
Abstract
Odontasteridae (Asteroidea: Echinodermata) (Verrill in Am J Sci, 1899) is placed within Valvatida, a derived assemblage of sea stars. Odontasterids are found in the Southern, Atlantic, and Pacific Oceans and are concentrated in high southern latitudes. To date, the phylogenetic and evolutionary history of Odontasteridae as a whole has not been rigorously examined. We conducted molecular and morphological phylogenetic analyses of Odontasteridae to assess the interrelationships among and within recognized genera. We used mitochondrial 16S and cytochrome c oxidase subunit I molecular markers and 29 external morphological characters in an attempt to reconstruct the evolutionary history of the group. Generally, our results indicate that traditionally used external skeletal characters are not representative of phylogenetic history of Odontasteridae. We can conclude that species present in high latitudes in the Southern Hemisphere (i.e., Southern Ocean) are the most derived taxa. Additionally, mtDNA data suggest unrecognized lineages of odontasterids are present in high southern latitudes. A new species Odontaster cynthiae sp. nov. is described from the Galapagos Islands.








Similar content being viewed by others

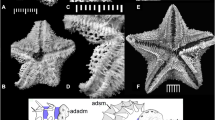

References
Arntz WE, Brey T, Gallardo VA (1994) Antarctic zoobenthos. Oceanogr Mar Biol 32:241–304
Bell FJ (1894) On the echinoderms collected during the voyage of H.M.S. Penguin and by H.M.S. Egeria, when surveying Macclesfield Bank. Proc Zool Soc Lond 1894:392–413
Blake DB (1987) Classification and phylogeny of post-Palaeozoic sea stars (Asteroidea: Echinodermata). J Nat Hist 21:481–528
Brandt A (1992) Origin of Antarctic Isopoda (Crustacea, Malacostraca). Mar Biol 113:415–423
Brandt A, Gooday AJ, Brandao SN, Brix S, Brokeland W, Cedhagen T, Choudhury M, Cornelius N, Danis B, DeMesel I et al (2007) First insights into the biodiversity and biogeography of the Southern Ocean deep sea. Nature 447:307–311
Briggs JC (2003) Marine centers of origin as evolutionary engines. J Biogeogr 20:1–18
Clark AM (1962) Asteroidea. Rep BANZ Antarct Res Exped 1929–1931 9:68–70
Clark AM, Downey ME (1992) Starfishes of the Atlantic. Chapman and Hall, London
Clark HES, McKnight DG (2001) The marine fauna of New Zealand: echinodermata: Asteroidea (Sea-Stars) order Valvatida. NIWA Biodivers Mem 117:142–194
Clarke A, Johnston NM (2003) Antarctic marine benthic diversity. Oceanogr Mar Biol 41:47–114
Crame JA (1993) Latitudinal range fluctuations in the marine realm through geological time. Trends Ecol Evol 8:162–166
Dayton PK, Robilliar GA, Paine RT, Dayton LB (1974) Biological accommodation in the benthic community at McMurdo Sound, Antarctic. Ecol Monogr 44:105–128
Dell RK (1972) Antarctic benthos. Adv Mar Biol 10:1–216
Fell HB (1953) The origin and migrations of Australasian echinoderm faunas since the Mesozoic. Trans Proc R Soc NZ 81:245–255
Fisher WK (1908) Some necessary changes in the generic names of starfishes. Zool Anz 33:356–359
Fisher WK (1911) Asteroidea of the North Pacific and adjacent waters. 1. Phanerozonia and Spinulosida. Bull US Nat Mus 76:xiii + 420 pp. 122 pls
Fisher WK (1940) Asteroidea. Discov Rep 20:69–306
Gebruk AV (1994) Two main stages in the evolution of the deep-sea fauna of elasipodid holothurians. In: David B, Guille B, Feral J-P, Roux M (eds) Echinoderms through time. Balkema, Rotterdam, pp 507–514
Hall TA (1999) Bioedit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acid Res 41:95–98
Hunter RL, Halanych KM (2008) Evaluating connectivity of the brooding brittle star Astrotoma agassizii across the Drake Passage in the Southern Ocean. J Hered 99:137–148
Jangoux DB, Massin C (1986) Catalogue commete des types d’Echinodermes actuels conserves dans les collections nationales belges. Bulletin de l’Institut Royal des Sciences Naturelles de Belguique, Biologie 55:83–97
Janosik AM, Halanych KM (2010) Unrecognized Antarctic biodiversity: a case study of the genus Odontaster (Odontasteridae; Asteroidea). Integr Comp Biol 50:981–992
Janosik AM, Mahon AR, Halanych KM (2011) Evolutionary history of Southern Ocean sea star species separated by the drake passage Odontasteridae; Asteroidea). Polar Biol 34:575–586
Lambert P (2000) The sea stars of British Columbia. British Columbia Provincial Museum Handbook, Province of British Columbia 39
Ludwig H (1903) Seesterne. Results du Voyage du S.Y. Belgica 1897–1899. Rapports Scientifiques Zoologie (1901–1913). Buschmann: Anvers. pp 1–72
Maddison WP, Maddison DR (2010) Mesquite: modular system for evolutionary analysis. Version 2.73 http://mesquiteproject.org
Mah C, Foltz D (2011) Molecular phylogeny of the Valvatacea (Asteroidea: Echinodermata). Zool J Linn Soc Lond 161:769–788
McClintock JB, Pearse JS, Bosch I (1988) Population structure and energetics of the shallow-water Antarctic sea star Odontaster validus in contrasting habitats. Mar Biol 99:235–246
McKnight DG (2006) The marine fauna of New Zealand, Echinodermata: Asteroidea (Sea-stars). 3. Orders Velatida, Spinulosida, Forcipulatida, Brisingida with addenda to Paxillosida, Valvatida. NIWA Biodiver Mem 120:1–187
Nylander JAA (2004) MRMODELTEST. Version 2. Distributed by the author. Evolutionary Biology Centre, Uppsala University, Sweden
Page RDM (2001) NEXUS data editor: http://taxonomy.zoology.gla.ac.uk/rod/rod.html
Palumbi SR, Martin A, Romano S, McMillan WO, Stice L, Grabowski G (1991) The simple fool’s guide to PCR. Department of Zoology, University of Hawaii, Honolulu, Hawaii
Pawlowski J, Bowser SS, Gooday A (2007) A note on the genetic similarity between shallow- and deep-water Epistominella vitrea (Foraminifera) in the Antarctic. Deep Sea Res Part II 54:1720–1727
Pawson DL, Ahearn CA (2000) Bathyal echinoderms of the Galapagos Islands Echinoderms. In: Barker M (ed) Echinoderms 2000. A.A Balkema, Rotterdam, pp 41–46
Perrier E (1875) Revisiond e la Collection de Stellerides du Museum d’Historie Naturelle de Paris. Reinwald, Paris
Perrier E (1891) Echinodermes de la mission scientifique du Cap Horn. Stelleroides. Mission Scientifique du Cap-Horn. Zoologie 6:1–168
Perrier E (1894) Stellerides. Expeditions scientifiques du Travailleur et du Talisman pendant les annees 1880, 1881,1882, 1883, vol. 3. G. Masson, Paris, pp 1–431
Ronquist F, Huelsenbeck JP (2003) MRBAYES 3: bayesian phylogenetic inference under mixed models. Bioinformatics 19:1572–1574
Shaw PW, Arkhipkin AI, Al-Khairulla H (2004) Genetic structuring of Patagonian toothfish populations in the Southwest Atlantic Ocean: the effect of the Antarctic Polar Front and deep-water troughs as barriers to genetic exchange. Mol Ecol 13:3293–3303
Sladen WP (1889) Report on the Asteroidea. Report on the scientific results of the voyage of H.M.S. Challenger during the years 1873–1876. Zoology 30, xlii + 893 pp, 118 pis
Strugnell JM, Rogers AD, Prodohl PA, Collins MA, Allcock AL (2008) The thermohaline expressway: the Southern Ocean as a centre of origin for deep-sea octopuses. Cladistics 24:853–860
Thornhill DJ, Mahon AR, Norenburg JL, Halanych KM (2008) Open-ocean barriers to dispersal: a test case with the Antarctic Polar Front and the ribbon worm Parborlasia corrugatus (Nemertea:Lineidae). Mol Ecol 17:5104–5117
Verrill AE (1880) Notice of the remarkable marine fauna occupying the outer banks off the southern coast of New England. Am J Sci 20:390–403
Verrill AE (1899) Revision of certain genera and species of starfishes with descriptions of new forms. Trans Conn Acad 10:145–234
Verrill AE (1909) Description of new genera and species of starfishes from the North Pacific coast of America. Am J Sci 28:59–70
Acknowledgments
Dr. Christopher Mah from the Smithsonian Institution National Museum of Natural History kindly helped AMJ while visiting to study the Odontasteridae collection. Special thanks go to Cynthia Ahearn from the USNM for lending and coordinating specimens used in this study. Loans were also provided by the California Academy of Sciences and the National Institute of Water and Atmospheric Research for loan of specimens. We are grateful for the help and logistical support by the crew and participants of the 2004 and 2006 Antarctic cruise aboard the R/V Laurence M. Gould. This project was supported by National Science Foundation grants (OPP-9910164, OPP-0338087, and OPP-0338218, ANT-1043745) to K.M Halanych and R.S. Scheltema. This is AU Marine Biology Program contribution #95 and Molette Lab contribution #9.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Uthicke.
Appendices
Appendix 1
See Table 3.
Appendix 2
See Fig. 9.

Bayesian inference topology for 16S sequence data. Analysis details are provided in text. Number next to node indicates Bayesian posterior probabilities
Appendix 3
See Fig. 10.

Bayesian inference topology for COI sequence data. Analysis details are provided in text. Number next to node indicates Bayesian posterior probabilities
Appendix 4
See Fig. 11.

All morphological characters mapped onto Bayesian inference tree. Analysis details are provided in text. Bars indicate where a change has occurred, and squares indicate character reversals or loss of a character. Asterisks indicate character states that were equally parsimonious at a node
Appendix 5. Morphological Character analysis of Odontasteridae
- 1.:
-
Recurved spine on oral plates: 0 = absent, 1 = one spine, 2 = two spines
- :
-
Whether one or two recurved spines were present in the last common ancestor of Odontasteridae cannot be determined here. Hoplaster lacks a recurved spine, which is a lost character. Acodontaster, Eurygonias, Odontaster have one recurved, glassy spine per oral plate; Diplodontias has two recurved, glassy spines per oral plate, while the recurved, glassy spines are missing in Diabocilla and Hoplaster. Number of changes on tree (changes) = 2; Consistency Index (CI) = 1.0
- 2.:
-
Abactinal plates: 0 = tabulate, 1 = paxillate, 2 = highly paxillate
- :
-
Tabulate abactinal plates are inferred as an ancestral character, with a change occurring in Eurygonias hyalacanthus and Odontaster species. A change to highly paxillate occurs in O. validus. Acodontaster has tabulate abactinal plates. Diplodontias has sub-tabulate abactinal plates. Eurygonias has abactinal plates that are paxillar and club-shaped. Hoplaster has abactinal plates that are tabulate. Overall, Odontaster has somewhat paxillate abactinal plates, although some species tend to have a more tabulate look. Changes = 3; CI = 0.67
- 3.:
-
Abactinal spines per plate: 0 = (5–10), 1 = (11–15), 2 = (16–20), 3 = (21–25), 4 = (26–30), 5 = (30 and above)
- :
-
Diplodontias and Eurygonias have the most spines, while a change occurs in Diplodontias singularis from a category 5 to 4. Changes = 10; CI = 0.45
- 4.:
-
Abactinal spine: 0 = smooth spines, 1 = rough spines
- :
-
Whether the last common ancestor of Diplodontias had either smooth or rough abactinal spines cannot be determined. Acodontaster, Eurygonias, Hoplaster, and Odontaster share a common ancestor that likely had rough abactinal spines. Smooth or rough texture of spines on abactinal spines is not genus specific and varies greatly from species to species. Changes = 5; CI = 0.167
- 5.:
-
Glassy granules on abactinal plate: 0 = absent, 1 = present
- :
-
Glassy granules are a derived character present only in Diplodontias miliaris, Diplodontias dilatatus, Hoplaster spinosus, O. aucklandensis, O. australis, O. cynthiae. Changes = 2; CI = 0.5
- 6.:
-
Abactinal spine shape: 0 = granular, 1 = short, stout, 2 = short, slender, 3 = clavate, 4 = long, slender, 5 = long, slender prominent spine in middle
- :
-
Granular abactinal spines are inferred as the most ancestral state. A change occurs in Eurygonias hyalacanthus, which has short, slender spines. A change also occurs at the base of the Odontaster clade to clavate spines. A reversal to granular spines occurs in O. penicillatus. A change occurs in O. validus and O. robustus to long, slender spines. Acodontaster and Diplodontias species all have granular abactinal spines. Diabocilla and Hoplaster species have clavate or club-shaped spines, while Eurygonias has short, slender spines. Odontaster species tend to have a variation of spine shapes. Changes = 7; CI = 0.71
- 7.:
-
Papulae on abactinal surface: 0 = restricted to arms and central disk, not found interradially, 1 = absent from disks center and interradial area, 2 = covering entire abactinal surface
- :
-
Papulae covering the entire abactinal surface are inferred as an ancestral character in Eurygonias hyalacanthus and Diplodontias species. Odontaster and Acodontaster species have papulae restricted to the arms and central disk. A reversal has occurred in Hoplaster kupe, which has papulae covering the entire abactinal surface. Changes = 3; CI = 0.67
- 8.:
-
Marginal plate border: 0 = plates form even border with abactinals, 1 = plates form slightly raised border with abactinals, 2 = plates form distinct border with abactinals
- :
-
The inferred ancestral character is marginal plates that form a slightly raised border with the abactinals, while a change to forming an even border occurs at the base of the Acodontaster and Odontaster clade. A character reversal to a slightly raised border occurs in O. crassus, O. meridionalis, O. pearsei, and O. roseus. Acodontaster hodgsoni has marginal plates that form a distinct border with the abactinal plates. Changes = 4; CI = 0.50
- 9.:
-
Grooves between marginal plates: 0 = grooves not distinct, 1 = deep grooves between plates
- :
-
Diplodontias and Eurygonias hyalacanthus have deep grooves between plates. It is equally parsimonious that either deep grooves or grooves not distinct between marginal plates were present in the last common ancestor of Acodontaster and Odontaster. Odontaster species have deep grooves between marginal plates, while Acodontaster do not. Changes = 4; CI = 0.25
- 10.:
-
Marginal plate shape: 0 = wider than long, 1 = square (block-like), 2 = wedge-shaped, 3 = rectangular round
- :
-
Square-shaped marginal plates are inferred as ancestral, with change occurring in Acodontaster conspicuus and Eurygonias hyalacanthus to rectangular round-shaped plates. Diplodontias miliaris, O. crassus, O. benhami, O. meridionalis, O. pearsei, O. penicillatus, and O. roseus have plates that are wider than long. A reversal occurs in O. validus. Changes = 7; CI = 0.43
- 11.:
-
Superomarginal plates: 0 = densely covered in spines of same length, 1 = densely covered in spines getting longer toward the edge of the plate
- :
-
With the exception of Hoplaster kupe, O. crassus, and O. hispidus, all members of Odontasteridae have superomarginal plates densely covered in spines of the same length. Acodontaster, Diplodontias, Eurygonias species have superomarginal plates densely covered in spines of the same shape. Diabocilla, Hoplaster, and Odontaster species vary between densely covered in spines of the same shape and densely covered in spines getting longer toward the edge of the plate. Changes = 3; CI = 0.33
- 12.:
-
Spines on superomarginal plates: 0 = granules, 1 = spinelets
- :
-
Granules for spines on the superomarginal plates are inferred as the ancestral character state. A change occurs at the base of the Odontaster clade. Reversals occur in Odontaster crassus, O. mediterraneus, and O. penicillatus. Acodontaster, Diplodontias, and Eurygonias have granules for spines on the superomarginal plates. Diabocilla and Hoplaster have spinelets on the superomarginal plates, while Odontaster is varied, with some species having granules and some with spinelets. Changes = 5; CI = 0.20
- 13.:
-
Inferomarginal plates: 0 = densely covered in spines of the same length, 1 = densely covered in spines getting longer toward the edge of the plate
- :
-
With the exception of Hoplaster kupe, O. crassus, and O. hispidus, and O. penicillatus, all members of Odontasteridae have inferomarginal plates densely covered in spines of the same length. Acodontaster, Diplodontias, and Eurygonias have granules for spines on the inferomarginal plates. Diabocilla has spinelets on the superomarginal plates, while Hoplaster and Odontaster are varied, with some species having granules and some with spinelets. Changes = 4; CI = 0.25
- 14.:
-
Spines on inferomarginal plates: 0 = same as superomarginal plates, 1 = longer than superomarginal plates, 2 = granules, with one long, prominent spine toward the outside edge of the plate
- :
-
Diplodontias, Eurygonias hyalacanthus, and Acodontaster have spines on inferomarginal plates that are the same as the superomarginal plates. A change to longer spines occurs in O. meridionalis, O. pearsei, O. penicillatus, and O. roseus. Acodontaster marginatus has one long, prominent spine toward the outside edge of the plate. Most Acodontasterspecies have spines that are the same length as the spines on the superomarginal plates. Changes = 4; CI = 0.50
- 15.:
-
Glassy granules on superomarginal plates: 0 = absent, 1 = present, 2 = present only on plates toward the arm tips
- :
-
The presence of glassy granules on the superomarginal plates is inferred as an ancestral character found in Diplodontias and Eurygonias hyalacanthus. Absence of glassy granules occurs at the base of Acodontaster and Odontaster, with changes in O. benhami, O. crassus and O. mediterraneus. Odontaster hispidus has glassy granules present only on the plates toward the arm tips. Changes = 5; CI = 0.40
- 16.:
-
Glassy granules on inferomarginal plates: 0 = absent, 1 = present, 2 = present only on plates toward the arm tips
- :
-
The ancestral state of the glassy granules on the inferomarginal plates cannot be determined. Glassy granules are present in hyalacanthus, Diplodontias dilatatus, D. miliaris, Odontaster benhami, O. crassus, and O. mediterraneus. Odontaster hispidus has glassy granules, but only on the plates toward the arm tips. Changes = 6; CI = 0.33
- 17.:
-
Number of chevrons on actinal surface: 0 = 3, 1 = 4, 2 = 5, 3 = 6, 4 = 7
- :
-
The number of chevrons present on the actinal surface is quite variable across and within all genera (character not mapped on Fig. 8)
- 18.:
-
Spines per plate on actinal surface: 0 = (4–9), 1 = (10–15), 2 = (16–20), 3 = (8–10 with one prominent longer spine)
- :
-
The number of spines per plate on the actinal surface is varied across genera and species within genera. Sixteen to twenty spines per plate on the actinal surface was inferred as the ancestral state. Changes occur at the base of the Diplodontias miliaris and D. dilatatus clade and several times throughout Acodontaster and Odontaster clades. Changes = 10; CI = 0.10
- 19.:
-
Glassy granules on actinal surface: 0 = absent, 1 = present
- :
-
Acodontaster, Diabocilla, Diplodontias, Eurygonias, and Hoplaster are all lacking in glassy granules on the actinal surface. Only two members of Odontaster have glassy granules present on the actinal surface. Changes = 1; CI = 1.0
- 20.:
-
Number of furrow spines: 0 = (1–2), 1 = (2–3), 2 = (3–4), 3 = (4–5)
- :
-
The number of furrow spines is varied across and within taxa. The majority of taxa have 2–3 furrow spines. Changes = 9; CI = 0.33
- 21.:
-
Furrow spine shape: 0 = smooth, cylindrical, 1 = smooth, pointy, 2 = rough, cylindrical, 3 = rough, pointy
- :
-
Diplodontias species and E. hyalacanthus have smooth, cylindrical furrow spines. A change to smooth, pointy spines occurs at the base of the Acodontaster and Odontaster clade. Further changes occur within the Odontaster and Acodontaster clades. Character reversal to smooth, cylindrical furrow spines occurs in O. benhami, O. hispidus, and O. penicillatus. Changes = 14; CI = 0.21
- 22.:
-
Adambulacral plate shape: 0 = rectangle, 1 = square
- :
-
The ancestral character state is inferred as rectangular adambulacral plates. Character state changes occur in lineages leading to Acodontaster hodgsoni, A. marginatus, Hoplaster kupe, Odontaster penicillatus, and O. robustus. Changes = 5; CI = 0.20
- 23.:
-
Pedicellariae: 0 = absent, 1 = present
- :
-
All Acodontaster, except A. capitatus, have pedicellariae. Diabocilla, Hoplaster, and Diplodontias (except D. singularis) are lacking pedicellariae. Eurygonias and several members of Odontaster have pedicellariae. Type and appearance of pedicellariae are variable. Changes = 3; CI = 0.33
- 24.:
-
Body shape outline: 0 = pentagonal, 1 = sub-pentagonal, 2 = pentagonal-stellate, 3 = sub-stellate, 4 = stellate
- :
-
Determination of the body shape outline in the last common ancestor of Odontasteridae is not possible. Changes = 12; CI = 0.33
- 25.:
-
Interradial arc: 0 = rounded, 1 = sub-linear, 2 = linear
- :
-
A rounded interradial arc is the inferred ancestral character state. All Acodontaster and the majority of Diplodontias species have rounded interradial arcs. Diabocilla and Hoplaster have sub-linear interradial arcs, while Odontaster have either rounded or sub-linear interradial arcs. Eurygonias has a completely linear interradial arc. Changes = 6; CI = 0.33
- 26.:
-
Arm length: 0 = short, 1 = medium, 2 = elongate
- :
-
Acodontaster have elongate arms. Diabocilla has medium length arms, and Diplodontias, Hoplaster, and Odontaster have a variety of arm length, while Eurygonias have very short arms. Changes = 12; CI = 0.17
- 27.:
-
Number of apical spines per oral plate: 0 = 2, 1 = 3, 2 = 4, 3 = 5, 4 = 6
- :
-
The majority of taxa have four apical spines per oral plate. Changes occur at terminal nodes within Acodontaster and Diplodontias. Several changes occur throughout the Odontaster clade (character not mapped)
- 28.:
-
Number of suboral spines per oral plate: 0 = 2, 1 = 3, 2 = 4, 3 = 5, 4 = 6+
- :
-
The character state of last common ancestor of Odontasteridae is equivocal with either two or three suboral spines per oral plate. The number of suboral spines per oral plate is variable across and within genera (character not mapped).
- 29.:
-
Number of marginal spines per oral plate: 0 = (2–3), 1 = (3–4), 2 = (4–5), 3 = (5–6), 4 = (6–7)
The number of marginal spines per oral plate is variable across and within genera (character not mapped)
Rights and permissions
About this article
Cite this article
Janosik, A.M., Halanych, K.M. Seeing stars: a molecular and morphological investigation into the evolutionary history of Odontasteridae (Asteroidea) with description of a new species from the Galapagos Islands. Mar Biol 160, 821–841 (2013). https://doi.org/10.1007/s00227-012-2136-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-012-2136-x