
Scientific name Cephalochordata | Order Amphioxiformes Higher classification Chordate | |
 | ||
Similar Cephalochordate, Branchiostoma, Chordate, Tunicate, Lamprey | ||
The lancelets — also known as amphioxi (singular, amphioxus) — comprise some 32 species of fish-like marine chordates in the order Amphioxiformes, with a global distribution in shallow temperate (as far north as Scotland) and tropical seas, usually found half-buried in sand. They are the modern representatives of the subphylum Cephalochordata, formerly thought to be the sister group of the craniates. In Asia, they are harvested commercially as food for humans and domesticated animals. They are an important object of study in zoology as they provide indications about the evolutionary origins of the vertebrates. Lancelets serve as an intriguing comparison point for tracing how vertebrates have evolved and adapted. Although lancelets split from vertebrates more than 520 million years ago, their genomes hold clues about evolution, particularly how vertebrates have employed old genes for new functions. They are regarded as similar to the archetypal vertebrate form.
Contents
- Anatomy
- Nervous system
- Visual system
- Joseph cells and Hesse organs
- Frontal eye
- Digestive system
- Other systems
- Model organism
- Taxonomy
- References
The first representative organism of the group to be described was Branchiostoma lanceolatum. It was described by Peter Simon Pallas in 1774 as molluscan slugs in the genus Limax. It was not until 1834 that Gabriel Costa brought the phylogenetic position of the group closer to the agnathan vertebrates (hagfish and lampreys), including it in the new genus Branchiostoma (from the Greek, branchio = "gills", stoma = "mouth").
In 1836, Yarrel renamed the genus as Amphioxus (from the Greek: "pointed on both sides"), now considered an obsolete synonym of the genus Branchiostoma. Today, the term "amphioxus" is still used as a common name for the Amphioxiformes, along with "lancelet", especially in the English language. A non-technical review of all aspects of lancelet biology is: Stokes, M. D. and Holland, N. D. 1998. American Scientist 86: 552-560. The genome of the Florida lancelet (Branchiostoma floridae) has been sequenced.

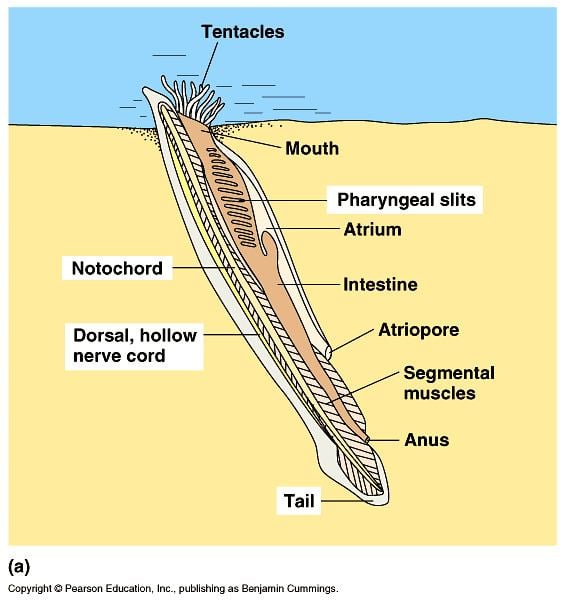
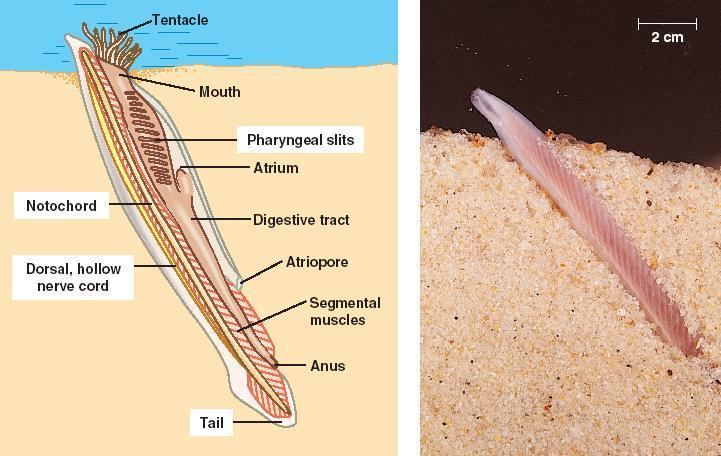
Anatomy

Lancelets are typically 5 centimetres (2.0 in) long, or 7 centimetres (2.8 in) at the longest. They have a translucent, somewhat fish-like body, but without any paired fins or other limbs. A relatively poorly developed tail fin is present, so they are not especially good swimmers. While they do possess some cartilage-like material stiffening the gill slits, mouth, and tail, they have no true skeleton.
Nervous system
In common with vertebrates, lancelets have a hollow nerve cord running along the back, pharyngeal slits and a tail that runs past the anus. Also like vertebrates, the muscles are arranged in blocks called myomeres.

Unlike vertebrates, the dorsal nerve cord is not protected by bone but by a simpler notochord made up of a cylinder of cells that are closely packed to form a toughened rod. The lancelet notochord, unlike the vertebrate spine, extends into the head. This gives the subphylum its name (cephalo- meaning 'relating to the head'). The nerve cord is only slightly larger in the head region than in the rest of the body, so that lancelets do not appear to possess a true brain. However, developmental gene expression and transmission electron microscopy indicate the presence of a diencephalic forebrain, a possible midbrain, and a hindbrain.
Visual system
Lancelets have two known kinds of light-sensing structures, Joseph cells, and Hesse organs, as well as a "frontal eye" and lamellar body. The frontal eye is widely thought to be light-sensitive, although this has not been confirmed by electrophysiological measurement. The lamellar body, a possible homologue of the pineal body, was once thought to be light-sensitive, although this has been called into question. All of these organs and structures are located in the neural tube, with the frontal eye at the front, followed by the lamellar body, the Joseph cells, and the Hesse organs.
Joseph cells and Hesse organs
Joseph cells are bare photoreceptors surrounded by a band of microvilli. These cells bear the opsin melanopsin. The Hesse organs (also known as dorsal ocelli) consist of a photoreceptor cell surrounded by a band of microvill and bearing melanopsin, but half enveloped by a cup-shaped pigment cell. The peak sensitivy of both cells is ~470 nm (blue).
Both the Joseph cells and Hesse organs are in the neural tube, the Joseph cells forming a dorsal column, the Hesse organs in the ventral part along the length of the tube. The Joseph cells extend from the caudal end of the anterior vesicle (or cerebral vesicle) to the boundary between myomeres 3 and 4, where the Hesse organs begin and continue nearly to the tail.
Frontal eye
The frontal eye consists of a pigment cup, a group of putative photoreceptor cells (termed Row 1), three rows of neurons (Rows 2–4), and glial cells. The frontal eye, which expresses the PAX6 gene, has been proposed as the homolog of vertebrate paired eyes, the pigment cup as the homolog of the RPE (retinal pigment epithelium), the putative photoreceptors as homologs of vertebrate rods and cones, and Row 2 neurons as homologs of the retinal ganglion cells.
The pigment cup is oriented concave dorsally. Its cells contain the pigment melanin.
The putative photoreceptor cells, Row 1, are arranged in two diagonal rows, one on either side of the pigment cup, symmetrically positioned with respect to the ventral midline. The cells are flask-shaped, with long, slender Ciliary processes (one cilium per cell). The main bodies of the cells lie outside of the pigment cup, while the cilia extend into the pigment cup before turning and exiting. The cells bear the opsin c-opsin 1, except for a few which carry c-opsin 3.
The Row 2 cells are serotonergic neurons in direct contact with Row 1 cells. Row 3 and 4 cells are also neurons. Cells of all four rows have axons that project into the left and right ventrolateral nerves. For Row 2 neurons, axon projections have been traced to the tegmental neuropil. The tegmental neuropil has been compared with locomotor control regions of the vertebrate hypothalamus, where paracrine release modulates locomotor patterns such as feeding and swimming.
Digestive system
Lancelets also have oral cirri, thin tentacle-like strands that hang in front of the mouth and act as sensory devices and as a filter for the water passing into the body. Water passes from the mouth into the large pharynx, which is lined by numerous gill-slits. The ventral surface of the pharynx contains a groove, called the endostyle, which, connected to a structure known as Hatschek's pit, produces a film of mucus. Ciliary action pushes the mucus in a film over the surface of the gill slits, trapping suspended food particles as it does so. The mucus is collected in a second, dorsal, groove, and passed back to the rest of the digestive tract. Having passed through the gill slits, the water enters an atrium surrounding the pharynx, then exits the body via the atriopore.
Both adults and larvae exhibit a "cough" reflex to clear the mouth or throat of debris or items too large to swallow. In larvae the action is mediated by the pharyngial muscles while in the adult animal it is accomplished by atrial contraction.
The remainder of the digestive system consists of a simple tube running from the pharynx to the anus. The hepatic caecum, a single blind-ending caecum, branches off from the underside of the gut, with a lining able to phagocytize the food particles, a feature not found in vertebrates. Although it performs many functions of a liver, it is not considered a true liver but a homolog of the vertebrate liver.
Other systems
Lancelets have no respiratory system, breathing solely through their skin, which consists of a simple epithelium. Despite the name, little if any respiration occurs in the gill slits, which are solely devoted to feeding. The circulatory system does resemble that of primitive fish in its general layout, but is much simpler, and does not include a heart. There are no blood cells, and no haemoglobin.
The excretory system consists of segmented "kidneys" containing protonephridia instead of nephrons, and quite unlike those of vertebrates. Also unlike vertebrates, there are numerous, segmented gonads.
Model organism
Lancelets became famous in the 1860s when Ernst Haeckel ("the German Darwin") began promoting them as a model for the ancestor of all vertebrates. By 1900 lancelets had become a model organism. By the mid-20th century they had fallen out of favor for a variety of reasons, including a decline of comparative anatomy and embryology, and due to the belief that lancelets were more derived than they appeared, e.g., the profound asymmetry in the larval stage.
With the advent of molecular genetics lancelets are once again regarded as a model of vertebrate ancestors, and are in process of being revived as a model organism.
Taxonomy
The Cephalochordata is traditionally seen as a sister subphylum to the vertebrates, with which it is grouped together into a clade (sometimes called Notochordata) which in turn is the sister group to the simpler still Urochordata. Newer research suggests this is not the case. The Cephalochordata is the most basal subphylum of the chordates, while the sister group of the vertebrates are the tunicates, previously known as the urochordates. However, older molecular studies placed cephalochordates nearer to vertebrates, and "[m]ost authors regard amphioxus as the closest relative of the Vertebrata on the basis of 10–15 [morphological] features that are not seen in tunicates".
The following are the species recognised by ITIS. Other sources, for instance Tudge, show that there might be up to thirty species.