
Abstract
This study deals mainly with the biostratigraphy, paleoecology and paleobiogeogrphy of the Middle Eocene benthic foraminifera from an outcrop exposed at Qattamiya area, Cairo–Suez district, Egypt. This district is covered by relatively thick Eocene sequences composed of limestone intercalated with shales and marl. Stratigraphically, the oldest rock unit exposed in the study area is the Observatory Formation which is followed by the Qurn Formation. The examination of the collected samples led to the recognition of 27 species belonging to 17 genera and 10 families. Three benthic foraminiferal biozones are recognized Elphidium trompi-Quinqueloculina carinata Assemblage Zone, Bulimina jacksonensis Zone, and Quinqueloculina ludwigi Zone. These local biozones are correlated with their equivalents inside Egypt. Therefore, the age is assigned to the Middle Eocene. To detect the paleoecology of the recorded foraminiferal community, some parameters (Foraminiferal abundance, richness, and the percentages of epifaunal and infaunal species) are calculated. Three local paleoecologic ecozones are proposed signifying a hyper-to hyposaline inner neritic environment for the lower part of the Observatory, the middle shelf with low oxygen conditions for the upper part of the Observatory Formation, and hypersaline inner neritic environment for the Qurn Formation. Recently, multivariate analyses are considered as a valuable tool in establishing the paleoecology and paleobiogeographical provinces of the Paleogene microfossils. In this study, a matrix composed of 18 species from six countries (Tunisia, Egypt, Libya, France, Spain and Italy) is suggested for multivariate analyses. This matrix is subjected to the principal component analysis (PCA) and the Q-mode cluster analysis to detect the possible provinces. Consequently, two significant provinces are detected, the Southern Tethyan Province (Tunisia, Libya and Egypt) and the Western Tethyan Province (Spain, Italy, and France). Based on the similarity index, there is a strong similarity between the two provinces, which proposes a marine connection between them during Eocene.
Similar content being viewed by others

Introduction
Benthic foraminifera were broadly used for reconstruction of paleoecological parameters, comprising depth, salinity, temperature, and oxygen content (van der Zwaan et al. 1999; Gooday 2003; Murray, 2006; Boscolo Galazzo et al. 2013; Ostad-Ali-Askari et al. 2020). Several investigations were done on the paleoenvironment, taxonomy, and biostratigraphy of the Middle Eocene foraminifera from Egypt (e.g., El Dawy, 1997; Shahin, 2000; Elewa, 2004; Nassif and Korin, 2018; Abd El-Gaied, 2019; Farouk et al. 2020).
El Dawy (1997) subdivided the Middle Eocene outcrops in the Nile Valley into three biozones Uvigerina nakkadyi-Anomalinoides fayoumensis Zone, Brizaline cookie Zone, and Nonion scaphum-Pararotalia audouini Zone. Shahin (2000) subdivided the Middle Eocene rocks in western Sinai into Norcottia danvillensis-Alistoma aegyptiaca Zone and Uvigerina rippensis-Uvigerina churchi Zone. Ali et al. (2011) dealt with the taxonomy of Eocene foraminifera from the Nile Valley and identified 128 species. Nassif and Korin (2018) subdivided the outcrops in the North Eastern Desert into three subzones, Quinqueloculina carinata, Uvigerina rippensis, and Textularia adalta. Moreover, Abd El-Gaied et al. (2019) subdivided the sequences at Cairo-Helwan area into five biozones. Farouk et al. (2020) concentrated on the evaluation of the Middle Eocene foraminifera at Wadi Nukhul, Sinai, to detect the fluctuations in paleoproductivity and paleobathymetry.
This paper concentrated on the Middle Eocene small benthic foraminifera from Qattamiya area, Cairo–Suez district, Egypt. It represents an attempt to identify the relationship between the Egyptian benthic foraminifera and those from the southern Tethys (Libya, Tunisia) from one side, and those from the western Tethys (Spain, Italy, and France) from the other side. Recently, multivariate analyses represent an essential tool in detecting the paleobiogeographical provinces of the microfossils (e.g., Elewa and Mohamed, 2014; Elewa, 2018; El Baz, 2019; Shahin and El Baz, 2021). Therefore, this work is carried out using multivariate analyses (principal component analysis, Q-mode cluster analysis and Jaccard index). The examined foraminifera include Lagena sulcata, L. striata, Nuttalides truempyi, Marginulinopsis tuberculata, Gaudryina pyramidata, Cibicidoides beadnelli, C. eocaenus, C. libycus, Bulimina jacksonensis, Orthokarstenia nakkadyi, Uvigerina mexicana, Cancris subconicus, Bolivina carinata, Spiroplectammina carinata, Globulina gibba, Cibicides lobatulus, Oridorsalis umbonatus, and Lenticulina cultrata. Also, this work aims to establish the benthic foraminiferal biozones and to detect the paleoenvironment.
Geological setting
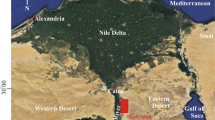
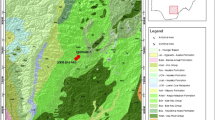
The considered outcrop occurs in the Qattamiya area (Fig. 1), Cairo-Suez district (latitude 29° 53′ 56″ N and longitude 31° 24′ 13″ E). This district is covered by relatively thick Eocene sequences composed of limestones intercalated with shales and marl. The study area was subjected to the stresses resulted from the collision between European and African plates during Lutetian time (Issawi et al. 1999). This tectonic event produced variable sedimentary environments, each with characteristic rock units and benthic foraminiferal assemblages.

Location and geologic maps (modified after the Geological Survey of Egypt, 2014)
Material and methods
Thirty-two rock samples were collected to examine their foraminiferal content. Approximately 100 g of dry rock sample was saturated with a 5% H2O2 solution, washed through a 63-μm mesh sieve, and then dried and sieved. About 1 g of each washed residue was examined. The picked individuals were identified and classified following Loeblich and Tappan (1988). Some selected foraminiferal species were photographed by the scanning electronic microscope (Fig. 2). The proposed biozones are correlated with their equivalents in Egypt. Furthermore, the paleoecological conditions that prevailed during the Middle Eocene depended mainly on the characters of the recorded assemblage, such as abundance, richness, and percentages of Rotaliina, Textulariina, and Miliolina. Also, the percentages of epifaunal and infaunal species were calculated to deduce the oxygen content. In this work, multivariate analyses were used to detect the paleobiogeographic provinces. The principal component analysis and Q-mode cluster analysis (based on Ward’s method) were applied on a matrix composed of 19 foraminiferal species from six countries (Tunisia, Egypt, Libya, France, Spain, and Italy). Furthermore, the similarity index (Jaccard index) was calculated to identify the relation between these regions.

Vertical distribution of the identified species
Results and discussion
Lithostratigraphy
The whole Middle Eocene succession is subdivided into the Observatory Formation (at the base) and the Qurn Formation (at the top).
Observatory Formation
This rock unit was firstly described by Farag and Ismail (1959). It was subdivided into (from base to top) the Upper Building Stone and Guishi members. In this study, it is overlain conformably by the Qurn Formation. The base of this rock unit is unexposed. It is composed of hard fossiliferous limestone and marly limestone. The thickness reaches 32 m. The identified benthic foraminiferal assemblage includes Eponides ellisorae, Neoeponides schreibersi, Cancris subconicus, Lobatula lobatulus, Baggina brady, Textularia agglutinans, Nonionella spissa, Nonionella auris, Nonion scaphum, Idalina cuvillieri, Triloculina gibba, Triloculina trigonula, Pyrgo elongata, Bolivina carinata, Brizalina cookei, Uvigerina jacksonensis, Uvigerina rippensis, Uvigerina multistriata, and Uvigerina mediterranea. The occurrence of larger benthic (Nummulites spp.) is restricted to some levels.
The documented macrofossils include small branched corals, Vulsella sp., and gastropod moulds. Based on the studies of Shahin et al. (2007), Nassif and Korin (2018), Sallam et al. (2018), and Abd El-Gaied et al. (2019), the Observatory Formation is assigned to the Middle Eocene (Lutetian).
Qurn Formation
This rock unit was originally described by Farag and Ismail (1959). The upper part of this formation is unexposed. The exposed part is 20 m thick, consisting mainly of hard limestone and dolomitic limestone. Benthic foraminiferal assemblages of this unit are relatively low and include Quinqueloculina ludwigi Zone, lobatula lobatulus, Elphidium trompi, Triloculina gibba, Triloculina trigonula, Pyrgo elongata, and Textularia agglutinans. In some intervals, some moulds of bivalves and gastropods occur. Based on the studies of Shahin et al. (2007), Nassif and Korin (2018), Sallam et al. (2018), and Abd El-Gaied et al. (2019), the Qurn Formation is assigned to the Middle Eocene (Bartonian).
Systematic of foraminifera
According to the classification of Loeblich and Tappan (1988), 27 benthic foraminiferal species are identified. Also, the stratigraphic distribution of each species is briefly discussed.
Order: Foraminiferida Eichwald, 1830
Suborder: Textulariina Delage et Herouard, 1896
Superfamily: Textulariacea Ehrenberg, 1838
Family: Textulariidae Ehrenberg, 1838
Subfamily: Textulariinae Ehrenberg, 1838
Genus: Textularia Defrance, 1824
Textularia agglutinans d’Orbigny, 1839
(Pl. 1, Fig. 1 )
1839 Textularia agglutinans d’Orbigny, pl. 1, figs. 17, 18, 32–34.
1963 Textularia agglutinans d’Orbigny-Said and Metwalli, pl. 1, Fig. 13.
Occurrence: Obervatory Formation.
Stratigraphic distribution: Eocene of Belgium (Kaasschieter, 1961) and Middle Eocene of Egypt (e.g., Shahin et al. 2007).
Suborder: Miliolina Delage et Hérouard, 1896
Superfamily: Miliolacea Ehrenberg, 1839
Family: Hauerinidae Schwager, 1876
Subfamily: Hauerininae Schwager, 1876
Genus: Quinqueloculina d’Orbigny, 1826
Quinqueloculina carinata d’Orbigny, 1850
(Pl. 1, Fig. 2)
1990 Quinqueloculina carinata d’Orbigny-Helal, pl. 1, figs. 9–13.
2019 Quinqueloculina carinata d’Orbigny-Abd El-Gaied et al., Fig. 4.11.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of France (Le Clavez, 1949) and Egypt (Shahin et al. 2007; Nassif and Korin, 2018), Middle to Late Eocene of Egypt (Abu Ellil. 2004; Abd El-Gaied et al. 2019; Hassan and Korin, 2019), and Late Eocene of Egypt (Helal, 1990).
Quinqueloculina ludwigi Reuss, 1866
(Pl. 1, Fig. 3)
1866 Quinqueloculina ludwigi Reuss, p. 126, pl. 1, Fig. 12.
1970 Quinqueloculina ludwigi Reuss-Boukhary, pl. 15, Fig. 3.
2019 Quinqueloculina ludwigi Reuss-Abd El-Gaied et al., Fig. 4.14.
2021 Quinqueloculina ludwigi Reuss-Ramadan et al., Fig. 6.26.
Occurrence: Qurn Formation.
Stratigraphic distribution: In Egypt, it was reported from the Middle Eocene (Boukhary, 1970; Nassif and Korin, 2018), the Early Eocene (Shamah and Helal, 1993), the Middle-Late Eocene (Abd El-Gaied et al. 2019; Hassan and Korin, 2019), and Late Eocene (Ramadan et al. 2021). In Italy, it was reported from the Late Eocene (Barbin and Keller-Grünig, 1991).
Subfamily: Miliolinellinae Vella, 1957
Genus: Pyrgo Defrance, 1824
Pyrgo elongata (d’Orbigny, 1850)
(Pl. 1, Fig. 4)
1850 Biloculina elongata d’Orbigny, p. 298, pl. 16, figs. 5,6 and 7.
2019 Pyrgo elongata (d’Orbigny) -Abd El-Gaied et al., Fig. 4.17.
Occurrence: Observatory and Qurn formations.
Stratigraphic distribution: Eocene of Belgium (Kaasschieter, 1961), Middle Eocene of Egypt (Abu Ellil, 2004; Shahin et al. 2007; Nassif and Korin, 2018), and Late Eocene of Egypt (Helal, 1990; Shamah and Helal, 1993).
Genus: Idalina Schlumberger and Munier-Chalmas, 1884
Idalina cuvillieri Bignot, 1992
2004 Idalina cuvillieri Bignot-Abu Ellil, p. 60, pl. 6, figs. 5, 6, 8; pl. 14, figs. 1–8; pl. 18, Fig. 8.
2019 Idalina cuvillieri Bignot-Hassan and Korin, Fig. 11.9.
Occurrence: Observatory and Qurn Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Strougo et al.1992; Abu Ellil, 2004; Hassan and Korin, 2019).
Genus: Triloculina d’Orbigny, 1826
Triloculina gibba d’Orbigny, 1846
1846 Triloculina gibba d’Orbigny, p. 274, pl. 16, figs. 22–24.
1993 Triloculina gibba d’Orbigny-Shamah & Helal, p. 253, pl. 2, Fig. 15.
Occurrence: Observatory and Qurn formations.
Stratigraphic distribution: Middle Eocene of France (Le Calvez, 1949) and Egypt (Helal, 1990), and the Middle-Upper Eocene of Egypt (Abu Ellil, 2004).
Triloculina trigonula (Lamarck, 1804)
(Pl. 1, Fig. 5 )
1961 Triloculina trigonula (Lamarck)-Kaasschieter, p. 165, pl. 5, figs. 8 -11.
2019 Triloculina trigonula (Lamarck)-Abd El-Gaied, et al., Fig. 4.20.
Occurrence: Observatory and Qurn formations.
Stratigraphic distribution: Eocene of Belgium (Kaasschieter, 1961) and Middle Eocene of Egypt (El Dawy, 1997; Shahin et al. 2007).
Suborder: Rotaliina Delage et Hérouard, 1896
Superfamily: Bolivinacea Glaessner, 1937
Family: Bolivinidae Glaessner, 1937
Genus: Bolivina d’Orbigny, 1839
Bolivina carinata Terquem, 1882
(Pl. 1, Fig. 6)
1882 Bolivina carinata Terquem, p. 149, pl. 15, Fig. 19.
1970 Bolivina carinata Terquem-Boukhary, p. 142, pl. 19, Fig. 15.
2019 Bolivina carinata Terquem-Abd El-Gaied et al., Fig. 4.38.
2021 Bolivina carinata Terquem-Ramadan et al., Fig. 7.20.
Occurrence: Observatory Formation.
Stratigraphic distribution: Eocene of France (Terquem, 1882) and south Atlantic Ocean (Müller-Merz and Oberhänsli, 1991), Middle-Late Eocene of Egypt (Boukhary, 1970), the Middle Eocene of Egypt (El Dawy, 1997) and Early-Middle Eocene of Libya (Abd El-Gaied and Abd El-Aziz, 2020) and Spain (Ortiz and Thomas, 2006), and Late Eocene of Egypt (Ramadan et al. 2021).
Genuus: Brizalina Costa, 1826
Brizalina cookei (Cushman, 1922)
1922 Bolivina cookei Cushman, p. 126, pl. 8, figs. 25–26.
1997 Brizalina cookei (Cushman)-El Dawy, pl. 2, figs. 7–8.
2002 Brizalina cookei (Cushman)-Helal, pl. 2, Fig. 10.
2019 Brizalina cookei (Cushman)-Abd El-Gaied, et al., figs. 4.36–4.37.
2021 Brizalina cookei (Cushman) -Ramadan et al., Fig. 7.22.
Occurrence: Observatory Formation.
Stratigraphic distribution: Eocene of Belgium (Kaasschieter, 1961), Middle Eocene of Egypt (Helal, 2002; Hassan and Korin, 2019), and Late Eocene of Egypt (Shahin, 2000; Ramadan et al. 2021).
Superfamily: Buliminacea Jones, 1875
Family: Buliminidae Jones, 1875
Genus: Bulimina d’Orbigny, 1826
Bulimina elongata d’Orbigny, 1826
(Pl. 1, Fig. 7)
1826. Bulimina elongata d’Orbigny, p. 269.
2019 Bulimina elongata d’Orbigny-Hassan and Korin, Fig. 11.5.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Hassan and Korin, 2019).
Bulimina jacksonensis Cushman, 1925
(Pl. 1, Fig. 8)
1925 Bulimina jacksonensis Cushman, p. 6, pl. 1, figs. 6 and 7.
1970 Bulimina jacksonensis Cushman-Boukhary, pl. 18, Fig. 17.
1992 Bulimina jacksonensis Cushman-Hussein, figs. 2.10–2.11.
2021 Bulimina jacksonensis Cushmans-Ramadan et al., figs. 7.23–7.24.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Libya (Barr and Berggren, 1980), Tunisia (Amami-Hamdi et al. 2014) and Egypt (Elewa, 2004; Farouk et al. 2020), the Middle-Late Eocene of Egypt (Fahmy, 1975; Hassan and Korin, 2019), and the Late Eocene of Egypt (Shahin, 2000; Ramadan et al. 2021).
Family: Uvigerinidae Haeckel, 1894
Subfamily: Uvigerininae Haeckel, 1894
Genus: Uvigerina d’Orbigny, 1826
Uvigerina compressa Ansary, 1955
1955 Uvigerina mediterranea Hofker var. compressa Ansary-Ansary, p. 96,
pl. 3, Fig. 17.
1994 Uvigerina compressa Ansary-Anan, Fig. 9.1.
Occurrence: Observatory Formation.
Stratigraphic distribution: Late Eocene of Egypt (Ansary, 1955) and Middle-Late Eocene of Egypt (Anan, 1994).
Uvigerina jacksonensis Cushman, 1925
(Pl. 1, Fig. 9)
1925 Uvigerina jacksonensis Cushman, pl. 10, Fig. 13.
2000 Uvigerina jacksonensis Cushman-Abul Nasr, Fig. 12.6.
2019 Uvigerina jacksonensis Cushman-Hassan and Korin, Fig. 11.22.
2021 Uvigerina jacksonensis Cushman-Ramadan et al., Fig. 7.28.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Elewa, 2004; Hassan and Korin, 2019) and Late Eocene of Egypt (Abul Nasr, 2000; Ramadan et al. 2021).
Uvigerina mediterranea Hofker, 1932
1932 Uvigerina mediterranea Hofker, p. 119, tf. 32.
1970 Uvigerina mediterranea Hofker-Boukhary, pl. 19, Fig. 16.
2021 Uvigerina mediterranea Hofker-Ramadan et al., Fig. 7.29.
Occurrence: Observatory Formation.
Stratigraphic distribution: Late Eocene of Egypt (Ansary, 1955; Ramadan et al. 2021), Middle Eocene of Egypt (Boukhary, 1970).
Uvigerina multistriata Hantken, 1871
1983 Uvigerina multistriata Hantken-Sztrakos, pl. 1, figs. 7–13.
2000 Uvigerina multistriata Hantken-Abul Nasr, figs. 12.1–12.3.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Abul Nasr, 2000; Shahin et al. 2007).
Uvigerina rippensis Cole, 1927
1927 Uvigerina rippensis Cole, pl. 2, Fig. 16.
2004 Uvigerina rippensis Cole-Elewa, pl. 1, Fig. 2.
2019 Uvigerina rippensis Cole-Hassan and Korin, Fig. 11.23.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Libya (Barr and Berggren, 1980) and Egypt (Hussein, 1994; Elewa, 2004; Nassif and Korin, 2018), and Middle-Late Eocene of Egypt (Anan, 1994), Late Eocene of Egypt (Shahin, 2000).
Superfamily: Discorbacea Ehrenberg, 1838
Family: Bagginidae Cushman, 1927
Subfamily: Baggininae Cushman, 1927
Genus: Baggina Cushman, 1926
Baggina bradyi (Brotzen, 1936)
1936 Valvulineria bradyi Brotzen, p. 154.
2019 Baggina bradyi (Brotzen)-Abd El-Gaied, et al., Fig. 5.6.
Occurrence: Observatory Formation.
Stratigraphic distribution: In Egypt, it was recorded from the Late Eocene (Ansary, 1955), Middle Eocene (Abd El-Gaied, et al. 2019), and Middle-Late Eocene (Anan, 1994).
Genus: Cancris De Montfort, 1808
Cancris subconicus (Terquem, 1882)
(Pl. 1, Fig. 10)
1882 Rotalina subconicus Terquem, pl. 4, Fig. 5.
2002 Cancris subconicus (Terquem)-Helal, pl. 3, Fig. 11.
2019 Cancris subconicus (Terquem)-Abd El-Gaied, et al., Fig. 5.9.
Occurrence: Observatory Formation.
Stratigraphic distribution: Eocene of France (Terquem, 1882), the Middle Eocene of Egypt (Boukhary, 1970; Abdel Ghany, 1990), the Late Eocene of Italy (Barbin and Keller-Grünig, 1991), and the Middle-Late Eocene of Egypt (Anan, 1994; Abd El-Gaied, et al. 2019).
Family: Eponididae Hofker, 1951
Subfamily: Eponidinae Hofker, 1951
Genus: Eponides De Montfort, 1808
Eponides ellisorae Garrett, 1939
1939 Eponides ellisorae Garrett, pl. 66, figs. 6–8.
1997 Eponides ellisorae Garrett-El Dawy, p. 435.
Occurrence: Observatory Formation.
Stratigraphic distribution: In Egypt, it was reported from the Late Eocene (Helal, 2002), Middle Eocene (El Dawy, 1997), and Middle-Late Eocene (Anan, 1994).
Genus: Neoeponides Reiss, 1960
Neoeponides schreibersi (d’Orbigny, 1846)
1846 Rotalina schreibersi d’Orbigny, pl. 8, figs. 4–6.
1985 Eponides schreibersi (d’Orbigny)-Abd Elshafy et al., pl. 3, Fig. 16.
2021 Neoeponides schreibersi (d’Orbigny)-Ramadan et al., figs. 8.12–8.14.
Occurrence: Observatory Formation.
Stratigraphic distribution: In Egypt, it was reported from the Middle Eocene (El Dawoody, 1971), Middle-Late Eocene (Abd El-Gaied et al. 2019), and Late Eocene (Ramadan et al. 2021).
Family: Cibicididae Cushman, 1927
Subfamily: Cibicinae Cushman, 1927
Genus: Lobatula Fleming, 1828
Lobatula lobatulus (Walther and Jacob, 1798)
1798 Nautilus lobatulus Walther and Jacob, pl. 14, Fig. 36.
1963 Cibicides lobatulus (Walther and Jacob)-Said and Metwalli, pl. 3, Fig. 5.
2019 Lobatula lobatulus (Walker and Jacob)-Abd El-Gaied et al., Fig. 5.24.
2021 Lobatula lobatulus (Walker and Jacob)-Ramadan et al., Fig. 8.22.
Occurrence: Observatory and Qurn formations.
Stratigraphic distribution: Eocene of France (Le Clavez, 1949) and Egypt (Ansary, 1955), the Late Eocene of Italy and (Barbin and Keller-Grünig, 1991) Egypt (Ramadan et al. 2021), and the Middle Eocene of Egypt (Shahin et al. 2007).
Superfamily: Nonionacea Schultze, 1854
Family: Nonionidae Schultze, 1854
Subfamily: Nonioninae Schultze, 1854
Genus: Nonion De Montfort, 1808
Nonion acutidorsatum ten Dam, 1944
1944 Nonion acutidorsatum ten Dam, p. 108, pl. 3, Fig. 19.
1994 Nonion acutidorsatum ten Dam-Anan, p. 226.
Occurrence: Observatory Formation.
Stratigraphic distribution: Late Eocene of Egypt (Ansary, 1955), Middle Eocene of Egypt (Anan, 1994; Shahin et al. 2007), and Eocene of Neatherland (Ten Dam, 1944).
Nonion scaphum (Fichtel and Moll, 1798)
(Pl. 1, Fig. 11)
1798 Nautilus scaphum Fichtel and Moll, p. 105, pl. 19, figs. e, f.
2019 Nonion scaphum (Fichtel and Moll)-Abd El-Gaied et al., Fig. 5.33.
2021 Nonion scaphum (Fichtel and Moll)-Ramadan et al., Fig. 8.25.
Occurrence: Observatory Formation.
Stratigraphic distribution: In Egypt, it was reported from the Late Eocene (Anan, 1994), Middle Eocene (Shamah and Helal, 1993), and Middle-Late Eocene (Boukhary, 1970).
Genus: Nonionella Cushman, 1926
Nonionella auris (d’Orbigny, 1839)
1839 Valvulina auris d’Orbigny, p. 47, pl. 2, figs. 15–17.
1990 Nonionella auris (d’Orbigny)-Abdel Ghany, p. 86, pl. 3, figs. 11 a, b.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Shahin et al. 2007), and Late Eocene of Egypt (Abdel Ghany, 1990).
Nonionella spissa Cushman, 1931
1961 Nonionella spissa Cushman-Kaasschieter, p. 206, pl. 11, figs. 7–8.
2019 Nonionella spissa Cushman-Abd El-Gaied et al., Fig. 5.37.
Occurrence: Observatory Formation.
Stratigraphic distribution: Eocene of Belgium (Kaasschieter, 1961), Middle Eocene of Egypt (Shamah and Helal, 1993; Abd El-Gaied et al. 2019).
Superfamily: Rotaiiacea Ehrenberg, 1839
Family: Elphidiidae Galloway, 1933
Subfamily: Elphidiinae Galloway, 1933
Genus: Elphidium De Montfort, 1808
Elphidium ancestrum Le Calvez, 1950
2019 Elphidium ancestrum Le Calvez-Abd El-Gaied et al., Fig. 6.10.
2021 Elphidium ancestrum Le Calvez-Ramadan et al., Fig. 8.34.
Occurrence: Observatory and Qurn formations.
Stratigraphic distribution: Middle Late Eocene of Egypt (Abd El-Gaied et al. 2019), and Late Eocene of Egypt (Ramadan et al. 2021).
Elphidium trompi (Ansary, 1955 )
(Pl. 1, Fig. 12)
1955 Nonion trompi Ansary, p. 73, pl. 2, Fig. 25.
1975 Elphidium trompi (Ansary)-Fahmy, p. 313, pl. 2, Fig. 14.
Occurrence: Observatory Formation.
Stratigraphic distribution: Middle Eocene of Egypt (Shahin et al. 2007), and Late Eocene of Egypt (Ansary, 1955; Fahmy, 1975).
Biostratigraphy
Based on the stratigraphical distribution of the documented foraminifera, three biozones are established and correlated with their equals in Egypt (Table 1). The proposed biozones are arranged from base to top as follows:
Elphidium trompi-Quinqueloculina carinata Assemblage Zone
Definition: The base of this zone is not exposed, while the upper boundary is placed at the first existence of Bulimina jacksonensis. The thickness reaches 12 m and occupies the lower part of the Observatory Formation.
Characteristic species: Besides the proposed species, this assemblage comprises Eponides ellisorae, Neoeponides schreibersi, Cancris subconicus, Lobatula lobatulus, Baggina brady, Nonionella spissa, Idalina cuvillieri Triloculina gibba, Pyrgo elongata, Nonionella auris, and Triloculina trigonula (Fig. 3).

Statistics of the foraminiferal community in the Qattamia section
Equivalents and age: This zone is equivalent to the Nummulites gizehensis Zone of Mansour et al. (1982), the base of the Nummulites cf. syrticus Zone of Strougo et al. (1992), the lower part of the Uvigerina nakkadyi-Anomalinoides fayoumensis Zone of El Dawy (1997), the base of the Bulimina jacksonensis-Uvigerina jacksonensis Zone of Elewa et al. (1998), the base of Nummulites aff. puchellas Zone of Boukhary et al. (2002), the base of the Globigerinatheka kugleri-Morozovella aragonensis Zone of Hassan and Korin (2019), the lower part of Dictyoconus aegyptiensis Zone of El Baz (2019), and both the Elphidium cherifi, and the base of the Quinqueloculina seminula zones of Abd El-Gaied, et al. (2019). Moreover, Uvigerina rippensis, U. jacksonensis, and Nonion scaphum are distinctive taxa for the Middle Eocene (Elewa et al. 1998; Shahin et al. 2007). Also, Strougo (1979) suggested that the top of the Upper Building Stone Member marks the end of Lutetian. Accordingly, this zone belongs to the Lutetian age.
Bulimina jacksonensis Interval Zone
Definition: It is defined as an interval from the first existence of Bulimina jacksonensis to the first existence of Quinqueloculina ludwigi. The thickness reaches 17 m and occupies the top of the Observatory Formation.
Characteristic species: Bolivina carinata, Brizalina cookei, Uvigerina jacksonensis, Uvigerina rippensis, Textularia agglutinans, Nonionella spissa, N. auris, Nonion scaphum, Uvigerina multistriata, and Uvigerina mediterranea.
Equivalents and age: This zone is equal to the Orbitolites complanatus Zone of Mansour et al. (1982), the Morozovella lehneri Zone of Allam et al. (1991), both the upper part of the Uvigerina nakkadyi-Anomalinoides fayoumensis and the Brizalina cooki zones of El Dawy (1997), the top of the Bulimina jacksonensis-Uvigerina jacksonensis Zone of Elewa et al. (1998), the upper part of Globigerinatheka kugleri-Morozovella aragonensis Zone of Hassan and Korin (2019), and the upper part of the Quinqueloculina seminula and the Nonionella africana Zones of Abd El-Gaied et al. (2019). Accordingly, this zone belongs to Late Lutetian.
Quinqueloculina ludwigi Zone
Definition: It is defined from the first occurrence of Quinqueloculina ludwigi, while the top is not exposed. It occupies the whole Qurn Formation and the thickness reaches 20 m.
Characteristic species: Besides the proposed species, this assemblage comprises lobatula lobatulus, Elphidium trompi, Triloculina gibba, Pyrgo elongata, Triloculina trigonula, and Textularia agglutinans.
Equivalents and age: This zone is equal to the Nummulites beaumonti Zone of Mansour et al. (1982), both the Fabularia schwagerinides and Idalina cuvillieri zones of Strougo et al. (1992), both the upper part of Brizalina cooki and Nonion scaphum-Pararotalia audouin zones of El Dawy (1997), the Uvigerina eocaena-Uvigerina steyeri Zone of Abul Nasr (2000), the Nummulites sp. cf. beaumonti/Nummulites aff. pulchellus Assemblage Zone of Boukhary et al. (2002), the Uvigerina mediterranea Zone of Shahin et al. (2007), the Palmula ansaryi Zone of Aly et al. (2011), the upper part of Uvigerina rippensis and the Textularia adalta subzones of Nassif and Korin (2018), and the Brizalina cooki and Nonion scaphum Zones of Abd El-Gaied et al. (2019). Moreover, Issawi et al. (1999) assigned the Qurn Formation to Bartonian age. Accordingly, this zone belongs to Bartonian.
Paleoecology
To detect the paleosalinity, paleobathymetry and the oxygen conditions, some foraminiferal parameters, including foraminiferal number, richness, the percentages of Rotaliina, Miliolina, Textulariina, and epifaunal and infaunal species, were calculated. In this study, the Middle Eocene rocks are classified into three ecozones, involving the base of the Observatory Formation, the top of the Observatory Formation, and the Qurn Formation.
Ecozone 1
Ecozone 1 represents the base of the Observatory Formation. It is characterized by a low foraminiferal number (17–198 tests per gram; average 74 tests) as shown in Table 2. The foraminiferal richness is also low (2–10 species). It is noted that the foraminiferal assemblage is dominated only by calcareous and epifaunal species (Fig. 4). Planktonic foraminifera and agglutinated foraminifera are completely absent. In order to detect the paleosalinity, the percentages of Rotaliina, Miliolina and Textulariina were calculated. Rotaliina is represented by the following genera Elphidium, Eponides, Cancris, Lobatula, Baggina, and Nonionella. The average percentage of Rotaliina reaches 80%. Miliolina (Triloculina gibba, Quinqueloculina carinata, Idalina cuvillieri, and Pyrgo elongata) reaches about 18%. The foraminiferal plots of this ecozone on the ternary diagram of Murray (1973) are placed on the line of Rotaliina-Miliolina close to the Rotaliina corner (Fig. 5), indicating hyper saline lagoonal conditions. Also, the occurrence of the following foraminiferal assemblage Triloculina, Quinqueloculina, Idalina, Pyrgo, and Elphidium indicates shallow marine environment (Phleger and Parker, 1951; Kaasschieter, 1961; Haynes, 1981). Moreover, the occurrence of Lobatula lobatulus and Cancris subconicus refers to inner-middle shelf (Murray et al. 1981). Furthermore, the presence of small branched corals reflects shallow marine environment (Wells, 1956). Therefore, the deposition of this ecozone occurred in a hyper-to hyposaline, inner neritic environment.

Foraminiferal plots of the studied ecozones on the ternary diagram of Murray (1973)
Ecozone 2
It represents the top of the Observatory Formation. It is characterized by a low foraminiferal number (16–225 tests per gram; average 71 tests). Also, the number of foraminifera species is low (2–13 species). This ecozone is dominated by calcareous species, while only one agglutinated species is recorded. Rotaliina is represented by the following genera Bolivina, Bulimina, and Uvigerina. The average percentage of Rotaliina reaches 96%. Textulariina includes only one species (Textularia agglutinans), and it represents about 3.5%. Miliolina (Triloculina and Quinqueloculina) reaches about 0.5%. The foraminiferal plots of this ecozone are placed on the Rotaliina corner (Fig. 5), indicating hyposaline lagoonal conditions. Moreover, to detect the oxygen content, the percentages of infaunal and epifaunal species are calculated. Infaunal species represents about 66.6%, while the epifaunal species represents 33.4%. Furthermore, the genera Bulimina, Bolivina, and Uvigerina indicate outer neritic environment with low oxygen conditions (e.g., Barr and Berggren, 1980; Miller and Lohmann 1982). The genus Textularia indicates deep inner shelf ((Boersma, 1984). Nonion extends from inner to outer shelf (Murray, 2006). Therefore, the deposition of this ecozone occurred in the middle shelf with low oxygen conditions.
Ecozone 3
This ecozone represents the whole the Qurn Formation. It is characterized by a low foraminiferal number (8–117 tests per gram; average 34 tests). Also, a low number foraminiferal species is recorded (2–9 species). This ecozone is dominated by calcareous species, whereas only one agglutinated species is recorded. Planktonic foraminifera are completely absent. The average percentage of epifaunal species reaches 97.8%, while the average percentage of infaunal species reaches 2.2%. Miliolina (Triloculina, Quinqueloculina and Pyrgo) reaches about 71.2%. Rotaliina reaches 26.6%, and is represented by the following genera Elphidium, Bulimina, and Cibicides. Textulariina represents 2.2% and includes only one species (Textularia agglutinans). The foraminiferal plots of this ecozone are placed on the line of Rotaliina-Miliolina near the Miliolina corner (Fig. 5), indicating hyper saline lagoonal conditions. The dominance of Miliolina indicates hypersaline environment and water depths extending from 12 to 18 m (e.g., Parker and Gischler, 2015). Also, Elphidium indicates very shallow environment (Murray, 2006). Therefore, the deposition of this ecozone occurred in a hypersaline inner neritic environment.

1st principal component axis versus 2nd principal component axis of the studied 6 countries
Paleobiogeograpgy and multivariate analyses
Many of the identified foraminiferal species were previously documented from the Southern and Western Tethys areas (e.g., Le Calvez, 1949; Kaasschieter, 1961; Said and Metwalli, 1963; Barbin and Keller-Grünig, 1991; Ortiz and Thomas, 2006; Shahin et al. 2007; Amami-Hamdi et al. 2014; Farouk et al. 2020; Abd El-Gaied and Abd El-Aziz, 2020). The paleobiogeography of the studied foraminifera was done with the aid of multivariate analyses (the principal component analysis and Q-mode cluster analysis). Firstly, the principal component analysis was applied on the suggested matrix (Table 3) that consists of 18 foraminiferal species from six regions, including France, Spain, Italy, Tunisia, Egypt, and Libya.
The results of PCA are based on the first vector (34.17%) and the second vector (21.27%) as clarified in Table 4 and Fig. 6. It is noted that the examined countries could be classified into two provinces, the first one called the Southern Tethyan Province (STP), which includes Tunisia, Libya and Egypt, while the second province called the Western Tethyan Province (WTP) and includes Spain, France and Italy.

Dendrogram resulting from cluster analysis in Q-mode using the Ward's method
Furthermore, the Q-mode cluster analysis was applied on the same matrix. Therefore, the resulting dendrogram (Plate 1) shows the separation of two clusters (A and B) at a distance of about 5. Cluster A represents the STP and is characterized by a great similarity between the Egyptian foraminiferal assemblage and those from Libya (70%) and Tunisia (55%) as noted in Table 5. The common species between these regions are Lagena sulcata, Nuttalides truempyi, Marginulinopsis tuberculata, Cibicidoides libycus, Cibicidoides beadnelli, Bulimina jacksonensis, and Uvigerina mexicana.

1, Textularia agglutinans d’Orbigny, Observatory Formation; 2, Quinqueloculina carinata d’Orbigny, Observatory Formation; 3, Q. ludwigi Reuss, Qurn Formation; 4, Pyrgo elongata (d’Orbigny), Observatory Formation; 5, Triloculina trigonula (Lamarck), Observatory Formation; 6, Bolivina carinata (Terquem), Observatory Formation; 7, Bulimina elongata d’Orbigny, Observatory Formation; 8, Bulimina jacksonensis Cushman, Observatory Formation; 9, U. jacksonensis Cushman, Observatory Formation; 10, Cancris subconicus (Terquem), Observatory Formation; 11, Nonion scaphum (Fichtel and Moll), Observatory Formation; 12, Elphidium trompi (Ansary), Observatory Formation. (Scale bar = 100 µm except for 2 and 3 = 200 µm)
On the other hand, Cluster B represents the WTP and is characterized by a considerable similarity between Italy and Spain (35%), 25% between Spain and France, and 23% between Italy and France. The common species between these regions involve Gaudryina pyramidata, Nuttalides truempyi, Cibicidoides eocaenus, Orthokarstenia nakkadyi, Bolivina carinata, Spiroplectammina carinata, Cibicides lobatulus, and Oridorsalis umbonatus. Moreover, there is a strong similarity between Egypt and Italy (50%), Spain (44%), and France (35%). These results indicate that there was a marine connection between the Southern and Western Tethyan regions through Eocene.
Conclusions
The examined Middle Eocene succession is classified into two rock units, the Observatory Formation (at the base) and Qurn Formation (at the top). The examination of the foraminiferal assemblages leads to the recognition of 27 species belonging to 3 suborders (Textulariina, Miliolina and Rotaliina), 10 families, and 17 genera. Three local biozones are suggested Elphidium trompi-Quinqueloculina carinata Assemblage Zone, Bulimina jacksonensis Zone, and Quinqueloculina ludwigi Zone. Based on the stratigraphic position of the studied two rock units, the comparison between the suggested biozones with their equivalents in Egypt and the presence of some characteristic species as Idalina cuvillieri and Nummulites spp., the age of the two rock units is supposed to the Middle Eocene. Three local paleoecologic ecozones are suggested and denoted a hyper-to hyposaline inner neritic environment for the lower part of the Observatory, the middle shelf with low oxygen conditions for the upper part of the Observatory Formation, and hypersaline inner neritic environment for the Qurn Formation.
Moreover, the paleogeographic distribution of the studied benthic foraminifera was documented. With the aid of multivariate analysis, the paleobiogeographic framework is done. The results of PCA show that there are two provinces, the Southern Tethyan Province (Tunisia, Libya and Egypt) and the Western Tethyan Province (Spain, France and Italy). Also, the results of cluster analysis support this conclusion. The similarity between the two provinces suggests a marine connection between them through Eocene.
References
Abd El-Gaied IM, Abd El-Aziz SM (2020) Stratigraphy and paleoenvironment of the Lower-Middle Eocene succession in the Darnah area, northeast Libya. J. Afr. Ear. Sci. 169, 103774.
Abd El-Gaied IM, Attia, GM, Mahmoud AA, Abu Bakr S (2019) Foraminiferal biostratigraphy and paleoenvironment of the middle and Upper Eocene succession at Cairo-Helwan area, north Eastern Desert, Egypt. J. Afr. Ear. Sci. 158, 103516.
Abu Ellil MM (2004) Stratigraphy of the Middle Eocene rocks in the area of Qattamiya-Northern Galala stretch, north Eastern Desert, Egypt. Ph. D. Thesis, Fac. of. Sci., Zagazig Univ., 236 pp
Abul Nasr RA (2000) Middle-Upper Eocene benthonic Foraminifera of Wadi Tayiba and Wadi Bagha (western Sinai): A comparative study. M.E.R.C. Ain Shams University Ear Sci Ser 14:49–76
Allam A, Shamah K, Zalat A (1991) Biostratigraphy of the middle Eocene succession at Gabal Mishgigah, Wadi Rayan, Libyan desert Egypt. J Afr Ear Sci 12(3):449–459
Anan HS (2019) On the variability of benthic foraminiferal species of the genus Pleurostomella in the Tethys. J Microbiol Experiment 7(3):173–181
Ansary SE (1955) Report on the foraminiferal fauna from the Upper Eocene of Egypt. Publ. Inst. Desert Egypt 6: 160 pp.
Barbin V, Keller-Grünig A (1991) Benthic foraminiferal assemblages from the Brendola Section (Priabonian Stage Stratotype area, northern Italy): distribution, palaeoenvironment and palaeoecology. Marine Micropalentol 17:237–254
Boukhary M (1970) Facies, paleontology and biostratigraphy of some Eocene rocks of the Cairo-Minia reach of the Nile Valley. M. Sc. Thesis, Fac. Sci., Ain Shams Univ., P 227.
Boscolo Galazzo F, Giusberti L, Luciani V, Thomas E (2013) Paleoenvironmental changes during the Middle Eocene Climatic Optimum (MECO) and its aftermath: the benthic foraminiferal record from the Alano section (NE Italy). Palaeo., Palaeo. Palaeo 378:22–35
El Baz SM (2019) Middle Eocene larger benthic foraminifera from Egypt and their similarity to the southern tethyan provinces. J Afr Ear Sci 152:95–100
El Dawy MH (1997) Middle Eocene benthic foraminiferal biostrstigraphy and paleoecology of east Beni Mazar area, Nile Valley. Egypt Egy J Geol 41(1):413–464
Elewa AT (2004) Quantitative analysis and palaeoecology of Eocene ostracoda and benthonic foraminifera from Gebel Mokattam, Cairo. Egypt Palaeogeogr Palaeoclimatol Palaeoecol 221(3–4):309–323
Elewa AT (2018) Cretaceous Ostracods of North Africa and the Middle East: Paleogeography and Paleoecology. Int J Res Environ Scie 4(2):11–21
Elewa AMT, Mohamed O (2014). Migration Routes of the Aptian to Turonian Ostracod Assemblages from North Africa and the Middle East. Paleontol J v. 2014, 7 p.
Elewa AAM, Omar AA, Dakrory AM (1998) Biostratigraphical and paleoenvironmental studies on some Eocene ostracodes and foraminifera from the Fayoum depression, Western Desert. Egypt Egypt J Geol 42(2):439–469
Farag IAM, Ismail MM (1959) Contribution to the stratigraphy of the Wadi Hof area (northeast of Helwan). Bull Faculty Sci Cairo Univ 34:147–168
Farouk S, Jainb S, Belal N, Omran M, Al-Kahtany K (2020) Quantitative Middle Eocene benthic foraminiferal biofacies from west-central Sinai, Egypt: implications to paleobathymetry and sequence stratigraphy. Marine Micropaleont. 155: 101823.
Gooday AJ (2003) Benthic foraminifera (Protista) as tools in deep-water palaeoceanography: a review of environmental influences on faunal characteristics. Adv. Mar. Biol.: 46–90.
Hassan HF, Korin AH (2019) Contribution to the biostratigraphy of the Middle-Upper Eocene rock units at North Eastern Desert; an integrated micropaleontological approach. Heliyon 5: e01671.
Haynes JR (1981) Foraminifera. Macmillan pub l. LTD., London and Basingstoke, p 391.
Issawi B, El-Hinnawi M, Francis M, Mazher A (1999) The Phanerozoic geology of Egypt, A geodynamic approach. Geol. Surv. Egypt, Special Publication 76: 462.
Kaasschieter JPH (1961) Foraminifera of the Eocene of Belgium, vol. 147. Institut Royal des Sciences Naturelles de Belgique, Memoires: p. 271.
Le Calvez Y(1949) Revision des foraminifers Lutetiens du Bassin de Paris. Mem. Expl. Carte, Geol. Det. France., pt. 2, Rotaliidae et Familles affines.
Loeblich AR, Tappan H (1988) Foraminiferal Genera and Their Classification. Van Nosrand Reinhold Co., New York 2: p. 970.
Murray JW (1973) Distribution and ecology of living benthic foraminiferids. Crane and Russak, New York: p. 274.
Murray JW (2006) Ecology and applications of benthic foraminifera. Cambrige University Press, p. 462.
Murray JW, Curry D, Haynes JR, King C (1981) Paleogene. In: second ed. In: Jenkins, D.G., Murray, J.W. (Eds.), Stratigraphical atlas of fossil foraminifera, Brit. Micropaleont. Soc. Ser. 10: 490–536.
Nassif MS, Korin AE (2018) Middle-Upper Eocene benthic foraminiferal biostratigraphy across Cairo-Sukhna district, North Eastern Desert. Egypt IOSR Journal of Applied Geology and Geophysics 6(1):43–53
Ostad-Ali-Askari et al., (2020), Effect of climate change on precipitation patterns in an arid region using GCM models: case study of Isfahan-Borkhar Plain. Natural Hazards Review. ASCE-Am Soc Civil Eng Pub 21(2) 2020. https://doi.org/10.1061/(ASCE)NH.1527-6996.0000367.
Parker JH, Gischler E (2015) Modern and relict foraminiferal biofacies from a carbonate ramp, offshore Kuwait, northwest Persian Gulf. Facies 61(3):1–22
Phleger FB, Parker FI (1951) Ecology of foraminifera northwest Gulf of Mexico. Geol. Soc. Am. Mem. 46.
Ramadan AA, Abd El-Gaied, IM, Saber, SG, Salama, YF (2021) Foraminiferal biostratigraphy and paleoenvironment evolution recorded in the Upper Eocene succession in northeastern Desert, Egypt. J Sediment Environ https://doi.org/10.1007/s43217-021-00065-4.
Said R, Martin L (1964) Cairo area, geological excursion notes, In Trip to Egypt Petrol. Explor. Soc. Libya, Annu. 6th field Conf.: 107–121.
Sallam ES, Ozgen N, Sinanoğlu D, Ruban DA (2018) Mid-Eocene (Bartonian) larger benthic foraminifera from southeastern Turkey and northeastern Egypt: new evidence for the palaeobiogeography of the Tethyan carbonate platforms. J Afr Earth Sc 141:70–85
Shahin A (2000) Biostratigraphic significance, paleobiogeography and paleobathymetry of tertiary buliminacea and bolivinacea in the western Sinai, Egypt- N. Jb. Geol. Palaont., Abh. 216 ( 2): 195–231.
Shahin A, Bassal A, El-Halaby O, El Baz SM (2007) Middle Eocene benthic foraminiferal biostratigraphy and paleoenvironment at the Qattamiya area, northern Eastern Desert, Egypt. Egy. J. Paleont. 7, 29 pp.
Shahin A, El Baz SM (2021). Biostratigraphy and paleobiogeography of the Early-Middle Miocene ostracods and foraminifera from the northern part of the Gulf of Suez, Egypt. J African EarthSci 182, 1–18.
Strougo A, Bignot G, Abd-Allah AM (1992) Biostratigraphy and paleoenvironments of Middle Eocene benthonic foraminiferal assemblages of the north central Eastern Desert. Egypt Mid East Res Center Ain Shams Univ Earth Sci 6:1–12
Sztrákos K (2000) Eocene foraminifers in the Adour Basin (Aquitaine, France): biostratigraphy and taxonomy. Rev De Micropaléont 43(1):71–172
Van der Zwaan G, Duijnstee I, Den Dulk M, Ernst S, Jannink N, Kouwenhoven T (1999) Benthic foraminifers: proxies or problems?: a review of paleocological concepts. Earth Sci Rev 46:213–236
Wells JW (1956) Scleractinia. In: Moore RC (ed) Treatise on invertebrate paleontology. Geological Society of America and University of Kansas Press, Lawrence, pp 328–444
Acknowledgements
The author would like to thank the editor and the reviewers for their constructive and useful comments that helped to improve the manuscript.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The author declares no competing interests.
Additional information
Responsible Editor: Zakaria Hamimi
This article is part of the Topical Collection on Current Advances in Geological Research of Egypt
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baz, S.M.E. Middle Eocene benthic foraminifera from Qattamiya area, Cairo–Suez district, Egypt: biostratigraphy, paleoecology, and their relation to the Southern and Western Tethyan Provinces. Arab J Geosci 15, 749 (2022). https://doi.org/10.1007/s12517-022-09933-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12517-022-09933-3