCPDPO-IRIUXVKKSA-N Thr-Tyr-Glu Chemical compound [H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(O)=O)C(O)=O REJRKTOJTCPDPO-IRIUXVKKSA-N 0.000 description 2
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 2
- 108700009124 Transcription Initiation Site Proteins 0.000 description 2
- 102100034738 Transcription factor LBX1 Human genes 0.000 description 2
- HHFMNAVFGBYSAT-IGISWZIWSA-N Tyr-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H](CC1=CC=C(C=C1)O)N HHFMNAVFGBYSAT-IGISWZIWSA-N 0.000 description 2
- 102000006943 Uracil-DNA Glycosidase Human genes 0.000 description 2
- 108010072685 Uracil-DNA Glycosidase Proteins 0.000 description 2
- 101710172430 Uracil-DNA glycosylase inhibitor Proteins 0.000 description 2
- LNYOXPDEIZJDEI-NHCYSSNCSA-N Val-Asn-Leu Chemical compound CC(C)C[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)[C@H](C(C)C)N LNYOXPDEIZJDEI-NHCYSSNCSA-N 0.000 description 2
- QSPOLEBZTMESFY-SRVKXCTJSA-N Val-Pro-Val Chemical compound CC(C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=O QSPOLEBZTMESFY-SRVKXCTJSA-N 0.000 description 2
- RTJPAGFXOWEBAI-SRVKXCTJSA-N Val-Val-Arg Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N RTJPAGFXOWEBAI-SRVKXCTJSA-N 0.000 description 2
- SSKKGOWRPNIVDW-AVGNSLFASA-N Val-Val-His Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N SSKKGOWRPNIVDW-AVGNSLFASA-N 0.000 description 2
- JSOXWWFKRJKTMT-WOPDTQHZSA-N Val-Val-Pro Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N1CCC[C@@H]1C(=O)O)N JSOXWWFKRJKTMT-WOPDTQHZSA-N 0.000 description 2
- LLJLBRRXKZTTRD-GUBZILKMSA-N Val-Val-Ser Chemical compound CC(C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(=O)O)N LLJLBRRXKZTTRD-GUBZILKMSA-N 0.000 description 2
- JVGDAEKKZKKZFO-RCWTZXSCSA-N Val-Val-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C(C)C)NC(=O)[C@H](C(C)C)N)O JVGDAEKKZKKZFO-RCWTZXSCSA-N 0.000 description 2
- 102000005789 Vascular Endothelial Growth Factors Human genes 0.000 description 2
- 108010019530 Vascular Endothelial Growth Factors Proteins 0.000 description 2
- 108700005077 Viral Genes Proteins 0.000 description 2
- 230000002159 abnormal effect Effects 0.000 description 2
- OIRDTQYFTABQOQ-KQYNXXCUSA-N adenosine Chemical compound C1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](CO)[C@@H](O)[C@H]1O OIRDTQYFTABQOQ-KQYNXXCUSA-N 0.000 description 2
- 108010076324 alanyl-glycyl-glycine Proteins 0.000 description 2
- 108010086434 alanyl-seryl-glycine Proteins 0.000 description 2
- 108010041407 alanylaspartic acid Proteins 0.000 description 2
- 108010044940 alanylglutamine Proteins 0.000 description 2
- KOSRFJWDECSPRO-UHFFFAOYSA-N alpha-L-glutamyl-L-glutamic acid Natural products OC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=O KOSRFJWDECSPRO-UHFFFAOYSA-N 0.000 description 2
- 230000004075 alteration Effects 0.000 description 2
- 125000000539 amino acid group Chemical group 0.000 description 2
- 230000003321 amplification Effects 0.000 description 2
- 238000002583 angiography Methods 0.000 description 2
- 108010013835 arginine glutamate Proteins 0.000 description 2
- 108010093581 aspartyl-proline Proteins 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 210000004155 blood-retinal barrier Anatomy 0.000 description 2
- 230000004378 blood-retinal barrier Effects 0.000 description 2
- 108010006025 bovine growth hormone Proteins 0.000 description 2
- 210000004556 brain Anatomy 0.000 description 2
- 125000002091 cationic group Chemical group 0.000 description 2
- 210000000170 cell membrane Anatomy 0.000 description 2
- 238000002659 cell therapy Methods 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 239000003153 chemical reaction reagent Substances 0.000 description 2
- 101150100265 cif-1 gene Proteins 0.000 description 2
- 230000024203 complement activation Effects 0.000 description 2
- 238000001514 detection method Methods 0.000 description 2
- 230000004069 differentiation Effects 0.000 description 2
- 238000007847 digital PCR Methods 0.000 description 2
- 239000002552 dosage form Substances 0.000 description 2
- 230000034431 double-strand break repair via homologous recombination Effects 0.000 description 2
- 239000003937 drug carrier Substances 0.000 description 2
- 230000009977 dual effect Effects 0.000 description 2
- 230000008482 dysregulation Effects 0.000 description 2
- 238000002337 electrophoretic mobility shift assay Methods 0.000 description 2
- 210000000981 epithelium Anatomy 0.000 description 2
- 230000000763 evoking effect Effects 0.000 description 2
- 238000010195 expression analysis Methods 0.000 description 2
- 239000013604 expression vector Substances 0.000 description 2
- 201000006321 fundus dystrophy Diseases 0.000 description 2
- 230000004927 fusion Effects 0.000 description 2
- 238000003205 genotyping method Methods 0.000 description 2
- 108010042598 glutamyl-aspartyl-glycine Proteins 0.000 description 2
- 108010049041 glutamylalanine Proteins 0.000 description 2
- 108020004445 glyceraldehyde-3-phosphate dehydrogenase Proteins 0.000 description 2
- 108010051307 glycyl-glycyl-proline Proteins 0.000 description 2
- 108010050848 glycylleucine Proteins 0.000 description 2
- 230000006801 homologous recombination Effects 0.000 description 2
- 238000002744 homologous recombination Methods 0.000 description 2
- 208000017532 inherited retinal dystrophy Diseases 0.000 description 2
- 230000000670 limiting effect Effects 0.000 description 2
- 150000002632 lipids Chemical class 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- 108010009298 lysylglutamic acid Proteins 0.000 description 2
- 108010054155 lysyllysine Proteins 0.000 description 2
- 238000012423 maintenance Methods 0.000 description 2
- 239000011159 matrix material Substances 0.000 description 2
- 238000002493 microarray Methods 0.000 description 2
- 238000010208 microarray analysis Methods 0.000 description 2
- 238000009126 molecular therapy Methods 0.000 description 2
- 230000000877 morphologic effect Effects 0.000 description 2
- 229910052754 neon Inorganic materials 0.000 description 2
- GKAOGPIIYCISHV-UHFFFAOYSA-N neon atom Chemical compound [Ne] GKAOGPIIYCISHV-UHFFFAOYSA-N 0.000 description 2
- 210000005157 neural retina Anatomy 0.000 description 2
- 238000003199 nucleic acid amplification method Methods 0.000 description 2
- 150000002894 organic compounds Chemical class 0.000 description 2
- 238000004806 packaging method and process Methods 0.000 description 2
- 108091008695 photoreceptors Proteins 0.000 description 2
- 102000054765 polymorphisms of proteins Human genes 0.000 description 2
- 230000023603 positive regulation of transcription initiation, DNA-dependent Effects 0.000 description 2
- 230000002265 prevention Effects 0.000 description 2
- 230000001737 promoting effect Effects 0.000 description 2
- 238000003753 real-time PCR Methods 0.000 description 2
- 238000011084 recovery Methods 0.000 description 2
- 230000022532 regulation of transcription, DNA-dependent Effects 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 238000007894 restriction fragment length polymorphism technique Methods 0.000 description 2
- 238000012216 screening Methods 0.000 description 2
- 108010048818 seryl-histidine Proteins 0.000 description 2
- 238000001228 spectrum Methods 0.000 description 2
- RWQNBRDOKXIBIV-UHFFFAOYSA-N thymine Chemical compound CC1=CNC(=O)NC1=O RWQNBRDOKXIBIV-UHFFFAOYSA-N 0.000 description 2
- 230000009466 transformation Effects 0.000 description 2
- 230000007704 transition Effects 0.000 description 2
- 239000013638 trimer Substances 0.000 description 2
- 230000007306 turnover Effects 0.000 description 2
- 241000701447 unidentified baculovirus Species 0.000 description 2
- 230000002792 vascular Effects 0.000 description 2
- 230000004393 visual impairment Effects 0.000 description 2
- 238000011179 visual inspection Methods 0.000 description 2
- JNTMAZFVYNDPLB-PEDHHIEDSA-N (2S,3S)-2-[[[(2S)-1-[(2S,3S)-2-amino-3-methyl-1-oxopentyl]-2-pyrrolidinyl]-oxomethyl]amino]-3-methylpentanoic acid Chemical compound CC[C@H](C)[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(O)=O JNTMAZFVYNDPLB-PEDHHIEDSA-N 0.000 description 1
- CWFMWBHMIMNZLN-NAKRPEOUSA-N (2s)-1-[(2s)-2-[[(2s,3s)-2-amino-3-methylpentanoyl]amino]propanoyl]pyrrolidine-2-carboxylic acid Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O CWFMWBHMIMNZLN-NAKRPEOUSA-N 0.000 description 1
- GZCWLCBFPRFLKL-UHFFFAOYSA-N 1-prop-2-ynoxypropan-2-ol Chemical compound CC(O)COCC#C GZCWLCBFPRFLKL-UHFFFAOYSA-N 0.000 description 1
- 101150028074 2 gene Proteins 0.000 description 1
- VLEIUWBSEKKKFX-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid Chemical compound OCC(N)(CO)CO.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O VLEIUWBSEKKKFX-UHFFFAOYSA-N 0.000 description 1
- MZZYGYNZAOVRTG-UHFFFAOYSA-N 2-hydroxy-n-(1h-1,2,4-triazol-5-yl)benzamide Chemical compound OC1=CC=CC=C1C(=O)NC1=NC=NN1 MZZYGYNZAOVRTG-UHFFFAOYSA-N 0.000 description 1
- OSJPPGNTCRNQQC-UWTATZPHSA-N 3-phospho-D-glyceric acid Chemical compound OC(=O)[C@H](O)COP(O)(O)=O OSJPPGNTCRNQQC-UWTATZPHSA-N 0.000 description 1
- 108020003589 5' Untranslated Regions Proteins 0.000 description 1
- 108010004483 APOBEC-3G Deaminase Proteins 0.000 description 1
- 102100022142 Achaete-scute homolog 1 Human genes 0.000 description 1
- 102000013563 Acid Phosphatase Human genes 0.000 description 1
- 108010051457 Acid Phosphatase Proteins 0.000 description 1
- 101000860090 Acidaminococcus sp. (strain BV3L6) CRISPR-associated endonuclease Cas12a Proteins 0.000 description 1
- 241000186361 Actinobacteria <class> Species 0.000 description 1
- 241001655883 Adeno-associated virus - 1 Species 0.000 description 1
- 241000202702 Adeno-associated virus - 3 Species 0.000 description 1
- 241000580270 Adeno-associated virus - 4 Species 0.000 description 1
- 241001634120 Adeno-associated virus - 5 Species 0.000 description 1
- 241000972680 Adeno-associated virus - 6 Species 0.000 description 1
- 241001164823 Adeno-associated virus - 7 Species 0.000 description 1
- 241001164825 Adeno-associated virus - 8 Species 0.000 description 1
- 241000649045 Adeno-associated virus 10 Species 0.000 description 1
- 241000649046 Adeno-associated virus 11 Species 0.000 description 1
- RLMISHABBKUNFO-WHFBIAKZSA-N Ala-Ala-Gly Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)NCC(O)=O RLMISHABBKUNFO-WHFBIAKZSA-N 0.000 description 1
- CXRCVCURMBFFOL-FXQIFTODSA-N Ala-Ala-Pro Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N1CCC[C@H]1C(O)=O CXRCVCURMBFFOL-FXQIFTODSA-N 0.000 description 1
- YYSWCHMLFJLLBJ-ZLUOBGJFSA-N Ala-Ala-Ser Chemical compound C[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O YYSWCHMLFJLLBJ-ZLUOBGJFSA-N 0.000 description 1
- KQFRUSHJPKXBMB-BHDSKKPTSA-N Ala-Ala-Trp Chemical compound C1=CC=C2C(C[C@H](NC(=O)[C@H](C)NC(=O)[C@@H](N)C)C(O)=O)=CNC2=C1 KQFRUSHJPKXBMB-BHDSKKPTSA-N 0.000 description 1
- FXKNPWNXPQZLES-ZLUOBGJFSA-N Ala-Asn-Ser Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O FXKNPWNXPQZLES-ZLUOBGJFSA-N 0.000 description 1
- LJFNNUBZSZCZFN-WHFBIAKZSA-N Ala-Gly-Cys Chemical compound N[C@@H](C)C(=O)NCC(=O)N[C@@H](CS)C(=O)O LJFNNUBZSZCZFN-WHFBIAKZSA-N 0.000 description 1
- PNALXAODQKTNLV-JBDRJPRFSA-N Ala-Ile-Ala Chemical compound C[C@H](N)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](C)C(O)=O PNALXAODQKTNLV-JBDRJPRFSA-N 0.000 description 1
- CFPQUJZTLUQUTJ-HTFCKZLJSA-N Ala-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@H](C)N CFPQUJZTLUQUTJ-HTFCKZLJSA-N 0.000 description 1
- SUHLZMHFRALVSY-YUMQZZPRSA-N Ala-Lys-Gly Chemical compound NCCCC[C@H](NC(=O)[C@@H](N)C)C(=O)NCC(O)=O SUHLZMHFRALVSY-YUMQZZPRSA-N 0.000 description 1
- XHNLCGXYBXNRIS-BJDJZHNGSA-N Ala-Lys-Ile Chemical compound [H]N[C@@H](C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XHNLCGXYBXNRIS-BJDJZHNGSA-N 0.000 description 1
- IPZQNYYAYVRKKK-FXQIFTODSA-N Ala-Pro-Ala Chemical compound C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O IPZQNYYAYVRKKK-FXQIFTODSA-N 0.000 description 1
- VJVQKGYHIZPSNS-FXQIFTODSA-N Ala-Ser-Arg Chemical compound C[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCN=C(N)N VJVQKGYHIZPSNS-FXQIFTODSA-N 0.000 description 1
- XQNRANMFRPCFFW-GCJQMDKQSA-N Ala-Thr-Asn Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)O)NC(=O)[C@H](C)N)O XQNRANMFRPCFFW-GCJQMDKQSA-N 0.000 description 1
- IETUUAHKCHOQHP-KZVJFYERSA-N Ala-Thr-Val Chemical compound CC(C)[C@H](NC(=O)[C@@H](NC(=O)[C@H](C)N)[C@@H](C)O)C(O)=O IETUUAHKCHOQHP-KZVJFYERSA-N 0.000 description 1
- XAXMJQUMRJAFCH-CQDKDKBSSA-N Ala-Tyr-Lys Chemical compound NCCCC[C@@H](C(O)=O)NC(=O)[C@@H](NC(=O)[C@@H](N)C)CC1=CC=C(O)C=C1 XAXMJQUMRJAFCH-CQDKDKBSSA-N 0.000 description 1
- 241000710929 Alphavirus Species 0.000 description 1
- 241001142141 Aquificae <phylum> Species 0.000 description 1
- UISQLSIBJKEJSS-GUBZILKMSA-N Arg-Arg-Ser Chemical compound NC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CO)C(O)=O UISQLSIBJKEJSS-GUBZILKMSA-N 0.000 description 1
- TTXYKSADPSNOIF-IHRRRGAJSA-N Arg-Asp-Phe Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=O TTXYKSADPSNOIF-IHRRRGAJSA-N 0.000 description 1
- GDVDRMUYICMNFJ-CIUDSAMLSA-N Arg-Cys-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(O)=O)C(O)=O GDVDRMUYICMNFJ-CIUDSAMLSA-N 0.000 description 1
- YWENWUYXQUWRHQ-LPEHRKFASA-N Arg-Cys-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CS)NC(=O)[C@H](CCCN=C(N)N)N)C(=O)O YWENWUYXQUWRHQ-LPEHRKFASA-N 0.000 description 1
- PNIGSVZJNVUVJA-BQBZGAKWSA-N Arg-Gly-Asn Chemical compound NC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=O PNIGSVZJNVUVJA-BQBZGAKWSA-N 0.000 description 1
- YBIAYFFIVAZXPK-AVGNSLFASA-N Arg-His-Arg Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O YBIAYFFIVAZXPK-AVGNSLFASA-N 0.000 description 1
- WMEVEPXNCMKNGH-IHRRRGAJSA-N Arg-Leu-His Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N WMEVEPXNCMKNGH-IHRRRGAJSA-N 0.000 description 1
- YCYXHLZRUSJITQ-SRVKXCTJSA-N Arg-Pro-Pro Chemical compound NC(=N)NCCC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 YCYXHLZRUSJITQ-SRVKXCTJSA-N 0.000 description 1
- VRTWYUYCJGNFES-CIUDSAMLSA-N Arg-Ser-Gln Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(N)=O)C(O)=O VRTWYUYCJGNFES-CIUDSAMLSA-N 0.000 description 1
- ISJWBVIYRBAXEB-CIUDSAMLSA-N Arg-Ser-Glu Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(O)=O ISJWBVIYRBAXEB-CIUDSAMLSA-N 0.000 description 1
- BECXEHHOZNFFFX-IHRRRGAJSA-N Arg-Ser-Tyr Chemical compound [H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=O BECXEHHOZNFFFX-IHRRRGAJSA-N 0.000 description 1
- WOZDCBHUGJVJPL-AVGNSLFASA-N Arg-Val-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCCN=C(N)N)N WOZDCBHUGJVJPL-AVGNSLFASA-N 0.000 description 1
- 101150074374 Ascl2 gene Proteins 0.000 description 1
- UHGUKCOQUNPSKK-CIUDSAMLSA-N Asn-Leu-Cys Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CC(=O)N)N UHGUKCOQUNPSKK-CIUDSAMLSA-N 0.000 description 1
- RZNAMKZJPBQWDJ-SRVKXCTJSA-N Asn-Lys-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CC(=O)N)N RZNAMKZJPBQWDJ-SRVKXCTJSA-N 0.000 description 1
- BKFXFUPYETWGGA-XVSYOHENSA-N Asn-Phe-Thr Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H]([C@@H](C)O)C(O)=O BKFXFUPYETWGGA-XVSYOHENSA-N 0.000 description 1
- MKJBPDLENBUHQU-CIUDSAMLSA-N Asn-Ser-Leu Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(O)=O MKJBPDLENBUHQU-CIUDSAMLSA-N 0.000 description 1
- BCADFFUQHIMQAA-KKHAAJSZSA-N Asn-Thr-Val Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O BCADFFUQHIMQAA-KKHAAJSZSA-N 0.000 description 1
- YSYTWUMRHSFODC-QWRGUYRKSA-N Asn-Tyr-Gly Chemical compound [H]N[C@@H](CC(N)=O)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(O)=O YSYTWUMRHSFODC-QWRGUYRKSA-N 0.000 description 1
- XBQSLMACWDXWLJ-GHCJXIJMSA-N Asp-Ala-Ile Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O XBQSLMACWDXWLJ-GHCJXIJMSA-N 0.000 description 1
- ICAYWNTWHRRAQP-FXQIFTODSA-N Asp-Arg-Cys Chemical compound C(C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CC(=O)O)N)CN=C(N)N ICAYWNTWHRRAQP-FXQIFTODSA-N 0.000 description 1
- SOYOSFXLXYZNRG-CIUDSAMLSA-N Asp-Arg-Gln Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=O SOYOSFXLXYZNRG-CIUDSAMLSA-N 0.000 description 1
- MFMJRYHVLLEMQM-DCAQKATOSA-N Asp-Arg-His Chemical compound C1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CC(=O)O)N MFMJRYHVLLEMQM-DCAQKATOSA-N 0.000 description 1
- VIRHEUMYXXLCBF-WDSKDSINSA-N Asp-Gly-Glu Chemical compound [H]N[C@@H](CC(O)=O)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O VIRHEUMYXXLCBF-WDSKDSINSA-N 0.000 description 1
- QCVXMEHGFUMKCO-YUMQZZPRSA-N Asp-Gly-Leu Chemical compound CC(C)C[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H](N)CC(O)=O QCVXMEHGFUMKCO-YUMQZZPRSA-N 0.000 description 1
- QHHVSXGWLYEAGX-GUBZILKMSA-N Asp-His-Gln Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CC(=O)O)N QHHVSXGWLYEAGX-GUBZILKMSA-N 0.000 description 1
- LDLZOAJRXXBVGF-GMOBBJLQSA-N Asp-Ile-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCSC)C(=O)O)NC(=O)[C@H](CC(=O)O)N LDLZOAJRXXBVGF-GMOBBJLQSA-N 0.000 description 1
- PWAIZUBWHRHYKS-MELADBBJSA-N Asp-Phe-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC2=CC=CC=C2)NC(=O)[C@H](CC(=O)O)N)C(=O)O PWAIZUBWHRHYKS-MELADBBJSA-N 0.000 description 1
- BWJZSLQJNBSUPM-FXQIFTODSA-N Asp-Pro-Asn Chemical compound OC(=O)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CC(N)=O)C(O)=O BWJZSLQJNBSUPM-FXQIFTODSA-N 0.000 description 1
- PDIYGFYAMZZFCW-JIOCBJNQSA-N Asp-Thr-Pro Chemical compound C[C@H]([C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CC(=O)O)N)O PDIYGFYAMZZFCW-JIOCBJNQSA-N 0.000 description 1
- WAEDSQFVZJUHLI-BYULHYEWSA-N Asp-Val-Asp Chemical compound [H]N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=O WAEDSQFVZJUHLI-BYULHYEWSA-N 0.000 description 1
- 241000182988 Assa Species 0.000 description 1
- 241000193830 Bacillus <bacterium> Species 0.000 description 1
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 1
- 239000002126 C01EB10 - Adenosine Substances 0.000 description 1
- 208000010482 CADASIL Diseases 0.000 description 1
- 101150030967 CFH gene Proteins 0.000 description 1
- 238000010446 CRISPR interference Methods 0.000 description 1
- 101100381481 Caenorhabditis elegans baz-2 gene Proteins 0.000 description 1
- 101100290713 Caenorhabditis elegans mef-2 gene Proteins 0.000 description 1
- 108090000565 Capsid Proteins Proteins 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- 208000021910 Cerebral Arterial disease Diseases 0.000 description 1
- 208000033221 Cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy Diseases 0.000 description 1
- 208000033935 Cerebral autosomal dominant arteriopathy-subcortical infarcts-leukoencephalopathy Diseases 0.000 description 1
- 102100023321 Ceruloplasmin Human genes 0.000 description 1
- 238000010196 ChIP-seq analysis Methods 0.000 description 1
- 241000192733 Chloroflexus Species 0.000 description 1
- 102100021615 Class A basic helix-loop-helix protein 15 Human genes 0.000 description 1
- 102000003780 Clusterin Human genes 0.000 description 1
- 108090000197 Clusterin Proteins 0.000 description 1
- 101710094648 Coat protein Proteins 0.000 description 1
- 108010053085 Complement Factor H Proteins 0.000 description 1
- MIKUYHXYGGJMLM-GIMIYPNGSA-N Crotonoside Natural products C1=NC2=C(N)NC(=O)N=C2N1[C@H]1O[C@@H](CO)[C@H](O)[C@@H]1O MIKUYHXYGGJMLM-GIMIYPNGSA-N 0.000 description 1
- ISWAQPWFWKGCAL-ACZMJKKPSA-N Cys-Cys-Glu Chemical compound [H]N[C@@H](CS)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(O)=O)C(O)=O ISWAQPWFWKGCAL-ACZMJKKPSA-N 0.000 description 1
- VIRYODQIWJNWNU-NRPADANISA-N Cys-Glu-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)[C@H](CS)N VIRYODQIWJNWNU-NRPADANISA-N 0.000 description 1
- CVLIHKBUPSFRQP-WHFBIAKZSA-N Cys-Gly-Ala Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](C)C(O)=O CVLIHKBUPSFRQP-WHFBIAKZSA-N 0.000 description 1
- GCDLPNRHPWBKJJ-WDSKDSINSA-N Cys-Gly-Glu Chemical compound [H]N[C@@H](CS)C(=O)NCC(=O)N[C@@H](CCC(O)=O)C(O)=O GCDLPNRHPWBKJJ-WDSKDSINSA-N 0.000 description 1
- KJJASVYBTKRYSN-FXQIFTODSA-N Cys-Pro-Asp Chemical compound C1C[C@H](N(C1)C(=O)[C@H](CS)N)C(=O)N[C@@H](CC(=O)O)C(=O)O KJJASVYBTKRYSN-FXQIFTODSA-N 0.000 description 1
- KSMSFCBQBQPFAD-GUBZILKMSA-N Cys-Pro-Pro Chemical compound SC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1 KSMSFCBQBQPFAD-GUBZILKMSA-N 0.000 description 1
- 102000005381 Cytidine Deaminase Human genes 0.000 description 1
- NYHBQMYGNKIUIF-UHFFFAOYSA-N D-guanosine Natural products C1=2NC(N)=NC(=O)C=2N=CN1C1OC(CO)C(O)C1O NYHBQMYGNKIUIF-UHFFFAOYSA-N 0.000 description 1
- 102100038076 DNA dC->dU-editing enzyme APOBEC-3G Human genes 0.000 description 1
- 230000033616 DNA repair Effects 0.000 description 1
- 238000001712 DNA sequencing Methods 0.000 description 1
- 102000016928 DNA-directed DNA polymerase Human genes 0.000 description 1
- 108010014303 DNA-directed DNA polymerase Proteins 0.000 description 1
- 102000004163 DNA-directed RNA polymerases Human genes 0.000 description 1
- 108090000626 DNA-directed RNA polymerases Proteins 0.000 description 1
- 241001135761 Deltaproteobacteria Species 0.000 description 1
- 206010012689 Diabetic retinopathy Diseases 0.000 description 1
- 101100477411 Dictyostelium discoideum set1 gene Proteins 0.000 description 1
- 206010061818 Disease progression Diseases 0.000 description 1
- 101100229963 Drosophila melanogaster grau gene Proteins 0.000 description 1
- 102000016942 Elastin Human genes 0.000 description 1
- 108010014258 Elastin Proteins 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 241000701959 Escherichia virus Lambda Species 0.000 description 1
- 241000206602 Eukaryota Species 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 108010073385 Fibrin Proteins 0.000 description 1
- 102000009123 Fibrin Human genes 0.000 description 1
- BWGVNKXGVNDBDI-UHFFFAOYSA-N Fibrin monomer Chemical compound CNC(=O)CNC(=O)CN BWGVNKXGVNDBDI-UHFFFAOYSA-N 0.000 description 1
- 101000860092 Francisella tularensis subsp. novicida (strain U112) CRISPR-associated endonuclease Cas12a Proteins 0.000 description 1
- 102100028652 Gamma-enolase Human genes 0.000 description 1
- 101710191797 Gamma-enolase Proteins 0.000 description 1
- 208000003098 Ganglion Cysts Diseases 0.000 description 1
- 108700007698 Genetic Terminator Regions Proteins 0.000 description 1
- 102100039289 Glial fibrillary acidic protein Human genes 0.000 description 1
- 101710193519 Glial fibrillary acidic protein Proteins 0.000 description 1
- MLZRSFQRBDNJON-GUBZILKMSA-N Gln-Ala-Lys Chemical compound C[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](CCC(=O)N)N MLZRSFQRBDNJON-GUBZILKMSA-N 0.000 description 1
- GMGKDVVBSVVKCT-NUMRIWBASA-N Gln-Asn-Thr Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O GMGKDVVBSVVKCT-NUMRIWBASA-N 0.000 description 1
- XSBGUANSZDGULP-IUCAKERBSA-N Gln-Gly-Lys Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CCCCN)C(O)=O XSBGUANSZDGULP-IUCAKERBSA-N 0.000 description 1
- HWEINOMSWQSJDC-SRVKXCTJSA-N Gln-Leu-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O HWEINOMSWQSJDC-SRVKXCTJSA-N 0.000 description 1
- GURIQZQSTBBHRV-SRVKXCTJSA-N Gln-Lys-Arg Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O GURIQZQSTBBHRV-SRVKXCTJSA-N 0.000 description 1
- QFXNFFZTMFHPST-DZKIICNBSA-N Gln-Phe-Val Chemical compound CC(C)[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](CCC(=O)N)N QFXNFFZTMFHPST-DZKIICNBSA-N 0.000 description 1
- HMIXCETWRYDVMO-GUBZILKMSA-N Gln-Pro-Glu Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCC(O)=O)C(O)=O HMIXCETWRYDVMO-GUBZILKMSA-N 0.000 description 1
- HLRLXVPRJJITSK-IFFSRLJSSA-N Gln-Thr-Val Chemical compound [H]N[C@@H](CCC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(O)=O HLRLXVPRJJITSK-IFFSRLJSSA-N 0.000 description 1
- KKCUFHUTMKQQCF-SRVKXCTJSA-N Glu-Arg-Leu Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(O)=O KKCUFHUTMKQQCF-SRVKXCTJSA-N 0.000 description 1
- DSPQRJXOIXHOHK-WDSKDSINSA-N Glu-Asp-Gly Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(O)=O)C(=O)NCC(O)=O DSPQRJXOIXHOHK-WDSKDSINSA-N 0.000 description 1
- IESFZVCAVACGPH-PEFMBERDSA-N Glu-Asp-Ile Chemical compound CC[C@H](C)[C@@H](C(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](N)CCC(O)=O IESFZVCAVACGPH-PEFMBERDSA-N 0.000 description 1
- CKOFNWCLWRYUHK-XHNCKOQMSA-N Glu-Asp-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)O)NC(=O)[C@H](CCC(=O)O)N)C(=O)O CKOFNWCLWRYUHK-XHNCKOQMSA-N 0.000 description 1
- OPAINBJQDQTGJY-JGVFFNPUSA-N Glu-Gly-Pro Chemical compound C1C[C@@H](N(C1)C(=O)CNC(=O)[C@H](CCC(=O)O)N)C(=O)O OPAINBJQDQTGJY-JGVFFNPUSA-N 0.000 description 1
- COSBSYQVPSODFX-GUBZILKMSA-N Glu-His-Cys Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CCC(=O)O)N COSBSYQVPSODFX-GUBZILKMSA-N 0.000 description 1
- VSRCAOIHMGCIJK-SRVKXCTJSA-N Glu-Leu-Arg Chemical compound OC(=O)CC[C@H](N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCN=C(N)N)C(O)=O VSRCAOIHMGCIJK-SRVKXCTJSA-N 0.000 description 1
- VGBSZQSKQRMLHD-MNXVOIDGSA-N Glu-Leu-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O VGBSZQSKQRMLHD-MNXVOIDGSA-N 0.000 description 1
- UGSVSNXPJJDJKL-SDDRHHMPSA-N Glu-Leu-Pro Chemical compound CC(C)C[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](CCC(=O)O)N UGSVSNXPJJDJKL-SDDRHHMPSA-N 0.000 description 1
- NJCALAAIGREHDR-WDCWCFNPSA-N Glu-Leu-Thr Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H]([C@@H](C)O)C(O)=O NJCALAAIGREHDR-WDCWCFNPSA-N 0.000 description 1
- YKBUCXNNBYZYAY-MNXVOIDGSA-N Glu-Lys-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O YKBUCXNNBYZYAY-MNXVOIDGSA-N 0.000 description 1
- QJVZSVUYZFYLFQ-CIUDSAMLSA-N Glu-Pro-Ala Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C)C(O)=O QJVZSVUYZFYLFQ-CIUDSAMLSA-N 0.000 description 1
- FGGKGJHCVMYGCD-UKJIMTQDSA-N Glu-Val-Ile Chemical compound [H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H]([C@@H](C)CC)C(O)=O FGGKGJHCVMYGCD-UKJIMTQDSA-N 0.000 description 1
- NZAFOTBEULLEQB-WDSKDSINSA-N Gly-Asn-Glu Chemical compound C(CC(=O)O)[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)N)NC(=O)CN NZAFOTBEULLEQB-WDSKDSINSA-N 0.000 description 1
- XRTDOIOIBMAXCT-NKWVEPMBSA-N Gly-Asn-Pro Chemical compound C1C[C@@H](N(C1)C(=O)[C@H](CC(=O)N)NC(=O)CN)C(=O)O XRTDOIOIBMAXCT-NKWVEPMBSA-N 0.000 description 1
- XTQFHTHIAKKCTM-YFKPBYRVSA-N Gly-Glu-Gly Chemical compound NCC(=O)N[C@@H](CCC(O)=O)C(=O)NCC(O)=O XTQFHTHIAKKCTM-YFKPBYRVSA-N 0.000 description 1
- LHRXAHLCRMQBGJ-RYUDHWBXSA-N Gly-Glu-Phe Chemical compound C1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CCC(=O)O)NC(=O)CN LHRXAHLCRMQBGJ-RYUDHWBXSA-N 0.000 description 1
- JSNNHGHYGYMVCK-XVKPBYJWSA-N Gly-Glu-Val Chemical compound [H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O JSNNHGHYGYMVCK-XVKPBYJWSA-N 0.000 description 1
- UQJNXZSSGQIPIQ-FBCQKBJTSA-N Gly-Gly-Thr Chemical compound C[C@@H](O)[C@@H](C(O)=O)NC(=O)CNC(=O)CN UQJNXZSSGQIPIQ-FBCQKBJTSA-N 0.000 description 1
- OLPPXYMMIARYAL-QMMMGPOBSA-N Gly-Gly-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)CNC(=O)CN OLPPXYMMIARYAL-QMMMGPOBSA-N 0.000 description 1
- SXJHOPPTOJACOA-QXEWZRGKSA-N Gly-Ile-Arg Chemical compound NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@H](C(O)=O)CCCN=C(N)N SXJHOPPTOJACOA-QXEWZRGKSA-N 0.000 description 1
- NSTUFLGQJCOCDL-UWVGGRQHSA-N Gly-Leu-Arg Chemical compound NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N NSTUFLGQJCOCDL-UWVGGRQHSA-N 0.000 description 1
- GMTXWRIDLGTVFC-IUCAKERBSA-N Gly-Lys-Glu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(O)=O GMTXWRIDLGTVFC-IUCAKERBSA-N 0.000 description 1
- MHXKHKWHPNETGG-QWRGUYRKSA-N Gly-Lys-Leu Chemical compound [H]NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(C)C)C(O)=O MHXKHKWHPNETGG-QWRGUYRKSA-N 0.000 description 1
- QSQXZZCGPXQBPP-BQBZGAKWSA-N Gly-Pro-Cys Chemical compound C1C[C@H](N(C1)C(=O)CN)C(=O)N[C@@H](CS)C(=O)O QSQXZZCGPXQBPP-BQBZGAKWSA-N 0.000 description 1
- 102100021181 Golgi phosphoprotein 3 Human genes 0.000 description 1
- 208000034508 Haemangioma of retina Diseases 0.000 description 1
- 102100021888 Helix-loop-helix protein 1 Human genes 0.000 description 1
- 229920000209 Hexadimethrine bromide Polymers 0.000 description 1
- LYSMQLXUCAKELQ-DCAQKATOSA-N His-Asp-Arg Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N LYSMQLXUCAKELQ-DCAQKATOSA-N 0.000 description 1
- CYHWWHKRCKHYGQ-GUBZILKMSA-N His-Cys-Glu Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(=O)O)C(=O)O)N CYHWWHKRCKHYGQ-GUBZILKMSA-N 0.000 description 1
- 102100023823 Homeobox protein EMX1 Human genes 0.000 description 1
- 102100028404 Homeobox protein Hox-B4 Human genes 0.000 description 1
- 102100040228 Homeobox protein Hox-D3 Human genes 0.000 description 1
- 102100021086 Homeobox protein Hox-D4 Human genes 0.000 description 1
- 102100033791 Homeobox protein aristaless-like 3 Human genes 0.000 description 1
- 102000010029 Homer Scaffolding Proteins Human genes 0.000 description 1
- 108010077223 Homer Scaffolding Proteins Proteins 0.000 description 1
- 101000901099 Homo sapiens Achaete-scute homolog 1 Proteins 0.000 description 1
- 101100178820 Homo sapiens HTRA1 gene Proteins 0.000 description 1
- 101000897691 Homo sapiens Helix-loop-helix protein 1 Proteins 0.000 description 1
- 101001048956 Homo sapiens Homeobox protein EMX1 Proteins 0.000 description 1
- 101000839788 Homo sapiens Homeobox protein Hox-B4 Proteins 0.000 description 1
- 101001037158 Homo sapiens Homeobox protein Hox-D3 Proteins 0.000 description 1
- 101001041136 Homo sapiens Homeobox protein Hox-D4 Proteins 0.000 description 1
- 101000779611 Homo sapiens Homeobox protein aristaless-like 3 Proteins 0.000 description 1
- 101000619927 Homo sapiens LIM/homeobox protein Lhx9 Proteins 0.000 description 1
- 101000576323 Homo sapiens Motor neuron and pancreas homeobox protein 1 Proteins 0.000 description 1
- 101000738966 Homo sapiens POU domain, class 6, transcription factor 1 Proteins 0.000 description 1
- 101000687346 Homo sapiens PR domain zinc finger protein 2 Proteins 0.000 description 1
- 101001069723 Homo sapiens Paired mesoderm homeobox protein 2 Proteins 0.000 description 1
- 101000609255 Homo sapiens Plasminogen activator inhibitor 1 Proteins 0.000 description 1
- 101000670189 Homo sapiens Ribulose-phosphate 3-epimerase Proteins 0.000 description 1
- 101000658622 Homo sapiens Testis-specific Y-encoded-like protein 2 Proteins 0.000 description 1
- 101001041525 Homo sapiens Transcription factor 12 Proteins 0.000 description 1
- 101000750380 Homo sapiens Ventral anterior homeobox 1 Proteins 0.000 description 1
- 101000854936 Homo sapiens Visual system homeobox 1 Proteins 0.000 description 1
- 101000854931 Homo sapiens Visual system homeobox 2 Proteins 0.000 description 1
- 101000760226 Homo sapiens Zinc finger protein 333 Proteins 0.000 description 1
- 241000701044 Human gammaherpesvirus 4 Species 0.000 description 1
- UNDGQKWQNSTPPW-CYDGBPFRSA-N Ile-Arg-Met Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCSC)C(=O)O)N UNDGQKWQNSTPPW-CYDGBPFRSA-N 0.000 description 1
- LEDRIAHEWDJRMF-CFMVVWHZSA-N Ile-Asn-Tyr Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1 LEDRIAHEWDJRMF-CFMVVWHZSA-N 0.000 description 1
- JQLFYZMEXFNRFS-DJFWLOJKSA-N Ile-Asp-His Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)N JQLFYZMEXFNRFS-DJFWLOJKSA-N 0.000 description 1
- MQFGXJNSUJTXDT-QSFUFRPTSA-N Ile-Gly-Ile Chemical compound N[C@@H]([C@@H](C)CC)C(=O)NCC(=O)N[C@@H]([C@@H](C)CC)C(=O)O MQFGXJNSUJTXDT-QSFUFRPTSA-N 0.000 description 1
- NZGTYCMLUGYMCV-XUXIUFHCSA-N Ile-Lys-Arg Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)N NZGTYCMLUGYMCV-XUXIUFHCSA-N 0.000 description 1
- UDBPXJNOEWDBDF-XUXIUFHCSA-N Ile-Lys-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(=O)O)N UDBPXJNOEWDBDF-XUXIUFHCSA-N 0.000 description 1
- AGGIYSLVUKVOPT-HTFCKZLJSA-N Ile-Ser-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)N AGGIYSLVUKVOPT-HTFCKZLJSA-N 0.000 description 1
- MUFXDFWAJSPHIQ-XDTLVQLUSA-N Ile-Tyr Chemical compound CC[C@H](C)[C@H]([NH3+])C(=O)N[C@H](C([O-])=O)CC1=CC=C(O)C=C1 MUFXDFWAJSPHIQ-XDTLVQLUSA-N 0.000 description 1
- UYODHPPSCXBNCS-XUXIUFHCSA-N Ile-Val-Leu Chemical compound CC[C@H](C)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CC(C)C UYODHPPSCXBNCS-XUXIUFHCSA-N 0.000 description 1
- JZBVBOKASHNXAD-NAKRPEOUSA-N Ile-Val-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CO)C(=O)O)N JZBVBOKASHNXAD-NAKRPEOUSA-N 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 229930010555 Inosine Natural products 0.000 description 1
- UGQMRVRMYYASKQ-KQYNXXCUSA-N Inosine Chemical compound O[C@@H]1[C@H](O)[C@@H](CO)O[C@H]1N1C2=NC=NC(O)=C2N=C1 UGQMRVRMYYASKQ-KQYNXXCUSA-N 0.000 description 1
- 108091092195 Intron Proteins 0.000 description 1
- PMGDADKJMCOXHX-UHFFFAOYSA-N L-Arginyl-L-glutamin-acetat Natural products NC(=N)NCCCC(N)C(=O)NC(CCC(N)=O)C(O)=O PMGDADKJMCOXHX-UHFFFAOYSA-N 0.000 description 1
- IBMVEYRWAWIOTN-UHFFFAOYSA-N L-Leucyl-L-Arginyl-L-Proline Natural products CC(C)CC(N)C(=O)NC(CCCN=C(N)N)C(=O)N1CCCC1C(O)=O IBMVEYRWAWIOTN-UHFFFAOYSA-N 0.000 description 1
- 102100022141 LIM/homeobox protein Lhx9 Human genes 0.000 description 1
- 241001112693 Lachnospiraceae Species 0.000 description 1
- IBMVEYRWAWIOTN-RWMBFGLXSA-N Leu-Arg-Pro Chemical compound CC(C)C[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N1CCC[C@@H]1C(O)=O IBMVEYRWAWIOTN-RWMBFGLXSA-N 0.000 description 1
- QKIBIXAQKAFZGL-GUBZILKMSA-N Leu-Cys-Gln Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(N)=O)C(O)=O QKIBIXAQKAFZGL-GUBZILKMSA-N 0.000 description 1
- HYIFFZAQXPUEAU-QWRGUYRKSA-N Leu-Gly-Leu Chemical compound CC(C)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC(C)C HYIFFZAQXPUEAU-QWRGUYRKSA-N 0.000 description 1
- RZXLZBIUTDQHJQ-SRVKXCTJSA-N Leu-Lys-Asp Chemical compound [H]N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(O)=O RZXLZBIUTDQHJQ-SRVKXCTJSA-N 0.000 description 1
- XXXXOVFBXRERQL-ULQDDVLXSA-N Leu-Pro-Phe Chemical compound CC(C)C[C@H](N)C(=O)N1CCC[C@H]1C(=O)N[C@H](C(O)=O)CC1=CC=CC=C1 XXXXOVFBXRERQL-ULQDDVLXSA-N 0.000 description 1
- JGAMUXDWYSXYLM-SRVKXCTJSA-N Lys-Arg-Glu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=O JGAMUXDWYSXYLM-SRVKXCTJSA-N 0.000 description 1
- GKFNXYMAMKJSKD-NHCYSSNCSA-N Lys-Asp-Val Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O GKFNXYMAMKJSKD-NHCYSSNCSA-N 0.000 description 1
- DCRWPTBMWMGADO-AVGNSLFASA-N Lys-Glu-Leu Chemical compound [H]N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CC(C)C)C(O)=O DCRWPTBMWMGADO-AVGNSLFASA-N 0.000 description 1
- KNKJPYAZQUFLQK-IHRRRGAJSA-N Lys-His-Arg Chemical compound C1=C(NC=N1)C[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NC(=O)[C@H](CCCCN)N KNKJPYAZQUFLQK-IHRRRGAJSA-N 0.000 description 1
- PPNCMJARTHYNEC-MEYUZBJRSA-N Lys-Tyr-Thr Chemical compound NCCCC[C@H](N)C(=O)N[C@H](C(=O)N[C@@H]([C@H](O)C)C(O)=O)CC1=CC=C(O)C=C1 PPNCMJARTHYNEC-MEYUZBJRSA-N 0.000 description 1
- 101150094019 MYOG gene Proteins 0.000 description 1
- 206010025421 Macule Diseases 0.000 description 1
- 101710125418 Major capsid protein Proteins 0.000 description 1
- 241000124008 Mammalia Species 0.000 description 1
- WXHHTBVYQOSYSL-FXQIFTODSA-N Met-Ala-Ser Chemical compound CSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CO)C(O)=O WXHHTBVYQOSYSL-FXQIFTODSA-N 0.000 description 1
- UOENBSHXYCHSAU-YUMQZZPRSA-N Met-Gln-Gly Chemical compound [H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=O UOENBSHXYCHSAU-YUMQZZPRSA-N 0.000 description 1
- XOFDBXYPKZUAAM-GUBZILKMSA-N Met-Met-Ser Chemical compound CSCC[C@@H](C(=O)N[C@@H](CCSC)C(=O)N[C@@H](CO)C(=O)O)N XOFDBXYPKZUAAM-GUBZILKMSA-N 0.000 description 1
- HLZORBMOISUNIV-DCAQKATOSA-N Met-Ser-Leu Chemical compound CSCC[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CC(C)C HLZORBMOISUNIV-DCAQKATOSA-N 0.000 description 1
- 102000006890 Methyl-CpG-Binding Protein 2 Human genes 0.000 description 1
- 108010072388 Methyl-CpG-Binding Protein 2 Proteins 0.000 description 1
- 241001590997 Moolgarda engeli Species 0.000 description 1
- 102100025170 Motor neuron and pancreas homeobox protein 1 Human genes 0.000 description 1
- 241000699666 Mus <mouse, genus> Species 0.000 description 1
- 101100219625 Mus musculus Casd1 gene Proteins 0.000 description 1
- 241000699670 Mus sp. Species 0.000 description 1
- YBAFDPFAUTYYRW-UHFFFAOYSA-N N-L-alpha-glutamyl-L-leucine Natural products CC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=O YBAFDPFAUTYYRW-UHFFFAOYSA-N 0.000 description 1
- XMBSYZWANAQXEV-UHFFFAOYSA-N N-alpha-L-glutamyl-L-phenylalanine Natural products OC(=O)CCC(N)C(=O)NC(C(O)=O)CC1=CC=CC=C1 XMBSYZWANAQXEV-UHFFFAOYSA-N 0.000 description 1
- 108091007491 NSP3 Papain-like protease domains Proteins 0.000 description 1
- 241000588650 Neisseria meningitidis Species 0.000 description 1
- 206010028980 Neoplasm Diseases 0.000 description 1
- 206010029113 Neovascularisation Diseases 0.000 description 1
- 101710141454 Nucleoprotein Proteins 0.000 description 1
- 108700026244 Open Reading Frames Proteins 0.000 description 1
- 238000010222 PCR analysis Methods 0.000 description 1
- 101150012394 PHO5 gene Proteins 0.000 description 1
- 102100037483 POU domain, class 6, transcription factor 1 Human genes 0.000 description 1
- 102100024885 PR domain zinc finger protein 2 Human genes 0.000 description 1
- 102100033829 Paired mesoderm homeobox protein 2 Human genes 0.000 description 1
- 102100041030 Pancreas/duodenum homeobox protein 1 Human genes 0.000 description 1
- 102000035195 Peptidases Human genes 0.000 description 1
- 108091005804 Peptidases Proteins 0.000 description 1
- 108010077524 Peptide Elongation Factor 1 Proteins 0.000 description 1
- 108010067902 Peptide Library Proteins 0.000 description 1
- DFEVBOYEUQJGER-JURCDPSOSA-N Phe-Ala-Ile Chemical compound N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)O DFEVBOYEUQJGER-JURCDPSOSA-N 0.000 description 1
- 108091000080 Phosphotransferase Proteins 0.000 description 1
- 102100039418 Plasminogen activator inhibitor 1 Human genes 0.000 description 1
- 206010063381 Polypoidal choroidal vasculopathy Diseases 0.000 description 1
- VXCHGLYSIOOZIS-GUBZILKMSA-N Pro-Ala-Arg Chemical compound NC(N)=NCCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 VXCHGLYSIOOZIS-GUBZILKMSA-N 0.000 description 1
- XQLBWXHVZVBNJM-FXQIFTODSA-N Pro-Ala-Ser Chemical compound OC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H]1CCCN1 XQLBWXHVZVBNJM-FXQIFTODSA-N 0.000 description 1
- SFECXGVELZFBFJ-VEVYYDQMSA-N Pro-Asp-Thr Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)O)C(O)=O SFECXGVELZFBFJ-VEVYYDQMSA-N 0.000 description 1
- SKICPQLTOXGWGO-GARJFASQSA-N Pro-Gln-Pro Chemical compound C1C[C@H](NC1)C(=O)N[C@@H](CCC(=O)N)C(=O)N2CCC[C@@H]2C(=O)O SKICPQLTOXGWGO-GARJFASQSA-N 0.000 description 1
- VYWNORHENYEQDW-YUMQZZPRSA-N Pro-Gly-Glu Chemical compound OC(=O)CC[C@@H](C(O)=O)NC(=O)CNC(=O)[C@@H]1CCCN1 VYWNORHENYEQDW-YUMQZZPRSA-N 0.000 description 1
- FMLRRBDLBJLJIK-DCAQKATOSA-N Pro-Leu-Ala Chemical compound OC(=O)[C@H](C)NC(=O)[C@H](CC(C)C)NC(=O)[C@@H]1CCCN1 FMLRRBDLBJLJIK-DCAQKATOSA-N 0.000 description 1
- GFHXZNVJIKMAGO-IHRRRGAJSA-N Pro-Phe-Ser Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CO)C(O)=O GFHXZNVJIKMAGO-IHRRRGAJSA-N 0.000 description 1
- KBUAPZAZPWNYSW-SRVKXCTJSA-N Pro-Pro-Val Chemical compound CC(C)[C@@H](C(O)=O)NC(=O)[C@@H]1CCCN1C(=O)[C@H]1NCCC1 KBUAPZAZPWNYSW-SRVKXCTJSA-N 0.000 description 1
- GMJDSFYVTAMIBF-FXQIFTODSA-N Pro-Ser-Asp Chemical compound [H]N1CCC[C@H]1C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(O)=O GMJDSFYVTAMIBF-FXQIFTODSA-N 0.000 description 1
- 101710083689 Probable capsid protein Proteins 0.000 description 1
- 239000004365 Protease Substances 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 108010067787 Proteoglycans Proteins 0.000 description 1
- 102000016611 Proteoglycans Human genes 0.000 description 1
- 108010026552 Proteome Proteins 0.000 description 1
- 101710183548 Pyridoxal 5'-phosphate synthase subunit PdxS Proteins 0.000 description 1
- 238000010357 RNA editing Methods 0.000 description 1
- 230000026279 RNA modification Effects 0.000 description 1
- 101100372762 Rattus norvegicus Flt1 gene Proteins 0.000 description 1
- 102000003661 Ribonuclease III Human genes 0.000 description 1
- 108010057163 Ribonuclease III Proteins 0.000 description 1
- 238000012300 Sequence Analysis Methods 0.000 description 1
- RDFQNDHEHVSONI-ZLUOBGJFSA-N Ser-Asn-Ser Chemical compound OC[C@H](N)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H](CO)C(O)=O RDFQNDHEHVSONI-ZLUOBGJFSA-N 0.000 description 1
- CTRHXXXHUJTTRZ-ZLUOBGJFSA-N Ser-Asp-Cys Chemical compound C([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](CO)N)C(=O)O CTRHXXXHUJTTRZ-ZLUOBGJFSA-N 0.000 description 1
- SWSRFJZZMNLMLY-ZKWXMUAHSA-N Ser-Asp-Val Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=O SWSRFJZZMNLMLY-ZKWXMUAHSA-N 0.000 description 1
- LQESNKGTTNHZPZ-GHCJXIJMSA-N Ser-Ile-Asn Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(N)=O)C(O)=O LQESNKGTTNHZPZ-GHCJXIJMSA-N 0.000 description 1
- QYSFWUIXDFJUDW-DCAQKATOSA-N Ser-Leu-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O QYSFWUIXDFJUDW-DCAQKATOSA-N 0.000 description 1
- GJFYFGOEWLDQGW-GUBZILKMSA-N Ser-Leu-Gln Chemical compound CC(C)C[C@@H](C(=O)N[C@@H](CCC(=O)N)C(=O)O)NC(=O)[C@H](CO)N GJFYFGOEWLDQGW-GUBZILKMSA-N 0.000 description 1
- NNFMANHDYSVNIO-DCAQKATOSA-N Ser-Lys-Arg Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=O NNFMANHDYSVNIO-DCAQKATOSA-N 0.000 description 1
- NQZFFLBPNDLTPO-DLOVCJGASA-N Ser-Phe-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CC1=CC=CC=C1)NC(=O)[C@H](CO)N NQZFFLBPNDLTPO-DLOVCJGASA-N 0.000 description 1
- HHJFMHQYEAAOBM-ZLUOBGJFSA-N Ser-Ser-Ala Chemical compound [H]N[C@@H](CO)C(=O)N[C@@H](CO)C(=O)N[C@@H](C)C(O)=O HHJFMHQYEAAOBM-ZLUOBGJFSA-N 0.000 description 1
- 108020004682 Single-Stranded DNA Proteins 0.000 description 1
- 108091027967 Small hairpin RNA Proteins 0.000 description 1
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 1
- 241000589970 Spirochaetales Species 0.000 description 1
- 241001180364 Spirochaetes Species 0.000 description 1
- 241000194017 Streptococcus Species 0.000 description 1
- 241000194020 Streptococcus thermophilus Species 0.000 description 1
- 102100028706 Synaptophysin Human genes 0.000 description 1
- 108090001076 Synaptophysin Proteins 0.000 description 1
- 208000005400 Synovial Cyst Diseases 0.000 description 1
- 101710137500 T7 RNA polymerase Proteins 0.000 description 1
- 108700026226 TATA Box Proteins 0.000 description 1
- 102100034917 Testis-specific Y-encoded-like protein 2 Human genes 0.000 description 1
- 241000223257 Thermomyces Species 0.000 description 1
- XGFYGMKZKFRGAI-RCWTZXSCSA-N Thr-Val-Arg Chemical compound C[C@@H](O)[C@H](N)C(=O)N[C@@H](C(C)C)C(=O)N[C@H](C(O)=O)CCCN=C(N)N XGFYGMKZKFRGAI-RCWTZXSCSA-N 0.000 description 1
- 101001023030 Toxoplasma gondii Myosin-D Proteins 0.000 description 1
- 108010073062 Transcription Activator-Like Effectors Proteins 0.000 description 1
- 102100037116 Transcription elongation factor 1 homolog Human genes 0.000 description 1
- 102100021123 Transcription factor 12 Human genes 0.000 description 1
- 101710195626 Transcriptional activator protein Proteins 0.000 description 1
- 108010074506 Transfer Factor Proteins 0.000 description 1
- 101800005109 Triakontatetraneuropeptide Proteins 0.000 description 1
- 208000026487 Triploidy Diseases 0.000 description 1
- SAKLWFSRZTZQAJ-GQGQLFGLSA-N Trp-Ile-Ser Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CO)C(=O)O)NC(=O)[C@H](CC1=CNC2=CC=CC=C21)N SAKLWFSRZTZQAJ-GQGQLFGLSA-N 0.000 description 1
- 108020004417 Untranslated RNA Proteins 0.000 description 1
- 102000039634 Untranslated RNA Human genes 0.000 description 1
- SLLKXDSRVAOREO-KZVJFYERSA-N Val-Ala-Thr Chemical compound C[C@H]([C@@H](C(=O)O)NC(=O)[C@H](C)NC(=O)[C@H](C(C)C)N)O SLLKXDSRVAOREO-KZVJFYERSA-N 0.000 description 1
- JIODCDXKCJRMEH-NHCYSSNCSA-N Val-Arg-Gln Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CCC(=O)N)C(=O)O)N JIODCDXKCJRMEH-NHCYSSNCSA-N 0.000 description 1
- CGGVNFJRZJUVAE-BYULHYEWSA-N Val-Asp-Asn Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)N)C(=O)O)N CGGVNFJRZJUVAE-BYULHYEWSA-N 0.000 description 1
- FPCIBLUVDNXPJO-XPUUQOCRSA-N Val-Cys-Gly Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CS)C(=O)NCC(O)=O FPCIBLUVDNXPJO-XPUUQOCRSA-N 0.000 description 1
- ZXAGTABZUOMUDO-GVXVVHGQSA-N Val-Glu-Lys Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCCCN)C(=O)O)N ZXAGTABZUOMUDO-GVXVVHGQSA-N 0.000 description 1
- UKEVLVBHRKWECS-LSJOCFKGSA-N Val-Ile-Gly Chemical compound CC[C@H](C)[C@@H](C(=O)NCC(=O)O)NC(=O)[C@H](C(C)C)N UKEVLVBHRKWECS-LSJOCFKGSA-N 0.000 description 1
- VHRLUTIMTDOVCG-PEDHHIEDSA-N Val-Ile-Ile Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)O)NC(=O)[C@H](C(C)C)N VHRLUTIMTDOVCG-PEDHHIEDSA-N 0.000 description 1
- SDUBQHUJJWQTEU-XUXIUFHCSA-N Val-Ile-Lys Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](CCCCN)C(=O)O)NC(=O)[C@H](C(C)C)N SDUBQHUJJWQTEU-XUXIUFHCSA-N 0.000 description 1
- JZWZACGUZVCQPS-RNJOBUHISA-N Val-Ile-Pro Chemical compound CC[C@H](C)[C@@H](C(=O)N1CCC[C@@H]1C(=O)O)NC(=O)[C@H](C(C)C)N JZWZACGUZVCQPS-RNJOBUHISA-N 0.000 description 1
- DJQIUOKSNRBTSV-CYDGBPFRSA-N Val-Ile-Val Chemical compound CC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H](C(C)C)N DJQIUOKSNRBTSV-CYDGBPFRSA-N 0.000 description 1
- RWOGENDAOGMHLX-DCAQKATOSA-N Val-Lys-Ala Chemical compound C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](C(C)C)N RWOGENDAOGMHLX-DCAQKATOSA-N 0.000 description 1
- VPGCVZRRBYOGCD-AVGNSLFASA-N Val-Lys-Val Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C(C)C)C(O)=O VPGCVZRRBYOGCD-AVGNSLFASA-N 0.000 description 1
- BGXVHVMJZCSOCA-AVGNSLFASA-N Val-Pro-Lys Chemical compound CC(C)[C@@H](C(=O)N1CCC[C@H]1C(=O)N[C@@H](CCCCN)C(=O)O)N BGXVHVMJZCSOCA-AVGNSLFASA-N 0.000 description 1
- LTTQCQRTSHJPPL-ZKWXMUAHSA-N Val-Ser-Asp Chemical compound CC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N[C@@H](CC(=O)O)C(=O)O)N LTTQCQRTSHJPPL-ZKWXMUAHSA-N 0.000 description 1
- VIKZGAUAKQZDOF-NRPADANISA-N Val-Ser-Glu Chemical compound CC(C)[C@H](N)C(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCC(O)=O VIKZGAUAKQZDOF-NRPADANISA-N 0.000 description 1
- BZDGLJPROOOUOZ-XGEHTFHBSA-N Val-Thr-Cys Chemical compound C[C@H]([C@@H](C(=O)N[C@@H](CS)C(=O)O)NC(=O)[C@H](C(C)C)N)O BZDGLJPROOOUOZ-XGEHTFHBSA-N 0.000 description 1
- JAIZPWVHPQRYOU-ZJDVBMNYSA-N Val-Thr-Thr Chemical compound C[C@H]([C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)O)NC(=O)[C@H](C(C)C)N)O JAIZPWVHPQRYOU-ZJDVBMNYSA-N 0.000 description 1
- 201000004810 Vascular dementia Diseases 0.000 description 1
- 102100021166 Ventral anterior homeobox 1 Human genes 0.000 description 1
- 241001261005 Verrucomicrobia Species 0.000 description 1
- 241001661641 Verrucosa Species 0.000 description 1
- 241000251539 Vertebrata <Metazoa> Species 0.000 description 1
- 102100020673 Visual system homeobox 1 Human genes 0.000 description 1
- 102100020676 Visual system homeobox 2 Human genes 0.000 description 1
- 108010031318 Vitronectin Proteins 0.000 description 1
- 102100035140 Vitronectin Human genes 0.000 description 1
- 208000000208 Wet Macular Degeneration Diseases 0.000 description 1
- PTFCDOFLOPIGGS-UHFFFAOYSA-N Zinc dication Chemical compound [Zn+2] PTFCDOFLOPIGGS-UHFFFAOYSA-N 0.000 description 1
- 101710185494 Zinc finger protein Proteins 0.000 description 1
- 102100024772 Zinc finger protein 333 Human genes 0.000 description 1
- 102100023597 Zinc finger protein 816 Human genes 0.000 description 1
- 230000010933 acylation Effects 0.000 description 1
- 238000005917 acylation reaction Methods 0.000 description 1
- 229960005305 adenosine Drugs 0.000 description 1
- 230000002411 adverse Effects 0.000 description 1
- 230000029936 alkylation Effects 0.000 description 1
- 238000005804 alkylation reaction Methods 0.000 description 1
- 230000003281 allosteric effect Effects 0.000 description 1
- 230000009435 amidation Effects 0.000 description 1
- 238000007112 amidation reaction Methods 0.000 description 1
- 230000001775 anti-pathogenic effect Effects 0.000 description 1
- 108010008355 arginyl-glutamine Proteins 0.000 description 1
- 108010029539 arginyl-prolyl-proline Proteins 0.000 description 1
- 108010062796 arginyllysine Proteins 0.000 description 1
- 108010060035 arginylproline Proteins 0.000 description 1
- 108010038633 aspartylglutamate Proteins 0.000 description 1
- 108010047857 aspartylglycine Proteins 0.000 description 1
- 108010092854 aspartyllysine Proteins 0.000 description 1
- 230000002238 attenuated effect Effects 0.000 description 1
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 1
- 230000004071 biological effect Effects 0.000 description 1
- 208000002352 blister Diseases 0.000 description 1
- 210000004369 blood Anatomy 0.000 description 1
- 239000008280 blood Substances 0.000 description 1
- 230000017531 blood circulation Effects 0.000 description 1
- 210000004204 blood vessel Anatomy 0.000 description 1
- 239000007853 buffer solution Substances 0.000 description 1
- 201000011510 cancer Diseases 0.000 description 1
- 101150055766 cat gene Proteins 0.000 description 1
- 210000005056 cell body Anatomy 0.000 description 1
- 210000004671 cell-free system Anatomy 0.000 description 1
- 108091092356 cellular DNA Proteins 0.000 description 1
- 210000003169 central nervous system Anatomy 0.000 description 1
- 208000016886 cerebral arteriopathy with subcortical infarcts and leukoencephalopathy Diseases 0.000 description 1
- 206010008118 cerebral infarction Diseases 0.000 description 1
- 238000012512 characterization method Methods 0.000 description 1
- 238000007385 chemical modification Methods 0.000 description 1
- 238000004587 chromatography analysis Methods 0.000 description 1
- 230000001886 ciliary effect Effects 0.000 description 1
- 238000010367 cloning Methods 0.000 description 1
- 238000004891 communication Methods 0.000 description 1
- 230000004154 complement system Effects 0.000 description 1
- 238000009833 condensation Methods 0.000 description 1
- 230000005494 condensation Effects 0.000 description 1
- 108091036078 conserved sequence Proteins 0.000 description 1
- 239000013068 control sample Substances 0.000 description 1
- 230000001276 controlling effect Effects 0.000 description 1
- 238000011461 current therapy Methods 0.000 description 1
- 235000018417 cysteine Nutrition 0.000 description 1
- 125000000151 cysteine group Chemical class N[C@@H](CS)C(=O)* 0.000 description 1
- 108010060199 cysteinylproline Proteins 0.000 description 1
- 230000006378 damage Effects 0.000 description 1
- 230000004300 dark adaptation Effects 0.000 description 1
- 230000007547 defect Effects 0.000 description 1
- 230000007850 degeneration Effects 0.000 description 1
- 230000002939 deleterious effect Effects 0.000 description 1
- 239000003085 diluting agent Substances 0.000 description 1
- 238000010790 dilution Methods 0.000 description 1
- 239000012895 dilution Substances 0.000 description 1
- 238000006471 dimerization reaction Methods 0.000 description 1
- LOKCTEFSRHRXRJ-UHFFFAOYSA-I dipotassium trisodium dihydrogen phosphate hydrogen phosphate dichloride Chemical compound P(=O)(O)(O)[O-].[K+].P(=O)(O)([O-])[O-].[Na+].[Na+].[Cl-].[K+].[Cl-].[Na+] LOKCTEFSRHRXRJ-UHFFFAOYSA-I 0.000 description 1
- 108010054812 diprotin A Proteins 0.000 description 1
- 230000005750 disease progression Effects 0.000 description 1
- 238000009826 distribution Methods 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 229920002549 elastin Polymers 0.000 description 1
- 238000001962 electrophoresis Methods 0.000 description 1
- 238000005538 encapsulation Methods 0.000 description 1
- 239000002158 endotoxin Substances 0.000 description 1
- 238000005516 engineering process Methods 0.000 description 1
- 230000032050 esterification Effects 0.000 description 1
- 238000005886 esterification reaction Methods 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 238000011156 evaluation Methods 0.000 description 1
- 230000007717 exclusion Effects 0.000 description 1
- 239000013613 expression plasmid Substances 0.000 description 1
- 239000003889 eye drop Substances 0.000 description 1
- 229940012356 eye drops Drugs 0.000 description 1
- 229950003499 fibrin Drugs 0.000 description 1
- 238000009472 formulation Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 108010006664 gamma-glutamyl-glycyl-glycine Proteins 0.000 description 1
- 108010063718 gamma-glutamylaspartic acid Proteins 0.000 description 1
- 239000000499 gel Substances 0.000 description 1
- 238000003633 gene expression assay Methods 0.000 description 1
- 210000005046 glial fibrillary acidic protein Anatomy 0.000 description 1
- 108010078144 glutaminyl-glycine Proteins 0.000 description 1
- 108010013768 glutamyl-aspartyl-proline Proteins 0.000 description 1
- 230000002414 glycolytic effect Effects 0.000 description 1
- 108010062266 glycyl-glycyl-argininal Proteins 0.000 description 1
- 108010078326 glycyl-glycyl-valine Proteins 0.000 description 1
- 239000003102 growth factor Substances 0.000 description 1
- 229940029575 guanosine Drugs 0.000 description 1
- 230000036541 health Effects 0.000 description 1
- 238000010438 heat treatment Methods 0.000 description 1
- 125000000487 histidyl group Chemical group [H]N([H])C(C(=O)O*)C([H])([H])C1=C([H])N([H])C([H])=N1 0.000 description 1
- 206010020718 hyperplasia Diseases 0.000 description 1
- 238000003384 imaging method Methods 0.000 description 1
- 230000001771 impaired effect Effects 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 238000007901 in situ hybridization Methods 0.000 description 1
- 238000011534 incubation Methods 0.000 description 1
- MOFVSTNWEDAEEK-UHFFFAOYSA-M indocyanine green Chemical compound [Na+].[O-]S(=O)(=O)CCCCN1C2=CC=C3C=CC=CC3=C2C(C)(C)C1=CC=CC=CC=CC1=[N+](CCCCS([O-])(=O)=O)C2=CC=C(C=CC=C3)C3=C2C1(C)C MOFVSTNWEDAEEK-UHFFFAOYSA-M 0.000 description 1
- 229960004657 indocyanine green Drugs 0.000 description 1
- 210000004263 induced pluripotent stem cell Anatomy 0.000 description 1
- 230000001939 inductive effect Effects 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 150000002484 inorganic compounds Chemical class 0.000 description 1
- 229910010272 inorganic material Inorganic materials 0.000 description 1
- 229960003786 inosine Drugs 0.000 description 1
- 102000028416 insulin-like growth factor binding Human genes 0.000 description 1
- 108091022911 insulin-like growth factor binding Proteins 0.000 description 1
- 230000003993 interaction Effects 0.000 description 1
- 230000010039 intracellular degradation Effects 0.000 description 1
- 238000001990 intravenous administration Methods 0.000 description 1
- 230000002427 irreversible effect Effects 0.000 description 1
- 108010044374 isoleucyl-tyrosine Proteins 0.000 description 1
- 239000010978 jasper Substances 0.000 description 1
- 238000011813 knockout mouse model Methods 0.000 description 1
- 108010057821 leucylproline Proteins 0.000 description 1
- 238000001638 lipofection Methods 0.000 description 1
- 230000004807 localization Effects 0.000 description 1
- 208000018769 loss of vision Diseases 0.000 description 1
- 231100000864 loss of vision Toxicity 0.000 description 1
- 230000004777 loss-of-function mutation Effects 0.000 description 1
- 108010003700 lysyl aspartic acid Proteins 0.000 description 1
- 108010064235 lysylglycine Proteins 0.000 description 1
- 108010017391 lysylvaline Proteins 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 238000013507 mapping Methods 0.000 description 1
- 230000013011 mating Effects 0.000 description 1
- 230000008720 membrane thickening Effects 0.000 description 1
- 108010016686 methionyl-alanyl-serine Proteins 0.000 description 1
- 108010056582 methionylglutamic acid Proteins 0.000 description 1
- 108010085203 methionylmethionine Proteins 0.000 description 1
- 238000012737 microarray-based gene expression Methods 0.000 description 1
- 238000000520 microinjection Methods 0.000 description 1
- 239000011859 microparticle Substances 0.000 description 1
- 229930014626 natural product Natural products 0.000 description 1
- 210000005036 nerve Anatomy 0.000 description 1
- 210000000461 neuroepithelial cell Anatomy 0.000 description 1
- 210000002569 neuron Anatomy 0.000 description 1
- 230000007935 neutral effect Effects 0.000 description 1
- 108091027963 non-coding RNA Proteins 0.000 description 1
- 102000042567 non-coding RNA Human genes 0.000 description 1
- 230000006780 non-homologous end joining Effects 0.000 description 1
- 238000010899 nucleation Methods 0.000 description 1
- 210000004940 nucleus Anatomy 0.000 description 1
- 201000005111 ocular hyperemia Diseases 0.000 description 1
- 238000002577 ophthalmoscopy Methods 0.000 description 1
- 230000003204 osmotic effect Effects 0.000 description 1
- 230000007310 pathophysiology Effects 0.000 description 1
- 238000002823 phage display Methods 0.000 description 1
- 239000000546 pharmaceutical excipient Substances 0.000 description 1
- 108010084572 phenylalanyl-valine Proteins 0.000 description 1
- 239000002953 phosphate buffered saline Substances 0.000 description 1
- 102000020233 phosphotransferase Human genes 0.000 description 1
- 230000004310 photopic vision Effects 0.000 description 1
- 239000000049 pigment Substances 0.000 description 1
- 210000002826 placenta Anatomy 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 239000002243 precursor Substances 0.000 description 1
- 125000002924 primary amino group Chemical group [H]N([H])* 0.000 description 1
- 230000008569 process Effects 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 108010070643 prolylglutamic acid Proteins 0.000 description 1
- 230000000069 prophylactic effect Effects 0.000 description 1
- 235000019419 proteases Nutrition 0.000 description 1
- 230000012743 protein tagging Effects 0.000 description 1
- 238000000746 purification Methods 0.000 description 1
- 230000001172 regenerating effect Effects 0.000 description 1
- 238000007634 remodeling Methods 0.000 description 1
- 239000000790 retinal pigment Substances 0.000 description 1
- 210000003583 retinal pigment epithelium Anatomy 0.000 description 1
- 238000012954 risk control Methods 0.000 description 1
- 102220130916 rs1061147 Human genes 0.000 description 1
- 102220005751 rs2274700 Human genes 0.000 description 1
- 102200031793 rs800292 Human genes 0.000 description 1
- 210000001423 scleral cell Anatomy 0.000 description 1
- 230000004296 scotopic vision Effects 0.000 description 1
- 238000007790 scraping Methods 0.000 description 1
- 230000028327 secretion Effects 0.000 description 1
- 238000002864 sequence alignment Methods 0.000 description 1
- 210000002966 serum Anatomy 0.000 description 1
- 239000004055 small Interfering RNA Substances 0.000 description 1
- 239000011780 sodium chloride Substances 0.000 description 1
- 241000894007 species Species 0.000 description 1
- 238000004611 spectroscopical analysis Methods 0.000 description 1
- 230000006641 stabilisation Effects 0.000 description 1
- 238000011105 stabilization Methods 0.000 description 1
- 210000000130 stem cell Anatomy 0.000 description 1
- 235000000346 sugar Nutrition 0.000 description 1
- 150000008163 sugars Chemical class 0.000 description 1
- 239000013589 supplement Substances 0.000 description 1
- 230000002195 synergetic effect Effects 0.000 description 1
- 238000007910 systemic administration Methods 0.000 description 1
- 210000001550 testis Anatomy 0.000 description 1
- 230000004797 therapeutic response Effects 0.000 description 1
- 108010061238 threonyl-glycine Proteins 0.000 description 1
- 229940104230 thymidine Drugs 0.000 description 1
- 229940113082 thymine Drugs 0.000 description 1
- 230000000451 tissue damage Effects 0.000 description 1
- 231100000827 tissue damage Toxicity 0.000 description 1
- 230000000699 topical effect Effects 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 230000001988 toxicity Effects 0.000 description 1
- 231100000419 toxicity Toxicity 0.000 description 1
- 230000005030 transcription termination Effects 0.000 description 1
- 230000037426 transcriptional repression Effects 0.000 description 1
- 238000012546 transfer Methods 0.000 description 1
- 230000001131 transforming effect Effects 0.000 description 1
- 238000003146 transient transfection Methods 0.000 description 1
- 238000002054 transplantation Methods 0.000 description 1
- RTKIYFITIVXBLE-QEQCGCAPSA-N trichostatin A Chemical compound ONC(=O)/C=C/C(/C)=C/[C@@H](C)C(=O)C1=CC=C(N(C)C)C=C1 RTKIYFITIVXBLE-QEQCGCAPSA-N 0.000 description 1
- NMEHNETUFHBYEG-IHKSMFQHSA-N tttn Chemical compound C([C@@H](C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CO)C(=O)N[C@@H](CC=1NC=NC=1)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](C(C)C)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(N)=O)C(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N1[C@@H](CCC1)C(=O)NCC(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(O)=O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(O)=O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CCSC)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@H](CC(O)=O)NC(=O)[C@@H](NC(=O)[C@H]1N(CCC1)C(=O)[C@H](CCC(N)=O)NC(=O)[C@@H](N)[C@@H](C)O)[C@@H](C)O)C1=CC=CC=C1 NMEHNETUFHBYEG-IHKSMFQHSA-N 0.000 description 1
- 230000009452 underexpressoin Effects 0.000 description 1
- 230000006490 viral transcription Effects 0.000 description 1
- 230000004304 visual acuity Effects 0.000 description 1
Images











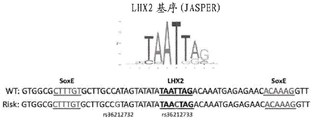
































Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K48/00—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy
- A61K48/005—Medicinal preparations containing genetic material which is inserted into cells of the living body to treat genetic diseases; Gene therapy characterised by an aspect of the 'active' part of the composition delivered, i.e. the nucleic acid delivered
- A61K48/0066—Manipulation of the nucleic acid to modify its expression pattern, e.g. enhance its duration of expression, achieved by the presence of particular introns in the delivered nucleic acid
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K31/00—Medicinal preparations containing organic active ingredients
- A61K31/70—Carbohydrates; Sugars; Derivatives thereof
- A61K31/7088—Compounds having three or more nucleosides or nucleotides
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P27/00—Drugs for disorders of the senses
- A61P27/02—Ophthalmic agents
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/11—DNA or RNA fragments; Modified forms thereof; Non-coding nucleic acids having a biological activity
- C12N15/113—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing
- C12N15/1137—Non-coding nucleic acids modulating the expression of genes, e.g. antisense oligonucleotides; Antisense DNA or RNA; Triplex- forming oligonucleotides; Catalytic nucleic acids, e.g. ribozymes; Nucleic acids used in co-suppression or gene silencing against enzymes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C12N15/86—Viral vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases RNAses, DNAses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/48—Hydrolases (3) acting on peptide bonds (3.4)
- C12N9/50—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25)
- C12N9/64—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue
- C12N9/6421—Proteinases, e.g. Endopeptidases (3.4.21-3.4.25) derived from animal tissue from mammals
- C12N9/6424—Serine endopeptidases (3.4.21)
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
- C07K2319/80—Fusion polypeptide containing a DNA binding domain, e.g. Lacl or Tet-repressor
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2310/00—Structure or type of the nucleic acid
- C12N2310/10—Type of nucleic acid
- C12N2310/20—Type of nucleic acid involving clustered regularly interspaced short palindromic repeats [CRISPRs]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2320/00—Applications; Uses
- C12N2320/30—Special therapeutic applications
- C12N2320/34—Allele or polymorphism specific uses
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2740/00—Reverse transcribing RNA viruses
- C12N2740/00011—Details
- C12N2740/10011—Retroviridae
- C12N2740/16011—Human Immunodeficiency Virus, HIV
- C12N2740/16041—Use of virus, viral particle or viral elements as a vector
- C12N2740/16043—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2750/00—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA ssDNA viruses
- C12N2750/00011—Details
- C12N2750/14011—Parvoviridae
- C12N2750/14111—Dependovirus, e.g. adenoassociated viruses
- C12N2750/14141—Use of virus, viral particle or viral elements as a vector
- C12N2750/14143—Use of virus, viral particle or viral elements as a vector viral genome or elements thereof as genetic vector
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/40—Systems of functionally co-operating vectors
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2830/00—Vector systems having a special element relevant for transcription
- C12N2830/008—Vector systems having a special element relevant for transcription cell type or tissue specific enhancer/promoter combination
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y304/00—Hydrolases acting on peptide bonds, i.e. peptidases (3.4)
- C12Y304/21—Serine endopeptidases (3.4.21)
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Organic Chemistry (AREA)
- Biomedical Technology (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Microbiology (AREA)
- Medicinal Chemistry (AREA)
- Plant Pathology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Pharmacology & Pharmacy (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Epidemiology (AREA)
- Virology (AREA)
- Mycology (AREA)
- Ophthalmology & Optometry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Abstract
本发明提供了用于治疗10号染色体驱动的年龄相关性黄斑变性的组合物和方法,其包括基因治疗以增加眼中视网膜色素上皮细胞中的HTRA1表达。
Description
技术领域
本发明涉及用于治疗年龄相关性黄斑变性的方法和组合物,以及其在生物和医疗领域的应用。
序列列表
本申请包含已经以ASCII格式电子提交的序列表,并据此以引用方式全文并入。于2020年4月9日形成的所述ASCII副本被命名为098846-1185529-000810PC_SL.txt并且大小为46,642字节。
相关申请的交叉引用
本申请要求2019年4月10日提交的美国临时申请62/832,182的权益,其全部内容以引用方式并入本文。
背景技术
年龄相关性黄斑变性(AMD)是发达国家不可逆视力丧失的主要原因(概述参见Zarbin,fur.Ophthalmol.8:199-206,(1998)),其影响约15%的60岁以上的人。估计有6亿人属于这个年龄人口。AMD的患病率随着年龄的增长而增加;在75岁及更老的人群中,近30%发生轻度或早期型,约7%发生晚期型(Vingerling et al.,Epidemiol.Rev.17(2):347-360,1995)。
传统疗法通常涉及两个月一次或根据需要玻璃体内注射治疗剂,例如抗VEGF剂。这些疗法需要反复进行侵入性手术,这给患者带来了极大的不适和不便。这些疗法中涉及的程序也可增加副作用的风险。此外,在大多数情况下,治疗剂仅在脉络膜新血管形成(CNV)发展后和黄斑显著损伤时施用。目前没有批准用于早期AMD或预防AMD发展的疗法
基于对AMD患者的广泛基因分型研究,现在了解到AMD包括两种不同的生物学疾病:染色体1指导的AMD(或“Chr 1AMD”),其由补体系统失调引起,包括补体因子H失调,以及10号染色体指导的AMD(或“Chr 10AMD”),其与染色体区域10q26中的遗传变体相关,该区域含有ARMS2和HTRA1基因。参见Keenan et al,2015,“ASSESSMENT OF PROTEINS ASSOCIATEDWITH COMPLEMENT ACTIVATION AND INFLAMMATION IN MACULAE OF HUMAN DONORSHOMOZYGOUS RISK AT CHROMOSOME 1CFH-TO-F13B”Invest Ophthalmol Vis Sci.56:487-79;Hageman,2015,“METHODS OF PREDICTING THE DEVELOPMENT OF AMD BASED ONCHROMOSOME1AND CHROMOSOME 10”,美国专利公布2015/0211065,其以引用方式并入。据报道,1号染色体和10号染色体上的这些基因座一起占白种人人群所有AMD风险的95%,并且染色体区域10q26与AMD风险增加密切相关(Fisher et al.,2005;Rivera et.al.2005)。风险区包含共享广泛连锁不平衡(LD)(D’=0.99)的基因(包括ARMS2和HTRA1)。
发明内容
本公开提供了用于治疗10号染色体驱动的年龄相关性黄斑变性(AMD)、预防其发展、减缓其进展、逆转或改善其症状和体征的方法。
在一些方面,本公开提供了一种通过施用增加受试者细胞(例如,RPE细胞、水平细胞或感光细胞)中的HTRA1表达的药剂来治疗、预防受试者的Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法。在一些情况下,受试者具有单个10号染色体风险等位基因。在一些情况下,受试者对于10号染色体风险等位基因是纯合的。在一些情况下,受试者表现出Chr 10AMD临床表型。在一些情况下,受试者不携带一个或多个1号染色体风险等位基因。
在一些方案中,药剂引起内源性HTRA1基因序列的转录的上调节。例如,药剂可以为结合HTRA1转录调节区或HTRA1启动子内的靶序列的转录激活子。在一些情况下,药剂为下列的融合蛋白:i)能够识别HTRA1转录调节区内的靶序列的DNA靶向蛋白,和ii)转录激活子。在一些情况下,DNA靶向蛋白通过与HTRA1转录调节区或HTRA1启动子区内的序列互补的指导RNA识别靶序列。在一些情况下,DNA靶向蛋白是无酶活性的Cas9蛋白(dCas9)。在一些情况下,转录激活子为VP16。在一些情况下,方法还包括施用与HTRA1转录调节区内的序列互补的sgRNA。
在一些情况下,施用药剂导致受试者细胞(包括,例如RPE细胞、水平细胞或感光细胞)中的外源性HTRA1蛋白的表达。在基因疗法方案中,药剂可以为载体,诸如病毒载体(例如,AAV或慢病毒),其递送优选地可操作连接到启动子的外源性HTRA1蛋白或编码HTRA1蛋白的核酸序列。在一些情况下,启动子为RPE特异性启动子。
在一些情况下,使用药剂治疗受试者导致受试者的RPE细胞中的染色体DNA的改性。例如,基因组编辑方法可用于改变HTRA1基因或转录调节区中的序列或者将一个或更多个风险等位基因转换成对应的一个或多个非风险等位基因。在一些情况下,药剂为基于DNA核酸内切酶的体系。在一些情况下,DNA核酸内切酶为Cas9核酸内切酶、锌指核酸酶、转录激活子样效应子核酸酶、归巢核酸内切酶、大范围核酸酶或Cre重组酶。在一些情况下,药剂修饰染色体DNA并产生包含SEQ ID NO:34的染色体DNA序列。在一些情况下,药剂修饰染色体DNA并产生包含SEQ ID NO:14的染色体DNA序列。在一些情况下,将rs36212733处的风险等位基因序列(C)改变成非风险等位基因序列(t)。在一些情况下,通过视网膜下注射施用药剂。在一些情况下,通过脉络膜上层注射递送药剂。在一些情况下,通过玻璃体内注射递送药剂。在一些情况下,通过其他途径递送药剂。在一些情况下,一种或更多种药剂包含Cas9和一种或更多种sgRNA,其中一种或更多种sgRNA将Cas9引导到包含一种或更多种风险等位基因的染色体区,从而修改该染色体区,这导致一种或更多种风险等位基因被对应的非风险等位基因置换。
在一些情况下,药剂为增加RPE细胞中HTRA1表达或活性的小分子化合物、肽或核酸。
在一些方面,本公开提供了一种治疗Chr 10AMD的药物组合物,其包含(i)增加受试者的RPE细胞中的HTRA1表达的药剂,和(ii)药学上可接受的载体。
在一种形式中,本发明的组合物包含指导RNA(gRNA),其包含对应于HTRA1启动子或HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸。在一个方面,HTRA1启动子具有在SEQ ID NO:5、7、8或13中列出的序列,并且2kb调节区具有在SEQ ID NO:14中列出的序列。
在其他方面,本发明包含核蛋白(RNP)复合物,所述复合物包含:(a)指导RNA(gRNA),其包含对应于HTRA1启动子或HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸;以及(b)融合蛋白,其包含与转录激活子结构域融合的CRISPR-相关蛋白(Cas)结构域,其中Cas蛋白结构域缺乏核酸酶活性,并且HTRA1启动子具有在SEQ ID NO:5、7、8或13中列出的序列,并且2kb调节区具有在SEQ ID NO:14中列出的序列。在一个实施方案中,RNP复合物的Cas为dCas9或dCas12a。在一个方面,RNP复合物的靶序列在启动子中并且转录激活子选自VP16、VP64、VP160、MLL、E2A、HSF1、NF-IL6、NFAT1和NF-kB。在一个方面,RNP复合物的靶序列在2kb调节区中并且转录激活子为LHX2。
在另一方面,本发明包含核蛋白(RNP)复合物,所述复合物包含:(a)指导RNA(gRNA),其包含对应于HTRA1启动子或HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸的指导序列;以及(b)CRISPR-相关(Cas)蛋白,其中HTRA1 2kb调节区具有在SEQ ID NO:14中列出的序列。在一个实施方案中,RNP复合物的Cas为Cas9、Cas12a或Cas3。
在一个方面,gRNA或RNP复合物的指导序列包含对应于靶序列的至少15个连续核苷酸。在另一方面,gRNA或RNP复合物的指导序列包含对应于靶序列的至少20个连续核苷酸。在又一个方面,gRNA或RNP复合物的指导序列包含对应于靶序列的12-25各连续核苷酸。在一个实施方案中,gRNA或RNP复合物的靶序列与前间隔序列邻近基序(PAM)相邻,所述前间隔序列邻近基序为NGG。
在一个方面,gRNA或RNP复合物的指导序列包含SEQ ID NO:15-33中的任一者,或包含SEQ ID NO:15-33中任一者的至少15个连续碱基的序列。
在另一个方面,本发明包括gRNA或RNP复合物,其中(i)指导序列包含SEQ ID NO:36-49中的任一者;和/或(ii)靶序列包含或邻近选自s10490924、rs144224550、rs36212731、rs36212732、rs36212733、rs3750848、rs3750847和rs3750846处的风险的风险等位基因。
在另一个方面,本发明包括编码gRNA的多核苷酸,其中所述多核苷酸为DNA。在一个实施方案中,多核苷酸包含可操作地连接到编码gRNA的序列的启动子。
在另一个方面,本发明包括包含多核苷酸的病毒载体。
在另一方面,本发明提供了包含编码HTRA1多肽的多核苷酸,其中所述多核苷酸包含编码可操作地连接到启动子的HTRA1的人密码子优化的序列。在一个实施方案中,病毒载体为逆转录酶病毒、慢病毒、疱疹病毒或腺相关病毒(AAV)。
在一个实施方案中,多核苷酸的启动子或病毒载体为RPE特异性启动子。
在另一个方面,本发明包括HTRA1激活系统,其包含:(a)包含编码gRNA的DNA序列;和(b)包含编码融合蛋白的DNA序列的载体,融合蛋白包含融合到转录激活子结构域的Cas蛋白结构域,其中Cas蛋白结构域缺乏核酸酶活性。在一个实施方案中,(a)中的载体和(b)中的载体为不同的载体。
在另一方面,本发明包括HTRA1靶向系统,其包含:(a)包含编码本文所公开的gRNA的核酸的载体;和(b)包含编码Cas蛋白的核酸的载体。在一个方面,HTRA1靶向系统还包含:(c)包含编码模板修复序列的核酸的载体,所述模板修复序列任选地包含SEQ ID NO:87-94中的至少一者或SEQ ID NO:87-94中的至少一者的互补序列。在一个实施方案中,(a)和(b)是相同的载体,或(a)、(b)和(c)是相同的载体。
在另一个方面,本发明包含分离的细胞,其包含gRNA、RNP、多核苷酸、病毒载体、激活系统或靶向系统。
在一个方面,本发明提供了gRNA、分离的多核苷酸、载体、激活系统、靶向系统或分离的细胞用于制备用于治疗年龄相关性黄斑病变(AMD)的药物的用途。在另一方面,
指导RNA、分离的多核苷酸、载体、激活系统、靶向系统或分离的细胞用于制备用于治疗年龄相关性黄斑病变(AMD)的药物。在一个实施方案中,受治疗的受试者(a)表现出Chr10AMD临床表型;(b)具有10号染色体风险等位基因;(c)对于10号染色体风险等位基因是纯合的;或(d)
不具有1号染色体风险等位基因。
本发明还包括一种用于增加细胞中HTRA1表达的方法,所述方法包括在细胞中表达激活系统或靶向系统。
在另一方面,本发明包括一种治疗受试者的Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法,所述方法包括施用增加受试者RPE细胞或水平细胞中HTRA1表达的一种或多种药剂。在一个实施方案中,受试者表现出Chr 10AMD临床表型。在另一个实施方案中,受试者具有10号染色体风险等位基因。在另一个实施方案中,受试者对于10号染色体风险等位基因是纯合的。在另一实施方案中,受试者不具有1号染色体风险等位基因。在一个方面,内源HTRA1基因序列的转录增加。在一个实施方案中,药剂为核蛋白复合物,其包含:(a)无酶活性的Cas蛋白结构域和转录激活子结构域的融合蛋白和(b)指导RNA。在该方面的一个实施方案中,无酶活性的Cas蛋白为dCas9。在一个实施方案中,核蛋白复合物结合在HTRA1启动子区域中。在另一实施方案中,核蛋白复合物结合在HTRA1增强子区域中。在一个实施方案中,转录激活子结构域结合LHX2结合基序。在另一实施方案中,药剂为核糖核酸复合物,其包含指导RNA和Cas蛋白。在另一方面,受试者携带HTRA1基因增强子中的风险等位基因,并且药剂为包含下列物质的组合:(a)包含指导RNA和Cas蛋白的核糖核酸复合物,和(ii)包含对应于风险等位基因的非风险等位基因序列的模板修复多核苷酸。
附图说明
图1示出了人视网膜中HTRA1 mRNA的原位杂交。
图2示出了Newman数据集中HTRA1 mRNA表达的微阵列分析。
图3A和3B示出了DiaxonHit(A)和Newman(B)数据集中HTRA1 mRNA表达的qRT-PCR分析(人眼部组织)
图4A和4B分别示出了HTRA1在人眼部组织(RPE-脉络膜和神经褶)和RPE刮取物中的等位基因特异性表达。测定相对于具有非风险等位基因的HTRA1 mRNA的量,包含rs1049331处的风险等位基因的HTRA1 mRNA的量。
图5A和5B示出了hTERT-RPE1和分化的人胎儿RPE细胞中HTRA1蛋白的极化表达。
图6示出了来自人眼部的各种组织提取物中检测的HTRA1蛋白的量。
图7示出了长的非编码RNA lncSCTM1(LOC105378525)的预测和观察到的同种型。
图8示出了RPE-脉络膜和视网膜中人HTRA1 mRNA和lncSCTM1RNA的等位基因特异性表达。
图9示出了LHX2蛋白的一致性结合基序以及被rs3621273多态位点处的风险等位基因破坏的基序。
图10A和10B示出了LHX2与在rs36212733处包含非风险(WT)序列的Chr10探针的结合,但较弱结合到风险序列且不结合到杂乱序列。
图11示出了非风险Chr10探针序列相对于风险序列的优先结合亲和性。
图12A和12B示出了按Chr10基因型基团计,人视网膜中的HTRA1蛋白和mRNA含量。
图13A和13B示出了按Chr10基因型基团计,RPE-脉络膜中的HTRA1蛋白和mRNA含量。
图14示出了用于靶向HTRA1启动子的sgRNA序列。
图15示出了使用从用基于CRISPR的SAM组分处理72小时的h1RPE7细胞中分离的总RNA的HTRA1和IL1B mRNA含量(定量RT-PCR)。
图16示出了用LentiMPH质粒(MPH)、LentiSAM质粒(SAM)或两者(MPH+SAM)模拟转染或转染72小时的h1RPE7细胞的总RNA中的HTRA1mRNA含量(定量RT-PCR)。
图17A和17B示出了用HTRA1-靶向的P7 sgRNA-LentiSAM(P7)或P18 sgRNA(P18)或具有或不具有LentiMPH质粒(MPH)的IL1B-靶向的P2sgRNA-LentiSAM转染的h1RPE7细胞的总RNA中的HTRA1(图17A)和IL1B(图17B)mRNA的含量。
图18A和18B示出了用不同量的P7 sgRNA-LentiSAM质粒(SAM)转染的h1RPE7细胞的总RNA中HTRA1 mRNA的含量(定量RT-PCR)。
图19A和19B示出了HTRA1 ELISA的结果,(A)HTRA1蛋白质的标准曲线;(B)用不同量(2.5μg、5.0μg和7.5μg)的Ctrl sgRNA-或P18sgRNA-LentiSAM转染3天、4天和5天的h1RPE7的细胞培养物上清液中的HTRA1蛋白质含量。
图20A和20B示出了用不同量(2.5μg、5.0μg和7.5μg)的Ctrl sgRNA-LentiSAM或P18 sgRNA转染3天、4天和5天的h1RPE7细胞中标称化HTRA1蛋白质含量(A)和HTRA1 mRNA含量(B)的结果。
图21示出了以MOI=20用Ctrl-或P18-LentiSAM病毒颗粒转染3天、6天和9天的h1RPE7的细胞培养物上清液中的HTRA1蛋白含量。数据按照相对于Ctrl-LentiSAM的倍数增长绘图。
图22示出了以MOI=20用Ctrl-或P18-LentiSAM病毒颗粒转染3天、6天和9天的h1RPE7的细胞培养物上清液中的ENPP-2蛋白含量(ng/ml)。
图23A和23B示出了用Ctrl sgRNA-或P18 sgRNA-LentiSAM的MOI 20转染3天、6天和9天的h1RPE7的细胞中标称化HTRA1蛋白含量(A)和HTRA1 mRNA含量(B)的结果。
图24示出了相对于对照pCTM259载体,使用脂质体3000用无启动子(无)、BEST1、RPE65或CMV-HTRA1质粒转染72小时的HTRA1敲除RPE1细胞中的HTRA1 mRNA含量。
图25示出了相对于对照pCTM259载体,使用脂质体3000用无启动子(无)、BEST1、RPE65或CMV-HTRA1质粒转染72小时的RPE1细胞中的HTRA1 mRNA含量。
图26示出了用由EST1-、RPE65-或CMV-来源的启动子驱动的AAV2-HTRA1质粒转染的RPE1(HTRA1 KD)细胞中的HTRA1 mRNA表达的时程动力学。
图27A-27C示出了用由EST1-、RPE65-或CMV-来源的启动子驱动的AAV2-HTRA1质粒转染的RPE1(HTRA1 KD)细胞中的HtrA1蛋白表达的时程动力学。
图28示出了包含ARMS2基因的Chr10风险基因座内的区域的等位基因特异性缺失的示例。
图29示出了AMD 10号染色体基因座上的“原因”(调节)区和lncSCTM1。Chr10调节区为大约2-4kb。鉴定出新型lncRNA(命名为lncSCTM1),其与该调节区重叠。新型lncRNA包含ARMS2 rs10490924。箭头示出IncSCTM1从HTRA1启动子以反义取向转录并且与HTRA1共享分歧启动子。
图30示出了人外黄斑RPE脉络膜(图30A)、外黄斑视网膜(图30B)、黄斑RPE脉络膜(图30C)和黄斑视网膜(图30D)中,用外显子靶向探针对HTRA1 mRNA表达进行微阵列分析,其比较了Chr10非风险(GG)供体与杂合(GT)和纯合风险(TT)供体。
具体实施方式
发明人已发现,增加眼(例如,视网膜色素上皮细胞)中的HTRA1 mRNA和/或蛋白质含量向患有或处于形成年龄相关性黄斑变性(AMD)风险的受试者提供有益效果。具体地讲,增加HTRA1 mRNA和/或蛋白含量向患有10号染色体指导的AMD(或“Chr 10AMD”)表现或者遗传上倾向于发展10号染色体指导的AMD的患者提供有益效果。
本公开提供了通过增加需要治疗的受试者眼部中的HTRA1表达或含量来治疗年龄相关性黄斑变性(AMD)、预防其发展、减缓其进展、逆转或改善其症状和体征的方法和试剂。
描述了治疗Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法。该方法包括向受试者施用药剂,其中所述药剂增加受试者的RPE细胞、或者另选地水平细胞或感光细胞中的HTRA1 mRNA和/或蛋白含量。下文描述了用于增加HTRA1 mRNA和/或蛋白含量的示例性方法。在一种方案中,HTRA1表达在受试者的视网膜色素上皮(RPE)细胞中增加。用于增加眼部中的HTRA1 mRNA和/或蛋白含量的方法包括HTRA1的转录调节,包括使用CRISPRa将内源性HTRA1的表达上调,风险区中的CRISPR-介导的修复,将HTRA1蛋白编码序列导入细胞中的基因疗法,减少HTRA1在细胞内的降解以及细胞疗法。然而,本发明不限于特定方法并且可使用任何治疗有效的方法。如本文所用,“增加HTRA1的表达”是指通过增加来自内源性或外源性基因的转录和基因产品(mRNA或蛋白质)的产生来增加HTRA1蛋白的含量。因此,取决于上下文,HTRA1“表达”可指HTRA1 mRNA或HTRA1蛋白的制备。“增加表达”也可指通过除了转录之外的机制增加HTRA1蛋白量的方法,诸如增加细胞中HTRA1蛋白或mRNA的稳定性,增加HTRA1 mRNA的翻译率,mRNA招募,以及将外源性HTRA1蛋白引入细胞中。读者将意识到“增加HTRA1表达”可指增加所需种类的HTRA1蛋白的量。例如,在表达与风险基因型相关的内源性HTRA1蛋白的细胞中,“增加HTRA1表达”可指增加具有与风险基因型无关的序列的HTRA1蛋白的量。在一种方法中,HTRA1丝氨酸蛋白酶活性增加。术语“上调”可用于指将转录(例如,mRNA生产的量)相对于不用药剂处理的对照细胞增加至少10%、或至少20%、或至少30%、或至少50%。
预料不到的是发现增加HTRA1含量向患者提供有益效果。AMD领域中一致认为HTRA1的过表达而不是欠表达与发展AMD的风险增加相关,并且应当减少或抑制HTRA1含量或活性来治疗AMD。例如,Dewan等人(2006)记录了在rs11200638处的HTRA1启动子SNP的风险等位基因(A)与培养的RPE细胞中的HTRA1转录增强相关。Yang等人(2006)还记录了与来自非风险等位基因(GG)纯合子个体的淋巴细胞相比,来自风险等位基因(AA)纯合子个体的循环淋巴细胞表达更高含量的HTRA1 mRNA。Chan等人(2007)记录了在患有AMD的患者的存档者中HTRA1表达在mRNA含量方面上调并且得出结论HTRA1表达增强是湿性AMD的斑点损伤中活性新血管形成的原因。同样,Vierkotten等人(2011)记录了HTRA1的过表达与Bruch膜的弹性层中的超微结构变化相关,并且提出HTRA1对AMD的病理生理学起作用。Jones等人(2011)记录了小鼠视网膜色素上皮细胞中过表达人HTRA1与诱发的息肉状脉络膜血管病变(“PVC”)主特征相关,包括脉络膜血管的分支网络、息肉状病变、弹性层的严重退化和脉络膜血管的中膜。DeAngelis(美国专利公布2013/0122016)提出通过降低HTRA1基因的表达或降低HTRA1基因产物的生物活性来减缓受试者的年龄相关性黄斑变性的进展。Wu等人(美国专利公布2013/0129743)提出使用结合HTRA1并抑制HTRA1酶活性的单克隆抗体来治疗AMD。
1.与AMD相关的HTRA1调控元件
已经鉴定了通过增加HTRA1水平来治疗AMD的靶标和方法。该发现部分地基于实施例中描述的研究。
使用来自对rs10490924(GT,TT)处的AMD风险等位基因纯合或杂合的人类受试者和来自对于非风险(GG)纯合的对照受试者的供体眼,观察到与对照相比,具有AMD风险等位基因的供体具有较低的HTRA1 mRNA表达。重要的是,数据展示出风险患者的HTRA1 mRNA水平的降低是组织特异性的:在RPE中检测到该降低,但在神经视网膜或脉络膜中未检测到。此外,在具有或不具有Chr10基因座风险的供体中,人黄斑外视网膜和RPE-脉络膜中的HTRA1蛋白和mRNA水平根据年龄变化的比较结果展示出,视网膜HtrA1水平随年龄相对不变,与受试者的Chr10风险状态无关,但在RPE脉络膜中,与具有风险等位基因的供体相比,不具有Chr10风险的供体的HtrA1 mRNA和蛋白质水平随年龄显著增加。
4 kb AMD风险区域
使用HTRA1等位基因特异性表达测定来缩小10号染色体区域,该区域是AMD的原因。该测定使用来源于人供体眼睛的mRNA,其中罕见的重组事件在AMD相关的ARMS2/HTRA1LD块内。将与HTRA1 mRNA减少相关的区域映射到HTRA1编码序列上游的大约4kb调节区(介于rs11200632和rs3750846之间并包括rs11200632和rs3750846)。4kb区域包括rs10490924(ARMS2 A69S)。通过Grassmann等人(Genetics 2017)在其案例/对照研究中对重组单倍型的分析发现相同的遗传区域与AMD疾病的风险升高有关。发现被鉴定为与HTRA1的等位基因特异性表达相关的4kb区域与AMD风险相关的区域匹配强烈地暗示HTRA1表达的风险相关性降低导致AMD发病率增加。该区域被命名为“4kb AMD风险区域”,或等同地“4kb风险区域”、“4kb调控区域”、“4kb因果区域”或“4kb区域”。
2 kb AMD风险区域
使用描述转录激活的表观遗传标记物的数据,鉴定出大约2kb的基因组区域(对应于Chr10:122454508-122456564(hg38)),其与4kb区域重叠并调节HTRA1转录。在RPE细胞中,该区域的特征在于转录激活的标志物,包括H3K27乙酰化,但该区域在视网膜组织或使用各种非RPE细胞系的ENCODE数据中未示出H3K27乙酰化,从而表明该区域的染色质在RPE组织中是活跃的,但在其他细胞类型中不是活跃的。2kb AMD风险区域”(SEQ ID NO:14)等效地称为“2kb风险区域”、“2kb调控区域”、“2kb因果区域”或“2kb区域”。2kb AMD风险区域也称为“HTRA1增强子区域”。
HTRA1启动子
HTRA1“启动子序列”包含CRISPRa(CRISPR激活)靶序列和其他特征。启动子区序列在以下序列中提供。除非另有说明或由上下文显而易见,本文对“启动子”的提及旨在指这些序列范围中的每一个。SEQ.ID NO:8(300bp)示出了HTRA1初级启动子序列,其包括自推定的转录起始位点起的5'300bp。SEQ.ID NO:5(469bp)示出具有附加上游序列的扩展启动子序列。SEQ.ID NO:7(400bp)示出初级启动子序列加上100bp的5'UTR序列。“天然启动子区域”是具有启动子和UTR序列的SEQ ID NO:13(853bp)。
1.通过转录激活增加HTRA1表达
在一些方法中,基因疗法用于增强RPE细胞中内源性HTRA1的表达。在一些方法中,基因治疗涉及递送一种转录激活子,该转录激活子可与HTRA1的转录调控区结合并促进HTRA1的转录。在一种方法中,使用结合HTRA1启动子区域并将转录激活子定位在HTRA1启动子区域附近的靶向部分(例如,Cas蛋白-指导RNA复合物)。在相关方法中,靶向部分结合HTRA1增强子区域并将转录激活子定位在转录激活子结合基序附近。
在一些方法中,转录激活子活性由转录激活子和DNA靶向蛋白的融合蛋白提供。本文所公开的DNA靶向蛋白可以为结合到HTRA1转录调控区的DNA靶向蛋白。可以使用使用DNA靶向蛋白的各种平台,如下所述。在某些实施方案中,使用“CRISPRa”(CRISPR激活)方法。在一些情况下,转录激活子作为融合蛋白提供,其中核酸酶缺陷型II CRISPR相关蛋白(Cas)是DNA靶向蛋白。一般来讲,Cas被修饰使得其缺乏核酸内切酶活性(即“核酸酶死亡Cas”或“dCas”)。在一些方法中,Cas是dCas9。在一些方法中,Cas是dCas12a。在一些方法中,Cas是任何Cas蛋白,经过修饰使其缺乏核酸酶。如上文和本文其他地方所述,本发明不限于这种激活方法,并且可使用将转录激活子引导至启动子或增强子区域的非Cas蛋白或部分。
因此,在本发明的一些方面,dCas9用于在CRISPR激活(“CRISPRa”)系统中介导HTRA1表达的上调。与广泛用于II型CRISPR/Cas系统并在被引导至目标序列时在基因组DNA中产生双链断裂的Cas9相比,dCas9缺乏核酸酶活性并且不在DNA中产生双链断裂。相反,dCas9可连接到转录激活子并与指导RNA(例如sgRNA)复合,所述指导RNA与HTRA1启动子区中的序列特异性杂交,以实现精确且稳健的RNA指导的转录调控。dCas9介导的基因激活系统的使用是众所周知的,并在例如Dominquez et al.,2016,“Beyond editing:repurposing CRISPR Cas9for precision genome regulation and interrogation,”Nat.Rev.Mol.Cell.Biol.17:5-15中进行了描述。
其他核酸酶缺陷型Cas蛋白也可用于将转录激活子募集到启动子或增强子位点。例如,dCas12a结构域可融合到转录激活子结构域以用于本发明的方法中(参见,Sarstedt,“Spotlight on Cas12 A search for more type V Cas12 family members turns upunexpected functionality”,Nature Methods,2019,16:213 219;Pickar-Oliver etal.,“The next generation of CRISPR Cas technologies and applications”,NatureReviews Molecular Cell Biology,2019,20:490-507;Kleinstiver et al.,“EngineeredCRISPR Cas12a Variants with Increased Activities and Improved TargetingRanges for Gene,Epigenetic And Base Editing”,Nature Biotechnology,2019,37:276282;Xu et al.,2018,“A CRISPR dCas Toolbox for Genetic Engineering andSynthetic Biology”,J.Mol.Biol.doi.org/10.1016/j.jmb.2018.06.037)。在一种方法中,Cas 12a为酸氨基球菌属BV3L6(AsCas12a)。
因此,在一个实施方案中,本文所公开的方法包括向患者施用药剂,其中施用导致dCas-转录激活子融合蛋白和一种或多种sgRNA递送至需要治疗的患者的眼部(例如,RPE)。融合蛋白在一个或多个单指导RNA(sgRNA)的指导下与HTRA1调节区结合,从而上调HTRA1转录。在一种方法中,dCas9来源于金黄色葡萄球菌(“Sa-dCas9”)。在一种方法中,dCas9来源于化脓性葡萄球菌(“Sp-dCas9”)。dCas9转录激活子融合蛋白可以采用不同的构型,例如Dominguez et al.,Nat Rev Mol Cell Biol.Jan;17(1):5 15(2016)中所述的那些。在一个示例中,融合蛋白可由多个转录激活子的拷贝和一个dCas9拷贝组成。
(i)用于转录激活的融合蛋白
可用于增加HTRA1 mRNA或蛋白质表达的转录因子结合在HTRA1启动子或增强子区中,并且该结合导致受试者细胞(例如RPE细胞、水平细胞或感光细胞)中的转录增加。术语“转录因子”包括结合在启动子区中的因子,包括通过反式激活域(“TAD”)激活转录的因子和在增强子区结合的因子,包括通过支架(诸如LIM元件)或通过其他机制激活转录的因子,所述支架可募集和组装附加的转录因子和染色质重塑蛋白以启动转录。参见,例如,Hiraiet al.,Structure and functions of powerful transactivators:VP16,MyoD and FoxAInt.J.Dev.Biol.2010;54(11-12):1589-1596。两类转录因子的成员包括将转录因子引导至靶启动子或增强子元件的DNA结合域和效应子域(例如,TAD或支架等)。
一方面,本发明利用融合蛋白,所述融合蛋白将核酸酶缺陷型Cas/指导RNA系统的DNA结合和识别特性与转录因子的转录效应子特性结合。本发明中所用的转录因子效应子结构域可以来自任何源,但通常来源于人或病毒转录因子或其工程化衍生物。可用于上调HTRA1转录的示例性转录因子包括:VP16、VP64、VP160(VP64由VP16的两个或更多个拷贝组成,而VP160由VP16的10个串联拷贝组成);MLL(UniProt ID Q00613)、E2A(UniProt IDP15923)、HSF1(UniProt ID Q00613)、NF-IL6(UniProt ID P17676)、NFAT1(UniProt IDQ13469)、NFIX(UniProt ID Q14938)、NF-kB(UniProt ID Q04206);MEF2A(Potthoff&Olson(2007),“MEF2:a central regulator of diverse developmental programs,”Development 2007;134(23):4131 4140;UniProt ID Q02078));和YY1(Weintraub et al.(2017),“YY1 Is a Structural Regulator of Enhancer-Promoter Loops”,Cell 2017;171:1573 88;UniProt ID P25490)、LHX2(ZIBETTI et al.,“Epigenomic profiling ofretinal progenitors reveals LHX2 is required for developmental regulation ofopen chromatin”,Communications Biology,April 25,2019,Pages 1-13,2,UniProt IDP50458)。
人类转录因子(TFs)或转录激活子是众所周知的且充分表征的。例如,Lambert etal.describe over 1,600likely human transcription factors(Lambert et al.,2018,“The Human Transcription Factors”Cell 172:650-665)。还参见Fulton et al.(2009),“TFCat:the curated catalog of mouse and human transcription factors”,GenomeBiol 2009;10:R29,Vaquerizas et al.(2009),“A census of human transcriptionfactors:function,expression and evolution”,Nat.Rev.Genet.2009;10:252-263,Wingender et al.(2015),“TFClass:a classification of human transcriptionfactors and their rodent orthologs”,Nucleic Acids Res.2015;43:D97-D102。转录因子结合基序是已知的,并在以下集合中进行了描述,诸如:TRANSFAC(Matys et al.(2006),“TRANSFAC and its module TRANSCompel:transcriptional gene regulation ineukaryotes”,Nucleic Acids Res.2006;34:D108-D110),JASPAR(Mathelier et al.(2016),“JASPAR 2016:a major expansion and update of the open-access databaseof transcription factor binding profiles”,Nucleic Acids Res.2016;44:D110-D115),HT-SELEX(Jolma et al.(2013),“DNA-binding specificities of humantranscription factors”,Cell 2013;152:327-339;Jolma et al.(2015),“DNA-dependent formation of transcription factor pairs alters their bindingspecificity”,Nature 2015;527:384-388;Yin et al.(2017),“Impact of cytosinemethylation on DNA binding specificities of human transcription factors”,Science 2017;356:eaaj2239),UniPROBE(Hume et al.(2015),“UniPROBE,update 2015:new tools and content for the online database of protein-binding microarraydata on protein-DNA interactions”.Nucleic Acids Re.2015;43:D117-D122),以及CisBP(Weirauch et al.(2014),“Determination and inference of eukaryotictranscription factor sequence specificity”,Cell 2014;158:1431-1443)。
其中转录因子效应子结构域与核酸酶缺陷型DNA结合结构域组合的融合蛋白也可使用除Cas蛋白之外的可编程核酸酶制备,包括例如利用TALEN和ZFN。
3.2指导RNA
在一些方法中,靶向部分包括具有与DNA靶互补的区域的RNA。RNA可被称为指导RNA(gRNA),且互补区域可被称为指导序列。gRNA的“指导序列”是赋予靶特异性的序列。它与靶序列的相反链杂交(即,它是其反向互补链)。在自然界中,许多CRISPR系统包括两个RNA分子:tracrRNA,其结合Cas核酸内切酶;以及crRNA,其结合DNA靶序列。一些CRISPR系统(例如,CRISPR Cas12a/Cpf1)仅需要crRNA。在研究和生物医学应用中,更典型的是使用嵌合单指导RNA(“sgRNA”),其是crRNA-tracrRNA融合,一般不需要RNase III和crRNA加工,并结合Cas和靶标两者。应当理解,除非从上下文显而易见,否则提及“sgRNA”包括使用具有适当结合特异性的任何合适的指导RNA(例如,sgRNA、crRNA或包含指导序列的其他RNA)的任何靶向方法。
最常用的sgRNA长度大约为100个碱基对。可编程靶向序列在sgRNA的5'端或附近包含大约20个碱基。通过对该序列进行编程,CRISPR Cas9系统或其他Cas系统可针对与该序列互补的任何基因组区域进行靶向。
用于设计靶向特定基因组区域的sgRNA的方法是本领域众所周知的。参见,Doenchet al.Nature Biotechnology 34:184-191,2016;Horlbeck et al.eLife.5,e19760(2016);Doench et al.,Nt.Biotechnol 34(2):184-191(2016);Cui et al.,“Review ofCRISPR/Cas9 sgRNA Design Tools.Interdiscip.Sci.2018,10:455465;Jensen,2018,Design principles for nuclease-deficient CRISPR-based transcriptionalregulators”FEMS Yeast Research,18:4,以及PCT专利公布WO2018107028。用于设计此类sgRNA的方法和工具也可例如从Dharmacon,Inc.商购获得;如在以下网站上公布,诸如:dharmacon.horizondiscovery.com/applications/crispra-transcriptional-activation-for-gene-overexpression or benchling.com/academic;以及Broadinstitute.org/gpp/public/analysis-tools/sgrna-design-help-crisprai。
一方面,本发明提供了指导RNA(gRNA),其包含具有至少10个对应于HTRA1启动子中的靶序列的连续核苷酸的指导序列。在一些情况下,指导RNA是sgRNA。在一些方法中,指导序列包含至少10个、至少15个、至少20个或至少25个核苷酸。在一些情况下,指导序列的长度为20个核苷酸。在一些实施方案中,本发明提供包含指导序列的指导RNA,其中指导RNA与Cas(例如Cas9)复合,所述Cas缺乏核酸酶活性(例如dCas9)。在一些情况下,本发明提供了与包括Cas DNA结合域和转录激活子的Cas融合蛋白复合的指导RNA,诸如sgRNA。通常,Cas是缺乏核酸酶的dCas(诸如dCas9)。
在一些情况下,DNA靶序列(例如,在HTRA1启动子或增强子中)与被Cas蛋白识别的前间隔序列邻近基序(PAM)相邻。例如,Cas9通常需要PAM基序NGG以获得活性。因此,在一些系统中,基于靶序列与PAM的相近性,某些靶序列(以及相应的某些指导序列)将是优选的。然而,一些Cas蛋白,包括Cas9的变体,具有灵活的PAM要求(参见Karvekis et al.,2019,“PAM recognition by miniature CRISPR-Cas14 triggers programmable double-stranded DNA cleavage.” bioRxiv.https://doi.org/10.1101/654897;Legut et al.,2020, “High-Throughput Screens of PAM-Flexible Cas9”,Cell Reports 30:28592868;Jakimo et al.,2018Cas9 with Complete PAM Recognition for AdenineDinucleotides bioRxiv doi.org/10.1101/429654;Esvelt KM,Mali P,et al.(2013)Orthogonal Cas9 proteins for RNA-guided gene regulation and editing.NatureMethods,10(11):1116 1121;Gleditzsch et al.,2019,PAM identification by CRISPR-Cas effector complexes:diversified mechanisms and structures.RNABiol.2019Apr;16(4):504 517;Tang et al.,2019,Efficient cleavage resolves PAMpreferences of CRISPR-Cas in human cells Cell Regeneration 8:44-50),和其他Cas蛋白是PAM非依赖性的(例如Cas14a1)。示例性PAM包括来自化脓性链球菌NGG的SpCas9;来自化脓性链球菌NRG的SpCas9;来自嗜热链球菌NNAGAAW的StCas9;来自脑膜炎奈瑟菌NNNNGATT的NmCas9;来自金黄色葡萄球菌NNGRRT的SaCas9;SaCas9变体(KKH SaCas9)NNNRRT;SpCas9 D1135E变体NGG;SpCas9 VRER变体NGCG;SpCas9 EQR变体NGAG;SpCas9 VQR变体NGAN或NGNG 3';来自酸胺球菌的AsCpf1,来自毛螺菌科TTTN的LbCpf1;来自新生弗朗西斯菌株U112 TTN和/或CTA的FnCpf1;来自四种主要分类群的C2c1:杆菌(Bacilli)、疣微菌门(Verrucomicrobia)、变形菌门(proteobacteria)和d-变形菌门TTN(N=A、T、C或G;R=A或G;W=A或T),其描述于Zhao et al.,2017,CRISPR-offinder:a CRISPR guide RNAdesign and off-target searching tool for user-defined protospacer adjacentmotif.Int J Biol Sci 2017;13(12):1470-1478中。因此,虽然常用的Cas9蛋白需要靶序列和连续NGG PAM,但其他天然存在或工程化Cas蛋白具有宽松的或不具有PAM要求。因此,通过明智地选择CRISPR/Cas系统,从业者将能够选择HTRA1转录调控区域中的几乎任何子序列作为招募转录激活子的靶。在一种方法中,HTRA1启动子靶存在于SEQ ID NO:13中。在一些方法中,HTRA1启动子靶存在于SEQ ID NO:5中。在一些方法中,HTRA1启动子靶存在于SEQ ID NO:8中。在一些方法中,HTRA1启动子靶存在于SEQ ID NO:13中。在一些方法中,HTRA1启动子靶存在于SEQ ID NO:7中。在一种方法中,HTRA1增强子靶存在于SEQ ID NO:14(2kb区)中。在一些方法中,HTRA1靶存在于SEQ ID NO:14(2kb区)或SEQ ID NO:34(4kb区)中。
根据本发明,指导序列与靶序列杂交,该靶序列包括HTRA1启动子序列(SEQ IDNO:5、SEQ ID NO:7、SEQ ID NO:8或SEQ ID NO:13)或SEQ ID NO:5、SEQ ID NO:7、SEQ IDNO:8或SEQ ID NO:13或HTRA1增强子序列(SEQ ID NO:14或34)的反向互补序列或SEQ IDNO:14或34的反向互补序列的至少10个、至少15个、至少20个或约20个连续核苷酸。
在一些情况下,指导序列包含上文给出的启动子序列的10个或更多个连续碱基。包含SEQ NO:13的10个连续碱基的序列的示例包括:GTCCCAACGG、TCCCAACGGA、CCCAACGGAT、CCAACGGATG、CAACGGATGC等,或其反向互补。包含SEQ ID NO:13的10个或更多个连续碱基的序列包括编码SEQ ID NO:13的第1至10位碱基、编码SEQ ID NO:13的第2-11位碱基、编码SEQ ID NO:13的第3-12位碱基的序列等。包含SEQ ID NO:13的10个或更多个连续碱基的序列包括其中核苷酸为SEQ ID NO:13的碱基X的序列,其中X为1至300,且10个连续碱基延伸至SEQ ID NO:13的X+Y,其中Y为9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25或更多,前提条件是指导序列长度通常小于50个碱基。可以用相同的方式描述15个或至少20个连续核苷酸的序列。例如,在一些情况下,指导序列包含SEQ NO:13的15个或更多个连续碱基(GTCCCAACGGATGC、TCCCAACGGATGCAC、CCCAACGGATGCACC、CCAACGGATGCACCA、CAACGGATGCACCAA)或其反向互补序列。
应当认识到,10、15或20个核苷酸序列中的每一个及其互补序列都可以在表中列出,并且单独的序列或序列的组合可独立地选自此类表中以包含在一个集合中,或从一个集合排除。即,此类表将描述并提供选择或排除单个序列和组合的基础。应当理解,以上描述旨在说明这些表格的位置并具有相同的内容。
如实施例9(§14.9)中所述,设计了sgRNA,其指导序列对应于SEQ ID NO:13中的靶区域,表示为SEQ ID NO:15-33,并测试了它们从HTRA1启动子激活转录的能力。在一些方法中,本方法中使用的gRNA靶向包含一个SEQ ID NO:15-33的序列。
在一些实施方案中,感兴趣的基因中的靶序列可与sgRNA的指导区互补。一般来讲,gRNA的指导序列与其对应的靶序列之间存在精确的互补性或同一性,所述互补性或同一性可小于100%。在一些实施方案中,sgRNA的指导区与其对应的靶序列之间的互补性或同一性的程度可小于100%,但需要100%的同一性以避免脱靶效应。在一些实施方案中,sgRNA的指导区和感兴趣的基因的靶区域可以至少95%相同(例如,20个中的一个错配)、至少90%相同或至少85%相同。
3.3递送
将与转录激活子蛋白和gRNA融合的Cas蛋白(例如,Cas9和dCas9)递送至细胞的方法是已知的。融合蛋白可以蛋白质形式递送(例如,通过显微注射)。更常见的是,融合蛋白以DNA形式递送,例如以可引入RPE或脉络膜细胞中的合适载体形式。一般来讲,将编码gRNA的DNA克隆到启动子(例如U6启动子)下游的载体中进行表达。在一些方法中,递送通过病毒,例如慢病毒、腺相关病毒(AAV)进行,如Byrne et al.,Methods Enzymol.546,119-38(2014);Cong et al.,Science(80).339,819-823;Hirsch et al.,Mol.Ther.18,6-8(2010)中所述的。在一些方法中,递送通过来源于细胞的纳米囊泡或其他方法来进行。另外参见下文第7节。
3.3协同激活介质
在一些方法中,称为协同激活介质(SAM)的CRISPRa系统用于增加Chr 10AMD患者中HTRA1的表达。SAM系统使用多个转录因子,其可进一步提高Cas9介导的基因激活的效力。参见,Konermann et al.,Nature,January 29;517(7536)2015),相关的公开内容以引用方式并入本文。在一些情况下,该系统使用两种质粒,一种编码sgRNA和dCas9转录激活子,例如dCas9-VP64分子,而另一种编码MS2-TAD融合蛋白。MS2-TAD包含MS2多肽和至少一个转录激活域。在一些方法中,MS2-TAD融合蛋白包含MS2多肽(SEQ ID NO:56)和p65和HSF1的反式激活结构域(MS2-p65-HSF1)。用于SAM系统中的sgRNA包含一个或多个MS2结合序列(例如,SEQ ID NO:55),其可结合到MS2-TAD融合蛋白。在一些实施方案中,gRNA中包括两个MS2结合序列:一个在四环中,一个在gRNA的茎环上。编码MS2-TAD(例如,MS2-p65-HSF1)的质粒可商购获得,例如LentiMPH质粒(Addgene,Watertown,MA,USA)。可通过将合适的sgRNA编码序列克隆到携带dCas9-VP64编码序列的质粒中来构建编码sgRNA和dCas9-VP64融合蛋白的质粒。携带dCas9-VP64编码序列的质粒也是可商购获得的,例如来自Addgene(Watertown,MA,USA)的LentiSAM质粒。sgRNA编码序列可克隆到LentiSAM质粒中。
3.5增强子结合位点
在一些方法中,通过将转录激活子与HTRA1 2kb(增强子)区域中的位点结合,可增加从内源性HTRA1基因的转录。如下文详细讨论的,发明人已经鉴定了转录激活子LHX2的结合基序,其与rs36212733的风险等位基因重叠。几条遗传和实验证据表明,相对于与非风险等位基因的结合,LHX2结合到风险等位基因不存在或减弱,这导致HTRA1的表达减少。在一种方法中,将与Cas蛋白融合的LHX2 LIM结构域与适当的指导RNA结合,并募集到携带风险等位基因的患者的LHX2风险结合位点。据信,LHX2传递到风险等位基因结合位点附近导致基因表达增加,因为转录激活子的束缚有效地增加了LHX2的局部浓度,消除或减轻了风险变化对结合的负面影响,并导致HTRA1表达增加。在另一种方法中,LHX2转录激活子被修饰以有效结合风险等位基因序列,从而导致转录增加。
部分基于以下发现:与转录激活子结合位点一致的风险等位基因可降低或以其他方式影响转录激活子的结合并导致HTRA1表达降低,并且将相应的转录激活子募集到该位点可增加HTRA1表达,对2kb区域进行了查看以鉴定与转录激活子结合基序一致的其他风险相关序列变异。对rs144224550、rs10490924、rs3750848和rs6212733处的风险等位基因(即风险相关变体)的分析鉴定出若干与风险相关的SNP一致的结合基序(除了LXH2基序之外)。
rs144224550(10:122455084-122455085);Nkx2-5(var2);NFIX;
rs36212733(10:122455695);MEF2A;HOXB4;ALX3;HOXD3;LBX1;HOXD4;VAX1;LBX1;VSX1;VSX2,LHX9;MNX1;PDX1;EMX1;PRRX2;BARX2;
rs10490924(10:122454932)RHOXF;Myog;TCF12;ASCL1;Ascl2;NHLH1;BHLHA15var2
rs3750848(10:122455799);YY1;BARX2;
根据本发明的方面,来自上文列出的转录激活子的效应子结构域,诸如来自LXH2的LIM结构域,被定位(例如,使用CRISPRa)至相应的结合基序以增加HTRA1表达。值得注意的是,基于RNA-Seq数据,NFIX、MEF2A和YY1在FPKM大于1的RPE中表达。在本发明的各个方面,HTRA1表达通过NFIX、MEF2A和YY1中的任一个与其相应结合基序的结合而增加。
一般来讲,合适的指导RNA的指导序列将结合靶序列,其中末端在风险相关核苷酸的20个碱基以内,有时在50个碱基以内,有时在100个碱基以内。
3.6其他可编程内切核酸酶
本发明的方法一般在CRISPR/Cas系统的上下文中有所描述。也可使用用于(a)核酸的序列特异性(“可编程”)修饰和(b)转录激活子到启动子和增强子区域的序列特异性募集的其他系统。此类系统的示例包括锌指核酸酶系统、转录激活子样效应核酸酶(TALEN)系统、归巢核酸内切酶系统、大范围核酸酶系统或Cre重组酶系统(例如,隐蔽loxP位点之间的Cre诱导的重组)。使用锌指核酸酶、TALEN、大范围核酸酶和DNA指导的多肽(诸如格雷戈里纳诺杆菌(NgAgo))来修饰眼中细胞中的序列或表达在Yanik et al.,2017,In vivogenome editing as a potential treatment strategy for inherited retinaldystrophies Progress in Retinal and Eye Research56:1-18中有所描述。还参见,Lloyd et al.,Frontiers in Immunology,4(221),1-7(2013);Urnov et al.,2010,“Genome editing with engineered zinc finger nucleases,”Nat Rev Genet.11(9):636-46;Sun et al.,2013,“Transcription activator-like effector nucleases(TALENs):a highly efficient and versatile tool for genome editing,”BiotechnolBioeng.110(7):1811-21;Sengupta et al.,2017,“Viral Cre-LoxP tools aid genomemodification in mammalian cells”J.Biological Engineering 11:45(描述了Cre-Lox的慢病毒和腺相关病毒传递系统)以及Nagy 2000,“Cre recombinase:the universalreagent for genome tailoring,”Genesis,26(2):99-109,每篇文献以引用方式并入本文。这些修饰包括通过同源重组、非同源末端连接、同源定向修复、组蛋白修饰、转录激活、RNA编辑、转录抑制来编辑的靶序列。此外,Thakore et al.2018in“RNA-guidedtranscriptional silencing in vivo with S.aureus CRISPR-Cas9 repressors”NatCommun.9(1):1674以及Amabile et al.2016in“Inheritable Silencing of EndogenousGenes by Hit-and-Run Targeted Epigenetic Editing”Cell 167(1):219232.e14描述了不具有切割的工程化示例。
在一些实施方案中,靶向DNA的分子包含一种或多种锌指蛋白(ZFP)或其结构域,其以序列特异性方式结合到DNA并与核酸酶融合。ZFP或其结构域是通过一个或多个锌指以序列特异性方式结合DNA的蛋白质或较大蛋白质内的结构域,其结构通过锌离子的配位而稳定的结合结构域内的氨基酸序列区域。术语锌指DNA结合蛋白通常缩写为锌指蛋白或ZFP。
在ZFP中,有靶向特定DNA序列的人工ZFP结构域,通常为9-18个核苷酸长,由单独指的组装产生。ZFP包括其中单指结构域的长度为大约30个氨基酸并包含α螺旋的那些,该螺旋包含两个不变的组氨酸残基,其通过锌与单一β转角的两个半胱氨酸配位,并具有两个、三个、四个、五个或六个指。通常,ZFP的序列特异性可通过在锌指识别螺旋上的四个螺旋位置(-1、2、3和6)进行氨基酸取代来改变。因此,在一些实施方案中,ZFP或含有ZFP的分子是非天然存在的,例如,被工程化成与选择的靶位点结合。参见,例如,Beerli et al.(2002)Nature Biotechnol.20:135-141;Pabo et al.(2001)Ann.Rev.Biochem.70:313-340;Isalan et al.(2001)Nature Biotechnol.19:656-660;Segal et al.(2001)Curr.Opin.Biotechnol.12:632-637;Choo et al.(2000)Curr.Opin.Struct.Biol.10:411-416;美国专利6,453,242;6,534,261;6,599,692;6,503,717;6,689,558;7,030,215;6,794,136;7,067,317;7,262,054;7,070,934;7,361,635;7,253,273以及美国专利公布2005/0064474;2007/0218528;2005/0267061,所有文献全文以引用方式并入本文。
在一些实施方案中,靶向DNA的分子为或包含与DNA切割域融合以形成靶向核酸酶的锌指DNA结合域、TALEN或其他靶向DNA的蛋白质。在一些实施方案中,融合蛋白包含来自至少一种IIS型限制酶和一种或多种DNA靶向蛋白的切割结构域(或切割半结构域)。在一些实施方案中,切割结构域来自IIS型限制性内切核酸酶Fok I。Fok I通常催化DNA的双链切割,在一条链上距其识别位点9个核苷酸处和在另一条链上距其识别位点13个核苷酸处。参见,例如,美国专利5,356,802;5,436,150和5,487,994;以及Li et al.(1992)Proc.Natl.Acad.Sci.USA89:4275-4279;Li et al.(1993)Proc.Natl.Acad.Sci.USA 90:2764-2768;Kim et al.(1994)Proc.Natl.Acad.Sci.USA 91:883-887;Kim et al.(1994)J.Biol.Chem.269:31,978-31,982。
术语“可编程核酸酶”可指CRISPR家族Cas核酸酶或其衍生物、转录激活子样效应核酸酶(TALEN)或其衍生物、锌指核酸酶(ZFN)或其衍生物、以及归巢核酸内切酶(HE)或其衍生物。
4.用非风险形式替换风险形式的基因编辑
在一些方法中,治疗Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法包括施用药剂以通过利用对应的非风险等位基因转化或置换风险等位基因中的一种或多种来修饰患者RPE细胞的基因组DNA。在某些实施方案中,等位基因在2.0kb风险区域中(参见,例如,实施例5和图13)。
4.1风险等位基因的修复
在一些方法中,使用了靶向2kb区域中一个或多个风险等位基因的gRNA。这些风险等位基因包括但不限于,rs10490924(风险等位基因为T)、rs144224550(风险等位基因为GT插入)、rs36212731(风险等位基因为T)、rs36212733(风险等位基因为C)、rs875084(风险等位基因为G)、rs3750847(风险等位基因为T)和rs3750846(风险等位基因为C)。可用于靶向这些风险等位基因的示例性sgRNA/指导序列示于下表1中。核苷酸以粗体形式存在于风险等位基因中。

可将包含非风险等位基因序列的sgRNA、Cas蛋白和模板修复多核苷酸引入一种或多种病毒载体中的RPE细胞。模板修复序列一般需要一定量的切割位点的每一侧上的重叠(同源性)。对于单个核苷酸的修复而言,供体模板的“同源臂”应约为200-500个核苷酸。对于更大的修复(例如整个2kb区域)而言,每个同源臂应当为大约500-800个核苷酸。这导致风险等位基因被野生型等位基因取代,从而将Chr10 AMD患者RPE细胞中的HTRA1表达恢复到正常水平。示例性修复序列如表2所示。
表2:示例性修复序列
| 靶SNP | 修复序列 | SEQ ID NO: |
| rs10490924 | gatcccagctGctaaaatcca | 87 |
| rs144224550 | attctggagtGGtgccctgcag | 88 |
| rs36212731 | atattctcacGgctttccagt | 89 |
| rs36212732 | tgtgcttgccAtagtatatat | 90 |
| rs36212733 | gtatatataaTtagacaaatg | 91 |
| rs3750848 | tgattcaatgTtaaaccattt | 92 |
| rs3750847 | caagacctttCggtggctgcc | 93 |
| rs3750846 | ggactgctggTctcatgcaac | 94 |
应当理解,指导序列不必与靶SNP直接重叠。如果sgRNA靠近或邻近SNP,这将足以允许对有缺陷的SNP进行同源定向修复。在这种情况下,“邻近”是指与指导序列核苷酸杂交的最接近的核苷酸在距SNP或其他修复位点25个核苷酸内,优选在20个核苷酸内,并且有时在15或10个核苷酸内。
4.2HTRA1转录调控区域中大区域的置换
在一些方法中,在CRISPR/Cas9系统中使用一对gRNA以去除来自Chr10 AMD患者的RPE细胞中的整个2kb风险区域。该sgRNA对被设计成靶向两个核苷酸位置,由所述两个位置定义的区域包含2kb风险区域。在一些方法中,sgRNA是SEQ ID NO:50和SEQ ID NO:52。在一些方法中,sgRNA对是SEQ ID NO:51和SEQ ID NO:52。将sgRNA和Cas9对引入RPE细胞导致包含2kb区域的区域被去除。将编码与删除区域相对应的非风险序列并在5'切割位点上游和3'切割位点下游包含500-800nt附加基因组序列的质粒引入RPE细胞,使得非风险序列通过同源定向修复插入缺失区域,从而导致2kb风险区域被非风险序列置换。sgRNA对、Cas9和编码非风险模板序列的质粒可引入相同或不同的病毒载体中,并且可同时或依次引入以增加Chr10 AMD患者的RPE细胞中的HTRA1表达。

5.CRISPR/Cas系统
在一些方法中,CRISPR技术用于将一个或多个核苷酸置换引入携带Chr 10风险等位基因的个体中的RPE或脉络膜细胞的基因组DNA中,从而用相应的非风险等位基因置换一个或多个风险等位基因。“CRISPR/Cas”系统是指一类广泛用于防御外来核酸的细菌系统。CRISPR/Cas系统包括I型、II型和III型亚型。野生型II型CRISPR/Cas系统使用RNA介导的核酸酶,例如Cas9,其与指导和激活RNA复合以识别和切割外源核酸。Cas蛋白的非限制性示例包括Cas1、Cas1B、Cas2、Cas3、Cas4、Cas5、Cas6、Cas7、Cas8、Cas9(也称为Csn1和Csx12)、Cas10、Csy1,Csy2、Csy3、Cse1、Cse2、Csc1、Csc2、Csa5、Csn2、Csm2、Csm3、Csm4、Csm5、Csm6、Cmr1、Cmr3、Cmr4、Cmr5、Cmr6、Csb1、Csb2、Csb3、Csx17、Csx14、Csx10、Csx16、CsaX、Csx3、Csx1、Csx15、Csf1、Csf2、Csf3、Csf4、其同系物或其经修饰的版本。这些酶是已知的。例如,酿脓链球菌Cas9蛋白的氨基酸序列可在SwissProt数据库中找到,登录号为Q99ZW2。Cas9蛋白中突变的非限制性示例是本领域已知的(参见,例如WO2015/161276),根据所提供的方法,这些突变中的任一个均可包括在CRISPR/Cas9系统中。Cas9同系物存在于多种真细菌中,其包括但不限于以下分类群的细菌:放线菌(Actinobacteria)、产水菌(Aquificae)、拟杆菌-绿菌(Bacteroidetes-Bhlorobi)、衣原体-疣微菌(Bhlamydiae-Verrucomicrobia)、绿屈挠菌(Chlroflexi)、蓝藻细菌(Cyanobacteria)、厚壁菌(Firmicutes)、变形菌(Proteobacteria)、螺旋体(Spirochaetes)、和热孢菌(Thermotogae)。示例性cas9蛋白是酿脓链球菌(streptococcu spyogenes)cas9蛋白。其他cas9蛋白和其同源物描述于,例如Chylinksi,et al.,RNA Biol.2013May 1;10(5):726 737;Nat.Rev.Microbiol.2011June;9(6):467-477;Hou,et al.,Proc Natl Acad Sci U S A.2013Sep 24;110(39):15644-9;Sampson et al.,Nature.2013May9;497(7448):254-7;以及Jinek,et al.,Science.2012Aug17;337(6096):816-21.Cas9。Cas9及其同源物可与sgRNA一起使用将特异性修饰引入对感兴趣的基因组区域中,例如,包含一个或多个Chr 10风险等位基因的基因组区域。
还可用于CRISPR/Cas系统以转换2kb风险区域中的风险等位基因的其他RNA介导的核酸酶包括,例如Cas 12a和Cascade/Cas3。参见,Pickar-Oliver and Gersbach,Nature,vol.20,August 2019,其相关部分以引用方式并入本文。Cas 12a识别与在3'PAM旁边定位的crRNA(sgRNA)中的间隔区互补的靶序列。目标识别导致产生交错的DNA双链断裂。Cascade/Cas 3是多聚体复合物,其靶向DNA,该DNA与crRNA的间隔部分具有互补性,并且位于3'PAM旁边并产生单链切口。
因此,在一些方法中,该方法包括将CRISPR系统引入个体的RPE细胞中,其中该系统可包含与靶序列杂交的Cas9蛋白和指导RNA(例如,sgRNA)。sgRNA和Cas9可从系统的相同或不同载体表达。此外,编码与切割位点重叠的非风险核苷酸序列的供体载体与CRISPR-Cas9质粒共递送。指导RNA靶向包含或接近风险等位基因的序列,并且Cas9蛋白切割基因组DNA分子。使用供体序列质粒通过同源重组修复经切割的基因组DNA,从而导致基因组序列从风险等位基因向非风险等位基因的改变。上述风险等位基因中的任一者均可转化为非风险等位基因。在一些方法中,整个2.0kb区域(参见实施例5)或整个4kb AMD风险区域(参见实施例3)被转化,使得其不包含Chr 10风险等位基因,这恢复了RPE细胞中的HTRA1表达。
5.1Cas9切口酶
在一些方法中,可使用基于Cas9切口酶的CRISPR/Cas系统来纠正单独的风险等位基因。与未经修饰的Cas9蛋白相比,cas9切口酶蛋白在其核酸酶结构域中的一者中包含突变。因此,Cas9切口酶蛋白将“切割”靶DNA,即在靶位点导致单链DNA切割,而不是像未修饰的Cas9那样导致双链切割。在一个示例中,dCas9在其两个核酸酶结构域中的每一者中都包含突变,然而Cas9切口酶仅在其一个核酸酶结构域中包含突变。与本文所述的其他CRISPR方法一样,指导RNA能够将Cas9切口酶靶向到特异性基因组位置。cas9切口酶蛋白与碱基编辑蛋白一起引入,诸如胞苷脱氨酶(例如APOBEC1、AID、APOBEC3G或CDA1),其将胞嘧啶碱基转化为尿嘧啶碱基(其具有胸腺嘧啶碱基的碱基配对特性)和腺嘌呤脱氨酶,其将腺苷碱基转化为肌苷碱基(其具有鸟苷的碱基配对特性)。这些转换具有将C-G碱基对变为T-A碱基对(胞苷脱氨酶)或将A-T碱基对变为G-C碱基对(腺嘌呤脱氨酶)的效果。在一些情况下,基于CRISPR/Cas系统的Cas9切口酶还包括细胞DNA修复反应的抑制剂,以促进碱基编辑效率。例如,尿嘧啶DNA糖基化酶抑制剂可用于防止细胞中的尿嘧啶DNA糖基化酶催化将尿嘧啶碱基从DNA中去除。去除尿嘧啶可导致编辑后的尿嘧啶回复为胞嘧啶。基于Cas9切口酶的基因编辑系统是众所周知的并且描述于例如Komor等人,Nature,533(7603):420-424(2016)中。在一个示例中,该系统包含融合至具有16个氨基酸残基的XTEN接头的Cas9切口酶的N端的胞苷脱氨酶(APOBEC1),以及融合至Cas9切口酶的C端的尿嘧啶DNA糖基化酶抑制剂(UGI)(例如,Komor Et al.,Nature,533(7603):420-424(2016)中的编辑子BE3)。此类系统能够以低插入缺失(即小的缺失或插入)形成率和脱靶活性(即在除由指导RNA靶向的基因座之外的基因组位置进行编辑或插入缺失形成),有效地将体内的C-G碱基对编辑为T-A碱基对。在一些情况下,与Cas9切口酶连接的编辑蛋白可靶向多个碱基,这具体取决于接头序列的长度。例如,与具有16个残基XTEN接头的dCas9融合的APOBEC1可在大约5个核苷酸的窗口内(通常从原型间隔序列中的位置4到8,将原型间隔序列的远端末端计数为位置1)使胞嘧啶碱基脱氨基(参见,例如,Komor et al.,Nature,533(7603):420-424(2016))。在一些方法中,可将指导RNA序列设计成在此类窗口内靶向一个或多个碱基。
在一些方法中,表达Cas9切口酶和指导RNA的载体序列作为转基因整合到宿主细胞DNA中。例如,编码上述APOBEC1-XTEN-Cas9切口酶-UGI融合物的基因序列,连同指导RNA,可整合到质粒中并转染到群体中的细胞中。在一些方法中,CRISPR碱基编辑系统组件的递送通过病毒,例如慢病毒或腺相关病毒(AAV)。在一些情况下,编码CRISPR碱基编辑系统组分的DNA序列太大而无法封装在AAV中,所述AAV具有小于5千碱基的基因组包装尺寸限制。在一种方法中,可使用采用内含肽序列的双AAV策略来递送CRISPR基础编辑系统组分。内含肽是蛋白质的片段,其能够自我切除并将剩余蛋白质部分拼接在一起。前体蛋白的内含肽可来自两个基因,在这种情况下,它们被称为断裂内含肽。在此类示例双AAV策略的一个中,如Levy et al.,Nat.Biomed.Eng.,4(1):97-110(2020)中所述,胞嘧啶碱基编辑器(例如上述BE3)可以分成两半,其中每一半与快速剪接分裂内含子的一半融合。编码这些产物的序列可与表达指导RNA的序列一起整合到两个单独的AAV基因组中,并共同转导到细胞中。当两种产物在细胞中表达时,它们可以拼接在一起,从而重建全长碱基编辑器诸如BE3。
6.通过引入外源性HTRA1蛋白编码多核苷酸使用基因疗法增加HTRA1表达
在一个实施方案中,治疗AMD涉及增强RPE细胞中HTRA1表达的基因疗法。基因治疗是一种众所周知的技术,并描述于例如,Moore et al.,2017,“Gene therapy for age-related macular degeneration”Expert Opinion on Biological Therapy 17:10:1235-1244;Aponte-Ubillus et al.,2018,“Molecular design for recombinant adeno-associated virus(rAAV)vector production”Applied microbiology andbiotechnology 102.3:1045-1054;Ochakovski et al.,2017,“Retinal Gene Therapy:Surgical Vector Delivery in the Translation to Clinical Trials”Frontiers inNeuroscience 11; et al.,2015, “Retinal gene delivery by adeno-associatedvirus(AAV)vectors:Strategies and applications” European Journal ofPharmaceutics and Biopharmaceutics 95:343 352;Naso et al.,2017,“Adeno-Associated Virus(AAV)as a Vector for Gene Therapy”BioDrugs 31:317;Dunbar etal.,2018,“Gene therapy comes of age”Science 359:6372;Penaud-Budloo et al.,2018.,“Pharmacology of Recombinant Adeno-associated Virus Production”Molecular Therapy:Methods&Clinical Development 8:166-180中,所述文献中的每一者以引用方式并入用于所有目的。
et al.,2015, “Retinal gene delivery by adeno-associatedvirus(AAV)vectors:Strategies and applications” European Journal ofPharmaceutics and Biopharmaceutics 95:343 352;Naso et al.,2017,“Adeno-Associated Virus(AAV)as a Vector for Gene Therapy”BioDrugs 31:317;Dunbar etal.,2018,“Gene therapy comes of age”Science 359:6372;Penaud-Budloo et al.,2018.,“Pharmacology of Recombinant Adeno-associated Virus Production”Molecular Therapy:Methods&Clinical Development 8:166-180中,所述文献中的每一者以引用方式并入用于所有目的。
6.1外源性HTRA1蛋白的表达
在一些方法中,进行基因治疗以在RPE细胞中引入外源性HTRA1蛋白。在一些方法中,引入的外源性HTRA1具有与天然HTRA1蛋白(SEQ ID NO:2)相同的氨基酸序列。在一些方法中,外源性HTRA1蛋白具有与SEQ ID NO:2不同但与SEQ ID NO:2的氨基酸序列共享至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%或至少99%的氨基酸序列同一性。在一些方法中,外源性HTRA1蛋白不同于SEQ ID NO:2(即HTRA1变体)但保持HTRA1的丝氨酸蛋白酶活性。HTRA1的丝氨酸蛋白酶活性可使用本领域众所周知的方法测量,例如,如Grau et al.,Proc.Natl.Acad.Sci.U.S.A.April 26,102(17)6021-6026(2005)中所述的。出于本公开的目的,保持丝氨酸蛋白酶活性是指外源性HTRA1蛋白具有至少50%、至少60%、至少70%、至少80%、至少90%或至少95%天然HTRA1蛋白(SEQ ID NO:2)的丝氨酸蛋白酶活性。
在一些方法中,基因疗法包括使用包含核酸序列(“货物”)的载体,所述核酸序列编码SEQ ID NO:2的外源性HTRA1蛋白或如上所述的变体,诸如具有HTRA1的丝氨酸蛋白酶活性的变体。在一些方法中,核酸序列包括SEQ ID NO:1。在一些方法中,核酸序列共享显著的序列同一性,例如与SEQ ID NO:1的至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%或至少99%的核酸序列同一性。在一些方法中,编码外源性HTRA1蛋白的核酸序列与SEQ ID NO:2具有显著的序列同一性,例如与SEQ ID NO:2的至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%或至少99%序列同一性。在一些情况下,编码HTRA1蛋白的核酸包括包含相对于天然HTRA1核酸序列的缺失、插入或置换的序列,这仍然产生编码具有丝氨酸蛋白酶活性的多肽的多核苷酸。
HTRA1核酸和多肽也可使用本领域技术人员众所周知的方法通过化学或酶促改变进行修饰。例如,可通过添加脂质、糖、肽、有机或无机化合物,通过包含经修饰的核苷酸或氨基酸等来修饰序列。因此,HTRA1核酸和蛋白质也可与另一部分(例如,报告基因蛋白)缀合,其存在可有利于检测或用于其他目的。
6.2用于基因疗法的载体
适用于引入HTRA1编码序列的病毒载体包括但不限于腺病毒、AAV2病毒、慢病毒、牛乳头瘤病毒(BPV-1)或Epstein-Barr病毒(pHEBo、pREP衍生的和p205)。可以使用非病毒系统,诸如配制成微粒的裸DNA,参见见下文第7节。
6.3用于基因疗法载体的启动子
在一些方法中,递送的转基因(例如,外源HTRA1基因,或重组核酸,其在RPE细胞中的表达可增加内源HTRA1基因的表达)包含可操作连接到启动子序列的蛋白质编码序列。在一些方法中,启动子与HTRA1多核苷酸是异质的。在一些方法中,启动子是天然HTRA1启动子(例如,SEQ ID NO:13)。在一些方法中,启动子是诱导型启动子。在一些方法中,启动子是组成型启动子。启动子可以为天然存在的启动子或结合多于一种启动子的元素的杂合启动子。在一些方法中,启动子是组织特异性启动子。一般来讲,启动子是人内源性RPE特异性启动子序列的缩短版本(例如,RPE65-5022核苷酸和BEST1-5479核苷酸)。RPE特异性启动子的非限制性示例包括BEST1-EP-454;RPE65-EP-415;smCBA;CBA;RPE65-EP-419;sctmCBA;或VMD2,如国际专利公布WO2020019002中所述。其他启动子或修饰的启动子(包括天然的和合成的)也可用于控制本文所公开的治疗产物的表达,包括但不限于UBC、GUSB、NSE、突触蛋白、MeCP2、GFAP、PAI1、ICAM、flt-1和CFTR(参见,Papadakis et al 2004;Promoters andControl Elements:Designing Expression Cassettes for Gene Therapy in CurrentGene Therapy,2004,4,89-113;Gray&Samulski 2011;Vector Design andConsiderations for CNS Applications in Gene Vector Design and Application toTreat Nervous System Disorders,ed.J.Glorioso(Washington,DC:Society forNeuroscience),1-9.;Trapani et al2014;Vector platforms for gene therapy ofinherited retinopathies Progress in Retinal and Eye Research 43(2014)108e128;Powell and Gray 2015)。Viral Expression Cassette Elements to Enhance TransgeneTarget Specificity and Expression in Gene Therapy Discov.Med.2015January;19(102):49 57,其各自以引用方式并入本文。还可使用的其他启动子包括SV40的早期和晚期启动子、tet启动子、腺病毒或巨细胞病毒立即早期启动子、RSV启动子、由T7 RNA聚合酶指导表达的T7启动子、λ噬菌体的主要操纵子和启动子区域,fd外壳蛋白的控制区、3-磷酸甘油酸激酶或其他糖酵解酶的启动子、酸性磷酸酶的启动子(例如Pho5)、酵母α-交配因子的启动子、杆状病毒系统和已知控制原核或真核细胞或其病毒及其各种组合的基因表达的其他序列的多面体启动子。
外源HTRA1基因或转基因也可受其他调控序列的控制,诸如增强子或激活子序列、前导序列或信号序列、核糖体结合位点、转录起始和终止序列以及聚腺苷酸化序列。可用于本发明方法的增强子包括但不限于:SV40增强子、巨细胞病毒(CMV)增强子、延伸因子1(EF1)增强子、酵母增强子、病毒基因增强子等。终止控制区可包含或来源于合成序列、合成聚腺苷酸化信号、SV40晚期聚腺苷酸化信号、SV40聚腺苷酸化信号、牛生长激素(BGH)聚腺苷酸化信号、病毒终止子序列等。
示例性启动子和增强子核苷酸序列以SEQ ID NO:6、11、12和13(“启动子序列”)提供。本领域技术人员将理解,调控(启动子/增强子)序列可容忍一定程度的变异,同时保留调控特性。在其中调用启动子/增强子的本文所述的某些方法中,基本上相同的序列(例如,在整个启动子/增强子序列范围内,具有至少约90%同一性,优选至少约91%、92%、93%、94%、95%、96%、97%、98%、99%核苷酸同一性)被认为是调用序列的合适替代物。
7.CRISPR、CRISPRa、核酸内切酶、修复模板和其他组件的递送
用于递送蛋白质和核酸(包括CRISPR元件)的系统(“递送系统”)是本领域众所周知的。这些系统可用于向细胞(例如、RPE、感光细胞和水平细胞)递送Cas蛋白(具有或不具有核酸酶活性)、Cas切口酶、sgRNA或其他指导RNA、Cas转录激活子融合蛋白、HTRA1蛋白编码序列、模板修复序列等。递送至细胞的物质有时在本文中被称为“转基因”或“货物”。Hageman,G and Richards,B.,International Patent Publication WO2020019002,andYanik et al.,2017,In vivo genome editing as a potential treatment strategyfor inherited retinal dystrophies Progress in Retinal and Eye Research 56:1-18描述了可适于本发明的用于将组分递送至眼部中的细胞以用于转基因表达、基因修复、基因激活等的方法。在一些方法中,递送通过病毒,例如慢病毒、腺相关病毒(AAV)进行,如Byrne et al.,Methods Enzymol.546,119-38(2014);Cong et al.,Science(80).339,819-823;Hirsch et al.,Mol.Ther.18,6-8(2010)中所述的。
在一些方法中,使用rAAV2表达载体递送货物(例如,HTRA1转基因)。在一种方法中,使用rAAV2系统将转基因或其他组分(例如外源性HTRA1基因,或在RPE细胞中的表达可增加内源性HTRA1表达的重组核酸)递送至RPE能够以高效率转导RPE细胞。除AAV2之外,其他基于腺相关病毒的载体包括AAV1、AAV2、AAV3、AAV4、AAV5、AAV6、AAV7、AAV8、AAV9、AAV10、AAV11和假型AAV。还可以采用逆转录病毒、慢病毒、腺病毒、杆状病毒等,如Lau and Suh(2017)doi:10.12688/f1000research.11243.1中所公开的,其相关公开内容通过引用并入本文。
为了将转基因包装到AAV载体中,ITR是与转基因相同的构建体中所需的顺式的唯一AAV组件。cap和rep基因可以反式提供。因此,可以设计DNA构建体,使得AAV ITR位于抗病原体构建体(或其亚基,或其与作为可调节启动子的一部分的二聚化结构域的亚基)的编码序列的侧部,从而定义扩增和包装的区域,唯一的设计限制是待包装的DNA大小的上限(约4.5kb)。
除了AAV载体之外,可使用的其他病毒载体包括但不限于逆转录病毒、腺病毒(AdV)、慢病毒、痘病毒、甲病毒和疱疹病毒。
可使用本领域已知的方法(包括本文引用的出版物中描述的方法)生产、收集和纯化携带带有HTRA1转基因或其他货物的表达盒的病毒载体(例如,rAAV2、慢病毒载体)。对于AAV方法,参见,Zolotukin et al.,2002,Production And Purification Of Serotype 1,2,And 5Recombinant Adeno-Associated Viral Vectors”Methods 28:158-167,incorporated by reference,and,Aponte-Ubillus et al.,2018;Naso et al.,2017;以及Penaud-Budloo et al.,2018;所有文献均以引用方式并入本文并在上文中引用。对于涉及基因疗法(包括对表达和递送系统的描述)的一般综述,还参见,Moore et al.,2017,"Gene Therapy For Age-Related Macular Degeneration"Expert Opinion onBiological Therapy17:10:1235-1244;Aponte-Ubillus et al.,2018,"MolecularDesign For Recombinant Adeno-Associated Virus(Raav)Vector Production"Appliedmicrobiology and biotechnology 102.3:1045-1054;Ochakovski et al.,2017,"Retinal Gene Therapy:Surgical Vector Delivery In The Translation To ClinicalTrials"Frontiers in Neuroscience 11;Schon et al.,2015,"Retinal Gene DeliveryBy Adeno-Associated Virus(Aav)Vectors:Strategies And Appucations"EuropeanJournal of Pharmaceutics and Biopharmaceutics 95:343-352;Naso et al.,2017,"Adeno-Associated Virus(Aav)As A Vector For Gene Therapy"BioDrugs 31:317;Dunbar et al.,2018,"Gene Therapy Comes Of Age"Science 359:6372;Penaud-Budlooet al.,2018.,"Pharmacology of Recombinant Adeno-Associated Virus Production"Molecular Therapy:Methods&Clinical Development 8:166-180;其中每一者以引用方式并入用于所有目的。
非病毒载体或方法也可用于递送货物。这些包括使用病毒样颗粒(VLP)递送、使用阳离子脂质体施用、细胞衍生的纳米囊泡、直接核酸注射、流体动力学注射、使用核酸缩合肽和非肽、阳离子脂质体和脂质体中的封装。在一种方法中,病毒样颗粒(VLP)用于递送货物。VLP包含病毒载体的工程化版本,其中核酸或非核酸货物通过另选的机制(例如,mRNA募集、蛋白质融合、蛋白质-蛋白质结合)包装到VLP中。参见,See Itaka and Kataoka,2009,"Recent development of nonviral gene delivery systems with virus-likestructures and mechanisms,"Eur J Pharma and Biopharma 71:475-483;以及Keeleret al.,2017,“Gene Therapy2017:Progress and Future Directions”Clin.Transl.Sci.(2017)10,242 248,以引用方式并入。
8.用于增加HTRA1表达或活性的其他疗法
8.1细胞疗法
在一种方法中,干细胞(例如,iPSC)在体内和体外修饰以移植到RPE(参见Peddleet al.,“CRISPR Interference Potential Application in Retinal Disease”,Int.J.Mol.Sci.,2020 21:1-14)。
8.2其他试剂
在一些方法中,该方法包括用可增加HTRA1表达的小分子化合物治疗患者。本文所公开的小分子化合物是指有机化合物,通常具有小于5,000道尔顿、小于1,000道尔顿、小于900道尔顿或小于800道尔顿的分子量。治疗Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法可包括向患者施用小分子化合物,其中所述药剂增加HTRA1的表达。示例性化合物包含曲古抑菌素A(TSA),I类和II类组蛋白脱乙酰酶的抑制剂,其被视为将HTRA1的mRNA表达增加约两倍。Wang et al.,Plos|One 2012,https://doi.org/10.1371/journal.pone.0039446。
在一些情况下,可通过使用定量体外转录测定筛选化合物库来鉴定合适的化合物。体外转录测定可在无细胞系统中进行,该系统包含含有HTRA1和调节元件的质粒。在一些方法中,鉴定可增加HTRA1表达的药剂包括将药剂库与表达HTRA1的细胞接触,测量HTRA1的表达水平或活性,并选择增加HTRA1表达或活性的药剂。可使用本领域众所周知的方法筛选化合物库并且测量HTRA1的转录。例如,转录可以在放射性标记或荧光标记的核苷酸的存在下进行,并且标记的转录物可以在凝胶上沉淀,通过电泳分离,然后定量。另选地,HTRA1mRNA可通过定量RT-PCR或数字PCR来测量。然后选择并测试增加HTRA1转录的试剂。选择可激活转录的药剂(包括化合物或肽)的方法是本领域众所周知的,例如,如美国专利6,174,722中所述,其全部内容以引用方式并入本文。
构建可用于筛选可激活HTRA1转录的药剂的文库的方法也是众所周知的。例如,可制备多种类型的化合物的组合文库,所述化合物可以逐步方式合成。可通过WO 95/12608、WO 93/06121、WO 94/08051、WO95/35503和WO 95/30642中所述的编码合成文库(ESL)方法构建化合物的大型组合文库。肽文库也可通过噬菌体展示方法产生(参见,例如,Devlin,WO91/18980)。细菌、真菌、植物和动物提取物形式的天然化合物文库可从商业来源获得或在现场收集。已知的药剂可进行定向或随机化学修饰,诸如酰化、烷基化、酯化、酰胺化以制备结构类似物。
在一些方法中,测试药剂可以为天然存在的蛋白质或其片段。测试药剂也可以为肽,例如约5至约30个氨基酸的肽,优选约5至约20个氨基酸,并且特别优选约7至约15个氨基酸。肽可以为天然存在的蛋白质、随机肽或“偏向”的随机肽的消化物。测试药剂也可为具有各种长度和序列的核酸。
在一些方法中,可用于治疗Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的药剂可通过将药剂与可操作地连接到报告基因的HTRA1启动子接触来鉴定,并基于其促进报告基因表达的能力来选择试剂。
9.患者群体
9.1患者群体
本发明的组合物和方法特别适于治疗患有10号染色体AMD或具有发展10号染色体AMD风险的受试者。如上所述,已知Chr 10AMD与染色体区域10q26的遗传病变相关,该区域含有ARMS2和HTRA1基因。参见,Hageman et al,2015,“METHODS OF PREDICTING THEDEVELOPMENT OF AMD BASED ON CHROMOSOME 1AND CHROMOSOME 10”,美国专利2015/0211065,其以引用方式并入本文。患有Chr 10AMD的患者可基于疾病表现和/或基于基因型进行鉴定。在一种方法中,治疗的候选者表现出Chr 10AMD临床表型。因此,在一种方法中,接受治疗的受试者表现出Chr 10临床表型。在一种方法中,接受治疗的受试者携带一个或两个10号染色体风险等位基因。在一种方法中,接受治疗的受试者在10号染色体风险等位基因的一个或两个拷贝上携带一个风险等位基因。在一种方法中,受试者具有Chr 10AMD临床表型或基因型并且不具有任何1号染色体风险等位基因。AMD的1号染色体风险等位基因包括rs529825、rs800292、rs3766404、rs1061147、rs203674、rs0161170、rs2274700、rs375046、rs946296和rs94629676。
Chr 10AMD患者主要表现出经典的脉络膜新生血管(CNV),很少有隐匿性CNV和视网膜血管瘤增生(RAP),通常导致严重、快速的视力丧失。Chr 10AMD患者通常比Chr 1AMD患者具有更少的玻璃疣、视网膜内液(囊肿)、快速的地区性萎缩(GA)生长率和视网膜/脉络膜变薄。Chr 10AMD患者也具有减小的视网膜和脉络膜(包括脉络膜毛细血管)血管密度。
在一些方面,通过基因疗法来实现增强HTRA1表达。在一些方法中,增强HTRA1通过施用药剂(例如一种小分子化合物、肽或核酸)来实现的,以上调内源性HTRA1基因的转录。上调HTRA1转录的基因治疗可通过不同的方式进行。在一些方法中,基因疗法上调内源性HTRA1表达(mRNA或蛋白质表达)。在一些方法中,基因疗法引入了外源性HTRA1基因,该基因在RPE细胞中表达。在一些方法中,基因治疗方法用于将细胞基因组中的Ch10风险等位基因转化为相应的非风险等位基因,如下所述。
如下所述,患有Chr 10AMD的个体通常携带一个或多个与疾病发展相关的风险等位基因(“Chr 10风险等位基因”)。参见实施例1和3。发明人已惊奇地发现,相对于健康或低风险对照,Chr 10AMD患者在RPE中特异性地降低了HTRA1 mRNA表达(图2)。此外,发明人已经鉴定了导致HTRA1降低的基因组区域,其对应于与AMD风险增加相关的区域。
对风险等位基因纯合的患者被示为在RPE细胞中具有减少的HTRA1蛋白,这也可能影响RPE和Bruch膜之间界面(“亚RPE空间”)中的HTRA1水平。据信,HTRA1表达的降低,例如亚RPE空间中HTRA1表达的降低,有助于Chr 10AMD的发展。HTRA1是丝氨酸蛋白酶,并可降解在亚RPE空间中高度富集的细胞外基质(ECM)蛋白。不旨在受特定理论或机制的束缚,HTRA1表达的减少可损害ECM蛋白的加工、维持或周转,这导致受损、错误折叠和/或聚集蛋白的积累。继而,这些蛋白质的积累可破坏RPE组织与其基底层和/或Bruch膜的附着,从而导致血-视网膜屏障的丧失并促进AMD的发展。表4示出某些表型与Chr10风险相关的奇数比和p值。在10号染色体风险等位基因纯合的供者的眼部中观察到脉络膜纤维化和基底层状沉积(BLD)。BLD是位于RPE质膜和基底层之间的异常细胞外物质,并且先前已示为与晚期AMD密切相关。此外,导致其错误调节的ECM结构蛋白中的突变导致AMD样疾病,其包括L-ORD(C1qTNF5)、Sorsby眼底营养不良(TIMP3)、Ehlers Danlos综合征VI型(PLOD1)或多氏蜂窝状视网膜营养不良(EFEMP1)。参见,Hayward et al.,Hum.Mol.Genet.12:2657-67(2003);Weber et al.,Nat.Genet 8(4):352-6;and Marmorstein et al.,PNAS.99(20):13067-72。这些证据表明,ECM的改变和基底层状沉积物的形成搞那个像共同的病因,并且HTRA1表达的降低导致受损、错误折叠和聚集的蛋白质的积累以及AMD的发展。
表4
Chr10 AMD中的组织学相关性

1使用调整性别和年龄的多元回归模型计算。
因此,在一些方面,本公开提供了增加RPE细胞中HTRA1表达以防止或减缓Chr10AMD发展的方法。HTRA1疗法可在基于基因谱确定患者具有发展Chr 10AMD的风险之后进行,并且在一些情况下,在AMD的任何临床症状表现之前进行。这种早期干预可避免与AMD相关联的广泛组织损伤。本文所公开的方法需要最少的手术,通常在患者的寿命期间仅进行一次或两次,从而最大限度地减少与常规治疗所需的重复手术相关联的不适和不利影响。
9.1.1治疗对象的选择
在一些方法中,基于用于治疗的临床表型或遗传因素来选择用于治疗的患者。在一些方法中,通过基因分型来对它们进行评估以确定它们的个体遗传学(例如,通过评估如上所公开的Chr 10风险等位基因的存在)和相关的疾病风险。此外,它们可通过临床检查进行评估,包括但不限于以下内容:成像和形态学评估(例如但不限于,彩色眼底摄影、SD-OCT、OCT-A、吲哚青绿血管造影、荧光血管造影术和共焦激光扫描眼底镜(例如海德堡光谱系统),包括近红外反射(NIR)、蓝光自发荧光、绿光自发荧光;以及功能测试(例如但不限于视力、最佳矫正视力(使用ETDRS图表的BCVA)、低亮度BCVA(LLVA,使用带有ETDRS图表的中性密度滤光片)、阅读速度(单目/双目)、微视野(MAIA),包括固视稳定性、暗适应微视野(S-MAIA),暗视和中视微视野灵敏度、视觉诱发电位(VEP)评估和多焦ERG。
其他指标包括形态和功能信息的组合(视力、阅读速度、弱光视力、固视、视网膜电图等)。
此外,可基于多种表型和生物标志物对患者进行评估。施用本文所公开的疗法增加了HTRA1表达,并且当在由眼睛解剖结构和外观的变化或某些生物标志物水平的变化定义的特定表型窗口中施用时,还为患者提供有益效果,其包括但不限于:几乎没有玻璃疣(玻璃疣小且硬)、视网膜内液(囊肿)、快速的GA生长率和视网膜/脉络膜变薄。
一方面,在Chr 10AMD进展的非常早期阶段施用本文所公开的治疗剂可提供优异的治疗有益效果。例如,对出现Chr 10AMD的体征或症状之前的患者(例如,不存在以下症状中的任一者:玻璃疣、视网膜内液(囊肿)、GA快速生长率和视网膜/脉络膜变薄),具体地讲由于具有一个或多个Chr 10风险等位基因而具有高遗传风险的患者进行治疗。因此,本文提供了用于预防Chr 10AMD的发展以及减缓Chr 10AMD的进展、逆转或改善其症状和体征的方法和组合物。在一些方法中,患者不具有AMD的症状(即无症状)。在一些方法中,在第一次施用治疗剂时,患者不表现出Chr 10AMD的临床表型中的任一者。
在一些方法中,患者具有Chr 1和Chr 10、Chr1和其他次要AMD相关基因(C3、CFB、C2等)的组合或所有基因的组合,并且患者用本发明用于治疗Chr 10AMD、预防或减缓其症状和体征的进展的疗法,以及治疗Chr 1AMD、预防或减缓其进展的第二种药剂来治疗。
9.2与HTRA1表达降低和Chr 10AMD相关的遗传因素
9.2.1Chr 10风险等位基因
除了临床表型之外,还可仅基于遗传因素将个体鉴定为处于发展Chr 10AMD的高风险。
因此,本文所公开的方法和组合物可用于用增加受试者RPE细胞中HTRA1表达的药剂治疗具有这些风险等位基因的患者,从而治疗Chr 10AMD、预防其发展、减缓其进展或逆转或改善其症状和体征。
如实施例中所述,与降低的HTRA1 mRNA相关的区域可位于HTRA1的上游调节区,介于rs11200632和rs3750846之间并包括rs11200632和rs3750846(“4kb AMD风险区域”)。示例性Chr 10风险等位基因位于rs11200632(风险等位基因为G)、rs11200633(风险等位基因为T)、rs61871746(风险等位基因为C)、rs61871747(风险等位基因为T)、rs10490924(风险等位基因为T)、rs36212731(风险等位基因为T)、rs36212732(风险等位基因为G)、rs36212733(风险等位基因为C)、rs3750848(风险等位基因为G)、rs3750847(风险等位基因为T)和rs3750846(风险等位基因为C)。表5示出了这些多态位点的风险等位基因(顶部)和非风险等位基因(底部)。表5中示出了具有在4kb风险区域内的rs10490924的处于优异LD(r2=1)或非常高LD的SNP的完整列表。本文所公开的方法和组合物可用于治疗具有这些风险等位基因的一种或多种的患者。
表5.在4kb调控区域内的变体(SNP)和LD


在一些方法中,可从本文所公开的疗法中获益的个体可携带一种或多种风险等位基因,其导致与一种或多种转录激活子的结合减少和HTRA1基因的转录减少。如实施例所示,发明人已发现位于Chr10:122454508-Chr10:122456564处的2kb区域(“2kb风险区域”),该区域在RPE细胞中具有转录活性,并且据信负责结合可激活HTRA1转录的转录激活子。该2kb区域包含活性转录增强子元件的表观遗传标记,其包括H3K4单甲基化和H3K27乙酰化。例如,LHX2与该区域内的序列基序结合,该序列基序具有核苷酸序列TTGCCATAGTATATATAATTAGACAAAT(其包含非风险等位基因T,带下划线,位于rs36212733处)。LHX2与TTGCCGTAGTATATATAACTAGACAAAT(包含风险等位基因C,带下划线,位于rs36212733处)的结合较差。参见图9(图10和11)。因此,在一些方法中,本文所公开的方法包括向患者施用药剂,其在该2.0kb区域中的基因组DNA与HTRA1的转录激活子的结合亲和力降低,并且药剂的施用增加RPE细胞中的HTRA1表达。在一些方法中,患者示出与LHX2的结合减少或完全丧失。在一些方法中,患者可具有在rs36212733处的风险等位基因C(即TTGCCATAGTATATATAACTAGACAAAT),这导致其失去与LHX2的结合。在一些方法中,治疗方法包括向患者施用药剂,其中该药剂通过促进转录激活子(例如,LHX2)与HTRA1的转录调控区的结合来增加患者RPE细胞中的HTRA1表达。
9.2.2IncSCTM1表达与HTRA1表达呈负相关
在一些方法中,与对照相比,可从本文所公开的治疗中获益的个体示出非编码RNA、lncSCTM1(也称为LOC105378525)或其同种型的等位基因特异性表达增加。IncSCTM1从与HTRA1共享相同LD块的DNA序列转录而来。如图7和示例中所示,lncSCTM1与HTRA1共享不同的启动子,并且从HTRA1启动子以反义方向转录。lncSCTM1的等位基因特异性表达与HTRA1的等位基因特异性表达呈负相关。在杂合子患者中,lncSCTM1的风险等位基因(例如rs11200638)的mRNA水平是比lncSCTM1的非风险等位基因的mRNA水平更高的水平。相比之下,HTRA1的风险等位基因的mRNA水平低于非风险等位基因的mRNA水平(图8)。
在一些方法中,可从本文所公开的疗法获益的患者携带一种或多种剪接形式的lncSCTM1,其与HTRA1的表达降低相关。如实施例所示,IncSCTM1作为不同的剪接变体存在(图7)并在不同的眼部组织中表达(图14)。
在一些方法中,本文所公开的方法和组合物可用于治疗具有Chr 10AMD的一种或多种临床表型的个体中的Chr 10AMD,例如几乎没有玻璃疣、视网膜内液(囊肿),表现出快速的GA生长速率,视网膜/脉络膜变薄。在一些方法中,患者具有如上所述的一个或多个Chr10风险等位基因。在一些方法中,患者具有Chr 10AMD的一种或多种临床表型和一种或多种Chr 10风险等位基因两者。
在一些情况下,除了具有Chr 10AMD临床表型和/或具有Chr10风险等位基因外,患者还具有Chr 1风险等位基因和/或Chr 1驱动的AMD临床表型。Chr 1风险等位基因/单倍型。参见美国专利7,867,727,其通过引用方式并入本文。在一些情况下,患者不具有AMD的Chr 1风险等位基因/单倍型。
10.给药方法和剂量
如上所述,本发明的各方面包括向需要治疗的受试者施用药剂以增加HTRA1表达或编辑基因组区域以将风险等位基因转化为非风险等位基因的方法。因此,本发明的各方面包括在受试者中药剂的递送导致对受试者健康的一个或多个方面产生有益影响的条件下,使受试者与一种或多种治疗剂接触,例如如上所述的病毒载体、化合物、肽或其组合。本发明不限于特定的给药部位或方法。例如,出于说明而非限制性的目的,可通过全身给药(例如静脉注射或输注)、局部注射或输注(例如,视网膜下、脉络膜上、玻璃体内、经巩膜或其他眼部注射或输注),通过使用渗透压泵,通过电穿孔,通过应用(例如,滴眼液)和通过其他方式。预期本发明的转基因可被引入并表达于多种细胞类型中,包括神经视网膜细胞类型,例如视杆细胞、视锥细胞、RPE和神经节细胞、睫状上皮细胞、巩膜细胞、脉络膜细胞和其他眼细胞。
本文所公开的治疗剂可悬浮在生理相容性载体中以用于对人给药。给药可通过眼部或非眼部(例如,玻璃体内、血管内、眼外给药)途径来进行。考虑到递送途径,本领域技术人员可容易地选择合适的载体。例如,一种合适的载体包括盐水,其可与多种缓冲溶液(例如磷酸盐缓冲盐水)一起配制。
10.1眼部给药
10.1.1视网膜下注射和其他注射
在RPE-脉络膜界面水平附近引入治疗剂可在Chr 10AMD的早期阶段更好地控制HTRA1调节,并防止与晚期地区性萎缩和脉络膜新生血管形成相关的失明。在一种方法中,通过在视网膜下产生疱疹或水疱,将病毒载体直接输送到受疾病影响的视网膜区域,经由视网膜下注射施用药剂。一种或多种疱疹可在眼部的一个象限或多个象限中产生以确保治疗剂的有效分布。参见,Xue et al.,“Technique of retinal gene therapy:delivery ofviral vector into the subretinal space”Eye 31:1308-1316,2017。还参见Moore etal.2017,Ochakovski et al.2017, et al.2015,如上所述。
et al.2015,如上所述。
在另一种方法中,药剂通过脉络膜上注射给药,从而到达RPE的基底表面。参见Ding et al.,“AAV8-vectored suprachoroidal gene transfer produces widespreadocular transgene expression”,J Clin Invest 129(11):4901-4911,2019。另外参见Emami-and Yiu,Medical and Sugical Applications for the Suprachoroidal Space,Int Ophthalmol Clin 59(1):195-207,2019。在另一种方法中,可将药剂注射到玻璃体内。这种方法可能对于使药剂到患有地区性萎缩或CNV的Chr 10AMD患者的RPE中而言特别有用。该技术是本领域技术人员众所周知的。参见,Kansar et al.,“SuprachoroidalDelivery of Viral and Nonviral Gene Therapy for Retinal Disease”,J OcularPharmacol Ther DOI:10.1089/jop.2019.0126.2020。
剂量
值得注意的是,剂量值可能会随着产品的性质和病症的严重程度而变化。还应当理解,对于任何特定受试者,可根据个体需要和施用或监督组合物施用的人的专业判断随时间推移来调整具体的剂量方案,并且本文所述的剂量范围仅是示例性的且不旨在限制要求保护的组合物的范围或实践。
施用的药剂的量将是“有效量”或“治疗有效量”,即在必要的剂量和时间段内有效实现期望结果的量。期望的结果将包括靶细胞(例如,RPE细胞)中HTRA1表达或活性的改善或与HTRA1表达降低相关的症状的可检测的改善,其包括但不限于AMD症状或体征的改善,优选统计学上的显著改善。另选地,如果预防性地使用药物组合物,则期望的结果将包括对Chr 10AMD的一种或多种症状的明显预防,其包括但不限于AMD的症状或体征,优选地统计学上的显著预防。此类组合物的治疗有效量可根据诸如个体的疾病状态、年龄、性别和体重或病毒载体在个体中引发期望反应的能力等因素而变化。可调整剂量方案以提供最佳反应。治疗有效量也是这样的量,其中药剂(例如病毒载体)的任何毒性或有害作用被治疗有益作用抵消。组合物中病毒载体的量可根据诸如个体的疾病状态、年龄、性别和体重等因素而变化。
可调整剂量方案以提供最佳治疗反应。例如,可施用单次推注,可随着时间推移施用若干分开的剂量,或者可根据治疗情况的紧急情况按比例减少或增加剂量。在治疗剂是AAV颗粒的情况下,优选的人剂量可以为每次注射108至1012个AAV基因组,体积为100-300μl/视网膜下疱。每个眼睛可能会产生多于一个疱。任何给定的个体其寿命中都可能需要多次治疗。
11.治疗结果
在合适的患者中进行HTRA1基因治疗,包括治疗处于发展AMD风险或处于疾病早期阶段的个体,可稳定、改善或逆转患者的AMD症状或体征。例如但不限于,提供外源性HTRA1蛋白,向Chr 10风险等位基因杂合或纯合的患者引入转录激活子可稳定和/或减缓甚至逆转疾病的进展,如由各种眼部生物标志物所证实的。在一种方法中,用HTRA1相关疗法治疗的患者的主要期望治疗结果是与Chr 10AMD相关的一种或多种症状的可检测改善,即具有玻璃疣、视网膜内液(囊肿)、快速的GA生长率和视网膜/脉络膜变薄、视网膜和脉络膜血管密度降低、黄斑区脉络膜毛细血管影(OCT-A上无血流)、黄斑区脉络膜纤维化、外黄斑区脉络膜纤维化、黄斑区内Bruch基底层沉积(BLD)、黄斑区内的BLD、黄斑区内的Bruch膜增厚、外黄斑区的BLD。
预期用HTRA1相关疗法治疗的患者的预期治疗结果,其也可以是一项或多项功能性测量的可检测改善,其包括但不限于:视敏度(早期治疗糖尿病视网膜病变研究,或ETDRS);最佳矫正视力(或BCVA);微视野(黄斑完整性评估,或MAIA);暗适应;阅读速度;视觉诱发电位(VEP);和多焦视网膜电图(mfERG)等。指示稳定、减缓或逆转AMD进展的其他生物标志物包括但不限于:BCVA变化;GA变化区域(平方根变换或其他方式);注视;阅读速度;GA新领域%;感光器高度;个体德鲁斯特征。
12.药物组合物
本发明的另一方面涉及本发明载体的药物组合物。在一个实施方案中,组合物包含有效量的药剂和药学上可接受的载体。在一些方法中,无菌可注射溶液可通过掺入所需量的载体(例如病毒载体),任选地与适用于注射到人患者中的稀释剂或赋形剂来制备。提供了单位剂型,诸如单次使用、预填充注射器或其他注射装置,具有例如足够的AAV颗粒或化合物以用于对患者进行单次给药。本发明的任何药物制剂均可包装或附有关于该制剂及其治疗AMD的用途的信息。
13.定义和惯例
在更详细地描述本发明之前,应当理解,本发明不限于所述特定方法,因此当然可以变化。还应当理解,本文所用的术语仅用于描述特定的方法,而不旨在进行限制,因为本发明的范围将仅由所附权利要求来限制。
如本文所用,术语“转基因”与“外源基因”可互换使用,是指使用基因治疗载体引入细胞中以导致表达的重组多核苷酸构建体。
如本文所用,术语“天然启动子”是指天然和/或最初存在于细胞中的启动子,并且其通常被指定用于特定基因的表达。例如,SEQ ID NO:13是天然HTRA1启动子。基因的非天然启动子是不与基因天然相关的启动子。例如,VMD2启动子(SEQ ID NO:6)不是天然HTRA1启动子。
如本文所用,术语“天然转录激活子”是指天然和/或最初存在于细胞中的转录激活子,其通常被指定用于调节特定基因的转录。例如,LHX2是HTRA1启动子的天然转录激活子,然而VP16不是HTRA1的天然转录激活子。
如本文所用,“基因治疗载体”是指用于将转基因引入细胞中的病毒衍生的序列元件。
如本文所用,“病毒载体”是指包括衣壳蛋白的基因治疗载体,其用于将转基因递送至细胞。
如本文所用,术语“启动子”是指能够控制(例如,增加)编码序列或功能性RNA的表达的DNA序列。启动子可包括最小启动子(由TATA盒和用于指定转录起始位点的其他序列组成的短DNA序列)。增强子序列(例如,上游增强子序列)是可以与启动子相互作用以控制(例如增加)编码序列或功能性RNA的表达的调节元件。如本文所用,提及“启动子”可包括增强子序列。
当启动子和其他调控序列影响转基因或转基因产物(例如,mRNA或蛋白质)的表达或稳定性时,它们与转基因“可操作地连接”。
如本文所用,在基因治疗的上下文中,术语“引入”或“引入的”是指在多核苷酸进入细胞并在细胞中表达以产生蛋白质的条件下,施用组合物,所述组合物包含例如编码HTRA1多肽的多核苷酸(DNA)、可增加HTRA1表达的转录激活子或DNA核酸内切酶。可使用病毒(例如,AAV2)载体,使用非病毒载体系统或通过其他方法将多核苷酸作为裸DNA引入。
术语“对应于”和语法等价物在本文中使用以指相似或同源蛋白质或核苷酸序列中的位置,无论确切位置与测量相似性或同源性的分子是相同还是不同。例如,假定第一蛋白质的长度为100个残基,且第二个蛋白质除了在氨基末端缺失5个氨基酸之外与第一个蛋白质相同,则第一个蛋白质的第12位将“对应”于第二种蛋白质的第7位。
“腺相关病毒2(AAV2)”和“重组腺相关病毒2(rAAV2)”等效使用。示例性AAV2载体源自腺相关病毒2基因组,并在科学文献中进行了广泛的描述。参见,例如,Srivastava,etal,1983,J.Virol.45:555-564,其以引用方式和下文引用的其他参考文献并入本文。
如本文所用,“慢病毒”是指可用于将转基因转导到细胞中的基因治疗载体(慢病毒载体)。参见,例如,Keeker et al.,2017,Clin Transl Sci.10:242 248,其以引用方式和下文引用的其他参考文献并入本文。
在两个或更多个核酸或多肽序列的上下文中,术语“相同”或百分比“同一性”是指两个或更多个序列或子序列是相同(“同一”)的,或具有特定百分比的相同的氨基酸残基或核苷酸(即,当通过手动比对和视觉检查或使用具有下文所述的默认参数的BLAST或BLAST2.0序列比较算法所测量的,在比较窗口或指定区域上进行比较和比对以获得最大对应时,在指定区域的整个序列范围内的至少约70%同一性、至少约75%同一性、至少80%同一性、至少约90%同一性,优选至少约91%、92%、93%、94%、95%、96%、97%、98%、99%或更高同一性(参见,例如,NCBI网站ncbi.nlm.nih.gov/BLAST/等))。此类序列然后被认为是“基本上相同的”
术语“受试者”或“患者”是指接受预防性或治疗性治疗的人或动物(特别是哺乳动物)和其他生物体。例如,受试者可以为非人类灵长类动物。
如下所述,优选算法可以考虑间隙等。优选地,同一性存在于长度至少约25个氨基酸或核苷酸的区域上,或更优选地存在于长度为50-100个或更多个氨基酸或核苷酸的区域上。对于序列比较,通常一个序列充当参考序列,测试序列与所述参考序列进行比较。在一些方法中,确定与选自SEQ ID NO:1(HTRA1的核苷酸序列)或SEQ ID NO:2(HTRA1的氨基酸序列)的参考序列的全长相关的百分比同一性。当使用序列比较算法时,将测试和参考序列输入计算机中,必要时指定子序列坐标,并指定序列算法程序参数。优选地,可使用默认程序参数,或者可指定替代参数。然后,序列比较算法基于程序参数计算测试序列相对于参考序列的序列同一性百分比。如本文所用,“比较窗”包括指选自由20至600,通常约50至约200,更通常约100至约150个组成的组中的多个连续位置中任一个的片段。,其中在两个序列最佳比对后,可将序列与具有相同数量了连续位置的参考序列进行比较。用于比较的序列比对方法是本领域众所周知的。可通过以下方法进行用于比较的序列的最佳比对:例如,Smith&Waterman(Adv.Appl.Math.2:482(1981))的局部同源算法,Needleman&Wunsch(J.Mol.Biol.48:443(1970))的同源性比对算法,Pearson&Lipman(Proc.Nat'l.Acad.Sci.USA 85:2444(1988))的搜索相似性方法,这些算法的计算机化实施(威斯康星遗传学软件包中(Genetics Computer Group,575Science Dr.,Madison,Wis.)中的GAP、BESTFIT、FASTA和TFASTA),或者手动比对和目视检查(参见,例如,Current Protocols inMolecular Biology(Ausubel et al.,eds.1995,增刊))。适用于确定百分比序列同一性和序列相似性的算法是BLAST和BLAST 2.0算法,它们分别在Altschul et al.,Nuc.AcidsRes.25:3389-3402(1977)和Altschul et al.,J.Mol.Biol.215:403-410(1990)中有所描述。BLAST和BLAST 2.0与本文所述的参数一起使用以确定本发明的核酸和蛋白质的百分比序列同一性。用于执行BLAST分析的软件可通过国家生物技术信息中心(www.ncbi.nlm.nih.gov/)公开获得。该算法首先通过鉴定查询序列中长度较短的词来鉴定高评分序列对(HSP),当与数据库序列中相同长度的词比对时,这些词匹配或满足某个正值阈值分数T。T被称为邻域词得分阈值(Altschul等人,同上)。这些初始邻域词命中充当种子,用于启动搜索以找到包含它们的更长的HSP。单词命中沿每个序列在两个方向上扩展,只要累积比对分数能够增加。对于核苷酸序列,使用参数M(一对匹配残基的奖励分数;总是>0)和N(错配残基的惩罚分数;总是<0)来计算累积分数。对于氨基酸序列,使用评分矩阵来计算累积评分。在以下情况下,单词在每个方向上的扩展会停止:累积比对分数从其最大实现值下降了数量X;由于一个或多个负评分残基比对的累积,累积分数变为零或更低;或到达任一序列的末端。BLAST算法参数W、T和X决定比对的灵敏度和速度。BLASTN程序(用于核苷酸序列)使用默认字长(W)为11,期望值(E)为10,M=5,N=-4以及两条链的比较结果。对于氨基酸序列,BLASTP程序默认使用字长为3、期望值(E)为10和BLOSUM62评分矩阵(参见Henikoff&Henikoff,Proc.Natl.Acad.Sci.USA89:10915(1989))比对(B)为50,期望值(E)为10,M=5,N=-4,以及两条链的比较结果。
“玻璃膜疣”是位于RPE基底层和Bruch膜之间的局灶性细胞外沉积物,包括脂质、流体、各种蛋白质,包括补体途径相关蛋白质。玻璃膜疣在检眼镜下作为白色/黄色点可见,并且可使用多种本领域已知的方法进行检测,包括Wu et al.,2015,“Fundusautofluorescence characteristics of nascent geographic atrophy in age-relatedmacular degeneration”Invest Ophthalmol Vis Sci.56:1546-52和该参考文献的参考文献1-8。如本文所用,术语“小玻璃疣”和“小硬玻璃疣”是指直径小于约63μm的不同玻璃疣。术语“大玻璃膜疣”、“软玻璃膜疣”和“大软玻璃膜疣”是指直径大于约125μm的玻璃膜疣,它们通常是成簇的。直径在63μm和125μm之间的玻璃膜疣可以称为“中间玻璃膜疣”。RPE的局部脱离,通常称为色素上皮脱离(PED),通常称为玻璃疣。
如本文所用,“单倍型”是指存在于染色体上不同位点处的DNA序列或DNA序列的组合,它们一起传递;单倍型可以为一个基因座、几个基因座或整个染色体,这取决于在一组给定基因座之间发生的重组事件的数量。
术语“多态性”是指一个或多个基因决定的替代序列或等位基因在群体中出现。“多态位点”是发生序列分歧的位点。多态位点至少有一个等位基因。二等位基因多态性有两个等位基因。三等位基因多态性具有三个等位基因。二倍体生物体对于等位基因形式可以是纯合的或杂合的。多态位点可小到一个碱基对。多态位点的示例包括:限制性片段长度多态性(RFLP)、可变数量串联重复序列(VNTR)、高变区、小卫星、二核苷酸重复序列、三核苷酸重复序列、四核苷酸重复序列和简单序列重复。如本文所用,提及“多态性”可包括一组多态性(即,单倍型)。
“单核苷酸多态性(SNP)”可以发生在由单个核苷酸占据的多态位点处,这是等位基因序列之间的变异位点。该位点可在等位基因的高度保守序列之前和之后。由于在多态性位点处用一种核苷酸置换另一种核苷酸,可能产生SNP。一种嘌呤被另一种嘌呤置换或一种嘧啶被另一种嘧啶置换被称为转变。嘌呤被嘧啶取代或反之亦然被称为颠换。同义SNP是指在不改变编码多肽的氨基酸序列的编码区中用一个核苷酸置换另一个核苷酸。非同义SNP是指在编码区用一个核苷酸置换另一个核苷酸,从而改变所编码多肽的氨基酸序列。SNP也可源自相对于参考等位基因的一个或多个核苷酸的缺失或插入。
如本文所用,“连锁不平衡”或“LD”是两个或多个基因座上等位基因的非随机关联,其不一定在同一染色体上。它与基因连锁不同,上所述基因连锁描述了染色体上两个或多个基因座的关联,它们之间的重组有限。基因连锁不平衡描述了一种情况,在这种情况下,等位基因或遗传标记的某些组合在群体中的出现频率高于或低于基于等位基因随机形成单倍型的频率所预期的不同位点处的多态性之间的非随机关联通过连锁不平衡(LD)的程度来测量。连锁不平衡的水平会受到许多因素的影响,其包括遗传连锁、重组率、突变率、随机漂移、非随机交配和种群结构。因此,“连锁不平衡”或“等位基因关联”是指特定等位基因或遗传标记与另一个特定等位基因或遗传标记的非随机关联,其频率高于种群中任何特定等位基因频率的偶然所预期的。与信息性标记(诸如本文描述的SNP、单倍型或双倍型中的一种)的连锁不平衡的标记可用于检测对Chr 10驱动的AMD的易感性。
“ARMS2”是指AMD易感性2基因。ARMS2基因由两个外显子组成并且理论上编码107氨基酸蛋白,且与已知的蛋白质基序没有同源性,也没有已知的功能。ARMS2蛋白在细胞内的表达和定位仍然难以捉摸,并且使用缺乏特征的抗体导致了相互矛盾的报告(Fritsche等人,2008;Kanda等人,2007;Kortvely等人,2010;Wang等人,2012)。此外,RNA-Seq分析表明,除睾丸(TPM为大约6.9)和胎盘(未示出qRT-PCR数据)外,大多数组织中ARMS2 mRNA的表达异常低(TPM<1.0)(Lonsdale等人,2013;The GTEx Consortium,2015)。
“HTRA1”是指HtrA丝氨酸肽酶1,其mRNA和蛋白质分别由基因库登录号NM_002775和NP_002766表示。HTRA1在几乎所有检查的细胞和组织中普遍表达,并且在视网膜的光感受器和水平细胞、视网膜色素上皮(RPE)和脉络膜中的各种细胞类型中大量表达(图1)。与视网膜和RPE相比,HTRA1富集在来源于Bruch膜和脉络膜的提取物中(图6)。HTRA1用作分泌型丝氨酸蛋白酶,并且主要以大约150kDa的三聚体形式存在于溶液中。HTRA1由变构机制激活(Cabrera等人,2017)并裂解多种细胞外基质蛋白、蛋白聚糖和许多生长因子,诸如TGFb、FGF和IGFBP。在HTRA1敲除小鼠模型中,HTRA1的缺失导致许多ECM蛋白增加,包括脑血管蛋白质组中的TIMP3、凝聚素、弹性蛋白、玻连蛋白和纤维蛋白3(Zellner等人,2018)。由于大脑中的年龄相关性血管系统缺陷,损害HtrA1蛋白酶活性或降低mRNA表达的功能缺失突变也可导致CARASIL(伴有皮质下梗塞和白质脑病的常染色体显性隐性脑动脉病)(Hara等人,2009年;Fukutake,2011年)。
如本文所用,术语“治疗”或其任何语法变体(例如,治疗(treat)、治疗中(treating)、治疗(treatment)等)包括但不限于减轻疾病或病症的症状;和/或减少、阻止、抑制、减轻、改善或影响疾病或病症的进展、严重性和/或范围。
如本文所用,术语“转录调控区”是指HTRA1启动子和HTRA1增强子。
如本文所用,术语“增强子”是指HTRA1 2kb区域。
如本文所用,序列上下文中的术语“对应于”是指互补物、等同于DNA序列但U置换T的RNA序列,以及从上下文显而易见的类似物。
如本文所用,“密码子优化”具有其在本领域中的通常含义。与使用非优化序列产生的转录物相比,密码子优化可用于提高翻译速度或产生具有期望特性的重组RNA转录物,诸如更长的半衰期和更高的表达效率。在一些实施方案中,本公开内容提供已被工程化以最大化表达效率的HTRA1编码序列。用于密码子优化的方法很容易获得,例如优化器,可在http://genomes.urv.es/OPTIMIZER处免费访问,以及得自DNA 2.0(Newark,California)的 表达优化技术。在特定实施方案中,使用来自GenScript(Piscataway,NewJersey)的OptimumGeneTM算法对编码序列进行密码子优化以在人中表达。
表达优化技术。在特定实施方案中,使用来自GenScript(Piscataway,NewJersey)的OptimumGeneTM算法对编码序列进行密码子优化以在人中表达。
如本文所用,“缺乏核酸酶活性”的Cas蛋白相对于野生型等同物具有活性的至少50%降低,有时至少80%降低,有时至少95%降低,有时至少99%降低。
如本文所用,术语“水平细胞”具有本领域中的正常含义,参见,例如,Pochéetal.,“Retinal horizontal cells:challenging paradigms of neural development andcancer biology”,Development,2009,136,Pages 2141-2151。“水平细胞”是横向互连的神经元,其细胞体位于脊椎动物眼睛的视网膜的内核层中。它们整合并调节来自多个感光细胞的输入。
如本文所用,术语“感光细胞”是存在于神经视网膜的最外层中的特化神经上皮细胞。它们能够进行视觉光转导,并具有两种主要类型,介导暗视觉的视杆细胞和介导明视觉的视锥细胞。
如本文所用,“可编程”核酸内切酶是可通过选择相关分子(例如,用于Cas蛋白的gRNA)、融合蛋白序列或其他方式来特异性靶向特定DNA序列的核酸酶。
本文使用以下约定。DNA“目标序列”[A]与第一条DNA链上的PAM[P]相邻。目标序列[C]的互补序列位于互补DNA链上。gRNA的指导序列[G]与靶的互补序列杂交并互补,并且具有靶的序列[A],不同的是DNA中的胸苷被RNA中的尿嘧啶取代。gRNA的指导序列可通过[C]的转录产生。
除非另有说明,核苷酸序列从5'到3'呈现。
除非另有定义,本文所用的所有技术和科学术语与本发明所属领域的普通技术人员通常理解的含义相同。虽然与本文所述的那些相似或等效的任何方法和材料也可用于本发明的实践或测试,但现在描述代表性的说明性方法和材料。
14.实施例
14.1HTRA1在具有10号染色体风险等位基因的人供体RPE中的表达降低
该实施例示出,与对照相比,来自具有AMD风险等位基因的个体的人供体眼部具有较低的mRNA HTRA1表达。
使用两组独立的人供体眼部组织(分别包括80个和200个供体样品)进行基于微阵列的基因表达分析。结果示出,相对于非风险等位基因纯合的非风险供体,对于在rs10490924处的风险等位基因纯合或杂合的人供体的RPE脉络膜组织中HTRA1 mRNA水平降低(图2和图30)。通过qRT-PCR分析证实了RPE-脉络膜组织中HTRA1 mRNA水平降低的结果(图3A和3B)。使用来自杂合供体RPE组织样本的mRNA进行的等位基因特异性表达分析也示出,相对于野生型等位基因mRNA,风险等位基因mRNA显著且可重复降低。使用数字PCR和Taqman SNP测定确定具有非风险等位基因相对于风险等位基因的mRNA拷贝数。在来自相同供体样本的脉络膜或视网膜中没有检测到等位基因特异性mRNA变化(图4)。从等位基因特异性HTRA1表达测定中获得的这些结果强烈支持了AMD相关风险等位基因导致HTRA1 mRNA表达降低。
结果还示出,HTRA1 mRNA的减少是组织特异性的—其仅在RPE中检测到,但在视网膜中未检测到。图2、图3A、图3B和图30示出,相对于具有两个、野生型、非风险等位基因的非风险供体,在rs10490924处具有一个或两个风险等位基因的患者的RPE-脉络膜组织中HTRA1 mRNA水平降低,但HTRA1表达在视网膜中保持不变。
14.2HTRA1表达在来自人非AMD样本的眼部组织中的Bruch膜和脉络膜中富集
该实施例示出,HTRA1 mRNA以极化方式分泌,并在来自非AMD样本的人眼部组织中的Bruch膜和脉络膜中富集。
在极化的hTERT-RPE1细胞(人RPE细胞系,参见Bodnar et al.,1998,Science279:349-52)和人胎儿RPE细胞中,HTRA1蛋白以极化方式分泌,其中70%的HtrA1顶端分泌且30%基础分泌(图5A和5B)。HTRA1的基础分泌可能是老年人中的亚RPE HTRA1的唯一来源,因为由于Bruch膜和RPE基底层的选择性渗透,预期HTRA1三聚体不从脉络膜毛细血管进入亚RPE空间(Moore,DJ等人,2001)。RPE和Bruch膜之间的界面高度富集细胞外基质(ECM)蛋白,并且很可能的是具有被证明的降解各种ECM蛋白的能力的HTRA1(分泌型丝氨酸蛋白酶)在该空间中具有年龄相关性基本功能。相对于视网膜、RPE和脉络膜,该数据表明HTRA1蛋白富集在来自Bruch膜的提取物中,所述提取物具有或不具有来自不患有AMD的患者的样本(图6)。不旨在受特定理论或机制的束缚,这些结果与模型一致,其中亚RPE空间中HTRA1蛋白减少的纯合型风险患者改变了ECM蛋白的加工、维持或周转,从而导致损坏、错误折叠和/或聚集的蛋白质的积累。这可能破坏RPE组织与其基底层和/或Bruch膜的附着,从而导致血-视网膜屏障的丧失。为了支持这一点,已证明脉络膜纤维化和基底层状沉积(BLD)(位于RPE质膜和基底层之间的异常细胞外物质,并示出与晚期AMD密切相关)在对10号染色体风险纯合的供体中发生(上表1)。导致其错误调节的几种ECM结构蛋白中的突变导致AMD样疾病,其包括L-ORD(C1qTNF5)、Sorsby眼底营养不良(TIMP3)、Ehlers Danlos综合征VI型(PLOD1)或多氏蜂窝状视网膜营养不良(EFEMP1)。参见Hayward et al.,Hum.Mol.Genet.12:2657-67(2003);Weber et al.,Nat.Genet 8(4):352-6;andMarmorstein et al.,PNAS.99(20):13067-72。这表明ECM的改变和BLD的形成可具有共同的病因。
14.3人眼组织中的HtrA1蛋白
AAV-HTRA1疗法的目标是向眼部递送治疗量的HtrA1以恢复功能。为此,有必要了解人类和AGM眼部组织中的HtrA1浓度。通过ELISA测定从各种眼部组织的提取物中测量HtrA1的浓度。表6总结了人眼组织中的浓度。
表6:人血清和眼组织中HtrA1蛋白的浓度


黄斑外视网膜和RPE-脉络膜中HtrA1蛋白和mRNA水平随Chr10基因座处具有或不具有风险的供体的年龄变化的比较结果展示出,HtrA1水平在视网膜中随着年龄相对不变,而与Chr10风险状态无关(图12)),但在不具有Chr10风险的供体的RPE脉络膜中随年龄显著增加(图13A)。相比之下,具有Chr10风险的供体在非风险供体的RPE脉络膜中HtrA1的年龄依赖性增加方面表现出显著减少。这与非风险供体中的HTRA1mRNA的类似年龄依赖性增加一致,但与具有风险的供体不一致(图13B)。不旨在受特定理论或机制的束缚,这些结果与模型一致,其中对HtrA1的需求随年龄增加,这通过HTRA1 mRNA的转录上调来满足,从而导致更多HtrA1蛋白的翻译,但在对于风险纯合的供体中,存在mRNA或蛋白质水平上调HTRA1的失败。
14.4包含导致HTRA1 mRNA表达降低的调节元件的4Kb区域也与AMD风险相关
该示例示出,导致HTRA1降低的基因组区域匹配与发展为AMD的风险相关的区域。
为了缩小10号染色体上负责AMD的区域(风险区域),HTRA1等位基因特异性表达测定使用来源于供体的mRNA,在AMD相关的ARMS2/HTRA1 LD块内发生罕见的重组事件。供体在HTRA1外显子1中的rs1049331SNP处是杂合的,但对于LD块内的上游SNP的一部分中的风险或非风险等位基因是纯合的。通过在等位基因特异性表达测定中使用这些供体并经由DNA测序绘制每个供体的重组位点,HTRA1启动子SNP和ARMS2插入-缺失(indel)区域被排除在HTRA1 mRNA表达降低的因果驱动因素之外。相反,与减少的HTRA1 mRNA相关的区域被映射到上游调控区,所述上游调控区介于rs11200632和rs3750846SNP(包括rs10490924(ARMS2A69S))之间且包括rs11200632和rs3750846SNP(表8)。由Grassmann等人(Genetics2017)在病例/对照研究中对重组单倍型的分析发现,相同的遗传区域与AMD疾病风险升高有关。与HTRA1等位基因特异性表达相关的区域匹配与AMD风险相关的同一区域的发现强烈暗示HTRA1表达的风险相关降低导致AMD发病率增加。该区域被指定为4kb AMD风险区域。
下面的表7示出ARMS2区域的九个单倍型,其各自占群体的2%以上。单倍型H1、H2、H3和H6都示出增加的Chr 10AMD风险,并且全部包含位于SNP rs10490924处的T等位基因。这指示rs10490924(A69S)风险等位基因的存在与Chr 10AMD的发展相关。
表7:ARMS2区域内的单倍型

表7(续)


表7(续)

表8
对人类供体的RPE脉络膜中的HTRA1等位基因特异性表达的分析,其中基因重组在与AMD相关联的10q26 GWAS LD块内


14.5HTRA1和IncSCTM1 mRNA表达之间的反比关系
这个例子展示出HTRA1和incSCTM1等位基因特异性mRNA表达之间的反比关系。鉴定出长非编码RNA(LOC105378525)并将其映射到与4kb AMD风险区域重叠的区域。我们的分析示出,lncRNA有四种预测的变体,包括XR_946382、XR_946383、XR_946384和XR_946385(图7)。表9中示出在hTERT-RPE1细胞、视网膜和RPE组织中检测到的lncSCTM1的各种同种型。该lncRNA从HTRA1启动子开始以反义方向转录,其显示出HTRA1和LOC105378525可能共享重叠的发散启动子。我们将该lncRNA命名为lncSCTM1。来自视网膜和RPE的RNA的RACE(cDNA末端的快速扩增)PCR分析指示,映射在XR_946382变体1中的外显子3比所报道的更远延伸大约1kb。这由来自GTEx联盟的RNA-seq数据支持,其中读取被映射到各种组织中的这个扩展区域。RACE PCR还指示,映射在变体4(XR_946385)中的替代外显子3从hg38 Chr 10:122,455,021(alt ex3a)或Chr 10:122,454,857(Alt ex3b)开始,并在反义方向延伸至Chr 10:122,454,457。序列分析指示alt ex3a变体包含rs10490924SNP,其风险形式与AMD风险升高相关,然而alt ex3b则不相关。此外,包含外显子3而不是替代外显子3的变体在4kb AMD风险区域中不包含rs10490924或任何其他SNP。lncSCTM1替代外显子3几乎与ARMS2外显子1完全重叠。
表9
IncSCTM1同种型的表达


由于lncRNA有时会调节附近基因的表达,因此使用HTRA1(rs1049331,外显子1)和lncSCTM1(rs11200638,外显子1)的等位基因特异性表达测定来检查HTRA1 mRNA和lncSCTM1 ncRNA水平之间的关系。来自杂合供体的mRNA用于测定中,并且与预期的50:50比率的任何偏差都表明等位基因特异性表达。基因特异性引物用于cDNA合成,以确保转录物来自预期的链。据观察,HTRA1的等位基因特异性表达与lncSCTM1的等位基因特异性表达呈负相关。结果还示出,来源于lncSCTM1风险等位基因的mRNA表达水平高于来源于非风险等位基因的mRNA表达水平,然而来源于HTRA1风险等位基因的mRNA相对于非风险等位基因的mRNA表达水平降低(图8)。图8中分析的风险等位基因位于rs11200638,其位于与AMD相关的LD块中,包括rs10490924。rs11200638也位于lncSCTM1的外显子1内,并存在于lncSCTM1的所有同种型中。视网膜和RPE的RNA-seq分析指示HTRA1 mRNA的表达水平比lncSCTM1 mRNA高>400倍(数据未示出)。如果lncSCTM1调节HTRA1表达,则似乎不太可能的是它能够反式调节HTRA1,相反,最有可能的是IncSCTM1可通过对HTRA1转录的顺式作用调节HTRA1的表达。
14.6包含导致HTRA1 mRNA表达减少的调节元件的2kb AMD风险区
表观遗传标记
该例子描述了新鉴定的2kb区域,该区域负责调节HTRA1转录以及与之相关的转录激活子。对源自人类胎儿RPE组织或诱导多能干细胞(iPSC-RPE)的公开的表观遗传数据(ATAC-Seq和DNase-Seq)的检查指示,大约2.0kb的开放染色质区域与4kb AMD风险区重叠。此外,这个相同的2kb区域包含活跃转录的表观遗传标记,包括H3K27乙酰化。该2kb区域与lncSCTM1的ARMS2外显子1-内含子1和替代外显子3(图10)以及4kb AMD风险区中的其他SNP(包括rs36212731、rs36212732、rs36212733和rs3750848)重叠。
视网膜和RPE提取物的ChIP-Seq分析使用靶向与增强子元件相关的组蛋白标记的抗体进行,包括单甲基化组蛋白H3赖氨酸4(H3K4me1)和乙酰化组蛋白H3赖氨酸27(H3K27Ac)。在RPE提取物中,两个标记都存在于与开放染色质区域重叠的峰中,然而视网膜中仅存在H3K4me1标记。这些结果支持如下假设:ARMS2内含子内的这个大约2kb区域作为组织特异性增强子元件起作用。
位于2kb区域中的SNP
8个SNP(rs10490924、rs144224550、rs36212731、rs36212732、rs36212733、rs3750848、rs3750847、rs3750846)在Chr10 LD块中。
14.7 2Kb区域包括存在于野生型中而不是风险等位基因中的LHX2共有结合基序
使用HOMER、JASPER和TRANSFAC工具扫描已知的转录因子结合基序的序列,比较每个AMD相关SNP的非风险基因型和风险基因型。该分析将ARMS2内含子中的rs 36212733变体鉴定为转录因子结合位点,其具有预测的LHX2、POU6F1和/或ZNF33的基因型依赖性差异结合。其中,RPE中的LHX2 mRNA表达中等(FPKM>20),而POU6F1和ZNF333表达较低(FPKM<2)。在筛选的那些中,仅LHX2具有位于野生型而不是风险序列内的共有结合基序,并且该序列基序在ARMS2和lncSCTM1的内含子中几乎完美匹配(图9)。LHX2也在视网膜和脉络膜的一些细胞类型中表达,因此不是RPE特异性转录因子。与AMD风险相关的rs36212733SNP通过将第6位的“T”转换为“C”来破坏该基序中的一个关键残基。LHX2位点被SoxE转录因子家族成员的两个潜在结合位点包围,但这些位点不与Chr10相关SNP重叠。
不旨在受特定理论或机制的束缚,这些数据与模型一致,其中LHX2结合到基序,该基序与位于HTRA1上游的4kb AMD风险区中的rs36212733重叠并与ARMS2和lncSCTM1基因重叠。在非风险RPE细胞中,LHX2结合该基序以及RPE特异性辅因子,并作为HTRA1转录的增强子,从而以RPE特异性方式将HTRA1的表达增加到高于基础水平。目前尚不清楚RPE特异性辅因子是什么,其表达是否受年龄相关因素或其他外部信号的影响。如果LHX2与该增强子的结合阻碍了RNA聚合酶进行lncSCTM1转录的能力,则这将导致lncSCTM1基因的表达降低。由于预测LHX2仅结合非风险等位基因,这导致等位基因失衡,杂合供体中非风险等位基因相对于风险等位基因的水平降低。这种假设由我们的发现所支持,即RPE细胞中HTRA1和lncSCTM1的等位基因特异性表达之间存在反比关系(图8)。最后,在非RPE细胞中,由于缺乏RPE特异性辅因子,LHX2与非风险序列的结合不会增强HTRA1表达。然而,它仍然可干扰lncSCTM1转录,从而导致非风险等位基因的表达水平较低。因此,在非RPE组织中,只要细胞表达LHX2,就不存在HTRA1的等位基因特异性表达,但lncSCTM1的等位基因特异性表达仍然存在。
14.8 LHX2与野生型探针的结合比与包含风险序列的探针的结合更强
为了测试LHX2是否可结合该DNA序列,使用来自HEK293细胞的核提取物进行电泳迁移率变化测定(EMSA),所述HEK293细胞用编码LHX2的质粒或空载体以及含有Chr10区域的非风险(WT)序列或风险序列(RISK)(包括rs36212732和rs 36212733)的生物素化寡探针进行转染。还使用了包含该区域的乱序序列(SCR,阴性对照)或先前报道的LHX2结合位点(POS,阳性对照)(Muralidharan et al.(2017)J.Neurosci37(46):11245-54)的探针。结果示出LHX2结合到野生型和风险探针以及阳性对照探针,但不结合到乱序探针(图10A)。向反应中加入抗LHX2抗体导致LHX2带的超移,从而证实了LHX2的身份,并且未标记探针的加入导致带的丢失(图10B)。最后,通过在反应中添加不断增加量的探针来比较LHX2对野生型相对于对风险探针的结合亲和力。LHX2与野生型探针的结合比与包含风险序列的探针的结合更强(图11)。总的来说,这些结果示出LHX2可与包含rs36212733的序列结合,并且风险等位基因的存在显著降低了LHX2与该序列的结合。结合上述表观遗传数据和rs36212733与HTRA1表达的eQTL关联,结果支持如下假设:该区域作为RPE细胞中HTRA1表达的增强子起作用,并且LHX2可能有助于HTRA1表达。
14.9 CRISPRa系统用于上调HTRA1 mRNA表达的用途(慢病毒质粒)
供体RPE组织的微阵列和qRT-PCR分析展示出,与不具有风险的供体相比,在Chr10处具有纯合风险的供体中,HTRA1 mRNA的水平下调了大约30%。选择SAM CRISPRa系统以将RPE组织中HTRA1的表达恢复到与非风险供体相似的水平,同时最大限度地减少对基因表达的任何脱靶效应。SAM系统由两个质粒组成:LentiSAM质粒(SAM;Addgene#92062),其编码sgRNA和dCas9-VP64融合蛋白,以及LentiMPH(MPH;Addgene#92065)质粒,其编码由MS2亲和标签和p65以及HSF1转录因子组成的融合蛋白。
使用Benchling软件,设计sgRNA以覆盖HTRA1的启动子区域(图14)。这些序列连同已公布的靶向IL1B的sgRNA序列一起被克隆到邻近LentiSAM v2(Puro)质粒(Addgene)内的U6启动子的BsmBI位点。在具有或不具有等摩尔量的Addgene(Neo)质粒(总共5μg)的情况下,通过电穿孔用sgRNA-LentiSAMv2(Puro)质粒转染细胞。在24-96小时后,使用RNeasy试剂盒(Qiagen,目录号#74106)提取和纯化总RNA。使用1μg的总RNA和SuperScript IV VILOMaster Mix试剂盒(Invitrogen,目录号#11756050)生成互补DNA。使用50ng cDNA和TaqMan基因表达测定(Applied Biosystems),按照标准方案对HTRA1(Hs01016151_m1)、IL1B(Hs01555410_m1)和GAPDH(hs03929097_g1)进行定量PCR。
测试了表10中列出的sgRNA使用SAM CRISPRa系统上调HTRA1表达的能力(Konerman et al.,Nature,517(7536)2015)。
表10
Crispr激活的指导序列


图15示出大多数sgRNA将HTRA1上调到一定水平。P7和P18sgRNA示出HTRA1水平的最高增加(P7和P18 sgRNA分别增加了大约3.6倍和3.2倍)。
为了评估质粒中的任一者是否对mRNA水平具有脱靶效应,利用SAM组分单独地(MPH或SAM)或一起(MPH+SAM)转染h1RPE7细胞。在这些实验中,SAM质粒中的sgRNA不靶向任何已知的人类基因(Ctrl sgRNA)。如图16所示,相对于模拟转染细胞,在不存在HTRA1靶向sgRNA的情况下,MPH质粒将HTRA1的水平增加了大约1.8倍。类似地,在仅用SAM质粒转染的细胞中,HTRA1增加了大约1.6倍,而这并不由于MPH转录因子的存在而进一步增加。这些数据指示SAM系统组分可促进转录的普遍增加。
接着,进行实验以确定HTRA1的上调是否需要MPH质粒。在不存在或存在MPH质粒的情况下,测试最强的HTRA1靶向sgRNA(P7和P18,图17)对h1RPE7细胞中HTRA1表达的影响。据发现,即使在没有MPH质粒的情况下,SAM质粒也增加HTRA1水平(P7为5倍,P18为2.5倍),尽管与包含MPH时的水平(P7为7倍且P18为5倍)相比处于降低的水平。因此,看起来编码HTRA1靶向sgRNA和dCas9-VP64反式激活子的单一质粒足以上调HTRA1表达。此外,如果SAM系统浓度依赖于HTRA1水平,则效果随着SAM质粒量的减少而降低(图18)。
14.10 HTRA1 mRNA的增加与HTRA1蛋白水平的增加相关
进行初步实验以测试上述HTRA1 mRNA的增加是否与HtrA1蛋白水平的增加相关联。使用ELISA测定,测量了用LentiSAM质粒转染的h1RPE7细胞的细胞培养上清液中的HtrA1蛋白浓度。对于不同的时间点(3天、4天和5天)用不同量的P18 sgRNA-LentiSAM质粒(2.5、5.0和7.5μg)转染h1RPE7细胞。还用非靶向sgRNA(Ctrl)转染细胞作为对照。图19示出在转染后3天和4天,HTRA1蛋白水平在P18 sgRNA-LentiSAM转染的细胞中特异性增加。使用5μg LentiSAM质粒时,蛋白质水平达到其最大值。图20示出归一化为ENPP-2蛋白后的HTRA1蛋白水平。这些初始结果数据指示,相对于对照,在转染后3天和4天,在用P18 sgRNA-LentiSAM质粒转染的细胞中,HTRA1蛋白水平特异性增加。
上述数据展示出,在用P18 sgRNA-LentiSAM质粒瞬时转染的h1RPE7细胞中,HTRA1mRNA和蛋白质水平以剂量依赖性方式上调。
14.11慢病毒递送
构建了表11中所示的慢病毒颗粒。这些颗粒包含P18-lentiSAM质粒
表11.sgRNA-LentiSAM质粒的包装慢病毒
| 质粒 | 滴度(TU/mL) | 量(μl) | 数量 |
| Ctrl-LentiSAM | 3.18x10^8 | 25 | 4 |
| P18-LentiSAM | 2.33x10^8 | 25 | 4 |
| GFP | 1.49x10^9 | 50 | 1 |
测试了基于CRISPR的SAM系统的慢病毒递送是否可诱导RPE细胞中的HTRA1表达。用编码P18-LentiSAM质粒的慢病毒颗粒以20的MOI转导h1RPE7细胞,并在转导后每3天收集细胞培养上清液。作为对照,用编码非靶向sgRNA(Ctrl-LentiSAM)的慢病毒颗粒转导细胞。还用不具有病毒的含聚凝胺的培养基(模拟)处理细胞。在转导后3天、6天和9天收集的细胞培养上清液中的HTRA1蛋白水平通过HTRA1 ELISA测定法进行测量。如图21所示,与用Ctrl-LentiSAM病毒颗粒转导的细胞相比,在转导后6天和9天,在用P18-LentiSAM转导的细胞中,HTRA1蛋白水平增加大约40%(相对于Ctrl-LentiSAM增加1.4倍)。
为了解释每个单独样品转导之后细胞数量可能存在的差异,将HtrA1蛋白水平归一化为ENPP2蛋白水平,ENPP2蛋白水平在所有样品中随时间增加,但在对照和经P18-LentiSAM处理的细胞之间没有差异(数据未显示)。图23示出归一化为ENPP-2蛋白后的HTRA1蛋白水平。在转导后第6天和第9天,使用P18 sgRNA-LentiSAM转导之后,HtrA1的水平仍显示约30-40%的增加。为了测试HTRA1 mRNA表达和HTRA1分泌蛋白水平之间的相关性,从转导后9天提取的总RNA测量HTRA1的mRNA水平(图23)。与用Ctrl-LentiSAM处理的细胞相比,用P18-LentiSAM处理的细胞中的HTRA1 mRNA水平增加大约2倍。因此,响应于P18-LentiSAM处理,HTRA1 mRNA和蛋白质水平均增加。这指示基于CRISPR的SAM系统的病毒传递可成功诱导HTRA1。
用于上调HTRA1的AAV2-HTRA1质粒
本实施例中的实验旨在开发一种基于AAV2的载体,该载体携带在合适的RPE特异性启动子控制下的HTRA1基因,以恢复AMD患者的RPE中HTRA1的野生型表达水平。使用KpnI和SphI限制性位点将HTRA1基因从pCTM16质粒亚克隆到pCTM295中以生成pTR-HTRA1(pCTM289)。BEST1和RPE65启动子的片段使用具有Acc65I(正向)和BamHI限制性位点(反向)的引物进行PCR扩增。PCR产物用Acc65I和BamHI消化,并在这些位点亚克隆到pCTM289中,以在BEST1和RPE65来源的启动子片段的控制下产生HTRA1表达质粒。
表12示出用于由BEST1-启动子(SEQ ID NO:11)和RPE65启动子片段(SEQ ID NO:12)制备启动子片段中每一者的引物。稳定表达HTRA13-UTR靶向的shRNA质粒(并因此表达低水平的内源性HTRA1)的RPE1细胞,通过在含有10,000个细胞/孔的96孔板中,使用100μlNeon吸头电穿孔或通过脂质转染利用5μg指定质粒转染。总共使用了0.15μlLipofectamine 3000、0.2μl P3000和100ng DNA。转染后24-96小时收集细胞培养物上清液和RNA。执行我们的标准HTRA1 ELISA以测量蛋白质水平。使用RNeasy试剂盒(Qiagen,目录号74106)从细胞中提取总RNA。使用500ng总RNA和SuperScript IV VILO Master Mix试剂盒(Invitrogen,目录号11756050)生成互补DNA。对于HTRA1(Hs01016151_m1)和GAPDH(hs03929097_g1),使用50ng cDNA和TaqMan基因表达分析(Applied Biosystems)进行定量PCR。
表12
克隆到pTR-HTRA1质粒(pCTM289)中的BEST1和RPE65启动子的不同截断

测试了这些AAV-HTRA1质粒在汇集的RPE1克隆体群(7-6和7-7)中的HTRA1过表达,使用电穿孔稳定敲除HTRA1(>90%)。由于这些细胞中的HTRA1水平较低,因此信噪比得到改善,并将增加检测的灵敏度。作为阴性对照,使用pCTM259,其在与我们的质粒相同的AAV载体骨架中编码smCBA驱动的CFH基因。CFH过表达不影响HTRA1表达(数据未显示)。图24示出,与用pCTM259转染的细胞相比,所有BEST1-和RPE65-驱动的HTRA1构建体都不同程度地增加了HTRA1。
还在亲本RPE1细胞中测试了这些质粒。如图25所示,与阴性对照质粒(pCTM259)相比,在用BEST1-和RPE65-HTRA1质粒转染的细胞中HTRA1 mRNA增加。相对于彼此的HTRA1表达的模式在两种细胞系中是相似的。
选择了六个构建体,包括BEST1_723、BEST_699、BEST1_418、BEST1_340、RPE65_316和RPE65_146,用于进一步测试。进行时程分析以测量用最强候选质粒转染的细胞中HTRA1的mRNA和蛋白质水平。使用由不含内毒素的最大制备试剂盒制备的质粒DNA通过电穿孔进行转染以降低毒性,同时最大限度地提高转染效率。相对于对照质粒(pCTM259),由BEST1_723、BEST1_340和RPE65_146启动子驱动的几种AAV-HTRA1质粒显著增加了HTRA1 mRNA的水平(图26)。利用这些质粒中的每一者,mRNA水平在24小时达到峰值,并在48小时和72小时时间点逐渐下降。利用阳性对照CMV驱动的启动子,HTRA1水平也在24小时达到峰值,但在48小时和72小时时间点急剧下降。
我们检查了HTRA1蛋白表达的动力学并将其归一化为不受HTRA1过表达影响的VEGF水平(将pCTM259对照与图27B中的每个其他样品进行比较)。HTRA1的水平随时间增加,即使在pCTM259对照样品中也是如此。然而,HTRA1蛋白的相对增加显著大于用由BEST1_723、BEST1_340和RPE65_146启动子驱动的AAV-HTRA1质粒处理的细胞中的背景(图27A)。在归一化为VEGF后,HtrA1蛋白表达相对于对照质粒增加4至6倍,在24小时达到峰值并逐渐降低(图27C)。
这些数据一起展示出,通过基于BEST1或RPE65的启动子驱动的AAV2-HTRA1质粒的瞬时转染,可以上调mRNA和蛋白质水平上的HTRA1表达。
14.12 CRISPR介导的编辑
本实施例示出进行实验从而以等位基因特异性方式选择性删除Chr10上包含HTRA1调节区的DNA区域。具体地讲,CRISPR/Cas9系统和CRISPR指导RNA对的组合用于从RPE1细胞中的风险或野生型等位基因中特异性去除围绕rs10490924SNP的大部分DNA。
使用IDT Alt-R CRISPR系统进行转染。以下表13中列出了各个指导RNA的序列。将crRNA重新悬浮在Tris-EDTA溶液中,最终浓度为200μM。每个crRNA与tracrRNA(IDT目录号#1072532)在不含核酸酶的双链体缓冲液(IDT目录号#11-01-03-01)中以1:1的比率结合,并通过在95℃下加热2分钟进行退火,并逐渐冷却。crRNA:tracrRNA复合物用不含核酸酶的双链体缓冲液稀释,并将spCas9-3NLS(IDT目录号#1074181)加入到复合物中。样品在室温下温育20分钟以允许RNP形成。在温育期间,收获RPE1细胞并以5x107/ml的浓度重新悬浮在电穿孔缓冲液“R”缓冲液中。细胞与1.8μM的3'和5'RNP复合物连同1.8μM的载体ssDNA混合。对于每个反应,最终解决方案包含2μM spCas9-3NLS、1.8μM crRNA:tracrRNA和1.8μM载体ssDNA。使用10μl Neon移液管尖端在1300伏、脉冲宽度20和2个脉冲下对细胞-RNP混合物进行电穿孔,然后将其转移到6孔板中的3ml培养基中。24小时后,将转染的细胞连续稀释至15个细胞/ml的浓度。对于每个样品,将100μl(大约1.5个细胞/孔)接种到3x96孔板中。
表13.靶向rs10490924区域的CRISPR药剂

使用DNeasy Blood&Tissue试剂盒(QIAGEN,目录号#69504)从克隆培养物中纯化基因组DNA。使用200nM T7E1-Reg8-F(5'CTT ACCACCCTCGCTACATC3')和INDEL-DEL-R1(5'CCAGGGTGGTGTAATCC ATC3')引物在含有50ng基因组DNA、200μM dNTP(Thermo Fisher,目录号#18427)和Q5热启动高保真DNA聚合酶(NEB,目录号#M0493)的Q5 PCR缓冲液(NEB,目录号#B9027)中进行PCR。PCR产物通过琼脂糖凝胶电泳可视化。
RPE1细胞是染色体10q26的三倍体,其包含rs10490924野生型等位基因的两个拷贝和风险等位基因的单个拷贝。使用未经处理的RPE1细胞进行该区域的PCR扩增产生两个PCR产物,其对应于野生型等位基因的3.5kb带和风险等位基因的3.2kb带。琼脂糖凝胶电泳显示野生型相对于突变等位基因在密度方面的偏差为2:1。当细胞用针对野生型等位基因具有特异性的CRISPR指导对进行处理时,出现三种可能的结果。单个野生型等位基因可能被删除,如由1:1的3.5kb/3.2kb带连同从片段被移除的切割等位基因产生的600-800bp带所表明的。如果两个野生型等位基因都被切割,则PCR将仅从突变等位基因产生3.2kb带,从切割的野生型等位基因产生600-800bp带。相比之下,用针对突变等位基因具有特异性的CRISPR指导对进行处理的细胞应仅示出更密集的3.5kb野生型带和对应于切割突变等位基因的600-800bp带。
我们确定crRNA指导物的靶向对可以以等位基因依赖性方式有效去除大部分rs10490924区域(表14和图28)。这种效应在每个CRISPR指导对的大量转染RPE1细胞中得到证实(数据未示出)。基于这些结果,我们通过限制稀释大量种群来分离和建立单克隆培养物,以准确评估我们删除2kb区域的能力。通过切割位点周围的rs10490924区域的PCR扩增筛选来自单克隆培养物的基因组DNA,并通过琼脂糖凝胶电泳进行分析。图28示出一个示例,其中A2-WT2 crRNA的组合导致Chr10非风险等位基因(等位基因A)的缺失,而A2-INDEL1crRNA的组合导致Chr10风险等位基因(等位基因B)的缺失。crRNA的其他组合产生了预期的等位基因特异性缺失(数据未示出)。这表明我们可以以等位基因特异性方式特异性靶向染色体区域的去除。
表14.用于从rs10490924区域去除DNA片段的RNP复合物的组合


表15.命名法
| 简称 | 名称 | 删除 |
| A2 | ARMS2-2 | Allele A |
| WT2 | ARMS2-WT-3UTR-2 | 2868bp del |
| A2 | ARMS2-2 | Allele B |
| INDEL1 | ARMS2-INDELK-1 | 2535bp del |
15.参考文献
Cabrera AC,Melo E,Roth D,Topp A,Delobel F,Stucki C,Chen C,Jakob P,Banfai B,Dunkley T,Schilling O,Huber S,Iacone R,Petrone P.(2017)HtrA1activation is driven by an allosteric mechanism of intermonomercommunication.BioRxiv doi:10.1101/163717。
Chan C,Shen D,Zhou M,Ross R,Ding X,Zhang K,Green R,Tuo J.(2007)Humanhtra1 in the archived eyes with age-related macular degeneration.Trans AmOphthalmol Soc.2007;105:92 98。
Chowers I,Meir T,Lederman M,Goldenberg-Cohen N,Cohen Y,Banin E,Averbukh E,Hemo I,Pollack A,Axer-Siegel R,Weinstein O,Hoh J,Zack DJ,GalbinurT.(2008)Sequence variants in HTRA1 and LOC387715/ARMS2 and phenotype andresponse to photodynamic therapy in neovascular age-related maculardegeneration in populations from Israel.Mol Vis 14:2263-71。
Colijn JM,Buitendijk GHS,Prokofyeva E.et al.(2017)Prevalance of age-related macular degeneration in Europe:The past and the future.Ophthalmology124:1753-63。
Dewan A,Liu M,Hartman S,Zhang SS,Liu DT,Zhao C,Tam PO,Chan WM,Lam DS,Snyder M,Barnstable C,Pang CP,Hoh J.(2006)HTRA1 promoter polymorphism in wetage-related macular degeneration.Science:314:989-92。
Fisher SA,Abecasis GR,Yashar BM,et.al.(2005)Meta-analysis of genomescans of age-related macular degeneration.Hum Mol Genet 14:2257-64。
Flaxman SR,Bourne RRA,Resnikoff S,et al.(2017)Global causes ofblindness and distance vision impairment 1990-2020:a systematic review andmeta-analysis.Lancet Glob Health 5(12):e1221-e1234.doi:10.1016/S2214-109X(17)30393-5。
Fritsche LG,Loenhardt T,Janssen A,Fisher SA,Rivera A,Keilhauer CN,Weber BH.(2008)Age-related macular degeneration is associated with anunstable ARMS2(LOC387715)mRNA.Nature Genetics 40:892-6。
Fukutake,T.(2011)Cerebral Autosomal Recessive Arteriopathy withSubcortical Infarcts and Leukoencephalopathy(CARASIL):From Discovery to GeneIdentification.J Stroke Cerebrovasc Dis 20:85-93。
Lonsdale J,Thomas J,Salvatore M et al.(2013)The Genotype-TissueExpression(GTEx)project.Nat Genet 45:580-5。
The GTEx Consortium(2015)The Genotype-Tissue Expression(GTEx)pilotanalysis:Multitissue gene regulation in humans.Science 348,648 660。
Grassman F,Heid IM,Weber BH,International AMD Genomics Consortium.(2017)Recombinant haplotypes narrow the ARMS/HTRA1 association signal forage-related macular degeneration.Genetics 205:919-924。
Hara K,Shiga A,Fukutake T et al.(2009)Association of HTRA1 mutationsand familial sschemic cerebral small-vessel disease.N Engl J Med 360:1729-39。
Holz FG,Tadayoni R,Beatty S,Berger A,Cereda MG,Cortez R,Hoyng CB,Hykin P,Staurenghi G,Heldner S,Bogumil T,Heah T,Sivaprasad S.(2015)Multi-country real-life experience of anti-vascular endothelial growth factortherapy for wet age-related macular degeneration.Br J Ophthalmol 99(2):220-6。
Jones A,Kumar S,Zhang N,Tong Z,Yang JH,Watt C,Anderson J,Amrita,Fillerup H,McCloskey M,Luo L,Yang Z,Ambati B,Marc R,Oka C,Zhang K,Fu Y.(2011)Increased expression of multifunctional serine protease,HTRA1,in retinalpigment epithelium induces polypoidal choroidal vasculopathy in mice.Proc.Natl.Acad.Sci.U.S.A.Aug.30;108(35):14578-83。
Kanda A,Chen W,Othman M,Branham KE,Brooks M,Khanna R,He S,Lyons R,Abecasis GR,Swaroop A.(2007)A variant of mitochondrial protein LOC387715/ARMS2,not HTRA1,is strongly associated with age-related maculardegeneration.PNAS 104:16227-32。
Kortvely E,Hauck SM,Duetsch G,Gloeckner CJ,Kremmer E,Alge-PriglingerCS,Deeg CA,Ueffing M.(2010)ARMS2 is a constituent of the extracellular matrixproviding a link between familial and sporadic age-related maculardegenerations.Invest Ophtalmol Vis Sci 51:79-88。
Lau C and Suh Y.(2017)In vivo genome editing in animals using AAV-CRISPR system:applications to translational research of human disease.(2017)F1000 Res.2017;6:2153;doi:10.12688/f1000research.11243.1。
Lambert SA,Jolma A,Campitelli LF,Das PK,Yin Y,Albu M,Chen X,TaipaleJ,Hughes TR,Weirauch MT(2018)Cell172(4):650-665。
Moore DJ and Clover GM.(2001)The effect of age on the macromolecularpermeability of human Bruch’s membrane.Invest Ophthalmol Vis Sci 42:2970-5。
Peng Y,Shekhar K,Yan W,Herrmann D,Sappington A,Bryman GS,vanZyl T,DoMTH,Regev A and Sanes JR.2019 Molecular classification and comparativetaxonomics of foveal and peripheral cells in primate retina.Cell 176:5:1222-1237。
Regillo CD,Busbee BG,Ho AC,Ding B,Haskova Z(2015)Baseline predictorsof 12-month treatment response to ranibizumab in patients with wet age-related macular degeneration.Am J Ophthalmol 160(5):1014-23。
Rivera A,Fisher SA,Fritsche LG,Keilhauer CN,Lichtner P,Meitinger T,Weber BH.(2005)Hypothetical LOC387715 is a second major susceptibility genefor age-related macular degeneration,contributing independently of complementfactor H to disease risk.Hum Mol Genet 14:3227-36。
Vierkotten S,Muether P,Fauser S(2011)Overexpression of HTRA1 Leads toUltrastructural Changes in the Elastic Layer of Bruch's Membrane via Cleavageof Extracellular Matrix Components.PLoS One.2011;6(8):e22959。
Wang N,Eckert K,Zomorrodi A,Xin P,Pan W,Shearer D,Weisz J,Maranus C,Clawson G,(2012)Plos|One,https://doi.org/10.1371/journal.pone.0039446。
Wang G,Scott WK,Whitehead P,Court BL,Kovach JL,Schwartz SG,Agarwal A,Dubovy S,Haines JL,Pericak-Vance MA.(2012)A novel ARMS2 splice variant isidentified in human retina.Exp.Eye Res 94:187-91。
Wong WL,Su X,Li X,Cheung CM,Klein R,Cheng CY,Wong TY(2014)Globalprevalence of age-related macular degeneration and disease burden projectionfor 2020and 2040:a systematic review and meta-analysis.Lancet Glob Health 2(2):e106-16.doi:10.1016/S2214-109X(13)70145-1。
Yang Z,Camp NJ,Sun H,Tong Z,Gibbs D,Cameron DJ,Chen H,Zhao Y,PearsonE,Li X,Chien J,Dewan A,Harmon J,Bernstein PS,Shridhar V,Zabriskie NA,Hoh J,Howes K,Zhang K.(2006)A variant of the HTRA1 gene increases susceptibility toage-related macular degeneration.Science 314:992-3。
Zellner A,Scharrer E,Arzberger T et al.(2018)CADASIL brain vesselsshow a HTRA1 loss-of-function profile.Acta Neuropathol 136:111-125。
表16.序列汇总


尽管为了清楚理解的目的,已经通过说明和示例的方式对前述发明进行了一些详细描述,但是本领域技术人员将理解,可在所附权利要求的范围内实施某些改变和修改。
本发明可以在不脱离其结构、方法或其他基本特征的情况下以其他具体形式体现,如本文中广泛描述的和在下文中要求保护的。所述方法在所有方面都被认为仅是说明性的,而不是限制性的。因此,本发明的范围由所附权利要求而不是由前述说明指示。在权利要求的等效含义和范围内的所有变化都应包含在其范围内。
本说明书中引用的所有出版物和专利均以引用方式并入本文,就好像每个单独的出版物或专利都被明确地且单独地指示为以引用方式并入那样,并以引用方式并入本文以公开和描述与出版物所引用的方法和/或材料相关的方法和/或材料。任何出版物的引用都是为了其在申请日之前的公开内容,并且不应被解释为承认本发明无权因在先发明而早于此类出版物。此外,所提供的出版日期可能与实际出版日期不同,所述实际出版日期可能需要独立确认。
序列表
<110> 犹他大学研究基金会
<120>用于治疗 AMD 的 HTRA1 调节
<130> 098846-1185529-000810PC
<140>
<141>
<150> 62/832,182
<151> 2019-04-10
<160> 94
<170>专利版本 3.5
<210> 1
<211> 1374
<212> DNA
<213> 智人
<400> 1
cagctgtccc gggccggccg ctcggcgcct ttggccgccg ggtgcccaga ccgctgcgag 60
ccggcgcgct gcccgccgca gccggagcac tgcgagggcg gccgggcccg ggacgcgtgc 120
ggctgctgcg aggtgtgcgg cgcgcccgag ggcgccgcgt gcggcctgca ggagggcccg 180
tgcggcgagg ggctgcagtg cgtggtgccc ttcggggtgc cagcctcggc cacggtgcgg 240
cggcgcgcgc aggccggcct ctgtgtgtgc gccagcagcg agccggtgtg cggcagcgac 300
gccaacacct acgccaacct gtgccagctg cgcgccgcca gccgccgctc cgagaggctg 360
caccggccgc cggtcatcgt cctgcagcgc ggagcctgcg gccaagggca ggaagatccc 420
aacagtttgc gccataaata taactttatc gcggacgtgg tggagaagat cgcccctgcc 480
gtggttcata tcgaattgtt tcgcaagctt ccgttttcta aacgagaggt gccggtggct 540
agtgggtctg ggtttattgt gtcggaagat ggactgatcg tgacaaatgc ccacgtggtg 600
accaacaagc accgggtcaa agttgagctg aagaacggtg ccacttacga agccaaaatc 660
aaggatgtgg atgagaaagc agacatcgca ctcatcaaaa ttgaccacca gggcaagctg 720
cctgtcctgc tgcttggccg ctcctcagag ctgcggccgg gagagttcgt ggtcgccatc 780
ggaagcccgt tttcccttca aaacacagtc accaccggga tcgtgagcac cacccagcga 840
ggcggcaaag agctggggct ccgcaactca gacatggact acatccagac cgacgccatc 900
atcaactatg gaaactcggg aggcccgtta gtaaacctgg acggtgaagt gattggaatt 960
aacactttga aagtgacagc tggaatctcc tttgcaatcc catctgataa gattaaaaag 1020
ttcctcacgg agtcccatga ccgacaggcc aaaggaaaag ccatcaccaa gaagaagtat 1080
attggtatcc gaatgatgtc actcacgtcc agcaaagcca aagagctgaa ggaccggcac 1140
cgggacttcc cagacgtgat ctcaggagcg tatataattg aagtaattcc tgatacccca 1200
gcagaagctg gtggtctcaa ggaaaacgac gtcataatca gcatcaatgg acagtccgtg 1260
gtctccgcca atgatgtcag cgacgtcatt aaaagggaaa gcaccctgaa catggtggtc 1320
cgcaggggta atgaagatat catgatcaca gtgattcccg aagaaattga ccca 1374
<210> 2
<211> 458
<212> PRT
<213> 智人
<400> 2
Gln Leu Ser Arg Ala Gly Arg Ser Ala Pro Leu Ala Ala Gly Cys Pro
1 5 10 15
Asp Arg Cys Glu Pro Ala Arg Cys Pro Pro Gln Pro Glu His Cys Glu
20 25 30
Gly Gly Arg Ala Arg Asp Ala Cys Gly Cys Cys Glu Val Cys Gly Ala
35 40 45
Pro Glu Gly Ala Ala Cys Gly Leu Gln Glu Gly Pro Cys Gly Glu Gly
50 55 60
Leu Gln Cys Val Val Pro Phe Gly Val Pro Ala Ser Ala Thr Val Arg
65 70 75 80
Arg Arg Ala Gln Ala Gly Leu Cys Val Cys Ala Ser Ser Glu Pro Val
85 90 95
Cys Gly Ser Asp Ala Asn Thr Tyr Ala Asn Leu Cys Gln Leu Arg Ala
100 105 110
Ala Ser Arg Arg Ser Glu Arg Leu His Arg Pro Pro Val Ile Val Leu
115 120 125
Gln Arg Gly Ala Cys Gly Gln Gly Gln Glu Asp Pro Asn Ser Leu Arg
130 135 140
His Lys Tyr Asn Phe Ile Ala Asp Val Val Glu Lys Ile Ala Pro Ala
145 150 155 160
Val Val His Ile Glu Leu Phe Arg Lys Leu Pro Phe Ser Lys Arg Glu
165 170 175
Val Pro Val Ala Ser Gly Ser Gly Phe Ile Val Ser Glu Asp Gly Leu
180 185 190
Ile Val Thr Asn Ala His Val Val Thr Asn Lys His Arg Val Lys Val
195 200 205
Glu Leu Lys Asn Gly Ala Thr Tyr Glu Ala Lys Ile Lys Asp Val Asp
210 215 220
Glu Lys Ala Asp Ile Ala Leu Ile Lys Ile Asp His Gln Gly Lys Leu
225 230 235 240
Pro Val Leu Leu Leu Gly Arg Ser Ser Glu Leu Arg Pro Gly Glu Phe
245 250 255
Val Val Ala Ile Gly Ser Pro Phe Ser Leu Gln Asn Thr Val Thr Thr
260 265 270
Gly Ile Val Ser Thr Thr Gln Arg Gly Gly Lys Glu Leu Gly Leu Arg
275 280 285
Asn Ser Asp Met Asp Tyr Ile Gln Thr Asp Ala Ile Ile Asn Tyr Gly
290 295 300
Asn Ser Gly Gly Pro Leu Val Asn Leu Asp Gly Glu Val Ile Gly Ile
305 310 315 320
Asn Thr Leu Lys Val Thr Ala Gly Ile Ser Phe Ala Ile Pro Ser Asp
325 330 335
Lys Ile Lys Lys Phe Leu Thr Glu Ser His Asp Arg Gln Ala Lys Gly
340 345 350
Lys Ala Ile Thr Lys Lys Lys Tyr Ile Gly Ile Arg Met Met Ser Leu
355 360 365
Thr Ser Ser Lys Ala Lys Glu Leu Lys Asp Arg His Arg Asp Phe Pro
370 375 380
Asp Val Ile Ser Gly Ala Tyr Ile Ile Glu Val Ile Pro Asp Thr Pro
385 390 395 400
Ala Glu Ala Gly Gly Leu Lys Glu Asn Asp Val Ile Ile Ser Ile Asn
405 410 415
Gly Gln Ser Val Val Ser Ala Asn Asp Val Ser Asp Val Ile Lys Arg
420 425 430
Glu Ser Thr Leu Asn Met Val Val Arg Arg Gly Asn Glu Asp Ile Met
435 440 445
Ile Thr Val Ile Pro Glu Glu Ile Asp Pro
450 455
<210> 3
<211> 1440
<212> DNA
<213> 智人
<400> 3
atgcagatcc cgcgcgccgc tcttctcccg ctgctgctgc tgctgctggc ggcgcccgcc 60
tcggcgcagc tgtcccgggc cggccgctcg gcgcctttgg ccgccgggtg cccagaccgc 120
tgcgagccgg cgcgctgccc gccgcagccg gagcactgcg agggcggccg ggcccgggac 180
gcgtgcggct gctgcgaggt gtgcggcgcg cccgagggcg ccgcgtgcgg cctgcaggag 240
ggcccgtgcg gcgaggggct gcagtgcgtg gtgcccttcg gggtgccagc ctcggccacg 300
gtgcggcggc gcgcgcaggc cggcctctgt gtgtgcgcca gcagcgagcc ggtgtgcggc 360
agcgacgcca acacctacgc caacctgtgc cagctgcgcg ccgccagccg ccgctccgag 420
aggctgcacc ggccgccggt catcgtcctg cagcgcggag cctgcggcca agggcaggaa 480
gatcccaaca gtttgcgcca taaatataac tttatcgcgg acgtggtgga gaagatcgcc 540
cctgccgtgg ttcatatcga attgtttcgc aagcttccgt tttctaaacg agaggtgccg 600
gtggctagtg ggtctgggtt tattgtgtcg gaagatggac tgatcgtgac aaatgcccac 660
gtggtgacca acaagcaccg ggtcaaagtt gagctgaaga acggtgccac ttacgaagcc 720
aaaatcaagg atgtggatga gaaagcagac atcgcactca tcaaaattga ccaccagggc 780
aagctgcctg tcctgctgct tggccgctcc tcagagctgc ggccgggaga gttcgtggtc 840
gccatcggaa gcccgttttc ccttcaaaac acagtcacca ccgggatcgt gagcaccacc 900
cagcgaggcg gcaaagagct ggggctccgc aactcagaca tggactacat ccagaccgac 960
gccatcatca actatggaaa ctcgggaggc ccgttagtaa acctggacgg tgaagtgatt 1020
ggaattaaca ctttgaaagt gacagctgga atctcctttg caatcccatc tgataagatt 1080
aaaaagttcc tcacggagtc ccatgaccga caggccaaag gaaaagccat caccaagaag 1140
aagtatattg gtatccgaat gatgtcactc acgtccagca aagccaaaga gctgaaggac 1200
cggcaccggg acttcccaga cgtgatctca ggagcgtata taattgaagt aattcctgat 1260
accccagcag aagctggtgg tctcaaggaa aacgacgtca taatcagcat caatggacag 1320
tccgtggtct ccgccaatga tgtcagcgac gtcattaaaa gggaaagcac cctgaacatg 1380
gtggtccgca ggggtaatga agatatcatg atcacagtga ttcccgaaga aattgaccca 1440
<210> 4
<211> 480
<212> PRT
<213> 智人
<400> 4
Met Gln Ile Pro Arg Ala Ala Leu Leu Pro Leu Leu Leu Leu Leu Leu
1 5 10 15
Ala Ala Pro Ala Ser Ala Gln Leu Ser Arg Ala Gly Arg Ser Ala Pro
20 25 30
Leu Ala Ala Gly Cys Pro Asp Arg Cys Glu Pro Ala Arg Cys Pro Pro
35 40 45
Gln Pro Glu His Cys Glu Gly Gly Arg Ala Arg Asp Ala Cys Gly Cys
50 55 60
Cys Glu Val Cys Gly Ala Pro Glu Gly Ala Ala Cys Gly Leu Gln Glu
65 70 75 80
Gly Pro Cys Gly Glu Gly Leu Gln Cys Val Val Pro Phe Gly Val Pro
85 90 95
Ala Ser Ala Thr Val Arg Arg Arg Ala Gln Ala Gly Leu Cys Val Cys
100 105 110
Ala Ser Ser Glu Pro Val Cys Gly Ser Asp Ala Asn Thr Tyr Ala Asn
115 120 125
Leu Cys Gln Leu Arg Ala Ala Ser Arg Arg Ser Glu Arg Leu His Arg
130 135 140
Pro Pro Val Ile Val Leu Gln Arg Gly Ala Cys Gly Gln Gly Gln Glu
145 150 155 160
Asp Pro Asn Ser Leu Arg His Lys Tyr Asn Phe Ile Ala Asp Val Val
165 170 175
Glu Lys Ile Ala Pro Ala Val Val His Ile Glu Leu Phe Arg Lys Leu
180 185 190
Pro Phe Ser Lys Arg Glu Val Pro Val Ala Ser Gly Ser Gly Phe Ile
195 200 205
Val Ser Glu Asp Gly Leu Ile Val Thr Asn Ala His Val Val Thr Asn
210 215 220
Lys His Arg Val Lys Val Glu Leu Lys Asn Gly Ala Thr Tyr Glu Ala
225 230 235 240
Lys Ile Lys Asp Val Asp Glu Lys Ala Asp Ile Ala Leu Ile Lys Ile
245 250 255
Asp His Gln Gly Lys Leu Pro Val Leu Leu Leu Gly Arg Ser Ser Glu
260 265 270
Leu Arg Pro Gly Glu Phe Val Val Ala Ile Gly Ser Pro Phe Ser Leu
275 280 285
Gln Asn Thr Val Thr Thr Gly Ile Val Ser Thr Thr Gln Arg Gly Gly
290 295 300
Lys Glu Leu Gly Leu Arg Asn Ser Asp Met Asp Tyr Ile Gln Thr Asp
305 310 315 320
Ala Ile Ile Asn Tyr Gly Asn Ser Gly Gly Pro Leu Val Asn Leu Asp
325 330 335
Gly Glu Val Ile Gly Ile Asn Thr Leu Lys Val Thr Ala Gly Ile Ser
340 345 350
Phe Ala Ile Pro Ser Asp Lys Ile Lys Lys Phe Leu Thr Glu Ser His
355 360 365
Asp Arg Gln Ala Lys Gly Lys Ala Ile Thr Lys Lys Lys Tyr Ile Gly
370 375 380
Ile Arg Met Met Ser Leu Thr Ser Ser Lys Ala Lys Glu Leu Lys Asp
385 390 395 400
Arg His Arg Asp Phe Pro Asp Val Ile Ser Gly Ala Tyr Ile Ile Glu
405 410 415
Val Ile Pro Asp Thr Pro Ala Glu Ala Gly Gly Leu Lys Glu Asn Asp
420 425 430
Val Ile Ile Ser Ile Asn Gly Gln Ser Val Val Ser Ala Asn Asp Val
435 440 445
Ser Asp Val Ile Lys Arg Glu Ser Thr Leu Asn Met Val Val Arg Arg
450 455 460
Gly Asn Glu Asp Ile Met Ile Thr Val Ile Pro Glu Glu Ile Asp Pro
465 470 475 480
<210> 5
<211> 469
<212> DNA
<213> 智人
<400> 5
gcgcgtgacc ggggtccgcg gtgccgcaac gccccgggtc tgcgcagagg cccctgcagt 60
ccctgcccgg cccagtccga gcttcccggg cgggccccca gtccggcgat ttgcaggaac 120
tttccccggc gctcccacgc gaagccgccg cagggccccc ttgcaaagtt ccattagttt 180
gaaggacgcg aatctcagcg agagaacctg cggaaagcga atatgtgggg cgcgcagacg 240
gggaaactga gtcccgcgag agggccggcc tgtgcgctgc cccgcccgcg ccccgccagc 300
accgccgtgc ccgggcgcgc ccgccctgcc ccctccgcgg gcggtcccgg tccagccgcc 360
cgccctccct cccgccatcc ggccagcccc catcccgggc gccgtgcccg tccccaaggc 420
ggctcgtcac cgctgcgagg ccaatgggct gggccgcgcg gccgcgcgc 469
<210> 6
<211> 796
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 6
caattctgtc attttactag ggtgatgaaa ttcccaagca acaccatcct tttcagataa 60
gggcactgag gctgagagag gagctgaaac ctacccggcg tcaccacaca caggtggcaa 120
ggctgggacc agaaaccagg actgttgact gcagcccggt attcattctt tccatagccc 180
acagggctgt caaagacccc agggcctagt cagaggctcc tccttcctgg agagttcctg 240
gcacagaagt tgaagctcag cacagccccc taacccccaa ctctctctgc aaggcctcag 300
gggtcagaac actggtggag cagatccttt agcctctgga ttttagggcc atggtagagg 360
gggtgttgcc ctaaattcca gccctggtct cagcccaaca ccctccaaga agaaattaga 420
ggggccatgg ccaggctgtg ctagccgttg cttctgagca gattacaaga agggaccaag 480
acaaggactc ctttgtggag gtcctggctt agggagtcaa gtgacggcgg ctcagcactc 540
acgtgggcag tgccagcctc taagagtggg caggggcact ggccacagag tcccagggag 600
tcccaccagc ctagtcgcca gaccggggat cctctagagg atccggtact cgaggaactg 660
aaaaaccaga aagttaactg gtaagtttag tctttttgtc ttttatttca ggtcccggat 720
ccggtggtgg tgcaaatcaa agaactgctc ctcagtggat gttgccttta cttctaggcc 780
tgtacggaag tgttac 796
<210> 7
<211> 400
<212> DNA
<213> 智人
<400> 7
tccattagtt tgaaggacgc gaatctcagc gagagaacct gcggaaagcg aatatgtggg 60
gcgcgcagac ggggaaactg agtcccgcga gagggccggc ctgtgcgctg ccccgcccgc 120
gccccgccag caccgccgtg cccgggcgcg cccgccctgc cccctccgcg ggcggtcccg 180
gtccagccgc ccgccctccc tcccgccatc cggccagccc ccatcccggg cgccgtgccc 240
gtccccaagg cggctcgtca ccgctgcgag gccaatgggc tgggccgcgc ggccgcgcgc 300
actcgcaccc gctgcccccg aggccctcct gcactctccc cggcgccgct ctccggccct 360
cgccctgtcc gccgccaccg ccgccgccgc cagagtcgcc 400
<210> 8
<211> 300
<212> DNA
<213> 智人
<400> 8
tccattagtt tgaaggacgc gaatctcagc gagagaacct gcggaaagcg aatatgtggg 60
gcgcgcagac ggggaaactg agtcccgcga gagggccggc ctgtgcgctg ccccgcccgc 120
gccccgccag caccgccgtg cccgggcgcg cccgccctgc cccctccgcg ggcggtcccg 180
gtccagccgc ccgccctccc tcccgccatc cggccagccc ccatcccggg cgccgtgccc 240
gtccccaagg cggctcgtca ccgctgcgag gccaatgggc tgggccgcgc ggccgcgcgc 300
<210> 9
<211> 102
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 9
cgccgtgccc gtccccaagg gttttagagc tagaaatagc aagttaaaat aaggctagtc 60
cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt tt 102
<210> 10
<211> 59
<212> DNA
<213> 智人
<400> 10
gtggcgcttt gtgcttgcca tagtatatat aaytagacaa atgagagaac acaaaggtt 59
<210> 11
<211> 723
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 11
ctctgaagca acttactgat gggccctgcc agccaatcac agccagaata acgtatgatg 60
tcaccagcag ccaatcagag ctcctcgtca gcatatgcag aattctgtca ttttactagg 120
gtgatgaaat tcccaagcaa caccatcctt ttcagataag ggcactgagg ctgagagagg 180
agctgaaacc tacccggggt caccacacac aggtggcaag gctgggacca gaaaccagga 240
ctgttgactg cagcccggta ttcattcttt ccatagccca cagggctgtc aaagacccca 300
gggcctagtc agaggctcct ccttcctgga gagttcctgg cacagaagtt gaagctcagc 360
acagccccct aacccccaac tctctctgca aggcctcagg ggtcagaaca ctggtggagc 420
agatccttta gcctctggat tttagggcca tggtagaggg ggtgttgccc taaattccag 480
ccctggtctc agcccaacac cctccaagaa gaaattagag gggccatggc caggctgtgc 540
tagccgttgc ttctgagcag attacaagaa gggactaaga caaggactcc tttgtggagg 600
tcctggctta gggagtcaag tgacggcggc tcagcactca cgtgggcagt gccagcctct 660
aagagtgggc aggggcactg gccacagagt cccagggagt cccaccagcc tagtcgccag 720
acc 723
<210> 12
<211> 750
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 12
atactctcag agtgccaaac atataccaat ggacaagaag gtgaggcaga gagcagacag 60
gcattagtga caagcaaaga tatgcagaat ttcattctca gcaaatcaaa agtcctcaac 120
ctggttggaa gaatattggc actgaatggt atcaataagg ttgctagaga gggttagagg 180
tgcacaatgt gcttccataa cattttatac ttctccaatc ttagcactaa tcaaacatgg 240
ttgaatactt tgtttactat aactcttaca gagttataag atctgtgaag acagggacag 300
ggacaatacc catctctgtc tggttcatag gtggtatgta atagatattt ttaaaaataa 360
gtgagttaat gaatgagggt gagaatgaag gcacagaggt attaggggga ggtgggcccc 420
agagaatggt gccaaggtcc agtggggtga ctgggatcag ctcaggcctg acgctggcca 480
ctcccaccta gctcctttct ttctaatctg ttctcattct ccttgggaag gattgaggtc 540
tctggaaaac agccaaacaa ctgttatggg aacagcaagc ccaaataaag ccaagcatca 600
gggggatctg agagctgaaa gcaacttctg ttccccctcc ctcagctgaa ggggtgggga 660
agggctccca aagccataac tccttttaag ggatttagaa ggcataaaaa ggcccctggc 720
tgagaacttc cttcttcatt ctgcagttgg 750
<210> 13
<211> 853
<212> DNA
<213> 智人
<400> 13
gtcccaacgg atgcaccaaa gattctccag tgggaaatca aatttttgat aacaagtgtt 60
ttgaaataat cacgcacttg gtgaaaaatc aaaagagggg aaaacccctt tcccatctga 120
gaccgctcca ccctcgccag ttacgagctg ccgagccgct tcctaggctc tctgcgaata 180
cggacacgca tgccacccac aacaactttt taaaagaatc agacgtgtga aggattctat 240
tcgaattact tctgctctct gcttttatca cttcactgtg ggtctgggcg cgggctttct 300
gccagctccg cggacgctgc cttcgtccgg ccgcagaggc cccgcggtca gggtcccgcg 360
tgcggggtac cgggggcaga accagcgcgt gaccggggtc cgcggtgccg caacgccccg 420
ggtctgcgca gaggcccctg cagtccctgc ccggcccagt ccgagcttcc cgggcgggcc 480
cccagtccgg cgatttgcag gaactttccc cggcgctccc acgcgaagcc gccgcagggc 540
ccccttgcaa agttccatta gtttgaagga cgcgaatctc agcgagagaa cctgcggaaa 600
gcgaatatgt ggggcgcgca gacggggaaa ctgagtcccg cgagagggcc ggcctgtgcg 660
ctgccccgcc cgcgccccgc cagcaccgcc gtgcccgggc gcgcccgccc tgccccctcc 720
gcgggcggtc ccggtccagc cgcccgccct ccctcccgcc atccggccag cccccatccc 780
gggcgccgtg cccgtcccca aggcggctcg tcaccgctgc gaggccaatg ggctgggccg 840
cgcggccgcg cgc 853
<210> 14
<211> 2056
<212> DNA
<213> 智人
<400> 14
gatactaggg acctctgttg cctcctctgg cagagcagga ctgaggggtg gaccctccct 60
gagaccaccc aacaattcag ggtggagtta tcagggcgcc ctgactcctg ggggcatttt 120
tgtgtgacgg gaaaagacaa tgctcctggc tgagtgagat ggcagctggc ttggcaaggg 180
gacagcacct ttgtcaccac attatgtccc tgtaccctac atgctgcgcc tatacccagg 240
accgatggta actgaggcgg aggggaaagg agggcctgag atggcaagtc tgtcctcctc 300
ggtggttcct gtgtccttca tttccactct gcgagagtct gtgctggacc ctggagttgg 360
tggagaagga gccagtgaca agcagaggag caaactgtct ttatcacact ccatgatccc 420
agctgctaaa atccacactg agctctgctt accagccttc ttctctcctg ctggaaccca 480
gaggaggttc cagcagcctc agcaccacct gacactggta agaaatgcag atgatcaggc 540
cttaccccag acctattgaa tcagaaattc tggagtggtg ccctgcagct tgcattttaa 600
ccagccttca ggtgcttctg atgcatgctc aggtttgagc accactggcc acagggaggc 660
ctaggcaatt cagccttcct ctggttgaat agctggagaa ttgggaatat cagtaaatac 720
ttccaatgca cctgctacat gccagaaaaa ggaaacaaga agacgcagta ggtctgagaa 780
agtgatgggg tgagcagaaa cccaaagctt atagaaggcc atctgagtgg cccctcaagc 840
cggtgaattg gctttagggt ttactgaagg aggtggaaac ctcagcctgc ttctcgtccg 900
ggttgttaga ggagtcattt agaaagctgt accattcttt caatattctc acggctttcc 960
agtgctcatt tttcctgctc atttatggat taaaaaaaat ccttggctgt atgtgtaaga 1020
aaacaacaat gcaagtttgt agagaaagaa tctgggcctt acaggtcacg ttggtttaaa 1080
atttagacat caagcagctt agagaccatg ttgccaaata agcttagtaa atgctttcta 1140
atgcttacgg aactgtggcg ctttgtgctt gccatagtat atataattag acaaatgaga 1200
gaacacaaag gttgaacccc ttccctctct taatttttgt tttttacaag cagatttaaa 1260
attctggctc ataatgtcct tgattcaatg ttaaaccatt ttgcctaaat ggcagcatgt 1320
tctaaatgtg agcgcgctca gcttttcaag ctgctcccga gtgacagaaa ttgacaagct 1380
gtcattcaag acctttcggt ggctgcctgg ggctctgttt gaattgtatc tgtctgatac 1440
tttaccatgg agagtgaaaa atttgatcac atgccatgct tttaattttc taaagcaaat 1500
atgttggaag gagccaatta atgcaaagat ggactgctgg tctcatgcaa ctgatttagg 1560
ggaagggttc gcctaaatta ataaaagatc tgaattatag atcttaacaa atacatagaa 1620
tgtaaaggct taaaaggaaa ctgagcagca gcaggcctgg ggttggcttt taagtatcta 1680
tatttaacta atagacatga atgttttgat ttgatattag aaatgctagt gctggagtct 1740
ctgagcctac tctggctcga gaggatgccc tatctaaaaa acaaaaaaca aaaaaaaaaa 1800
agaaaaaaga aaaaaagaaa tgctggtatt gtaattctaa agtgcttcag aaattctcaa 1860
aaataggcca ggcatggtgg ctcatggctg tataccagca ctttgggagg ccaaggtggg 1920
caaatcactt gcagtcagga gtttgagacc agcctggtca acatggtgaa accccatctc 1980
tactaaaaat acagaaatta gccaggcatg gtggcagcac ctgtaatccc agctacttgg 2040
gaggctgagg caggag 2056
<210> 15
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 15
cacgcgggac cctgaccgcg 20
<210> 16
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 16
tgcccccggt accccgcacg 20
<210> 17
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 17
cccggggcgt tgcggcaccg 20
<210> 18
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 18
cctgcaaatc gccggactgg 20
<210> 19
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 19
ctgcggaaag cgaatatgtg 20
<210> 20
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 20
ggaaactgag tcccgcgaga 20
<210> 21
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 21
gggctggccg gatggcggga 20
<210> 22
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 22
agcggtgacg agccgccttg 20
<210> 23
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 23
actgagtccc gcgagagggc 20
<210> 24
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 24
cgcacaggcc ggccctctcg 20
<210> 25
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 25
gcgcgggcgg ggcagcgcac 20
<210> 26
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 26
cccgccagca ccgccgtgcc 20
<210> 27
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 27
agggcgggcg cgcccgggca 20
<210> 28
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 28
cgccctgccc cctccgcggg 20
<210> 29
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 29
ggaccgggac cgcccgcgga 20
<210> 30
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 30
ggagggcggg cggctggacc 20
<210> 31
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 31
gggcacggcg cccgggatgg 20
<210> 32
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 32
cgccgtgccc gtccccaagg 20
<210> 33
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 33
gtcaccgctg cgaggccaat 20
<210> 34
<211> 4090
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 34
tttaagcaat tcttttgcct cagcctctgg aattacaggc acgcgccacc atgccctgct 60
aatttttgta tttttagtag agacagggtt ccaccatgtt ggccaggctg atctggaact 120
cctgacctca agtgatctgc ccgcctcggc ctcccaaagt gctgggatta caggtgtgag 180
ccaccgcacc tggccaacta ttttaaaaaa ttttaagaga ctgagtctca ctctgtcacc 240
cagtctggtg cagtggcacg atcatggctt cagccttaaa gtcctgggct caagcaatcc 300
tcccacctca gccccccgag tagctgggaa tacaagtgtg cacagtcaca ctaaactaaa 360
aaaaaaaaaa aaaactaatt ttattttatt ttattttatt ttttatttat ttattttttg 420
tagggatggg tttttgctat gttgcctagg ctggtctcaa agtcctggcc tcaagcaatc 480
ctcctgcttc atcctcccaa agtgctgtga ttacaggcat gagccactgt tcccagcctc 540
tccccactct tctagcccta agcaagcaca aatctatttt ctgtatctgt agattttcct 600
gttatacaca tttcttatga atgaaatcat ataatatgtt gtcttttgta tctggcttct 660
ttcatttagc cccatgcttt cagggttcac catgttgtag tgggtatttt attctttttt 720
atggctgaat agtatttcat tttatggata tgccataatt tatctgttat cagttgatgg 780
acatttgggt tgtttctacc tttgggctgt tatgaatagt gctgctatga acatgtgtgt 840
tccagtttct gtatggacac atgttttcct ttctcttggg tagctaccta ggaatggaaa 900
tgctggatca tatattcact ctgtttgaag aactgccaga ctgttttcca aagcagctat 960
accattttac aatcccacta gcagtgcatg aggattctga tttctccaca tccttgctga 1020
tacttgttat catctgactt tttgattctg gctaccttag tgcctatgaa gtagtatctc 1080
aatgtggttt tgatttgttg atttgctgat gactagagat gccaagcatc ttctcatgtg 1140
tttatttgtg ctcttagagt ttttaattca gttggtctgg aatagttttt tttttccttt 1200
tattttttat ttttttgaga cagagtcttg ctctgtcacc aagctggagt gcagtggcgt 1260
gatcttggct cactgcaacc tctgactccc tggttcgggc tattctcctg cctcagcctc 1320
ccgagtaact gggattacag gcacgcacca ccatgcccag ctaatttttg tatttttagt 1380
agagccagga ttccaccatg ttagccatga tggtctcgat ctcctgagct cgtgatctgt 1440
ccaccttggc ttcccaaagt ggtgggatga caggcgtgag ccatggcgcc cggctttttt 1500
tttcctttta aaaaagtgtt tggctgggag aagtggttca cacctgtaat cccaacactt 1560
tgggaggccg aggcaggagg atcacttgag cccaggactt caagacctgc ctgggcaaca 1620
tagtaagacc ccatctccaa aaaaaaaaaa aaaaagtaaa ttagccaggc gtggtgctgc 1680
acacctgtgg tcccagctac ttgggaggct gaggtgggag gatcacctga gccgaggagt 1740
atgaggctgc agtgagccat gatcatgcca ctggattcca gcctgggtga caaagtgaga 1800
ccctgtctca gaaaaaaaaa aaaaggctgg ggggcctggg ctaggattat gcctgtaatc 1860
ctagcacttt tgcaggtaga ttgcttgagc tcggagttcg ataccagcct ggcaacatgg 1920
caaaacctgg tctctacaaa aaacataaaa aatagctggg cgtggtctgt agctgtagtc 1980
ccagctactc aggtggctga ggtaggagga tcaccttagc ccgaggatgt tgaggccaca 2040
ataagctgtg gtcataccac tgcacaccag cctggacaac agaatgagac cccatctcaa 2100
aaaaaaaagt tttttatttt gaaataattt taggtttaga gaaaagttga aaaatagtaa 2160
aacaacaaca acaacaaaaa aacaacaaaa aatcccaaaa cccccaaaac tctcattgac 2220
ccttatctca gatttcccga atgcttacca ccctcgctac atcattcaag ttcttggaaa 2280
catttaaagc atgtgaaaca tttaaaacat ttaagttgga ggctttaagt tgcacgtcct 2340
ttatttctaa atatttcagt gtgtttttct taaaaaaaat tttctcatac cacagtaaca 2400
tgatcaaaat tggaaaatca acattgattc aatactatga tctacaatca aggttttttt 2460
tttttttcaa atccctgggt ctctgcattt tttaaaagct tcacagatga tttcaatgga 2520
tactagggac ctctgttgcc tcctctggca gagcaggact gaggggtgga ccctccctga 2580
gaccacccaa caattcaggg tggagttatc agggcgccct gactcctggg ggcatttttg 2640
tgtgacggga aaagacaatg ctcctggctg agtgagatgg cagctggctt ggcaagggga 2700
cagcaccttt gtcaccacat tatgtccctg taccctacat gctgcgccta tacccaggac 2760
cgatggtaac tgaggcggag gggaaaggag ggcctgagat ggcaagtctg tcctcctcgg 2820
tggttcctgt gtccttcatt tccactctgc gagagtctgt gctggaccct ggagttggtg 2880
gagaaggagc cagtgacaag cagaggagca aactgtcttt atcacactcc atgatcccag 2940
ctgctaaaat ccacactgag ctctgcttac cagccttctt ctctcctgct ggaacccaga 3000
ggaggttcca gcagcctcag caccacctga cactggtaag aaatgcagat gatcaggcct 3060
taccccagac ctattgaatc agaaattctg gagtggtgcc ctgcagcttg cattttaacc 3120
agccttcagg tgcttctgat gcatgctcag gtttgagcac cactggccac agggaggcct 3180
aggcaattca gccttcctct ggttgaatag ctggagaatt gggaatatca gtaaatactt 3240
ccaatgcacc tgctacatgc cagaaaaagg aaacaagaag acgcagtagg tctgagaaag 3300
tgatggggtg agcagaaacc caaagcttat agaaggccat ctgagtggcc cctcaagccg 3360
gtgaattggc tttagggttt actgaaggag gtggaaacct cagcctgctt ctcgtccggg 3420
ttgttagagg agtcatttag aaagctgtac cattctttca atattctcac ggctttccag 3480
tgctcatttt tcctgctcat ttatggatta aaaaaaatgc cttggctgta tgtgtaagaa 3540
aacaacaatg caagtttgta gagaaagaat ctgggcctta caggtcacgt tggtttaaaa 3600
tttagacatc aagcagctta gagaccatgt tgccaaataa gcttagtaaa tgctttctaa 3660
tgcttacgga actgtggcgc tttgtgcttg ccatagtata tataattaga caaatgagag 3720
aacacaaagg ttgaacccct tccctctctt aatttttgtt ttttacaagc agatttaaaa 3780
ttctggctca taatgtcctt gattcaatgt taaaccattt tgcctaaatg gcagcatgtt 3840
ctaaatgtga gcgcgctcag cttttcaagc tgctcccgag tgacagaaat tgacaagctg 3900
tcattcaaga cctttcggtg gctgcctggg gctctgtttg aattgtatct gtctgatact 3960
ttaccatgga gagtgaaaaa tttgatcaca tgccatgctt ttaattttct aaagcaaata 4020
tgttggaagg agccaattaa tgcaaagatg gactgctggt ctcatgcaac tgatttaggg 4080
gaagggttcg 4090
<210> 35
<211> 1443
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成多肽
<400> 35
atgcagattc ctagagctgc tctgctgccc ctgctgctgt tgcttcttgc tgctcctgct 60
agcgcccagc tgtccagagc tggaagatct gctcctctgg ccgctggctg tcctgataga 120
tgcgaacctg ccagatgtcc tcctcagcct gagcactgtg aaggcggcag agccagagat 180
gcctgcggct gttgtgaagt gtgcggagca cctgaaggcg ccgcttgtgg acttcaagaa 240
ggaccttgcg gagagggcct gcagtgcgtt gtgccttttg gagtgcctgc ctctgccaca 300
gttaggcgga gagcacaagc tggcctgtgt gtgtgcgcct cttctgagcc agtgtgtggc 360
tccgacgcca acacctacgc caatctgtgt cagctgagag ccgccagcag aagaagcgag 420
agactgcaca gacctccagt gatcgtgctg cagagaggcg cttgcggaca gggacaagag 480
gaccccaata gcctgcggca caagtacaac tttatcgccg acgtggtgga aaagatcgcc 540
cctgccgtgg tgcacatcga gctgttcaga aagcttccat tcagcaagcg cgaggtgcca 600
gtggcctctg gatctggctt tatcgtgtcc gaggacggcc tgatcgtgac aaatgcccac 660
gtggtcacca acaagcacag agtgaaggtg gaactgaaga acggcgccac ctacgaggcc 720
aagatcaagg acgtggacga gaaggccgac attgccctga tcaagatcga ccaccagggc 780
aagctgccag tgctgctgct tggcagaagc agcgaactca gacctggcga gtttgtggtg 840
gccatcggaa gccctttcag cctgcaaaac accgtgacca ccggcatcgt gtccaccaca 900
caaagaggcg gcaaagagct gggcctgaga aacagcgaca tggactacat ccagaccgac 960
gccatcatca actacggcaa ctctggcggc cctctggtca acctggatgg cgaagtgatc 1020
ggcatcaaca ccctgaaagt gacagccggc atcagcttcg ctatccccag cgataagatc 1080
aagaagttcc tgaccgagag ccacgaccgg caggccaagg gaaaagccat caccaagaag 1140
aagtacatcg gaatccggat gatgagcctg accagcagca aggccaaaga actgaaggac 1200
cggcacagag acttccccga tgtgatctct ggcgcctaca tcattgaagt gatccccgac 1260
acaccagccg aagccggcgg actgaaagaa aacgacgtga tcatcagcat caacggccag 1320
agcgtggtgt ccgccaacga tgtgtctgat gtgatcaagc gggaaagcac cctgaacatg 1380
gttgtgcgga ggggcaacga ggatatcatg atcaccgtga ttcccgagga aatcgacccc 1440
tga 1443
<210> 36
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 36
ctggtaagca gagctcagtg 20
<210> 37
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 37
aaggagccag tgacaagcag 20
<210> 38
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 38
tggttaaaat gcaagctgca 20
<210> 39
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 39
ttgaatcaga aattctggag 20
<210> 40
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 40
tgagcaggaa aaatgagcac 20
<210> 41
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 41
ccgtgagaat attgaaagaa 20
<210> 42
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 42
ttgtctagtt atatatacta 20
<210> 43
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 43
agacaaatga gagaacacaa 20
<210> 44
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 44
aatggtttac cattgaatca 20
<210> 45
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 45
gcttgtcaat ttctgtcact 20
<210> 46
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 46
agccccaggc agccaccaaa 20
<210> 47
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 47
tacaattcaa acagagcccc 20
<210> 48
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 48
gcctcatgca actgatttag 20
<210> 49
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 49
ttaatgcaaa gatggactgc 20
<210> 50
<211> 20
<212> DNA
<213> 智人
<400> 50
gaaggacaca ggaaccaccg 20
<210> 51
<211> 20
<212> DNA
<213> 智人
<400> 51
cccaggaccg atggtaactg 20
<210> 52
<211> 20
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 52
taataaccgg gagagaaagg 20
<210> 53
<211> 28
<212> DNA
<213> 智人
<400> 53
ttgccatagt atatataayt agacaaat 28
<210> 54
<211> 81
<212> RNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 54
guuuuagagc uagaaauagc aaguuaaaau aaggcuaguc cguuaucaac uugaaaaagu 60
ggcaccgagu cggugcuuuu u 81
<210> 55
<211> 26
<212> RNA
<213> 人工序列
<220>
<223> 人工序列描述:合成寡核苷酸
<400> 55
aacaugagga ucacccaugu cugcag 26
<210> 56
<211> 130
<212> PRT
<213> 人工序列
<220>
<223> 人工序列描述: 合成
多肽
<400> 56
Met Ala Ser Asn Phe Thr Gln Phe Val Leu Val Asp Asn Gly Gly Thr
1 5 10 15
Gly Asp Val Thr Val Ala Pro Ser Asn Phe Ala Asn Gly Val Ala Glu
20 25 30
Trp Ile Ser Ser Asn Ser Arg Ser Gln Ala Tyr Lys Val Thr Cys Ser
35 40 45
Val Arg Gln Ser Ser Ala Gln Lys Arg Lys Tyr Thr Ile Lys Val Glu
50 55 60
Val Pro Lys Val Ala Thr Gln Thr Val Gly Gly Val Glu Leu Pro Val
65 70 75 80
Ala Ala Trp Arg Ser Tyr Leu Asn Met Glu Leu Thr Ile Pro Ile Phe
85 90 95
Ala Thr Asn Ser Asp Cys Glu Leu Ile Val Lys Ala Met Gln Gly Leu
100 105 110
Leu Lys Asp Gly Asn Pro Ile Pro Ser Ala Ile Ala Ala Asn Ser Gly
115 120 125
Ile Tyr
130
<210> 57
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 57
cggggtaccc cctgccagcc aatcaca 27
<210> 58
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 58
cgcggatcca aatccagagg ctaaaggatc tg 32
<210> 59
<211> 30
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 59
cggggtaccc tctgaagcaa cttactgatg 30
<210> 60
<211> 26
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 60
cgcggatccg gtctggcgac taggct 26
<210> 61
<211> 26
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 61
cggggtaccc ctgccagcca atcaca 26
<210> 62
<211> 31
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 62
cgcggatccc tgtgctgagc ttcaacttct g 31
<210> 63
<211> 26
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 63
cggggtaccc ctgccagcca atcaca 26
<210> 64
<211> 26
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 64
cgcggatccg gtctggcgac taggct 26
<210> 65
<211> 29
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 65
cggggtaccc aatcagagct cctcgtcag 29
<210> 66
<211> 31
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 66
cgcggatccc tgtgctgagc ttcaacttct g 31
<210> 67
<211> 28
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 67
cggggtaccg aacactggtg gagcagat 28
<210> 68
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 68
cgcggatcca ctccctggga ctctgtg 27
<210> 69
<211> 31
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 69
cggggtaccg aaattcccaa gcaacaccat c 31
<210> 70
<211> 29
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 70
cgcggatcct tcttcttgga gggtgttgg 29
<210> 71
<211> 35
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 71
cggggtaccc tggttcatag gtggtatgta ataga 35
<210> 72
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 72
cgcggatcca gagaatggtg ccaaggt 27
<210> 73
<211> 31
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 73
cggggtacca tactctcaga gtgccaaaca t 31
<210> 74
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 74
cgcggatccc caactgcaga atgaagaagg aa 32
<210> 75
<211> 29
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 75
cggggtaccg agggttagag gtgcacaat 29
<210> 76
<211> 30
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 76
cgcggatcca aaggagttat ggctttggga 30
<210> 77
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 77
cggggtaccg agaatgaagg cacagaggta tt 32
<210> 78
<211> 34
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 78
cgcggatccc caaggagaat gagaacagat taga 34
<210> 79
<211> 31
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 79
cggggtacca acctggttgg aagaatattg g 31
<210> 80
<211> 34
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 80
cgcggatccc caaggagaat gagaacagat taga 34
<210> 81
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 81
cggggtaccg agaatgaagg cacagaggta tt 32
<210> 82
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 82
cgcggatccc caactgcaga atgaagaagg aa 32
<210> 83
<211> 27
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 83
cggggtacca gagaatggtg ccaaggt 27
<210> 84
<211> 32
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 84
cgcggatccc caactgcaga atgaagaagg aa 32
<210> 85
<211> 29
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 85
cggggtaccg agggttagag gtgcacaat 29
<210> 86
<211> 34
<212> DNA
<213> 人工序列
<220>
<223> 人工序列描述: 合成
引物
<400> 86
cgcggatccc caaggagaat gagaacagat taga 34
<210> 87
<211> 21
<212> DNA
<213> 智人
<400> 87
gatcccagct gctaaaatcc a 21
<210> 88
<211> 22
<212> DNA
<213> 智人
<400> 88
attctggagt ggtgccctgc ag 22
<210> 89
<211> 21
<212> DNA
<213> 智人
<400> 89
atattctcac ggctttccag t 21
<210> 90
<211> 21
<212> DNA
<213> 智人
<400> 90
tgtgcttgcc atagtatata t 21
<210> 91
<211> 21
<212> DNA
<213> 智人
<400> 91
gtatatataa ttagacaaat g 21
<210> 92
<211> 21
<212> DNA
<213> 智人
<400> 92
tgattcaatg ttaaaccatt t 21
<210> 93
<211> 21
<212> DNA
<213> 智人
<400> 93
caagaccttt cggtggctgc c 21
<210> 94
<211> 21
<212> DNA
<213> 智人
<400> 94
ggactgctgg tctcatgcaa c 21
Claims (43)
1.一种指导RNA(gRNA),其包含对应于HTRA1启动子或HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸的指导序列。
2.根据权利要求1所述的gRNA,其中所述HTRA1启动子具有SEQ ID NO:5、7、8或13中列出的序列,并且所述2kb调节区具有SEQ ID NO:14中列出的序列。
3.一种核糖核蛋白(RNP)复合物,其包含:
a)指导RNA(gRNA),其包含对应于HTRA1启动子或HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸的指导序列以及
b)融合蛋白,其包含与转录激活子结构域融合的CRISPR相关蛋白(Cas)结构域,
其中所述Cas蛋白结构域缺乏核酸酶活性并且
其中所述HTRA1启动子具有SEQ ID NO:5、7、8或13中列出的序列,并且所述2kb调节区具有SEQ ID NO:13中列出的序列。
4.一种核糖核蛋白(RNP)复合物,其包含
a)指导RNA(gRNA),其包含对应于HTRA1 2kb调节区中的靶序列的至少10个连续核苷酸的指导序列以及,
ii)CRISPR-相关(Cas)蛋白,
其中所述2kb调节区具有SEQ ID NO:13中列出的序列。
5.根据权利要求1所述的gRNA或根据权利要求3或权利要求4所述的RNP复合物,其中所述指导序列包含对应于所述靶序列的至少15个连续核苷酸。
6.根据权利要求1所述的gRNA或根据权利要求3或权利要求4所述的RNP复合物,其中所述指导序列包含对应于所述靶序列的至少20个连续核苷酸。
7.根据权利要求1所述的gRNA或根据权利要求3或权利要求4所述的RNP复合物,其中所述指导序列包含对应于所述靶序列的至少15-25个连续核苷酸。
8.根据权利要求1所述的gRNA或根据权利要求3或权利要求4所述的RNP复合物,其中所述靶序列与为NGG的前间隔序列邻近基序(PAM)相邻。
9.根据权利要求3所述的RNP复合物,其中Cas为dCas9或dCas12a。
10.根据权利要求4所述的RNP复合物,其中Cas为Cas9、Cas12a或Cas3。
11.根据权利要3所述的RNP复合物,其中所述靶序列在启动子中并且所述转录激活子选自VP16、VP64、VP160、MLL、E2A、HSF1、NF-IL6、NFAT1和NF-kB。
12.根据权利要3所述的RNP复合物,其中所述靶序列在所述2kb调节区中并且所述转录激活子为LHX2。
13.根据权利要求1-2或4-7所述的gRNA或根据权利要求3所述的RNP复合物,其中所述指导序列包含SEQ ID NO:15-33中的任一者,或包含SEQ ID NO:15-33中任一者的至少15个连续碱基的子序列。
14.根据权利要求1-2所述的gRNA或根据权利要求4所述的RNP复合物,其中:
i)指导序列包含SEQ ID NOS:36-49中的任一者,
ii)靶序列包含风险等位基因,所述风险等位基因选自下列中任一者处的风险:rs10490924、rs144224550、rs36212731、rs36212732、rs36212733、rs3750848、rs3750847以及rs3750846,
iii)靶序列邻近下列中的任一者:rs10490924、rs144224550、rs36212731、rs36212732、rs 36212733、rs3750848、rs3750847以及rs3750846。
15.一种多核苷酸,其编码根据权利要求1-2、5-8或14-15中任一项所述的gRNA,其中所述多核苷酸为DNA。
16.根据权利要求16所述的多核苷酸,其包含可操作地连接到编码gRNA的序列的启动子。
17.一种病毒载体,其包含根据权利要求16或17所述的多核苷酸。
18.一种病毒载体,其包含编码HTRA1多肽的多核苷酸,其中所述多核苷酸包含与启动子可操作地连接的编码HTRA1的人密码子优化序列。
19.根据权利要求18或权利要求19所述的病毒载体,其为逆转录病毒、慢病毒、疱疹病毒或腺相关病毒(AAV)。
20.根据权利要求17所述的多核苷酸或根据权利要求19-20所述的病毒载体,其中所述启动子是RPE特异性启动子。
21.一种HTRA1激活系统,其包含:
a)包含编码根据权利要求1、2、5-8或14-15中任一项所述的gRNA的DNA序列的载体,以及
b)包含编码融合蛋白的DNA序列的载体,所述融合蛋白包含与转录激活子结构域融合的Cas蛋白结构域,其中所述Cas蛋白结构域缺乏核酸酶活性。
22.根据权利要求22所述的HTRA1激活系统,其中(a)中的载体和(b)中的载体是不同的载体。
23.一种HTRA1靶向系统,其包含:
(a)包含编码根据权利要求1、2、5-8或14-15中任一项所述的gRNA的核酸的载体,以及
(b)包含编码Cas蛋白质的核酸的载体。
24.根据权利要求24所述的HTRA1靶向系统,其还包含:
(c)包含编码模板修复序列的核酸的载体,所述模板修复序列任选地包含根据SEQ IDNO:87-94中的至少一个或SEQ ID NO:87-94中至少一个的互补序列。
25.根据权利要求25所述的HTRA1靶向系统,其中(a)和(b)是相同的载体,或(a)、(b)和(c)是相同的载体。
26.一种分离的细胞,其包含根据权利要求1或2所述的gRNA,根据权利要求3或4所述的RNP复合物,根据权利要求16、17或21所述的多核苷酸、根据权利要求18-20所述的病毒载体、根据权利要求22-23所述的激活系统、或根据权利要求24-26所述的靶向系统。
27.根据权利要求1-2、4-7或14-15中任一项的gRNA,根据权利要求16所述的分离的多核苷酸,根据权利要求18-20所述的载体,根据权利要求22-23所述的激活系统,根据权利要求24-26所述的靶向系统,或根据权利要求27所述的分离细胞用于制备用于治疗年龄相关性黄斑变性(AMD)的药物的用途。
28.根据权利要求1-2、4-7或14-15中任一项的指导RNA,根据权利要求16所述的分离的多核苷酸,根据权利要求18-20所述的载体,根据权利要求22-23所述的激活系统,根据权利要求24-26所述的靶向系统,或根据权利要求27所述的分离细胞,其用于制备用于治疗年龄相关性黄斑变性(AMD)的药物。
29.根据权利要求28所述的用途或根据权利要求29所述的指导RNA,其中被治疗的受试者:
a)表现出Chr 10AMD临床表型;
b)具有10号染色体风险等位基因;
c)对于10号染色体风险等位基因是纯合的;或者
d)不具有1号染色体风险等位基因。
30.一种用于增加细胞中HTRA1表达的方法,所述方法包括在细胞中表达根据权利要求22-23所述的激活系统或根据权利要求24-26所述的靶向系统。
31.一种治疗受试者的Chr 10AMD、预防其发展、减缓其进展、逆转或改善其症状和体征的方法,所述方法包括施用增加受试者RPE细胞或水平细胞中HTRA1表达的一种或多种药剂。
32.根据权利要求32所述的方法,其中所述受试者表现出Chr 10AMD临床表型。
33.根据权利要求32或33所述的方法,其中所述受试者具有10号染色体风险等位基因。
34.根据权利要求34所述的方法,其中所述受试者对于10号染色体风险等位基因是纯合的。
35.根据权利要求34或35所述的方法,其中所述受试者不具有1号染色体风险等位基因。
36.根据权利要求32-36中任一项所述的方法,其中内源性HTRA1基因序列的转录增加。
37.根据权利要求37所述的方法,其中所述药剂为核糖核蛋白复合物,其包含(a)无酶活性的Cas蛋白结构域和转录激活子结构域的融合蛋白和(b)指导RNA。
38.根据权利要求38所述的方法,其中所述无酶活性的Cas蛋白是dCas9。
39.根据权利要求38所述的方法,其中所述核糖核蛋白复合物结合在HTRA1启动子区中。
40.根据权利要求38所述的方法,其中核糖核蛋白复合物结合在HTRA1增强子区中。
41.根据权利要求39所述的方法,其中所述转录激活子结构域结合LHX2结合基序。
42.根据权利要求37所述的方法,其中所述药剂为包含指导RNA和Cas蛋白的核糖核酸复合物。
43.根据权利要求37所述的方法,其中所述受试者携带在所述HTRA1基因增强子中的风险等位基因,并且所述药剂是包含下列的组合:(a)包含指导RNA和Cas蛋白的核糖核酸复合物和(ii)模板修复多核苷酸,其包含对应于风险等位基因的非风险等位基因的序列。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US201962832182P | 2019-04-10 | 2019-04-10 | |
| US62/832,182 | 2019-04-10 | ||
| PCT/US2020/027802 WO2020210724A1 (en) | 2019-04-10 | 2020-04-10 | Htra1 modulation for treatment of amd |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| CN113966401A true CN113966401A (zh) | 2022-01-21 |
Family
ID=72750902
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN202080042929.4A Pending CN113966401A (zh) | 2019-04-10 | 2020-04-10 | 用于治疗amd的htra1调节 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US20230165976A1 (zh) |
| EP (1) | EP3953485A4 (zh) |
| JP (1) | JP2022526429A (zh) |
| CN (1) | CN113966401A (zh) |
| CA (1) | CA3136119A1 (zh) |
| WO (1) | WO2020210724A1 (zh) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN113512564A (zh) * | 2021-08-05 | 2021-10-19 | 呈诺再生医学科技(珠海横琴新区)有限公司 | 以iPSC来源的细胞为载体、基因治疗湿性年龄相关性黄斑变性 |
| WO2023023278A2 (en) * | 2021-08-18 | 2023-02-23 | University Of Utah Research Foundation | Multigene constructs for treatment of age-related macular degeneration and other complement dysregulation-related conditions |
| WO2023086663A2 (en) * | 2021-11-15 | 2023-05-19 | The Children's Medical Center Corporation | Methods for tauopathy diagnosis and treatment |
Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2008013893A2 (en) * | 2006-07-26 | 2008-01-31 | Yale University | Diagnosis and treatment of age related macular degeneration |
| US20110111407A1 (en) * | 2008-04-08 | 2011-05-12 | Ehrmann Michael | Method for analysing the epigenetic status of the htra 1 gene in a biological sample |
| WO2012125869A1 (en) * | 2011-03-15 | 2012-09-20 | University Of Utah Research Foundation | Methods of diagnosing and treating vascular associated maculopathy and symptoms thereof |
| WO2013055998A1 (en) * | 2011-10-14 | 2013-04-18 | Genentech, Inc. | ANTI-HtrA1 ANTIBODIES AND METHODS OF USE |
| WO2018169983A1 (en) * | 2017-03-13 | 2018-09-20 | President And Fellows Of Harvard College | Methods of modulating expression of target nucleic acid sequences in a cell |
Family Cites Families (33)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE69128362T2 (de) | 1990-06-01 | 1998-05-20 | Chiron Corp | Zusammensetzungen und verfahren zur identifizierung von molekülen mit biologischer wirksamkeit |
| CA2118806A1 (en) | 1991-09-18 | 1993-04-01 | William J. Dower | Method of synthesizing diverse collections of oligomers |
| US5487994A (en) | 1992-04-03 | 1996-01-30 | The Johns Hopkins University | Insertion and deletion mutants of FokI restriction endonuclease |
| US5356802A (en) | 1992-04-03 | 1994-10-18 | The Johns Hopkins University | Functional domains in flavobacterium okeanokoites (FokI) restriction endonuclease |
| US5436150A (en) | 1992-04-03 | 1995-07-25 | The Johns Hopkins University | Functional domains in flavobacterium okeanokoities (foki) restriction endonuclease |
| CA2143848C (en) | 1992-10-01 | 2007-09-11 | W. Clark Still | Complex combinatorial chemical libraries encoded with tags |
| JPH09508353A (ja) | 1993-11-02 | 1997-08-26 | アフィマックス テクノロジーズ ナムローゼ フェンノートシャップ | 分子多様性の合成及びスクリーニング |
| WO1995030642A1 (en) | 1994-05-06 | 1995-11-16 | Pharmacopeia, Inc. | Combinatorial dihydrobenzopyran library |
| US5663046A (en) | 1994-06-22 | 1997-09-02 | Pharmacopeia, Inc. | Synthesis of combinatorial libraries |
| DE19710159A1 (de) | 1997-03-12 | 1998-10-01 | Hoechst Ag | In vitro Transkriptionsverfahren zum Screening von Naturstoffen und anderen chemischen Substanzen |
| US6534261B1 (en) | 1999-01-12 | 2003-03-18 | Sangamo Biosciences, Inc. | Regulation of endogenous gene expression in cells using zinc finger proteins |
| US6599692B1 (en) | 1999-09-14 | 2003-07-29 | Sangamo Bioscience, Inc. | Functional genomics using zinc finger proteins |
| US6453242B1 (en) | 1999-01-12 | 2002-09-17 | Sangamo Biosciences, Inc. | Selection of sites for targeting by zinc finger proteins and methods of designing zinc finger proteins to bind to preselected sites |
| US7070934B2 (en) | 1999-01-12 | 2006-07-04 | Sangamo Biosciences, Inc. | Ligand-controlled regulation of endogenous gene expression |
| US7030215B2 (en) | 1999-03-24 | 2006-04-18 | Sangamo Biosciences, Inc. | Position dependent recognition of GNN nucleotide triplets by zinc fingers |
| US6794136B1 (en) | 2000-11-20 | 2004-09-21 | Sangamo Biosciences, Inc. | Iterative optimization in the design of binding proteins |
| ATE309536T1 (de) | 1999-12-06 | 2005-11-15 | Sangamo Biosciences Inc | Methoden zur verwendung von randomisierten zinkfingerprotein-bibliotheken zur identifizierung von genfunktionen |
| WO2001059450A2 (en) | 2000-02-08 | 2001-08-16 | Sangamo Biosciences, Inc. | Cells expressing zinc finger protein for drug discovery |
| US7067317B2 (en) | 2000-12-07 | 2006-06-27 | Sangamo Biosciences, Inc. | Regulation of angiogenesis with zinc finger proteins |
| US7262054B2 (en) | 2002-01-22 | 2007-08-28 | Sangamo Biosciences, Inc. | Zinc finger proteins for DNA binding and gene regulation in plants |
| US7361635B2 (en) | 2002-08-29 | 2008-04-22 | Sangamo Biosciences, Inc. | Simultaneous modulation of multiple genes |
| US7888121B2 (en) | 2003-08-08 | 2011-02-15 | Sangamo Biosciences, Inc. | Methods and compositions for targeted cleavage and recombination |
| US7972854B2 (en) | 2004-02-05 | 2011-07-05 | Sangamo Biosciences, Inc. | Methods and compositions for targeted cleavage and recombination |
| AU2005233550B2 (en) | 2004-04-08 | 2010-11-18 | Sangamo Therapeutics, Inc. | Treatment of neuropathic pain with zinc finger proteins |
| CA2561714A1 (en) | 2004-04-08 | 2005-10-27 | Sangamo Biosciences, Inc. | Methods and compositions for treating neuropathic and neurodegenerative conditions |
| CA2659392A1 (en) | 2006-07-13 | 2008-01-17 | University Of Iowa Research Foundation | Methods and reagents for treatment and diagnosis of vascular disorders and age-related macular degeneration |
| US7972787B2 (en) | 2007-02-16 | 2011-07-05 | Massachusetts Eye And Ear Infirmary | Methods for detecting age-related macular degeneration |
| US20150211065A1 (en) | 2012-09-14 | 2015-07-30 | University Of Utah Research Foundation | Methods of predicting the development of amd based on chromosome 1 and chromosome 10 |
| LT3066201T (lt) * | 2013-11-07 | 2018-08-10 | Editas Medicine, Inc. | Su crispr susiję būdai ir kompozicijos su valdančiomis grnr |
| RU2685914C1 (ru) * | 2013-12-11 | 2019-04-23 | Регенерон Фармасьютикалс, Инк. | Способы и композиции для направленной модификации генома |
| KR20230152175A (ko) | 2014-04-18 | 2023-11-02 | 에디타스 메디신, 인코포레이티드 | 암 면역요법을 위한 crispr-cas-관련 방법, 조성물 및 구성성분 |
| SG10202106058WA (en) | 2016-12-08 | 2021-07-29 | Intellia Therapeutics Inc | Modified guide rnas |
| WO2020019002A1 (en) | 2018-07-20 | 2020-01-23 | University Of Utah Research Foundation | Gene therapy for macular degeneration |
-
2020
- 2020-04-10 CA CA3136119A patent/CA3136119A1/en active Pending
- 2020-04-10 CN CN202080042929.4A patent/CN113966401A/zh active Pending
- 2020-04-10 WO PCT/US2020/027802 patent/WO2020210724A1/en unknown
- 2020-04-10 EP EP20787458.7A patent/EP3953485A4/en active Pending
- 2020-04-10 US US17/599,979 patent/US20230165976A1/en active Pending
- 2020-04-10 JP JP2021559892A patent/JP2022526429A/ja active Pending
Patent Citations (5)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2008013893A2 (en) * | 2006-07-26 | 2008-01-31 | Yale University | Diagnosis and treatment of age related macular degeneration |
| US20110111407A1 (en) * | 2008-04-08 | 2011-05-12 | Ehrmann Michael | Method for analysing the epigenetic status of the htra 1 gene in a biological sample |
| WO2012125869A1 (en) * | 2011-03-15 | 2012-09-20 | University Of Utah Research Foundation | Methods of diagnosing and treating vascular associated maculopathy and symptoms thereof |
| WO2013055998A1 (en) * | 2011-10-14 | 2013-04-18 | Genentech, Inc. | ANTI-HtrA1 ANTIBODIES AND METHODS OF USE |
| WO2018169983A1 (en) * | 2017-03-13 | 2018-09-20 | President And Fellows Of Harvard College | Methods of modulating expression of target nucleic acid sequences in a cell |
Also Published As
| Publication number | Publication date |
|---|---|
| JP2022526429A (ja) | 2022-05-24 |
| EP3953485A1 (en) | 2022-02-16 |
| EP3953485A4 (en) | 2023-05-17 |
| WO2020210724A1 (en) | 2020-10-15 |
| US20230165976A1 (en) | 2023-06-01 |
| CA3136119A1 (en) | 2020-10-15 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US20210017509A1 (en) | Gene Editing for Autosomal Dominant Diseases | |
| US20200255859A1 (en) | Cellular models of and therapies for ocular diseases | |
| EP3289080B1 (en) | Gene therapy for autosomal dominant diseases | |
| CN113966401A (zh) | 用于治疗amd的htra1调节 | |
| US11492614B2 (en) | Stem loop RNA mediated transport of mitochondria genome editing molecules (endonucleases) into the mitochondria | |
| JP2022505139A (ja) | ゲノム編集の方法及び構築物 | |
| KR20210049833A (ko) | Mma의 치료를 위한 비-붕괴적 유전자 요법 | |
| KR20200095462A (ko) | Hbb 유전자 기능 회복을 위한 아데노-연관 바이러스 조성물 및 이의 사용 방법 | |
| WO2019134561A1 (en) | High efficiency in vivo knock-in using crispr | |
| JP2021520837A (ja) | トランススプライシング分子 | |
| CN111718420B (zh) | 一种用于基因治疗的融合蛋白及其应用 | |
| US20170355956A1 (en) | Compositions for increasing survival of motor neuron protein (smn) levels in target cells and methods of use thereof for the treatment of spinal muscular atrophy | |
| Liu et al. | Allele-specific gene-editing approach for vision loss restoration in RHO-associated retinitis pigmentosa | |
| US20230313235A1 (en) | Compositions for use in treating autosomal dominant best1-related retinopathies | |
| WO2020187272A1 (zh) | 一种用于基因治疗的融合蛋白及其应用 | |
| LLADO SANTAEULARIA | THERAPEUTIC GENOME EDITING IN RETINA AND LIVER | |
| WO2021163550A1 (en) | Haplotype-based treatment of rp1 associated retinal degenerations | |
| WO2024069144A1 (en) | Rna editing vector | |
| WO2023212273A1 (en) | Treatments for age-related macular degeneration | |
| CA3216825A1 (en) | Promoters for viral-based gene therapy |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination |