
Abstract
Human-induced water level fluctuations (WLFs) are among the major pressures threatening lake ecosystems. Their effect on meiobenthic species of the littoral zone has been poorly investigated. In this study, we aimed at assessing the effects of human-induced WLFs on the composition and functionality of the benthic copepod assemblages of the littoral zone of Lake Maggiore, Italy and Switzerland. From 1942 to present, the water level of Lake Maggiore has been regulated through the Miorina Dam. We monitored copepod assemblages during different water levels defined within the period of regulation by the dam, using taxonomy- and trait-based metrics. We observed variation in the overall abundance and biomass of copepods, as well as in the number of individuals belonging to some trait classes such as ovigerous females, opportunists, omnivores and deposit feeders. None of the investigated trait class was completely lost. The results of our study suggest that ecosystem services supplied by the littoral zone of Lake Maggiore are likely altered by human-induced WLFs. Our findings provide a first picture of WLF impacts on benthic copepod assemblages, which can be useful for future research expanding on other functional traits.
Similar content being viewed by others

Introduction
Freshwater lakes are among the most important water reserves for human beings and provide provisioning (e.g. fisheries, transportation, irrigation, thermoelectric and hydroelectric water uses), regulating (e.g. climate regulation, flood control, nutrient cycling, pollution dilution), supporting and cultural (e.g. recreational, touristic, spiritual and cultural benefits) ecosystem services (e.g. Allan et al., 2015; Zhong et al., 2019; Sterner et al., 2020). Climate change, dam building and other anthropogenic pressures have led to lake ecosystem dysfunctions, irreversible species loss and severe changes in community composition (e.g. Hallstan et al., 2013; Cantonati et al., 2020, 2021; Mammides, 2020) with inevitable effects on human well-being (Millennium Ecosystem Assessment, 2005).
The littoral zone of lakes is characterised by heterogeneous ecological conditions. Seasonal variations in temperature, light and current cause relevant changes in littoral assemblages (Caramujo et al., 2008; Callieri et al., 2014; Kreuzinger-Janik et al., 2015; Carmignani & Roy, 2017). Human-altered hydrological regimes are a strong anthropogenic component affecting the composition and functionality of the littoral communities (e.g. Brauns et al., 2011). Human-induced water level fluctuations (WLFs; e.g. Zohary & Ostrovsky, 2011; Evtimova & Donohue, 2016) are one of the determinants of benthic assemblage changes in the littoral zone of lakes (e.g. Leira & Cantonati, 2008; Wantzen et al., 2008; Cantonati et al., 2020, 2021; Mammides, 2020). This is because human-induced drawdowns occur with a different magnitude and frequency relative to natural WLFs (e.g. Leira & Cantonati, 2008). Mesocosm studies have demonstrated that alterations of natural WLFs can reduce both the density and taxonomic distinctness of littoral benthic metazoan assemblages (Evtimova & Donohue, 2014). Rapid WLFs induce horizontal shoreline displacements and hydrodynamic stress on organisms living in the shore region and on the sediment surface (Hofmann et al., 2008). Evtimova & Donohue (2016) concluded that altered WLFs are an increasingly pervasive pressure on lake ecosystems worldwide. However, the effects of WLFs, whether natural or human-induced, on the functionality of littoral benthic communities of freshwater lakes remain understudied (e.g. Brancelj et al., 2000; Heino, 2008; Leira & Cantonati, 2008; Evtimova & Donohue, 2016). Many studies to date have concerned macroinvertebrates (e.g. Scheifhacken et al., 2007; Leira & Cantonati, 2008; Poikane et al., 2020; Špoljar et al., 2021) and showed that human-induced WLFs have, in many cases, more substantial impacts than the natural ones (e.g. Vermaat et al., 2022). For instance, Brauns et al. (2008) observed that macroinvertebrate species richness was not significantly affected by natural WLFs in East-German lowland lakes. Similarly, the functionality of the assemblages, namely the abundances of filterers, piercers, predators, scrapers, shredders and xylophagous, varied mainly under the different types of substrates rather than following WLFs (Brauns et al., 2008). On the contrary, Baumgärtner et al. (2008) found significant correlations between the duration of flooding from human-induced WLFs and macroinvertebrate assemblages of the littoral zone of Lake Constance.
The effect of human-induced WLFs on meiobenthic animals has been much less investigated, despite their potential as bioindicators highlighted by previous studies (e.g. Caramujo et al., 2008; Kreuzinger-Janik et al., 2015; Majdi et al., 2020; Traunspurger et al., 2020). Copepods are one of the most abundant groups of meiofaunal organisms in freshwater benthic habitats of streams, springs and lakes (e.g. Dole-Olivier et al., 2000; Galassi, 2001), as well as in groundwaters (e.g. Galassi et al., 2009a, b; Stoch & Galassi 2010; Iannella et al., 2020; Di Cicco et al., 2021). Copepods occupy numerous trophic niches in the freshwater food web serving as prey for larger predators (e.g. Dole-Olivier et al., 2000; Galassi, 2001) and feeding on bacteria, algae, detritus, and even preying on other animals (Reid & Williamson, 2010). Lake benthic copepods remain understudied compared to nematodes, especially when considering functional trait variation potentially induced by WLFs (Majdi et al., 2020; Cifoni et al., 2021). Functional traits are defined as morpho–physio-phenological features that impact the fitness of individual species via their effects on growth, reproduction, locomotion, behaviour, feeding habits and survival (Violle et al., 2007). Functional traits have been relatively well defined in marine planktonic copepods to the point of depicting global biogeography of key traits for pelagic species (Brun et al., 2016). Similarly, trait studies concerning freshwaters have been mainly focused on planktonic copepods (e.g. Colina et al., 2016; Cifoni et al., 2021). The insufficient research carried out on benthic copepods so far is mainly due to challenging taxonomic identification in field studies and difficulties in experimental manipulation through in vivo experiments (Fonseca et al., 2018; Majdi et al., 2020). However, it is worth overcoming these issues in order to explore the role of copepods as bioindicators in benthic freshwater environments, given that the group is very abundant in this type of environment, exhibiting a wide range of trait guilds potentially responding to different natural and anthropogenic stressors (e.g. Di Lorenzo et al., 2021a, b and references therein).
In this study, we aimed at assessing the effects of late spring–summer human-induced WLFs on the composition and functionality of benthic copepod assemblages of the littoral zone of Lake Maggiore, an extensively studied lake and a fundamental water resource for the whole River Po Catchment in northern Italy (Arfè et al., 2019). Since 1943 to present, the water level of Lake Maggiore has been regulated by the Miorina Dam. We used both taxonomy and trait-based metrics of response to late spring–summer human-induced WLFs, as opposed to only taxonomy, to capture key aspects of copepod functional diversity (e.g. Gerino et al., 2003; Descloux et al., 2014; Di Lorenzo et al., 2021a). Since we expected that a set of potential environmental determinants would affect copepod composition and functionality, we also investigated the biotic response to thirteen environmental variables which were measured and analysed. First, we hypothesised that WLFs of Lake Maggiore have a significant effect on the environmental parameters as well as the composition and functionality of the littoral copepod assemblages and that the effects override those induced by sampling stations and site types (Hypothesis H1). Secondly, considering the findings of previous studies on the topic (e.g. Caramujo et al., 2008; Callieri et al., 2014; Kreuzinger-Janik et al., 2015; Martínez et al., 2021), we also hypothesised that variation in environmental parameters, including reactive and total phosphorous, nitrogen compounds and reactive silica, could play a major role in determining variation in composition and functionality of copepod assemblages in Lake Maggiore (Hypothesis H2). The main conclusions of our study provide the base for further research on the effect of human-induced WLFs on benthic copepod assemblages of regulated lakes.
Materials and methods
Study area
Lake Maggiore (area = 213 km2; altitude = 193 m a.s.l.; max depth = 370 m; trophic status = oligo-mesotrophic) is located in the Regio Insubrica in the subalpine area of the west-southern Alps. Lake Maggiore, formed by glaciers and fluvial activities, is the second-largest and deepest lake in Italy. It belongs to the northern Italian lake district including also Como, Garda and Iseo Lakes. All these lakes are part of the Long-Term Ecological Research (LTER) network (Site LTER_EU_IT_008) and, therefore, have been subjected to continuous environmental monitoring since the 1970s. The Lake Maggiore watershed (6,599 km2) is half-shared with Switzerland (Fig. 1). Lake Maggiore has 35 tributaries, with the River Toce (Italy) and the River Ticino Inlet (Switzerland) the main two. River Ticino also acts as the lake outlet (River Ticino Outlet, Italy). The watershed of Lake Maggiore is characterised by relatively high precipitation (yearly average precipitation of about 1,700 mm; sub-littoral alpine rainfall regime, characterised by two maxima in spring and autumn and two minima in winter and summer; Ambrosetti et al., 2006). Since 1942, the regulation of the lake levels through the Miorina Dam was designed to keep the lake level in the range + 1.00 and − 0.50 m, measured at the hydrometer of Sesto Calende, where the dam is located (Boggero et al., 2021). The regulation was designed to satisfy agriculture and industry needs and to control floods and drought events. During the 1960s, the regulation limits were increased for the winter period (from November to March) to + 1.50 m. Finally, in 2014, the authority in charge of water level regulation at the Miorina Dam (Consorzio del Ticino) asked to extend the water level to + 1.50 m during the late spring–summer period (Boggero et al., 2021). Lake levels and precipitation are continuously measured by a wide meteorological and hydrographic network. For our work needs, we considered three hydrometer stations, those nearest to the chosen sampling stations (namely, Locarno representative of the Bolle di Magadino lake-level conditions, Pallanza for the Fondo Toce conditions, and Sesto Calende for the Angera-Sesto Calende conditions). Due to the morphological and geographic position of these three hydrometer stations, the measured lake level may differ and may have a different influence on ecological conditions at the sampling stations. Usually, the lake levels are highest in late spring (mainly, May–July) and gradually decrease, due to water use by stakeholders, low summer precipitations and scarce snow inputs, until reaching a minimum value in the early autumn season (Ciampittiello et al., 2019). Lake Maggiore WLFs are exacerbated by lake water level regulation at the outlet to match downstream water requirements and flood control (Carrara et al., 2009; Callieri et al., 2014).

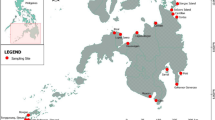
Map of sampling stations located in three protected areas and distributed according to the north–south direction of the lake: Bolle di Magadino (BM; yellow triangles) in the northern part, Fondo Toce (FT; green triangles) in the central part and Angera-Sesto Calende (A; red triangles) in the southern part. Monitoring was conducted at each sampling station, in two sites, of which one was permanently wet (W) and the other subjected to drought (D). The border between Italy and Switzerland is marked by the yellow line; that between the Piedmont and Lombardia regions is indicated in white
Lake Maggiore has been subjected to continuous environmental monitoring since 1978 within the limnological campaigns funded by the International Commission for the Protection of Swiss-Italian Waters (CIPAIS; http://www.cipais.org/). Long-term datasets concerning water chemistry, biology (plankton and fish), hydrology, and climate are available from synoptic studies (e.g. Jackson & Fuereder, 2006; Manca & Bertoni, 2014; Rogora et al., 2015; Fenocchi et al., 2017; Morabito et al., 2018). Since 2019, Lake Maggiore has also been monitored within the framework of the INTERREG Project Parchi Verbano Ticino (PVT, 2019–2023, ID 481668, https://www.parchiverbanoticino.it/). The project, which has been funded by the European Regional Development Fund (ERDF) through the INTERREG Cooperation Programme between Italy and Switzerland (2014–2020), aims at proposing common strategies for shared and sustainable management of the freshwater resources of the lake and the downstream River Ticino. The assessment of impacts of late spring–summer human-induced WLFs on the littoral meiofauna, the object of this work, is one of the planned project activities.
Sampling survey
We monitored the lake during the summer period at three sampling stations along the north–south axis of the lake (Fig. 1): Bolle di Magadino (BM; North), Fondo Toce (FT; Centre), Angera-Sesto Calende (A; South). The three sampling stations are included in an Emerald area (Switzerland; Bolle di Magadino) and two Natura 2000 sites (Italy; Fondo Toce and Angera-Sesto Calende), which are nature reserves ecologically and economically sustainably managed. We monitored two different sandy sites at each station, namely a site permanently flooded throughout the year (W) and a site occasionally subjected to drought during low lake water levels (D). Each site covered a sandy shore of about 25 m, up to 20 m towards open waters and up to a maximum depth of 1.10 m, depending on the shore slopes and lake levels. Sediment granulometry was similar across stations and sites, dominated by sand (63 µm < φ ≤ 2 mm; range 93–95%), followed by clay-silt (φ ≤ 63 µm; range 1–7%) and gravel (2 mm < φ ≤ 64 mm; range 1–3%). We measured the lake level three times each month at the meteorological stations of CNR-IRSA near the three sampling stations. The data are reported in the Supplementary File (Table S1). In line with our previous observations, during the survey, the highest water level (H) was recorded in July 2020 (193.91 ± 0.21 m a.s.l.), the intermediate (M) one in August 2019 (193.44 ± 0.24 m. a.s.l.) and the lowest (L) in September 2019 (193.33 ± 0.25 m a.s.l.). In total, we collected 18 samples for chemical analyses and 52 biological samples. The outline of the sampling survey is reported in the Supplementary File (Fig. S1).
Environmental survey
We collected 1 l of water for chemical analyses at 10 m from the shoreline where we also measured water temperature (T). The following variables were analysed: pH, electrical conductivity (EC), alkalinity (potentiometric methods), nitrate (ion chromatography), ammonium, total nitrogen (TN), reactive and total phosphorus, and reactive silica (UV–VIS spectrophotometry). Organic nitrogen (ON) was calculated as the difference between TN and inorganic nitrogen (N-NO3− + N-NH4+). All the analyses were performed in the laboratory of CNR-IRSA in Verbania according to standard methods for freshwater samples (APHA, AWWA, WEF, 2012; APAT IRSA-CNR, 2003). Dissolved oxygen data were available from long-term studies on Lake Maggiore funded by the CIPAIS (Rogora et al., 2018). At each site, dissolved oxygen had been monitored monthly in a previous campaign (Rogora et al., 2018). Overall, 18 environmental samples were collected during the survey (3 sampling stations × 3 water levels × 2 site types; Fig. S1).
Biological survey
At each site, we collected three spatial replicates (R1, R2 and R3) along a diagonal transect about 20 m wide reaching a maximum depth of 1.10 m. We took fixed reference points (trees or trunks fallen into the water) on the shoreline and ensured that R1 was taken around 5 m from the fixed point, R2 at about 10 m and R3 at about 20 m. Semi-quantitative biological samples were collected by disturbing an area of about 625 cm2 with a hand net following the methods in Pozojević et al. (2019). Differently from Pozojević et al. (2019), who used a mesh of 300 µm and disturbed the sediments up to 5 cm, we used a 60-µm mesh net and disturbed the substrate by foot up to 7 cm, where possible, and for 30 s to standardise the sampling time (Ausden, 1997). After sediment disturbance, the net was quickly dragged over the disturbed area opposite to the waves and then closed at the opening before being withdrawn to the surface to avoid filtering the water column. Once collected, the samples were preserved in 80% ethanol solution and bottled until being sorted in the laboratory. In July 2020, we could not collect the spatial replicates R3 in the W and D sites of the Fondo Toce sampling station because the water level was too high to allow sampling. Hence, 52 samples were collected during the biological survey (3 sampling stations × 3 water levels × 2 site types × 3 spatial replicates − 2 spatial replicates not collected at Fondo Toce station).
In the laboratory, semi-quantitative samples were standardised by sorting sediments and picking 150 meiobenthic animals individually using a glass pipette under a stereomicroscope at × 16. The size of the sub-samples (i.e. 150 ind.) was determined based on a preliminary sampling survey performed in May 2019 aimed at maximising meiobenthic taxa richness at the class/order level (Metzeling & Miller, 2001). Nine out of the 52 collected samples were completely sorted because they contained, each, exactly 150 meiobenthic individuals, while the remaining samples were only partially sorted, until the standard size of 150 individuals was reached. Copepods were identified to the species level following literature (e.g. Kiefer, 1960; Boxshall & Halsey, 2004; Dussart & Defaye, 2006). The exhaustiveness of the sampling effort for copepod assemblages was assessed using five non-parametric (Chao1, Chao2, Jackknife1, Jackknife2 and Bootstrap) and one parametric (Michaelis–Menten) estimators (Magurran & McGill, 2011 and references therein) by applying 999 randomisations without replacement.
The length and width of each collected copepod individual were measured at the stereomicroscope with the software LAS (Leica Application Suite, Version 4.7.1 of Leica Microsystems, Wetzlar, Germany). Copepod lengths were measured from the tip of the cephalic shield to the end of caudal rami, whereas width was measured at the larger somite-bearing legs. Afterwards, for each individual, body dimensions (length and width) were converted to species-specific biovolume (BV in nl) using Eq. 1 for both adult and copepodite stages, and Eq. 2 for naupliar stages (Feller & Warwick, 1988):
where BV is biovolume, a is length (mm), b is width (mm), and CF is a correction factor accounting for the body shape. CF was 560 for cyclopoids and 490 for harpacticoids, as indicated by Feller & Warwick (1988) and Schmid & Schmid-Araya (2007). Biomass was computed from the biovolume and converted into individual fresh mass assuming species-specific gravities of 1.1 (Feller & Warwick, 1988). Carbon content was estimated by assuming a dry/wet mass ratio of 0.25 and a dry mass carbon content of 40% (Feller & Warwick, 1988; Reiss & Schmid-Araya, 2008, 2010).
We selected five functional traits that we expected to respond to WLFs following Descloux et al. (2014), Di Lorenzo et al. (2021a) and Robertson et al. (2021) and allocated each collected copepod individual into one or more trait classes. We selected the following traits (for trait definitions, see Table S2): (1) diet (four classes: detritus, living microphytes, living invertebrates, omnivores), which provides information on the food items that copepods are enzymatically and behaviourally capable of using (Heino, 2008); (2) feeding habits (five classes: deposit feeders, scrapers, predators, grazers and opportunists), which refer to the functional feeding mechanisms of the copepods (Heino, 2008); (3) life history (four classes: adults, ovigerous females, copepodites and nauplii), which provides information on the age-specific schedule of development and maturity; (4) locomotion and substrate relation (two classes: swimmers and burrowers), which refers to the movement pattern of individual copepods (Heino, 2008; Martínez et al., 2021); (5) biomass, which provides information on the volume of each individual copepod (see, for instance, Descloux et al., 2014 and Neury-Ormanni et al., 2020). Biomass was treated as a continuous variable while the trait profile of each sample was obtained by weighting the individual trait class of each species by their abundance in the sample. Information on the functional traits of the copepod species collected in this study was obtained from the available literature (Fryer, 1957; Gophen, 1977; Sarvala, 1979; Hansen & Santer, 1996; Dussart & Defaye, 2001; Jersabek et al., 2001; Monakov, 2003; Tõnno et al., 2016; Adamczuk, 2021) and personal expert-based knowledge.
Data analyses
Testing Hypothesis H1
To test if late spring–summer human-induced WLFs have a significant effect on the composition and functionality of the copepod assemblages of the littoral zone of Lake Maggiore, overriding the effects induced by the geographic distribution of the sampling stations and/or the site types, we explored the environmental (Hypothesis H1a), taxonomic (Hypothesis H1b) and functional (Hypothesis H1c) data by univariate and multivariate permutational analyses of variance (PERMANOVA; Anderson et al., 2008) based on resemblance matrices and setting permutations equal to 999. Three fixed and orthogonal factors were used, namely sampling stations (three levels: BM, FT, A), water levels (three levels: L, M, H) and site type (two levels: D, W). The model equation was as follows:
where SS is the sum of squares and SSres is the residual sum of squares (the sum of squared deviations of observations from their own group mean); WL (water level), ST (sampling station) and SI (site type) are the factors. Type I sum of squares and permutation of residuals under a reduced model were applied to both univariate and multivariate models as this setting yields the best power and the most accurate type I error for unbalanced, multi-factorial designs (Anderson, 2001). The statistical design provided only one replicate per level of each factor. This implied that the WL × ST × SI interaction had to be considered null and excluded a priori from the PERMANOVA routine. Pairwise comparisons among all pairs of levels of the factors ST and WL were obtained by doing additional separate runs of the PERMANOVA routine using a direct multivariate analogue to the univariate t statistic. The analysis is comparable to a traditional univariate analysis performing a two-sample (two-tailed) t-test; however, unlike traditional statistics packages, P-values for all pairwise tests are obtained using permutations, not tables (Anderson et al., 2008). Levene’s tests were performed to check for homogeneity of variances by using the PERMDISP routine for testing the homogeneity of multivariate dispersions and the data were log(x + 1)-transformed, when necessary, prior to the analyses (Anderson, 2001). Biological and environmental data were analysed separately. The resemblance matrices of environmental data were based on Euclidean distances of normalised environmental data, whereas Bray–Curtis distances, after log(x + 1) transformation for the taxonomic and trait data, were used. A Draftsman’s plot was used to examine the correlation among environmental parameters prior to the analyses (Clarke & Green, 1988; Clarke & Gorley, 2006). Redundant variables showing > 90% correlation were excluded from the analyses. When deemed useful to enhance the result comprehension, visual complements to the significant multivariate outputs of the environmental data were provided by principal component analysis (PCA). The statistical significance for all the analyses was set at α = 0.05 since no additional corrections for multiple comparisons are required for permutational tests (Anderson et al., 2008).
Testing Hypothesis H2
To assess if the variation in environmental parameters induced by WLFs would drive composition and functionality of copepod assemblages of Lake Maggiore, we modelled univariate and multivariate linear relationships among the biological and environmental variables. The multivariate and univariate analyses were performed by a DistLM (Distance-based Linear Models) routine that models linear relationships between dissimilarity matrices of biological data and predictor environmental variable(s). This routine allows fitting one or more environmental predictors to one or more biological variables. Multivariate data using the AICc estimator of prediction error and the BEST selection procedure (Anderson et al., 2008) were explored. The AICc estimator (Akaike, 1974) was preferred over the AIC estimator since it has a better performance when the number of samples is < 40, as it is in this study (Burnham & Anderson, 2004). Statistical significance of results of the DistLM routine was assessed by permutation tests whereby each set of samples was randomly permuted 999 times relative to the others (Anderson et al., 2008). Variance explained by the models (R2) was used to assess fit of the best-solution models. Univariate linear models (marginal tests; Anderson et al., 2008) were also explored by assessing the Pearson correlation between each taxon or trait classes and the environmental variable taken one at a time. P-values obtained through 999 permutations were used to assess fit of the univariate models (statistical significance set at α = 0.05).
All analyses and plots were performed with E-PRIMER and PERMANOVA + software v. 6 (Anderson et al., 2008).
Results
Hypothesis H1a: effect of WLFs on physical and chemical variables
Raw data of the physical and chemical parameters are reported in Table 1. Long-term data available for Lake Maggiore highlighted that the lake waters are well oxygenated (values usually between 9 and 11 mg O2/L). The Draftsman’s plot highlighted a correlation > 90% between EC and total alkalinity, therefore alkalinity was excluded from analyses as a redundant variable. Results of the univariate PERMANOVAs are shown in Table S3 while the mean and standard deviation values, as well as significant pairwise post hoc t-tests, are shown in Table 2. EC, nitrate and reactive silica showed significant differences when explored under the WL factor (Table S3). However, post hoc t-tests were significant only for nitrates, which showed higher concentrations in the medium than the low level (M vs. L: t = 7.01, P-value = 0.030; Table 2) and in the high than in the low level (L vs. H: t = 12.61, P-value = 0.028; Table 2), and reactive silica, which occurred with higher concentrations at the low than the medium level (M vs. L: t = 6.96, P-value = 0.037; Table 2). EC, ammonium, reactive and total phosphorous and reactive silica showed significant differences when explored under the ST factor (Table S3). However, post hoc t-tests were significant only for EC, which was higher in Bolle di Magadino than in the other two sampling stations (FT vs. BM: t = 7.168, P-value = 0.039; A vs. BM: t = 10.45, P-value = 0.028; Table 2). Post hoc t-tests were also significant for ammonium (FT vs. BM: t = 10.97, P-value = 0.035; A vs. BM: t = 5.58, P-value = 0.047; Table 2) and reactive silica (FT vs. BM: t = 8.28, P-value = 0.032; A vs. BM: t = 13.31, P-value = 0.024; Table 2) while total phosphorous was significantly higher in Bolle di Magadino than in Angera-Sesto Calende (A vs. BM: t = 7.48, P-value = 0.032; Table 2). Total phosphorous also showed significant differences between drought and wet sites, irrespectively of the water level and sampling station (Table S3). In contrast, EC showed differences under the interaction WL × ST factors (Table S3), however, without significant differences among the sampling stations within groups of water levels.
The main test of the multivariate PERMANOVA (Table S3) highlighted a significant difference in the overall physical and chemical variables due to WL and ST factors. However, post hoc t-tests showed significant differences between sampling stations FT and BM (t = 3.39; P-value = 0.042; Fig. 2a) and between medium and low levels only (t = 3.23; P-value = 0.048; Fig. 2b). Accordingly, the PCA explained 66.2% of the cumulative variance of the physical and chemical data on the first two axes (Fig. 2a). Samples collected at Bolle di Magadino showed the highest EC values, and the highest concentrations of ammonium, reactive and total phosphorus, as well as reactive silica and were clearly separated from the samples collected at Angera-Sesto Calende and Fondo Toce on the first PCA axis which accounted for 46.2% of the data variability (Fig. 2a). Samples collected during the medium water level period showed the highest concentrations of nitrate and TN and the highest temperature. These samples were separated, though not completely, from those collected during the low and high water level periods on the second PCA axis which accounted, however, for only 20% of the data variability (Fig. 2b).

Plots of the principal components analysis showing the separation of the samples according to: a sampling stations (ST), and b water level (WL). For the ST factor in a: Bolle di Magadino (light blue circles), Fondo Toce (green triangles), Angera-Sesto Calende (white squares). For the WL factor in b: medium water level (grey triangles), low water level (red circles) and high water level (green squares). Variables abbreviation as in Table 1
Hypothesis H1b: effect of WLFs on copepod assemblage composition
Overall, we sorted 7,800 meiobenthic animal individuals, of which 1,375 were copepods. The remaining individuals were rotifers, tardigrades, nematodes (mainly), oligochaetes, bivalves, mites, and early larval stages of insects (ephemeropterans and chironomids, i.e. “temporary meiofauna” according to Giere, 2009). Out of the 1,375 copepods, 856 were adults or copepodites attributed to 11 species, of which 6 belonged to the order Cyclopoida and 5 to the order Harpacticoida: Macrocyclops albidus (Jurine, 1820), Paracyclops fimbriatus fimbriatus (Fischer, 1853), Eucyclops serrulatus serrulatus (Fischer, 1851), Eucyclops lilljeborgi (Sars G.O., 1918), Eucyclops macrurus macrurus (Sars G.O., 1863), Mesocyclops leuckarti (Claus, 1857), Attheyella (Attheyella) crassa (Sars G.O., 1863), Bryocamptus (Echinocamptus) echinatus (Mrázek, 1893), Nitokra hibernica hibernica (Brady, 1880), Epactophanes richardi (Mrázek, 1893) and Bryocamptus (Echinocamptus) hoferi (Van Douwe, 1908). Since nauplii are all very similar, they were not assigned to any species. Copepod abundances (adults and copepodites) are reported in Table 3 and expressed as the sum of abundances of the three spatial replicates at each sampling station, site type and water level. All the estimators indicated that we captured the whole expected copepod biodiversity (expected number of taxa: Chao1: 11; Chao2: 11; Jacknife1: 11; Jacknife2: 10 and Michaelis–Menten: 11; Fig. S2).
Results of univariate and multivariate PERMANOVAs are shown in Table S4, while significant pairwise post hoc t-tests are shown in Table 2. Abundances of the individual species did not show significant changes, except for the abundances of P. fimbriatus under the WL factor and those of E. lilljeborgi under the ST factor (Table S4). However, post hoc t-tests showed significant differences only for P. fimbriatus, which was more abundant in medium than in high water level (M vs. H: t = 15.003, P-value = 0.024; Table 2) and in the low than in the high level (L vs. H: t = 4.929, P-value = 0.041; Table 2). Abundances of the whole copepod assemblage (multivariate scenario) significantly changed when explored under both WL and ST factors (Table S4); however, the pairwise post hoc t-tests did not reveal significant differences among levels of the ST factor, while abundances were significantly higher in the high than in the low level (L vs. H: t = 2.47, P-value = 0.027; Table 2).
Hypothesis H1c: effect of WLFs on copepod assemblage functionality
Abundances of different trait classes are reported in Table 3. Results of the univariate and multivariate PERMANOVAs are shown in Table S4, while significant pairwise post hoc t-tests are shown in Table 2. Biomass showed significant differences under the WL factor (Table S4): biomasses were significantly higher in the high than in the low level (H vs. L: t = 8.77, P-value = 0.026; Table 2). Biomasses were also significantly different when explored under the interaction ST × SI (Table S4); however, post hoc t-tests were not significant. For the trait diet only, abundances of omnivores showed significant differences under the ST and WL factors as well as under their interaction (Table S4). Post hoc t-tests revealed that abundances were higher in the low than in the high level (L vs. H: t = 5.54, P-value = 0.037; Table 2) and in the medium than in the high level (M vs. H: t = 11.27, P-value = 0.024; Table 2) but not among the sampling stations. In the multivariate scenario, classes of the trait diet showed no differences (Table S4). For the trait feeding habits, only the deposit feeders and opportunists showed differences when explored under the WL factor (Table S4). Abundances of the deposit feeders were significantly higher in medium than in high level (M vs. H: t = 6.39, P-value = 0.027; Table 2) and in low than in high level (L vs. H: t = 4.11, P-value = 0.039; Table 2) as well as the abundances of the opportunists (M vs. H: t = 15.00, P-value = 0.018; L vs. H: t = 4.92, P-value = 0.045; Table 2). In the multivariate scenario, classes of the trait feeding habits varied according to the water level and under the interaction ST × SI (Table S4). However, the differences were significant only between the high and medium levels (M vs. H: t = 3.94, P-value = 0.036). Overall abundances of this trait were higher in the medium level than in the other two. For the trait life history, abundances of copepodites, nauplii and ovigerous females showed significant differences when explored under the WL factor (Table S4), while copepodites showed differences under the interaction ST × SI, as well. However, post hoc t-tests showed significant differences only in abundance of ovigerous females that were higher in the high than in the medium level (M vs. H: t = 4.78, P-value = 0.040; Table 2). In the multivariate scenario, classes of the trait life history varied according to the water level (Table S4). However, pairwise post hoc t-tests were not significant. Finally, for the trait locomotion and substrate relation, abundances of burrowers and swimmers did not vary either in the univariate or in the multivariate scenario (Table S4).
Hypothesis H2: effect of environmental parameters on copepod assemblages
Concerning the copepod assemblage composition, the best DistLM consisted of the parameter EC (AICc = 134.08), which explained 15% of the total variance of copepod abundances. When individually taken, the species E. lilljeborgi showed significant correlations with the environmental variables, particularly EC (r = − 0.60; P-value = 0.004) and ammonium concentrations (r = − 0.55; P-value = 0.009). The abundances of this species were lower in the samples characterised by higher values of the two environmental parameters. Regarding the copepod assemblage functionality, the best multivariate DistLM for biomass consisted of the parameter nitrate (AICc = 50.06) which explained 22.7% of the total variation. Univariate analyses showed significant correlation of biomass with nitrate (r = 0.48; P-value = 0.047). The best multivariate DistLM for the trait diet consisted of the sole parameter reactive phosphorus (AICc = 121.9) which, however, explained only 9% of total variation. Univariate models explored for each class of the trait diet were not significant. Similarly, the best multivariate DistLM for the trait feeding habits consisted of the sole parameter EC (AICc = 120.3) which, however, explained only 9% of total variation. Univariate models were not significant. Reactive silica was the sole parameter of the best DistLM model (AICc = 106.7) for the trait life history which, however, explained 8% of the total variation, while TN was the sole parameter explaining the highest variation (9%) of the trait locomotion and substrate relation (AICc = 98.08). Univariate models explored for each class of the traits life history and locomotion and substrate relation were not significant.
Discussion
The results of this study suggest that late spring–summer human-induced WLFs in Lake Maggiore drive significant variability in the ecology of the littoral zone, affecting not only the composition and functioning of the benthic copepod assemblages but also environmental conditions.
Effect of WLFs on physical and chemical parameters
In this study, we found significant variation, due to human-induced WLFs, in 2 out of the 13 analysed physical and chemical variables, namely nitrate and reactive silica. Higher nitrate concentrations occurring during the high and medium water levels were likely due to nitrification increasing in the re-flooded sediments, as observed in previous studies (e.g. Carmignani & Roy, 2017). On the other hand, since the atmospheric deposition was the main source of nitrate to Lake Maggiore (Rogora et al., 2006), the higher nitrates occurring in the medium and high water levels might also depend on precipitation inputs occurring prior to the sampling survey in July and August. The observed pattern of reactive silica showed higher concentrations in the low rather than in the medium water level. Silica, generated through the chemical weathering of silicate minerals, is transported to the lake by tributaries and is utilised by diatoms (Wang et al., 2010). Due to dissolution phenomena affecting sedimentation of diatom frustules, silica concentrations usually increase with water level (and, consequently, with depth) in lakes (Cantonati et al., 2009 and references therein). Disturbance due to human-induced WLFs may alter this pattern (e.g. Li et al., 2021), as observed in this study.
Effect of WLFs on copepod assemblage composition
Four out of the 11 benthic copepod species collected in this study are new records for Lake Maggiore, namely: B. hoferi, E. richardi, N. hibernica and E. lilljeborgi (Italian Ministry of Environment, 2000). The occurrence of E. lilljeborgi represents the first record of this species from Italy. Although E. lilljeborgi is not generally considered a common species, it occurs in the littoral epibenthos of ponds and lakes almost worldwide. Its distribution covers the Palearctic, Nearctic and Afrotropical regions. Overall species richness did not vary significantly among the three water levels. However, overall diversity was lower than that observed in the littoral zones of other oligotrophic lakes not affected by human-induced WLFs (e.g. Sarvala, 1986). Relatively low species richness observed in this study seemed independent of the sampling methodology since the saturation of species accumulation curves indicated that sampling effort was adequate to capture the whole expected copepod diversity.
Copepod abundances varied significantly among the three water levels used in this study. A low density of crustacean species at all sampling depths in freshwater lakes with altered WLFs was observed in previous studies (e.g. Witthöft-Mühlmann et al., 2007; Evtimova & Donohue 2014). The detrimental effect of altered WLFs on abundances of meiobenthic taxa has also been observed for benthic nematodes, chironomids, oligochaetes, mites, platyhelminths and gastropods that tend to reduce their abundances drastically during low water level periods (e.g. Witthöft-Mühlmann et al., 2007; Mastrantuono et al., 2008), as also observed in this study. Reduced abundances during the low water levels could be due to an avoidance reaction: in fact, most (but not all) meiofauna react to disturbance due to drawdowns by attempting to escape through migration (Giere, 2009 and references therein). For instance, marine benthic harpacticoids show avoidance reactions to increasing wave action due to the vibration of the sediments (Foy & Thistle, 1991). In this study, an avoidance reaction could explain the low overall abundances observed during the low water level when the littoral zone of the Lake Maggiore was more disturbed by waves. Indeed, the sudden exposure of sites during the low water level period determines sediment mobilisation, erosion, suspension and alteration of the interstitial water flow, which is usually accompanied by reduced meiofaunal diversity and abundances (Giere, 2009 and references therein). On the other hand, sediment disturbance due to our sampling methodology, which seemed not to affect the species richness, could play a role in the overall abundances as well. Phenology of the collected species seemed not to determine the observed pattern since 10 out of the 11 collected species are known to have a peak abundance in August or September (Table 4), corresponding to the medium and low water-level periods in this study.
Effect of WLFs on copepod assemblage functionality
We found significant variation in functional composition of benthic copepod assemblages due to late spring–summer human-induced WLFs, at least concerning biomass and some trait classes. We observed variation in four functional classes (ovigerous females, deposit feeders, opportunists and omnivores) due to human-induced WLFs but no significant changes due to sampling stations or site type. Copepod biomass was significantly lower in the low water level. Similarly, in a study concerning water-level regulation in Finnish lake littoral zones, Palomäki (1994) observed that variation in light penetration in the whole littoral area due to human-induced WLFs disturbed the zonation of the benthos and the biomass of the zoobenthos. In this study, copepod biomass showed a significant positive correlation with nitrate. According to the findings of previous studies (e.g. Vakkilainen et al., 2004; Ristau et al., 2015; Sodré & Bozelli, 2019), higher biomass found in the high water level period could be due to mechanisms related to an increase in nutrients which, in turn, are triggered by WLFs. For instance, Sodré & Bozelli (2019) observed that biomass of planktonic microcrustaceans, including copepods, is mainly dependent on temperature and predation pressure, but also on nutrients. Similarly, Vakkilainen et al. (2004) found that biomass of the littoral invertebrate assemblage across six European lakes was positively related to nutrient enrichment. The increase of copepod biomasses with nutrient increase found by Ristau et al. (2015) seems to be in line with our results as well.
Omnivores (trait diet), deposit feeders and opportunists (trait feeding habits) varied significantly in their abundances during this study, being more abundant during the medium–low water levels than in the high one, as also observed in previous studies examining the effect of human-induced WLFs on benthic invertebrates (e.g. Evtimova & Donohue, 2016). For instance, under low water level conditions (that is, when the habitat is more disturbed by waves due to air exposure), increased abundances of deposit-feeder nematodes were observed at the river mouth area of Lake Constance (Witthöft-Mühlmann et al., 2007). As postulated by Death & Winterbourn (1995), variability in abundances of deposit feeders, omnivores and opportunists may be indicative of the role of colonisers able to resist, and even thrive, under disturbance events in dynamic environments. Variable densities of generalist feeders in lakes with altered WLFs could be attributed to their ability to employ various feeding strategies to cope with changes in food availability owing to their broad range of potential food sources. In this study, higher abundances of opportunists and omnivores in the medium/low water levels were mainly due to P. fimbriatus, a common species in benthic habitats of the littoral and profundal zone of lakes (Gaviria, 1998). Paracyclops fimbriatus burrows among sediment particles, preferring sandy substrates (Särkkä, 1996; Sarvala, 1988; Dole-Olivier et al., 2000) and has a fast development (65 days at 11.5 °C). Being an opportunist, P. fimbriatus may interact with different trophic levels by feeding on algae (Hopp & Maier, 2005), or preying on ciliates (Agasild et al., 2013) and smaller metazooplankters (Hansen & Santer, 1996). It is possible that the varied feeding behaviour of this species was an advantage in the medium–low water levels.
The higher number of ovigerous females observed in the high water level seemed not to be related to the phenology of the species since most of them have a peak of abundances in August or September (see Sect. 4.2; Table 4). Hart (1985) observed that seasonality of zoobenthos and zooplankton is often related to hydrological events such as water fluctuations that potentially affect food and habitat availability. Cifoni et al. (2017) observed that abundances of ovigerous females of E. serrulatus tended to increase with resource availability and temperature in laboratory conditions. Accordingly, the increase in nitrates observed in this study during high water level suggests an increase in nutrients which, in turn, might have boosted egg production, as it seems from the positive (although weak) correlation of nitrates with the abundance of ovigerous females observed in our study. The thermal regime, which is a determinant of copepod productivity (e.g. Hart, 1985; Sarvala, 1986), likely had no role in this study. For instance, Fefilova (2007) observed that abundance of ovigerous females of harpacticoid species (among these, A. crassa and E. richardi also found in our study) in the northeast lakes of European Russia were controlled primarily by water temperature. High temperatures determined the formation of egg-sacs in females and influenced the speed and duration of ontogenesis so that abundances of ovigerous females reached their peak between July and August. However, this pattern does not fit our data because water temperature did not vary significantly among the three water levels.
Other determinant factors
The location of the sampling stations seemed to determine patterns of EC and nutrients, organic matter and solutes in general (affecting conductivity) that were higher in the Bolle di Magadino than in the other two stations. However, these differences in nutrients were likely due to a cormorant nest located near the permanently wet site of the sampling station Bolle di Magadino and should be considered a temporary event. Other factors, such as proximity to inflows, may also affect solutes and other environmental parameters and determine the high values of EC in this station.
Apart from ammonium, none of the above parameters had a significant correlation, linear at least, with the composition and functionality of copepod assemblages. On the other hand, a negative correlation between ammonium and copepod abundances was found for E. lilljeborgi. This result is in line with our previous studies in which we demonstrated that even low concentrations of ammonium (a few µg/L) might be detrimental for freshwater copepod species, affecting their survival, development, metabolism, biomass, reproduction and dispersal (e.g. Di Marzio et al., 2009, 2013, 2018; Di Lorenzo et al., 2014, 2015, 2016).
Despite the absence of significant variation in our taxonomic variables (species richness and abundance partition) among our sampling stations, the composition of copepod assemblages at the three sampling stations differed in terms of the type of species. In detail, E. macrurus was found only in Angera-Sesto Calende station, while B. hoferi occurred at Fondo Toce only. Macrocyclops albidus and E. richardi were collected from Angera-Sesto Calende and Fondo Toce but not from Bolle di Magadino. The differences in the assemblages might be due to further factors not investigated in this study. For instance, the composition of microbial communities might determine the occurrence of deposit feeders and scrapers such as E. richardi and B. hoferi, while the dimension of the interstitial voids might represent an impediment for grazers, such as E. macrurus, and predators such as M. albidus. Since microbial communities significantly differ among habitats in Lake Maggiore, potentially because of specific mechanisms creating and maintaining habitat filtering and structure (Eckert et al., 2020), correlations between microbial communities and benthic copepod assemblages should be further investigated.
Conclusions
Our results indicated that the late spring–summer human-induced WLFs of Lake Maggiore drive significant differences in the ecology of the littoral zone, affecting the structure and functioning of benthic copepod assemblages. In particular, overall abundance and biomass, as well as the abundance of ovigerous females, opportunists, omnivores and deposit feeders, significantly changed during the survey, albeit none of the investigated trait classes was completely lost. Since variation in the composition and functionality of a taxonomic assemblage can have cascading effects on the whole community, the results of our study suggest that the ecosystem services provided by the littoral zone of Lake Maggiore are likely altered due to human-induced WLFs. This study supports mixed (taxonomy-based and trait-based) approaches for assessing the effect of human-induced WLFs on meiobenthic communities. Further functional traits, such as resting stage, number of cycles per year and reproduction, would be helpful to strengthen the conclusions of this study. Resting stage, in particular, seems to be a promising trait because of its cascading effects on reproduction, population dynamics, as well as the phenology of predators and living food items. Finally, the sampling effort of our survey seemed to have caught the diversity of the benthic copepod assemblages of Lake Maggiore, also providing four new species records. However, increasing the number of the sampling stations and extending the monitoring for a further year would be useful to confirm the observed patterns.
Data, material and/or code availability
All data and material and code generated or analysed during this study are included in this published article and its Supplementary Information files.
References
Adamczuk, M., 2021. Demographic consequences of mitigating strategies in planktonic invertebrates facing global browning of freshwater ecosystem. Aquatic Sciences 83: 31.
Agasild, H., P. Zingel, K. Karus, K. Kangro, J. Salujõe & T. Nõges, 2013. Does metazooplankton regulate the ciliate community in a shallow eutrophic lake? Freshwater Biology 58: 183–191.
Akaike, H., 1974. A new look at the statistical model identification. IEEE Transactions on Control Systems Technology 19: 716–723.
Allan, J. D., S. D. Smith, P. B. McIntyre, C. A. Joseph, C. E. Dickinson, A. L. Marino, R. G. Biel, J. C. Olson, P. J. Doran, E. S. Rutherford, J. E. Adkins & A. O. Adeyemo, 2015. Using cultural ecosystem services to inform restoration priorities in the Laurentian Great Lakes. Frontiers in Ecology and the Environment 13: 418–424.
Ambrosetti, W., L. Barbanti & A. Rolla, 2006. The climate of Lago Maggiore area during the last fifty years. Journal of Limnology 65: 1–62.
Anderson, M. J., 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26: 32–46.
Anderson, M. J., R. N. Gorley & K. R. Clarke, 2008. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods. PRIMER-Ev Ltd., Plymouth: 214.
APAT IRSA-CNR, 2003. Metodi analitici per le acque. APAT Manuali e Linee Guida 29/2003.
APHA, AWWA, WEF, 2012. Standard Methods for the Examination of Water and Wastewater, 22nd ed. American Public Health Association, Washington, DC:
Arfè, A., P. Quatto, A. Zambon, H. J. MacIsaac & M. Manca, 2019. Long-term changes in the zooplankton community of Lake Maggiore in response to multiple stressors: a functional principal components analysis. Water 11(5): 962.
Ausden, M., 1997. Invertebrates. In Sutherland, W. J. (ed), Ecological Census Techniques: A Handbook Cambridge University Press, Cambridge: 139–177.
Baumgärtner, D., M. Mörtl & K. O. Rothhaupt, 2008. Effects of water-depth and water-level fluctuations on the macroinvertebrate community structure in the littoral zone of Lake Constance. Hydrobiologia 613: 97–107.
Boggero, A., M. Ciampittiello, M. Cifoni, T. Di Lorenzo, L. Kamburska, D. Paganelli, N. Riccardi, M. Rogora & S. Zaupa, 2021. Lago Maggiore: il futuro della gestione dei livelli. Nimbus 85(1).
Boxshall, G. A. & S. H. Halsey, 2004. An Introduction to Copepod Diversity, 166 (Part I). Ray Society, London: 1–421.
Brancelj, A., M. Sisko, I. R. Brancelj, Z. Jeran & R. Jacimovic, 2000. Effects of land use and fish stocking on a mountain lake-evidence from the sediment. Periodicum Biologorum 102: 259–268.
Brauns, M., X. F. Garcia & M. T. Pusch, 2008. Potential effects of water-level fluctuations on littoral invertebrates in lowland lakes. Hydrobiologia 613: 5–12.
Brauns, M., B. Gücker, C. Wagner, X. F. Garcia, N. Walz & M. T. Pusch, 2011. Human lakeshore development alters the structure and trophic basis of littoral food webs. Journal of Applied Ecology 48: 916–925.
Brun, P., M. R. Payne & T. Kiørboe, 2016. Trait biogeography of marine copepods – an analysis across scales. Ecology Letters 19: 1403–1413.
Burnham, K. P. & D. R. Anderson, 2004. Multi-model inference: understanding AIC and BIC in model selection. Sociological Methods and Research 33: 261–304.
Callieri, C., R. Bertoni, M. Contesini & F. Bertoni, 2014. Lake level fluctuations boost toxic cyanobacterial “Oligotrophic Blooms.” PLoS ONE 9(10): e109526.
Cantonati, M., S. Scola, N. Angeli, G. Guella & R. Frassanito, 2009. Environmental controls of epilithic diatom depth-distribution in an oligotrophic lake characterized by marked water-level fluctuations. European Journal of Phycology 44(1): 15–29.
Cantonati, M., S. Poikane, C. M. Pringle, L. E. Stevens, E. Turak, J. Heino, J. S. Richardson, R. Bolpagni, A. Borrini, N. Cid, M. Čtvrtlíková, D. M. P. Galassi, M. Hájek, I. Hawes, Z. Levkov, L. Naselli-Flores, A. A. Saber, M. Di Cicco, B. Fiasca, P. B. Hamilton, J. Kubečka, S. Segadelli & P. Znachor, 2020. Characteristics, main impacts, and stewardship of natural and artificial freshwater environments: consequences for biodiversity conservation. Water 12: 260.
Cantonati, M., R. Zorza, M. Bertoli, P. Pastorino, G. Salvi, G. Platania, M. Prearo & E. Pizzul, 2021. Recent and subfossil diatom assemblages as indicators of environmental change (including fish introduction) in a high-mountain lake. Ecological Indicators 125: 107603.
Caramujo, M.-J., C. R. B. Mendes, P. Cartaxana, V. Brotas & M.-J. Boavida, 2008. Influence of drought on algal biofilms and meiofaunal assemblages of temperate reservoirs and rivers. Hydrobiologia 598: 77–94.
Carmignani, J. R. & A. H. Roy, 2017. Ecological impacts of winter water level drawdowns on lake littoral zones: a review. Aquatic Science 79: 803–824.
Carrara, E. A., L. Barbanti & W. Ambrosetti, 2009. Deep waters warming in Italian lakes: secular variations. In AIOL/S.it.E. Congress, Ancona, 2007: 555–561.
Ciampittiello, M., C. Dresti, H. Saidi & D. Manca, 2019. Indagini sul bacino imbrifero. Caratteristiche idrologiche - Ricerche sull'evoluzione del Lago Maggiore. Aspetti limnologici. Programma triennale 2016–2018. Campagna 2018 e relazione triennio 2016–2018. Commissione Internazionale per la protezione delle acque italo-svizzere (eds): 9–21 and 96–100. ISSN 1013-8099.
Cifoni, M., D. M. P. Galassi, C. Faraloni & T. Di Lorenzo, 2017. Test procedures for measuring the (sub)chronic effects of chemicals on the freshwater cyclopoid Eucyclops serrulatus. Chemosphere 173: 89–98.
Cifoni, M., A. Boggero, D. M. P. Galassi & T. Di Lorenzo, 2021. An overview of studies on meiofaunal traits of the littoral zone of lakes. Water 13: 473.
Clarke, K. R. & R. N. Gorley, 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth: 192.
Clarke, K. R. & R. H. Green, 1988. Statistical design and analysis for a ‘biological effects’ study. Marine Ecology 46: 213–226.
Colina, M., D. Calliari, C. Carballo & C. Kruk, 2016. A trait-based approach to summarize zooplankton–phytoplankton interactions in freshwaters. Hydrobiologia 767: 221–233.
Death, R. G. & M. J. Winterbourn, 1995. Diversity patterns in stream benthic invertebrate communities: the influence of habitat stability. Ecology 76: 1446–1460.
Descloux, S., T. Datry & P. Usseglio-Polatera, 2014. Trait-based structure of invertebrates along a gradient of sediment colmation: benthos versus hyporheos responses. Science of the Total Environment 466–467: 265–276.
Di Cicco, M., T. Di Lorenzo, M. Iannella, I. Vaccarelli, D. M. P. Galassi & B. Fiasca, 2021. Linking hydrogeology and ecology in Karst landscapes: the response of Epigean and obligate groundwater copepods (Crustacea: Copepoda). Water 13: 2106.
Di Lorenzo, T., W. D. Di Marzio, M. E. Sáenz, M. Baratti, A. Dedonno, A. Iannucci & D. M. P. Galassi, 2014. Sensitivity of hypogean and epigean freshwater copepods to agricultural pollutants. Environmental Science and Pollution Research 21: 4643–4655.
Di Lorenzo, T., M. Cifoni, P. Lombardo, B. Fiasca & D. M. P. Galassi, 2015. Ammonium threshold values for groundwater quality in the EU may not protect groundwater fauna: evidence from an alluvial aquifer in Italy. Hydrobiologia 743: 139–150.
Di Lorenzo, T., S. Cannicci, D. Spigoli, M. Cifoni, M. Baratti & D. M. P. Galassi, 2016. Bioenergetic cost of living in polluted freshwater bodies: respiration rates of the cyclopoid Eucyclops serrulatus under ammonia-N exposures. Fundamental and Applied Limnology 188: 147–156.
Di Lorenzo, T., B. Fiasca, M. Di Cicco, M. Cifoni & D. M. P. Galassi, 2021a. Taxonomic and functional trait variation along a gradient of ammonium contamination in the hyporheic zone of a Mediterranean stream. Ecological Indicators 132: 108268.
Di Lorenzo, T., B. Fiasca, M. Di Cicco & D. M. P. Galassi, 2021b. The impact of nitrate on the groundwater assemblages of European unconsolidated aquifers is likely less severe than expected. Environmental Science and Pollution Research 28: 11518–11527.
Di Marzio, W. D., D. Castaldo, C. Pantani, A. Di Cioccio, T. Di Lorenzo, M. E. Sáenz & D. M. P. Galassi, 2009. Relative sensitivity of hyporheic copepods to chemicals. Bulletin of Environmental Contamination and Toxicology 82: 488–491.
Di Marzio, W. D., D. Castaldo, T. Di Lorenzo, A. Di Cioccio, M. E. Sáenz & D. M. P. Galassi, 2013. Developmental endpoints of chronic exposure to suspected endocrine-disrupting chemicals on benthic and hyporheic freshwater copepods. Ecotoxicology and Environmental Safety 96: 86–92.
Di Marzio, W. D., M. Cifoni, M. E. Sáenz, D. M. P. Galassi & T. Di Lorenzo, 2018. The ecotoxicity of binary mixtures of Imazamox and ionized ammonia on freshwater copepods: implications for environmental risk assessment in groundwater bodies. Ecotoxicology and Environmental Safety 149: 72–79.
Dole-Olivier, M. J., D. M. P. Galassi, P. Marmonier & M. Creuzé Des Châtelliers, 2000. The biology and ecology of lotic microcrustaceans. Freshwater Biology 4: 63–91.
Dussart, B. H. & D. Defaye, 2001. Introduction to the Copepoda. In Dumont, H. J. (ed), Guides to the Identification of the Macroinvertebrates of the Continental Waters of the World, Vol. 16. Backhuys, Leiden: 344.
Dussart, B. H. & D. Defaye, 2006. World Directory of Crustacea Copepoda of Inland Waters. II – Cyclopiformes. Backhuys Publishers, Leiden
Eckert, E. M., S. Amalfitano, A. Di Cesare, C. Manzari, G. Corno & D. Fontaneto, 2020. Different substrates within a lake harbour connected but specialised microbial communities. Hydrobiologia 847(7): 1689–1704.
Evtimova, V. V. & I. Donohue, 2014. Quantifying ecological responses to amplified water level fluctuations in standing waters: an experimental approach. Journal of Applied Ecology 51: 1282–1291.
Evtimova, V. V. & I. Donohue, 2016. Water-level fluctuations regulate the structure and functioning of natural lakes. Freshwater Biology 61: 251–264.
Fefilova, E., 2007. Seasonal development of harpacticoid copepods in the North-East of European Russia. Fundamental and Applied Limnology 170: 65–75.
Feller, R. J. & R. M. Warwick, 1988. Energetics. In Higgins, R. P. & H. Thiel (eds), Introduction to the Study of Meiofauna Smithsonian Institution Press, Washington, DC: 181–196.
Fenocchi, A., M. Rogora, S. Sibilla & C. Dresti, 2017. Relevance of inflows on the thermodynamic structure and on the modelling of a deep subalpine lake (Lake Maggiore, Northern Italy/Southern Switzerland). Limnologica 63: 42–56.
Fonseca, G., D. Fontaneto & M. Di Domenico, 2018. Addressing biodiversity shortfalls in meiofauna. Journal of Experimental Marine Biology and Ecology 502: 26–38.
Foy, M. S. & D. Thistle, 1991. On the vertical distribution of a benthic harpacticoid copepod: field, laboratory, and flume results. Journal of Experimental Marine Biology and Ecology 153: 153–164.
Fryer, G., 1957. The food of some freshwater cyclopoid copepods and its ecological significance. Journal of Animal Ecology 26: 263–286.
Galassi, D. M. P., 2001. Groundwater copepods (Crustacea: Copepoda): diversity patterns over ecological and evolutionary scales. Hydrobiologia 453: 227–253.
Galassi, D. M. P., R. Huys & J. W. Reid, 2009a. Diversity, ecology and evolution of groundwater copepods. Freshwater Biology 54: 691–708.
Galassi, D. M. P., F. Stoch, B. Fiasca, T. Di Lorenzo & E. Gattone, 2009b. Groundwater biodiversity patterns in the Lessinian Massif of northern Italy. Freshwater Biology 54: 830–847.
Gaviria, S., 1998. Colonization of a new man-made river (Marchfeld Canal, Lower Austria) by benthic copepods. Journal of Marine Systems 15: 127–134.
Gerino, M., G. Stora, F. François-Carcaillet, F. Gilbert, J. Poggiale, F. Mermillod-Blondin, G. Desrosiers & P. Vervier, 2003. Macro-invertebrate functional groups in freshwater and marine sediments: a common mechanistic classification. Vie et Milieu 53: 221–231.
Giere, O., 2009. Meiobenthology. The Microscopic Motile Fauna of Aquatic Sediments. Springer, Berlin.
Gophen, M., 1977. Food and feeding habits of Mesocyclops leuckarti (Claus) in Lake Kinneret (Israel). Freshwater Biology 7: 513–518.
Hallstan, S., C. Trigal, K. S. L. Johansson & R. K. Johnson, 2013. The impact of climate on the geographical distribution of phytoplankton species in boreal lakes. Oecologia 173: 1625–1638.
Hansen, A. M. & B. Santer, 1996. The influence of food resources on the development, survival and reproduction of the two cyclopoid copepods: Cyclops vicinus and Mesocyclops leuckarti. Journal of Plankton Research 17: 631–646.
Hart, R. C., 1985. Embryonic development times of entomostracan zooplankton from Lake le Roux (Orange River, South Africa), and their possible relationships to seasonal succession. Hydrobiologia 127: 17–26.
Heino, J., 2008. Patterns of functional biodiversity and function–environment relationships in lake littoral macroinvertebrates. Limnology and Oceanography 53: 1446–1455.
Hofmann, H., A. Lorke & F. Peeters, 2008. Temporal scales of water-level fluctuations in lakes and their ecological implications. Hydrobiologia 613: 85–96.
Hopp, U. & G. Maier, 2005. Survival and development of five species of cyclopoid copepods in relation to food supply: experiments with algal food in a flow-through system. Freshwater Biology 50: 1454–1463.
Iannella, M., B. Fiasca, T. Di Lorenzo, M. Biondi, M. Di Cicco & D. M. P. Galassi, 2020. Spatial distribution of stygobitic crustacean harpacticoids at the boundaries of groundwater habitat types in Europe. Scientific Reports 10: 19043.
Italian Ministry of Environment, 2000. Checklist of the Species of the Italian Fauna [available on internet at: https://www.faunaitalia.it/checklist/]. Accessed 20 April 2022.
Jackson, J. K. & L. Fuereder, 2006. Long-term studies of freshwater macroinvertebrates: a review of the frequency, duration and ecological significance. Freshwater Biology 51: 591–603.
Jersabek, C. D., A. Brancelj, F. Stoch & R. Schabetsberger, 2001. Distribution and ecology of copepods in mountainous regions of the Eastern Alps. Hydrobiologia 453(454): 309–324.
Kiefer, F., 1960. Beiträge zur Copepodenkunde (XX). Zoologischer Anzeiger 165: 37–45.
Kreuzinger-Janik, B., F. Schroeder, N. Majdi & W. Traunspurger, 2015. Depth-related effects on a meiofaunal community dwelling in the periphyton of a mesotrophic lake. PLoS ONE 10: e0137793.
Leira, M. & M. Cantonati, 2008. Effects of water-level fluctuations on lakes: an annotated bibliography. Hydrobiologia 613: 171–184.
Li, T., B. He, Y. Zhang, S. Fu, Y. Zhang & X. Long, 2021. Spatiotemporal dynamics of coupled dissolved silica and bicarbonate in a dam-induced urban lake system in the Three Gorges Reservoir Area. Journal of Hydrology 597: 126181.
Magurran, A. E. & B. J. McGill, 2011. Biological Diversity: Frontiers in Measurement and Assessment, University Press, Oxford:
Majdi, N., J. M. Schmid-Araya & W. Traunspurger, 2020. Preface: patterns and processes of meiofauna in freshwater ecosystems. Hydrobiologia 847: 2587–2595.
Mammides, C., 2020. A global assessment of the human pressure on the world’s lakes. Global Environmental Change 63: 102084.
Manca, M. & R. Bertoni, 2014. Seventy-five years of limnology at the Istituto Italiano di Idrobiologia in Pallanza. Journal of Limnology 73: 5–19.
Martínez, A., G. García-Gómez, A. García-Herrero, N. Sánchez, F. Pardos, A. Izquierdo-Muñoz, D. Fontaneto & S. Mammola, 2021. Habitat differences filter functional diversity of low dispersive microscopic animals. Hydrobiologia 848: 681–2698.
Mastrantuono, L., A. Solimini, P. Noges & M. Bazzanti, 2008. Plant-associated invertebrates and hydrological balance in the large volcanic Lake Bracciano (Central Italy) during two years with different water levels. Hydrobiologia 599: 143–152.
Metzeling, L. & J. Miller, 2001. Evaluation of the sample size used for the rapid bioassessment of rivers using macroinvertebrates. Hydrobiologia 444: 159–170.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-Being – Biodiversity Synthesis. Report.
Monakov, A. V., 2003. In Monakov, A. V. (ed.), Feeding of Freshwater Invertebrates. Kenobi Productions, Ghent: 373.
Morabito, G., M. Rogora, M. Austoni & M. Ciampittiello, 2018. Could the extreme meteorological events in Lake Maggiore watershed determine a climate-driven eutrophication process? Hydrobiologia 824: 163–175.
Neury-Ormanni, J., J. Vedrenne, M. Wagner, G. Jan & S. Morin, 2020. Micro-meiofauna morphofunctional traits linked to trophic activity. Hydrobiologia 847: 2725–2736.
Palomäki, R., 1994. Response by macrozoobenthos biomass to water level regulation in some Finnish lake littoral zones. Hydrobiologia 286: 17–26.
Poikane, S., F. Salas Herrero, M. G. Kelly, A. Borja, S. Birke & W. de Bunda, 2020. European aquatic ecological assessment methods: a critical review of their sensitivity to key pressures. Science of the Total Environment 740: 140075.
Pozojević, I., L. Juršić, N. Vučković, V. Dorić, S. Gottstein, I. Ternjej & Z. Mihaljević, 2019. Is the spatial distribution of lentic water mite assemblages (Acari: Hydrachnidia) governed by prey availability? Experimental and Applied Acarology 77: 487–510.
Reid, J. W. & C. E. Williamson, 2010. Copepoda. In Thorp, J. H. & A. P. Covich (eds), Ecology and Classification of North American Freshwater Invertebrates 3rd ed. Academic, Elsevier: 829–899.
Reiss, J. & J. M. Schmid-Araya, 2008. Existing in plenty: abundance, biomass and diversity of ciliates and meiofauna in small streams. Freshwater Biology 53: 652–668.
Reiss, J. & J. M. Schmid-Araya, 2010. Life history allometries and production of small fauna. Ecology 91: 497–507.
Ristau, K., N. Spann & W. Traunspurger, 2015. Species and trait compositions of freshwater nematodes as indicative descriptors of lake eutrophication. Ecological Indicators 53: 196–205.
Robertson, A. L., D. M. Perkins, J. England & T. Johns, 2021. Invertebrate responses to restoration across benthic and hyporheic stream compartments. Water 13: 996.
Rogora, M., R. Mosello, A. Calderoni & A. Barbieri, 2006. Nitrogen budget of a subalpine lake in North-Western Italy: the role of atmospheric input in the upward trend of nitrogen concentrations. Verhandlungen des Internationalen Verein Limnologie 29: 2027–2030.
Rogora, M., R. Mosello, L. Kamburska, N. Salmaso, L. Cerasino, B. Leoni, L. Garibaldi, V. Soler, F. Lepori, L. Colombo & F. Buzzi, 2015. Recent trends in chloride and sodium concentrations in the deep subalpine lakes (Northern Italy). Environmental Science and Pollution Research 22: 19013–19026.
Rogora, M., F. Buzzi, C. Dresti, B. Leoni, F. Lepori, R. Mosello, M. Patelli & N. Salmaso, 2018. Climatic effects on vertical mixing and deep-water oxygen content in the subalpine lakes in Italy. Hydrobiologia 824: 33–50.
Särkkä, J., 1996. Meiofauna of the profundal zone of the northern part of Lake Ladoga as an indicator of pollution. In: Simola, H., M. Viljanen, T. Slepukhina, & R. Murthy (eds), The First International Lake Ladoga Symposium. Developments in Hydrobiology, vol. 113. Springer, Dordrecht.
Sarvala, J., 1979. Effect of temperature on the duration of egg, nauplius and copepodite development of some freshwater benthic Copepoda. Freshwater Biology 9: 515–534.
Sarvala, J., 1986. Patterns of benthic copepod assemblages in an oligotrophic lake. Annales Zoologici Fennici 23: 101–130.
Sarvala, J., 1998. Ecology and role of benthic copepods in northern lakes. Journal of Marine Systems 15: 75–86.
Scheifhacken, N., C. Fiek & K. O. Rothhaupt, 2007. Complex spatial and temporal patterns of littoral benthic communities interacting with water level fluctuations and wind exposure in the littoral zone of a large lake. Fundamental and Applied Limnology 169: 115–129.
Schmid, P. E. & J. M. Schmid-Araya, 2007. Body size and scale invariance: multifractals in invertebrate communities. In Hildrew, A. G., D. G. Raffaelli & R. Edmonds-Brown (eds), Body Size: The Structure and Function of Aquatic Ecosystems Cambridge University Press, Cambridge: 140–166.
Sodré, E. D. O. & R. L. Bozelli, 2019. How planktonic microcrustaceans respond to environment and affect ecosystem: a functional trait perspective. International Aquatic Research 11: 207–223.
Špoljar, M., M. S. Perić, H. Wang, C. Zhang, N. Kuczyńska-Kippen, J. Fressl & Z. Ercegovac, 2021. Does the size structure of the littoral community reflect water level fluctuations in shallow waterbodies? Ecological Indicators 132: 108330.
Sterner, R. W., B. Keeler, S. Polasky, R. Poudel, K. Rhude & M. Rogers, 2020. Ecosystem services of Earth’s largest freshwater lakes. Ecosystem Service 41: 101046.
Stoch, F. & D. M. P. Galassi, 2010. Stygobiotic crustacean species richness: a question of numbers, a matter of scale. In Naselli-Flores, L. & G. Rossetti (eds), Fifty Years after the “Homage to Santa Rosalia”: Old and New Paradigms on Biodiversity in Aquatic Ecosystems. Developments in Hydrobiology 213, vol 213. Springer, Dordrecht..
Tõnno, I., H. Agasild, T. Kõiv, R. Freiberg, P. Nõges & T. Nõges, 2016. Algal diet of small-bodied crustacean zooplankton in a cyanobacteria-dominated eutrophic lake. PLoS ONE 11: e0154526.
Traunspurger, W., B. Wilden & N. Majdi, 2020. An overview of meiofaunal and nematode distribution patterns in lake ecosystems differing in their trophic state. Hydrobiologia 847: 2665–2679.
Vakkilainen, K., T. Kairesalo, J. Hietala, D. M. Balayla, E. Bécares, W. J. Van de Bund, E. Van Donk, M. Fernández-Aláez, M. Gyllström, L.-A. Hansson, M. R. Miracle, B. Moss, S. Romo, J. Rueda & D. Stephen, 2004. Response of zooplankton to nutrient enrichment and fish in shallow lakes: a pan-European mesocosm experiment. Freshwater Biology 49: 1619–1632.
Vermaat, J. E., V. Biberdžić, V. Braho, B. B. Gjoreska, M. Cara, Z. Dana, et al., 2022. Relating environmental pressures to littoral biological water quality indicators in Western Balkan lakes: can we fill the largest gaps? Science of the Total Environment 804: 150160.
Violle, C., M. L. Navas, D. Vile, E. Kazakou, C. Fortunel, I. Hummel & E. Garnier, 2007. Let the concept of trait be functional! Oikos 116: 882–892.
Wang, F. S., Y. X. Yu, C. Q. Liu, B. L. Wang, Y. C. Wang, J. Guan & H. Y. Mei, 2010. Dissolved silicate retention and transport in cascade reservoirs in Karst area, Southwest China. Science of the Total Environment 408(7): 1667–1675.
Wantzen, K. M., K. O. Rothhaupt, M. Mörtl, M. Cantonati, G. László & P. Fischer, 2008. Ecological effects of water-level fluctuations in lakes: an urgent issue. Hydrobiologia 613: 1–4.
Witthöft-Mühlmann, A., W. Traunspurger & K. O. Rothhaupt, 2007. Combined influence of river discharge and wind on littoral nematode communities of a river mouth area of Lake Constance. Aquatic Ecology 41: 231–242.
Zhong, S., Y. Geng, Y. Qian, C. Wei & P. Hengyu, 2019. Analyzing ecosystem services of freshwater lakes and their driving forces: the case of Erhai Lake, China. Environmental Science and Pollution Research 26: 10219–10229.
Zohary, T. & I. Ostrovsky, 2011. Ecological impacts of excessive water level fluctuations in stratified freshwater lakes. Inland Waters 1: 47–59.
Acknowledgements
We thank Lyudmila Kamburska, Silvia Zaupa and Daniele Paganelli (CNR-IRSA) for the cooperation in the fieldwork and the staff of the CNR-IRSA Hydrochemical Lab performing the chemical analyses. In addition, we are indebted to four anonymous reviewers who provided helpful information to improve the manuscript. We are grateful to Editor Eric R. Larson for edits of English language grammar and style, which highly improved the readability of this paper.
Funding
This research was funded by INTERREG Project V-A ITALIA-SVIZZERA 2014–2020, Parchi Verbano Ticino, ID 481668 and MUR-FOE-PROJECT Capitale Naturale – Task Biodiversità.
Author information
Authors and Affiliations
Contributions
MC and TDL contributed to conceptualisation and formal analysis; MC, AB and TDL contributed to methodology; TDL, DMPG, BF and AM contributed to validation; MC, AB, MR, MC and TDL contributed to investigation; AB and TDL contributed to resources, project administration and funding acquisition; MC, AB, MR, MC, AM, DMPG, BF and TDL contributed to data curation; MC contributed to writing—original draft preparation; and TDL contributed to writing—review and editing. .
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical approval
The research complies with ethical standards.
Informed consent
I can confirm that the manuscript has been read and approved by all named authors and that there are no other persons who satisfied the criteria for authorship but are not listed. I further confirm that the order of authors listed in the manuscript has been approved by all of us.
Additional information
Handling Editor: Eric R. Larson
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Cifoni, M., Boggero, A., Rogora, M. et al. Effects of human-induced water level fluctuations on copepod assemblages of the littoral zone of Lake Maggiore. Hydrobiologia 849, 3545–3564 (2022). https://doi.org/10.1007/s10750-022-04960-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-022-04960-3