
Abstract
The chalara-like anamorphs of Leotiomycetes are phialidic hyphomycetes with cylindrical collarettes and deeply seated sporulating loci, and hyaline, aseptate or septate, cylindrical conidia. They are commonly found on plant litters in both terrestrial and submerged environments, and with broad geographical distribution. This paper reports our research result of diversity, taxonomy and phylogeny of these fungi in China, which is based on a systematic study by using an integrated approach of literature study, morphological observation and phylogenetic analyses of 153 chalara-like fungal species with diversified morphology in conidiomata, setae, conidiophores, phialides and conidia. The phylogenetic analyses employing different datasets of SSU, LSU and ITS sequences of 116 species showed that these chalara-like fungi were paraphyletic and scattered in 20 accepted genera belonging to five families of Leotiomycetes: Arachnopezizaceae, Hamatocanthoscyphaceae, Helotiaceae, Neolauriomycetaceae and Pezizellaceae. Additional six genera, Ascoconidium, Bioscypha, Chalarodendron, Didonia, Phaeoscypha and Tapesina, all reported with chalara-like anamorphs in literatures, are also accepted as members of Pezizellaceae or Leotiomycetes genera incertae sedis. Among of these 26 accepted genera of chalara-like fungi in Leotiomycetes, 17 genera are asexually typified genera (Ascoconidium, Bloxamia, Chalara, Chalarodendron, Constrictochalara, Cylindrochalara, Cylindrocephalum, Leochalara, Lareunionomyces, Minichalara, Neochalara, Neolauriomyces, Nagrajchalara, Parachalara, Stipitochalara, Xenochalara and Zymochalara), and 9 are sexually typified genera (Bioscypha, Bloxamiella, Calycellina, Calycina, Didonia, Hymenoscyphus, Mollisina, Phaeoscypha and Tapesina). The phylogenetic significance of conidial septation in generic delimitation was further confirmed; while other morphologies such as conidiomata, setae, conidiophores, phialides, conidial length, and conidial ornamentation have little phylogenetic significance, but could be used for species delimitation. The polyphyletic genus Chalara s. lat. is revised with monophyletic generic concepts by redelimitation of Chalara s. str. in a narrow concept, adaption of the emended Calycina to also include asexually typified chalara-like fungi, reinstatement of Cylindrocephalum, and introduction of six new genera: Constrictochalara W.P. Wu & Y.Z. Diao, Leochalara W.P. Wu & Y.Z. Diao, Minichalara W.P. Wu & Y.Z. Diao, Nagrajchalara W.P. Wu & Y.Z. Diao, Parachalara W.P. Wu & Y.Z. Diao and Stipitochalara W.P. Wu & Y.Z. Diao. Chaetochalara becomes a synonym of Chalara s. str., and the known species are disassembled into Chalara s. str. and Nagrajchalara. The polyphyletic genus Bloxamia is also redefined by introducing the new genus Bloxamiella W.P. Wu & Y.Z. Diao for B. cyatheicola. Five existing species of Chalara s. lat. were excluded from Leotiomycetes and reclassified: Chalara breviclavata as Chalarosphaeria breviclavata W.P. Wu & Y.Z. Diao gen. et sp. nov. in Chaetosphaeriaceae, C. vaccinii as Sordariochalara vaccinii W.P. Wu & Y.Z. Diao gen. et sp. nov. in Lasiosphaeriaceae, and three other Chalara species with hyaline phialides, C. hyalina, C. schoenoplecti and C. siamense as combinations of Pyxidiophora in Pyxidiophoraceae. For biodiversity of these fungi in China, a total of 80 species in 12 genera, including 60 new species, 17 new records and 1 new name, were discovered and documented in this paper. In addition, five species including three new species are reported from Japan. In connection to this revision, a total of 44 new combinations are made. The identification keys are provided for most of these genera. Future research area of these fungi should be the phylogenetic relationship of several sexually typified genera such as Bioscypha, Calycellina, Calycina, Didonia, Phaeoscypha, Rodwayella and Tapesina, and systematic revision of existing names under the genera Bloxamia, Chaetochalara and Chalara.
Similar content being viewed by others



Table of contents
Introduction
Materials and methods
Result and discussion
Molecular phylogeny
Morphology
Taxonomy
Pezizellaceae
Genera with aseptate conidia
Bloxamia
Bloxamiella
Calycellina
Calycina
Chalara
Mollisina
Parachalara
Zymochalara
Genera with septate conidia
Nagrajchalara
Cylindrocephalum
Neochalara
Phaeoscypha
Rodwayella
Tapesina
Arachnopezizaceae
Leochalara
Hamatocanthoscyphaceae
Constrictochalara
Cylindrochalara
Infundichalara
Stipitochalara
Xenochalara
Helotiaceae
Hymenoscyphus
Neolauriomycetaceae
Exochalara
Lareunionomyces
Minichalara
Neolauriomyces
Leotiomycetes genera incertae sedis
Ascoconidium
Bioscypha
Chalarodendron
Didonia
Excluded species from Leotiomycetes
Chaetosphaeriaceae
Chalarosphaeria
Lasiosphaeriaceae
Sordariochalara
Pyxidiophoraceae
Pyxidiophora
Sclerococcaceae
Rhopalophora
Pseudosclerococcum
Discussion
A list of new names proposed in this study
References
Introduction
The chalara-like fungi with cylindrical collarettes and deeply seated conidiogenous loci are paraphyletic and phylogenetically connected to several ascomycetous orders such as Chaetosphaeriales, Dothideales, Helotiales, Laboulbeniales, Microascales, Mytilinidiales, Sclerococcales, Sordariales and Trichosphaeriales (Holubová-Jechová 1973, 1993; Matsushima 1975; Nag Raj and Kendrick 1975; Upadhyay 1981; Gams and Philippi 1992; Paulin and Harrington 2000; Seifert et al. 2011; Johnston et al. 2019; Crous et al. 2021). Within Leotiomycetes, the chalara-like fungi are hyphomycetous anamorphs, and characterized by sporodochial or synnematous conidiomata, or solitary conidiophores, sessile or stalked phialidic conidiogenous cell with a basal venter and a cylindrical collarette bearing a deep-seated conidiogenous locus, and hyaline, cylindrical or obclavate or oblong, aseptate or septate conidia extruded in short or long chain (Nag Raj and Hughes 1974; Nag Raj and Kendrick 1975; Minter et al. 1982, 1983; Seifert et al. 2011). In nature they are commonly found as saprobes on decaying plant material such as leaves, branches, stems, barks, woods, and fruits from terrestrial and submerged environments (Willoughby and Archer 1973; Nag Raj and Kendrick 1975; Tsui et al. 2001; McKenzie et al. 2002; Baschien et al. 2008; Koukol 2011; Friggens et al. 2017). However, they were reported as plant pathogens of trees and ferns, fungicolous or lichenicolous, endophytic fungi of plants and lichens, associated with roots of plants and eggs of nematodes, or from soil (Nag Raj and Kendrick 1975; Gams and Holubová-Jechová 1976; Morgan-Jones et al. 1984; Samuels and Rogerson 1990; Christiansen 1993; Etayo and Diererich 1996; Liu and Zhang 1998; Kowalski 2006; Etayo and Sancho 2008; Husson et al. 2011; Koukol et al. 2012; Gross and Han 2015; Gross et al. 2015 Guatimosim et al. 2016; Friggens et al. 2017; Crous et al. 2020; Etayo and Silanes 2020; Newsham et al. 2020).
Most of the chalara-like anamorphs within Leotiomycetes are classified into asexually typified genera such as Ascoconidium Seaver, Bloxamia Berk. & Broome, Chalara (Corda) Rabenh. s. lat., Chaetochalara B. Sutton & Piroz., Chalarodendron C.J.K. Wang & B. Sutton, Lareunionomyces Crous & M.J. Wingf., Neochalara Crous, Neolauriomyces Crous, Xenochalara M.J. Wingf. & Crous and Zymochalara Guatim., R.W. Barreto & Crous (Nag Raj and Kendrick 1975; Wang and Sutton 1984; Coetsee et al. 2000; Crous et al. 2016b, 2018a, b; Guatimosim et al. 2016). However, some other species are known with both chalara-like anamorphs and discomycete teleomorphs, thus assigned to the sexually typified genera such as Bioscypha Syd., Calycellina Höhn., Calycina Nees ex Gray, Crocicreas Fr., Cyathicula De Not., Didonia Velen., Hamatocanthoscypha Svrček, Hymenoscyphus Gray, Mollisia (Fr.) P. Karst., Phaeoscypha Spooner, Phialea (Pers.) Gillet, Rodwayella Spooner, Sageria A. Funk and Tapesina Lambotte (Nag Raj and Kendrick 1975; Wang and Sutton 1984; Samuels and Rogerson 1990; Seifert et al. 2011; Crous et al. 2016a, b, 2018a, b; Guatimosim et al. 2016). Connections of these anamorphs with their teleomorphs in Leotiomycetes were established only for a few species and limited to four genera such as Ascoconidium, Bloxamia, Chalara and Chaetochalara (Funk 1966a, b, 1975; Nag Raj and Kendrick 1975; Hennebert and Bellemere 1979; Graddon 1980; Samuels and Rogerson 1990; Gams and Philippi 1992; Baral 1989, 2002; Hosoya and Otanl 1997; Paulin and Harrington 2000; Paulin et al. 2002; Guatimosim et al. 2016; Hosoya and Zhao 2016). The two known species of Ascoconidium, A. purpurascens and A. tsugae, were connected with Sageria in Helotiaceae (Funk 1966a, b, 1975). Three species of Bloxamia are connected with members in Pezizellaceae (Bloxamia cyatheicola, Calycina discedens and C. sulfurina) (Johnston 1988; Guatimosim et al. 2016; Hosoya and Zhao 2016; Johnston et al. 2019). The known teleomorphs of Chaetochalara and Chalara s. str. (Leotiomycetes) are members of Bioscypha, Calycina, Calycellina, Cyathicula, Didonia, Hamatocanthoscypha, Mollisina, Phaeoscypha, Phialea and Rodwayella in different families (Nag Raj and Kendrick 1975; Samuels and Rogerson 1990; Gams and Philippi 1992; Svrček 1992a, b; Hosoya and Otani 1997; Baral 2002; Paulin and Harrington 2000; Han et al. 2014). The phylogenetic connections between all these asexually typified genera (except for Chalarodendron) with Leotiomycetes were confirmed by molecular phylogenetic analyses (Paulin and Harrington 2000; Paulin et al. 2002; Cai et al. 2009; Crous et al. 2016a, b, 2018a, b, 2019, 2021; Guatimosim et al. 2016; Ekanayaka et al. 2019; Johnston et al. 2019).
Among all these genera, Chaetochalara and Chalara are by far the largest genera with 157 legitimate names and demonstrate high diversity on morphology (MycoBank, accessed on July 6th, 2022). The two genera were monographed by Nag Raj and Kendrick (1975) who accepted 66 species, including some species correctly placed under Thielaviopsis. Since then another 58 new species have been formally described under the two genera (Gams and Holubová-Jechová 1976; Morgan-Jones and Ingram 1976; Arambarri et al. 1981, 2007; Holubová-Jechová 1982, 1984; McKenzie 1982, 1993, 1997; Kiffer and Delon 1983; Carris and Glawe 1984; Kirk 1984, 1985, 1986; Morgan-Jones et al. 1984; Etayo 1996; Subramanian and Sudha 1986; Kile and Walker 1987; Carris 1988; Cabello 1989; Morgan-Jones et al. 1992; Christiansen 1993; Matsushima 1993; Sutton 1993; Kowalski and Halmschlager 1996; Kile et al. 1996; Gadhil and Dick 1999; Taylor et al. 2001; McKenzie et al. 2002; Wu 2004; Kowalski 2006; Shabunin 2007; Koukol 2011; Pratibha et al. 2005; Silva et al. 2015; Crous et al. 2016a, b, 2019, 2021; Etayo and Silanes 2020). Species concepts in the two genera are mainly based on limited morphological characters of setae, conidiophores (absent or present, shape, size, color, septation, smooth or verrucose), conidiogenous cells (shape, size, color, venter/collarette ratio), and conidia (shape, size, color, septation, basal frill). All other asexually typified genera of chalara-like fungi in Leotiomycetes are recently established monotypic or with only few known species (Nag Raj and Kendrick 1975; Wang and Sutton 1984; Guatimosim et al. 2016; Crous et al. 2016a, b, 2018a, b, 2019, 2021). These fungi are rarely studied with less than 15 species reported from China (Matsushima 1980; Guo 1989, 1997; Lu et al. 2000; Wu 2004; Li et al. 2013a, b).
Phylogenetic analyses showed that Chaetochalara and Chalara s. lat. were polyphyletic, while other recently established genera such as Lareunionomyces, Neochalara, Neolauriomyces and Zymochalara were monophyletic (Paulin and Harrington 2000; Cai et al. 2009; Koukol 2011, 2012; Réblová et al. 2011; Baral et al. 2014; Guatimosim et al. 2016; Suija and Motiejünaité 2016; Ekanayaka et al. 2019; Crous et al. 2020, 2021; Hosoya 2021; Mitchell et al. 2022). In their phylogenetic analyses of chalara-like fungi, Paulin and Harrington (2000) concluded that the 16 Chalara species without known teleomorphs had leotialian affinities, but the relationship among these Chalara specie and among the Leotiales were not resolved. Part of the reason for this was that very few strains were the ex-type materials and often two studied strains identified as the identical Chalara species were shown to have different rDNA sequences, and such discrepancies were seen between LSU sequences of two strains of C. microspora, C. microchona, C. constricta, C. crassipes and C. affinis. Cai et al. (2009) reported that both Chaetochalara and Chalara species reside in Helotiales were paraphyletic, and these species were divided into two well supported clades and other unsupported clades. Furthermore, Cai et al. (2009) also found that conidial width and septation were, to some extent, indicative for referring phylogenetic relationships, while other morphological features, such as conidial length and ornamentation, and form of collarette and venter, had little phylogenetic significance, as species sharing similar features scattered in the tree and did not constitute monophyletic clades. Similar results on potential correlation of morphology and evolution were obtained by several other researchers, although this was not brought into their considerations (Koukol 2011; Guatimosim et al. 2016; Suija and Motiejünaité 2016; Crous et al. 2020, 2021). Réblová et al. (2011) stated that the genus Chalara s. lat. is highly polyphyletic and the analyzed species of Chalara formed four separate groups in the phylogenetic tree. Some of the published species (for example, Chalara fraxinea, Chalara longissima, Chalara microchona) were segregated into monophyletic taxa in distantly related genera and orders. Johnston et al. (2019) in their phylogenetic study of Leotiomycetes reported that Chalara clidemiae and C. longipes belong to different families of Leotiomycetes, i.e., C. clidemiae in Pezizellaceae and C. longipes in Hyaloscyphaceae. Based on molecular phylogeny, Crous et al. (2020, 2021) described two new chalara-like fungi, Neochalara spiraeae and Hamatocanthoscypha podocarpi, under different generic names, although they were morphologically indistinguishable from Chalara s. lat. Despite these findings, no systematic revision was made for the genus Chaetochalara and Chalara, mainly due to small sampling size of species included in their study.
In study of anamorphic fungi from plant litter in China, a large collection of herbarium specimens (> 300) and living strains (293) of chalara-like fungi were made. Initial morphological study showed that these collections represented high diversity in taxa and morphology (conidiomata, setae, conidiophores, conidiogenous cells, and conidia), which were good material for a systematic study and revision of these fungi. With this inspiration, we systematically studied them by an integrated approach of literature review, morphological observation and molecular phylogeny with the following objectives: (1) to further elucidate the phylogenetic relationships of chalara-like fungi in Leotiomycetes; (2) to provide a framework in generic level towards monophyletic genera for future revision of these genera and species; and (3) to monographically document these fascinating fungi from China. The research results are reported in this publication.
Materials and methods
Collection of chalara-like fungi
Fresh specimens of decaying plant materials including leaf litter, dead branches, bark, rotten wood, and fruits from a variety of plants in various environments including forest, botanical garden and agriculture field were collected in many localities in different part of China. Samples were taken to the laboratory in Zip-lock plastic or paper bags for examination; some of these specimens were also incubated in sterile moist chambers in the laboratory to induce more Leotiomycetes fungi. Type specimens were preserved in the Herbarium Mycologicum Academiae Sinicae (HMAS), Institute of Microbiology, Chinese Academy of Science, Beijing. Ex-type strains were preserved in the Chinese General Microbiological Culture Collection Center (CGMCC), Beijing.
Morphological study and description
Morphological study is based on examinations of microscopic preparations made from both fresh specimens collected by the author and living cultures isolated from fresh specimens. For microscopic preparation and identification, the fresh material was mounted in distilled water. Semi-permanent slide preparations were also made by mounting the specimens in cotton blue or lactophenol and sealing with nail vanish, for observation of conidiogenous cells and conidia (Sutton 1980; Nag Raj 1993). The morphological characters (conidiomata, conidiophores, conidiogenous cells, conidia) were examined using a Nikon ECLIPSE 80i and 90i compound microscope and photographed with a Canon 550 D and 600 D digital camera fitted to the microscope. Measurements of morphological structures were made with the Tarosoft (R) v.0.9.7 Image Frame Work program. Photographic plates were edited and combined using Adobe Photoshop CS6 Extended version 13.0.1 software (Adobe Systems, USA).
For description of genera and species, the terms used were based on Seifert et al. (2011). The citation of authors and fungal names followed Index Fungorum. For description of new fungal genera and species, we followed the recommendation by Aime et al. (2021). MycoBank was used for registering all the new fungal names.
Fungal strains, growth, and preservation
Pure cultures were obtained from freshly collected specimens by single spore isolation method. Germinated spores were observed with a stereomicroscope and transferred to potato dextrose agar (PDA) for examination of culture characteristics, sporulation, extraction of DNA and preservation. Colony color was determined according to the color charts of Rayner (1970). The cultures were grown on potato-dextrose agar for morphological study and preservation. All cultures were preserved in 15% glycerin under minus 80 ℃.
Genomic DNA extraction from the fungal strains
DNA extraction was performed from fresh fungal mycelia. Isolates were grown on PDA at 25 °C under dark condition for 7–14 days, or until the colony with enough mycelium for DNA extraction. The mycelium was scraped off and collected in a 1.5 ml micro-centrifuge tube. Mycelium was ground to a fine powder in liquid nitrogen and genomic DNA was extracted using the Fungal gDNA Kit (BioMIGA, USA) according to the manufacturer’s instructions or a modified protocol of Doyle and Doyle (1987). The DNA products were kept at 4 °C for regular use and duplicated at − 20 °C for long-term storage.
DNA amplification and sequencing
DNA amplification was performed by polymerase chain reaction (PCR). Primers pairs NS1 (5′-GTA GTC ATA TGC TTG TCT C-3′) & NS4 (5′-CTT CCG TCA ATT CCT TTA AG-3′) as defined by White et al. (1990) were used to amplify a region spanning approximately 1100 nucleotides from the small subunit of the rDNA (rDNA). LR0R (5′-ACCCGCTGAACTTAAGC-3′) and LR5 (5′-TCCTGAGGGAAACT TCG-3′) primer pairs as defined by Vilgalys and Hester (1990) were used to amplify a segment of the large subunit 28S rDNA (approx. 850 nucleotides). ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) or ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) primer pairs as defined by White et al. (1990) were used to amplify a segment of the ITS1, 5.8 s and ITS2 regions. The amplification was performed in a 50 μl reaction volume as follows: 1X PCR buffer, 0.2 mM d’NTP, 0.3 μM of each primer; 1.5 mM MgCl2, 0.8 units Taq Polymerase and 5–10 ng DNA. The PCR products were checked on 1% agarose electrophoresis gels stained GelRed® Nucleic Acid Gel Stain.
PCR products were purified using minicolumns, purification resin and buffer according to the manufacturer’s protocols (Amersham product code: 27-9602-01). DNA sequencing was performed using the primers mentioned above in an Applied Biosystem 3730 DNA Analyzer by SinoGenoMax Company Limited. For each fungal strain, sequences generated from the respective primers (LROR & LR5, NS1 & NS4, ITS1, ITS5 and ITS2) were manually aligned to obtain an assembled sequence using one of the following programs, VectorNTI, Bioedit (Hall 1999) or SnapGene.
Gene markers, alignment and phylogenetic analyses
Representatives of Leotiomycetes have traditionally been studied using ITS, LSU, SSU and more recently also TEF and RPB2 loci (Han et al. 2014; Ekanayaka et al. 2019; Johnston et al. 2019; Hosoya 2021; Kosonen et al. 2021; Mitchell 2022). The same loci ITS, LSU and SSU were analyzed in the present study. Original sequences from the studied strains were checked using various bioinformatics programs including Vector NTIO, BioEdit, SnapGene, along with reference sequences originated from previous publications. The relevant homogenous sequences were obtained by BLAST searches from GenBank. The SSU and LSU sequences were successfully generated from selected strains representing all morphotypes of chalara-like fungi. The ITS sequences were successfully generated from all living strains obtained in this study. All the newly generated sequences were deposited in GenBank. Strains, their sources, and the GenBank accession numbers of sequences of ITS, LSU and SSU determined in this study are listed in Table 1. Accession numbers for sequences retrieved from GenBank and published in other studies are listed in the Supplementary Table 1.
Alignments of each dataset were done in MAFFT v7.307 online version (Katoh and Standley 2016) and manually verified in MEGA v.7 (Tamura et al. 2013; Kumar et al. 2016) to allow maximum alignment and minimum gaps. For the phylogenetic analyses of chalara-like anamorphs in Leotiomycetes, three datasets were analyzed. Fuscocatenula variegata #12520 (Chaetosphaeriaceae) was selected as outgroup. The first concatenated dataset of the two loci (SSU and LSU) contained 95 sequences of only the representative members of different families. The second concatenated dataset of the two loci (SSU and LSU) contained 159 sequences of many members of different families. The third concatenated dataset of the two loci (ITS and LSU) contained 265 sequences of many members of different families in Leotiomycetes.
The files for Bayesian inference analyses and maximum likelihood analyses were formatted with Mesquite v3.04. Phylogenetic analyses were performed using Bayesian Inference (BI) and Maximum-Likelihood (ML) approaches with MrBayes v. 3.2.6 (Ronquist et al. 2012), and PhyML v. 3.0 (Thompson et al. 1997; Guindon et al. 2010), respectively. For Bayesian inference analyses, the best evolutionary model for each locus was determined using MrModelTest v. 2.3 (Huelsenbeck and Ronquist 2001; Nylander 2004). Posterior probabilities (PP) (Rannala and Yang 1996; Zhaxybayer and Gogarten 2002) were calculated by Markov Chain Monte Carlo sampling (MCMC), using the estimated evolutionary models. Two analyses of four MCMC chains were run from random trees for 180,000,000 generations and sampled every 1000 generations. The first 25% of trees were discarded as the burn-in phase of each analysis and posterior probabilities determined from the remaining trees. For the ML analyses, the general time reversible model was applied with an invariable gamma–distributed rate variation (GTR + I + G). Phylogenetic trees were drawn with Figtree v1.4.3. All the multi-locus alignments and trees are deposited in TreeBASE (Submission Number: 29633).
Results
Molecular phylogeny
Phylogenetic relationships of chalara-like fungi in Leotiomycetes were partly resolved based on the analyses of three different datasets (2 for LSU + SSU, 1 for LSU + ITS). The phylogenetic trees were generated by BI and ML analyses and were largely congruent. All phylogenetic analyses with inclusion of a broad diversity of chalara-like fungi in this study clearly demonstrated that the chalara-like fungi in Leotiomycetes were paraphyletic and scattered in more than 4 different families of Leotiomycetes.
LSU and SSU dataset (I)
For the first dataset with only a few representative species under each family of Leotiomycetes, the combined nuclear LSU and SSU sequences were analyzed to determine the phylogenetic relationship of the chalara-like fungi within the Leotiomycetes. The final alignment included 95 strains representing 82 species in 44 genera and 11 families of the Leotiomycetes. which consisted of 1897 characters including gaps (deposited in TreeBASE 29633). The ML tree is shown in Fig. 1. In ML and BI analyses, 52 species of chalara-like fungi scattered on 16 distinct lineages that represented genera or natural groups of species; furthermore, these species clustered in 7 distinct clades. The first clade (Pezizellaceae) includes 13 species of Chalara s. lat. with septate conidia and +/− setae (named as Nagrajchalara in this study), Bloxamia cyatheicola, Zymochalara cyatheae, Z. lygodii and one unnamed chalara-like fungus, all with aseptate conidia and no setae. The second clade included two strains identified as Chalara fusidioides (the type species of Chalara s. lat.) and Xenochalara juniperi, characterized by aseptate conidia and no setae. The third clade (Hamatocanthoscyphaceae) included Chalara austriaca, C. constricta, Infundichalara microchona, and an undescribed chalara-like species, all characterized by aseptate conidia and absence of setae. The fourth clade (Pezizellaceae) consisted of two strongly supported subgroups, one with Bloxamia elongata, B. truncata, and several Chalara s. lat. species (C. africana, Chalara crassipes, C. eucalypticola, C. fungorum, C. parvispora and C. riisgaardii), all characterized by aseptate conidia and +/− setae; the other one consisted of Chalara aurea, C. hughesii, C. kendrickii and an unnamed chalara-like fungus, all characterized by septate conidia and +/− setae. The fifth clade included all known genera of Neolauriomycetaceae (Exochalara, Lareunionomyces, Neolauriomyces, and an undescribed chalara-like species). The sixth clade (Helotiaceae) consisted of four specie of Hymenoscyphus and three of them, H. albidus, H. albidoides and H. globus were with chalara-like anamorphs. The seventh clade consisted of two Chalara s. lat. species, C. hyalina and C. schoenoplecti, characterized by absence of setae, hyaline to subhyaline conidiophores and aseptate conidia without basal frills. These two species did not belong to Leotiomycetes and should be correctly placed in Pyxidiophoraceae under Pyxidiophorales.

Maximum likelihood (ML) tree based on 18S rDNA and 28S rDNA sequence data for the chalara-like anamorphic fungi in Leotiomycetes. Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Fuscocatenula variegata Wu12520 was chosen as the outgroup. Highlighted generic names are members of chalara-like fungi
LSU and SSU dataset (II)
Analyses of the second dataset of combined nuclear SSU and LSU sequences including more species and genera in Leotiomycetes obtained similar tree topologies as the first dataset. The final alignment included 159 strains representing 118 species in 47 genera and 11 families of Leotiomycetes, which consisted of 1901 characters including gaps (deposited in TreeBASE 29633). The ML tree is shown in Fig. 2. The overall results were very similar to those from the first dataset (Fig. 1). The 86 species of chalara-like fungi of Leotiomycetes were grouped on 16 distinct lineages that represent genera or natural groups of species in seven distinct clades with strong support. The first clade (92 bs/0.95 pp) (Pezizellaceae) consisted of 40 species of Chalara s. lat. with septate conidia and +/− setae, Bloxamia cyatheicola, Zymochalara cyatheae, Z. lygodii and an unnamed chalara-like fungus, all with aseptate conidia and absence of setae. The second clade (84 bs/– pp) (Hamatocanthoscyphaceae) included Chalara austriaca, C. constricta, C. ellisii, C. holubovae, C. hyalocuspica, C. recta, C. longipes, Infundichalara microchona, and an undescribed chalara-like species, all characterized by absence of setae and aseptate conidia without basal frill. The third clade (96 bs/– pp) (Pezizellaceae) consisted of Bloxamia elongata, B. truncata, Calycina populina, C. vulgaris, eleven Chalara s. lat. species (Chalara affinis, C. africana, C. brevipes, C. clidemiae, C. crassipes, C. eucalypticola, C. fungorum, C. parvispora, C. pseudoaffinis, C. qiandaohuensis, C. riisgaardii), and 5 undescribed chalara-like species, all characterized by aseptate conidia and +/− setae; the fourth clade (96 bs/0.98 pp) was with Chalara aurea, C. hughesii, and C. kendrickii and one undescribed species, characterized by uniseptate conidia and +/− setae. The fifth clade (89 bs/0.98 pp) (Helotiaceae) consisted of six specie of Hymenoscyphus and four of them, H. albidus, H. albidoides, H. globus, H. koreanus were with chalara-like anamorphs, characterized by reduced conidiophores, and aseptate, short-cylindrical or globose conidia. The sixth clade (90 bs/– pp) was with a strain of Chalara fusidioides among other genera. The seventh clade (97 bs/– pp) included all known genera of Neolauriomycetaceae (Exochalara, Lareunionomyces, Minichalara, Neolauriomyces). Finally, C. vaccinii formed an independent lineage, which was not related to Leotiomycetes and should be correctly placed into Lasiosphaeriaceae (Sordariales). The two Chalara s. lat. species C. hyalina and H. schoenoplecti, with hyaline conidiophores and conidiogenous cells and members of Pyxidiophoraceae, were not included in the analysis due to their far distance from other species.



Maximum likelihood (ML) tree based on 18S rDNA and 28S rDNA sequence data for the chalara-like anamorphic fungi in Leotiomycetes. Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Fuscocatenula variegata Wu12520 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
LSU and ITS dataset
For the third dataset with inclusion of broad diversity of species and genera in Leotiomycetes, the combined nuclear 28S and ITS sequences were analyzed, and the ML tree was shown in Fig. 3. The final alignment included the sequences of 265 strains of 169 species in Leotiomycetes, which consisted of 1591 characters including gaps (deposited in TreeBASE 29633). The result was very similar to those from the first two datasets (Figs. 1, 2). This further confirmed that the chalara-like fungi within Leotiomycetes affinity were paraphyletic. The 101 species of chalara-like fungi in Leotiomycetes were grouped on 19 distinct lineages that represented genera or natural groups of species, scattered in 4 distinct clades that represented different families of Leotiomycetes.






Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the chalara-like anamorphic fungi in Leotiomycetes. Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Neotainosphaeria microsperma 44779 and Parabahusutrabeeja minima 55337 were chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Clade I (87 bs/0.91 pp): consisted of members of Pezizellaceae, and within this clade six strongly supported subclades formed. The first subclade (Subclade I, 97 bs/– pp) consisted of 49 species of Chalara s. lat., all (except for C. strobilina) with septate conidia and +/– setae, named as Nagrajchalara in this work, and Calycellina leucella; the second subclade (Subclade II, 99 bs/1 pp) consisted of Calycellina fagina, Mollisina uncinata, Phialina lachnobrachyoides and P. ulmariae, and among them only M. uncinata was known to produce chalara-like anamorph with solitary conidiophores and aseptate conidia; the third subclade (Subclade III, 94 bs/0.99 pp) consisted of Bloxamia cyatheicola, Zymochalara cyatheae, Z. lygodii and one new chalara-like fungus named as Parachalara olekirkii in this study, characterized by solitary conidiophores or sporodochial conidiomata, and aseptate conidia; the fourth subclade (Subclade IV, 99 bs/1 pp) consisted of 3 species of Bloxamia, 11 species of Calycina (including the type species C. herbarum, and C. alstrupii, C. claroflava, C. cortegadensis, C. discreta, C. lactea, C. languida, C. marina, C. populina, C. sulfurina, C. vulgaris), 9 species of Chalara s. str. with aseptate species (except for C. eucalypticola with 0–1-septate conidia), and Mollisina rubi; The fifth subclade (Subclade V, 93 bs/1 pp) consisted of 8 species of Chalara s. str., all with aseptate conidia and +/– setae; the sixth subclade (Subclade VI, 100 bs/1 pp) consisting of two species of Calycina, C. citrina and C. shangrillana, no chalara-like anamorph were reported for them; the seventh subclade (Subclade VII, 97 bs/0.98 pp) consisted of 6 species of Chalara s. lat., all with septate conidia and +/– setae, named as Cylindrocephalum in this study. In addition, Brachyalara straminea (the type species), Neochalara spiraeae (the type species) and Remieria rhododendricola scattered among these subclades.
Clade II (100 bs/0.87 pp): included all known genera of Neolauriomycetaceae (Exochalara, Lareunionomyces, Neolauriomyces) and the new genus Minichalara. Roseodiscus sinicus (only ITS sequence was available) also clustered into this clade, but this needs to be further validated in future study.
Clade III (99 bs/1 pp): consisted of 7 species of Hymenoscyphus in Helotiaceae and five of them, H. albidus, H. albidoides, H. fraxineus, H. globus, H. koreanus were reported to produce chalara-like anamorphs.
Clade IV (100 bs/– pp): included members of 8 species of Chalara s. lat. (C. constricta, C. ellisii, C. holubovae, C. hyalocuspica, C. longipes, C. recta, C. piceae-abietis, and one new species), all with aseptate conidia and absence of setae, and named under Constrictochalara, Cylindrochalara in this study, 3 species Hamatocanthoscypha (the type species H. laricionis, H. podocarpi and H. straminella), Infundichalara microchona (the type species) 2 species of Microscypha (M. ellisii and one unnamed species), and Xenochalara juniperi (the type species) in Hamatocanthoscyphaceae. The chalara-like anamorphs in this family are characterized by absence of setae, and aseptate and narrow conidia without basal frills.
Phylogenetic placement of chalara-like fungi in Leotiomycetes
Based on the above phylogenetic analyses, it can be concluded that the chalara-like fungi within Leotiomycetes are paraphyletic, and the analyzed species belong to 20 genera in five families of Leotiomycetes. The other six chalara-like genera (Ascoconidium, Chalarodendron, Bioscypha, Didonia, Phaeoscypha and Tapesina) were not included in the analysis due to lacking living strain or DNA sequence, and they were listed under Pezizellaceae or Leotiomycetes genera incertae sedis (Baral 2002; Ekanayaka et al. 2019; Johnston et al. 2019). In addition, three existing species, Chalara hyalina, C. schoenoplecti and C. vaccinii should be excluded from Leotiomycetes and reclassified.
Arachnopezizaceae: includes Leochalara, a newly created genus for a Chalara-like fungus with hyaline conidiophores and aseptate conidia. In a different phylogenetic analysis with another dataset of LSU and ITS sequence, the new genus Leochalara, was assigned to Arachnopezizaceae. Morphologically it is similar to Pyxidiophora but phylogenetically distinct. Morphologically it is similar to Pyxidiophora but phylogenetically distinct.
Hamatocanthoscyphaceae: includes Constrictochalara, Cylindrochalara, Infundichalara and Stipitochalara. The new genus Constrictochalara is introduced for Chalara constricta, C. ellisii, C. holubovae and one undescribed species. They were characterized by short conidiophores consisting of 1–2-celled basal stalk and a terminal phialide with sharp transition from venter to collarette, obvious constriction and darker between venters and collarettes, and hyaline, aseptate, clavate, subcylindrical or cylindrical conidia (Nag Raj and Kendrick 1975; Koukol 2011). Cylindrochalara is introduced for C. hyalocuspica which clustered together with Infundichalara microchona (the type species of the genus), but the two species differs in shape of collarette and conidia. In I. microchona, collarettes were typical funnel-shaped, and conidia were aseptate and clavate; while in C. hyalocuspica, collarettes and conidia were cylindrical. The new genus Stipitochalara is established for Chalara longipes, C. picea-abietis and C. recta, all characterized by multi-septate conidiophores, terminal phialides with abrupt transition from venter to collarette, and hyaline, aseptate, cylindrical conidia without basal frill. Other included genera in this family were Xenochalara, Microscypha, and two species of Hamatocanthoscypha.
Helotiaceae: includes Hymenoscyphus. Five species of Hymenoscyphus (H. albidus, A. albidoides, H. fraxineus, H. globus and H. koreanus) were known to produce Chalara-like anamorphs, such as. The Chalara-like anamorphs in this clade are characterized by brown and reduced conidiophores consisting of 1–2 basal cells and a terminal phialide, gradual or disrupt transition from venter to collarette, and hyaline, aseptate, subglobose, short cylindrical to cylindrical conidia. In H. globus, the phialides have obvious constriction between venter and collarette and the conidia are slightly clavate-cylindrical. All these species are well-connected with their Hymenoscyphus teleomorphs.
Neolauriomycetaceae: includes Exochalara, Lareunionomyces, Minichalara and Neolauriomyces. These four genera form a strongly supported clade as a distinct family in all phylogenetic analyses. Exochalara, Lareunionomyces and Neolauriomyces and are well-defined monophyletic genera. The three strains of Chalara microspora and one unidentified species from China clustered together as a subclade, and morphologically the two species are very similar in producing pale colored conidiophores, short conidiophores consisting of a 1–3-septate basal stalk and s terminal phialides with gradual transition from venter to collarette, and hyaline, aseptate, cylindrical conidia with rounded ends and no basal frill. The new genus Minichalara is established for the two species. None of these fungi in this family was known with teleomorph (Crous et al. 2016a, b, 2018a, b, 2019).
Pezizellaceae: includes Bloxamia, Bloxamiella, Calycina, Calycellina, Chalara s. str., Cylindrocephalum, Mollisina, Nagrajchalara, Parachalara, Phaeoscypha, Tapesina, Xenochalara and Zymochalara. In the phylogenetic trees, these genera scattered as a few strongly supported lineages under Pezizellaceae clade.
Leotiomycetes genera incertae sedis: include Ascoconidium, Chalarodendron, Bioscypha, Didonia, Phaeoscypha and Tapesina. All these genera are known with Chalara-like anamorphs, but their phylogenetic relationship within Leotiomycetes can’t be determined due to lacking living strain and DNA sequences for molecular phylogenetic analysis.
Preliminary analyses of ITS as barcode for chalara-like anamorphs
Lengths of ITS sequences of the analyzed chalara-like anamorphs (114 species) of Leotiomycetes were between 462 and 468 base pairs. The ITS barcode separated intraspecific from interspecific variability for almost all genera, although occasionally a few species in a given genus were closely related and the genetic distance between them was between intraspecific and interspecific variability observed in other species. The intraspecific variability of the analyzed species was usually below 0.86% in sequence identity, although it was higher (up to 2.16%) in three species of Nagrajchalara. The interspecific sequence divergences were usually > 0.86%, except for the four pathogenic species of Hymenoscyphus with chalara-like anamorphs, which were very similar in morphology and also identity of the ITS sequences (Supplementary Table 2–8).
For the genus Chalara s. str., the lengths of ITS sequences were between 464 and 467 base pairs of the 21 analyzed species. The ITS barcode gap separated intraspecific from interspecific variability. The intraspecific variability among the analyzed species was 0–0.86% in sequence identity. The interspecific sequence divergences were 1.72–22.76%: the minimum is 1.48% between C. fungorum and C. sessilis, while the maximum one was 22.76% between C. longiphora and C. oxenbolliae.
For the genus Nagrajchalara, the lengths of ITS sequences were between 464 and 473 base pairs among the 129 strains representing 39 species. The ITS barcode gap separated intraspecific from interspecific variability, except for the closely related species N. acuaria and N. acuariella; the genetic distance between these two closely species (0.86–1.08%) was between intraspecific and interspecific variability observed in other species. The intraspecific variability of the 39 analyzed species was 0–2.16% in sequence identity. The greatest intraspecific variability was found in N. cannonii (up to 1.28%). N. tubakii (up to 2.16%) and N. yongnianii (up to 1.51%). The interspecific sequence divergences were of the 39 species are 0.86–30.7%: the minimum was 0.86–1.08% between N. acuaria and N. acuariella, while the maximum one was 30.7% between N. jonesii and N. paraunicolor.
Morphology
Morphology of chalara-like fungi in Leotiomycetes was documented by Nag Raj and Kendrick (1975). The common diagnostic characters are pale to dark brown, obclavate, lageniform or subcylindrical phialides consisting of a variously shaped venter and a cylindrical collarette with a deeply seated sporulating locus, and hyaline, aseptate or septate, cylindrical, clavate or other shaped conidia usually with obtuse apex and truncate base, and extruded in short or long chains (Figs. 4, 5, 6, 7).

Conidiomata, conidiophores and conidiogenous cells of chalara-like fungi in Leotiomycetes. a, b Sporodochial conidiomata (a) and cylindrical phialides (b) of Bloxamia elegans. c Apically branched conidiophores in Neolauriomyces crousii. d, f Apically branched conidiophores of N. beijingensis. e Conidiophores bearing branches and terminal phialides in Lareunionomyces minimus. g Upper part of conidiophores bearing branches and terminal phialides L. syzgii. h Solitary conidiophores bearing phialides with significant constriction between collarette and venter in Constrictochalara clavatospora. i, j Sessile phialides with significant constriction between venter and collarette in Hymenoscyphus globus. k Well-developed and multiseptate conidiophores in Chalara longiphora. l Subhyaline conidiophores bearing terminal phialides with gradual transition from venter to collarette and significant constriction in Minichalara aseptata. m Setae of Nagrajchalara mutabilis. Scale bar: 20 μm for a, 10 μm for d, e, k, m; 5 μm for b, c, f–j, l

Conidiophores and conidiogenous cells of chalara-like fungi in Leotiomycetes. a Nagrajchalara inflatipes. b N. unicolor. c N. pulchra. d N. knudsonii. e N. puerensis. f N. yongnianii. g Cylindrocephalum clavatisetosum. Scale bar: 5 μm

Conidiophores and conidiogenous cells of chalara-like fungi in Leotiomycetes. a Nagrajchalara sivanesanii. b N. aspera. c N. yinglaniae. d N. septata. e N. tubakii. f Cylindrocephalum clavatisetosum. g N. mutabilis. h Chalara parilis. i N. cannonii. j C. pengii. k Pyxidiophora schoenoplecti. l C. affinis. m N. strobilina. Scale bar: 5 μm

Conidia of chalara-like fungi in Leotiomycetes. a Leochalara danxiashanensis. b Hymenoscyphus globus. c Lareunionomyces loeiensis. d Chalara crassipes. e Constrictochalara aseptata. f Pyxidiophora schoenoplecti. g Bloxamia elongata. h C. sporendocladioides. i, j Nagrajchalara angionacea. k N. acuaria. l N. yongnianii. m, n N. pulchra. o N. unicolor. p N. strobilina. Scale bar: 5 μm
Sporodochial or synnematous conidiomata are only found in Bloxamia, Bloxamiella, Chalarodendron and Chalara longipes, while the conidiophores in all other genera are solitary or aggregated in small groups (Nag Raj and Kendrick 1975; Koukol 2011; Guatimosim et al. 2016).
Sterile or fertile setae present in several species which were traditionally classified under Chaetochalara. The setae are usually associated with conidiophores at base, or scattered among conidiophores, dark brown, septate, longer than conidiophores, with sterile or fertile apex (Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975; Kendrick 1980; Seifert et al. 2011). Kendrick (1980) was in doubt of difference between the two closely related genera Chalara and Chaetochalara, differentiated by a single character of absence or presence of sterile setae. Kirk and Spooner (1984) merged the two genera and transferred all Chaetochalara species known by that time into Chalara. Cai et al. (2009) in their phylogenetic analyses further confirmed the congeneric of the two genera, and concluded presence or absence of sterile setae was not a reliable character in delimiting the two genera. This study with inclusion of more species with setae further confirmed that presence of setae among the conidiophores can’t serve as the diagnostic character for Chaetochalara, and the two genera are congeneric.
Conidiophores in the chalara-like fungi arise as lateral outgrowths of individual or aggregated vegetative hyphae or from short, broad, thick-walled cells of pseudoparenchymatous layers of synnemata or sporodochia (Nag Raj and Kendrick 1975). When borne directly on the vegetative hyphae, they are usually solitary and scattered or sometimes gregarious, but when they originate from the aggregated hyphae or cells of prosenchymatous layers, they are arranged in loose or compact fascicles as seen in C. connari. The conidiophores of chalara-like fungi are usually morphologically distinct from the vegetative hyphae. In Lareunionomyces and Neolauriomyces the conidiophores are apically branched and with multiple phialides bearing chained conidia; while other genera are usually with simple and unbranched conidiophore, consisting of a basal stalk and a terminal phialide. Percurrent or sympodial proliferation are seen only in a few species such as Chalara constricta, C. nigricollis and C. prolifera. Morphology of the basal stalks varies a lot among species and can be used as one of the characters to separate species. In some species such as C. ampullula. C. emodensis, C. fusidioides, C. curvata, C. microspora and C. sessilis, the basal stalk is absent and the phialides directly arise from superficial vegetable hyphae. However, in most other species the basal stalks are present and consist of one basal cell, and this is found in C. brevipes, C. dictyoseptata, C. hughesii and C. pulchra) to multiseptate and cylindrical (such as C. bicolor, C. cylindrosperma, C. inaequalis, C. insignis, C. longipes, C. nothofagi, C. stipitata, C. tubifera and C. urceolata. Morphology of conidiophores seems to have limited value indicating the phylogeny.
The characteristic conidiogenous cell of chalara-like fungi is a terminal phialide of peculiar morphology. The basic features of this cell are a more or less expanded lower portion, or venter, and a narrower, more or less tubular, open ended collarette bearing a deeply seated sporulating locus (Nag Raj and Kendrick 1975). Based on morphology of venter, Nag Raj and Kendrick (1975) defined four typical morphotypes of the conidiogenous cells in chalara-like fungi, namely ampulliform or lageniform with globose, subglobose or ellipsoidal venter, obclavate with ellipsoidal venter, subcylindrical with cylindrical venter and urceolate with obconical. However, shapes of venters are much more variable than those typical morphotypes among different species or even within the same species. Collarettes in shape of cylindrical, conical or obconical are morphologically distinct from venter in most species of chalara-like fungi, but the venters and collarettes are indistinguishable in Bloxamia and Lareunionomyces, and some species of Chalara s. lat. such as C. microspora (Nag Raj and Kendrick 1975). Transition from venter to collarette may be abrupt (often marked by a perceptible constriction as in C. ampullula, C. insignis, C. inaequalis, C. tubifera, etc.), gradual (C. agathidis, and many other species) or barely perceptible (C. microspora). Cell wall of venter and collarettes are usually smooth in most species, but verrucose appearances are also found in several species such as C. aspera, C. brunnipes, C. curvata, C. cylindrica, C. emodensis, C. bohemica, C. panamensis, and C. scabrida. The phialidic conidiogenous cells among the chalara-like fungi are concolorous or versicolorous, and from hyaline (Leochalara danxiashanensis), pale brown to dark brown. Within the cylindrical collarettes, the monosporulating loci are seated deeply at the basal part of collarette or in the transition region of venter and collarette. All these characters of the conidiogenous cells can be used to separate species, however, they have limited value indicating the phylogeny (Cai et al. 2009).
Conidia in chalara-like fungi with Leotiomycetes affinity are usually endogenous, hyaline, aseptate or septate, cylindrical or subcylindrical, and extruded in short or long chains. However, other conidial shapes such as clavate, ellipsoidal, short cylindrical, subglobose are also seen in some species. The ends of the conidia are usually blunt or truncate, but they are rounded at one end, or both ends in some species. The conidia with truncate base usually bear a minute but distinct marginal frills. The conidia in Chalara rubi are unique in having a fringe of wall material extending for some microns from both ends (Nag Raj and Kendrick 1975; Baral 2002). The shape, size and septation of conidia are the most important characters to delimit species. Aligned with the research result from Cai et al. (2009), the phylogenetic analyses in this study also show that the conidial septation is phylogenetically informative. In the phylogenetic trees majority of the chalara-like species with aseptate and septate conidia phylogenetically clustered into different clades with strong support. Based on this, the polyphyletic genera such as Bloxamia, Chaetochalara and Chalara are revised, and several new genera are established.
Most of the chalara-like fungi of Leotiomycetes can be cultivated in laboratory. The conidia usually germinate easily on PDA, but most of these fungi grow slowly in artificial media. Morphology of colony, including growth rate, color, pigmentation, aerial mycelium etc. is useful for distinguishing genera and species in some cases. For example, the living culture of Hymenoscyphus globus produces characteristic green colony on PDA plate; the living cultures of all Cylindrocephalum species produce orange-colored pigment diffused into agar on PDA plate, which is used as one of the characters to distinguish it from Nagrajchalara.
Taxonomy
Based on the systematic study with an integrated approach of literature study, morphological observation and phylogenetic analyses, 152 species of chalara-like fungi with diversified morphology and phylogeny (115 species were with living strains and DNA sequence data) were classified into 26 genera in more than 5 families, i.e., Leochalara in Arachnopezizaceae; Constrictochalara, Cylindrochalara, Stipitochalara and Xenochalara in Hamatocanthoscyphaceae; Hymenoscyphus in Helotiaceae; Exochalara, Lareunionomyces, Minichalara and Neolauriomyces in Neolauriomycetaceae; Bloxamia, Bloxamiella, Calycellina, Calycina, Chalara, Chalarodendron, Cylindrocephalum, Mollisina, Neochalara, Nagrajchalara, Parachalara, Phaeoscypha and Zymochalara in Pezizelaceae; and Ascoconidium, Bioscypha, Didonia and Tapesina as Leotiomycetes genera incertae sedis, in which the chalara-like anamorphs were reported in literatures, but no living strain or DNA sequence was available for phylogenetic analysis (Table 2; Ekanayaka et al. 2019; Johnston et al. 2019). The species of each genus formed a strongly supported monophyletic clade distinct from others in the phylogenetic trees. Most of these genera were also morphologically well-delimitated. However, some of these genera could hardly be distinguished from each other in morphology. For example, Neochalara was similar to Nagrajchalara, and Calycina, Cylindrochalara and Stipitochalara were similar to Chalara s. str. An identification key for all these accepted genera is provided below.
Among these 26 accepted genera, fifteen genera, Chalara, Chalarodendron, Constrictochalara, Cylindrochalara, Cylindrocephalum, Leochalara, Lareunionomyces, Minichalara, Neochalara, Neolauriomyces, Nagrajchalara, Parachalara, Stipitochalara, Xenochalara and Zymochalara, were asexually typified and without known teleomorphs; while the other eleven genera, Ascoconidium, Bioscypha, Bloxamia, Bloxamiella, Calycellina, Calycina, Didonia, Hymenoscyphus, Mollisina, Phaeoscypha and Tapesina were known with both anamorphs and teleomorphs.
Based on morphological and phylogenetic study of more than 90 species representing a high diversity of morphotypes, the polyphyletic genera Chaetochalara and Chalara with solitary conidiophores, cylindrical collarettes and deeply seated conidiogenous loci, and hyaline aseptate or septate conidia, were revised with monophyletic generic concepts and reclassified into 14 different genera by: (a). redelimitation of Chalara s. str. in narrow concept, (b). adaption of the emended Calycina to accommodate asexually typified chalara-like fungi, (c). reinstatement of Cylindrocephalum, (d). acceptance of four known genera (Ascoconidium, Hymenoscyphus, Neochalara, Zymochalara), and (e). introduction of seven new genera: Constrictochalara W.P. Wu & Y.Z. Diao, Cylindrochalara W.P. Wu & Y.Z. Diao, Leochalara W.P. Wu & Y.Z. Diao, Minichalara W.P. Wu & Y.Z. Diao, Nagrajchalara W.P. Wu & Y.Z. Diao, Parachalara W.P. Wu & Y.Z. Diao and Stipitochalara W.P. Wu & Y.Z. Diao. Chaetochalara became a synonym of Chalara s. str., and the known species were transferred into Chalara s. str. and Nagrajchalara. The polyphyletic genus Bloxamia was also redefined by introducing the new genus Bloxamiella W.P. Wu & Y.Z. Diao for Bloxamia cyatheicola. Chalara breviclavata and C. vaccinii were excluded from Leotiomycetes, and the new genera Chalarosphaeria W.P. Wu & Y.Z. Diao (Chaetosphaeriaceae) and Sordariochalara W.P. Wu & Y.Z. Diao (Lasiosphaeriaceae) in Sordariomycetes were established for them respectively. Two hyaline Chalara species, C. hyalina and C. schoenoplecti were also excluded from Leotiomycetes and reclassified as new combinations of Pyxidiophora (Pyxidiophoraceae) in Laboulbeniomycetes.
For biodiversity assessment a total of 147 species in 26 genera, including 63 new species and 1 new name, were documented in this paper. Among them, 80 species in 12 genera, including 60 new species, 17 new records and 1 new name, were discovered and documented from China (Table 3). In addition, five species including 3 new species were also reported from Japan. In connection to this revision, a total of 44 new combinations were made. All these species are described and illustrated, and identification keys are provided for most of these genera and species. The DNA barcodes (ITS and LSU) were generated for all studied genera and species with pure cultures preserved.
Future research area for these fungi should be the phylogenetic relationship of the several sexually typified genera such as Bioscypha, Calycellina, Calycina, Didonia, Phaeoscypha, Rodwayella and Tapesina, and systematic revision of existing species under the generic names Bloxamia, Chaetochalara and Chalara.
Key to genera with the chalara-like anamorphs in Leotiomycetes
(*difficult to be distinguished in anamorphs, but differs in teleomorph and/or phylogeny)
-
1.
Conidiomata sporodochial or synnematous………2
-
1.
Conidiomata absent; conidiophores solitary or loosely aggregated………4
-
2.
Conidiomata sporodochial/synnematous, composed of aggregated conidiophores………3
-
2.
Conidiomata synnematous………Chalarodendron
-
3.
Saprobe on decaying plant material………Bloxamia
-
3.
Pathogenetic on fern………Bloxamiella
-
4.
Conidiophores with penicillate branches at the apex………5
-
4.
Conidiophores unbranched, with a single terminal phialide………6
-
5.
Transition from venter to collarette gradual………Lareunionomyces
-
5.
Transition from venter to collarette abrupt………Neolauriomyces
-
6.
Conidiophores hyaline to subhyaline………Leochalara
-
6.
Conidiophores pale brown to dark brown………7
-
7.
Phialide ruptured by a vertical split at the apex; conidia phragmoconidia or dictyoconidia……Ascoconidium
-
7.
Phialide ruptured not by vertical split; conidia various………8
-
8.
Conidia septate………9
-
8.
Conidia aseptate………13
-
9.
Conidia with frayed fringes of wall material at both ends………Tapesina
-
9.
Conidia with fringes of wall material at the base………10
-
10.
Setae present; phialide cylindrical to obclavate, no clearly differentiated venter and collarette, transition from venter to collarette gradual………Phaeoscypha
-
10.
Setae absent or present; phialides usually with venter and collarette………11
-
11.
Conidia uniseptate, with both ends truncated………Neochalara
-
11.
Conidia uni- or multiseptate, apex usually obtuse and base truncated………12
-
12.
Colony in PDA with diffused orange-colored pigment………Cylindrocephalum
-
12.
Colony in PDA without orange-colored pigment………Nagrajchalara
-
13.
Conidia short obclavate, with rounded apex and truncated base………14
-
13.
Conidia short-cylindrical to cylindrical, with rounded or flattened ends………15
-
14.
Phialide with constriction between venter and collarette………Constrictochalara
-
14.
Phialide without constriction between venter and collarette………Xenochalara
-
15.
Conidia short-cylindrical to globose………Hymenoscyphus
-
15.
Conidia cylindrical………16
-
16.
Pathogenetic on fern………Bioscypha and Zymochalara*
-
16.
Saprobes not pathogenic on ferns………17
-
17.
Conidiophores very pale brown; transition from venter to collarette gradual………Minichalara
-
17.
Conidiophores brown to dark brown………18
-
18.
Conidiophores reduced to 0–1 basal cell and a phialide; conidia without basal frill………19
-
18.
Conidiophores well-developed or reduced; conidia cylindrical………20
-
19.
Venter cylindrical………Cylindrochalara
-
19.
Venter ellipsoidal………Parachalara
-
20.
Conidiophores well-developed, consisting of a multiseptate stalk and a terminal phialides; conidia no basal frill………Stipitochalara
-
20.
Conidiophores reduced or well-developed; conidia with or without basal frill………Calycina, Calycinella, Chalara s. str. & Mollisina*
Pezizellaceae Velen., Monogr. Discom. Bohem.: 154, 1934.
= Bloxamiaceae Locq., Mycol. Gén. Struct. (Paris): 209, 1984.
= Bloxamiaceae Locq. ex Hern.-Restr., Gené, R.F. Castañeda, J. Mena, Crous & Guarro, Stud. Mycol. 86: 81, 2017.
= Chalaraceae Nann., Repert. MicUomo: 433, 1934.
= Chalareae Sacc., Syll. Fung. (Abellini) 4: 238, 1886.
= Porodiplodiaceae Crous, in Crous et al., Persoonia 40: 363, 2018.
Type genus: Pezizella Fuckel.
Ecology/substrate/host: Saprobe on dead plant material, or pathogenic on plants.
Geographical distribution: Widely distributed worldwide.
Description and illustration: Velenovský (1934), Baral (2016), and Ekanayaka et al. (2019).
Accepted chalara-like fungal genera: Bloxamia, Bloxamiella, Calycina, Calycellina, Chalara, Cylindrocephalum, Mollisina, Nagrajchalara, Neochalara, Parachalara, Phaeoscypha, Tapesina and Zymochalara.
Notes: Many genera were included in Pezizellaceae, and some of these genera produced chalara-like anamorphs. Concept of the family has been constantly challenged and evolving (Han et al. 2014; Baral 2016; Sujia and Motiejünaité 2016; Ekanayaka et al. 2019; Johnston et al. 2019; Mitchell 2022). Results from several phylogenetic analyses of Pezizellaceae by using different datasets were not completely aligned in the literatures (Crous et al. 2018b; Ekanayaka et al. 2019; Johnston et al. 2019). In the Helotiales phylogenetic tree generated by LSU sequences from Crous et al. (2018b), members of Pezizellaceae were split into three different clades (clade I–III); and the chalara-like fungi scattered in two of those 4 clades, but also in other two strongly supported clades outside Pezizellaceae, namely Porodiplodiaceae and Incertae sedis. Crous et al. (2018b) established a new family Porodiplodiaceae for two genera, namely Porodiplodia and two chalara-like fungi, Chalara africana and C. clidemiae. Johnston et al. (2019), based on a 15-gene analysis, adapted a broader concept of the family Pezizellaceae; but the family was split into three clades in the ITS tree, and three chalara-like species Bloxamia truncata, Chalara clidemiae and Mollisina uncinata clustered into two of the three clades. Ekanayaka et al. (2019) accepted 20 genera in the family Pezizellaceae, and 6 chalara-like species (Bloxamia truncata, Bloxamia cyatheicola, Mollisina uncinata, Zymochalara cyatheae, and two unnamed Chalara species) were included in this family. Furthermore, they established a new family Hamatocanthoscyphaceae for those species included in one of the three clades from Johnston et al. (2019), and Chalara was regarded as member of this family. Porodiplodiaceae was treated as a synonym of Pezizellaceae by Johnston et al. (2019) and Ekanayaka et al. (2019). Recently Bisporella was treated as a later synonym of Bispora in Helotiaceae, and many described species under Bisporella were transferred to Calycina (Mitchell et al. 2022). The phylogenetic analyses in this study supported separation of Hamatocanthoscyphaceae and Pezizellaceae as two different families.
Many different species of chalara-like fungi were included in the phylogenetic analysis by using the combined LSU and IST sequences of family Pezizellaceae. In the phylogenetic tree (Fig. 8), the chalara-like fungi scattered in 5 strongly supported clades or lineages under Pezizellaceae, each represented one or several different genera.



Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the chalara-like anamorphic fungi in Pezizellaceae (Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Hymenoscyphus albidus CBS126533 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Nagrajchalara lineage (88 bs/– pp): includes Calycellina leucella, some existing species of Chalara s. lat. and many new species. In conidial septation, all species (except for N. Strobilina and N. xiaohuiae) in this lineage were with septate conidia, mostly uniseptate but also multiseptate as seen in N. angustata, N. insignis, N. inflatipes, N. paraunicolor and N. unicolor. Among these species, four species, N. aspera, N. mutabilis, N. jonesii and N. septata were with dark brown setae, while other species without. Very different morphotypes of conidiophores (shape, color, size, and septation), conidiogenous cells (shape, size and color), and conidia (shape, size and septation) were found among these species. Phylogenetically this clade was distinct from the ones with C. fusidioides and other chalara-like species producing aseptate conidia. The new genus Nagrajchalara was established for these species. Although many teleomorphs of Pezizellaceae were included in the analyses, Calycellina leucella (no anamorph reported, not the type species) and Crocicreas strobilinum (not the type species, anamorph known as Chalara strobilina) were the only species known with teleomorph.
Calycina/Chalara s. str. lineage (95 bs/1 pp): included several existing Chalara species, Bloxamia, Calycina, Calycellina, Neochalara and some new species. This lineage further divided into several subgroups (Figs. 8), the Calycina group (99% bs/1 pp) encompassing 8 species of the genus Chalara s. lat. (C. affinis, C. brevipes, C. crassipes, C. eucalypticola, C. fungorum, C. parvispora, C. pseudoaffinis and C. riisgaardii), 13 species of the genus Calycina (including the type species of the genus, C. herbarum (the type species), and C. alstrupii, C. claroflava, C. cortegadensis, C. discedens, C. discreta, C. lactea, C. languida, C. marina, C. parilis, C. populina C. rubi C. sulfurina and C. vulgaris), 3 species of the genus Bloxamia (including the type species of the genus B. truncata, B. discedens and B. elongata); the Chalara s. str. group (82% bs/0.89 pp) containing 9 species of the genus Chalara s. str. (C. africana, C. bambusicola, C. clidemiae, C. cylindrosperma, C. fungorum, C. longiphora, C. pengii, C. platanicola and C. qinlingensis) and 2 species of Porodiplodia; and the Neochalara group (98 bs/1 pp) consisting of Allophylaria subliciformis, N. spiraeae (the type species of Neochalara), and 2 species of Calycina (C. citrina and C. shangrilana). Four different genera, Bloxamia, Calycina, Chalara s. str. and Neochalara are accepted for the chalara-like fungi in the Calycina/Chalara s. str. lineage. Both Calycina and Chalara s. str. were emended according to the phylogenetic analyses in this study. Conidia of these 18 species of chalara-like fungi in this lineage are typically hyaline, cylindrical and aseptate (except for C. eucalypticola, with both aseptate and septate conidia). Most of these species are without setae, except for the two species, C. africana and C. qinlingensis. Same as in the Nagrajchalara clade, conidiophores and phialidic conidiogenous cells are variable among these species.
Cylindrocephalum lineage (90 bs/1 pp): included a small group of chalara-like species with uniseptate conidia, such as C. aurea (the type species of Cylindrocephalum) C. hughesii, C. kendrickii and two undescribed species. Morphologically these species were similar to those of Nagrajchalara, but phylogenetically they formed a distinct lineage with strong support in all phylogenetic analyses with different datasets (Figs. 2, 3, 8), and also supported by other phylogenetic analyses (Cai et al. 2009; Réblová et al. 2011). The genus Cylindrocephalum, typified by Cylindrocephalum aureum, is reinstated for these fungi. No teleomorph was connected experimentally or phylogenetically for them.
Mollisina lineage (95 bs/1 pp): consisted of Calycellina fagina, Mollisina uncinata, Phialina lachnobrachyoides and P. ulmariae. Among them only M. uncinata was known to produce chalara-like anamorph. The fungus was typified by the teleomorph, but a chalara-like anamorph was described from pure culture.
Zymochalara lineage (91 bs/1 pp): includes three pathogens of tropical ferns, Bloxamia cyatheicola, Zymochalara cyatheae and Z. lygodii, and one saprophytic species of Chalara s. lat. with reduced conidiophores and aseptate cylindrical conidia. Morphologically B. cyatheicola is very similar to B. truncata, the type species, in producing sporodochial conidiomata, but they are phylogenetically distinct. The new genus Bloxamiella is created for this pathogenic fungus on ferns. Two Zymochalara species are morphologically similar to species of Chalara s. str. but phylogenetically distinct. Bloxamia cyatheicola was described with both anamorph and teleomorph, while other species were known with only anamorphs. The new genus Parachalara is created for the saprophytic Chalara-like species within this lineage, which is morphologically similar to Chalara s. str. but phylogenetically distinct.
Based on the above phylogenetic analysis and literature study, a total of 13 genera were accepted for the chalara-like fungi in Pezizellaceae: Bloxamia, Bloxamiella, Calycina, Calycellina, Chalara, Cylindrocephalum, Mollisina, Nagrajchalara, Neochalara, Parachalara, Phaeoscypha, Tapesina and Zymochalara. These included two new genera, Bloxamiella and Parachalara, one reinstated genus Cylindrocephalum, and two emended genera Calycina and Chalara s. str. Many species of these genera were discovered from China and described here.
Genera with aseptate conidia
Bloxamia Berk. & Broome, Ann. Mag. nat. Hist., Ser. 2 13: 468, 1854.
= Endosporostilbe Subram., J. Indian bot. Soc. 37: 49, 1958.
Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Conidiomata synnematous or sporodochial, dark brown, composed of a basal parenchymatous stroma and the aggregated conidiophores. Conidiophores densely aggregated in a sporodochium or synnemata, erect, straight, cylindrical to clavate, septate, simple, subhyaline, pale brown to brown, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells cylindrical to subcylindrical, subhyaline, pale brown to brown, smooth, with collarette not visibly differentiated from the venter. Conidia endogenous, extruded in short or long chains and aggregated in wet spore mass, cuboid or short-cylindrical, oblong, with truncated or flattened ends, straight, mostly hyaline, aseptate, smooth, often provided with basal marginal frill or rarely fringes of wall material. Teleomorph: Apothecia gregarious, sessile or nearly so, shallow cup-shaped, disc flat, bright sulphur-yellow, pale yellow, exterior paler and minutely smooth, tough. Asci cylindric or clavate, usually 8-spore, pore not blued by iodine. Paraphyses cylindrical, containing yellow oil drops. Ascospore biseriate, ellipsoid to elliptic-fusiform, hyaline, 1-septate (For teleomorph, adapted from Dennis 1968).
Type species: Bloxamia truncata Berk. & Broome.
Ecology/substrate/host: Saprobe on dead branches, rotten wood.
Geographical distribution: Widely distributed worldwide.
Description and illustration: Pirozynski and Morgan-Jones (1968), Nag Raj and Kendrick (1975), and Dennis (1968).
Notes: Bloxamia, typified by B. truncata, is characterized by its conidiophores densely aggregated in a black sporodochia or synnemata and arising from a basal thin parenchymatous stroma, cylindrical or clavate phialidic conidiogenous cells, and hyaline or pale-colored cuboid, short-cylindrical or oblong conidia in wet spore mass (Ellis 1971; Pirozynski and Morgan-Jones 1968; Nag Raj and Kendrick 1975; Aramarri et al. 1992). The conidiophores are macronematous, erect, cylindrical, septate, sparsely branched, subhyaline, pale brown or brown, forming a palisade over the stroma and terminating in phialides. The conidiogenous cells are phialidic, cylindrical, subcylindrical or clavate, with a deep collarette extending from undifferentiated venter. The conidia are endogenously differentiated, short-cylindrical to cuboid, oblong with truncate ends, and unicellular, hyaline to subhyaline (Nag Raj and Kendrick 1975). It shares its enteroblastic-phialidic mode of conidial ontogeny with Chalara and Sporoschisma, but is distinct in its sporodochia or synnemata, and the invisibly differentiated phialides. Bloxamiaceae Locq. Was created by Locquin (1984) and validated by Hernández-Restrepo et al. (2017) for Bloxamia truncata Berk. & Broome. However, the phylogenetic analyses showed that the genus Bloxamia was a member of Pezizellaceae (Johnston et al. 2019).
The species assigned to this genus include: B. bohemica, B. cremea, B. cyatheicola, B. foliicola, B. hesterae, B. nilagirica, B. santae-insulae and B. truncata (Nag Raj and Kendrick 1975; Aramarri et al. 1992; Liu and Zhang 1998; Coppins and Minter 1981; Minter and Holubová-Jechová 1981; Seifert et al. 2011; Spooren 2014; Guatimosim et al. 2016). Morphologically these species can be difficult to be distinguished, since limited morphological characters, including conidiogenous cells and conidia, can be used for identification (Lizon and Korf 1995).
Several connections were established between the asexually typified genus Bloxamia and sexually typified species, such as Calycina claroflava, C. discedens and C. sulfurina (Berthet 1964; Carpenter 1975; Johnston 1988; Lizon and Korf 1995; Gamundí and Giaiotti 1998; Zhuang and Hyde 2001; Hosoya et al. 2011; Hosoya and Zhao 2016; Mitchell et al. 2022). Berthet (1964) reported a Bloxamia anamorph in culture from single ascospore isolations of Calycina sulfurina. Carpenter (1975) and Johnston (1988) reported a Bloxamia anamorph for Calycina discedens, where the anamorph was found on host tissue in association with the teleomorph, on the sides of the apothecia, and was also produced in culture from single ascospore. The two species, Calycina claroflava and C. sulfurina can hardly be distinguished from each other based on morphology (Johnston 1988). Guatimosim et al. (2016) described both anamorph and teleomorph under Bloxamia cyatheicola as a leaf pathogen of fern from Brazil.
In a recent study Mitchell et al. (2022) showed that species of Bisporella should be reassigned to at least four genera, and the species with chalara-like anamorphs such as B. claroflava and B. discedens were assigned to Calycina with broad concept. However, the phylogenetic analyses with different datasets of the integrated LSU/SSU and LSU/ITS sequences in this study showed that the genus Calycina was polyphyletic, and the analyzed species scattered in different subclades under the well-supported Calycina/Chalara clade. In the phylogenetic tree of Pezizellaceae (Fig. 8), the 13 species of Calycina scattered in at least 3 subclades under Calycina/Chalara s. str. clade: the first one consisted of C. herbarum (the type species), C. alstrupii, C. discreta, C. languida, C. lactea, C. populina, C. vulgaris, and 6 chalara-like species, the second one consisted of C. cortegadensis, C. marina, C. claroflava, C. sulfurina and three Bloxamia species, and the last one consisted of C. citrina and C. shangrilana. Similar results were also presented by other researchers (Baral and Rämä 2015; Guatimosim et al. 2016; Friggens et al. 2017; Suija and Motiejūnaité 2016; Crous et al. 2019; Karunarathna et al. 2021; Mitchell et al. 2022).
Within the second subclade consisting of Bloxamia truncata, C. cortegadensis, C. marina, C. claroflava, C. sulfurina and two other Bloxamia species, all species with Bloxamia anamorphs were nested together as one strongly supported clade (100 bs/1 pp). The similar result was obtained from another analysis by using the integrated LSU and ITS dataset of Calycina and Chalara. For this reason, the genus Bloxamia is accepted as a separate genus (Fig. 9), although they are closely related to Calycina. This result was well supported by other works (Karunarathna et al. 2021; Mitchell et al. 2022). In the phylogenetic tree provided by Karunarathna et al. (2021), C. discedens and C. sulfurina clustered together as a strong supported subclade (99 bs/ 1 pp) distinct from other Calycina species. Mitchell et al. (2022) showed that B. truncata and C. claroflava clustered together, although the authors adapted a broad concept of Calycina.

Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the Bloxamia and related fungi in Pezizellaceae (Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Catenulifera lururians CBS647.75 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Biologically, most of the described Bloxamia species were saprophytic fungi on dead branches, rotten needles and woods. However, B. foliicola was reported as plant pathogens causing leaf spot disease (Liu and Zhang 1998). Bloxamia foliicola and B. nilagirica were recorded in China, however no strain could be found for molecular phylogenetic study.
Key to all known species of Bloxamia (B.) and Bloxamiella (Bl.)
-
1.
Conidiomata synnematous………2
-
1.
Conidiomata sporodochial………3
-
2.
Conidia cubic, 6–9 × 5–8 µm………B. foliicola
-
2.
Conidia rectangular, 4–5 × 3–3.5 µm………B. nilagirica
-
3.
Conidiogenous cells < 15 µm long………4
-
3.
Conidiogenous cells > 15 µm long………5
-
4.
Conidiogenous cells 8–11 × 1.5–2 µm; conidia cylindrical, 3–5.5 × 1 µm………B. bohemica
-
4.
Conidiogenous cells 10–14 × 1.5–2.5 µm; conidia globose, 2 µm diam………B. sanctae-insulae
-
5.
Conidiogenous cells 14–24 × 2–3 µm; conidia oblong to clavate, 5–6 × 2–3 µm………B. hesterae
-
5.
Conidia rectangular, short cuboid, short-cylindrical ends………6
-
6.
Pathogenetic of fern; conidiogenous cells 17–41 × 1.5–3.5 µm; conidia 2.5–8 × 1–3 µm……Bl. cyatheicola
-
6.
Saprophytic on needle, litter, branches or wood; conidia cylindrical………7
-
7.
Conidia short-cylindrical, < 1.5 µm wide, with rounded apex and truncated base………8
-
7.
Conidia rectangular, short cuboid, < 1.5 µm wide, truncated at both ends………9
-
8.
Conidiogenous cells 15–22 × 2–2.5 µm; conidia 3–4 × 1–1.5 µm………B. elongata
-
8.
Conidiogenous cells 24–26 × 2.5–3 µm; conidia 3–4 × 1–1.5 µm……….B. cremea
-
9.
Conidia 1.7–2 × 1.8–2 µm………..B. discedens
-
9.
Conidia more than 2 µm long………10
-
10.
Conidia 2–2.5 × 1.4–1.6 µm………B. elegans
-
10.
Conidia 2.5–2.6 × 2.3–2.4 µm………B. truncata
Bloxamia discedens (P. Karst.) W.P. Wu, comb. nov., Fig. 10, MycoBank MB846920.

Bloxamia discedens (Wu16029). a–c Sporodochial conidiomata with cylindrical conidiophores and conidiogenous cell. d, e Conidia. Scale bar: 10 µm for a, 5 µm b–e
≡ Helotium discedens P. Karst., Hedwigia 28: 1889.
≡ Bisporella discedens (P. Karst.) S.E. Carp., Mycotaxon 2(1): 124, 1975.
≡ Calycina discedens (P. Karst.) Kunze, Revis. Gen. Pl. (Leipzig) 3(3): 448, 1898.
Description on the natural substrate: Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Sporodochia scattered, or gregarious and confluent, disciform, black, up to 45 μm high, 25–30 µm wide, composed of densely aggregated conidiophores and basal stroma composed of aggregated cells. Conidiophores densely aggregated in a sporodochium and arranged in a densely packed palisade layer, erect, straight, cylindrical, 1–3-septate, simple, basal part subhyaline to pale brown, upper part brown to dark brown, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells cylindrical to subcylindrical, pale brown to brown, smooth, with collarette not visibly differentiated from the venter, 20–25 × 2–2.5 µm. Conidia entoblastic-phialidic, formed in easily dispersible chain and aggregated in wet spore mass, short cuboid, 1.7–2 × 1.8–2 µm, with truncated ends. Teleomorph: Not observed.
Culture characteristics: Colony effuse, rounded, aerial mycelium poorly developed, white, reverse white to very pale brown, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangxi Province, Damingshan, on rotten wood, 19 December 1997, Wenping Wu, Wu1415e; Guangxi Province, Damingshan, on dead branches of unidentified plant, 19 December 1997, Wenping Wu, Wu1413a; Hubei Province, Shennongjia, on rotten wood, 20 September 2004, Wenping Wu, Wu8018, Wu8140b and Wu8165; Jilin Province, on dead branched of unidentified plant, 20 August 2002, Wenping Wu & Yan Huang, Wu4593b and Wu4644b; Yunnan Province, Xishuangbanna, on dead branched of unidentified plant, 16 October 1999, Wenping Wu & Yan Huang, Wu2711; Zhejiang Province, Huaian County, Qiandaohu, 18 October 2018, Wenping Wu, Wu15029. Living strains: CGMCC3.23420 (= 76346, from Wu16029), 46036 (from Wu2711), 46490 (from Wu4644b), 46577 (from 4593b), 47945 (from Wu8018), 48051 (from Wu8165), and 50581 (from 8140b).
Ecology/substrate/host: Saprobe on dead branches, rotten wood.
Geographical distribution: Widely distributed worldwide, including Brazil, China, Colombia, Guadeloupe, Haiti, Japan, New Zealand, Philippines, Puerto Rico, Switzerland, and Venezuela (Lizon and Korf 1995; Carpenter 1975; Johnston 1988; Hosoya and Zhao 2016).
Description and illustration: Carpenter (1975), Johnston (1988), and Hosoya and Zhao (2016).
Notes: Bloxamia discedens is very similar to Bloxamia truncata, but with smaller conidia (1.7–2 × 1.8–2 μm vs. 3–5 × 2–3 µm in Bloxamia truncata). The similar fungus reported from Japan was almost identical to Bloxamia discedens in both morphology and ITS sequences (Hosoya and Zhao 2016). Bloxamia discedens, a widely distributed species in tropical areas, was documented for both teleomorph and anamorph by Carpenter (1975), Hosoya et al. (2011) and Hosoya and Zhao (2016). Carpenter (1975) reported that Bloxamia anamorph often associated with apothecia and processing cylindrical phialides (24–26 × 2.5–3 µm) with a slightly bulbous bases and in group of 3–15 on out surface of the apothecia, and tangular but somewhat spherical conidia (2–2.5 × 2 µm). Hosoya and Zhao (2016) described the scattered pustulate sporodochia with white slimy spore mass on the substrate, numerous cylindrical conidiogenous cells (24–26 × 2.5–3 µm) closely packed together, and cubic to elongate rectangular conidia (2–3 × 2–2.5 µm). From the New Zealand material, Johnston (1988) reported cylindrical conidiogenous cells (20–28 × 2–2.5 µm) and rectangular to more or less square conidia (1.8–2 × 2–2.2 µm). Johnston (1988) also reported both types of conidiomata from different collections and reported that the way in which the conidiogenous cells of the anamorph of Calycina discedens were arranged varied according to substrates. On host tissue and in culture discoid sporodochia formed, while on the apothecia loose aggregations of small numbers of conidiogenous cells were found. The fungus from the Chinese specimens were closer to those from Japan in morphology and also ITS sequences.
Three identical ITS sequences were generated from the relevant strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 50581 included several strains of Bisporella discedens (LC169491, 100% identity), B. discedens (LC169492, 99% identity), B. discedens (MK584952, 99% identity), Bisporella sp. (LC169493, 99% identity), and Bloxamia truncata (KY853426, 94% identity).
Bloxamia foliicola Yun L. Wang & Z.Y. Zhang, Mycosystema 17(1): 7, 1998.
Description on the natural substrate: Anamorph: Conidiomata synnemata, 66–86 µm high, 64–150 µm wide at the upper part, 45–105 µm in the lower part. Conidiophores cylindrical to clavate, septate, simple, brown, smooth. Conidiogenous cells cylindrical to subcylindrical, 64–95 × 9.5–11 µm, brown, with collarette not visibly differentiated from the venter. Conidia formed in easily dispersible chain, cuboid, 6.2–9.2 × 5–8 µm, truncate at both ends, hyaline, aseptate, smooth- and thick-walled (Adapted from Liu and Zhang 1998). Teleomorph: Unknown.
Ecology/substrate/host: Pathogenic on living leaves of Oxyspora paniculata.
Geographical distribution: China (Liu and Zhang 1998).
Description and illustration: Liu and Zhang (1998).
Notes: Bloxamia foliicola resembles B. nilagirica in forming synnemata conidiomata, but can be distinguished from the latter species by colored conidiophores, dry spore mass and larger conidia. In addition, B. foliicola is a leaf pathogen, while B. nilagirica is a saprophytic fungus on wood (Liu and Zhang 1998; Nag Raj and Kendrick 1975).
Bloxamia nilagirica (Subram.) Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 167, 1975.
≡ Endosporostilbe nilagirica Subram., J. Indian bot. Sci. 37: 49, 1958.
Description in pure culture on PDA: Anamorph: Conidiomata synnemata, cylindrical, brown, up to 1625 µm high and 37.5 µm wide, composed of densely aggregated conidiophores formed from the mycelia. Conidiophores densely aggregated, cylindrical, septate, simple, pale brown to brown, smooth, 2–4.5 µm wide. Conidiogenous cells cylindrical to subcylindrical, pale brown, smooth, with collarette not visibly differentiated from the venter. Conidia formed in easily dispersible chain and aggregated as spore mass, cuboid to elongate, truncate at both ends, hyaline to pale brown, aseptate, smooth- and thick-walled, 2.5–6.5 × 2–4 µm (adapted from Xu and Zhang 2012). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead branches, rotten wood.
Geographical distribution: China (Nag Raj and Kendrick 1975; Xu and Zhang 2012).
Description and illustration: Subramanian (1958), Nag Raj and Kendrick (1975), and Xu and Zhang (2012).
Notes: No specimen was examined in this study and the above description is based on publication by Xu and Zhang (2012).
Bloxamia elegans W.P. Wu & Y.Z. Diao, sp. nov., Fig. 11, MycoBank MB845310.

Bloxamia elegans (Wu17130, holotype). a Sporodochial conidiomata from side view. b, c Sporodochial conidiomata from top view, showing the numerous phialidic conidiogenous cells with openings. d, e Conidiophores and cylindrical phialidic conidiogenous cells. f–j Conidia. Scale bar: 20 µm for a, b, 10 µm for c, 5 µm for d–j
Etymology: Refers to its beautiful conidiomata and conidial mass.
Typification: China, Sichuan Province, Qingchengshan, on rotten wood, 9 November 2019, Wenping Wu, Holotype HMAS 352173 (= Wu17130), ex-type strain CGMCC3.23438 (= NN77711).
Description on the natural substrate: Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Sporodochia scattered, or gregarious and confluent, disciform, black, 70–90 μm high, up to 210 µm wide, composed of densely aggregated conidiophores and basal stroma composed of aggregated cells. Conidiophores densely aggregated in a sporodochium and arranged in a densely packed palisade layer, erect, straight, cylindrical, 1–2-septate, simple, basal part subhyaline to pale brown, upper part brown to dark brown, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells cylindrical to subcylindrical, 17–20 × 2–2.5 µm, pale brown to brown, smooth, with collarette not visibly differentiated from the venter. Conidia entoblastic-phialidic, formed in easily dispersible chain and aggregated in wet spore mass, short cuboid, 2–2.5 × 1.4–1.5 µm, with truncated ends. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse concolorous, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Other living strains: 77712 and 77713 (both from Wu17130).
Ecology/substrate/host: Saprobe on dead branches, rotten wood.
Geographical distribution: China.
Notes: Morphologically Bloxamia elegans is similar to other known species of the genus, but differs in relatively large conidiomata, cylindrical phialides, and hyaline cuboid conidia with truncated ends (Pirozynski and Morgan-Jones 1968; Nag Raj and Kenrick 1975).
Three identical ITS sequences were generated from the relevant strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77711 included several strains of Bisporella discedens (LC169491, 97.63–98.7% identity), Bisporella sp. (LC169493, 99% identity), Bloxamia truncata (KY853426, 94% identity), and Bloxamia sp. (LC169493, 98.9% identity).
Bloxamia elongata W.P. Wu & Y.Z. Diao, sp. nov., Figs. 12, 13, MycoBank MB845311.

Bloxamia elongata (Wu7036, holotype). a, b Conidiophores and cylindrical phialidic conidiogenous cells. c–m Conidia. Scale bar: 5 µm

Bloxamia elongata (Wu1041f). a Sporodochial conidiomata. b Basal stroma and unmature conidiophores. c–e Conidiophores with cylindrical collarettes. f–i Conidia. Scale bar: 10 μm for a, 5 μm for b–i
Etymology: Refers to its elongate conidia.
Typification: China, Yunnan Province, Kunming, Kunming Botanical Garden, on rotten wood, 12 September 2002, Wenping Wu, Holotype HMAS 352174 (= Wu7036), ex-type strain CGMCC3.23448 (= NN47646).
Description on the natural substrate: Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Sporodochia scattered, or gregarious and confluent, disciform, black, 50–60 μm high, 100–200 µm wide, composed of densely aggregated conidiophores and basal stroma composed of long, loosely aggregated cells. Conidiophores densely aggregated in a sporodochium arising from upper layer of the basal stroma and arranged in a densely packed palisade layer; erect, straight, cylindrical to clavate, 1–4-septate, simple, subhyaline, pale brown to brown, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells cylindrical to subcylindrical, 15–22 × 2–2.5 µm, subhyaline, pale brown to brown, smooth, with collarette not visibly differentiated from the venter. Conidia entoblastic–phialidic, formed in easily dispersible chain and aggregated in wet spore mass of white color, short cylindrical, oblong, 3–4.5 × 1.3–1.5 µm, with truncate ends and basal fringes of wall material. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded or lobbed, aerial mycelium poorly developed, white to pale brown, reverse pale brown, with yellow pigment diffused into agar, sterile, up to 3 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Ningxia Province, Jingyuan County, Liupan Mountain, Liantianxia, on wood of undetermined plant, 25 August 1997, Wenping Wu, Wu and Wu1041f; Living strains: 44441 (from Wu1032c) and 44129 (from Wu1041f).
Ecology/substrate/host: Saprobe on dead branches, rotten wood.
Geographical distribution: China.
Notes: Morphologically, Bloxamia elongata is similar to other known species of the genus, but differs in sporodochial conidiomata, cylindrical and longer conidia. Most other sporodochial species are with cuboid conidia (Pirozynski and Morgan-Jones 1968; Nag Raj and Kenrick 1975). In this species, the basal stroma consists of long and loosely aggregated hyphae.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 47646 included several strains of Bisporella discedens (98–99% identity), Chalara sp. (KX096663, 98% identity), Chalara sp. (MK432782, 99% identity), and many unnamed fungi of Leotiomycetes.
Bloxamia truncata Berk. & Broome, Ann. Mag. nat. Hist., Ser. 2 13: 468, 1854. Figure 14.

Bloxamia truncata (Wu8259b). a Basal stroma and conidiophores of a sporodochial conidiomata. b, c, f Conidiophores and cylindrical phialidic conidiogenous cells. d, e Conidia. Scale bar: 10 µm for a, 5 µm for b–f
Description on the natural substrate: Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Sporodochia scattered, or gregarious and confluent, disciform, black, 55–67 μm high, composed of densely aggregated conidiophores and basal stroma; the basal stroma composed of long, subhyaline to pale brown and loosely aggregated cells, 2.3–2.5 μm wide. Conidiophores densely aggregated in a sporodochium arising from upper layer of the basal stroma and arranged in a densely packed palisade layer; erect, straight, cylindrical to slightly clavate, 1–2-septate, simple, subhyaline, pale brown to brown, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells cylindrical to subcylindrical, 18–22 × 1.8–2.3 µm, subhyaline, pale brown to brown, smooth, with collarette not visibly differentiated from the venter. Conidia entoblastic-phialidic, formed in easily dispersible chain and aggregated in wet spore mass of white color, cuboid or short cylindrical, oblong, 2.5–2.6 × 2.3–2.4 µm, with truncate ends, straight, hyaline, aseptate, smooth, often provided with basal marginal frill or rarely fringes of wall material. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse slightly yellow brown, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Hubei Province, Shennongjia, 20 September 2004, on wood of undetermined plant, Wenping Wu, Wu8259b. Living strain: CGMCC3.23380 (= 50624, from Wu8259b).
Ecology/substrate/host: Saprobe on dead branches and rotten wood.
Geographical distribution: Probably widely distributed (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Berkeley and Broome (1854), Pirozynski and Morgan-Jones (1968), Nag Raj and Kendrick (1975), Minter (1980), Glawe (1984), Kirk and Spooner (1984), and Ellis and Ellis (1987).
Notes: Bloxamia truncata was documented in literature (Pirozynski and Morgan-Jones 1968; Nag Raj and Kenrick 1975). It can be recognized by the sporodochial conidiomata, closely compacted conidiophores terminating in a phialidic conidiogenous cell and hyaline aseptate, cuboid or short cylindrical, oblong conidia. Nag Raj and Kendrick (1975) illustrated a broad diversity of conidial shape in different specimens, and most likely it was the specie complex was recognized by them.
Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 50624 included Bloxamia truncata (KY853426, 98% identity), Bisporella discedens (LC169491, 95% identity), B. sulfurina (MT435107, 95% identity), B. claroflava (KC412006, 95% identity), and many unnamed fungi of Leotiomycetes.
‘Calycina’ claroflava (Grev.) Kuntze, Revis. gen. pl. (Leipzig) 3(3): 448, 1898.
≡ Peziza claroflava Grev., Fl. Edin.: 424, 1824.
≡ Bisporella claroflava (Grev.) Lizoň & Korf, Mycotaxon 54: 474, 1995.
= Bisporella sulfurina (Quél.) S.E. Carp., in Korf. & Carpenter, Mycotaxon 1(1): 59, 1974.
= Calycella citrinocolor (P. Crouan & H. Crouan) Boud., Hist. Class. Discom. Eur. (Paris): 95, 1907.
≡ Calycella claroflava (Grev.) Boud., Hist. Class. Discom. Eur. (Paris):95 1907.
= Calycina citrinocolor (P. Crouan & H. Crouan) Kuntze, Revis. gen. pl. (Leipzig) 3(3): 448, 1898.
= Calycina citrinocolor P. Crouan & H. Crouan, Florule Finistère (Paris): 48, 1867.
= Calycina sulfurina (Quél.) Kunze, Revis. gen. pl. (Leipzig) 3(3): 449, 1898.
= Calycina sulfurina (Quél.) Kunze var. brassicae Grelet, Revue Mycol., Paris 12(2): 63, 1947.
= Helotium sulfurinum Quél., Grevillea 8(no. 47): 116, 1880.
Ecology/substrate/host: Saprobes on various plants.
Geographical distribution: Widely distributed worldwide including Brazil, France, New Zealand, Puerto Rico, Slovakia, UK (Lizon and Korf 1995).
Description and illustration: Berthet (1964), Dennis (1968), and Lizon and Korf (1995).
Notes: The species was carefully reviewed by Lizon and Korf (1995). Connection of Bloxamia anamorph and its teleomorph (under Calycina sulfurina) was reported by Berthet (1964). No formal transfer to Bloxamia is made in this study, mainly due to lacking confirmation of anamorph and teleomorph connection at this moment. Calycina claroflava and C. discedens were treated as conspecific, but the phylogenetic analyses showed they represented two different species (Karunarathna et al. 2021; Hosoya and Zhao 2016).
Bloxamiella W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845214.
Etymology: Refers to its similarity to Bloxamia in sporodochial conidiomata, chalara-like collarettes, and hyaline conidia.
Typification: Bloxamia cyatheicola Guatim., R.W. Barreto & Crous.
Pathogenic on fern, causing leaf spot. Anamorph: Conidiomata sporodochial, hypophyllous, erumpent, solitary or crowded along the margins of the pinnule, discoid, slimy, amber-colored, black; basal stroma of textura intricata, dark brown towards the host tissue, and paler towards the external side. Conidiophores often reduced to the phialidic conidiogenous cells arising from the stroma surface in a densely packed palisade, discrete, subcylindrical, pale brown, becoming paler towards the apex, smooth. Conidia endogenous, basipetal, extruded in short easily fragmenting chains, cylindrical, truncate at both ends, aseptate, hyaline, smooth. Teleomorph: Ascomata apothecial, hypophyllous, sometimes associated with the conidioma on the same pinnula, erumpent, scattered at the margin of the pinnulae, discoid or cupulate (when dry), solitary, sessile, slimy, tough, black; basal stroma of textura intricata; medullary excipulum of textura epidermoidea, composed of thin-walled hyphae, sub-hyaline to hyaline. Paraphyses unbranched, filiform, swollen at the tip, septate, hyaline, smooth. Asci unitunicate, subcylindrical or clavate, without croziers, straight to curved, 8-spored, with small euamyloid apical ring, hyaline, smooth. Ascospores uniseriate, rarely biseriate, fusoid, straight, 1-septate, with one cell slightly larger, biguttulate, hyaline, smooth. (Adapted from Guatimosim et al. 2016).
Ecology/substrate/host: Pathogenic on frond of ferns.
Geographical distribution: Brazil (Guatimosim et al. 2016).
Description and illustration: Guatimosim et al. (2016).
Notes: Bloxamia cyatheicola, the type species of the new genus, was originally described with both anamorph and teleomorph for a fern pathogen in Brazil. The anamorphic state resembled other species of Bloxamia in sporodochial conidiomata, cylindrical phialides densely packed into palisade, and hyaline, aseptate and cylindrical conidia (Pirozynski and Morgan-Jones 1968; Nag Raj and Kenrick 1975). In phylogenetic analyses from this study (Figs. 1, 2, 3, 8, 9), Bloxamia cyatheicola clustered together with the other two chalara-like fungi Zymochalara cyatheae and Z. lygodii, as a strongly supported group distinct from other Bloxamia and Chalara species. The genus Zymochalara is morphologically similar to Chalara in producing solitary conidiophores consisting of a venter and a collarette, and hyaline, aseptate and cylindrical conidia. The new genus Bloxamiella is created as a monotypic genus to accommodate B. cyatheicola, which is phylogenetically and morphologically related to Zymochalara, but belong to two different genera.
Bloxamiella cyatheicola (Guatim., R.W. Barreto & Crous) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845215.
≡ Bloxamia cyatheicola Guatim., R.W. Barreto & Crous, in Guatimosim, Schwartsburd, Barreto & Crous, Mycol. Progr. 15(12): 1249, 2016.
Ecology/substrate/host: Pathogenic on frond of fern.
Geographical distribution: Brazil (Guatimosim et al. 2016).
Description and illustration: Guatimosim et al. (2016).
Notes: The species was described and illustrated by Guatimosim et al. (2016). Most existing Chalara s. lat. and Bloxamia species with affinity to Leotiomycetes are saprophytic on decaying plant material, while B. cyatheicola is a pathogenic on fern and causes frond spot disease.
Calycellina Höhn., Sber. Akad. Wiss. Math.-naturw. Kl., Abt. 1 127(8–9): 601, 1918.
= Scutoscypha Graddon, Trans. Br. mycol. Soc. 74(2): 268, 1980.
Type species: Calycellina punctata (Fr.) Lowen & Dumont.
Ecology/substrate/host: Saprobe on dead branch of Betula spp.
Geographical distribution: Widely distribution.
Description and illustration: Höhnel (1919) and Lowen and Dumont (1984).
Notes: Several species of the genus Calycellina were reported with chalara-like anamorphs, such as Calycellina aspera (= C. carolinensis), C. betulina, C. chalarae and C. ochracea. Among these species, Calycellina aspera is the only species with septate conidia and phylogenetically placed into Nagrajchalara. The phylogenetic relationships of other species with aseptate conidia remains to be studied.
Calycellina betulina (Velen.) Huhtinen & Purhonen, Memor. Soc. Fauna Flora fenn. 92: 127, 2016.
≡ Belonium betulinum Velen., Monogr. Discom. Bohem.: 178, 1934.
Ecology/substrate/host: Saprobe on dead branch of Betula spp.
Geographical distribution: Czech Republic, Finland and France (von Bonsdorff et al. 2016; Domínguez 2020).
Description and illustration: Baral et al. (2005), von Bonsdorff et al. (2016), and Centro De Estudios Micologicos Asturianos (CEMAS).
Notes: The chalara-like anamorph of Calycellina betullina are with short and 1–2-septate conidiophores, pale brown and obclavate phialides, transition from venter to collarette abrupt, and hyaline, aseptate, rectangular conidia (Baral et al. 2005; von Bonsdorff et al. 2016; Domínguez 2020). It differs from the chalara-like anamorphs of Calycellina ochracea (conidia roundish) by conidia shape, and from Calycellina chalarae by the long and multiseptate conidiophores in the latter species (Svrček 1992a, b). No living strain or DNA sequence was available for molecular phylogenetic analyses. Its phylogenetic relationship with other chalara-like fungi remains to be studied in future study.
Calycellina chalarae Svrček, Česká Mykol. 46(1–2): 33, 1992.
Ecology/substrate/host: Saprobe on dead leaf of Betula pendula.
Geographical distribution: Czech Republic (Svrček 1982, 1992a).
Description and illustration: Svrček (1982, 1992a).
Notes: The chalara-like anamorph of Calycellina chalarae was described to be Chalara cylindrosperma (Svrček 1992a, b). However, the connection was only based on observation of the minute apothecia of C. chalarae occurring in small groups among conidiophores of chalara-like anamorph. No living strain or DNA sequence was available for molecular phylogenetic analyses. Its phylogenetic relationship with other chalara-like fungi remains to be studied in future study.
Calycellina ochracea (Grelet & Croz.) Dennis, Persoonia 2(2): 187, 1962.
≡ Arachnopeziza ochracea (Grelet & Croz.) Iturr. & Korf, Mycotaxon 31(1): 246, 1988.
≡ Belonidium ochraceum Grelet & Croz., Bull. Tremest. Soc. mycol. Fr. 44: 336, 1928.
Ecology/substrate/host: Saprobe on dead branch of Betula spp.
Geographical distribution: Europe.
Description and illustration: Dennis (1962).
Notes: The chalara-like anamorph of Calycellina ochracea was reported to be with the roundish conidia. No living strain or DNA sequence wase available for molecular phylogenetic analyses. Its phylogenetic relationship with other chalara-like fungi remains to be studied in future study.
Calycina Nees ex Gray, Nat. Art. Brit. Pl. 1: 669, 1821.
= Helotium subgen. Calycella Fr., Summa veg. Scand., Sectio Post. (Stockholm): 355, 1849.
= Calycella (Fr.) Boud., Bull. Sco. mycol. Fr. 1: 112, 1885.
= Cystopezizella Svrček, Ceská Mykol. 37: 67, 1983.
= Eubelonis Clem., Gen. fung. (Minneapolis): 87, 1909.
= Pezizella Fuckel, Jb. nassau. Ver. Naturk. 23–24: 299, 1870.
Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to brown, smooth, septate and branched hyphae. Anamorph: Stroma absent or present; if present, consisting of aggregated cells. Setae absent. Conidiophores macronematous, micronematous, scattered or aggregated from the basal stroma, erect, simple, or rarely branched, subcylindrical, pale brown, brown to very dark brown, smooth or verrucose, septate, consisting of a basal cell or septate stalk and a terminal phialide, with or without percurrent proliferations. Conidiogenous cells ampulliform, lageniform, obclavate, ellipsoidal, urceolate or subcylindrical, pale brown to dark brown, composed of a venter and a collarette, transition from venter to collarette gradual, abrupt or barely perceptible. Conidia holoblastic, cylindrical, straight, hyaline, aseptate, smooth, with rounded or truncate ends, often with basal marginal frill or rarely fringes of wall material slightly rounded at the apex, extruded in short chains. Teleomorph: Apothecia sessile or nearly so, smooth, tough, the outer tissue of the cup composed of rather thick-walled hyphae undulating and somewhat interwoven but in general lying more or less parallel at a high angel to the surface, whitish or yellow. Asci cylindric or clavate, usually 8-spore. Ascospore ellipsoid to fusoid, hyaline, for a long time simple later often becoming septate with one to several septa (For teleomorph, adapted from Seaver 1934 and Dennis 1968).
Type species: Calycina herbarum (Pers.) Gray.
Ecology/substrate/host: Saprobe on decaying plant material.
Geographical distribution: Widely distributed.
Description and illustration: Gray (1821), Seaver (1934), and Dennis (1968).
Notes: Calycina, typified by C. herbarum, were reported with a total of 235 legitimate names are listed under the genus Calycina (MycoBank, accessed on July 7th, 2022). The phylogenetic analyses clearly showed its affinity with Pezizellaceae (Han et al. 2014; Ekanayaka et al. 2019; Johnston et al. 2019). The phylogenetic analyses of this study showed that at least 13 chalara-like species, including 9 species known as Chalara s. lat., were congeneric with Calycina, since these chalara-like fungal species clustered together with the type species C. herbarum and 8 other species of Calycina as a strongly supported clade (Fig. 8). Similar results were reported by other researchers (Baral and Rämä 2015; Guatimosim et al. 2016; Friggens et al. 2017; Suija and Motiejūnaité 2016; Crous et al. 2019). Based on the literature study and phylogenetic analysis, Calycina was chosen as the generic name for these chalara-like fungi with three strong arguments. Firstly, in the phylogenetic tree the type species C. herbarum clustered together with these chalara-like fungi; secondary Calycina Nees ex Gray (1811) is an earlier name than Chalara (Corda) Rabenh. (1844); and thirdly Calycina seemed to be more frequently used in the academic world, supported by the Google Scholar search with 13,800 results for Calycina and 8200 results for Chalara (searched on 12 December 2022). Based on this result, all these 9 species known as Chalara s. lat. were reassigned to Calycina.
The phylogenetic analyses from different datasets of the integrated LSU/SSU and LSU/ITS sequences showed that the genus Calycina was polyphyletic, although all analyzed species clustered in the well supported Calycina/Chalara s. str. clade (Figs. 2, 3, 8). For example, in the phylogenetic tree of Pezizellaceae (Fig. 8), the 13 species of Calycina scattered in 3 subclades under Calycina/Chalara s. str. clade: the first one consisted of C. alstrupii, C. discreta, C. herbarum, C. languida, C. lactea, C. populina, C. vulgaris, and 6 chalara-like species, the second one consisted of C. cortegadensis, C. marina, C. claroflava, C. sulfurina and three Bloxamia species, and the last one consisted of C. citrina and C. shangrilana. This was also showed into other studies (Baral and Rämä 2015; Guatimosim et al. 2016; Friggens et al. 2017; Suija and Motiejūnaité 2016; Crous et al. 2019; Karunarathna et al. 2021; Mitchell et al. 2022). Future work is needed to clarify concept of the genus and also phylogenetic relationship of many existing species.
Although apparently less frequently reported, several Calycina species have been reported with similar asexual states as phialidic, dematiaceous hyphomycetes (Mitchell et al. 2022). Calycina herbarum, the type species, has been shown to be genetically similar or identical to a Phialophora-like asexual fungus isolated as a grass endophyte (Sánchez Márquez et al. 2007). “Bisporella” tetraspora, Calycina drosodes, C. lactea, C. parilis and Calycina vulgaris have been described as having or growing in close association with a Chalara-like asexual state (Morozova 2014; Bara and Rämä 2015; Friggens et al. 2017; Mitchell 2022); “Bisporella” polygoni is also found with a species of Cystodendron (Carpenter 1981; Morozova 2014). “Bisporella” maireana has been described as bearing brown phialides on the ectal excipulum (Galán 1993). Mitchell et al. (2022) also observed a similar asexual state growing on the excipulum of a collection of a species of Calycina from Ecuador. Even though these fungi have been assigned to several different form genera, the descriptions of these phialidic, dematiaceous asexual states are similar. These results further supported the assignment of those chalara-like anamorphs into Calycina.
In total 14 species of Calycina were recorded in this study (Figs. 15, 16, 17, 18). Ten of these species were known with only chalara-like anamorphs, including the nine species known as Chalara s. lat. (Chalara affinis, C. brevipes, C. brevispora, C. crassipes, C. dualis, C. eucalypticola, C. fungorum, C. parvispora, C. pseudoaffinis) and two new species (Calycina oxenbolliae and C. riisgaardii). The other three species, Calycina lactea, C. parilis and C. vulgaris, were known with both anamorphs and teleomorphs. For identification purpose, an identification key is provided for all chalara-like fungi with aseptate conidia.


Maximum likelihood (ML) tree based on ITS sequence data for the chalara-like anamorphic fungi in Chalara s. str. and Calycina (Pezizellaceae). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Hymenoscyphus albidus CBS126533 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels

Colony of Bloxamia and Calycina species on PDA in 4 weeks. a–d Bloxamia elegans (a, b 76346; c, d 77711). e, f B. elongata (ex-type strain 47646). g, h B. truncata (50624). i, j Calycina affinis (76338). k, l C. brevipes (50639). m, n C. crassipes (CBS829.71). o, p C. eucalypticola (43033)

Colony of Calycina and Chalara species on PDA in 4 weeks. a, b Calycina fungorum (CBS942.72). c, d C. parvispora (CBS385.94). e, f C. oxenbolliae (ex-type strain 50633). g, h C. riisgaardii (47715). i, j Chalara africana (76574). k, l C. bambusicola (ex-type strain 54418). m–p C. longiphora (m, n 76308; o, p ex-type strain 76442)

Colony of Chalara species on PDA in 4 weeks. a–d Chalara pengii (a, b 76196; c, d 76222). e, f C. platanicola (76305). g, h C. qinlingensis (76950). i, j C. cylindrosperma (CBS658.79). k, l Parachalara olekirkii (ex-type strain 43656)
Key to species of chalara-like fungi with aseptate conidia
(C. for Chalara s. lat. species with aseptate conidia, but most likely do not belong to the genus Chalara s. str. and need to be relocated with future molecular phylogenic analyses; Ca. for Calycina; Ch. for Chalara s. str.; Cs. for Constrictochalara; Cy. for Cylindrochalara; I. for Infundichalara; M. for Minichalara; N. for Nagrajchalara; P. for Pyxidiophora; Pc. for Parachalara; S. for Sordariochalara; St. for Stipitochalara; species name in bold are documented in this paper)
-
1.
Setae present………2
-
1.
Setae absent………4
-
2.
Phialides ampulliform, 21–37 µm long; venter globose; conidia 6.5–10 × 1.5–2 µm………Ch. bulbosa
-
2.
Phialides lageniform, obclavate; center lageniform, subellipsoidal………3
-
3.
Phialides 21–37 µm long; conidia 5–9 × 1.5–2 µm………Ch. africana
-
3.
Phialides 127–31 μm long; conidia 5–6–9 × 1.5–1.8 µm……….Ch. qinlingensis
-
4.
Conidia ellipsoidal or clavate………5
-
4.
Conidia cylindrical………11
-
5.
Phialoconidia exceeding 10 µm long………6
-
5.
Phialoconidia less than 10 µm long………7
-
6.
Conidia 9–12.5 × 2.5–3.2 µm………C. caribensis
-
6.
Conidia 10–20 × 3.5–4.5 µm………Chalarosphaeria breviclavata
-
7.
Collarette rough-walled; base of phialophore usually broadly conical and darker………C. brunnipes
-
7.
Collarette smooth; transition usually marked by a constriction………8
-
8.
Holoblastic phragmospores present; conidia 4–6 × 1.3–2.3 µm………S. vaccinii
-
8.
Holoblastic phragmospores absent………9
-
9.
Collarette cylindrical, constricted at the base, 7–11 µm long; phialide not proliferating; conidia cylindrical, 3.5–8 × 1–2 µm………Cs. constricta
-
9.
Collarette funnel-shaped; Conidia ellipsoidal………10
-
10.
Phialide no proliferation; Collarette 7–10 µm long; Conidia 3–4 × 2–2.1 µm………C. antarctica
-
10.
Phialide sympodially proliferating; Collarette 1–2.5 µm long; Conidia 3.5–5 × 1.3–2.5 µm; on Pinus………I. microchona
-
11.
Phialides cylindrical; conidia dark brown, 10–14 × 6–8 µm………C. phaeospora
-
11.
Conidia hyaline………12
-
12.
Conidiophores rough-walled………16
-
12.
Conidiophores smooth………13
-
13.
Transition from venter to collarette usually marked by a constriction………14
-
13.
Transition from venter to collarette non constriction………15
-
14.
Phialide smooth; collarette widening towards apex, 7–9.5 µm long; conidia 3–12 × 1.5–2.1 μm………Cs. ellisii
-
14.
Phialide verruculose; conidia 4–6.5 × 1.75–2 µm………Ch. verruculosa
-
15.
Collarette 6–15 × 1.5–3 µm; venter 4.5–6 µm wide; conidia 3–9.5 × l–1.5 µm………Ch. cylindrica
-
15.
Collarette 15–39 × 3–4.5 µm; venter 6–9.5 µm wide; conidia 9.5–13 × 2–3 µm………Ch. bohemica
-
16.
Conidiophores reduced to sessile phialides, or phialides with one or two stalk cells………17
-
16.
Conidiophores well-developed and multi-septate………50
-
17.
Conidiophores and phialides hyaline or subhyaline………18
-
17.
Conidiophores and phialides pigmented………24
-
18.
Conidia dimorphic, either cylindrical or ranging from ellipsoid or globose to pyriform; cylindrical conidia 6.6–11.6(–16.5) × 2.8–4.4 µm; conidia of other shapes 5–12.5 × 3.3–8.8(–12) µm; on nematode cysts………C. heteroderae
-
18.
Conidia of only one kind………19
-
19.
Conidiophores hyaline………20
-
19.
Conidiophores hyaline to pale brown………21
-
20.
Conidiogenous cells 35–42 µm long, arising from a single stalk cell; conidia (7–)9.3(–12) × (2.5–)2.8(–3) µm; on Eleiodoxa………P. siamensis
-
20.
Conidiogenous cells 22–32 µm long, sometimes arising from a single stalk cell which may give rise to 2 or 3 conidiogenous cells; conidia 10–11 × 3–4 µm; on nematode cysts………P. hyalina
-
21.
Conidia up to 12 μm long………22
-
21.
Conidia more than 12 μm long………23
-
22.
Conidiogenous cells 25–47 µm long, arising from a single stalk cell; conidia (9.4–)11.7(–12.6) × (3.7–)4(–4.7) µm; on Nothofagus………C. brevicaulis
-
22.
Conidiogenous cells 35–50 µm long, arising directly from a single stalk cell; conidia 6–10(–12) × 2.5–3.5 (–4) µm; on lichens………C. lobariae
-
23.
Conidia (8–)12–17.5 × 3–5 µm, cylindrical; first formed conidia subglobose or ellipsoid 5–6.25 × 4.5–5 µm; on Schoenoplectus………P. schoenoplecti
-
23.
Conidia 12.1–26.4 × 2.50–3.3 µm, cylindrical; first formed conidia turbinate, 6.5–9.5 × 4–6 µm; on Ficus………C. sibika
-
24.
Conidia 2–2.5 × 2 μm………Ca. lactea
-
24.
Conidia longer than 3 μm………25
-
25.
Conidia 0–1-septate, 10–17 × 2.5–2.7 μm………Ca. eucalypticola
-
25.
Conidia always aseptate………26
-
26.
Collarettes usually shorter than venter………27
-
26.
Collarettes as long as or longer than venter………30
-
27.
Venter subcylindrical………28
-
27.
Venter conical or ellipsoidal………29
-
28.
Collarette 6–8 µm long; phialides lageniform, 12–21 µm long………Ch. austriaca
-
28.
Collarette 6–17 µm long; phialides usually obclavate, 18–36 µm long………M. microspora
-
29.
Venter usually ellipsoidal, 4–6.5 µm wide; collarette 3–10 × 2–3 µm; transition usually abrupt and marked by a constriction; conidia 1.5–2.5 µm wide………Ch. ampullula
-
29.
Venter usually conic, 3–4 µm wide; collarette 10–11 × 1.5 µm; transition gradual and without constriction; conidia 1–1.5 µm wide………Ch. sessilis
-
30.
Venter usually globose………31
-
30.
Venter cylindrical, subcylindrical or ellipsoidal………34
-
31.
Conidia 4–6 μm long, less than 1.5 μm wide………32
-
31.
Conidia more than 6 μm long and 1.5 μm wide………33
-
32.
Venter 3.5–5 μm diam.; collarettes 5.5–8 × 1.4–1.6 μm; conidia 4–6 × 0.8–1.2 µm………Ch. dennisii
-
32.
Venter 5–6 × 4.5–5.5 μm; collarettes 20–25 × 1.2–1.5 μm; conidia 5–6 × 1–1.2 μm………Ca. oxenbolliae
-
33.
Conidia 13–15 × 2–2.5 µm………Ch. kirkii
-
33.
Conidia 4.5–12 × 1.5–3.5 µm………Ch. fusidioides
-
34.
Collarette darker at base………35
-
34.
Collarette paler than, or concolorous with venter………36
-
35.
Phialides 37–52 μm long; venter 14–24 × 5–7 μm; collarettes 23–31 × 2–2.5 μm; conidia 6–12 × 1.5–2 μm………Ca. brevipes
-
35.
Phialides 50–65 μm long; venter 16–30 × 4.5–5 μm; collarettes 25–30 × 2–2.5 μm; conidia 6–7 × 1–2 μm………Ca.. riisgaardii
-
36.
Conidia not exceeding 10 µm long………37
-
36.
Conidia up to 19 µm long………45
-
37.
Conidia 3.5–4 μm wide………38
-
37.
Conidia less than 2.5 μm wide………39
-
38.
Phialides subcylindrical; transition from venter to collarette gradual; conidia 5–7 × 3.5–4 µm……….Ch. clavatophora
-
38.
Phialide lageniform, 42–48 μm long; transition from venter to collarette abrupt; venter 21–28 × 6–8.5 μm; collarettes 23–27 × 4–5.5 μm; conidia 7–9 × 3.5–4 μm………Ca. parilis
-
39.
Phialides cylindrical; collarette 32–40 × 2.5–3 µm; conidia 7.5–10 × 1–1.5 µm………Ch. cylindrophora
-
39.
Phialides lageniform, obclavate………40
-
40.
Phialides obclavate, transition from venter to collarette gradual………41
-
40.
Phialides lageniform, transition from venter to collarette abrupt………42
-
41.
Collarettes 15–20 × 2–2.5 μm; conidia 7.5–8.5 × 1–1.2 μm………Ch. bambusicola
-
41.
Collarette 11–21 × 2.5–4 µm, conidia 5.5–8 × 2–2.5 µm wide………Ca. fungorum
-
42.
Phialide 35–45 μm long; venter 20–25 × 6–7 μm; collarettes 18–22 × 2.5–3 μm; conidia 6.5–11 × 2–2.5 μm (specimen from China)………Ca. parilis
-
42.
Phialides less than 30 μm long………43
-
43.
Phialide 22–27 μm long; venter 13–16 × 4.3–5.5 μm; collarettes 13.5–21 × 3–3.5 μm; conidia 6.5–9 × 1.8–2.2 μm………Ch. pengii
-
43.
Conidia less than 5 μm in average………44
-
44.
Phialide 15–25 μm long; conidia 3–5 × 2–2.5 μm………Ca. clidemiae
-
44.
Collarette 17–22 × 1.5–2 µm; conidia 3.5–6 × 1.5–1.8 µm……….………Ch. neglecta
-
45.
Phialides cylindrical; Ratio of mean length of collarette and venter > 5; collarettes 65–115 × 5–6.5 µm; conidia (13–)14–17 × 3.5–4.5 µm………Ch. graminicola
-
45.
Ratio of mean length of collarette and venter never more than 3………46
-
46.
Phialide 25–47 μm long; venter 18–24 × 7–9.5 μm; collarettes 14–24 × 4.7–6.5 μm; conidia 9.4–12.5 × 3.8–4.7 μm………Ch. brevicaulis
-
46.
Conidia less than 3.5 μm wide………47
-
47.
Venter 6.5–9.5 µm wide, collarette 17.5–30 × 3–3.5 µm; conidia 10–14 × 2–2.5 µm………Ch. sinensis
-
47.
Venter 5 µm wide………48
-
48.
Collarette 30–45 × 3 µm; conidia 10–15 × 2.2–2.5 µm………Ch. kobensis
-
48.
Collarette less than 30 μm long………49
-
49.
Collarette 12–32 × 2–3 µm wide, conidia 5.5–19 × 1.5–2.5 µm………Ca. affinis
-
49.
Phialide 50–60 μm long; venter 26–31 × 5.5–6.5 μm; collarettes 24–28 × 2.6–2.9 μm; conidia 14.5–17.5 × 2 μm………Pc. olekirkii
-
50.
Conidia 2.5–4 × 0.5–0.8 µm; on lichens………Ch. lichenicola
-
50.
Conidia more than 1 µm wide, not on lichens………51
-
51.
Venter longer than collarette………52
-
51.
Venter as long as or shorter than collarette………55
-
52.
Mean conidium l/w ratio = 1.5: 1; conidia 2–4 × 1.5–2 µm……….………Ca. brevispora
-
52.
Mean conidium l/w ratio = 4: 1 or more………53
-
53.
Collarette less than l0 µm long; transition gradual; conidia lacking basal frill; mean conidium l/w ratio = 4: 1………St. longipes
-
53.
Collarette more than 10 μm long, transition abrupt………54
-
54.
Collarette 25–32.5 × 8–10 μm long; conidia 20–30 × 5–6.5 μm, without basal frill………Ch. indica
-
54.
Collarette 10–15 µm long; conidia 5–8 × 1–1.5 μm………C. novae-zelandiae
-
55.
Conidiophores arising from basal stromatic aggregations of vegetative hyphae………Ch. germanica
-
55.
Conidiophores not arising from such aggregations………56
-
56.
Conidiophores branched at the apex, bearing several phialides………Ch. sporendocladioides
-
56.
Conidiophores simple, bearing a single terminal phialide………57
-
57.
Mean conidium l/w ratio = 2.6:1 or less……….………58
-
57.
Mean conidium l/w ratio more than 2.6:1………61
-
58.
Venter 6.5–9.5 µm wide; collarette 3.5–5 µm wide; conidia 2.5–4 µm wide………C. brachyspora
-
58.
Conidia less than 3 µm wide………..59
-
59.
Conidia 3–4.5 × 1.5–2 µm………Mollisina uncinata
-
59.
Conidia slightly longer but narrower………60
-
60.
Collarette 1.5–2.5 µm wide; conidia 3.5–6 × 1.5–2.5 µm………Ca. parvispora
-
60.
Collarette 2.5–2.8 µm wide; conidia 5–7 × 2.5 µm………Ch. versicolor
-
61.
Collarette darker than venter………..Ch. bacillaris
-
61.
Collarette and venter concolorous………62
-
62.
Conidia 2.5–5 μm long………63
-
62.
Conidia more than 5 µm long………64
-
63.
Conidia 2.5–4.5 × 1.5–2 µm………Ch. matsushimae
-
63.
Phialides 18–35 μm long; venter 5–12 × 3.5–4.5 μm; conidia 3–5 × 1–1.5 μm………N. strobilina
-
64.
Phialides 65–85 μm long; venter 5–15 × 6–10 μm; collarettes 50–70 × 3–4 μm; conidia 7–15 × 3–4 μm………Ch. myrsines
-
64.
Phialides less than 50 μm long………65
-
65.
Conidia less than 1.5 μm wide………66
-
65.
Conidia more than 1.5 μm wide………67
-
66.
Phialides 34.5–45.5 μm long; venter 18–23 × 3–5 μm; collarettes 16.5–23 × 1.5–2 μm; conidia 4–6.5 × 1–1.5 μm………Ca. pseudoaffinis
-
66.
Phialides 18–42 μm long; venter 8–27 × 4.5–5.5 μm; collarettes 9–24 × 1.5–2.5 μm; Conidia 4.5–6.5 × 1–1.5 µm………..Ca. crassipes
-
67.
Phialides 31–35 μm long; venter 16–17.5 × 5–6 μm; collarettes 14–17 × 2.5–2.7 μm; conidia 6–8 × 2–2.5 μm………Ch. longiphora
-
67.
Conidia more than 7 μm long in average………68
-
68.
Conidia up to 11 μm long………69
-
68.
Conidia up to 17 μm long………70
-
69.
Phialides 26–28 μm long; venter 11–12.5 × 5.7–6.5 μm; collarettes 17–18 × 2.8–3.3 μm; conidia 9–10.5 × 2.2–2.5 μm………Ch. platanicola
-
69.
Conidia 7–11.4 × 1.3–2.1 µm; on Nothofagus………Ca. dualis
-
70.
Conidia 5.5–17 × 1.5–2.5 µm wide, no basal frill………Ch. cylindrosperma
-
70.
Conidia 13–17 × 2.5–3 µm, with basal marginal frill………Ch. nothofagi
Calycina affinis (Sacc. & Berl.) W.P. Wu, comb. nov. Figure 19, MycoBank MB846910.

Calycina affinis (Wu16205). a–j, m–r Conidiophores and phialidic conidiogenous cells. k Superficial mycelium and basal part of conidiophores. l, s Conidia. Scale bars: 10 µm for a–b, 5 µm for c–s
≡ Chalara affinis Sacc. & Berl., Atti Istit. Veneto Sci. lett., ed Arti, Sér. 6, 3: 741, 1885.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores solitary or aggregated, formed from superficial hyphae or aggregated cells, simple, erect, straight or slightly curved, obclavate, 1–2-septate, medium to dark brown, smooth, 37–49 µm long, 4–5 µm wide at the base. Conidiogenous cells integrated, terminal, phialidic, lageniform to obclavate, brown, smooth, 33–43 µm long, transition from venter to collarette abrupt; venter subcylindrical or subellipsoidal, brown, 18–20 × 5–5.5 µm; collarette cylindrical, brown, concolorous with venter or slightly darker in the lower part, 14–18 × 1.8–2 µm; ratio of mean lengths of collarette and venter = 0.8:1. Conidia endogenous, extruded in easily dispersible chains, cylindrical, ends blunt or slightly rounded, hyaline, aseptate, 6–11 × 1.6–1.8 µm; mean conidium length/width ratio = 5:1. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded at margin, aerial mycelium poorly developed, white then become yellow, reverse slightly yellow brown, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Zhejiang Province, Huai An County, on dead leaves of Quercus sp., 18 October 2018, Wenping Wu (Wu16142, Wu16143, Wu16205); Zhejiang Province, Huai An County, on dead leaves of Fagus sp., 18 October 2018, Wenping Wu (Wu16147). Living strains: 76333 (from Wu16142a), 76334 (from Wu16142b), 76335 (from Wu16142c), CGMCC3.23419 (= 76338, from Wu16147a), 76372 (from Wu16147b), 76404 (from Wu16143a), 76405 (from Wu16143b), 76421 (from Wu16205), 76422 (from Wu16205a) and 76476 (from Wu16205b).
Ecology/substrate/host: Saprobes on dead leaf of Aesculus, Beilschmiedia, Corylus, Fagus, Eucalyptus, Laurus, Nothofagus, Pinus, Quercus, Rhododendron, Sycopsis and undetermined plants.
Geographical distribution: British Isles, China, Czech Republic, Italy, New Zealand, North America and UK (Nag Raj and Kendrick 1975; Kirk 1981; Holubová-Jechová, 1984; Li et al. 2013a, b; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975), Kirk (1981), and Holubová-Jechová (1984).
Notes: Calycina affinis is characterized by 1–2-septate conidiophores, obclavate phialidic conidiogenous cells with subcylindrical or subellipsoidal venter, and aseptate and cylindrical conidia (6–11 × 1.6–1.8 μm) without basal frill. It resembles C. brevipes, C. dualis and C. riisgaardii. Morphologically it differs from C. brevipes in having subcylindrical venters, shorter collarettes than venters (0.8:1) and longer and wider conidia that lack basal marginal frill, from C. dualis by the main stalk of conidiophores with only 1–3 cells, and from C. riisgaardii by shorter phialides (33–43 µm long in C. affinis vs. 45–55 µm in C. riisgaardii; Nag Raj and Kendrick 1975; McKenzie et al. 2002). This fungus found in China slightly differs from those reported by Nag Raj and Kendrick (1975) in phialidic conidiogenous cell with a basally fuliginous collarettes and narrower range of conidial size. The slightly darker basal parts of collarettes were observed in the four studied collections. It represents a new recorded species for China.
Identical ITS sequences were generated from the ten studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 76422 included Chalara pseudoaffinis (NR_154761, 98% identity), Calycina herbarum (JN033407, 97% identity) and C. dualis (EF029209, 97% identity).
Three living strains under the name Calycina affinis were found in CBS fungal collection, and the partial LSU sequences from two of those strains (CBS562.77 and CBS620.75) were available in GenBank and CBS fungal collection database. Based on our phylogenetic analyses and morphological study, the strain CBS562.77 was identified as C. affinis. Both ITS and LSU sequences were generated from the CBS620.75 (isolated from Tremella sp. on decaying wood from Netherland) in this study, and the generated LSU sequence is identical to the one deposited in GenBank. Phylogenetically this strain clusters together with other species of Chalara s. lat. with septate conidia. The closest match is Calycellina leucella (MT231682, 98% identity), for which no anamorph was reported in literature. Most likely this strain represents a new species of the genus Nagrajchalara, however no sporulation was observed in pure culture in this study.
Calycina brevipes (Nag Raj & W.B. Kendr.) W.P. Wu, comb. nov., Fig. 20. MycoBank MB846911.

Calycina brevipes (Wu8280). a–j, o Conidiophores and conidiogenous cells. k–n Conidia. Scale bar: 10 µm for a, b; 5 µm for c–o
≡ Chalara brevipes Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 95, 1975.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 1.5–2.5 µm wide. Anamorph: Stroma absent or poorly developed with a few aggregated cells. Setae absent. Conidiophores solitary or irregularly aggregated, erect, straight or slightly curved, consisting of a basal cell and a phialide; the basal cell cooperated into superficial hyphae, up to 8 µm long and 5 µm wide, medium brown, smooth. Conidiogenous cells integrated, terminal, determinate, erect, straight or slightly curved, obclavate, lageniform, 40–50 µm long, medium brown, smooth; transition from venter to collarette abrupt; venters ellipsoidal, narrow ovoid, 15–17 × 5–6 µm; collarettes cylindrical, 22–25 µm long, 2–2.2 µm wide, lower part slightly darker than upper part and venter, smooth; ratio of mean lengths of collarettes and venters = 1.5:1. Conidia endogenous, extruded in easily dispersible chains, cylindrical, 7–10 × 2–2.2 µm, ends truncate or obtuse, hyaline, aseptate; mean conidium length/width ratio = 4:1. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded at margin, aerial mycelium poorly developed, white, then slightly yellow brown, reverse while to slightly yellow brown, sterile, up to 9 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Hubei Province, Shennongjia, on dead leaves of unidentified broad leaf tree, 18 September 2004, Wenping Wu (Wu8280, Wu8302). Living strains: CGMCC3.23381 (= 50639, from Wu8280) and 50681 (from Wu8302).
Ecology/substrate/host: Saprobe on dead leaves of Podocarpus and other plants.
Geographical distribution: Argentina, China and New Zealand (Nag Raj and Kendrick 1975; Gaumundi et al. 1977).
Description and illustration: Nag Raj and Kendrick (1975) and Gamundi et al. (1977).
Notes: Calycina brevipes is characterized by the reduced conidiophore consisting of one basal stalk cell and a terminal phialidic conidiogenous cell, the clearly differentiated ellipsoidal venter and cylindrical collarette with darker lower part, and aseptate, cylindrical conidia (Nag Raj and Kendrick 1975). Morphologically it is similar to C. affinis, but differs in having ellipsoidal venter, collarettes longer than venter (1.5:1 in C. brevipes vs. 0.8:1 in C. affinis) and wider conidia (2–2.2 µm in C. brevipes vs. 1.6–1.8 µm in C. affinis).
The two Chinese collections of this fungus were with slightly shorter but wider conidia than those from the type specimen (conidia 6–12 × 1.5–2 µm). It represented a new record for China. ITS sequences of this species were generated for the first time from the two studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 50681 included C. pseudoaffinis (NR_154761, 97% identity), C. dualis (EF029209, 96% identity) and Calcycina herbarum (JN33407, 95% identity).
Calycina brevispora (Nag Raj & W.B. Kendr.) W.P. Wu, comb. nov., MycoBank MB846912.
≡ Chalara brevispora Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera: 97, 1975.
Description on the natural substrate: Anamorph: Conidiophores cylindrical, 41–145 μm long, multiseptate, dark brown, smooth. Conidiogenous cells phialidic, obclavate to lageniform, 19–45 × 2–3 μm, light brown; venter cylindrical, 10–24 × 3.5–6 μm; collarette cylindrical 9–20 × 2–3 μm; transition from venter to collarette abrupt. Conidia mostly in chains, cylindrical or short cylindrical, ends rounded or blunt, aseptate, hyaline, 2–4 × 1.5–2 μm. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on Quercus sp.
Geographical distribution: Canada, Cuba and Czechoslovakia (Nag Raj and Kendrick 1975; Holubová-Jechová 1984).
Description and illustration: Nag Raj and Kendrick (1975) and Holubová-Jechová (1984).
Notes: Calycina brevispora is characterized by well-developed conidiophores consisting of a multiseptate stalk and a terminal phialide, cylindrical venter, and aseptate conidia in small size (Nag Raj and Kendrick 1975). Only partial SSU sequence from the CBS95.94 was available in GenBank, and a megablast search of GenBank nucleotide database showed highest identity with Chalara aurea CBS880.73, which is most likely identical to C. longipes. Its position in Chalara needs to be further confirmed when additional DNA sequences of other marker genes are available.
Calycina crassipes (Preuss) W.P. Wu, comb. nov., Fig. 21. MycoBank MB846913.

Calycina crassipes (CBS829.71) on PDA in weeks (25 degrees). a–l, r, s Conidiophores and conidiogenous cells. m–q Conidia. Scale bar: 10 μm for a–f; 5 μm for g–s
≡ Cylindrosporium crassipes Preuss, Linnaea 24: 106, 1851.
≡ Chalara crassipes (Preuss) Sacc., Syll. Fung. (Abellini) 4: 335, 1886.
Culture characteristics: Colony on PDA in 4 weeks effuse, rounded at margin, aerial mycelium poorly developed, white, reverse concolorous or slightly pale brown, fertile in the margin, up to 13 mm on PDA at 25 °C in 4 weeks. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae, solitary or in small group, erect or slightly curved, simple, cylindrical, 29–75 µm long, 3.5–4.5 µm wide at the base, obclavate, 1–6-septate, brown to dark brown, smooth-walled, often with 1–3 percurrent proliferations. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 24–30 µm long, 3.5–4.5 µm wide at the base, brown, smooth; transition from venter to collarette gradual; venter subcylindrical, ellipsoidal, 14–17 µm long, 4–5.5 µm wide; collarette cylindrical, 12–16 × 2.2–2.6 µm, concolorous with venter; ratio of mean lengths of collarettes and venter = 0.9:1. Conidia endogenous, extruded singly or in easily dispersible chains, short cylindrical, clavate, hyaline, aseptate, 3.8–4.8 × 1.5–2.8 µm, apex obtuse, base truncated and with short basal frill; mean conidium length/width ratio = 2:1. Teleomorph: Unknown.
Material examined: CBS82971, isolated from dead petiole of Pteridium aquilinum from Netherland.
Ecology/substrate/host: Saprobe on dead conifer wood and petiole of Pteridium aquilinum.
Geographical distribution: Germany, Netherland and UK (Nag Raj and Kendrick 1971, 1975; Holubová-Jechová 1984).
Description and illustration: Nag Raj and Kendrick (1971, 1975) and Holubová-Jechová (1984).
Notes: Three strains under the name Chalara crassipes were preserved in the CBS fungal collection, including CBS 829.71 (from dead petiole of Pteridium aquilinum), CBS 216.84 (from dead leaf of Quercus petraea) and CBS 121.95 (contaminant of Pezizella subtilis strain). The partial LSU sequences were available in GenBank for CBS 216.84 and CBS 829.71, but they were very different and most likely these two strains represented different species. The strain CBS 829.71 was studied in pure culture, it sporulated well in PDA after two weeks. The above description was based on study of this strain. Compared with the lectotype on natural substrate, this strain produced slightly shorter but wider conidia (Nag Raj and Kendrick 1975).
ITS sequence was generated from the studied strain CBS82971. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain were Lanzia sp. (KP204262, 97% identity), Chalara sp. (HQ630988, 97% identity) and many unnamed fungi of Leotiomycetes. In the phylogenetic trees (Figs. 8, 9), this strain showed close relationship with C. parvispora with aseptate conidia.
Calycina dualis (Aramb. & Gamundí) W.P. Wu, comb. nov., MycoBank MB846914.
≡ Chalara dualis Aramb. & Gamundí, in Arambarri, Gumundí & Bucsinszky, Darwiniana 23: 333, 1981.
Description on the natural substrate: Anamorph: Conidiophores cylindrical, 16–60 × 3.8–4.5 µm, up to 7-septate. Conidiogenous cells lageniform to obclavate, 35–51 µm long; transition from venter to collarette abrupt; venter subcylindrical, 14.4–19.2 × 4.3–5.7 µm; collarette cylindrical, 21.6–31.6 × 2.2–2.8 µm. Conidia hyaline, aseptate, cylindrical, 7–11.4 × 1.3–2.1 µm, with flattened ends (Arambarri et al. 1981). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves of Nothofagus dombeyi.
Geographical distribution: Argentina and New Zealand (Arambarri et al. 1981).
Description and illustration: Arambarri et al. (1981).
Notes: Calycina dualis is characterized by well-developed conidiophores consisting of a multiseptated basal stalk and a terminal phialide with abrupt transition from venter to collarette, and aseptate, cylindrical conidia (Arambarri et al. 1981). It resembles Chalara cylindrosperma but differs in having longer venter and collarettes (Nag Raj and Kendrick 1975). The ITS sequence from the New Zealand strain is unique among the existing Chalara species.
Calycina eucalypticola (Crous) W.P. Wu, comb. nov., Fig. 22. MycoBank MB846915.

Calycina eucalypticola (Wu1483d). a–k, o Conidiophores and conidia. l–n, p Conidia. Scale bar: 10 µm for a–f, 5 µm for h–p
≡ Chalara eucalypticola Crous, in Crous et al., Persoonia 43: 265, 2019.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, solitary or in small group, erect, straight or slightly curved, 1–4-septate, obclavate, 45–60 µm long, 5–6 µm wide at the base, medium brown, smooth. Conidiogenous cells integrated, terminate, phialidic, obclavate, lageniform, 38–50 µm long, medium brown, smooth; transition from venter to collarette abrupt or gradual; venters subcylindrical, 21–26 µm long, 6.5–7.5 µm wide, brown, thin- and smooth-walled; collarettes cylindrical, 15–22 × 3.5–3.8 µm, versicolorous, slightly darker in the transition part between venter and collarettes; ratio of mean lengths of collarette and venter = 0.8:1. Conidia endogenous, extruded in short chains, cylindrical, 10–17 × 2.5–2.7 µm, hyaline, uniseptate, both ends truncate or flattened; mean conidium length/width = 5.2:1. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded at margin, aerial mycelium poorly developed, white, then pale to medium brown in the middle with white margin, reverse concolorous, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Guangxi Province, Shiwandashan, On dead leaves and fruits of Quercus sp., 28 Dec 1997, Wenping Wu, Wu1483d. Living strains: CGMCC3.23358 (= NN 43033) and 43157 (from Wu1483d).
Ecology/substrate/host: Saprobe on dead fruit of Quercus sp.
Geographical distribution: China and South Africa (Crous et al. 2019).
Description and illustration: Crous et al. (2019).
Notes: Calycina eucalypticola is a recently described species with reduced conidiophores and 0–1-septate conidia (Crous et al. 2019). The fungus from the Chinese specimen was uniformly with 1-septate conidia. In the phylogenetic trees generated from different datasets (Figs. 2, 3, 8), C. eucalypticola clearly clustered together with all species of Calycina cortegadensis as a sister group of Bloxamia. It is the only exceptional species with septate conidia in Calycina and Chalara s. str.
ITS sequences from the Chinese material were with only 3 bp differences from the one generated from the ex-type strain. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 43033 included C. eucalypticola (MN562114, 99% identity), Calycina cortegadensis (MN017444, 93% identity), Calycina alstrupii (NR_154846, 92% identity) and many unnamed fungi of Leotiomycetes.
Calycina fungorum (Sacc.) W.P. Wu, comb. nov., MycoBank MB846916.
≡ Cylindrium fungorum Sacc., Atti Soc. Veneto-Trent. Sci. Nat. 2: 225, 1873.
≡ Chalara fungorum (Sacc.) Sacc., Michelia 1: 80, 1877.
Description on the natural substrate: Anamorph: Conidiophores obclavate, subcylindrical, reduced to phialides or with 1–2 indistinct septa. Conidiogenous cells lageniform to obclavate, 25–45 × 3–6.5 µm; transition from venter to collarette gradual; venter subcylindrical, 12–20 × 5.5–7 µm; collarette cylindrical, 11–20 × 2.5–4 µm. Conidia hyaline, aseptate, cylindrical, with flattened ends, 5.5–8 × 2–2.5 µm (Nag Raj and Kendrick 1975). Teleomorph: Unknown.
Materials examined: Germany, Eifel, Gerolstein, isolated from decaying needles of Pinus sylvestris, beside Pezizella subtilis, September 1980, W. Gams, CBS 405.81; Sweden, Stockholm, isolated from damaged root of Picea abies, L. Beyer, CBS 942.72.
Ecology/substrate/host: Saprobes on Hydnum compactum, Picea abies and Pinus sylvestris.
Geographical distribution: Germany, Italy and Sweden (Nag Raj and Kendrick 1975; CBS strains database).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Calycina fungorum was redescribed by Nag Raj and Kendrick (1975). Among the studied CBS strains of Chalara species, two strains identified as Chalara fungorum (CBS 240.82 and CBS 942.72) and one strain identified as C. sessilis (CBS 405.81) had identical LSU sequences. In this study, identical ITS sequences were generated from the two strains, CBS 942.72 and CBS405.81.
Calycina lactea (Sacc.) Baral, R. Galán & Platas, Mycosystema 32(3): 423, 2013.
≡ Dasyscyphus lacteus Sacc. (as 'Dasyscypha lactea’), Syll. Fung. (Abellini) 8: 436, 1889.
≡ Bisporella lactea (Sacc.) Stadelmann, Nova Hedwigia 30: 830, 1979 (1978).
≡ Helotium lacteum Ellis & Everh., J. Mycol. 4(6): 56, 1888.
≡ Hymenoscyphus ellisii Dennis, Persoonia 3(1): 48, 1964.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: UK (Friggens et al. 2017).
Notes: The chalara-like anamorph with smaller conidia (2–2.5 × 2 μm) was reported by Friggens et al. (2017).
Calycina oxenbolliae W.P. Wu & Y.Z. Diao, sp. nov., Fig. 23, MycoBank MB845206.

Calycina oxenbolliae (Wu8271b, holotype). a Basal stroma. b–j Conidiophores and phialides. Scale bar: 10 μm for a, b; 5 µm for c–j
Etymology: Named after the former leader Karen Oxenbøll in Novo Nordisk, who gave strong support to this study.
Typification: China, Hubei Province, Wufengshan, on dead leaves of unidentified tree, 10 September 2004, Wu Wenping, Holotype HMAS 352178 (= Wu8271b1), ex-type strain CGMCC3.23450 (= NN50675).
Description on the natural substrate: Colonies effuse, pale brown, superficial, hairy. Mycelium partly immersed and partly superficial, composed of pale to medium brown, septate and branched hyphae with smooth and thin walls, 2–3.5 µm wide. Anamorph: Stroma present, composed of dark brown, thick-walled and irregularly shaped cells. Setae absent. Conidiophores reduced to conidiogenous cells, solitary or aggregated, arising from the basal stroma, erect, straight or slightly curved, simple, obclavate, 20–30 µm long, dark brown. Conidiogenous cells directly arising from the basal stroma, erect, straight, obclavate, lageniform, 20–30 µm long, brown to dark brown, versicolorous, darker in the lower part of collarettes, smooth-walled; transition from venter to collarette abrupt; venters bulbous, subglobose, 5–8 µm long, 4.5–5.5 µm wide; collarettes cylindrical, 20–23 × 1.2–1.5 µm; ratio of mean lengths of collarette and venter = 3.3:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 5–6 × 1–1.2 µm, hyaline, aseptate, smooth- and thin-walled, both ends obtuse or flattened; mean conidium length/width = 4.6:1. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded at margin, aerial mycelium poorly developed, brown to dark brown, with lighter margins, reverse concolorous, with pale yellow brown pigment diffused in agar, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Calycina oxenbolliae can easily be distinguished from the existing Calycina and Chalara s. lat. species by presence of basal stroma, obclavate conidiogenous cells directly arising from basal stroma, bulbous venters, and small-sized conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002).
ITS sequences were also generated from the ex-type strain. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 50633 included the endophytic fungus of Calluna vulgaris (FM172800, 94% identity), Calycellina fagina (OL752703, 94% identity) and many unnamed fungi of Leotiomycetes.
Calycina parilis (P. Karst.) Kuntze, Revis. gen. pl. (Leipzig) 3(3): 448, 1898. Figure 24.

Calycina parilis (Wu7520a). a–e, g, h Conidiophores and phialidic conidiogenous cells. f, i–k Conidia. Scale bar: 5 µm
≡ Helotium parilis P. Karst., Bidr. Känn. Finl. nat. Folk 19: 115, 1871.
≡ Hymenoscyphus parilis (P. Karst.) Dennis, Persoonia 3(1): 74, 1964.
≡ Peziza parilis P. Karst., Not. Sällsk. Fauna et Fl. Fenn. Förh. 10: 146, 1869.
≡ Pezizella parilis (P. Karst.) Dennis, Mycol. Pap. 62: 49, 1956.
Description on the natural substrate: Colonies effuse, pale brown, sparse. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, smooth-walled, branched hyphae of 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores macronematous, micronematous, arising from superficial hyphae, solitary or aggregated loosely, erect, straight or slightly curved, simple, 1–3-septate, subcylindrical, 40–50 × 5–6 µm, medium brown, thick- and smooth-walled, basal part dark brown and often with 1–2 percurrent proliferations. Conidiogenous cells integrated, terminal, straight or slightly curved, obclavate, lageniform, 35–45 µm long, medium brown, versicolorous, slightly darker in the lower part of collarettes, thin- and smooth-walled; transition from venter to collarette abrupt; venters cylindrical to subcylindrical, 20–25 × 6–7 µm, medium brown, smooth- and thin-walled; collarettes cylindrical, 18–22 × 2.5–3 µm, medium brown; ratio of mean lengths of collarettes and venter = 0.9:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 6.5–11 × 2–2.5 µm, hyaline, aseptate, smooth- and thin-walled, base truncate with a minus frill, apex truncate or flattened, guttulate; mean conidium length/width ratio = 3.9:1. Teleomorph: Unknown.
Material examined: China, Yunnan Province, Baoshan, Tengchong, Gaoligongshan, on dead leaves of unidentified tree, 17 October 2003, Wenping Wu, HMAS 352176 (= Wu7520a), CGMCC3.23375 (= NN47837); Yunnan Province, Baoshan, Tengchong, Gaoligongshan, on dead leaves of unidentified tree, 17 October 2003, Wenping Wu, Wu7569. Living strain: 47902 (from Wu7569).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China, Czech Republic and UK (Friggens et al. 2017).
Description and illustration: Svrček (1984) and Friggens et al. (2017).
Notes: The chalara anamorph of Calycina parilis was described from both natural substrate and living culture in literature. Its connection with Calycina parilis was based on its co-existing of both anamorph and teleomorph on the natural substrate. This needs to be proved experimentally in future study (Dennis 1956; Svrček 1984; Friggens et al. 2017). The two specimens with identical ITS sequences were assigned to C. parilis in this study.
Calycina parilis is characterized by short and 1–3-septate conidiophores with percurrent proliferations, versicolorous phialides, abrupt transition from venters to collarettes, and aseptate conidia. Chalara sessilis (phialides 18–23 μm long; venters 10–12.5 × 3–4 μm, collarettes 10–11 × 1.5 μm, conidia 3.5–6 × 1.5 μm) resembles C. parilis in morphology of conidiophores and conidia, but differs by its smaller phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Compared to the record Calycina parilis from UK (collarettes 23–27 × 4–5.5 μm; conidia 7–9 × 3.5–4 μm),, the Chinese specimens of this fungus produced shorter collarettes and narrower conidia (Friggens et al. 2017).
Both specimens from this study were collected in the same locality (Tengchong, Yunnan) and their ITS sequences from the relevant strains were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 47837 included Chalara anamorph of Calycina parilis (MF110620, 100% identity), Graphilbum sp. (MZ494998, 98% identity), Calycina sp. (OM456212, 97% identity), Calycina languida (F116599, 97% identity) and Graphilbum pleomorphum (MH861928, 97% identity).
Calycina parvispora (Nag Raj & S. Hughes) W.P. Wu, comb. nov., Fig. 25. MycoBank MB846917.

Calycina parvispora (CBS385.94) on PDA in weeks (25 degrees). a–c, h–l Conidiophores and conidiogenous cells. d–g Conidia. Scale bar: 10 μm for a; 5 μm for b–l
≡ Chalara parvispora Nag Raj & S. Hughes, N.Z. Jl Bot. 12:117, 1974.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, white, then pale to medium brown, reverse yellow brown, with paler margin, sterile, up to 14 mm on PDA at 25 °C in 4 weeks. Anamorph: Conidiophores formed from superficial hyphae, erect, straight of slightly flexuous, 2–5-septated, cylindrical, obclavate, 45–65 × 4–5 μm, dark brown, smooth, terminating in a phialides, occasionally with 1–2 percurrent proliferations. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, lageniform, 31–42 μm long, pale brown to medium brown, smooth, consisting of a venter and a collarette, concolorous; transition from venter to collarette gradual; venters subcylindrical, 19–25 × 4.5–5.6 µm, medium brown; collarettes cylindrical, 16–20 × 2.7–3 µm, pale to medium brown; ratio of mean lengths of collarette and venter = 0.8:1. Conidia endogenous, extruded singly or in persistent chains, short-cylindrical, 3.8–5.4 × 2.4–2.5 µm, base truncate with frill, apex rounded or flattened, hyaline, aseptate; mean conidium length/width ratio = 1.9:1. Teleomorph: Unknown.
Material examined: Czechoslovakia, isolated from decaying plant material, 1971, V. Holubová-Jechová, CBS 385.94.
Ecology/substrate/host: Saprobe on dead plant material.
Geographical distribution: Czechoslovakia and New Zealand.
Description and illustration: Nag Raj and Hughes (1974) and Nag Raj and Kendrick (1975).
Notes: Calycina parvispora is characterized by long and multiseptated conidiophores, terminal lageniform conidiogenous cells with abrupt transition from venter to collarette, and hyaline, aseptate, short-cylindrical conidia in small size (conidia 3.5–6 × 1.5–2 µm from the holotype; Nag Raj and Kendrick 1975). The studied strain CBS385.94 sporulated well on PDA, and the above description was based on the observation of pure culture. Compared with the morphology from natural substrate (conidiophores multiseptated, 95–190 μm long; conidiogenous cells lageniform, 34–43 μm long, transition from venter to collarette abrupt, venters subcylindrical and 15–20 × 4–7 µm, collarettes 18–26 × 1.5–2.5 µm; conidia short-cylindrical, 3.5–6 × 1.5–2 µm), the fungus in pure culture produced shorter conidiophores, slightly longer venter, and wider conidia (Nag Raj and Kendrick 1975).
ITS sequence was also generated from the studied strain. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain CBS 385.94 included Lanzia sp. (KP204262, 97% identity), Chalara sp. (HQ630988, 97% identity), Chalara sp. (MT133992, 97% identity), Calycina sp. (OM456212, 96% identity), Mollisia caespiticia (KY965813, 95% identity), and many other unnamed fungi of Leotiomycetes. In addition, it also showed high identity (with 7 bp differences) with the ITS sequence generated from C. crassipes CBS829.71 in this study.
Calycina pseudoaffinis (Koukol) W.P. Wu, comb. nov., MycoBank MB846918.
≡Chalara pseudoaffinis Koukol, Fungal Diversity 49: 86, 2011.
Description on the natural substrate: Anamorph: Conidiophores septate, (45)56–107(130) μm long and 3–4 μm wide. Conidiogenous cells narrowly lageniform, gradually tapering towards the collarette, (26.5)34.5–45.5(52.5) μm long and 3–5 μm wide; venter cylindrical, (15.5)18–23.5(27) μm long; collarette cylindrical, light brown, (12.5)16.5–23(25) μm long and 1.5–2 μm wide. Transition from venter to collarette gradual, sometimes hardly perceptible. Conidia cylindrical with rounded ends, one-celled, hyaline, 4–6.5 × 1–1.5 μm (Koukol 2011). Teleomorph: Unknown.
Ecology/substrate/host: Saprobes on surface of mite extracted from Pinus.
Geographical distribution: Czech Republic (Koukol 2011).
Description and illustration: Koukol (2011).
Notes: Calycina pseudoaffinis is a recently described species from surface of mite extracted from Pinus sylvestris litter (Koukol 2011). It is morphologically comparable to C. affinis, but differs by multiseptate and longer conidiophores, and smaller conidia.
Calycina riisgaardii W.P. Wu, sp. nov., Fig. 26, MycoBank MB845210.

Calycina riisgaardii (Wu7077 holotype). a–d, g–n Conidiophores and phialidic conidiogenous cells. e, f Basal part of conidiophores with percurrent proliferations. o–r Conidia. Scale bars: 10 µm for a–d, 5 µm for e–r
Etymology: Named after the former CEO of Novozymes A/S, Steen Riisgaard, who made great contribution in industrial biotechnology.
Typification: China, Yunnan Province, Kunming, Heilongtan, on dead branches of Quercus sp., 19 October 2003, Wenping Wu, Holotype HMAS 352181 (= Wu7077), ex-type strain CGMCC3.23369 (= NN47715).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin– or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores scattered or 2–3 aggregated at the base, directly arising from superficial hyphae, erect, straight or flexuous, simple, obclavate, subcylindrical, 50–65 µm long, 3.5–4.5 µm wide at the base, basal cells slightly swollen, dark brown at the base, becoming medium brown towards the upper part, 1–3-septatea, smooth, often with one to several percurrent proliferations at the base, terminating in a phialide. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, obclavate, subcylindrical, 45–55 µm longer, medium brown, become pale brown towards the apex, composed of a venter and a collarette; transition from venter to collarette abrupt; venters subcylindrical to subellipsoidal, slightly verruculose, 16–30 × 4.5–5 µm; collarettes cylindrical, 23–30 × 2–2.5 µm, brown, basal part slightly darker than upper part and venter, verruculose; ratio of mean lengths of collarettes and venter = 1.2:1. Conidia endogenous, extruded in short chains, cylindrical, 6–7 × 1–2.5 µm, straight, subtruncate at both end or slightly rounded at the apex, without a basal frill, hyaline, aseptate, smooth; mean conidium length/width ratio = 3.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed at margin, aerial mycelium poorly developed, constricted at surface, soil brown, reverse concolorous or slightly darker, with yellow pigment diffused into agar, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead branches of ?Quercus sp.
Geographical distribution: China.
Notes: Calycina riisgaardii is characterized by short conidiophores with percurrent proliferation, and consisting of a basal stalk and a phialide, abrupt transition from venter to collarette, verruculose phialides, and aseptate conidia. Chalara nigricollis and C. prolifera also produced short conidiophores with multiple percurrent proliferations, but the conidia in these two species were uniseptate. Calycina riisgaardii morphologically resembled C. affinis, C. crassipes and Chalara bohemica in morphology of conidiophores, phialides and conidia, but differed in size of phialides and conidia (Nag Raj and Kendrick 1975). In C. bohemica, the conidiogenous cells were with shorter but wider venters (6–18 µm long, 6–9.5 µm wide) and larger conidia (9.5–13 × 2–3 µm). In C. crassipes, the conidiogenous cells were in shorter size (18–42 µm long) and the conidia were in similar length but narrower (4.5–6.5 × 1–1.5 µm). In C. affinis, the conidiophores (33–81 µm long), conidiogenous cells (28–54 µm long) and conidia (5.5–19 × 1.5–2.5 µm) were with broad variation.
Based on a megablast of GenBank nucleotide database, the closest matches to the ex-type strain 47715 included Chalara dualis (EF029209, 98% identity), Calycina herbarum (JN033407, 96% identity), C. pseudoaffinis (NR_154761, 96% identity), Graphilbum pleomorphum (MH861928, 96% identity), Pezizella discreta (JF908571, 96% identity), and many unidentified fungi of Leotiomycetes.
Calycina vulgaris (Fr.) Baral, Beitr. Kenntn. Pilze Mitteleur. 5: 227, 1989.
≡ Peziza vulgaris Fr., Syst. Mycol. (Lundae) 2(1): 146, 1822.
≡ Pezizella vulgaris (Fr.) Sacc., Syll. Fung. (Abellini) 8: 278, 1889.
For other synonyms, see Baral (1989) and MycoBank.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: Widely distributed (Baral 1989).
Description and illustration: The chalara-like anamorph by Morozova (2014).
Notes: Chalara anamorph of Calycina vulgaris was illustrated by Morozova (2014), however, no description was provided.
Chalara (Corda) Rabenh. s. str., Deutschl. Krypt.-Fl. (Leipzig) 1: 38, 1844.
≡ Torula subgen. Chalara Corda, Icon. Fung (Prague) 2: 9, 1838.
= Chaetochalara B. Sutton & Piroz., Trans. Br. mycol. Soc. 48: 350, 1965.
Emended description: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to brown, smooth, septate and branched hyphae. Anamorph: Stroma absent or present; if present, consisting of aggregated cells. Setae absent or present; if present, solitary, erect, straight or flexuous, simple, cylindrical, brown to dark brown, septate, tapering towards apex, acute or obtuse at the apex. Conidiophores macronematous, micronematous, scattered or aggregated from the basal stroma, erect, simple, or rarely branched, subcylindrical, pale brown, brown to very dark brown, smooth or verrucose, septate, consisting of a basal cell or septate stalk and a terminal phialide, with or without percurrent proliferations. Conidiogenous cells ampulliform, lageniform, obclavate, ellipsoidal, urceolate or subcylindrical, pale brown to dark brown, composed of a venter and a collarette, transition from venter to collarette gradual, abrupt or barely perceptible. Conidia holoblastic, cylindrical, obclavate, straight, hyaline, aseptate, smooth, with rounded or truncate ends, often with basal marginal frill or rarely fringes of wall material slightly rounded at the apex, extruded in short chains. Teleomorph: unknown.
Type species: Chalara fusidioides (Corda) Rabenh.
Ecology/substrate/host: Saprobe on decaying plant materials including leaves, branches, stems, fruits and wood from terrestrial and submerged environments.
Geographical distribution: Widely distributed in worldwide (Nag Raj and Kendrick 1975).
Description and illustration: Ellis (1971) and Nag Raj and Kendrick (1975).
Notes: Based on the morphological and phylogenetic analyses in this study (Figs. 1, 2, 3, 8, 15, 16, 17, 18), the genus Chalara, typified by C. fusidioides, is emended as a monophyletic genus in narrower concept to accommodate eight chalara-like fungi (Chalara africana, C. bambusicola, C. clidemiae, C. cylindrosperma, C. longiphora, C. platanicola and C. qinlingensis), which clustered as a strongly supported clade distinct from other chalara-like fungi. Morphologically the emended Chalara s. str. is characterized by solitary and unbranched conidiophore, terminal phialide consisting of a venter and a cylindrical collarette with deeply seated sporulating locus, and hyaline, aseptate and cylindrical conidia. Morphologically several other chalara-like genera such as Calycina, Cylindrochalara, Leochalara, Minichalara, Stipitochalara, Constrictochalara and anamorphs of Hymenoscyphus, share some similarity with Chalara s. str., but can be distinguished by different combinations of morphological characters such as conidiophores, conidiogenous cells and conidia. Furthermore, the separation of these genera are strongly supported by the phylogenetic analyses. In Leochalara and Minichalara, the conidiophores and phialides are subhyaline to pale brown, and transition from venters to collarettes are gradual; in Constrictochalara the phialides are constricted between venters and collarettes, and the conidia are clavate or not typical cylindrical. Several species of Hymenoscyphus produce chalara-like anamorphs, but differs from Chalara by sessile conidiogenous cells, relatively short collarettes, and short-cylindrical or globose conidia. The genus Stipitochalara can hardly be distinguished from Chalara s. str. in morphology, but phylogenetically belong to different family in Leotiomycetes.
However, the above treatment for Chalara s. str. is still problematic since the phylogenetic relationship of the type species C. fusidioides with those eight species was not known. Currently only one SSU sequence from an unauthorized strain of C. fusidioides was available in GenBank (AF203463) and its identity with C. fusidioides was doubtful. Johnston et al. (2019) briefly notified that based on a comparison of SSU sequences, the specimen identified as C. fusidioides (AF203463) could be congeneric with C. longipes which belongs to Hyaloscyphaceae. The phylogenetic analyses with SSU/LSU dataset in this study also points to the same conclusion (Figs. 1, 2). Although it has been recorded with broad distribution in North American and Europe, no living strain was deposited in any of those public culture collections such as ATCC, BCCM, CABI Fungi, CBS, CGMCC, DSMZ, IFO, etc. Thus, the phylogenetic relationship of C. fusidioides and other chalara-like fungi couldn’t be determined. Future studies on collecting of fresh material and epitypification of the type species is needed.
Based on the morphological study and phylogenetic analyses, a total of 17 species were accepted in this emended Chalara s. str. (Figs. 15, 17, 18). Among of them, eight species, C. africana, C. bambusicola, C. clidemiae, C. cylindrosperma, C. longiphora, C. pengii, C. platanicola and C. qinlingensis, were confirmed by the phylogenetic analysis. While the other 9 species, the type species C. fusidioides, C. bacillaris, C. clavatophora, C. cylindrophora, C. kirkii, C. sessilis, C. sinensis, C. sporendocladioides and C. versicolor, were without available of living strains, and thus their phylogenetic relationship with other chalara-like fungi remains to be studied in future. Morphologically the species assigned to Chalara s. str. are very variable in setae, conidiophores, phialides and conidia. Two species, C. africana and C. qinlingensis, were with setae among the conidiophores. Most species were with reduced conidiophores consisting of 1–2-celled basal stalk and a terminal phialide, however long and well-developed conidiophores with multiseptated stalk were also seen in C. brevispora, C. cylindrosperma, C. parvispora and several new species. Conidia of these species were always cylindrical or short-cylindrical, hyaline, aseptate, with obtuse or truncate ends, and formed in short or long easily dispersible chains. The basal frills of conidia were only found in C. brevipes and C. parvispora (Nag Raj and Kendrick 1975).
Chalara africana (B. Sutton & Pirozy.) P.M. Kirk & Spooner, Kew Bull. 38(4): 580, 1984. Figure 27.

Chalara africana (Wu16240). a–d Setae and conidiophores. e–m Conidiophores bearing phialides. n, o conidia. p Developing conidiophores. Scale bar 20 µm for a–c, 5 µm for e–p
≡ Chaetochalara africana B. Sutton & Pirozy., Trans. Br. mycol. Soc. 48(1): 352, 1965.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Setae solitary, erect, straight, simple, up to 100 µm long, cylindrical, slightly inflated at the base to a width of up to 6 µm, gradually tapering towards the acute apex, 0–2-septate, dark brown to blackish, smooth. Conidiophores solitary or aggregated around a seta, erect, simple, clavate, subcylindrical, 30–45 µm long, medium brown, 0–1-septate, smooth. Conidiogenous cells integrated, determinate, phialidic, erect, straight or slightly curved, narrowly ampulliform, obclavate, subcylindrical, 30–40 µm long, medium brown, smooth; transition from venter to collarette gradual; venter subcylindrical, 12–14 µm long, 4–6 µm wide at the base; collarette cylindrical, concolorous with venter, 18–24 µm long, 2.5–3 µm wide; ratio of mean lengths of collarette/venter = 1.6:1. Conidia endogenous, extruded in easily dispersible chains, cylindrical, ends truncate or obtuse, hyaline, aseptate, 5–6.5 × 1.5–2 µm; mean conidium length/width ratio = 4.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, white then grey to grey brown, reverse dark brown with paler colored margin, sterile, up to 15 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangdong Province, Guangzhou, South China Botanical Garden, on dead deed pod of unidentified Leguminosae, 12 February 2018, Wenping Wu (Wu16239, Wu16240); Guangdong Province, Shenzhen, Lianhuashan Park, on dead deed of Acer sp., 11 November 2019, Wenping Wu (Wu17142); Guangdong Province, on dead leaves of unidentified plant, 9 October 1998, Wenping Wu (Wu2083I); Guangxi Province, Nanning, on dead leaves and stem of Magnolia sp., 3 January 1998, Wenping Wu (Wu1620e); Guangxi Province, Nanning, on dead leaves of Quercus sp., 3 January 1998, Wenping Wu (Wu1657a); Yunnan Province, Kunming, Kunming Botanical Garden, on dead leaves of Cinnamomum sp., 25 November 1995, Wenping Wu (Wu1006b). Living strains: 43684 (from Wu1657a), CGMCC3.23423 (= 76574, from Wu16239a), 76575 (from Wu16239b), 76576 (from Wu16240) and 76577 (from Wu16240a).
Ecology/substrate/host: Saprobe on dead leaves and seed pod of Brachystegia, Beilschmiedia, Cinnamomum, Magnolia, Quercus and unidentified trees.
Geographical distribution: China, New Zealand and Zambia (Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975; Wu 2004).
Description and illustration: Sutton and Pirozynski (1965), Nag Raj and Kendrick (1975), and Wu (2004).
Notes: Chalara africana morphologically resembles C. bulbosa and C. qinlingensis in presence of setae, sessile conidiogenous cells arising directly from the fertile hyphae, and cylindrical, aseptate conidia without basal frill (Nag Raj and Kendrick 1975; Sutton and Pirozynski 1965; McKenzie et al. 2002). It differs from C. bulbosa in shape of venter, i.e., bulbous in C. bulbosa vs. subcylindrical in C. africana; differs from C. qinlingensis in large-sized phialides and slightly smaller conidia.
Identical ITS sequences were generated from the five studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 76577 include Prorodiplodia vitis (NR_163376 97% identity), P. livistonae (NR_160355, 97% identity) and Chalara clidemiae (NR_145313, 96% identity). These sequences have 29 bp differences with those from C. qinlingensis.
Chalara africana was reported from China with a single collection by Wu (2004). Since then, six additional specimens were collected from different localities. It is possible that this species is a rather common one in tropical and subtropical areas.
Chalara bacillaris W.P. Wu & Y.Z. Diao, sp. nov., Fig. 28, MycoBank MB845305.

Chalara bacillaris (Wu12285, holotype). a Superficial mycelium. b–e Conidiophores and conidiogenous cells. f–k Conidia. Scale bar: 5 μm
Etymology: Refers to its rod-shaped conidia.
Typification: China, Guangdong Province, Guangzhou, South China Botanical Garden, on dead bark of unidentified tree, 4 March 2012, Wenping Wu, Holotype HMAS 352241 (= Wu12285).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin– or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores scattered or 2–3 aggregated from the basal hyphae, erect, straight, simple, obclavate, lageniform, 32–35 µm long, 3.5–4 µm wide at the base, pale brown to medium brown, becoming paler towards the apex, 1–2 septate, smooth, no percurrent proliferations, terminating in a phialide. Conidiogenous cells integrated, terminal, obclavate, lageniform, 27–32 µm long, medium brown, become pale brown towards the apex, thin- and smooth-walled, composed of a venter and a collarette; transition from venter to collarettes abrupt; venter subcylindrical to subellipsoidal, 10–12 × 3.5–4.5 µm; collarette cylindrical, 16–20 × 1.5–2 µm, concolorous with venter, smooth; ratio of mean length of collarette and venter = 1.6:1. Conidia endogenous, extruded in short chains, rod-shaped, 3–4.5 × 1 µm, both ends truncated, hyaline, aseptate, without a basal frill; mean conidium length/width ratio = 3.8:1. Teleomorph: Unknown.
Material examined: China, Guangdong Province, Guangzhou, South China Botanical Garden, on dead bark of unidentified tree, 4 March 2012, Wenping Wu (Wu12285).
Ecology/substrate/host: Saprobe on dead bark of unidentified tree.
Geographical distribution: China.
Notes: Chalara bacillaris is characterized by reduced conidiophores, pale colored conidiogenous cells, abrupt transition from venter to collarette, and aseptate, rod-shaped conidia in smaller size and with truncated ends. It resembles C. austriaca and C. sessilis, but differs from them by is its longer phialides and shorter conidia (Nag Raj and Kendrick 1975).
Chalara bambusicola W.P. Wu & Y.Z. Diao, sp. nov., Fig. 29, MycoBank MB845201.

Chalara bambusicola (Wu12360, holotype). a–e Conidiophores and phialidic conidiogenous cells. f–j Conidia. Scale bar: 5 µm
Etymology: Refers to its substrate as bamboo.
Typification: China, Guangdong Province, Guangzhou, Guangzhou Botanical Garden, on dead culm of bamboo, 4 March 2012, Wenping Wu, Holotype HMAS 352175 = Wu12360; ex-type strain CGMCC3.23394 (= NN54418).
Description on the natural substrate: Colonies effuse, pale brown, sparse. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, smooth-walled, branched hyphae of 2–3 µm wide. Anamorph: Stroma absent or poorly developed with a few irregularly aggregated cells. Setae absent. Conidiophores macronematous, micronematous, solitary, erect, straight or slightly curved, simple, obclavate, subcylindrical, 30–45 × 3–4 µm, 1–3-septate, pale brown to medium brown, thin- and smooth-walled. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, obclavate, subcylindrical, 27–37 µm long, 3–4 µm wide at the basal widest part, medium brown, smooth-walled; transition from venter to collarette gradual; venter subcylindrical, 13–15 × 3–4 µm, medium brown; collarette cylindrical, 15–20 × 2–2.5 µm, concolorous with venter; ratio of mean lengths of collarettes and venter = 1.3:1. Conidia endogenous, extruded in easily dispersible chains, cylindrical, (5–) 7.5–8.5 × 1–1.2 µm, ends truncate or obtuse, hyaline, aseptate, usually with one guttulate near to one or both ends; mean conidium length/width ratio = 7.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, white, then pale to medium brown, reverse yellow brown, with paler margin, sterile, up to 16 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead culm of bamboo.
Geographical distribution: China.
Notes: Morphologically C. bambusicola is like C. affinis and C. clidemiae in obclavate conidiogenous cells with gradual transition from venters to collarettes, and aseptate, cylindrical conidia without basal frills, but differs in size of conidiogenous cells and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002; Crous et al. 2016b). In C. affinis, the conidiogenous cells are with collarettes shorter than venters, and much wider (5–5.5 µm) venter than those in C. bambusicola. In C. clidemiae, the phialidic conidiogenous cells are shorter (25–30 µm), and the conidia are also shorter (3–5 × 2–2.5 µm) but wider than those in C. bambusicola.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 54418 include Chalara clidemiae (GenBank NR_145313, 95% identity) and many unnamed endophytic fungi of Heliotiales.
Chalara clavatophora W.P. Wu and Y.Z. Diao, sp. nov., Fig. 30a, b, MycoBank MB845202.

Chalara clavatophora (a, b from Wu1052a, holotype), C. cylindrophora (c, d from Wu984, holotype) and C. kirkii (e, f from Wu1042g, holotype). a, c, e Conidia. b, d, f Conidiophores and conidia. Scale bar: 20 μm
Etymology: Refers to its clavate-shaped conidiogenous cells.
Typification: China, Ningxia Province, Jingyuan County, Liupan Mountain, Erlonghe, on wood of unidentified plant, 24 August 1997, Wenping Wu, Holotype HMAS352242 (= Wu1052a).
Description on the natural substrate: Colonies effuse, pale brown, superficial, sparse. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae, 2–3 µm diam. Anamorph: Stroma absent or poorly developed. Setae absent. Conidiophores reduced to phialidic conidiogenous cells. Conidiogenous cells directly arising from cells of superficial hyphae or aggregated cells, scattered or aggregated at the bases, erect, straight, clavate, subcylindrical, 25–35 µm long, pale brown to brown, thin- and smooth-walled, straight; transition from venter to collarette gradual; venter subcylindrical, ellipsoidal, 6–15 × 7–9 µm; collarette cylindrical, 7–10 × 4–5 µm, smooth, concolorous with venter; ratio of mean lengths of collarette and venter = 0.8:1. Conidia endogenous, extruded in short chains, oblong to short-cylindrical, 5–7 × 3.5–4 µm, both ends flattened or subtruncated, hyaline, aseptate, smooth- and thin-walled; mean conidium length/width ratio = 1.6:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on rotten wood.
Geographical distribution: China.
Notes: Chalara clavatophora differs from all existing species by its reduced conidiophores, clavate conidiogenous cells directly arising from superficial hyphae or aggregated cells, gradual transition from venter to collarette, and oblong to short-cylindrical conidia in small size (Nag Raj and Kendrick 1975; McKenzie et al. 2002). There is hardly any species in the genus with similarity to C. clavatophora. In morphology of conidiogenous cells and conidia, C. clavatophora superficially resembles members of Bloxamia, but the latter differs by well-developed sporodochial conidiomata. No living strain is available for molecular phylogenetic study.
Chalara clidemiae Crous & M.J. Wingf. in Crous et al., Persoonia 36: 357, 2016.
Description on the natural substrate: Anamorph: Conidiophores 25–30 × 3–4 μm, 1–5-septate, subcylindrical, lageniform. Conidiogenous cells phialidic, 15–25 × 3.5–4 μm; venter cylindrical. Conidia aseptate, subcylindrical, apex obtuse, base truncate, (3–)4(–5) × (2–)2.5 μm, forming long, curvy chains or slimy masses on older phialides. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on twig of Clidemia hirta.
Geographical distribution: France (Crous et al. 2016b).
Description and illustration: Crous et al. (2016b).
Notes: Calycina clidemiae is a recently described species with short conidiophores and small conidia (Crous et al. 2016a, b). Phylogenetically it is closely related to C. africana, C. bambusicola and C. qinlingensis, but differs in lacking setae and producing short cylindrical conidia.
Chalara cylindrophora W.P. Wu & Y.Z. Diao, nomen. nov., Fig. 30c, d, MycoBank MB845211.
≡ Chalara minima W.P. Wu, Mycosystema 23: 318, 2004 (non Chalara minima Höhn., Öst. bot. Z. 55: 15, 1905).
Etymology: Refers to its cylindrical conidiophores.
Typification: China, Yunnan Province, Kunming, Kunming Botanical Garden, on dead leaves of undetermined leaf litter, 24 November 1995, Wu Wenping, Holotype Wu984a.
Description on the natural substrate: Colonies effuse, pale brown, superficial, hairy. Mycelium partly immersed and partly superficial, composed of pale to medium brown, septate and branched hyphae with smooth and thin walls, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores reduced to phialidic conidiogenous cells. Conidiogenous cells directly arising from superficial hyphae, determinate, scattered or occasionally aggregated at the bases, erect, straight or slightly curved, cylindrical, 40–56 µm long, medium brown to brown, smooth-walled; transition from venter to collarette gradual; venter subcylindrical, 5–8 µm long, 5–6.5 µm wide; collarette cylindrical, 32–50 µm long and 2.5–3 µm wide, concolorous with venter; ratio of mean lengths of collarette and venter = 6.3:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 7.5–10 × 1–1.5 µm, hyaline, aseptate, smooth– and thin–walled, both ends truncate except for the apical conidium which is obtuse at the apex; mean conidium length/width ratio = 7:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China (Wu 2004).
Description and illustration: Wu (2004).
Notes: Chalara minima, originally described by Wu (2004), is an illegitimate name due to existing of earlier name C. minima Höhn. (1904). However, it represents a different fungus (vs. C. minima Höhn.), thus the new name C. cylindrophora is proposed. Among all described species, C. dracophylli (= C. australis McKenzie 1993), C. brunnipes, C. graminicola, and C. phaeospora are morphologically similar to C. cylindrophora on sessile conidiogenous cells, cylindrical phialides without clearly separation between venters and collarettes, and aseptate, cylindrical conidia (Nag Raj and Hughes 1974; McKenzie 1982, 1993; Kirk 1985, 1986; McKenzie et al. 2002). The conidiogenous cells in C. phaeospora are very similar to those in C. minima, however they are much longer (95–125 µm) and wider (14–18 µm at the swollen base and 7–10 µm in other parts), and the conidia are dark brown (Kirk 1985). Chalara brunnipes can be easily distinguished from C. cylindrophora by its longer venter (venter/collarette = 1/0.8) and short clavate conidia in smaller size (3.5–5 × 1.5 µm) (Nag Raj and Kendrick 1975). In C. graminicola, the phialides are much longer (80–140 µm) and wider (15–21 µm wide in venter and 5–6.5 µm in collarette), and the conidia are longer (14–17 µm) and wider (3.5–4.5 µm), thus it can also be easily distinguished from the species described here. Chalara dracophylli was described and illustrated with similar conidiogenous cells, but in this species the conidia are uniseptate and much longer (18–20 × 2–2.5 µm).
Chalara cylindrosperma (Corda) S. Hughes, Can. J. Bot. 36: 747, 1958. Figure 31.

Chalara cylindrosperma (a, d from Wu969; b, c from Holotype in IMI; e, f from Wu987). a, c, e Conidia. b, d, f Conidiophores and conidiogenous cells. Scale bar: 20 μm for a, c, e; 40 μm for b, d, f
≡ Menispora cylindrosperma Corda, Icon. Fung. (Prague) 1: 16, 1837.
= Cylindrotrichum inflatum Bonord., Handb. Allgem. mykol. (Stuttgart): 88, 1851.
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent or poorly developed with a few aggregated cells. Setae absent. Conidiophores scattered or aggregated at the base, erect, straight or curved, simple, long clavate, subcylindrical, basal cells slightly swollen and lobed, medium to dark brown, formed in two layers with short and long conidiophores; the shorter conidiophores up to 75 µm high, 3.5–4.5 µm wide, pale brown to medium brown, 1–7- septate; the longer conidiophores up to 150 µm high, 3–4 µm wide, dark brown to blackish at the lower part, 8–12- septate. Conidiogenous cells integrated, terminal, lageniform, pale to medium brown, transition from venter to collarette abrupt; venter ellipsoid to subcylindrical, 10–12.5 × 5–7 µm, widest at the transition from venter to collarette; collarette cylindrical, 17.5–20 × 2.5–3.5 µm, concolorous with venter; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, cylindrical, truncate or obtuse at both ends, 10–17 × 1.5–2.5 µm, hyaline, aseptate; mean conidial length/width ratio = 6.8:1. Teleomorph: Unknown.
Materials examined: China, Hubei Province, Shennongjia, on dead leaves of unidentified plant, September 15 2004, Wenping Wu, Wu8177; Yunnan Province, Kunming, Jindian Park, on dead leaves of Cinnamomum sp., 24 November 1995, Wenping Wu, Wu974a; Yunnan Province, Kunming, Kunming Botanical Garden, on dead leaves of Eriobotrya japonica., 25 November 1995, Wenping Wu, Wu970c; China: Yunnan Province, Kunming, Kunming Botanical Garden, on dead leaves of undetermined tree, 25 November 1995, Wenping Wu, Wu969a and Wu987. Living strains: 50499 (= CBS658.79) and 50590 (from Wu8177).
Ecology/substrate/host: Saprobe on dead leaves, branches, rotting wood, decaying fruit, decaying cupules of Aconitum, Aesculus, Agathis australis., Betula alba–sinensis, Eriobotrya japonica., Fagus sylvatica, Ilex., Podocarpus dacrydioides, P. totara, and other plants.
Geographical distribution: Austria, Brazil, Canada, China, Cuba, Czechoslovakia, France, India, Kenya, New Zealand, Poland, Serbia, UK and USSR (Rao 1970; Ellis 1971; Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Mercado Sierra et al. 1997; Wu 2004; Catania and Romero 2009; Savič 2020).
Description and illustration: Rao (1970), Ellis (1971), Nag Raj and Kendrick (1975), Holubová-Jechová (1984), Mercado Sierra et al. (1997), Wu (2004), and Catania and Romero (2009).
Notes: Chalara cylindrosperma was redescribed by Ellis (1971) and Nag Raj and Kendrick (1975). Its relationship with several other species such as C. longipes, C. nothofagi and C. stipitata was discussed by Nag Raj and Kendrick (1975). The conidiophores of this species could be very short, pale brown, and with a fewer (up to 7) septa to very long, dark brown, and with more (up to 12) septa. The short conidiophores are the dominant type from the Chinese specimens.
Twenty-one collections from nine different hosts (Aconitum napeffus, Eucalyptus spp., Fagus sylvatica, Ilex denticulata, Picea glauca, Picea mariana, Pinus sylvestris, Quercus sp., and Podocarpus milanjianus held in Herbarium IMI were examined for comparison. The fungus from the Chinese collections matched well with those collections, including the Type material IMI 44549 from Corda's herbarium (PR).
For comparison, the ITS sequence was generated from the strain CBS658.79 of C. cylindrosperma and was almost identical with the one generated from the Chinese collection Wu8177.
Chalara fusidioides (Corda) Rabenh., Deutschl. Krypt.-Fl. (Leipzig) 1: 38, 1844.
≡ Torula fusidioides Corda, Icon. Fung. (Prague) 2: 9, 1838.
Description on the natural substrate: Anamorph: Stroma absent. Setae absent. Conidiophores lageniform, rarely obclavate, brown, smooth, consisting of a 1–3-septate basal stack and a terminal phialide. Conidiogenous cells phialidic, lageniform, 11–16 µm long, subhyaline to pale brown, smooth; transition from venter to collarette abrupt; venter globose, 5–11 × 3.5–9 µm; collarette cylindrical, 5.5–16 × 1.5–4 µm. Conidia endogenous, extruded in short chains, cylindrical, 4.5–12 × 1.5–3.5 µm, hyaline, aseptate, with truncate or flattened ends (Nag Raj and Kendrick 1975). Teleomorph: Unknown.
Ecology/substrate/host: Saprobes on decaying rotten leaves, bark, fronds, needles, and fungal fruitbody.
Geographical distribution: Canada, Czechoslovakia, Germany, India, Italy, New Zealand, North America and UK (Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975) and Holubová-Jechová (1984).
Notes: Chalara fusidioides, the type species of the genus Chalara, was redescribed by Nag Raj and Kendrick (1975). It is characterized by 1(–3)-septate conidiophores, lageniform phialides with globose venters and cylindrical collarettes, abrupt transition from venters to collarettes, and hyaline, aseptate, cylindrical conidia (4.5–12 × 1.5–3.5 μm) with rounded ends and no frill (Nag Raj and Kendrick 1975).
Only one partial SSU sequence of C. fusidioides was available for molecular phylogenetic analyses, and its affinity to Leotiomycetes was confirmed. However, this sequence was not from the Type material (Gernandt et al. 2001). Johnston et al. (2019) reported that the strain from which this SSU sequence was generated might belong to same family as Chalara longipes. A megablast search of GenBank nucleotide database using this sequence showed it had highest homology with Xenochalara juniperi (99%), Neofabraea malicorticis (99%) and Hyaloscypha (99%) and Encoeliopsis rhododendri (99%).
No living strain of C. fusidioides seems to be available from any of those public fungal strain collections such as ATCC, CABI fungal strains, CBS, CMGCC, etc. The phylogenetic relationship of this species with other Chalara s. str. and Calycina species remains to be studied.
Chalara kirkii W.P. Wu and Y.Z. Diao, sp. nov., Fig. 30e, f, MycoBank MB845203.
Etymology: Named after the former IMI mycologist, Paul Kirk.
Typification: China, Ningxia Province, Jingyuan County, Liupan Mountain, Liantianxia, on wood of unidentified plant, 25 August 1997, Wenping Wu, Holotype HMAS 352243 (= Wu1042g).
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae with thin and smooth wall, 2–3 µm diam. Anamorph: Stroma absent. Setae absent. Conidiophores reduced to a small basal cell and a phialide. Conidiogenous cells directly arising from cells of superficial hyphae, scattered or aggregated at the bases, erect, straight, ampulliform or occasionally lageniform, 30–40 µm long, pale brown to brown, thin- and smooth-walled; transition from venter to collarette abrupt; venter subglobose, globose, 12–14 µm long and 7–8 µm wide; collarette cylindrical, 20–30 µm long, 2.8–3 µm wide, smooth, concolorous with venter; ratio of mean lengths of collarette and venter = 1.9:1. Conidia endogenous, extruded in long and loose chains, cylindrical, 13–15 × 2–2.5 µm, both ends rounded, hyaline, aseptate, thin- and smooth-walled; mean conidium length/width ratio = 6.2:1. Teleomorph: Unknown.
Material examined: China, Ningxia Province, Jingyuan County, Liupan Mountain, Liantianxia, on wood of unidentified plant, 25 August 1997, Wenping Wu, W1042g (holotype).
Ecology/substrate/host: Saprobe on rotten wood.
Geographical distribution: China.
Notes: Of the existing species under the genus Chalara s. lat., C. kirkii resembles C. dennisii and C. fusidioides in globose venter and aseptate conidia (Nag Raj and Kendrick 1975; Kirk 1986; McKenzie et al. 2002). Chalara kirkii is somewhat larger in all dimensions than other two species. The conidiogenous cells in both C. dennisii and C. fusidioides are never longer than 30 µm. The conidia of C. dennisii are smaller (4–6 × 0.8–1.0 µm) than those of C. kirkii.
Chalara longiphora W.P. Wu & Y.Z. Diao, sp. nov., Figs. 32, 33, MycoBank MB845205.

Chalara longiphora (Wu16044, holotype). a–f Conidiophores and conidiogenous cells. g–l Upper part of conidiophores bearing terminal phialides. m–o Conidia. Scale bar: 10 µm for a–f, 5 µm for g–o

Chalara longiphora (Wu16209). a–d, g unbranched conidiophores; e, f Branched conidiophores. h–l, s Upper part of conidiophores bearing terminal phialides. m–r Conidia. Scale bar: 20 µm for e–g, 10 µm for a–d, 5 µm for h–s
Etymology: Refers to its long conidiophores.
Typification: China, Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Holotype HMAS 352177 (= Wu16044), ex-type strain CGMC3.23418 (= NN76308).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores scattered, or a few aggregated at the base, erect, straight or slightly curved, simple or occasionally branched, long clavate, subcylindrical, (72–)90–155 µm high, 4–5 µm wide, basal cells slightly swollen and lobed, 8–13-septate, dark brown at the lower part, paler towards upper part, smooth-walled, terminating in a phialide. Conidiogenous cells integrated, terminal, lageniform, 31–35 μm long, medium brown, smooth; transition from venters to collarettes abrupt; venters lageniform, 16–17.5 μm long, 5–6 µm wide at the widest part which is the transition from venter to collarette; collarettes cylindrical, 14–17 × 2.5–2.7 µm, concolorous with venter; ratio of mean lengths of collarette and venter = 0.9:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 6–8 × 2–2.5 µm, straight, slightly rounded at the apex, truncate at the base, hyaline, aseptate; mean conidium length/width ratio = 3.1:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed at margin, aerial mycelium poorly developed, dark yellow brown, reverse dark brown, with yellow brown pigment diffused in agar, sterile, up to 6 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Zhejiang Province, Deqing, Moganshan, Luhuadang, on dead seed pods of unidentified Leguminosae, 16 October 2019, Wenping Wu, Wu17054; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of Quercus sp., 18 October 2018, Wenping Wu, Wu16192; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu (Wu16209). Living strains: 76309 (from 16044a), 76437 (from 16192), 76442 (from 16209), 77640, 77641 and 77642 (from Wu17054).
Ecology/substrate/host: Saprobe on dead seed and leaves of different plants.
Geographical distribution: China.
Notes: Chalara longiphora resembles C. cylindrosperma and C. platanicola in morphology of conidiophores, phialides and conidia, but differs by longer venter and shorter conidia (Ellis 1971; Nag Raj and Kendrick 1975). Morphological variations on conidiophores and conidia were observed among the examined specimens, for example the conidiophores in Wu16209 were frequently branched, and the conidiophores in Wu16044 were with fewer septa and significantly shorter than those in Wu16209. Conidia from the specimen Wu16192 were cylindrical and longer than those from other specimens. The branched conidiophores were also reported in C. cylindrosperma (Catania and Romero 2009).
Identical ITS sequences were generated from 7 different strains. Based on a megablast search of GenBank nucleotide database, the closest matched to the ex-type strain 76308 include C. clidemiae (NR_145313, 93% identity), C. cylindrosperma (MH873005, 97% identity), Chalara sp. (MK584995, 95% identity) and many unidentified fungi in Helotiaceae.
Chalara pengii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 34, MycoBank MB845207.

Chalara pengii (a–l from Wu16835, holotype; m–o, s–w from Wu16010; p–r from 16046). a–h, m–r Conidiophores and phialidic conidiogenous cells. i–l, s–w Conidia. Scale bar: 5 µm
Etymology: Named after my classmate Prof. Jianqiang Peng, who gave strong support during earlier time of this work.
Typification: China, Jiangsu Province, Wuxi, Wuxi Forestry Park, on dead leaves of Cyclobalanopsis sp., 25 Aug. 2019, Wenping Wu, Holotype HMAS 352179 (= Wu16835), ex-type strain CGMCC3.23461 (= NN77176).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, solitary or in small group of 2–4, obclavate, 32–36 µm long, 3.5–4.5 µm at the base, 1–2-septate, medium brown, slightly paler towards the apex, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, 22–27 µm long, medium brown, smooth; transition from venter to collarette gradual or abrupt; venters conic, subellipsoidal, (10–)13–16 µm long, 4.3–5.5 µm wide, pale brown; collarettes cylindrical, 13.5–21 × 3–3.5 µm, medium brown; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded in short chains cylindrical, 6.5–9 × 1.8–2.2 µm, base truncate with small frills, apex rounded or flattened, hyaline, aseptate; mean conidium length/width ratio = 3.9:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, white, then pale to medium brown, reverse yellow to dark brown, with paler margin, sterile, 5–13 mm on PDA at 25 °C in 4 weeks. The two strains (76196 and 76222) have different appearance and growth rate on PDA, but their ITS sequences are identical.
Materials examined: China, Jiangsu Province, Wuxi, Wuxi Forestry Park, on dead leaves of Cyclobalanopsis sp., 25 Aug. 2019, Wenping Wu, Wu16836; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of Platanus occidentalis, 18 October 2018, Wenping Wu, Wu16010, Wu16011, Wu16036 and Wu16037; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu (Wu16046). Living strains: CGMCC3.23415 (= 76196), 76197 (from Wu16010b), 76198 (from 16011a), 76199 (from 16011b), 76208 (from Wu16011a), 76208 (from Wu16036a), 76209 (from Wu16036a), 76210 (from Wu16037), CGMCC3.23416 (= 76222, from Wu16046), 76223 (from Wu16046a), 77140 (from Wu16836a), 77141 (from Wu16836b) and 77177 (from Wu16835b).
Ecology/substrate/host: Saprobe on dead leaves of Cyclobalanopsis sp., Platanus occidentalis.
Geographical distribution: China.
Notes: Chalara pengii is characterized by uniseptate and shorter conidiophores, obclavate conidiogenous cells with an ellipsoidal venter, and aseptate, small-sized conidia (6.5–9 × 1.8–2.2 μm). It is similar to several known Chalara or Calycina species such as Calycina affinis (conidiophores 33–81 μm long, phialides 28–54 μm long, venters 12–29 × 3.5–7 μm, collarettes 12–32 × 2–2.5 μm, conidia 5.5–19 × 1.5–2.5 μm) and Calycina fungorum (phialides 25–40 μm long, venters 12–19 × 5.5–7.5 μm, collarettes 11–21 × 2.5–4 μm, conidia 5.5–8 × 2–2.5 μm), but differs from them by shorter phialides. In addition, the conidia of C. affinis are much longer (Nag Raj and Kendrick 1975).
Identical ITS sequences were generated from all studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76196 included Porodiplodia vitis (NR_163376 97% identity), P. livistonae (NR_160355, 97% identity), C. clidemiae (NR_145313, 97% identity), and many unidentified fungi of Leotiomycetes. The ITS sequences from C. pengii are also with high identity (97%) with those from C. africana, and morphologically these two species are also similar, but can easily be distinguished by processing sterile setae, longer conidiogenous cells (30–40 μm long) and shorter conidia (5–6.5 × 1.5–2 μm) in the former species (Nag Raj and Kendrick 1975).
Chalara platanicola W.P. Wu & Y.Z. Diao, sp. nov., Fig. 35, MycoBank MB845208.

Chalara platanicola (Wu16038, holotype). a–e, h–m Conidiophores and conidia. g, n–r Conidia. Scale bar: 10 µm for a–h, 5 µm for n–r
Etymology: Refers to its host plant Platanus occidentalis.
Typification: China, Zhejiang Province, Huaian County, Qiandaohu, on dead leaf of Platanus occidentalis, 18 Oct 2018, Wenping Wu, Holotype HMAS 352244 (= Wu15038), ex-type strain CGMCC3.23417 (= NN76305).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent or present; if present, consisting of aggregated irregular cells. Setae absent. Conidiophores arising from basal stroma or superficial hyphae, scattered, or a few aggregated at the base, erect, straight or flexuous, simple, long clavate, subcylindrical, 70–110 µm high, 4–5 µm wide, basal cells slightly swollen and lobed, dark brown at the lower part, paler towards upper part, 8–11-septate, terminating in a phialide. Conidiogenous cells integrated, terminal, lageniform, 26–28 μm long, medium brown; transition from venter to collarette abrupt; venters lageniform, 11–12.5 × 5.7–6.5 µm, widest at the transition from venter to collarette; collarettes cylindrical, 17–18 × 2.8–3.3 µm, concolorous with venter; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, cylindrical, 9–10.5(–13) × 2.2–2.5 µm straight, slightly rounded at the apex, truncate at the base, hyaline, aseptate, smooth; mean conidium length/width = 3.9:1. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded at margin, aerial mycelium poorly developed, soil brown, reverse concolorous, sterile, up to 6 mm on PDA at 25 °C in 4 weeks.
Other materials examined; China, Hubei Province, Shennongjia, on dead leave of unidentified tree, September 2004, Wenping Wu, Wu8177. Living strains: 50590 (from Wu8177) and 76306 (from Wu16038a).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Chalara platanicola is similar to C. cylindrosperma in morphology of conidiophores, phialides and conidia, but differs by shorter conidiophores and phialides (Ellis 1971; Nag Raj and Kendrick 1975). Chalara cylindrosperma was described with broad range in size of conidiophores, phialides and conidia, an indication of a species complex. Chalara platanicola also resembles C. longiphora (conidiophores (72–)90–155 µm long; conidiogenous cells 31–35 μm long; venters 16–17.5 μm long, 5–6 µm wide; collarettes 14–17 × 2.5–2.7 µm; ratio of mean lengths of collarette and venter = 0.9:1. Conidia 6–8 × 2–2.5 µm, mean conidium length/width ratio = 3.1:1), but differs from it in small-sized conidiophores conidiogenous cells, venters and conidia.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76305 included C. clidemiae (NR_145313, 92% identity), C. cylindrosperma (MH873005, 97% identity), Chalara sp. (MK584995, 95% identity), and many unnamed fungi of Leotiomycetes. In addition, the ITS sequence from this species has 17 bp differences from those of C. longiphora.
Chalara qinlingensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 36, MycoBank MB845209.

Chalara qinlingensis (Wu16647, holotype). a Setae and conidiophores. b–h Conidiophores and phialidic conidiogenous cells. i, j Setae. k, l Conidia. Scale bar: 10 µm for a, 5 µm for b–l
Etymology: refers to the location Qinling mountain in Shaanxi Province, where the type specimen was connected.
Type specimen: China, Shaanxi Province, Zhouzhi, Qinling Mountain, on dead seed pod of Cercis chinensis, 2 Aug. 2019, Wenping Wu, Holotype HMAS352180 (= Wu16647), ex-type strain CGMCC3.23426 (= NN76950).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Stroma present, consisting of aggregated irregular cells. Setae aggregated with conidiophores, erect, straight or flexuous, simple, cylindrical, 75–100 µm long, slightly inflated at the base to a width of 4–5.5 µm, gradually tapering towards the acute apex to a width of 1.5–2 µm, aseptate, dark brown to blackish, smooth, thick-walled. Conidiophores arising from superficial hyphae, solitary or aggregated with setae, erect, straight or slightly flexuous, simple, lageniform, subcylindrical, 27–31 µm long, medium brown, 0–1-septate, smooth, thin-walled. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, subcylindrical or obclavate, 25–32 µm long, pale to medium brown, smooth; transition from venter to collarette gradual; venter subcylindrical to conic, 8–10 × 3.2–4 µm; collarette cylindrical, 15–17 × 2.2–2.7 µm, concolorous with venter; ratio of mean lengths of collarette and venters = 1.8:1. Conidia endogenous, extruded in short chains, cylindrical, 6–9 × 1.5–1.8 µm, both ends flattened or rounded, hyaline, aseptate; mean conidium length/width = 4.5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, yellow brown with gray brown in the middle, reverse yellow brown to brown, with paler margin, with yellow pigment diffused into agar, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Shaanxi Province, Zhouzhi, Qinling Mountain, on dead seed pod of Cercis chinensis, 2 Aug. 2019, Wenping Wu, Wu16574, Wu16670. Living strains: 76918 (from Wu16574c), 76929 (Wu16574b), CGMCC3.23426 (= 76950, from Wu16647) and 76951 (from 16647b).
Ecology/substrate/host: Saprobe on dead seed pod of Cercis chinensis.
Geographical distribution: China.
Notes: Among the known Chalara species with setae, Chalara qinlingensis is mostly similar to C. africana in producing aseptate setae, reduced conidiophores, lageniform conidiogenous cells, and cylindrical, aseptate conidia (Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975; Wu 2004). The conidiophores, conidiogenous cells and conidia in C. qinlingensis are slightly longer in average than those in C. africana (Conidiophores 30–45 µm long; Conidiogenous cells 30–40 µm long; venter 12–14 µm long, 4–6 µm wide; collarette 18–24 µm long, 2.5–3 µm wide; ratio of mean lengths of collarette/venter = 1.6:1; Conidia 5–6.5 × 1.5–2 µm). In addition, the ITS sequence from this species has 31 bp differences from those in C. africana.
Identical ITS sequences were generated from all five studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76946 included C. clidemiae (NR_145313, 94% identity), Porodiplodia vitis (NR_163376 94% identity), Porodiplodia livistonae (NR_160355, 94% identity), and many unnamed fungi of Leotiomycetes. The ITS sequences from Chalara qinlingensis were also with high identity (95.7%) with those from C. pengii; morphologically these two species were also similar, but could easily be distinguished by absence of sterile setae, shorter conidiogenous cells (22–27 μm long) and venter (8–10 μm long), and wider conidia (6.5–9 × 1.8–2.2 μm) in the latter species (Nag Raj and Kendrick 1975).
Chalara sessilis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 134, 1975. Figure 37a, b.

Chalara sessilis (a, b from Wu1034), C. sinensis (c, d from Wu1037, Holotype), C. versicolor (e, f from Wu1015c, holotype) and Chalara sp. (g, h from Wu1542a). a, c, e, g Conidia. b, d, f, h Conidiophores and conidiogenous cells. Scale bar: a–d 10 μm, e–h 20 μm
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores reduced, consisting of a to single cell cooperated into hyphae and a phialide, pale to medium brown, smooth, the basal cell up to 5 µm long and 5 µm wide. Conidiogenous cells integrated, determinate, monoblastic, erect, straight or slightly curved, pale to medium brown, smooth, lageniform, 15–21 µm long, consisting of a venter and a collarette, transition gradually from venter to collarette; venter conical, to subcylindrical, 9–11 µm long and 3.5–4.5 µm wide; collarette cylindrical, concolorous with venter, smooth, about the same length with venter and 2–2.5 µm wide; collarette/venter = 1.1/1. Conidia endogenous, extruded in short chains, cylindrical, 3.5–6 × 1–1.5 µm, ends truncate, hyaline, aseptate; mean conidium length/width ratio = 3.8:1. Teleomorph: Unknown.
Material examined: China, Guangxi Province, Shangsi, Shiwandashan, on dead branches of unidentified tree, 2 Jan. 1998, Wenping Wu (Wu1268e); Yunnan Province, Kunming, Jindian Park, on dead needle of Picea sp., 24 November 1995, Wenping Wu, Wu1037. Living strain: CBS405.81 (50502), isolated from decaying needle of Pinus sylvestris, besides Pezizella subtilis, from Germany.
Ecology/substrate/host: Saprobe on dead needle of Knightia excelsa, Picea sp.
Geographical distribution: China, New Zealand.
Description and illustration: Nag Raj and Kendrick (1975).
Note: Chalara sessilis was described with sessile and shorter (18–23 µm long) phialides, and smaller conidia (3.5–6 × 1.5 µm) (Nag Raj and Kendrick 1975). The fungus from these Chinese collections is very similar to the original description and illustration on most aspects but with one difference, i.e., each conidiogenous cell is supported by a single basal cell (Nag Raj and Kendrick 1975).
Chalara sinensis W.P. Wu, Mycosystema 32: 320, 2004. Figure 37c, d.
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, branched hyphae with thin and smooth walls, 2–3.5 µm wide. Anamorph: Conidiophores solitary, or rarely aggregated, erect, straight, obclavate, consisting of one basal cell and a phialide, constricted at the septa; basal cell brown, thick- and smooth-walled, swollen and lobed at the base, 5–8 × 8–12.5 µm, arising from hyphae. Conidiogenous cells integrated, terminal, lageniform, obclavate, 40–56 µm long, pale brown to brown, smooth-walled, straight or slightly curved; transition from venter to collarette abrupt; venter subcylindrical or subellipsoidal, 20–24.5 µm long and 6.5–9.5 µm wide; collarette cylindrical, 17.5–30 µm long and 3–3.5 µm wide; ratio of mean lengths of collarette and venter = 1.1:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 10–14 × 2–2.5 µm, both ends truncate except for the apical conidium which is obtuse at the apex, hyaline, aseptate, smooth- and thin-walled; mean conidium length/width = 5.3:1. Teleomorph: Unknown.
Material examined: China, Xi'an Botanical Garden, Xi'an, Shaanxi Province, on dead leaves of Fagus sp., 10 October 1992, Wu Wenping, Holotype Wu673.
Ecology/substrate/host: Saprobe on dead leaves of Fagus sp.
Geographical distribution: China (Wu 2004).
Description and illustration: Wu (2004).
Notes: Chalara sinensis resembles Calycina affinis, Calycina brevipes, Calycina fungorum and Chalara sessili, but differs in different combination of shape and size of conidiophores, conidiogenous cells and conidia (Wu 2004). Nag Raj and Kendrick (1975) reported a wide range of conidial size (5.5–19 × 1.5–2.5 µm) in C. affinis and the differences between this species and C. sinensis were obvious in several aspects. Many collections of C. affinis in Herbarium IMI were examined in this study and they were aligned with the description and illustration provided by Nag Raj (1975). Except for the differences mentioned above, the conidia in all examined collections of C. affinis were obviously narrower (1.2–2 µm), with both ends rounded while in C. sinensis the conidia were much wider (2–2.8 µm) and with truncate ends. Calycina brevipes produced collarette with darker base and conidia with rounded apex and basal marginal frills (Nag Raj and Kendrick 1975). Calycina fungorum and C. sessilis have shorter conidia (less than 8 µm long) and also shorter conidiogenous cells (Nag Raj and Kendrick 1975).
One of the characteristics for this species was the basal cells with lobed margins, which was also reported from several other species of Chalara, such as C. brunnipes, C. alabamensis, C. distans and C. graminicola (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; McKenzie 1982, 1993).
Chalara sporendocladioides W.P. Wu & Y.Z. Diao, sp. nov., Fig. 38, MycoBank MB845212.

Chalara sporendocladioides (Wu5533a, holotype). a–f Conidiophores and conidiogenous cells. g, h Conidia. Scale bar: 20 μm for a; 10 μm for b, c; 5 µm for d–h
Etymology: Refers to its similarity with Sporendocladia in apically branched conidiophores.
Typification: China, Hainan Province, on dead branches of unidentified plant, 20 December 2002, Wenping Wu and Yan Huang, Holotype HMAS 352245 (= Wu5533a).
Description on the natural substrate: Colonies effuse, sparse, brown. Mycelium partly superficial, partly immersed in the substratum, composed of branched, septate, smooth, pale brown to brown hyphae 2–4 μm wide. Anamorph: Stroma absent. Setae absent. Conidiophores macronematous, mononematous, solitary or 1–3 aggregated at the base, erect, straight or flexuous, branched, 80–200 × 6–9 μm, consisting of a cylindrical and septate basal stalk and several branches bearing terminal conidiogenous cells; basal stalk cylindrical, 5–7-septate, smooth, dark brown at the base, medium brown at the upper part; Fertile region consisting of branches with terminal phialides in loose arrangement with a mono- to bi-verticillate branching pattern; primary branches cylindrical, 10–12 × 5–7 μm, brown, smooth, thick-walled. Conidiogenous cells formed at the apices of branches, ampulliform, lageniform, 37–45 μm long, pale brown to medium brown, smooth; transition from venter to collarette abrupt; venters conic, subellipsoidal, 13–17 × 8–10 μm; collarettes cylindrical, 20–30 × 3 μm; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, extruded in long chains, cylindrical, 10–12 × 2.5 μm, straight, both end flattened or truncated, hyaline, smooth, aseptate, guttulate; mean conidium length/width = 4.4:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead branches of unidentified tree.
Geographical distribution: China.
Notes: Chalara sporendocladioides differs from other known species of Chalara s. lat. in producing apically branched conidiophores, terminal phialides with clear differentiation of venter and collarette, and hyaline, aseptate and cylindrical conidia in long chains (Nag Raj and Kendrick 1975; McKenzie et al. 2002). The apically branched conidiophores with several phialides were also found in Sporendocladia, Lareunionomyces and Neolauriomyces, but in these genera the apical branches were penicillate with numerous phialides covering out layer of sporulating head and the conidia were produced in wet spore mass, thus they could easily be distinguished from Chalara (Kendrick 1961; Nag Raj and Kendrick 1975; Sutton 1975, 1993; Onofri and Zucconi 1984; Kirk 1985; Wingfield et al. 1987; Mouton and Wingfield 1993; Crous et al. 2016a, b, 2018).
Chalara versicolor W.P. Wu & Y.Z. Diao, sp. nov., Fig. 37e, f, MycoBank MB845213.
Etymology: Refers to its versicolorous conidiogenous cells with darker basal part of collarettes.
Typification: China, Ningxia Province, Jingyuan County, Liupan Mountain, Erlonghe, on wood of unidentified plant, 23 August 1997, Wenping Wu, Holotype HMAS 352246 (= Wu1015c).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores arising from superficial mycelium, scattered or 2–3 aggregated at the base, erect, straight or flexuous, simple, subcylindrical, 15–50 µm long, 5–7 µm wide at the base, basal cells slightly swollen, dark brown at the base, becoming medium brown towards the apex, 2–5-septate, smooth, with 1–2 percurrent proliferations, terminating in a phialide. Conidiogenous cells integrated, terminal, obclavate, lageniform, 47–50 µm longer, medium brown, become pale brown towards the apex; transition from venter to collarette abrupt; venter cylindrical, subcylindrical to subellipsoidal, 28–40 × 7–8 µm; collarette cylindrical, 27–30 × 3.5–4.5 µm, brown, significantly darker at the lower part; ratio of mean lengths of collarette and venter = 0.8:1. Conidia endogenous, extruded in short chains, cylindrical, 6.5–15 × 2.5–3 µm, straight, truncated at both ends or slightly rounded at the apex, without basal frill, smooth, hyaline, aseptate, mean conidium length/width ratio = 3.9:1. Teleomorph: Unknown.
Other material examined: China Ningxia Province, Jingyuan County, Liupan Mountain, Erlonghe, on dead branches of Quercus sp., 23 August 1997, Wenping Wu, Wu1013.
Ecology/substrate/host: Saprobe on dead branches and wood of Quercus and other plant.
Geographical distribution: China.
Notes: Chalara versicolor is characterized by 2–5-septate conidiophores, obclavate to lageniform conidiogenous cells with cylindrical, subcylindrical to subellipsoidal venter, cylindrical and versicolorous collarettes with darker base, and aseptate conidia with both ends truncated and bearing no frill. It resembles C. crassipes, C. cylindrosperma, C. ginkgonis, C. longipes and C. parvispora. However, in all these species, the venter and collarette were concolorous (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In addition, C. crassipes, C. longipes and C. parvispora have narrower conidia (< 1.5 µm) than C. versicolor.
Chalara sp.1, Fig. 37g, h.
Description on the natural substrate: Colonies effuse, brown, hairy, sparse. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or medium thick-walled, 2–3.5 µm wide. Stroma absent. Anamorph: Conidiophores macronematous, mononematous, single or a few aggregated at the base, erect, simple, cylindrical, basal cells slightly swollen, 70–110 µm high, brown to dark brown, 2–5-septate, with 0–2 percurrent proliferation. Conidiogenous cells integrated, terminal, cylindrical, brown, becoming pale brown towards the apex, smooth, 30–35 × 5–6 µm. Conidia endogenous, hyaline, aseptate, cylindrical, straight, apex rounded, base truncate and with a basal frill, smooth, 6–7.5 × 3.5–4 µm. Teleomorph: Unknown.
Material examined: China, Guangxi Province, Shiwandashan, on dead culm of bamboo, 30 December 1997, Wenping Wu, Wu1542a.
Ecology/substrate/host: Saprobe on dead culm of bamboo.
Geographical distribution: China.
Notes: Chalara sp.1 is similar to C. quercina and C. ungeri on multiseptate conidiophores and cylindrical conidiogenous cells with transition from venter to collarette almost imperceptible (Nag Raj and Kendrick 1975). Chalara sp.1 differs from C. quercina by its dark brown conidiophores (vs. subhyaline in C. quercina) and wider conidia; from C. ungeri (conidia 5.5–11 × 3.5–4.5 μm) by smaller conidia and cylindrical conidiophores with percurrent proliferations. Morphologically it represents a new taxon of Chalara s. lat. However, no living strain was obtained from the fresh specimen and its phylogenetic relationship with Chalara s. lat. couldn’t be determined.
Other known Chalara s. lat. species with aseptate conidia.
Except for the above accepted species, the following known species fit well to the emended generic concept of Chalara s. str. and should be maintained to the genus (Nag Raj and Kendrick 1975; Gadgil and Dick 1999; McKenzie et al. 2002). However, no living strain or DNA sequence was available for molecular phylogenetic analysis, thus their phylogenetic relationship within Leotiomycetes remains to be studied.
In addition, 16 other published Chalara species fits well to the emended generic concept of Chalara and should.
Chalara austriaca (Fautrey & Lambotte) Nag Raj & W.B. Kendr., Monogr Chalara Genera (Waterloo): 89, 1975.
≡ Chalara longipes f. austriaca Fautrey & Lambotte, Revue Mycol., Toulouse 17: 69, 1895.
Ecology/substrate/host: Saprobe on decaying needle of Pinus nigra subsp. austriaca.
Geographical distribution: France (Koukol 2011).
Description and illustration: Fautrey and Lambotte (1895) and Koukol (2011).
Notes: Chalara austriaca was recognized as a distinct species by Nag Raj and Kendrick (1975). Their description was, however, for a different fungus (conidiophores 12–27 × 3–4 μm; conidia 4–6.5 × 1–1.5 μm) and corrected by Koukol (2011). According to Koukol (2011), the conidiophores in C. austriaca were brown, erect, widened at the base in the shape of a flask, one-septate, 35–40 × 5–6 μm; the conidia were unicellular, cylindrical, with both ends clearly truncate, hyaline, 10–12 × 2 μm. As the holotype practically did not contain the fungus, the original illustration was designated as the lectotype of the species by Koukol (2011). An epitype could be assigned only after the species is re-discovered in the indicated locality after a targeted study. The phylogenetic relationship of this fungus with other chalara-like fungus remains to be resolved.
Chalara bohemica Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 93, 1975.
Ecology/substrate/host: Saprobe on decaying branches, stems and trunk of Rubus, Fagus and Ulmus.
Geographical distribution: Czechoslovakia and France (Nag Raj and Kendrick 1975; Holubová-Jechová 1984).
Description and illustration: Nag Raj and Kendrick (1975) and Holubová-Jechová (1984).
Notes: Chalara bohemica differs from other species by verrucose conidiophores and hyaline, aseptate and cylindrical conidia (6–13 × 2–3 µm) bearing no basal frill.
Chalara brachyspora Sacc., Michelia 1: 81, 1877.
Ecology/substrate/host: Saprobe on decaying wood including Corylus.
Geographical distribution: Italy (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara brevicaulis Arambarri & Gamundi, in Arambarri, Gamundí & Bucsinszky, Darwiniana 23(2–4): 330, 1981.
Ecology/substrate/host: Saprobe on Nothofagus dombayi.
Geographical distribution: Argentina (Arambarri et al. 1981).
Description and illustration: Arambarri et al. (1981).
Notes: Chalara brevicaulus differs from other species by reduced conidiophores composed of one basal cell and an obclavate phialides (25–47 µm long), cylindrical to ellipsoidal venter (18–24 × 7–9.4 µm), cylindrical and wider collarettes (14–24 × 4.7–6.5 µm), and wider conidia (9.4–12.6 × 4.8–4.7 µm) bearing no basal frill.
Chalara bulbosa (B. Sutton & Piroz.) P.M. Kirk, in Kirk & Spooner, Kew Bull. 38(4): 580, 1984.
≡ Chaetochalara bulbosa B. Sutton and Piroz., Trans. Br. mycol. Soc. 48(1): 351, 1965.
Ecology/substrate/host: Saprobe on decaying leaves of Ilex aquifolium.
Geographical distribution: UK (Sutton and Pirozynski 1965).
Description and illustration: Sutton and Pirozynski (1965).
Notes: Chalara bulbosa differs from other species by presence of aseptate and sterile setae, sessile conidiogenous cells with a bulbous venter and a cylindrical collarette, and hyaline, aseptate and cylindrical conidia (99–14 × 4–4.5 µm) without basal frill.
Chalara cylindrica P. Karst., Meddn Soc. Fauna Flora fenn. 14: 108, 1887.
Ecology/substrate/host: Saprobe on Abies, Eucalyptus and Picea.
Geographical distribution: Australia, Czechoslovakia, France, Germany, North America and UK (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara cylindrica resembles Chalara bohemica in producing rough-walled conidiophores, but differs from it by the smaller conidia (3–9.5 × 1–1.5 µm vs. 6–13 × 2–3 µm).
Chalara dennisii P.M. Kirk, Trans. Br. mycol. Soc. 86(3): 411, 1986.
Ecology/substrate/host: Saprobe on rotten cone of Pinus sp.
Geographical distribution: UK (Kirk 1986).
Description and illustration: Kirk (1986).
Notes: Chalara dennisii (venter 3.5–5 µm in diam, collarette 5.5–8 × 1.4–1.6 µm, conidia 4–6 × 0.8–1.2 µm) is similar to C. fusidioides (venter 5–11 × 3.5–9 µm, collarette 5.5–16 × 1.5–4 µm, conidia 4.5–12 × 1.5–3.5 µm) in producing sessile conidiogenous cells with a globose venter and a cylindrical collarette, and hyaline, aseptate and cylindrical conidia without basal frill, but differs in somewhat smaller in all dimensions such as phialides and conidia (Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Kirk 1986).
Chalara graminicola McKenzie, N.Z. Jl Bot. 20(3): 245, 1982.
Ecology/substrate/host: Saprobe on dead leaves of Chionochloa sp.
Geographical distribution: New Zealand (McKenzie 1982).
Description and illustration: McKenzie (1982).
Notes: Chalara graminicola (venter 14–17 × 15–21 µm, collarette 65–115 × 5–6.5 µm, conidia 14–17 × 3.5–4.5 µm) is similar to C. cylindrophora (venter 5–8 × 5–6.5 µm, collarette 32–50 × 2.5–3 µm, conidia 7.5–10 × 1–1.5 µm) in producing sessile conidiogenous cells with a very short venter and a longer cylindrical collarette, and hyaline, aseptate and cylindrical conidia without basal frill, but differs in longer phialides and conidia (McKenzie 1982; Wu 2004).
Chalara indica J. Pratibha, K.D. Hyde & Bhat, Cryptog. Mycol. 26(2): 97, 2005.
Ecology/substrate/host: Saprobe on decaying leaves of Areca catechu.
Geographical distribution: India (Pratibha et al. 2005).
Description and illustration: Pratibha et al. (2005).
Notes: Chalara indica is characterized by the clustered conidiophores, percurrently proliferated conidiogenous cells, ellipsoidal venter (32.5–40 × 11–16.5 µm), shorter collarettes (25–32.5 × 8–10 µm), and hyaline, aseptate and cylindrical conidia (20–30 × 5–6.5 µm) with rounded apex and truncate base without basal frill. It differs from any other known species in wider venters and collarettes, and large-sized conidia (Nag Raj and Kendrick 1975; McKenzie 2002).
Chalara kobensis McKenzie, in McKenzie, Pinnoi, Wong, Hyde & Jones, Fungal Diversity. 11: 133, 2002.
Ecology/substrate/host: Saprobe on decaying leaves of Pasaia edulis.
Geographical distribution: Japan (Matsushima 1975; McKenzie et al. 2002).
Description and illustration: Matsushima (1975) and McKenzie et al. (2002).
Notes: Chalara kobensis is characterized by the sessile conidiogenous cells, subcylindrical venter (25–40 × 5 µm), cylindrical collarettes (30–45 × 3 µm), and hyaline, aseptate and cylindrical conidia (10–15 × 2.2–2.5 µm) with both ends rounded and no basal frill.
Chalara lichenicola M.S. Christ., Nordic Jl Bot. 13(3): 310, 1993.
Ecology/substrate/host: Saprobe on the basal parts of the podetial of Cladonia gracilis and the immature basidiomata of Tremella sp.
Geographical distribution: Svalbard (Christiansen 1993).
Description and illustration: Christiansen (1993).
Notes: Chalara lichenicola was described as a lichenicolous hyphomycete from Svalbard and differs from other species by small-sized phialides and conidia (2.5–4 × 0.5–0.8 µm) bearing no basal frill.
Chalara lobariae Etayo, in Etayo & Diederich, Bull. Soc. Nat. luxemb. 97: 97, 1996.
Ecology/substrate/host: Saprobe on old or necrosed thallus parts of Lobaria pulmonaria.
Geographical distribution: France (Etayo and Diederich 1996).
Description and illustration: Etayo and Diederich (1996).
Notes: Chalara lobariae is characterized by sessile, obclavate conidiogenous cells (35–50 µm long, venter 6–9 µm wide, collarette 3.5–4 µm wide) and hyaline, aseptate and cylindrical conidia (6–10 × 2.5–3.5 µm) bearing no basal frill.
Chalara matsushimae McKenzie, in McKenzie, Pinnoi, Wong, Hyde & Jones, Fungal Diversity 11: 134, 2002.
Ecology/substrate/host: Saprobe and isolated from soil.
Geographical distribution: Japan (Matsushima 1975; McKenzie et al. 202).
Description and illustration: Matsushima (1975) and McKenzie et al. (2002).
Notes: Chalara matsushimae is characterized by short conidiophores (50–80 × 3–3.5 µm) composed of one basal cell and a phialide (37–45 µm long), cylindrical or obconic venter (20 × 2 µm), cylindrical collarette (18–32 × 2–2.4 µm), and hyaline, aseptate and short-cylindrical conidia (2.5–4.5 × 1.5–2 µm) bearing no basal frill. The conidial size is similar to those in C. crassipes and C. parvispora, but differs in shape and size of conidiophores and phialides (Nag Raj and Kendrick 1975; McKenzie et al. 2002).
Chalara myrsines Gadgil & M.A. Dick, N.Z. Jl For. Sci. 29(3): 443, 1999.
Ecology/substrate/host: Saprobe on Myrsines chathamicae.
Geographical distribution: New Zealand (Gadgil and Dick 1999).
Description and illustration: Gadgil and Dick (1999).
Notes: Chalara myrsines differs from other species by 3–5-septate conidiophores (30–70 × 8–10 µm) with a mail stipe and a terminal phialide (65–85 µm long), obclavate or subglobose venter (5–15 × 6–10 µm), and longer and cylindrical collarettes (50–70 × 3–4 µm), and hyaline, aseptate and cylindrical conidia (7–15 × 3–4 µm) without basal frill.
Chalara neglecta Hol.-Jech., Folia Geobot. Phytotax. 19(4): 411, 1984.
Ecology/substrate/host: Saprobe on buds of deciduous trees.
Geographical distribution: Czechoslovakia (Holubová-Jechová 1984).
Description and illustration: Holubová-Jechová (1984).
Notes: Chalara neglecta is characterized by sessile and concolorous conidiogenous cells (35–41 µm long), sharp transition from venter to collarette, ellipsoidal to subcylindrical venter (16–20 × 4.5–6 µm), cylindrical collarette (17–22 × 1.5–2 µm), and hyaline, aseptate and cylindrical conidia (3.5–6 × 1.5–1.8 µm) without basal frill. It is similar to C. affinis and C. brevipes, but differs in size of phialides and conidia (Nag Raj and Kendrick 1975; Holubová-Jechová 1984).
Chalara nothofagi Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera: 126, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri var. cliffortioides.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara nothofagi is characterized by multiseptate and concolorous conidiophores (115–168 × 5–6 µm) with a mail stipe and a terminal phialide (48–58 µm long), subcylindrical venter (20–27 × 6–8.5 µm), cylindrical collarettes (26–33 × 3.5–4 µm), and hyaline, aseptate and cylindrical conidia (13–17 × 2.5–3 µm) without basal frill. It is similar to C. cylindrosperma, but with longer venter and broader conidia.
Mollisina Höhn. ex Weese, in Weese, Mitt. bot. Inst. tech. Hochsch. Wien 3(2): 67, 1926.
= Dendrotrichoscypha Svrček, Ceská Mykol. 31: 9, 1977.
Type species: Mollisina rubi (Rehm) Höhn., in Weese, Mitt. bot. Inst. tech. Hochsch. Wien 3(2): 67, 1926.
Ecology/substrate/host: Saprobe on decaying plant material.
Geographical distribution: Widely distributed.
Description and illustration: Höhnel (1919).
Notes: A total of 15 legitimate names were listed under the genus Mollisina (MycoBank, accessed on July 7th, 2022), but only one species, M. uncinata, was reported with chalara-like anamorph. Its affinity to Pezizellaceae was confirmed by the phylogenetic analyses (Ekanayaka et al. 2019; Johnston et al. 2019).
Mollisina uncinata Arendh. & R. Sharma, Mycotaxon 20(2): 657, 1984.
Anamorph: Conidiophores gregarious, upright, straight, mostly 200–450 × 5–6.5 μm, multi-septate, brown; in culture, straight or curved to undulating, multiseptate, smooth, not constricted at the septa, dark brown, terminating in a phialide. Conidiogenous cells phialidic lageniform, 24–35 × 2–4 μm, with globose to subcylindrical venter and cylindrical collarette. Conidia borne in chains, bacilliform to short clavate, 3–4.5 × 1.5–2 μm, truncate at the base, truncate to slightly rounded at the apex, aseptate, hyaline. Teleomorph: see Hosoya and Otani (1997).
Ecology/substrate/host: Saprobe on dead leaves of Quercus sp.
Geographical distribution: India and Japan (Arendholz and Sharma 1984; Hosoya and Otani 1997).
Description and illustration: Arendholz and Sharma (1984) and Hosoya and Otani (1997).
Notes: Arendholz and Sharma (1984) reported association of the chalara-like fungus with M. uncinata. Hosoya and Otani (1997) confirmed the connection of its Chalara anamorph with M. uncinata teleomorph with a single-ascospore isolate. The LSU and ITS sequences of this species were available from three different strains in the GenBank (Hosoya and Otani 1997).
Parachalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845216.
Etymology: Refers to its morphological similarity with Chalara fungorum in reduced conidiophores and aseptate conidia.
Diagnosis: Similar to Chalara in phialides with cylindrical collarettes and aseptate, cylindrical conidia, but phylogenetically distinct. Also similar to Nagrajchalara, but differs in producing aseptate conidia.
Type species: Parachalara olekirkii W.P. Wu & Y.Z. Diao.
Description in pure culture on PDA: Colonies on PDA effuse, rounded or lobbed at margin, aerial mycelium poorly developed, brown, reverse dark brown to blackish. Mycelium partly immersed and partly superficial, composed of subhyaline, pale brown, septate and branched hyphae with smooth and thin wall. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, erect, straight or slightly flexuous, obclavate, subcylindrical, 1-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, obclavate, lageniform, medium brown to brown, smooth; transition from venter to collarette abrupt; venter cylindrical, subellipsoidal, medium brown; collarette cylindrical, brown, versicolorous, basal part darker than venter and other part of collarettes. Conidia endogenous, extruded in short chains, cylindrical, base truncate, apex rounded or flattened, hyaline, aseptate, guttulate. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: In the phylogenetic analyses with different datasets of combined sequences of SSU/LSU and LSU/ITS, the strain CGMCC3.23361 formed a distinct lineage in the phylogenetic trees (Figs. 1, 2, 3, 8, 9). Only in the tree generated from the dataset with only ITS sequences of Nagrajchalara, the species was nested among other Nagrajchalara species, but as a distinct lineage. Although also with aseptate conidia, it was phylogenetically closer to Nagrajchalara than Chalara s. str.. Based on both morphology and phylogeny, new genus Parachalara is established to accommodate this fungus.
Parachalara is morphologically similar with Chalara, Nagrajchalara, Neochalara and Stipitochalara, but phylogenetically distinct. The septate conidia in Nagrajchalara and Neochalara clearly distinguish them from Parachalara (the aseptate conidia). Stipitochalara also produces aseptate conidia, but in this genus the conidiophores are well-developed and composed of a basal stalk and a terminal phialide, while in Parachalara the conidiophores are reduced to a basal cell and a terminal phialide (Nag Raj and Kendrick 1975). Morphologically Parachalara is indistinguishable from some existing species of Chalara s. str. with the reduced conidiophores and aseptate conidia, they can only be distinguished by the phylogenetic analysis.
Parachalara olekirkii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 39, MycoBank MB845217.

Parachalara olekirkii (ex-type strain 43656 from Wu1559d, holotype). a–j Conidiophores and phialidic conidiogenous cells. k–p Conidia. q Superficial mycelium and conidiophores. Scale bar: 10 µm for a–d; 5 µm for e–q
Etymology: Named after Mr. Ole Kirk, the former Vice President in Novozymes, who gave his strong support for this work during last many years.
Typification: China, Guangxi Province, Shiwandashan, On dead leaves of ?Cinnamomum sp., 30 December 1997, Wenping Wu, Holotype HMAS 352182 (= Wu1559d), ex-type strain CGMCC3.23361 (= NN43656).
Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, erect, straight or slightly flexuous, obclavate, subcylindrical, 50–63 µm long, 4–5 µm wide at the base, 1-septate, occasionally 2–4-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, obclavate, lageniform, 50–60 µm long, medium brown to brown, smooth; transition from venter to collarette abrupt; venter cylindrical, subellipsoidal, 26–31 µm long, 5.5–6.5 µm wide, medium brown; collarette cylindrical, 24–28 × 2.6–2.9 µm, brown, versicolorous, basal part darker than venter and other part of collarettes; ratio of mean lengths of collarette and venter = 0.9:1. Conidia endogenous, extruded in short chains, 14.8–17.5(–19) × 2 µm, cylindrical, base truncate, apex rounded or flattened, hyaline, aseptate, guttulate; mean conidium length/width = 8:1. Teleomorph: Unknown.
Culture characteristics: Colonies on PDA effuse, rounded or lobbed at margin, aerial mycelium poorly developed, brown, reverse dark brown to blackish, sterile, up to 10 mm at 25 °C in 4 weeks. Mycelium partly immersed and partly superficial, composed of subhyaline, pale brown, septate and branched hyphae with smooth and thin wall, 1.5–3 µm wide.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Parachalara olekirkii is morphologically similar with a few existing Chalara s. str. species with reduced conidiophores consisting of one basal stalk cell and a terminal phialide, and aseptate conidia, but differs in a unique combination of shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975).
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 43656 included Phialina ulmariae (MZ159550, 90% identity), Calycellina fagina (OL752703, 90% identity) and many unnamed fungi of Leotiomycetes. Based on LSU blast in GenBank, the closest matches to the ex-type strain 43656 include Calycellina fagina (OL744073, 96% identity), Chalara sp. strain OC0015 (= 43656) (FJ176240, 100% identity), Phialea strobilina (EF596821, 95% identity), Sphaeridium vitellinum (MH872784, 95.5% identity) and many other fungi of Leotiomycetes.
Zymochalara Guatim., R.W. Barreto & Crous, in Guatimosim, Schwartsburd, Barreto & Crous, Mycol. Progrs. 15(12): 1261, 2016.
Anamorph: Conidiophores reduced to phialides. Conidiogenous cells phialidic, sessile, scattered, solitary, unbranched, lageniform, subulate or subcylindrical, aseptate, brown to cinnamon-brown, paler towards the apex, smooth; venter subcylindrical or ellipsoid, pedicellate or not; collarette cylindrical, transition from venter to collarette gradual. Conidia endogenous, basipetal, extruded singly or in somewhat long and easily fragmenting chains, cylindrical, truncate at both ends, aseptate, hyaline, biguttulate, smooth. Yeast-like in culture. Teleomorph: Unknown.
Type species: Zymochalara cyatheae Guatim., R.W. Barreto & Crous.
Ecology/substrate/host: Pathogenic on fronds of fern.
Geographical distribution: Brazil.
Description and illustration: Guatimosim et al. (2016).
Notes: Zymochalara was established for two chalara-like fungi, Zymochalara cyatheae and Z. lygodii, both as frond pathogens of fern in Brazil. Morphologically it was hardly to be distinguished from Chalara s. lat., but phylogenetically was distinct and formed a sister clade with Bloxamia cyatheicola in the phylogenetic tree (Guatimosim et al. 2016; Figs. 2, 3, 8). Chalara s. lat. was known to be polyphyletic and several genera with chalara-like morphology were created (Coetsee et al. 2000; Paulin et al. 2002; Cai et al. 2009; Crous et al. 2021). The phylogenetic analyses with inclusion of many species of Chalara-related fungi in this study supported the separation of the genus Zymochalara from other chalara-like genera. Two species were known for the genus.
Two existing fungi under the genus Bioscypha, B. cyatheae and B. pteridicola, also produced chalara-like anamorphs with aseptate and rectangular conidia in small size (3.5 × 1.5 μm in B. cyatheae; (3.5–)7–8.7 × 1.7–2.5 μm in B. pteridicola; Samuels and Rogerson 1990). These two fungi could be congeneric with Zymochalara, but this needs to be confirmed in future study.
Zymochalara cyatheae Guatim, R.W. Barreto & Crous, in Guatimosim, Schwartsburd, Barreto & Crous, Mycol. Progr. 15(12): 1261, 2016.
Ecology/substrate/host: Pathogenic on frond of Cyathea delgadii (fern).
Geographical distribution: Brazil (Guatimosim et al. 2016).
Description and illustration: Guatimosim et al. (2016).
Zymochalara lygodii Guatim., R.W. Barreto & Crous, in Guatimosim, Schwartsburd, Barreto & Crous, Mycol. Progr. 15(12): 1263, 2016.
Ecology/substrate/host: Pathogenic on frond of Lygodium volubile (fern).
Geographical distribution: Brazil.
Geographical distribution: Brazil (Guatimosim et al. 2016).
Description and illustration: Guatimosim et al. (2016).
Notes: The two accepted species under the genus Zymochalara are morphologically very similar, but distinct in ITS and LSU sequences (15 bp of variable sites for the ITS locus; and 10 bp of variable sites for LSU). Guatimosim et al. (2016) also distinguished them by different host species and size of phialides (29–38 μm in Z. cyatheae and 32–50 μm in Z. lygodii).
Genera with septate conidia
Nagrajchalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845218.
Etymology: Named after the mycologist T.R. Nag Raj, who made the monograph of chalara-like fungi as a foundation for this study.
Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Stroma absent or present; if present, consisted of brown and irregular cells. Setae absent or present; solitary or aggregated, often in clusters with conidiophores, erect, straight or flexuous, cylindrical, tapering towards the apex, brown to dark brown, aseptate or septate, thick- and smooth-walled. Conidiophores scattered or aggregated at the basal stroma, sometimes associated with setae at the base, erect, straight or flexuous, simple, obclavate, subcylindrical, pale brown to very dark brown, aseptate or septate, smooth or verruculose, with or without percurrent proliferations, terminating in a phialidic conidiogenous cell. Conidiogenous cells integrated or discrete, terminal or directly arising from cells of superficial hyphae, ampulliform, lageniform, obclavate, urceolate or subcylindrical, pale brown to dark brown, smooth or verruculose, composed of a venter and a collarette; transition from venter to collarette gradual, abrupt or barely perceptible. Conidia endogenous, extruded singly or in short or long chains, cylindrical, rarely obclavate or ellipsoid, straight, mostly hyaline, less frequently subhyaline or pale brown, septate, smooth or verruculose, with rounded or truncate ends, often provided with basal marginal frill or rarely fringes of wall material, slightly rounded at the apex. Teleomorph: Unknown.
Type species: Nagrajchalara yongnianii W.P. Wu & Y.Z. Diao.
Ecology/substrate/host: Saprobe on decaying leaves, branches, bark, fruit, and rotten wood.
Geographical distribution: Widely distributed (Hughes 1958; Nag Raj and Kendrick 1975).
Notes: The phylogenetic analyses with different datasets in this study clearly showed that the chalara-like fungi with aseptate and septate conidia were paraphyletic in Pezizellaceae. The species with septate conidia were further split into two distinct subclades, i.e., the Nagrajchalara agathidis clade and Cylindrocephalum aurea clade, both were with strong support in the phylogenetic analyses from different datasets (Figs. 1, 2, 3, 8, 9). The Nagrajchalara agathidis clade included majority of the studied species with septate conidia, while the C. aurea clade was only with five species (three known as Chalara aurea, C. hughesii, C. kendrickii and two as new species). These two clades were distinct from Chalara s. str. and other chalara-like genera; thus, they need to be correctly assigned to other genera.
Based on a literature study and supported by the phylogenetic analyses (Figs. 1, 2, 3, 40), Cylindrocephalum is reinstated for the Cylindrocephalum aurea clade, and Nagrajchalara is established as a new genus for the N. agathidis clade. Seven existing generic names were considered as potential generic name for this clade, and they were two asexually typified genera, Cylindrocephalum (1851) and Excioconidium (1925), and four sexually typified genera, Calycellina, Phaeoscypha, Rodwayella and Tapesina. Under the asexually typified Chalara s. lat., six asexually typified generic synonyms were included, Cylindrocephalum, Thielaviopsis, Stilbochalara, Chalaropsis, Excioconidium and Hugehsiella (Nag Raj and Kendrick 1975). Among them, the type species of Chalaropsis, Hugehsiella, Stilbochalara and Thielaviopsis were congeneric and now reclassified as Thielaviopsis in Ceratocystidaceae (Microascales). This made Cylindrocephalum (1851) and Excioconidium (1925) as the only potential choices (Hughes 1958; Nag Raj and Kendrick 1975). Cylindrocephalum, typified by C. aureum, was naturally chosen as the generic name for the Chalara aurea clade. Excioconidium was only known with the type species E. cibotti, and no living strain or molecular sequence data was available, thus its phylogenetic relationship with other chalara-like fungi remains to be studied in future. Morphologically E. cibotti, possessing 7-septate, cuneiform or obovoid, and thick-walled conidia with a broad, rounded apex, and a narrow, truncate base, is not a typical member of the N. agathidis clade.



Maximum likelihood (ML) tree based on ITS sequence data for the chalara-like anamorphic fungi in Nagrajchalara (Pezizellaceae, Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Hymenoscyphus fraxineus was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
The four sexually typified genera Calycellina, Phaeoscypha, Rodwayella and Tapesina brought into our consideration for the N. agathidis clade due to the established anamorph-teleomorph connection and phylogenetic analyses, but again none of them could be chosen as a satisfactory generic name for this clade. Five species of Chaetochalara or Chalara with septate conidia were reported to be associated with the teleomorphs of Leotiomycetes: Chaetochalara aspera associated with Calycellina carolinensis, Chaetochalara cladii associated with Phaeoscypha cladii, Chalara aotearoae associated with Rodwayella sessilis, and Chalara rubi associated with Tapesina griseovitellina (Nag Raj and Kendrick 1975; Lowen and Dumont 1984; Zhuang and Hyde 2001; Baral 2002). The connection between Chalara rubi and Tapesina griseovitellina was confirmed by pure culture study, however, Tapesina was easily excluded from the candidate name due to its remarkable difference of the teleomorph and anamorph (aseptate conidia with frills at both ends) (Baral 2002). Calycellina was excluded from the choice since it is phylogenetically distinct from the Nagrajchalara agathidis clade (Han et al. 2014; Ekanayaka 2019; Johnston et al. 2019). The genus Calycellina with 65 legitimate names (MycoBank, accessed on July 13th, 2022) was polyphyletic, indicated by the three analyzed species C. leucella, C. populina and C. punctata scattered in different clades of Pezizellaceae, i.e., C. populina and C. punctata (the type species) in the Calycina/Chalara s. str. clade (Figs. 3, 8, 9), and Calycellina leucella in the N. agathidis clade.
Phaeoscypha and Rodwayella were also excluded from the choice due to lacking solid evidence of the anamorph-teleomorph connection and clarity of the phylogenetic relationship with the type species (Han et al. 2014; Ekanayaka 2019; Johnston et al. 2019). Similar to Calycellina, the phylogenetic relationship of Phaeoscypha cladii in Leotiomycetes is yet to be determined. Rodwayella was brought into consideration due to one strain (voucher H.B. 9913, from European, not the type specimen) identified to be the type species R. sessilis of the genus clustered together with N. agathidis clade was, but was also rejected due to lacking living strain and DNA sequence from the type material. Rodwayella sessilis was originally discovered with only teleomorph from Australia. Baral (2002), on the basis of European specimen, briefly mentioned the connection of Chalara aff. aotearoae with Rodwayella, but no description was provided, and the connection was not proven experimentally. The two existing species of the genus, R. citrinula and R. sessilis seemed to be not congeneric (Fig. 8). Based on all these analyses, none of these generic names (Calycellina, Cylindrocephalum, Excioconidium, Phaeoscypha, Rodwayella and Tapesina) could be satisfactorily chosen as the generic name for the N. agathidis clade, and the new genus Nagrajchalara is created for them.
Based on morphological study and phylogenetic analyses in this study, a total of 43 species with available molecular sequences were accepted in this new genus (Figs. 41, 42, 43, 44, 45, 46, 47). Morphologically they are very variable in setae, conidiophores, phialides and conidia. Only three species, N. aspera, N. jonesii, and N. septata, are with setae among the conidiophores. The conidiophores are well-developed with multiseptate basal stalk or reduced to 1–2 basal stalk cell with a terminal phialide. The conidia are always cylindrical or short-cylindrical, with obtuse apex and truncate or obtuse base. The basal frills of conidia are often seen as well (Nag Raj and Kendrick 1975).

Colony of Nagrajchalara species on PDA in 4 weeks. a, b Nagrajchalara acuariella (ex-type strain 76100). c, d N. agathidis (78321). e, f N. angionacea (57692). g–j N. aspera (g, h 47789; i, j 76391). k, l N. aunstrupii (ex-type strain 78289). m–p N. conifericola (m, n ex-type strain 77636; o, p 50643)

Colony of Nagrajchalara species on PDA in 4 weeks. a, b Nagrajchalara curviphora (ex-type strain 76101). c–f N. ejneri (c, d ex-type strain 55373; e, f 48016). g–j N. ellispoidea (g, h ex-type strain 47768; i, j 59078). k, l N. tubakii (ex-type strain 77326). m, n N. guangcaii (ex-type strain 76112). o, p N. hangzhouensis (i, j ex-type strain 77457)

Colony of Nagrajchalara species on PDA in 4 weeks. a, b N. hangzhouensis (71055). c, d N. haituoshanensis (ex-type strain 78171). e, f N. mutabilis (50725). g, h N. inflatipes (55517). i, j N. japonica (ex-type strain 77628). k, l N. jonesii (ex-type strain 76451). m, n N. keqinii (ex-type strain 76020). o, p N. kundsonii (ex-type strain 76023)

Colony of Nagrajchalara species on PDA in 4 weeks. a, b N. nawawii (47788). c, d N. neonawawii (ex-type strain 75983). e, f N. ohmanii (ex-type strain 43147). g, h N. paraunicolor (ex-type strain 50666). i, j N. pseudoaurea (50701). k, l N. puerensis (ex-type strain 76120). m, n N. qingchengshanensis (77720). o, p N. selaginellae (43153)

Colony of Nagrajchalara species on PDA in 4 weeks. a–d Nagrajchalara sichuanensis (a, b ex-type strain 57620; c, d 57632). e, f N. sivanesanii (ex-type strain 76694). g, h N. tengii (ex-type strain 76061). i–l N. tropicalis (i, j ex-type strain 42844; k, l 45844). m, n N. tsuensis (ex-type strain 77469). o, p N. tsukairakuensis (ex-type strain 77323)

Colony of Nagrajchalara species on PDA in 4 weeks. a, b. N. septata (ex-type strain 76452). c–h Nagrajchalara tubakii (c, d 76708; e, f 75999; g, h ex-type strain 77258). i–l N. venicola (i, j ex-type strain 76011; k, l 76015). m–p N. unicolor (m, n 44014; o, p 45977)

Colony of Nagrajchalara species on PDA in 4 weeks. a–d N. xiaohuiae (a, b ex-type strain 78663; c, d 78665). e–h N. yinglanii (e, h 76400; g, h 78708). i–p N. yongnianii (i, j 78668; k, l 46066; m, n 77190; o, p ex-type strain 77394)
Except for the species accepted in this study, many existing species with septate conidia might well belong to the genus and should be transferred to the new genus. These species include Chalara alabamensis, C. aotearoae, C. bicolor, C. cibotti, C. cladii, C. curvata, C. connari, C. dictyoseptata, C. distans, C. dracophylli, C. emodensis, C. germanica, C. ginkgonis, C. gracilis, C. grandispora, C. inaequalis, C. magnispora, C. nigricollis, C. paramontellica, C. prolifera, C. pteridina, C. rhynchophiala, C. rostrata, C. scabrida, C. spiralis, C. stipitata, C. transkelensis, C. tubifera, C. urecolata, Chaetochalara proteae, and Chaetochalara ramosa. However, no living strain or DNA sequence from these species is currently available for molecular phylogenetic analyses, and their phylogenetic relationship with other species of Nagrajchalara needs to be further studied in future.
Key to species of chalara-like fungi with septate conidia
(A. for Ascoconidium; C. for Chalara s. lat. with septate conidia; Cy. for Cylindrocephalum; N. for Nagrajchalara; P. for Phaeoscypha; S. for Stipitochalara; species documented in this research are in bold)
Species with presence of setae and septate conidia
-
1.
Setae sometime developing into distinct cylindrical conidiophores with terminal phialides………2
-
1.
Setae sterile and not developing into conidiophores………4
-
2.
Setae narrowly clavate, upper part multiseptate and wider than lower part………Cy. clavatisetosum
-
2.
Setae cylindrical, tapering towards apex, widest at the lower part ………3
-
3.
Conidiophores 60–122 μm long; phialides 47–54 μm long; venter 19–21 × 4.5–6 μm; collarette 32–35 × 3–3.5 μm; conidia 16–18 × 2–2.3 µm………N. jonesii
-
3.
Conidiophores 45–57.5 μm long; phialides 32.5–50 μm long; venter 12.5–17.5 × 4.5–5 μm; collarette 17.5–25 × 2.5 μm; conidia 12.5–17.5 × 2–2.5 µm………………N. mutabilis
-
4.
Phialides verruculose………5
-
4.
Phialides smooth………6
-
5.
Phialides ampulliform, 48–77 µm long; conidia 10–27 × 2.5–4 µm………N. aspera
-
5.
Phialides subcylindrical, 70–90 µm long; conidia 9–14 × 5–7 µm………C. proteae
-
6.
Conidia 0–1-septate, 10–18 × 4–5 µm; phialides 30–45 µm long………P. cladii
-
6.
All conidia septate………7
-
7.
Phialides arising as branches of short conidiophores, subcylindrical, 25–38 µm long; conidia 9–14 × 3–4.5 µm………C. ramosa
-
7.
Phialides not arising as branches of short conidiophores………8
-
8.
Phialides ampulliform, 25–38 µm long; conidia 11–15 × 1.5–2.5 µm………N. setosa
-
8.
Phialides long then 40 μm………9
-
9.
Phialides long lageniform, 80–100 µm long; conidia 16.5–19 × 2.5–3 µm……….C. laevis
-
9.
Phialides 48–62 μm long; conidia 15–17 × 2 μm………N. septata
Species with phragmoconidia or dictyoconidia and absence of setae
-
1.
Dictyoconidia………2
-
1.
Phragmoconidia………4
-
2.
Phialides ruptured by a vertical split at the apex………3
-
2.
Phialides ruptured not by a vertical split at the apex………C. dictyoseptata
-
3.
Conidia consistently 3-septate, 30–35 × 8–10 µm……….A. purpurascens
-
3.
Conidia 3–7-septate, 40–60 × 10–14 µm………A. tsugae
-
4.
Conidia usually 3-septate………5
-
4.
Conidia usually 7-septate………11
-
5.
Conidiophores composed of a single stalk cell and a phialide; venter and collarette concolorous………6
-
5.
Conidiophores multi-septate; venter and collarette not concolorous………9
-
6.
Conidia 3–4-septate, 11–21 × 2–5 μm………C. connari
-
6.
Conidia mostly 3-septate, more than 5 μm wide………7
-
7.
Conidia 18–42 (–54) × 5–8 µm………N. unicolor
-
7.
Conidia more than 8 µm wide in average………8
-
8.
Conidia (25–)30–50(–70) × 10–12.5 µm, (2–)3(–7)-septate………C. grandispora
-
8.
Conidia 25–45(–56) × 7.5–10.5 µm, (3–)4-septate………C. magnispora
-
9.
Collarette lighter than venter, smooth-walled; phialides obclavate to cylindrical, 13–40 µm; conidia 8–18 × 2–3 µm, occasionally 0–2-septate………C. pteridina
-
9.
Collarette darker than venter………10
-
10.
Collarette rough with transverse striae; conidia 22–37 × 4–5 µm………N. inflatipes
-
10.
Collarette smooth; conidia 22–42 × 4.8–6(–7) µm, (2–)3(–6) septate………N. angustata
-
11.
Conidia clavate or obclavate………C. cibotii
-
11.
Conidia cylindrical………12
-
12.
Collarette and venter concolorous; conidia 39–56 × 8.5–11 µm………N. pulchra
-
12.
Collarette darker than venter………13
-
13.
Conidiophores mostly consisting of a single stalk cell and/or a phialide………14
-
13.
Conidiophores mostly consisting of one or more stalk cells and phialides………15
-
14.
conidia 30–45 × 6.5–7 µm………N. versicolor
-
14.
Conidia 35–50 × 3.7–4.5 μm……….N. wenyingiae
-
15.
Conidia 18–54 × 5–7 µm………N. insignis
-
15.
Conidia 50–66 × 5.5–6 µm………C. bicolor
Species with didymospore and absence of setae
-
1.
Conidia with conspicuous fringes of wall material at the base or at each end………Tapesina griseovitellina
-
1.
Conidia without such conspicuous terminal fringes………2
-
2.
Venter asperate………3
-
2.
Venter smooth………6
-
3.
Phialides with smooth-walled basal part and asperate collarettes………N. panamensis
-
3.
Phialides with basal part asperate and smooth-walled collarettes………4
-
4.
Phialides usually subcylindrical; transition from venter to collarette gradual; conidia 5–19 × 2.5–3 µm………C. scabrida
-
4.
Phialides usually ampulliform; transition from venter to collarette abrupt………4
-
5.
Ratio of mean lengths of collarette/venter = 1.9: 1; conidia 11–15 × 2.5–3 µm, without basal marginal frills………C. emodensis
-
5.
Ratio of mean lengths of collarette/venter = 5.3: 1; conidia 22–30 × 2.5–3 µm, bearing basal marginal frill………C. curvata
-
6.
Conidiophores never consisting of more than a single stalk cell and/or a phialide………7
-
6.
Conidiophores mostly consisting of one or more stalk cells and phialides………37
-
7.
Collarette darker than venter………8
-
7.
Collarette not so………17
-
8.
Collarette uniformly dark, conidial septum median………C. nigricollis
-
8.
Collarette darker in its basal part, but lighter above………9
-
9.
Transition from venter to collarette abrupt………10
-
9.
Transition from venter to collarette gradual………11
-
10.
Phialides 61–67 μm long; venter ellipsoidal, 22–28 × 7–8 μm; collarettes 34–36 × 3–3.5 μm; conidia 17–24 × 2.5–3 μm………N. agathidis
-
10.
Phialides 54–67 μm long; venter ellipsoidal, 15–20 × 6–7 μm; collarettes 39–50 × 3–3.5 μm; conidia 14–19 × 2.5–2.8 μm………N. sivanesanii
-
11.
Phialides 23–35 μm long; venter cylindrical, 11–15 × 5.5–6.5 μm; collarettes 18–23 × 2.3–2.5 μm; conidia 14–16 × 2.5–2.6 μm………N. cannonii & N. conifericola
-
11.
Phialides longer than 50 μm………12
-
12.
Phialides 85–100 μm long………13
-
12.
Phialides less than 80 μm long………14
-
13.
Phialides 85–93 μm long; venter ellipsoidal to lageniform, 17–25 × 5–7.5 μm; collarettes 37.5–50 × 3–3.5 μm; conidia 12.5–16 × 2–2.5 μm………N. pseudoaurea
-
13.
Phialides 93–100 μm long; venter ellipsoidal, 29–32 × 7–9 μm; collarettes 60–65 × 3.5–4.5 μm; conidia 15.5–20 × 2.8–3.2 μm………N. aunstrupii
-
14.
Phialides 46–58 μm long; venter ellipsoidal, 20–23 × 5.5–7.5 μm; collarettes 27–39 × 3–3.5 μm; conidia 10–13 × 2.8–3 μm………N. japonica
-
14.
Phialides longer than 60 μm; collarette longer than 40 μm………15
-
15.
Phialides 65–72 μm long; venter ellipsoidal, 23–26 × 6–8.5 μm; collarettes 45–50 × 3.2–3.5 μm; conidia 15–17 × 2.5 μm………N. tubakii
-
15.
Phialides longer than 75 μm; collarettes longer than 50 μm………16
-
16.
Phialides 75–86 μm long; venter ellipsoidal to subcylindrical, 27–30 × 6–7.6 μm; collarettes 52–55 × 3.3–3.5 μm; conidia 16.5–18.5 × 2.2–2.3 μm………N. knudsonii
-
16.
Phialides 75–87 μm long; venter ellipsoidal, 25–29 × 6.5–7.3 μm; collarettes 50–58 × 3–3.5 μm; conidia 14–17.5 × 2–2.5 μm………N. puerensis
-
17.
Transition from venter to collarette gradual………18
-
17.
Transition from venter to collarette abrupt………35
-
18.
Vegetative hyphae asperate; collarette width 2.5 µm or less; conidia 1.5–2 µm wide………C. gracilis
-
18.
Vegetative hyphae smooth………19
-
19.
Conidia less than 15 µm long………20
-
19.
Conidia usually more than 15 µm long………25
-
20.
Phialides 65–74 μm long; venter ellipsoidal, 23–29 × 6.8–7.2 μm; collarettes 46–48 × 3.5–4.5 μm; conidia 11–13 × 2.8–3 μm………N. tsukairakuensis
-
20.
Collarette shorter than 60 μm………21
-
21.
Phialides 23–39 μm long; venter lageniform, 15–17 × 5–5.5 μm; collarettes 16–20 × 3–3.3 μm; conidia 13–15 × 2.4–2.6 μm………N. qingchengshanensis
-
21.
Phialides usually more than 40 μm long………22
-
22.
Conidia 2–3 μm wide………23
-
22.
Conidia 3–3.5 μm wide………24
-
23.
Phialides 41–67 μm long; venter 10–28 × 4–11 µm; collarette 18–40 × 2.5–6 µm; conidia 8–13 × 2–4 µm………Cy. kendrickii
-
23.
Phialides 40–60 μm long; venter 20–35 × 4–5 µm; collarette 25–30 × 2.5–3 µm; conidia 11–14 × 2–2.5 µm………N. truncata
-
24.
Phialides 57–63 μm long; venter ellipsoidal, 20–29 × 5.5–6.4 μm; collarettes 30–34 × 3–3.3 μm; conidia 10–15 × 2.3–2.5 μm………N. haituoshanensis
-
24.
Phialides 45–57 μm long; venter ellipsoidal, 16–24 × 5–6 μm; collarettes 28–32 × 3–3.5 μm; conidia 13–15 × 2.2–2.5 μm………N. keqinii
-
25.
Conidia 3–4.5 µm wide………26
-
25.
Conidia 2–2.5 µm wide………30
-
26.
Conidia > 3.5 μm wide………27
-
26.
Conidia less than 3.5 µm wide………28
-
27.
Phialides 70–80 μm long; venter subcylindrical to subellipsoidal, 22–30 × 9–10 μm; collarettes 50–60 × 4–5.5 μm; conidia 18–20 × 4–4.5 μm………N. tengii
-
27.
Phialides 60–74 μm long; venter ellipsoidal, 30–34 × 7–8 μm; collarettes 35–40 × 3.7–4.5 μm; conidia 14.5–16 × 3.6–3.8 μm………N. tropicalis
-
28.
Phialides 41–51 μm long; venter ellipsoidal, 18.5–26 × 6–7 μm; collarettes 21–34 × 3.5–4.2 μm; conidia 15.5–20 × 2.8–3.2………N. hangzhouensis
-
28.
Phialides longer than 50 μm; collarettes longer than 35 μm………29
-
29.
Phialides 53–64 μm long; venter 16–23 × 7.6–9 μm; collarettes 35–40 × 3.5–4.5 μm; conidia 13–18 × 3–3.5 µm………N. angionacea
-
29.
Phialides 59–67 μm long; venter ellipsoidal, 21–26 × 6.5–7.5 μm; collarettes 37–44 × 3.8–4.5 μm; conidia 13–16.5 × 3–3.2 μm………..N. tubakii
-
30.
Venter less than 25 μm long………31
-
30.
Venter longer than 25 μm………33
-
31.
Phialides 25–40 µm long; venter 7–15 × 4.5–6 μm; collarettes 20–30 × 2.5–3 μm; conidia (15.5–)18–20(–22.5) × 2–2.5 µm………C. dracophylli
-
31.
Phialides long than 40 μm; collarettes longer than 30 μm in average………32
-
32.
Phialides 40–82 µm long; venter 14–17 × 6–7 μm; collarette 30–53 × 3–4 μm; conidia 15–18 × 2–2.5 µm………C. alabamensis
-
32.
Phialides 48–63 μm long; venter ellipsoidal, 20–22 × 5–7.5 μm; collarettes 25–37 × 3–3.5 μm; conidia 15–21 × 2.3–2.5 μm………N. tsuensis
-
33.
Phialides 89–96 μm long; venter ellipsoidal, 40–48 × 6.5– μm; collarettes 48–55 × 3.3–3.6 μm; conidia 15–18.5–2.5–2.8 μm………N. venicola
-
33.
Venter less than 35 μm long………34
-
34.
Phialides 75–85 μm long; venter ellipsoidal, 23–35 × 7.5–8.5 μm; collarettes 48–60 × 4–5 μm; conidia 14–18 × 2.3–2.5 μm………N. ohmanii
-
34.
Phialides 68–80 μm long; venter ellipsoidal, 25–30 × 7–9 μm; collarettes 40–55 × 3.5–4.5 μm; conidia 22–27 × 3.5 μm………N. yongnianii
-
35.
Venter ellipsoidal or subcylindrical; conidia 12–17 × 2.5–3.5 µm………Cy. hughesii
-
35.
Venter globose………36
-
36.
Venter 7–8.5 µm wide; conidia 11–15 × 2.5 µm………N. yunnanensis
-
36.
Venter globose, 7.5–10 µm wide; conidia 12–23 × 2.5–3.5 µm………C. rhynchophiala
-
37.
Transition from venter to collarette abrupt………38
-
37.
Transition from venter to collarette gradual………41
-
38.
Collarette darker than venter………39
-
38.
Collarette not so………40
-
39.
Collarette uniformly dark, conidia 9.5–18 × 2–2.5 µm………C. inaequalis
-
39.
Collarette darker in its basal part, but lighter above, conidia 11–19 × 2–2.5 µm………C. tubifera
-
40.
Conidiophores 78–230 × 4.5–8 μm; phialides urceolate, 28–105 μm long; venter obconic, 18–37 × 8–13 μm; collarettes 21–72 × 3–6 μm; conidia 10–18 × 2.5–4 μm………C. urceolata
-
40.
Phialides not urceolate, Conidia 10–17 × 2.4–3 µm………C. paramontellica
-
41.
Phialides versicolorous, basal part of collarette darker than other part………42
-
41.
Phialides concolorous………46
-
42.
Conidiophores 2–3-septate, 30–43 × 5–6 μm; phialides 27–37 μm long; venter cylindrical, 10–15 × 5–7 μm; collarettes 16–20 × 2.6–2.8 μm; conidia 14–16 × 2.4–2.6 μm………N. cannonii
-
42.
Conidiophores more than 45 μm long………43
-
43.
Conidiophores 1–6-septate, 60–85 × 4–5 μm; phialides 40–60 μm long; venter lageniform to cylindrical, 17–24 × 5–6.5 μm; collarettes 21–34 × 2.5–2.8 μm; conidia 11–15 × 2.3–2.5 μm………N. yinglaniae
-
43.
Conidia more than 15 μm in average………44
-
44.
Conidiophores 2–4-septate, 65–76 × 6–7 μm; phialides 60–70 μm long; venter subcylindrical to ellipsoidal, 23–27 × 9–11 μm; collarettes 45–47 × 4–4.5 μm; conidia 16.5–20 × 3–3.5 μm……N. ellipsoidea
-
44.
Conidiophores longer than 85 μm………45
-
45.
Conidiophores 3–4-septate, 110–135 × 4–5 μm; phialides 94–100 μm long; venter cylindrical, 41–54 × 6.5–8.5 μm; collarettes 51–54 × 3.7–4.3 μm; conidia 16.5–21 × 2.5–3 μm………N. guangcaii
-
45.
Conidiophores 3–5 septate, 90–110 × 5–6.5 μm; phialides 67–88 μm long; venter cylindrical to ellipsoidal, 29–34 × 8–9 μm; collarettes 37–52 × 3–3.6 μm; conidia 13.5–17 × 2.7–3 μm……N. curviphora
-
46.
Conidiophores with repeated percurrent proliferations, conidia 10–16 × 3–4.5 µm………C. prolifera
-
46.
Conidiophores not repeatedly proliferating………47
-
47.
Conidia more than 3.5 µm wide………48
-
47.
Conidia 2–3.5 µm wide………50
-
48.
Conidia 29–34 × 7–9.5 µm………N. novozymia
-
48.
Conidia less than 20 µm long and 7 µm wide………49
-
49.
Collarette narrowing toward apex, 25–29 × 4.5–6.5 µm; venter 14–17 × 6.5–8.5 µm; conidia 10–18 × 3.5–4 µm………N. selaginellae
-
49.
Collarette cylindrical, not narrowing towards apex, 31–42 × 5–6.5 µm; venter 22–33 × 8–10 µm; conidia 13–18 × 3.5–4 µm………C. aotearoae
-
50.
Conidia 5–12 µm long………51
-
50.
Conidia mostly more than 12 µm long………53
-
51.
Conidiophores pale brown, 1–3 septate; conidia 9–10 × 1.2–1.5 µm………N. intermedia
-
51.
Conidiophores dark brown, up to 6–8 septate………52
-
52.
Conidia always 1-septate, 8–12 × 1.8–2.5 µm; on Picea………S. piceae-abietis
-
52.
Conidia 0–1 septate, 5–12 × 2–3 µm, on Ginkgo………C. ginkgonis
-
53.
Conidia 2–2.5 µm wide………54
-
53.
Conidia more than 2.5 µm wide………58
-
54.
Conidiophores not arising from a thin stromatic layer of cells, 70–120 × 3.5–4 µm; phialides 30–34 µm long; venter 12–16 × 5–6 µm; collarette 17–19 × 2.5–3 µm; conidia 11–15 × 2–2.5 µm………C. stipitata
-
54.
Collarettes longer than 50 µm………55
-
55.
Conidiophores 4–5-septate, 95–135 × 4–5 μm; Phialides 55–67 μm long; venter cylindrical to ellipsoidal, 20–27 × 5.5–6.5 μm; collarettes 31–34 × 3.5–3.8 μm; conidia 13–15.5 × 2.5–2.6 μm………N. acuariella
-
55.
Conidiophores less than 95 μm long………56
-
56.
Conidiophores 1–3-septate, 50–70 × 4–5 μm; Phialides 45–57 μm long; venter lageniform to cylindrical, 25–35 × 6.5–6 μm; collarettes 28–30 × 3.5–4 μm; conidia 12–18 × 2.5–3 μm………N. morganjonesii
-
56.
Collarettes longer than 30 μm………57
-
57.
Conidiophores 1–4-septate, 60–85 × 5–6.5 μm; phialides 54–67 μm long; venter subcylindrical, 20–28 × 5–6.5 μm; collarettes 33–45 × 3–3.5 μm; conidia 12–15 × 2–2.5 μm………N. sichuanensis
-
57.
Conidiophores 2–6-septate, 60–90 × 4–5 μm; Phialides 45–55 μm long; venter lageniform to cylindrical, 18–23 × 5.5–6.5 μm; collarettes 30–38 × 3.5–4 μm; conidia 9–14 × 2–2.5 μm………Cy. aureum
-
58.
Conidiophores 1–2-septate, 82–110 × 4–5 μm; Phialides 73–92 μm long; venter ellipsoidal, 25–33 × 7–8 μm; collarettes 50–56 × 4–4.5 μm; conidia 11–15 × 3.5–3.6 μm………N. neonawawaii
-
58.
Collarettes less than 50 μm long………59
-
59.
Conidiophores well-developed, with cylindrical and multiseptate stipes and terminal collarettes……60
-
59.
Conidiophores reduced and without cylindrical stipes………64
-
60.
Conidiophores 56–135 long; phialides 42–63 long; venter 16–28 × 5.5–8; collarettes 21–37 × 2.5–4.5; conidia 17–28 × 2.5–3.5 µm often forming long, helicoids chains………C. spiralis
-
60.
Conidia not forming helicoids chains………61
-
61.
Conidiophores 110–195 × 4–6 µm; phialides 40–78 µm long; venter 18–28 × 6–9.5 µm; collarettes 26–47 × 3–6 µm; transition distinct; conidia 9–13 × 2.5–3 µm………C. rostrata
-
61.
Conidia more than 15 µm long………62
-
62.
Conidiophores 2-septate, 74–90 × 4–4.5 μm; Phialides 60–74 μm long; venter lageniform, obconic, 24–26 × 6–7 μm; collarettes 44–47 × 3.5–3.7 μm; conidia 15–18 × 3–3.6 μm………Cy. zhejiangense
-
62.
Conidiophores 2–7-septate, up to 125 μm long………63
-
63.
Conidiophores 50–120 × 5.5–9 µm; phialides 33–77 µm long; venter 15–26 × 5–11 µm; collarette 14–57 × 2.5–5 µm, widening toward apex; conidia 12–20 × 2–3.5 µm………N. acuaria
-
63.
Conidiophores 2–7-septate, 87–125 µm high, Phialides 70–80 μm long; venter subcylindrical to subellipsoidal, 22–30 × 9–10 μm; collarettes 50–60 × 4–5.5 μm; conidia 18–20 × 4–4.5 μm………N. ejneri
-
64.
Conidiophores 1–3-septate, 60–100 µm long; phialides 60–80 µm long; venter 20–30 × 6–10 µm; collarettes 35–50 × 3–4 µm; conidia 18–22 × 2.5–3.5 µm; on Dracophyllum………C. distans
-
64.
Venter more than 30 μm long………65
-
65.
Conidiophores up to 4-septate, up to 93 long; phialides 65–88 µm long; venter 30–35 × 6–8 µm; collarettes 40–55 × 3–4 µm; conidia 12–17 × 2.5–3 µm; on dead wood………C. transkeiensis
-
65.
Conidiophores 1–3-septate, 82–90 × 5–6 μm; phialides 65–86 μm long; venter subcylindrical to ellipsoidal, 25–37 × 5.5–6.5 μm; collarettes 30–47 × 3.5–4 μm; conidia 14–19 × 3–3.5 μm……N. nawawii
Nagrajchalara acuaria (Cooke & Ellis) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 48, MycoBank MB845271.

Nagrajchalara acuaria (b–f, k–u Wu8182b) and Nagrajchalara acuariella (a, g–j Wu15193, holotype). a–f Conidiophores and phialidic conidiogenous cells. g–u Conidia. Scale bar: 10 µm for a–f, 5 µm for g–u
≡ Chalara acuaria Cooke & Ellis, Grevillea 6(no. 39): 89, 1878.
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae, 2.5–3.5 µm wide. Anamorph: Stroma absent or poorly developed and consisted of a few aggregated irregular cells. Setae absent. Conidiophores directly arising from superficial hyphae or the basal stroma, scattered or aggregated, erect, simple, subcylindrical, obclavate, 45–60 × 5–8 µm, basal cells slightly swollen, dark brown at the base, becoming medium brown towards the apex, 3–6-septate, smooth or verruculose, with 1–2 percurrent proliferations, terminating in a phialide. Conidiogenous cells integrated, terminal, obclavate, 60–70 μm long, medium brown, slightly darker in the lower part of collarettes, smooth, composed of a venter and a collarette; transition from venter to collarette gradual; venters subcylindrical, subellipsoidal, 23–30 × 6–7 µm; collarettes cylindrical, long funnel-shaped, 38–42 × 3.5–5 µm, broadest at the opening, concolorous or slightly darker in the lower part; ratio of mean lengths of collarette and venter = 1.5:1. Conidia endogenous, extruded in short chains, cylindrical, 13–17.5 × 2.8–3 µm, straight, hyaline, 1-septate, smooth, slightly rounded at the apex, truncate at the base and with a basal frill; mean conidium length/width ratio = 5.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded at margin, aerial mycelium poorly developed, grey to brown, reverse concolorous or slightly darker, sterile, up to 5 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangxi Province, Daming Mountain, on dead leaves of ?Cinnamomum sp., 18 December 1997, Wenping Wu, Wu1382f; Hubei Province, Wufeng Mountain, on dead leaves of unidentified tree, 14 September 2004, Wenping Wu, Wu8182b. Living strains: 43205 (from Wu1382f) and CGMCC3.23383 (= 50664, from Wu8182b).
Ecology/substrate/host: Saprobe on dead leaves of Cinnamomum, needles of Abies, Pinus and Podocarpus.
Geographical distribution: China, New Zealand and USA (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Nagrajchalara acuaria is characterized by multiseptate conidiophore, abrupt transition from venter to collarette, subcylindrical to subellipsoidal venter, and 1-septate and cylindrical conidia with truncated base bearing a basal frill. Although it has been only reported on needles of coniferous trees, the fungus on decaying leaves of ?Cinnamomum sp. in China fits well to the species concept as described and illustrated by Nag Raj and Kendrick (1975). Variations on the length and septation of conidiophores were observed within the same collection and also between the two studied collections.
Identical ITS sequences were generated from the two studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 50664 included Calycellina fagina (OL752703, 94% identity) many unnamed fungi of Leotiomycetes.
Nagrajchalara acuariella W.P. Wu & Y.Z. Diao, sp. nov., Fig. 49, MycoBank MB845219.

Nagrajchalara acuariella (Wu15193, holotype). a–d, f–k Conidiophores and phialidic conidiogenous cells. l Basal part of conidiophores. e, m–p Conidia. Scale bar: 10 µm for a–d, 5 µm for e–p
Etymology: Refers to its morphological similarity with Chalara acuaria.
Typification: China, Yunnan Province, Xishuangbanna, on decaying leaves of unidentified tree, 6 Dec. 2018, Y. Zhang, Holotype HMAS352191 (= Wu15193), ex-type strain CGMCC3.23454 (= NN76100).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, scattered or aggregated at the basal stroma, erect, straight of slightly curved, simple, subcylindrical, (75–)95–135 μm high, 4–5 µm wide, basal stalk up to 62 μm high, basal cells slightly swollen up to 10 μm wide, dark brown at the base, becoming medium brown towards the apex, 4–5-septate, smooth, occasionally with 1–2 percurrent proliferations, terminating in a phialide. Conidiogenous cells integrated, terminal, phialidic, obclavate, lageniform, medium brown, becoming pale brown towards the apex, smooth, 55–67 μm long; transition from venter to collarette abrupt; venters cylindrical to lageniform, 20–27 × 5.5–6.5 μm; collarettes cylindrical, 31–34 × 3.5–3.8 µm, concolorous or slightly darker in the lower part; ratio of mean length of collarette and venter = 1.4:1. Conidia endogenous, extruded in short chains, cylindrical, 13–15.5 × 2.5–2.6 µm, straight, hyaline, 1-septate, smooth, truncate at the base and with a basal frill, obtuse or slightly rounded at the apex; mean conidium length/width = 5.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, brown to dark brown, reverse concolorous or slightly darker, with yellow brown pigment diffused into agar, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara acuariella is morphologically similar to N. acuaria, but differs in longer and narrower conidiophores, and narrower conidia (Nag Raj and Kendrick 1975). The ITS sequence from the ex-type strain had only 4 bp differences from those in N. acuaria.
Nagrajchalara agathidis (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 50, MycoBank MB845272.

Nagrajchalara agathidis (Wu17542). a–g, i–l Conidiophores and phialidic conidiogenous cells. h Basal part of conidiophores. m–p Conidia. Scale bar: 10 µm for a–g; 5 µm for h–p
≡ Chalara agathidis Nag Raj & W.B. Kendr., Monogr. Chalara Allied genera: 81, 1975.
Description on the natural substrate: Colonies effuse, pale brown, sparse. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, smooth-walled, branched hyphae of 1.5–3 µm wide. Anamorph: Stroma absent or poorly developed and consisting of a few aggregated irregular cells. Setae absent. Conidiophore directly arising from superficial hyphae or aggregated cells, reduced to sessile phialides or composed of a terminal phialide and a short stalk with 1–3 cells, solitary or aggregated loosely, simple, erect, straight, obclavate, subcylindrical, 80–95 × 6–7 µm, basal cells inflated, constricted at the septa, brown, thick- and smooth-walled. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, obclavate, lageniform, 80–90 µm long, medium brown to brown, smooth-walled, consisting of a venter and collarette, versicolorous, significantly darker in the transition region from venter to collarette and lower part of collarette; transition from venter to collarette gradual; venter subellipsoidal to subcylindrical, 22–27 × 6–8 µm, brown; collarettes cylindrical, 50–60 × 3–4 µm, lowers part distinctly darker than venter and upper part of collarette; ratio of mean lengths of collarette and venter = 2.2:1. Conidia endogenous, extruded in short or long chains, cylindrical, 19–23 × 2.5–3 µm, apex rounded or flattened, base subtruncate with a frill, hyaline, uniseptate, smooth- and thin-walled, guttulate; mean conidium length/width = 7.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, brown in the middle, becoming pale brown to white in the margins, reverse concolorous or slightly darker, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangdong Province, Shenzhen, Yangtaishan Forestry Park, on dead leaves of tree, 17 October 2020, Wenping Wu, Wu17542; Guangxi Province, Shiwandashan, on dead leaves of ?Magnolia sp., 29 December 1997, Wenping Wu, Wu1507a; Guangxi Province, Shiwandashan, on dead fruits of Leguminosae, 28 Dec 1997, Wenping Wu, Wu1481b. Living strains: 42964 (from Wu1481b), 43036 and 43162 (from Wu1507a), CGMCC3.23444 (= NN78321, from Wu17542).
Ecology/substrate/host: Saprobe on dead leaves of Agathis australis and ?Magnolia sp.
Geographical distribution: China and New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Nagrajchalara agathidis is characterized by simple conidiophore with a 1–3-celled basal stalk and a terminal phialide, versicolorous conidiogenous cells with gradual transition from venter to collarette, subellipsoidal to subcylindrical venter, and 1-septate and longer conidia (Nag Raj and Kendrick 1975). Morphologically N. agathidis superficially resembles N. angionacea, but differs from it by longer and narrower venter, collarette and conidia (Nag Raj and Kendrick 1975). It is also similar to Chalara alabamensis, but differs in its versicolorous phialides (Morgan-Jones and Ingram 1976). The Chinese collections were with significantly longer phialides and collarettes (phialides 61–67 µm long, venters 22–28 µm long, and collarettes 34–46 µm long in the original description, Nag Raj and Kendrick 1975).
Morphologically and phylogenetically N. agathidis is also closely related to several new species described in this work, such as N. aunstrupii, N. knudsonii, N. puerensis, N. tsukairakuensis and N. tubakii, but differs from them in longer phialides (80–90 µm long) and conidia (19–23 × 2.5–3 µm), and different ITS sequence. Nagrajchalara aunstrupii differs from N. agathidis in its slightly longer phialides and shorter conidia. In addition, the ITS sequences from these two species are with 15 bp differences. The phialides of N. knudsonii (75–86 µm long) and N. puerensis (75–87 µm long) are in similar shape and size with those of N. agathidis, but their conidia are shorter and narrower, and their ITS sequences have 14 and 33 bp differences respectively from the one in N. agathidis. In N. tubakii (conidia 13–16.5 × 3–3.2 µm), the phialides and conidia are much shorter than those in N. agathidis, and furthermore their ITS sequences have 15, 9 and 16 bp differences respectively from N. agathidis.
Identical ITS sequences were generated from the four studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 43162 included Chalara sp. (LT629156, 94% identity), Chalara sp. (LT629155, 94% identity), Epacris pulchella (AY627819, 95% identity), Hymenoscyphus serotinus (KU204568, 95% identity), and many unnamed fungi of Leotiomycetes. In the phylogenetic tree, N. agathidis clustered together with N. aunstrupii, N. selaginellae and N. tsukaiakuensis.
Nagrajchalara angionacea (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 51, MycoBank MB845273.

Nagrajchalara angionacea (Wu13248). a–f, h–m Conidiophores and phialidic conidiogenous cells. g Basal stroma. n–p Conidia. Scale bar: 10 µm for a–f; 5 µm for g–p
≡ Chalara angionacea Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera: 85, 1975.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma present, composed of dark brown, thick-walled and irregular cells. Conidiophore arising from basal stroma, 3–5 aggregated in clusters, erect, straight, obclavate, lageniform, 65–80 µm long and 4–5 µm wide, consisting of one basal stalk cell and a phialide, the basal cell incorporated into hyphae, medium brown to brown, smooth-walled. Conidiogenous cells integrated, erect, straight, obclavate, lageniform, 60–75 µm long, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual; venters ellipsoidal, 17–23 µm long and 7.5–9 µm wide; collarettes cylindrical, 39–42 µm long, 3–3.5 µm wide, concolorous with venter; ratio of mean lengths of collarette and venter = 2:1. Conidia endogenous, extruded in short or long chain, cylindrical, 16–22 × 3–3.5 µm, hyaline, aseptate, base truncated or flattened and with a basal frill, apex rounded or obtuse; mean conidium length/width = 5.9:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, soil brown, reverse dark brown, with yellow brown pigment diffused into agar, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Sichuan Province, Ya An, Yucheng, Wanguan, Bifengxia, on dead leaves of unidentified tree, 15 December 2013, Wenping Wu, Wu13248. Living strains: 57693 (from Wu13248a), CGMCC3.23400 (= NN57692, from Wu13248b).
Ecology/substrate/host: Saprobe on dead leaves of Beilschmiedia tawa and Knightia excelsa.
Geographical distribution: China and New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Compared with the original description (conidiophores 60–65 µm long, phialides 53–64 µm long, and conidia 13–18 × 3–3.5 µm long), this fungus from the Chinese specimens was with slightly longer conidiophores, phialides and conidia (Nag Raj and Kendrick 1975).
Identical ITS sequences were generated from the two studied strains. In the phylogenetic tree, N. angiomacea clustered together with N. hanzhouensis, N. morganjonesii and N. tsuensis. Morphologically N. angiomacea differs from N. hangzhouensis in its longer phialides and slightly larger conidia; and furthermore, the ITS sequences from these two species have 17 bp differences; differs from N. morganjonesii (venter 25–35 µm long, conidia 12–18 × 2.5–2.8 µm) in shorter venter, wider conidia and 11 base pairs in ITS sequence; and differs from N. tsuensis (conidia 15–21 × 2.3–2.5 µm) in wider conidia and 37 base pairs in ITS sequence. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 57693 included Hymenoscyphus serotinus (KU204568, 96% identity) and many unnamed fungi of Leotiomycetes.
Nagrajchalara angustata (T. Kowalski & Halmschl.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845275.
≡ Chalara angustata T. Kowalski & Halmschl., Mycol. Res. 100(9): 1112, 1996.
Description on the natural substrate: Anamorph: Conidiophores multiseptate, brown to dark brown, 1–4 (–15)-septate, 110–160 μm long, 6–11 μm wide at the base. Conidiogenous cells phialidic subcylindrical, occasionally lageniform, 96–125 μm long; transition from venter to collarette gradual; venter subcylindrical to ellipsoid, 38–60 × 10–12 μm; collarette cylindrical, 58–87 × 5.6–7 μm. Conidia cylindrical, 22–42 × 4.8–6 μm, apex obtuse, base truncate and with a marginal frill, hyaline, 3-septate (Kowalski and Halmschlager 1996). Teleomorph: Unknown.
Ecology/substrate/host: From roots of Quercus petraea. Teleomorph: Unknown.
Geographical distribution: Austria (Kowalski and Halmschlager 1996).
Description and illustration: Kowalski and Halmschlager (1996).
Notes: Morphologically N. angustata is closely related to N. unicolor (Nag Raj and Hughes 1974, 1975). Nagrajchalara unicolor is, however, easily distinguished from N. angustata by its conidiophores composed of a single stalk cell without constriction at the base, concolorous venter and collarette, and wider conidia (Nag Raj and Kendrick 1975). The phylogenetic analyses by using different datasets clearly placed N. angustata with 3-septated conidia and other Nagrajchalara species with 1-septate conidia into the same clade.
Nagrajchalara aspera (Pirozy. & Hodges) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 52, MycoBank MB845274.

Nagrajchalara aspera (Wu16924). a–c Setae and conidiophores. d–e Conidiophores and conidiogenous cells. f–i Part of conidiophores bearing phialidic conidiogenous cells. j–p Conidia. Scale bar: 10 µm for a–e, 5 µm for f–p
≡ Chaetochalara aspera Pirozy. & Hodges, Can. J. Bot. 51(1): 157, 1973.
≡ Chalara aspera (Pirozy. & Hodges) P.M. Kirk in Kirk & Spooner, Kew Bull. 38(4): 580, 1984.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3.5 µm wide. Anamorph: Stroma absent or present, composting of dark brown, irregular cells. Setae arising from superficial hyphae or aggregated cells, solitary or in clusters with conidiophores, erect, straight or slightly flexuous, simple, cylindrical, up to 300 µm long, 4–5 µm wide, slightly inflated at the base to a width of up to 6 µm, gradually tapering toward the apex to a width of 2–2.5 µm, apex acute or obtuse, 4–7 septate, dark brown to reddish brown, smooth, sterile, or occasionally terminating into a phialide. Conidiophores arising from superficial hyphae or aggregated cells, associated with setae, erect, straight or slightly flexuous, simple, cylindrical, up to 50 µm long and 5–5.5 µm wide, medium brown, 1–5-septate, smooth. Conidiogenous cells integrated, terminal, straight or slightly curved, subcylindrical, obclavate, lageniform, 65–90 µm long, pale to medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual; venters subcylindrical, 15–32 µm long and 5.5–6 µm wide; collarettes cylindrical, 40–50 µm long, 4–4.5 µm wide, concolorous with venter, sparsely aspirate; ratio of mean lengths of collarette and venter = 1.9:1. Conidia endogenous, extruded in short or long chains, cylindrical, 17–20 × 2.8–3 µm, base truncated, without basal frill, apex rounded or flattened, hyaline, 1-septate, guttulate; mean conidium length/width ratio = 6.4:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, reverse concolorous or slightly darker, sterile, up to 13 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangdong Province, Dinghushan, on dead leaves of unidentified plant, 9 October 1998, Wenping Wu, Wu1920a; Guangdong Province, Zhaoqing, Dinghushan, on unidentified plant, 3 Mar. 2012, Wenping Wu, Wu12121; Guangdong Province, Shenzhen, Lianhuashan Park, on dead leaves of unidentified tree, 11 November 2019, Wenping Wu, Wu17252; Guangxi Province, Shiwandashan, on dead leaves of ?Rhododendron sp., 30 Dec 1997, Wenping Wu, Wu1546c; Guangxi Province, Shiwandashan, on dead leaves of unidentified plant, 30 Dec 1997, Wenping Wu, Wu1554a; Guangxi Province, Shiwandashan, on dead leaves of unidentified plant, 29 Dec 1997, Wenping Wu, Wu1517a; Yunnan Province, Kunming, Heilongtan Park, on dead leaves of unidentified tree, 19 October 2003, Wenping Wu, Wu7130a; Zhejiang Province, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 November 2018, Wenping Wu, Wu16052; Japan, Mie Prefecture, Tsu, Mie Center for the Arts, on dead leaves of unidentified tree, 3 October 2019, Wenping Wu, Wu16897, Wu16907, Wu16908, Wu16924, Wu16925 and Wu16949. Living strains: 42933 (from Wu1517a), 43010 (from Wu1546c), 43132 (1546c), 44447 (from Wu1554a), 45184 (from Wu1920a), CGMCC3.273374 (= NN 47789, from Wu7300a), 54256 (from Wu12121), CGMCC3.23421 (= 76391, from Wu16052), 76449 (from Wu16052a), 77312 (from Wu16897), 77314 (from Wu16907), 77315 (from Wu16908), 77319 (from Wu16924r), 77396 (from Wu16925b) and 77727 (from Wu17252).
Ecology/substrate/host: Saprobe on dead leaves of different plants, including Knightia excelsa, Myrica cerifera, Persea borbonia, ?Rhododendron sp. another plants.
Geographical distribution: China, Japan, New Zealand and USA (Nag Raj and Kendrick 1975).
Description and illustration: Pirozynski and Hodges (1973) and Nag Raj and Kendrick (1975).
Notes: Nagrajchalara aspera differs from other species of Chalara s. lat. with setae in possessing distinctly developed conidiophores with, aspirate collarettes, and 1-septate conidia (Nag Raj and Kendrick 1975). The fungus usually produces conidiophores with a 1–2-celled basal stalk and a terminal phialide. However, in the specimen Wu16924, the conidiophores are 3–6-septate, longer in size, and frequently with percurrent proliferations. The ITS sequences generated from 15 different strains in this study were almost identical, except for the sequence from the strain 47798, which was slightly different from others. Nagrajchalara panamensis also has similar conidiophores with asperate collarettes and 1-septate conidia (Crous et al. 2016a, b), but differs from N. aspera by lacking sterile setae among conidiophores and lower identity (< 90%) in the ITS sequence.
Consistent associations between Chaetochalara aspera and its teleomorph Calycellina carolinensis were described by different authors (Nag Raj and Kendrick 1975; Lowen and Dumont 1984; Zhuang and Hyde 2001). However, none of these connections was based on pure culture or phylogenetic analysis, thus this needs to be further validated in future study. No living strain of Calycellina carolinensis could be found in any of those public fungal collections.
Nag Raj and Kendrick (1975) observed a constantly connection of Nagrajchalara aspera with Calycellina carolinensis on both the holotype specimen and other collections made in New Zealand. No DNA sequence data is available from C. carolinensis for molecular phylogenetic analyses. Our phylogenetic analyses shows that N. aspera and members of Calycellina including the type species of the genus Calycellina punctiformis, are grouped into two distinct groups.
Nagrajchalara aunstrupii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 53, MycoBank MB845220.

Nagrajchalara aunstrupii (Wu17537, holotype). a–e, h–k Conidiophores and phialidic conidiogenous cells. f, g, l, m Conidia. Scale bar: 10 µm for a–e; 5 µm for f–m
Etymology: Named after the former Vice President and microbiologist Knud Aunstrup from Novo Nordisk.
Typification: China, Guangdong Province, Shenzhen, Yangtaishan Forestry Park, on dead leaves of unidentified tree, 17 October 2020, W.P. Wu, Holotype HMAS352192 (= Wu17537), ex-type strain CGMCC3.23443 (= NN78289).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Stroma absent. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or in small group, reduced to sessile phialide or composed of a terminal phialide and a short basal stalk cell, erect, straight or slightly curved, obclavate, 98–105 µm long, 4–5 µm wide at the base, basal cell up to 8 μm wide, medium brown, smooth. Conidiogenous cells integrated or discrete, phialidic, erect, straight or slightly curved, lageniform to obclavate, 93–99 µm long, medium brown, versicolorous, lower part of collarettes significantly darker than venter and upper part of collarettes, smooth; transition from venter to collarette abrupt or gradual; venters ellipsoidal, subcylindrical, 29–32 µm long and 7–9 µm wide, brown; collarettes cylindrical, (42–) 60–65 × 3.5–4.5 µm versicolorous, lower part significantly darker; ratio of mean lengths of collarette and venter = 2:1. Conidia endogenous, extruded in short chains, cylindrical, 15.5–20 × 2.8–3.2 µm, base truncated with short frills, apex rounded or flattened, hyaline, uniseptate, guttulate, smooth- and thin-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, brown to dark brown in the middle part, becoming light brown to white towards the margins, reverse concolorous, sterile, up to 13 mm on PDA at 25 °C in 4 weeks.
Other living strains: 78290 (from holotype Wu17537).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara aunstrupii is characterized by reduced conidiophores, versicolorous and obclavate phialides with ellipsoidal venter, and 1-septate conidia (15.5–20 × 2.8–3.2 μm). Morphologically it is similar to N. agathidis (phialide 61–67 μm long, venter 22–28 × 7–8 μm, collarettes 34–46 × 3–3.5 μm long; conidia 17–24 × 2.5–3 μm), but differs by longer phialides and slightly shorter conidia (Nag Raj and Kendrick 1975). The ITS sequence was generated from the ex-type strain, and it had 15 bp differences from the one of N. agathidis.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 78298 included Hymenoscyphus serotinus (KU204568, 97% identity), Chalara sp. (LT629156, 95% identity) and many unnamed fungi of Leotiomycetes.
Nagrajchalara cannonii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 54, MycoBank MB845221.

Nagrajchalara cannonii (Wu16069, holotype). a–l, n Conidiophores and conidiogenous cells. m, o–q Conidia. Scale bar: 10 µm for a–f, 5 µm for g–r
Etymology: Named after the former mycologist Paul Cannon.
Typification: China, Zhejiang, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu (HMAS 352193 (= Wu16066), holotype; ex-type strain 76454.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly formed from superficial hyphae or aggregated cells, obclavate, 35–44 µm long, 4–5 µm wide at the base, solitary or in small group, 1-septate, medium brown, smooth. Conidiogenous cells integrated, determinate, phialidic, monoblastic, erect, straight or slightly curved, medium brown, smooth, lageniform to obclavate, 25–35 µm long, consisting of a venter and a collarette, transition from venter to collarette abrupt; venters cylindrical, 11–15 µm long and 5.5–6.5 µm wide brown; collarettes cylindrical, darker than venter, (13.5) 18–23 (–26) × 2.3–2.5 µm, medium brown; ratio of mean lengths of collarette and venter = 1.6:1. Conidia endogenous, cylindrical, 14–16 × 2.4–2.6 µm, hyaline, uniseptate, base truncate without frill, apex rounded or flattened, extruded in short chains; mean conidium length/width ratio = 6:1. Teleomorph: Unknown.
Other material examined: China, Zhejiang, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu (Wu16069). Living strains: 77455 (from Wu16066) and76549 (from 16069).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara cannonii is characterized by relatively short conidiophores with versicolorous phialides, abrupt transition from venter to collarette, cylindrical to lageniform venter, and relatively long 1-septate conidia without basal frill. Nagrajchalara agathidis is morphologically similar to N. cannonii, but differs in longer conidiophores (65–71 μm), collarettes (34–26 μm) and conidia (17–24 μm) (Nag Raj and Kendrick 1975). It also resembles Chalara hughesii, but differs in versicolorous phialide with slight darker collarette (Nag Raj and Kendrick 1975).
ITS sequences generated from the three strains (76454, 76455 and 76549) were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77454 included Calycellina fagina (OL752703, 94% identity) and many unnamed fungi of Leotiomycetes.
Nagrajchalara conifericola W.P. Wu & Y.Z. Diao, sp. nov., Fig. 55, MycoBank MB845465.

Nagrajchalara conifericola (a–k Wu8183; l–q 17025). a–o Conidiophores and conidiogenous cells. p, q Conidia. Scale bar: 10 µm for a; 5 µm for b–q
Etymology: Refers to its host plant as coniferous tree.
Typification: China, Zhejiang Province, Deqing, Moganshan, Luhuadang, on dead needle of pine tree, 16 October 2019, Wenping Wu, Holotype HMAS352194 (= Wu17025), ex-type strain CGMCC3.23437 (= 77636).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, reduced to sessile phialides or composed of a 1–3-celled stalk and a terminal phialide, erect, straight or slightly curved, obclavate, 30–43 µm long, 5–6 µm wide at the base, pale brown to medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, (21–) 27–37 µm long, pale brown to medium brown, versicolorous, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, 10–15 µm long and 5–7 µm wide, pale brown; collarettes cylindrical, 16–20 × 2.5–2.8 µm, significantly darker in the lower part, medium brown; ratio of mean lengths of collarette and venter = 1.5:1. Conidia endogenous, extruded in short chains, cylindrical, 11–15 × 2.2–2.7 µm, base truncated with short frills, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width ratio = 5.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, becoming brown, reverse concolorous or slightly darker, with yellow brown to brown pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks. The two strains (77636 and 50643) appear slightly different in pigments in the agar plate.
Other materials examined: China, Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 18 September 2004, Wenping Wu, Wu8183; Hubei Province, Shengnongjia, on dead leaves of unidentified plant, 18 September 2004, Wenping Wu, Wu8283b; Living strains: 50593 (from Wu8183), CGMCC3.23382 (= 50643, from Wu8283b) and 77634 (from Wu17025).
Ecology/substrate/host: Saprobe on decaying needles of pine tree and leaves of undetermined broad leaf tree.
Geographical distribution: China.
Notes: Nagrajchalara cannonii and N. conifericola are the two closely related species in producing reduced conidiophores, versicolorous and lageniform or obclavate conidiogenous cells, conical or subcylindrical venter, and uniseptate, cylindrical conidia. In fact, they can hardly be distinguished in shape and size of conidiophores, phialides and conidia. However, they can be distinguished by the slightly difference in the ITS sequences and morphology of pure culture on PDA.
Some morphological variations were observed among different specimens. For example, in the specimen Wu17025, the conidiophores were 1-septate and shorter (30–35 μm), while in the specimen Wu8183, they were 1–3-septate and longer (40–59 μm). Compared with other specimens, the specimen Wu8283b had slightly longer conidia (15–17 μm).
ITS sequences generated from eight different strains were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77636 includes Calycellina fagina (OL752703, 94% identity) and many unnamed fungi of Leotiomycetes. In addition, these ITS sequence of N. conifericola had 6–7 bp difference from those of N. cannonii.
Nagrajchalara curviphora W.P. Wu & Y.Z. Diao, sp. nov., Fig. 56, MycoBank MB845222.

Nagrajchalara curviphora (Wu15021, holotype). a–j Conidiophores and conidiogenous cells. k–m Conidia. Scale bar: 10 µm for a–j; 5 µm for k–m
Etymology: Refers to is curved conidiophores.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Holotype HMAS 352195 (= Wu15201), ex-type strain CGMCC3.23412 (= NN76101).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, solitary or in small group of 2–3, erect, curved, cylindrical to obclavate, 90–110 µm long, 5–6.5 µm wide at the basal part, 3–5-septate, brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or curved, lageniform, obclavate, 67–88 µm long, medium brown, versicolorous, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical to subcylindrical, 29–34 µm long and 8–9 µm wide, brown; collarettes cylindrical, 37–52 × 3–3.6 µm, brown, lower part darker than the upper part and venter; ratio of mean lengths of collarette and venter = 1.4. Conidia endogenous, extruded in short chains, cylindrical, 13.5–17 × 2.7–3 µm, base truncate with small frills, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width = 5.4:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse soil brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Other living strain: 76102 (from Wu15201).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara curviphora is characterized by 3–5-septate and curved conidiophores, lageniform or obclavate and versicolorous phialides, cylindrical to subcylindrical venter, and 1-septate conidia. Among the existing species of Chalara s. lat., only Chalara urceolata produces the curved conidiophores, but in this species the conidiophores are longer (78–230 μm), and the phialides are concolorous and with wider venters (8–13 μm) (Nag Raj and Kendrick 1975). Nagrajchalara ejneri, a new species described in this study, also has curved conidiophores and phialides, but differs from N. curviphora in longer conidiophores and phialides, narrower venter and collarettes, and smaller conidia.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76101 included Hymenoscyphus serotinus (KU204586, 95% identity) and many unnamed fungi of Leotiomycetes.
Nagrajchalara ejneri W.P. Wu, sp. nov., Fig. 57, MycoBank MB845223.

Nagrajchalara ejneri (a–e, g–m Wu8109c, holotype; f, n Wu12470). a–f, j, k, m, n Conidiophores and conidia. g Basal stroma and basal part of conidiophores in cluster. h, i, l Conidia. Scale bar: 10 µm for a–f, h–j, 5 µm for g–i, k–n
Etymology: Named after the former Vice President Ejner Bech Jensen from Novozymes, who gave strong encouragement to this study.
Typification: China, Hubei Province, Wufengshan, on dead leaves of unidentified tree, 10 September 2004, Wenping Wu, Holotype HMAS 352196 (= Wu8109c), ex-type strain CGMCC3.23376 (= NN48016).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or thick-walled, 2–3 µm wide. Anamorph: Stroma present, solitary, composed of dark brown, irregular-shaped and thick-walled cells. Setae absent. Conidiophores arising from the stroma, 5–7 in clusters, erect, straight or curved, simple, subcylindrical, long obclavate, 87–125 µm high, 4.5–5 µm wide at the lower part, basal cells slightly swollen and lobed up to 10 µm wide, medium to dark brown, smooth, 2–7 septate, terminating in a phialidic conidiogenous cell. Conidiogenous cells integrated, terminal, phialidic, long obclavate, 60–76 μm long, medium brown, concolorous, composed of a venter and a collarette; transition from venter to collarette gradual; venters narrowly lageniform, 17.5–20 × 4.5–5.5 µm; collarettes cylindrical, 47–63 × 2.5–3.2 µm, concolorous; ratio of mean lengths of collarette and venter = 2.9:1. Conidia endogenous, extruded in short or long chains, cylindrical, 10–14.5 × 2.5 µm, straight, thin- and smooth-walled, base truncated and with short basal frills, apex rounded or flattened, hyaline, 1-septate; mean conidium length/width ratio = 4.9:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, becoming brown, reverse concolorous, sterile, up to 6 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Guangdong Province, Shaoguan, Danxiashan, on dead leaves of unidentified tree, 25 December 2012, Wenping Wu, Wu12470. Living strains: CGMCC3.23396 (= 55373, from 12470).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara ejneri is characterized by multiseptate and curved conidiophores, long phialides with a gradual transition from cylindrical venter to long collarette, and 1-septate conidia. Morphologically N. ejneri is similar to N. aurea (conidiophores up to 8-septate and 33–97 × 3.5–7.5 μm, phialides 29–76 μm long, venter 8.5–27 × 3.5–8.5 μm, collarette 14–46 × 2–4 μm, conidia 8–19 × 2–2.5 μm) and Chalara rostrata (conidiophores 110–195 × 4–6 μm, phialides 40–78 μm long, venter 18–28 × 6–9.5 μm, collarette 26–47 × 3–6 μm, conidia 9–13 × 2.5–3 μm), but differs from N. aurea by its longer and curved conidiophores and phialides, from C. rostrata by its shorter and curved conidiophores and longer collarettes (Nag Raj and Kendrick 1975). It also resembles the new species N. curviphora in producing the curved conidiophores, but differs by its longer conidiophores and phialides, and shorter conidia.
The two strains from different localities had the identical ITS sequences. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 48016 included the endophytic fungus from Culluna vulgaris (FM172800, 95% identities), Hymenoscyphus serotinus (KU204586, 93% identities), and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara ellipsoidea W.P. Wu, sp. nov., Fig. 58, MycoBank MB845224.

Nagrajchalara ellipsoidea. (Wu7291b). a–f, k–o Conidiophores with cylindrical collarettes. g–j Conidia. Scale bar: 10 μm for a–e, 5 μm for f–o
Etymology: Refers to its ellipsoidal venter.
Typification: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead leaves of Castanopsis sp., 12 June 2015, Wenping Wu, Holotype HMAS 352197 (= Wu13297a), ex-type strain CGMCC3.23402 (= NN59078).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Stroma present, composed of dark brown, thick-walled cells, scattered. Setae absent. Conidiophores solitary or 3–5 in clusters, arising from the stroma, erect, straight or flexuous, reduced to sessile phialides or composed of a 1–3-celled stalk and a terminal phialidic conidiogenous cell, obclavate, 65–76(–103) µm long, basal cells 6–7 µm wide, brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight, obclavate, lageniform, 60–70 µm long, brown, versicolorous, slightly darker in the lower part of collarettes, smooth; transition from venter to collarette gradual; venters ellipsoidal, subcylindrical, 23–27 µm long, 9–11 µm wide in the widest part, 6.5–7.5 µm wide at the base, brown, smooth; collarettes cylindrical, 45–47 µm long, 4–4.5 µm wide, brown, slightly darker in the lower part, smooth; ratio of mean lengths of collarette and venter = 1.8:1. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 16.5–20 × 3–3.5 µm, base truncated or flattened, without basal frill, apex rounded or flattened, hyaline, smooth, uniseptate; mean conidium length/width ratio = 5.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, becoming brown in the middle part, reverse yellow brown to dark brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Yunnan Province, Baoshan, Tengchong, Gaoligongshan, on dead leaves of unidentified tree, 15 October 2003, Wenping Wu, Wu7291b; Yunnan Province, Jinghong, Xishuangbanna, on dead leaves of unidentified tree, 12 December 2016, Wenping Wu, Wu15140. Living strains: CGMCC3.23370 (= NN 47768, from Wu7291b) and 76004 (from Wu15140).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Morphologically Nagrajchalara ellispoidea resembles N. angionacea and Cylindrocephalum kendrickii, but differs in its versicolorous phialides and unique ITS sequence. In addition, conidia of N. ellispoidea are longer and wider than those in C. kendrickii. Some morphological variations were observed among different specimens, for example, the conidiophores in the specimen Wu15140 were relatively longer (70–103 μm) than those in other specimens.
ITS sequences from different strains were with 4–5 bp differences. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain NN59078 included Chalara sp. (LT629156, 94% identity), Hymenoscyphus serotinus (KU204586, 93% identity) and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara guangcaii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 59, MycoBank MB845225.

Nagrajchalara guangcaii (WuYN06, holotype). a–d, m–q Conidiophores and conidia. e–l Conidia. Scale bar: 10 µm for a–d, 5 µm for e–q
Etymology: Named after the former Chinese mycologist Prof. Guangcai Zhao from Southwest University of Forestry, Kunming.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 Dec. 2018, Y. Zhang, Holotype HMAS 352199 (= WuYN006), ex-type strain CGMCC3.23413 (= NN76112).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary, erect, straight or curved, (1–)3–4-septate, cylindrical, 110–135 µm long, 4–5 µm wide at the base, medium brown, smooth- and thin-walled; basal cell dark brown, irregular, flattened, 6–11 µm wide. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform to obclavate, 94–101 µm long, medium brown to bark brown, concolorous or versicolorous, slightly darker at the lower part of phialides, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual or abrupt; venters cylindrical, subcylindrical, subellipsoidal, 41–54 µm long and 6.5–8.5 µm wide, brown; collarettes cylindrical, 51–54 × 3.7–4.2 µm, basal part slightly darker than the upper part and venter, brown; ratio of mean lengths of collarette and venter = 1.1:1. Conidia endogenous, extruded in short chains, cylindrical, 16.5–21 × 2.5–3 µm, obtuse or flattened at both ends, without basal frill, hyaline, uniseptate, thin- and smooth-walled, guttulate; mean conidium length/width ratio = 7.5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium well–developed, grey, reverse brown in the middle, becoming paler towards the margin, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves of undetermined tree.
Geographical distribution: China.
Notes: Nagrajchalara guangcaii is similar to Chalara aotearoae (phialide 59–69 µm long, venter 22–33 × 8–10 µm, collarette 32–42 × 5–6.5 µm, conidia 13–18 × 3.5–5 µm) and C. tubifera (phialide 49–65 µm long, venter 19–26 × 6–7 µm, collarette 28–40 × 2.5–3 µm, conidia 11–19 × 2–2.5 µm) in possessing septate conidiophores, versicolorous phialides, and 1-septate conidia. Nagrajchalara guangcaii differs from them by longer phialides and different conidial size (Nag Raj and Kendrick 1975). It also resembles N. nawawii, N. puerensis and N. venicola, but differs in size of phialides and conidia. Nagrajchalara guangcaii differs from N. nawawii (65–86 µm long, venter 25–37 × 5.5–6.5 µm, collarette 30–47 × 3.5–4 µm, conidia 14–19 × 3–3.5 µm) in its longer phialides, venters and collarettes, and narrower conidia; differs from N. venicola (89–96 µm long, venter 40–48 × 6.5–7 µm, collarette 48–55 × 3.3–3.6 µm, conidia 15–18.5 × 2.5–2.8 µm) by slightly longer and wider conidia. Furthermore, the ITS sequence of N. guangcaii has 24 bp, 60 bp and 30 bp differences from those in N. nawawii, N. puerensis and N. venicola respectively. In N. puerensis, the collarettes are 2 times longer than venter, while the collarettes and venters are in equal size in N. guangcaii.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76112 included Chalara sp. (LT629156, 94% identity), Hymenoscyphus serotinus (KU204586, 96% identity) and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara haituoshanensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 60, MycoBank MB845226.

Nagrajchalara haituoshanensis (Wu17412, holotype). a–h, i–m Conidiophores and conidia. o–t Conidia. Scale bar: 10 µm for a–h, 5 µm for i–m
Etymology: Refers to the locality, Haituoshan in Chicheng, Hebei Province, China, where the fungus was discovered.
Typification: China, Hebei Province, Zhangjiakou, Chicheng, Haituoshan, on dead fruit of Quercus dentata, 25 September 2020, Wenping Wu, Holotype HMAS352200 (= Wu17412), ex-type strain CGMCC3.23441 (= NN78171).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or in small group of 2–3, erect, straight of slightly curved, obclavate, 58–82 µm long, 4.5–5.5 µm wide at the base, mostly 1-septate, occasionally 2–3-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, lageniform, 57–63 µm long, concolorous or versicolorous, basal part of collarette slightly darker, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, subcylindrical, 20–29 µm long and 5.5–6.4 µm wide; collarettes cylindrical, 30–34 × 3–3.2 µm medium brown, lower part slightly darker than upper part and venter; ratio of mean lengths of collarette and venter = 1.3:1. Conidia endogenous, cylindrical, 10–15 × 2.3–2.5 µm, extruded in short chains, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width ratio = 5.2:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, reverse light soil brown, becoming paler towards the margin, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Other living strains: 78172, 78173 and 78174 (all from the holotype Wu17412).
Ecology/substrate/host: Saprobe on decaying nuts of Quercus dentata.
Geographical distribution: China.
Notes: Nagrajchalara haituoshanensis is characterized by reduced conidiophores, obclavate or lageniform phialides with ellipsoidal to subcylindrical venters, cylindrical collarettes with slightly darker lower part, and 1-septate conidia (10–15 × 2.3–2.5 μm). It is similar to several existing species possessing short conidiophores with one basal stalk cell and 1-septate conidia, and they are Chalara alabamensis, Cylindrocephalum hughesii and C. kendrickii. Nagrajchalara haituoshanensis differs from C. alabamensis (conidia 15–18 × 2–2.5 μm) by shorter conidia, from C. hughesii (conidia 12–17 × 2–2.5 μm) by longer conidiophores and slightly shorter conidia, and from C. kendrickii (conidia 8–13 × 2–4 μm) by longer conidia (Nag Raj and Kendrick 1975).
Identical ITS sequences were generated from the four studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 78171 included Chalara sp. (LT629156, 94% identity), Hymenoscyphus serotinus (KU204586, 96% identity), and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara hangzhouensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 61, MycoBank MB845227.

Nagrajchalara hangzhouensis (Wu16904, holotype). a–h Conidiophores and conidia. i–k Conidia. Scale bar: 5 µm
Etymology: Refers to the locality, Hangzhou, Zhejiang Province, China, where the fungus was originally collected.
Typification: Japan, Mie Prefecture, Tsu, Mie Center of the Arts, on dead leaves, 3 Oct 2019, Wenping Wu, Holotype HMAS 352201 (= Wu16904), ex-type strain CGMCC3.23433 (= NN77457).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, reduced to sessile phialides or composing of one basal supporting cell and a phialide, erect, straight or slightly curved, obclavate, lageniform, 42–54 µm long, 4–5 µm wide at the base, 1-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 41–51 µm long, medium brown, concolorous or versicolorous, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 18.5–26 µm long and 6–7 µm wide, medium brown; collarettes cylindrical, 21–34 × 3.5–4.2 µm, slightly darker and constricted in the lower part, widest in the upper opening region, medium brown; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded in short chains, cylindrical, 11–16 × 3–3.2 μm, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 4.4:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, grey, reverse soil brown to brown, with paler margins, with brown pigment diffused into agar, sterile, up to 5 mm on PDA at 25 °C in 4 weeks.
Other material examined: China Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead leaves of Castanopsis sp., 6 Dec. 2015, Wenping Wu, Wu13295. Living strain: CGMCC3.23401 (= 71055, from Wu13295c).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China and Japan.
Notes: Nagrajchalara hangzhouensis resembles several known species such as Chalara alabamensis, Cylindrocephalum hughesii and C. kendrickii, but differs by size of phialides and conidia. Nagrajchalara hangzhouensis differs from C. alabamensis (conidia 15–18 × 2–2.5 μm) by slightly smaller conidia, from C. hughesii (conidia 12–17 × 2–2.5 μm) by wider conidia, and from C. kendrickii (conidia 8–13 × 2–4 μm) by longer conidia (Nag Raj and Kendrick 1975). Nagrajchalara hangzhouensis is also similar to three new species established in this study, N. follicola, N. tropicalis, N. tubakii in the reduced conidiophores, versicolorous phialide with an ellipsoidal venter and a cylindrical collarette, and 1-septate conidia, but differs by size of phialides and conidia. Same as in N. hangzhouensis, the collarettes in both N. follicola and N. tubakii are with constriction in the lower part and widest in the upper part. However, in N. hanzhouensis, the phialides are shorter (41–51 µm long) than those in N. follicola; and in N. tubakii (phialides 59–67 μm, venters 21–26 × 6.5–7.5 μm, collarettes 37–44 × 3.8–4.5 μm, and conidia 13–16.5 × 3–3.1 μm), the phialides and collarettes are longer than those in N. hangzhouensis. Compared with N. hangzhouensis, N. tropicalis (phialides 60–74 μm long, venters 30–34 × 7–8 μm, collarettes 35–40 × 3.7–4.5 μm, and conidia 14.5–16 × 3.6–3.8 μm) produces longer phialides and venter, and wider conidia; and the collarettes in the latter species are not constricted in the lower part.
ITS sequences generated from two different strains (China and Japan) were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77457 included Chalara sp. (LT629156, 94% identity), Hymenoscyphus serotinus (KU204586, 96% identity) and many unnamed endophytic or saprophytic fungi from different environmental samples.
Nagrajchalara inflatipes (Preuss.) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 62, MycoBank MB845276.

Nagrajchalara inflatipes (Wu12532). a–d Conidiophores and conidiogenous cells. e–n Conidia. Scale bar: 10 µm for m, 5 µm for a–l, n
≡ Cylindrosporium inflatipes Preuss., Linnaea 24: 106, 1851.
≡ Chalara inflatipes (Preuss) Sacc., Syll. Fung. (Abellini) 4: 385, 1886.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of brown to dark, septate and branched hyphae with smooth and thick wall, 2–4 µm wide. Anamorph: Stroma absent or poorly developed, composed of several dark brown and irregular cells. Setae absent. Conidiophores arise from superficial hyphae or aggreged cells, solitary or rarely 2–3 in small group, erect, straight or slightly curved, composing of a 2–4-celled basal stalk and a terminal phialide, obclavate, lageniform, 120–160 µm high, 7–10 µm wide, brown to dark brown, versicolorous, thick-walled, verruculose, 2–6 septate, with 1–4 percurrent proliferations at the base. Conidiogenous cells integrated, terminal, erect, straight, lageniform, obclavate, 50–120 µm long, brown, versicolorous, verruculose, consisting of a venter and a collarette; transition from venter to collarette gradual; venters cylindrical to subellipsoidal, 30–50 µm long and 11–12 µm wide, medium brown, verruculose, thin-walled; collarettes cylindrical, 45–75 µm long, 5–7 µm wide, uniformly dark brown, significantly darker than venter, verruculose, apex irregularly ruptured; ratio of mean lengths of collartte and venter = 1.5:1. Conidia endogenous, extruded in long chains, cylindrical, (21–)28–34 × 3.8–4.3 µm, apex rounded, base truncated or flattened with short frills, hyaline, 3-septate, smooth; mean conidium length/width ratio = 7.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to light brown, reverse soil brown to brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 4 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Guangdong Province, Shaoguan, Danxiashan, on dead material of unidentified palm species, 25 Dec. 2012, Wenping Wu, Wu12532. Living strain: CGMCC3.23453 (= 55517, from Wu12532).
Ecology/substrate/host: Saprobe on dead branches and rotten wood of Abies alba, Alnus sp., Picea abies and Sorbus aucuparia.
Geographical distribution: China, Czechoslovakia, Germany, North America and USSR (Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975) and Holubová-Jechová (1984).
Notes: Nagrajchalara inflatipes is characterized by the versicolorous and verruculose conidiogenous cells with abrupt transition from venters to collarettes, darker collarettes, and 3-septate conidia with rounded apex and truncate base (Nag Raj and Kendrick 1971, 1975). Apart from the slightly shorter conidiophores and conidiogenous cells, the fungus from the Chinese specimen agreed well with the description and illustration provided by Nag Raj and Kendrick (1975). Morphologically this species resembles N. angustata in producing versicolorous phialides and 3-septate conidia, but differs by the smooth collarette and wider conidia (22–42 × 4.8–6 µm) in the latter species (Kowalski and Halmschlager 1996). Nagrajchalara inflatipes also bears some similarity with N. unicolor, however, in the latter species the conidiophores are concolorous and composed of a single stalk cell and a phialides with gradual transition from venters to collarettes (Nag Raj and Kendrick 1975).
The ITS sequence of this species was generated for the first time in this study. In the phylogenetic analyses, N. inflatipes clustered together with two other species (N. angustata and N. insignis) with 3- or 7-septate conidia. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 55517 included Chalara angustata (NR_159786, 90% identity), Mollisina uncinata (JN033457, 89% identity) and many unnamed endophytic fungi or fungi from environmental samples.
Nagrajchalara insignis (Sacc., M. Rousseau & E. Bommer) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845277.
≡ Sporoschisma insigne Sacc., Rousseau & E. Bommer, Arch. Mus. Hist. Nat. Paris, ser. 6 6: 455, 1884.
≡ Chalara insignis (Sacc., Rousseau & E. Bommer) S. Hughes, Can. J. Bot. 31: 622, 1953.
Description on the natural substrate: Anamorph: Conidiophores cylindrical, 70–220 μm long, 8–13 μm wide at the base, multiseptate, with percurrent proliferation. Conidiogenous cells phialidic subcylindrical to lageniform, 66–145 μm long, transition from venter to collarette abrupt; venter subcylindrical, 38–60 × 10–12 μm, light brown; collarette cylindrical, 58–87 × 5.6–7 μm, dark brown. Conidia cylindrical, 18–54 × 5–6 μm, hyaline, 7-septate, apex obtuse, base truncate and with a marginal frill (Nag Raj and Kendrick 1975). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on Betula pendula, Corylus avellana and unidentified dead wood.
Geographical distribution: Canada, Czech Republic, Italy, Poland, UK and USA (Hughes 1958; Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Koukol et al. 2020).
Description and illustration: Hughes (1958), Nag Raj and Kendrick (1975), Holubová-Jechová (1984), and Koukol et al. (2020).
Notes: Nagrajchalara insignis resembles Chalara bicolor (conidiophores up to 170 μm long, phialides 132–165 μm long, venters 50–55 × 14.4–16.2 μm, collarettes 7–10 μm wide, conidia 7-septate, 50–66 × 5.9–6.6 μm), N. angustata (conidiophores 110–160 μm long, phialides 96–125 μm long, venters 38–60 × 10–12 μm, collarettes 58–87 × 5.6–7 μm, conidia 3-septate 22–42 × 4.8–6 μm), and N. inflatipes (conidiophores 140–250 μm long, phialides 105–145 μm long, venters 27–55 × 8.5–15 μm, collarettes 83–97 × 5.5–8.5 μm, conidia 3-septate 22–37 × 4–5 μm) in morphology of conidiophores and multi-septate conidia. Nagrajchalara insignis can be easily distinguished from N. angustata and N. inflatipes by its 7-septate conidia. Chalara bicolor, also with 7-septate conidia, is very close to N. insignis, but can be distinguished by its wider venters and longer conidia (Nag Raj and Kendrick 1976).
Nagrajchalara intermedia (W.P. Wu) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 63, MycoBank MB845278.

Chalara intermedia (Wu1006c, holotype). a Conidiophores and conidiogenous cells. b Conidia. Scale bar: 20 μm
≡ Chalara intermedia W.P. Wu, Mycosystema 23: 316, 2004.
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly superficial and partly immersed, composed of pale brown to medium brown, septate and branched hyphae with thin- and smooth-wall or slightly verruculose, 2–3.5 µm diam. Anamorph: Stroma absent. Setae absent. Conidiophores scattered, erect, straight, unbranched, cylindrical, 25–40 × 3–4 µm, very pale colored, smooth to slightly verrucose, 1–3-septate, terminating in a phialide. Conidiogenous cells integrated, terminal, phialidic, subcylindrical, lageniform to ampulliform, 15–22 µm long, base 3–3.5 µm wide, very pale colored, smooth- or slightly rough-walled, collarette 1.8–2.2 µm wide. Conidia endogenous, extruded in longer and loose chains, cylindrical, 9–10 × 1.2–1.5 µm, apex obtuse to rounded, base truncate, hyaline, 1-septate, smooth- and thin-walled. Teleomorph: Unknown.
Material examined: China, Yunnan Province, Kunming, Kunming Botanical Garden, on dead leaves of Cinnamomum sp., 24 November 1995, Wu Wenping (Wu1006c, holotype).
Ecology/substrate/host: Saprobe on dead leaves of Cinnamomum sp.
Geographical distribution: China (Wu 2004).
Description and illustration: Wu (2004).
Notes: None of existing Chalara s. lat. species with uniseptate conidia and simple conidiophores were recorded with hyaline to very pale-colored phialophores (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Although several species such as Chalara alabamensis, C. australis, C. gracilis, C. scabrida, C. transkeiensis N. acuaria, N. agathidis, N. angionacea, Cylindrocephalum aurea, C. kendrickii, were described with simple conidiophores and 1-septate conidia, all these species had the dark brown conidiogenous cells with well differentiated venter and collarette (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; Morgan-Jones et al. 1992; McKenzie 1993; Wu 2004). No living strain was available for molecular phylogeny, its phylogenetic relationship with other chalara-like fungi remains to be studied in future.
Nagrajchalara japonica W.P. Wu & Y.Z. Diao, sp. nov., Fig. 64, MycoBank MB845228.

Nagrajchalara japonica (Wu16921, holotype) a–h, j–n Conidiophores and conidiogenous cells. i, o–q Conidia. Scale bar: 10 µm for a–h, 5 µm for i–q
Etymology: Refers to the country from which this fungus was originally collected.
Typification: Japan, Mie Prefecture, Tsu, Mie Center of the Arts, on dead leaves of unidentified tree, 3 October 2019, Wenping Wu, Holotype HMAS 352204 (= Wu16921), ex-type strain CGMCC3.23436 (= NN77628).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma poorly developed, composed of several dark brown and irregular cells. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, reduced to a 1–2-celled basal stalk and a phialide, obclavate, lageniform, 49–60(–70) µm long, 4–5 µm wide at the base, 1–2-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 46–58 µm long, versicolorous, medium brown, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, subcylindrical, 20–23 µm long and 5.5–7.5 µm wide, medium brown; collarettes cylindrical, 27–39 × 3–3.5 µm, slightly constricted in the lower part, widest at the upper part, versicolorous, slightly darker in the lower part, medium brown; ratio of mean lengths of collarette and venter = 1.5:1. Conidia endogenous, extruded in short chains, cylindrical, 10–13 × 2.8–3.0 µm, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width ratio = 4:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white then pale brown, reverse yellow brown to soil brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Other materials examined: Japan, Mie Prefecture, Tsu, Mie Center of the Arts, on dead leaves of unidentified tree, 3 October 2019, Wenping Wu, Wu16923. Living strains: 77629 (from Wu16921a) and 77460 (from Wu16923b).
Ecology/substrate/host: Saprobe on dead leaves of undetermined tree.
Geographical distribution: Japan.
Notes: Nagrajchalara japonica morphologically resembles several new species described in this study, such as N. follicola, N. hangzhouensis, N. tropicalis and N. tubakii in the reduced conidiophores, versicolorous phialides, ellipsoidal venters and 1-septate conidia, but differs from them by size of phialides and conidia. Same as in N. japonica, the collarettes in N. follicola, N. hangzhouensis and N. tubakii are characterized with constriction in the lower part and widest in the upper part. Nagrajchalara japonica differs from all these three species by its smaller conidia. In addition, the phialides in N. follicola, N. tropicalis and N. tubakii are longer than those in N. japonica. Compared with N. japonica, N. tropicalis (phialides 60–74 μm long, venters 30–34 × 7–8 μm, collarettes 35–40 × 3.7–4.5 μm, and conidia 14.5–16 × 3.6–3.8 μm) produces longer phialides and wider conidia; and the collarettes in the latter species are not constricted in the lower part.
ITS sequences generated from the three studied strains were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77629 included Chalara sp. (LT629156, 92% identity) Hymenoscyphus serotinus (KU204586, 96% identity) and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara jonesii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 65, 66, 67, MycoBank MB845229.

Nagrajchalara jonesii (f, g from Wu16063, a–e, h from Wu16158). a–e Setae with sterile or fertile apex bearing a phialide. f, g Conidiophores and conidiogenous cells. h Sterile setae with a terminal phialide. Scale bar: 10 µm for a–d, 5 µm for e–h

Nagrajchalara jonesii (Wu16158, holotype). a–h Conidiophores and conidiogenous cells. i–m Conidia. Scale bar: 5 µm

Nagrajchalara jonesii (Wu16063). a–c Setae and conidiophores. d–j Conidiophores and conidiogenous cells. k–t Conidia. Scale bar: 10 µm for a–e, 5 µm for f–t
Etymology: Named after the British mycologist E.B.G. Garth Jones.
Typification: China, Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Holotype HMAS 352205 (= Wu16158), ex-type strain CGMCC3.23460 (= NN76551).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae solitary or aggregated at the base, erect, straight or slightly flexuous, cylindrical, 210–260 × 4.5–5 μm, dark brown to black, versicolorous, 9–12-septate, gradually tapering towards the apex; basal cell swollen and up to 10 μm wide; apex acute or obtuse, sterile or occasionally developing into a phialide. Conidiophores formed from superficial hyphae, obclavate, cylindrical, (48–)60–122 µm long, 4–5.5 μm wide at the base, 1–3-septate, medium brown, smooth, occasionally with percurrent proliferation. Conidiogenous cells integrated, terminal, phialidic, lageniform, obclavate, 47–54 µm long, concolorous, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, 19–21 µm long and 4.5–6 µm wide, brown; collarettes cylindrical, 32–35 µm long, 3–3.5 μm wide, medium brown; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, extruded in short chains, cylindrical, (13–)16–18 × 2–2.3 µm, base truncated, apex obtuse, hyaline, uniseptate; mean conidium length/width ratio = 8:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse concolorous, sterile, very slow growing and up to 3 mm on PDA at 25 °C in 4 weeks.
Other material examined: China: Zhejiang Province, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, HMAS 352205 (= Wu16063). Living strains: CGMCC3.23456 (= 76451, from Wu16063) and 76550 (from Wu16158).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Morphologically Nagrajchalara jonesii is similar to N. aspera, N. mutabilis and N. septata, but differs in the unique combination of morphological characters and ITS sequences. Nagrajchalara jonesii differs from N. aspera by its smooth-walled collarettes, from N. mutabilis and N. septata by longer conidiophores and collarettes (Nag Raj and Kendrick 1975; Silva et al 2015).
The two examined specimens were collected from the same locality and with identical morphology, and ITS and LSU sequences. The ITS sequence had relatively lower identity with those in N. mutabilis and N. septata. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76451 included Hyaloscypha gabretae (MZ520783, 90% identity), H. melinii (EF093175, 90% identity) and many unnamed endophytic fungi or saprophytic fungi.
Nagrajchalara keqinii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 68, MycoBank MB845230.

Nagrajchalara keqinii (Wu15166, holotype). a–j Conidiophores and conidiogenous cells. k–p Conidia. Scale bar: 10 µm for a–c, 5 µm for d–p
Etymology: Named after the Chinese mycologist Prof. Zhang Keqin from the Yunnan University, Kunming.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Holotype HMAS 352206 (= Wu15166), ex-type strain CGMCC3.23408 (= NN76020).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, solitary or occasionally aggregated, erect, straight or slightly curved, reduced to a 1–2-celled basal stalk and a terminal phialide, lageniform, obclavate, 50–62 µm long, 4–5 µm wide at the base 1–2-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 47–57 µm long, versicolorous, medium brown to brown, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt or gradual; venters ellipsoidal, 16–24 µm long and 5–6 µm wide, brown; collarettes cylindrical, 28–32 µm long, 3–3.5 μm wide, medium brown; ratio of mean lengths of collarette and venter = 1.5:1. Conidia endogenous, extruded in short chains, cylindrical, 13–15 × 2.2–2.5 µm, truncate at both ends, without frills, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to grey, then grey brown, reverse brown, becoming paler towards the margin, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 Dec. 2018, Y. Zhang, Wu15177 and YN016. Living strain: 76021 (from Wu15177), 76066 (from YN16) and 76077 (from 15177a).
Ecology/substrate/host: Saprobe on dead leaves of undetermined tree.
Geographical distribution: China.
Notes: Nagrajchalara keqinii is similar to N. agathidis and N. angionacea in the reduced conidiophores, obclavate or lageniform phialides, ellipsoidal venter and 1-septate conidia, but differs in size of phialides and conidia. In N. agathidis (conidia 17–24 × 2.5–3 μm) and N. angionacea (13–18 × 3–3.5 μm), the conidia are longer and wider. In addition, the phialides (53–64 μm) of N. angionacea are with longer collarettes (35–43 μm) than those in N. keqinii (Nag Raj and Kendrick 1975).
The ITS sequences from the three studied strains are identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76020 include Hymenoscyphus serotinus (KU204586, 94% identity) and many unnamed endophytic fungi or saprophytic fungi.
Nagrajchalara knudsonii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 69, 70, MycoBank MB845231.

Nagrajchalara knudsonii (a–g, l–o from Wu15185, holotype; h–k from Wu15180). a–g, l–o Conidiophores and conidiogenous cells. h–k Conidia. Scale bar: 10 µm for a–g, 5 µm for h–o

Nagrajchalara knudsonii (Wu15185, holotype). a–f Conidiophores and conidiogenous cells. g–l Conidia. Scale bar: 5 µm
Etymology: Named after the Danish mycologist Henning Knudson.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Wenping Wu, Holotype HMAS 352207 (= Wu15185), ex-type strain CGMCC3.23409 (= NN76053).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent or poorly developed, composed of a few aggregated irregular cells. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, solitary or in small group, erect, straight or slightly curved, reduced to a 1–2-celled stalk and a terminal phialide, obclavate, lageniform, 79–100 µm long, 4–5 µm wide at the lower part, base, 1–2-septate, medium brown, smooth. Conidiogenous cell integrated, terminate, phialidic, erect, straight or slightly curved, obclavate, lageniform, 75–86 µm long, medium brown, versicolorous, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, subcylindrical, 27–30 × 6–7.5 µm, brown; collarettes cylindrical, 52–55 × 3.3–3.6 µm, brown, lower part slightly darker than the upper part of collarettes and venter; ratio of mean lengths of collarette and venter = 1.9:1. Conidia endogenous, hyaline, uniseptate, cylindrical, base truncate without frill, apex rounded or flattened, 16.5–18.5 × 2.2–2.5 µm, extruded in short chains; mean conidium length/width ratio = 7.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, reverse concolorous, becoming paler towards the margin, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Other material examined: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Wenping Wu, Wu15180. Living strains: 76023 (from 15180).
Ecology/substrate/host: Saprobe on dead leaves of undetermined tree.
Geographical distribution: China.
Notes: Nagrajchalara knudsonii resembles N. agathidis in possessing reduced conidiophores, obclavate or lageniform phialides with ellipsoidal venter and cylindrical collarettes, and 1-septate conidia, but differs by shorter conidia (Nag Raj and Kendrick 1975).
Identical ITS sequences were generated from the two studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 76023 included Chalara sp. (LT629156, 94% identity), Chalara sp. (LT629155, 94% identity), Hymenoscyphus serotinus (KU204568, 96% identity) and many unnamed fungi of Leotiomycetes. In addition, the ITS sequences of these two species have 14 bp differences.
Nagrajchalara morganjonesii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 74d–g, MycoBank MB845232.
Etymology: Named after the mycologist G. Morgan-Jones who studied the phialidic hyphomycetes.
Typification: China, Guangxi Province, Shangsi, Shiwandashan, on dead leaves of Quercus sp., 31 December. 1997, Wenping Wu, Holotype HMAS 352186 (= Wu1609h), ex-type strain CGMCC3.23446 (= NN44060).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, smooth-walled, septate and branched hyphae of 2.5–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, solitary or aggregated, reduced to one incorporated basal cell and a phialide, erect, straight or slightly curved, obclavate, lageniform, pale to medium brown, smooth, basal cell up to 6 µm long and 5 µm wide. Conidiogenous cells integrated or discrete, terminate, erect, straight or slightly curved, obclavate, lageniform, 38–57 µm long, concolorous, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual; venter subellipsoidal to subcylindrical, 25–35 × 6.5–7 µm; collarette cylindrical, 28–30 × 3.5–4 µm; ratio of mean lengths of collarette and venter = 1:1. Conidia endogenous, extruded in short chains, cylindrical, 12–18 × 2.5–3 µm, base truncated or flattened and no frill, apex flattened or rounded, hyaline, 1-septate; mean conidium length/width ratio = 5.5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, grey, reverse yellow brown, with brown pigment diffused into agar, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Guangdong Province, Dinghushan, on dead leaves of unidentified plant, 10 October 1998, Wenping Wu, Wu1962a; Guangxi Province, Nanning, on dead leaves of Quercus sp., 3 January 1998, Wenping Wu, HMAS 352208 (= Wu1613d) and Wu1657c; Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead leaves of Quercus sp., 2 January 1998, Wenping Wu, Wu1229d; Yunnan Province, Xishuangbanna, 16 October 1999, Wenping Wu & Yan Huang, Wu2888a. Living strains: 43682 (from Wu1657c) and 44402 (from Wu1613d).
Ecology/substrate/host: Saprobe on dead leaves of Quercus nigra, Quercus spp. and other plants.
Geographical distribution: China and North American.
Notes: Nagrajchalara morganjonesii resembles Chalara alabamensis, Cylindrocephalum hughesii, C. kendrickii, N. agathidis and N. angionacea, but differs from them by shape and size of phialides and conidia (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; McKenzie et al. 2002). In C. alabamensis, N. agathidis and N. angionacea, the phialides are longer and also with long collarettes than those in N. morganjonesii. Cylindrocephalum hughesii differs from N. morganjonesii by its lageniform phialides with abrupt transition from venter to collarettes. The conidia of C. kendrickii (8–13 × 2–4 μm) are smaller than those in N. morganjonesii.
Identical ITS sequences were generated from three studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 44060 included Hymenoscyphus serotinus (KU204568, 96% identity) and many unnamed fungi of Leotiomycetes.
Nagrajchalara mutabilis (C.R. Silva, S.S. Silva, Gusmão & R.F. Castañeda) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 71, MycoBank MB845279.

Nagrajchalara mutabilis (Wu8330, holotype). a–d Setae with sterile or fertile apex. e–o Conidiophores and phialidic conidiogenous cells. s–u Conidia. Scale bar: 10 µm for a–m, 5 µm for n–u
≡ Chaetochalara mutabilis C.R. Silva, S.S. Silva, Gusmão & R.F. Castañeda, Mycotaxon 130(2): 506, 2015.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3.5 µm wide. Anamorph: Stroma present, composed of dark brown, thick- and smooth-walled cells. Setae arising from basal stroma or superficial hyphae, solitary or aggregated at the base, associated with conidiophores, simple, erect, straight or flexuous, cylindrical, 200–250 × 5–6.5 µm, upper part 2.5–3 µm wide, tapering towards the acute apex, multiseptate, dark brown to blackish brown, becoming paler towards the apex, smooth, slightly inflated at the base, sterile or fertile and terminating into a phialide. Conidiophores solitary or associated with setae at the base, erect, straight or slightly curved, lageniform, obclavate, 45–57.5 µm long, composing of a 2–3-celled stalk and a terminal phialide, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 32.5–50 µm long, concolorous, pale to medium brown, smooth, consisting of a venter and a collarette, occasionally with 1–2 percurrent proliferations; transition from venter to collarette abrupt; venters cylindrical, 12.5–17.5 µm long and 4.5–5 µm wide; collarettes cylindrical, 17.5–25 µm long, 2–3 µm wide, concolorous, smooth; ratio of mean lengths of collarette and venter = 1.4:1. Conidia endogenous, extruded in short chains, cylindrical, 12.5–17.5 × 2–2.5 µm, both ends rounded or flattened, hyaline, 1-septate; mean conidium length/width ratio = 6.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, then brown, reverse pale brown to brown, becoming paler towards the margin, sterile, up to 9 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Hubei Province, Shengnongjia, on dead leaves of unidentified plant, 16 September 2004, Wenping Wu, HMAS 352202 (= Wu8330). Living strain: CGMCC3.23387 (= 50725, from Wu8330).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: Brazil and China (Silva et al. 2015).
Description and illustration: Silva et al. (2015).
Notes: Chaetochalara mutabilis was originally described from decaying needle-like leaves of Araucaria angustifolia in Brazil (Silva et al. 2015). Compared with the original description (setae 5–14-septate and 150–345 μm long, conidiophores 1–3-septate and 40–82 μm long, phialides 24–57 μm long, venters 12–25 × 4–6 μm, collarettes 12–33 × 3 μm, and conidia 10–15 × 2–2.5 μm), the Chinese specimen has shorter setae and slightly longer conidia.
ITS sequence of this species was generated for the first time in this study. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77457 included Hymenoscyphus serotinus (KU204586, 96% identity), Chalara sp. (LT629156, 93% identity), Calycellina fagina (OL752703, 92% identity) and many unnamed endophytic fungi or saprophytic fungi from different environmental samples.
Nagrajchalara nawawii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 72, MycoBank MB845233.

Nagrajchalara nawawii (Wu15167, holotype). a–f, j–l Conidiophores and phialidic conidiogenous cells. i, m Conidia. Scale bar: 10 µm for a–f; 5 µm for i–m
Etymology: Named after the mycologist Dr. A. Nawawi.
Typification: China, Yunnan Province, Jinghong, Xishuangbanna, on dead leaves of unidentified tree, 6 December 2018, Wenping Wu, Holotype HMAS 352208 (= Wu15167), ex-type strain NN76006.
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin– or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent or poorly developed, composed of a few aggregated cells. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, scattered or 2–3 aggregated at the base, erect, straight or curved, reduced to a 1–2-celled basal stalk and a terminal phialide, simple, obclavate, lageniform, rarely subcylindrical, 82–90(–115) µm long, 5–6 µm wide, brown, becoming medium brown towards the apex, 1–3 septate, smooth, basal cells flattened and dark brown. Conidiogenous cells integrated or discrete, terminal, obclavate, subcylindrical, 65–86 µm longer, versicolorous, medium brown, slightly darker in the lower part of collarettes, become pale brown towards the apex, composed of a venter and a collarette; transition from venter to collarette abrupt; venters subcylindrical to subellipsoidal, 25–37 × 5.5–6.5 µm, smooth; collarettes cylindrical, 30–47 × 3.5–4 µm, smooth; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded in short chains, cylindrical, 14–19 × 3–3.5 µm, straight, base truncated or flattened and no basal frill, apex obtuse or flattened, smooth, hyaline, 1-septate, guttulate; mean conidium length/width ratio = 5.1:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, grey to brown, reverse dark brown, with yellow pigment diffused into agar, sterile, up to 6 mm on PDA at 25 °C in 4 weeks.
Other material examined: China, Yunnan Province, Baoshan, Lujiang, Bawan, Gaoligongshan, on dead branches, 15 October 2003, Wenping Wu, Wu7299a. Living strains: CGMCC3.23373 (= 47788, from Wu7299a) and 76049 (from Wu15167b).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara nawawii is similar to N. agathidis and N. knudsonii in the reduced conidiophores with 1–2 stalk cells and a phialide, versicolorous phialides, and 1-septate conidia, but differs by slightly shorter but significantly wider conidia (Nag Raj and Kendrick 1975). It is also similar to N. neonawawii and N. tropicalis, two new species described in this study, but differs in size of conidia. Nagrajchalara neonawawii has smaller conidia and slowly growing colony on PDA, N. tropicalis (14.5–16 × 3.6–3.8 µm) has slightly shorter but significantly wider conidia than those in N. nawawii.
Identical ITS sequences were generated from three studied strains. They have 9 bp and 17 bp differences from those of N. neonawawii and N. tropicalis respectively.
Nagrajchalara neonawawii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 73, MycoBank MB845234.

Nagrajchalara neonawawii (Wu15143, holotype). a–g, k–m Conidiophores and phialidic conidiogenous cells. h–j Conidia. Scale bar: 10 µm for a–g; 5 µm for h–m
Etymology: Refers to its similarity to Nagrajchalara nawawii.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Holotype HMAS 352209 (= Wu15143), ex-type strain CGMCC3.23403 (= NN75983).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or aggregated, erect, straight or slightly curved, reduced to a 1–2-celled stalk and a phialide, obclavate, lageniform, 82–110 µm long, 4–5 µm wide at the base, 1–2-septate, concolorous, brown to dark brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 73–92 µm long, concolorous, brown to dark brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 25–33 µm long and 7–8 µm wide; collarettes cylindrical, 50–56 × 4–4.5 µm; ratio of mean lengths of collarette and venter = 1.8:1. Conidia endogenous, extruded in short chains, cylindrical, 11–15 × 3.5–3.6 µm, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled, guttulate; mean conidium length/width ratio = 3.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse concolorous, sterile, up to 16 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead frond of fern, 26 August 2022, Wenping Wu, Wu18060 and Wu18061. Living strains: 78610 and 78611 (from Wu18060), and 78612 (from Wu18061).
Ecology/substrate/host: Saprobe on dead leaves of Smilax sp. and unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara neonawawii morphologically resembles N. nawawii in shape of conidiophores, phialides and conidia, but differs in slightly shorter conidia. These two species also share the very similar ITS sequences which are with only 9 bp differences. On PDA, the strain of N. neonawawii grows much faster and with white colony, while the strain of N. nawawii grows slowly and with dark brown colony. Two additional specimens (Wu18060 and Wu18061) were recently collected on dead stem of fern from Hangzhou, and four living strains were successfully obtained from them by using single spore isolation method. The obtained ITS sequences from these strains were identical to N. neonawawii from the ex-type strain. However, the fungus from these two specimens were with longer conidiogenous cells (83–110(–137) µm long), broader venters (32–36 × 8–9 µm), longer collarette 68–77 × 3.5–4 µm) and conidia (20–23 × 3–3.5 µm).
Nagrajchalara novozymia W.P. Wu & Y.Z. Diao, sp. nov., Fig. 74a–c, MycoBank MB845235.

Nagrajchalara morganjonesii (d, e Wu1613d, holotype; f, g Wu2888a) and N. novozymia (a–c Wu1360a, holotype). a, d, f Conidia. b, c, e, g Conidiophores and conidiogenous cells. Scale bar: 40 for b, 20 μm for a, c, d–g
Etymology: Refers to the company Novozymes A/S where the authors conducted this research.
Typification: China, Guangxi Province, Damingshan, on dead branches of unidentified plant, 18 December 1997, Wenping Wu, Holotype HMAS 352247 (= Wu1360a).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin– or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores macronematous, scattered or aggregated at the base, erect, straight or flexuous, simple, cylindrical or subcylindrical, 130–250 µm long, 9–11 µm wide, medium to dark brown, 5–10-septate, smooth, terminating in a conidiogenous cells. Conidiogenous cells integrated, terminal, phialidic, obclavate, lageniform, 75–85 µm long, concolorous, medium brown to brown, become very pale brown at apex, smooth, composed of a venter and a collarette; transition from venter to collarette gradual or abrupt; venter ellipsoidal to subcylindrical, 30–41 µm long, 13–14 µm wide at widest; collarette cylindrical, 40–45 µm long, 8–8.5 µm at apex, concolorous with venter; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded easily in short chains, cylindrical, 29–34 × 7–8.5 µm, straight, apex rounded and with thick-walled, base truncated with distinct frills, hyaline, uniseptate, wall thick and smooth; mean conidium length/width ratio = 4:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead branches of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara novozymia is characterized by its long and multiseptate conidiophores, fairly differentiated conidiogenous cells, and uniseptate, cylindrical and larger conidia with rounded and basal frills (Nag Raj and Kendrick 1975; McKenzie et al. 2002).
Nagrajchalara ohmanii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 75, MycoBank MB845236.

Nagrajchalara ohmanii (Wu1289d, holotype). a–d Conidiophores and phialidic conidiogenous cells. e–l Conidia. Scale bar: 5 µm
Etymology: Named after Mr. Anders Ohmans, the former director in Novozymes, who gave strong support to this work.
Typification: China, Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead leaves of Cinnamomum sp., 2 January 1998, Wenping Wu, Holotype HMAS 352210 (= Wu1289d), ex-type strain CGMCC3.23359 (= NN43147).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Stroma absent. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, reduced to one basal cell and a phialide, solitary, erect, straight or slightly curved, obclavate, lageniform, 80–95 µm long, medium brown, smooth- and thin-walled. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, lageniform, obclavate, 75–85 µm long, medium brown, concolorous or slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters lageniform, 23–35 µm long and 7.5–8.5 µm wide; collarettes cylindrical, 48–60 × 4–5 µm, concolorous or slightly darker in the lower part, brown; ratio of mean lengths of collarette and venter = 1.9:1. Conidia endogenous, extruded in short chains, cylindrical, 14–18 × 2.3–2.5 µm, base truncate without frill, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width ratio = 6.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, brown to dark brown, reverse concolorous, with brown pigment diffused into agar, sterile, up to 8 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara ohmanii resembles N. agathidis, N. angionacea, N. nawawii and N. tropicalis in producing the reduced conidiophores, obclavate or lageniform and versicolorous phialides with ellipsoidal venter, and uniseptate and cylindrical conidia, but differs by shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Compared to N. ohmanii, conidia of N. agathidis are longer and narrower, and conidia of N. angionacea, N. nawawii and N. tropicalis are significantly wider (> 3 µm).
Nagrajchalara panamensis (Koukol, T.A. Hofm. & M. Piepenbr.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845280.
≡ Chalara panamensis Koukol, T.A. Hofm. & M. Piepenbr., in Crous et al., Persoonia 37: 267, 2016.
Description on the natural substrate: Anamorph: Conidiophores reduced to a single cell bearing a phialide. Conidiogenous cells phialidic, lageniform, transition from venter to collarette gradual, 85–93 μm long and 5–6.5 μm wide; venter cylindrical, 24–28.5 μm long; collarette with distinct small warts covering the wall (at least upper two thirds), 59–69 μm long and 3.5–4 μm wide; ratio of the mean lengths of collarette and venter 2.:1. Conidia cylindrical with both ends rounded, 1-septate, hyaline, smooth-walled, (11–)14.5–20.5 μm long and 2.5–3 μm wide (Crous et al. 2016a, b). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on needle litter of Pinus cf. caribaea.
Geographical distribution: Panama (Crous et al. 2016a, b).
Illustration and description: Crous et al. (2016a, b).
Notes: Nagrajchalara panamensis differs from other chalara-like species by having longer phialides with asperate collarettes. It is morphologically similar to Nagrajchalara aspera (phialides 48–77 μm, collarette 30–47 μm, conidia 10–27 μm), but differs in lack setae and with longer collarettes (Nag Raj and Kendrick 1975). Chalara verruculosa also produces verrucose collarette, but its conidia are 1-celled (McKenzie et al. 2002). The phylogenetic analyses in this study showed that C. panamensis was closely related with other Nagrajchalara species with septate conidia.
Nagrajchalara paraunicolor W.P. Wu & Y.Z. Diao, sp. nov., Fig. 76, MycoBank MB845237.

Nagrajchalara paraunicolor (Wu8194b, holotype). a Superficial mycelium. b–g, n, o Conidiophores and conidiogenous cells. h–m Conidia. Scale bar: 10 µm for a–g, 5 µm for h–o
Etymology: Refers to its morphological similarity with Nagrajchalara unicolor in 3-septate conidia.
Typification: China, Hubei Province, Shengnongjia, on dead culms of bamboo, 16 September 2004, Wenping Wu, Holotype HMAS 352211 (= Wu8194b), ex-type strain CGMCC3.23384 (= NN50666).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 3–4 µm wide. Anamorph: Stroma well-developed, composed of dark brown, thick- and smooth-walled, irregular cells of 3–6 µm diam. Setae absent. Conidiophores arising from surface cells of basal stroma, 3–6 aggregated in cluster, erect, straight or slightly flexuous, reduced to a single cell bearing a phialide, obclavate, lageniform, fusiform, 75–125 µm long, basal cell 5–10 × 6–9 µm, concolorous, medium brown to brown. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, lageniform, fusiform, 75–90 µm long, concolorous, medium brown to brown, asperate in the middle part and smooth at lower and upper part, composed of a venter and a collarette; transition from venter to collarette gradual; venters ellipsoidal, 25–45 µm long, 10–12 µm wide in the widest part; collarettes cylindrical, 40–60 µm long, 4.5–6 µm wide, concolorous with venter, becoming paler towards the apex, apex irregularly ruptured with untruncated and rounded apical apparatus; ratio of mean lengths of collarette and venster = 1.4:1. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 22.5–30 × 4–5 µm, apex blunt or rounded, base truncated or slightly rounded with distinct marginal frills, hyaline, 3-septate; mean conidium length/width = 5.8:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, brown to dark brown, reverse dark brown, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead culm of bamboo.
Geographical distribution: China.
Notes: Nagrajchalara paraunicolor bears some similarity with N. unicolor in having reduced conidiophores and 3-septate conidia, but differs in shorter conidiophores and phialides. In addition, in N. unicolor, basal stroma are absent, conidiophores (115–155 × 8.3–11 µm) and phialides (66–140 µm long) are longer, and conidia (18–42 × 5–8 µm) are narrower than those in N. paraunicolor (Nag Raj and Kendrick 1975). Nagrajchalara inflatipes also produces 3-septate conidia in similar size, but in this species conidiophores are well-developed with a septate basal stalk and a terminal phialide, phialides are versicolorous and with darker collarettes, and conidia (18–54 × 5–7 µm) are wider (Nag Raj and Kendrick 1975). ITS sequence generated from the strain 50666 had significant difference from those of N. inflatipes and N. unicolor.
Nagrajchalara pseudoaurea W.P. Wu & Y.Z. Diao, sp. nov., Fig. 77, MycoBank MB845238.

Nagrajchalara pseudoaurea (Wu8272, holotype). a–h Conidiophores and phialidic conidiogenous cells. Scale bars: 10 µm for a–f, 5 µm for g, h
Etymology: Refers to its similarity with Chalara aurea.
Typification: China, Hubei Province, Wufengshan, on dead leaves of unidentified tree, 16 September 2004, Wenping Wu, Holotype HMAS 352212 (= Wu8272), ex-type strain CGMCC3.23386 (= NN50701).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae, 2.5–3.5 µm wide. Anamorph: Stroma present, scattered, composed of dark brown and thick-walled cells. Setae absent. Conidiophores aggregated and directly arising from cells of the basal stroma, erect, straight or slightly curved, simple, obclavate, lageniform, 62–80 µm long, the basal cell dark brown, phialides brown and slightly darker in the lower part of collarette, 1- or rarely 2-septate, smooth, terminating in a phialidic conidiogenous cell. Conidiogenous cells obclavate, lageniform, 58–78 µm long, concolorous or slightly darker in the lower part of collarette, medium brown, smooth, composed of a venter and a collarette; transition from venter to collarette gradual; venters ellipsoidal, subcylindrical, 17.5–25 × 5–7.5 µm; collarettes cylindrical, 37.5–50 × 3–3.5 µm, slightly darker in the lower part; ratio of mean lengths of collarette and venter = 2:1. Conidia endogenous, extruded in short chains, cylindrical, 12.5–16 × 2–2.5 µm straight, base truncated and with a basal frill, apex slightly rounded, hyaline, 1-septate, smooth; mean conidium length/width ratio = 6.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, brown with white margin, reverse concolorous, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara pseudoaurea is similar to N. aurea, but differs in producing short conidiophores with only one-celled stalk, and slightly shorter conidia. It also resembles Chalara alabamensis, but differs by its shorter venters and longer collarettes, and narrower conidia (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; McKenzie et al. 2002).
Nagrajchalara puerensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 78, MycoBank MB845239.

Nagrajchalara puerensis (Wu15215, holotype). a–d, e–j, m Conidiophores and phialidic conidiogenous cells. e, k, l, n–q Conidia. Scale bar: 10 µm for a–d, 5 µm for e–q
Etymology: refers to the locality, Puer in Yunnan Province, where the type specimen was collected.
Typification: China, Yunnan Province, Xishuangbanna, on decaying leaves of unidentified tree, 6 December 2018Y. Zhang, Holotype HMAS 352213 (= Wu15215). Living strains: CGMCC3.23414 (= NN76120).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 3–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae, solitary or aggregated in small group, erect, straight, reduced to a 1–2-celled stalk and a terminal phialide, occasionally well-developed, cylindrical, obclavate, 80–95 μm long, 3–4 µm wide at the base, 1–2-septate, occasionally up to 5-septate, brown to dark brown, smooth- and thin-walled. Conidiogenous cells integrated, terminal, phialidic, erect, straight, lageniform, obclavate, 75–87 µm long, versicolorous, brown, darker in the lower part of collarette, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters subcylindrical, subellipsoidal, 25–29 × 6.5–7.3 µm; collarettes cylindrical, 50–58 µm long, 3–3.5 μm wide, brown, lower part significantly darker than the upper part and venter; ratio of mean lengths of collarette and venter = 2:1. Conidia endogenous, extruded in short chains, cylindrical, base truncated, apex rounded or flattened, 14–17.5 × 2–2.5 µm, hyaline, uniseptate, thin- and smooth-walled, guttulate; mean conidium length/width ratio = 7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to yellow brown, reverse concolorous or fresh yellow brown, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Other material examined: China, Yunnan Province, Xishuangbanna, Jinghong, 6 December 2018, Y. Zhang, Wu15179. Living strains: 75210 (from Wu15215) and 76052 (from Wu15179).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara puerensis is similar to two existing species N. agathidis (conidia 17–24 × 2.5–3 μm) and N. angionacea (conidia 13–18 × 3–3.5 μm) in reduced conidiophores, ellipsoidal venter, abrupt transition from venter to collarette, and 1-septate conidia, but differs from them by longer phialides and narrower conidia (Nag Raj and Kendrick 1975).
Morphologically N. puerensis is also closely related to several new species described in this work, such as N. aunstrupii, N. follicola, N. knudsonii, N. tsukairakuensis and N. tubakii, but differs from them in size of phialides and conidia, and ITS sequence. Nagrajchalara aunstrupii (phialides 93–99 µm long, conidia 15.5–20 × 2.8–3.2 µm) differs from N. puerensis in its longer phialides and slightly longer and wider conidia. In N. follicola (phialides 65–72 µm long, conidia 19–23 × 2.5–3 µm), N. tsukairakuensis (phialides 65–74 µm long, conidia 11–13 × 2.8–3 µm) and N. tubakii (phialides 59–67 µm long, conidia 13–16.5 × 3–3.2 µm), the phialides are shorter and the conidia are wider than those of N. puerensis. The phialides and conidia of N. knudsonii (phialides 75–86 µm long, conidia 16.5–18.5 × 2.2–2.5 µm) are in similar shape and size with those of N. puerensis, but their ITS sequences have 28 bp differences from each other.
Some morphological variations were observed from two studied specimens. In the specimen Wu15179, the conidiophores were constantly with one basic stalk cell, while in the specimen Wu15215, the conidiophores are usually 2-septate. However, their phialides and conidia were almost identical in shape and size. The ITS sequences generated from living strains of these two specimens were almost identical (with 4 bp differences).
Nagrajchalara pulchra (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., Figs. 79, 80, MycoBank MB845281.

Nagrajchalara pulchra (Wu1354b). a, b Conidiophores and conidiogenous cells. c–l Conidia. Scale bar: 5 µm

Nagrajchalara pulchra (a–c from Wu1374b, d–c, l from Wu1441b). a–g Conidiophores and conidiogenous cells. h–l Conidia. Scale bar: 10 µm for a–f, 5 µm for g–l
≡ Chalara pulchra Nag Raj & S. Hughes, N.Z. Jl Bot. 12: 126, 1974.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cell, reduced to a single cell incorporated into hyphae and a phialide, solitary or 2–3 aggregated at the base, erect, straight or slightly curved, simple, subcylindrical, obclavate, 100–120 µm long, medium brown to dark brown, smooth; basal cells 8–15 × 6–9 µm, dark brown. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, subcylindrical, obclavate, 90–115 µm long, apex irregularly ruptured with untruncated and rounded apical apparatus, medium brown to dark brown, smooth or slightly asperate, subcylindrical; venters and collarettes barely differentiated, transition from venter to collarette gradual, 15–18 µm wide in the widest part of venter, 9–11 µm at apex of collarette. Conidia endogenous, extruded in readily seceding short chains, cylindrical, apex blunt or rounded, base slightly truncated and with distinct marginal frills, 45–53 × 8.5–9.5 µm, hyaline, 7–8 septate, smooth- and thick-walled; mean conidium length/width ratio = 5.4:1. Teleomorph: Unknown.
Materials examined: China, Guangxi Province, Damingshan, on dead culm of bamboo, 18 December 1997, Wenping Wu, Wu1354b and Wu1374a; Guangxi Province, Damingshan, on dead culms of bamboo, 19 December 1997, Wenping Wu, Wu1441b; Hunan Province, Mangshan, on dead culm of bamboo, 15 April 2002, Wenping Wu, Wu6107.
Ecology/substrate/host: Saprobe on dead branches, rotten wood of different host, including Weinmannia racemesa, bamboo and other plants.
Geographical distribution: China and New Zealand.
Description and illustration: Nag Raj and Hughes (1974) and Nag Raj and Kendrick (1975).
Notes: Nagrajchalara pulchra was originally described on rotten wood of Weinmannia racemosa and unidentified tree from New Zealand (Nag Raj and Hughes 1974). It is characterized by reduced conidiophores composing of a single basal cell and a phialide, uniformly colored but barely differentiated venters and collarette, and 7–8 septate conidia in large size. Among the known species in Chalara s. lat., Chalara bicolor (conidia 50–60 × 5.5–6 µm), Nagrajchalara insignis (conidia 18–54 × 5–7 µm), N. pulchra (conidia 39–56 × 8.5–11 µm) and N. versicolor (conidia 30–45 × 6.5–7 µm) are with 7-septate conidia. Nagrajchalara pulchra differs from them by uniformly colored conidiogenous cells and broader conidia. In most species of chalara-like fungi, venter and collarette are clearly differentiated in shape and size and wall attenuated toward collarette apex. However, in N. pulchra, conidiogenous cells can be hardly distinguished into venter and collarette, and apex of mature conidiogenous cell is irregularly ruptured. These characters were also found in Ascoconidium; thus, it was regarded as marginal between Chalara s. lat. and Ascoconidium by Nag Raj and Kendrick (1975). However, in Ascoconidium, cell wall of conidiogenous cells is uniformly thick and splitting vertically at apex of collarette to release conidia (Nag Raj and Kendrick 1975). Several specimens of N. pulchra were collected from China; however, no living strain was obtained for molecular phylogeny.
Nagrajchalara qingchengshanensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 81, MycoBank MB845240.

Nagrajchalara qingchengshanensis (a–c, e–n, r, s from Wu17211, holotype; d, o–q from Wu17212). a–j, q–s Conidiophores and phialidic conidiogenous cells. k–p Conidia. Scale bar: 10 µm for a–d, 5 µm for e–s
Etymology: Refers to the location Qingchengshan, Sichuan Province, China, where the fungus was discovered.
Typification: China, Sichuan Province, Chengdu, Qingchengshan, on dead leaf vein of unidentified tree, 9 November 2019, Wenping Wu, Holotype HMAS352214 (= Wu17221), ex-type strain CGMCC3.23439 (= NN77720).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, solitary or 2–3 aggregated at the base, erect, straight or slightly curved, obclavate, lageniform, ampulliform, 35–45 µm long, 5–6 µm wide at the base, 1-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, lageniform, ampulliform, 29–39 µm long, versicolorous, medium brown, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, lageniform, 15–17 m long and 5–5.5 µm wide, brown; collarettes cylindrical, 16–20 × 3–3.2 µm, medium brown, slightly darker in the lower part; ratio of mean lengths of collarette and venter = 1.1:1. Conidia endogenous, extruded in short chains, cylindrical, 13–15 × 2.4–2.6 µm, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 5.6:1.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, then grey to grey brown, reverse brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 12 mm on PDA at 25 °C in 4 weeks. Teleomorph: Unknown.
Other material examined: China, Sichuan Province, Chengdu, Qingchengshan, on dead leaf vein of unidentified tree, 9 November 2019, Wenping Wu, Wu17222. Living strains: 77693 (from Wu17221a) and 77721 (from Wu17222a).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara qingchengshanensis is similar to Cylindrocephalum hughesii, N. cannonii and N. conifericola in reduced conidiophores, shape and size of phialides and conidia, but differs in size of phialides and conidia. Morphologically N. qingchengshanensis differs from Cylindrocephalum hughesii in its slightly shorter phialides and narrower venter; and phylogenetically these two fungi were classified into two different genera in this study (Nag Raj and Kendrick 1975). Nagrajchalara qingchengshanensis has similar-sized phialides and conidia with N. cannonii and N. conifericola, but differs from them by shape of phialide and venter. In N. qingchengshanensis, phialides are with lageniform or cylindrical venters and without significant constriction in the septum between the venter and the basal cell; while in N. cannonii and N. conifericola, phialides are with ellipsoidal venter and with significant constriction in the septum between the venter and the basal cell. Further ITS sequence from N. qingchengshanensis has 17 bp and 21 bp differences from those of N. cannonii and N. conifericola respectively.
Nagrajchalara selaginellae (M.L. Farr) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845282.
≡ Chalara selaginellae M.L. Farr in Farr & Horner, Nova Hedwigia 15: 269, 1968.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Anamorph: Conidiophores erect, straight or slightly curved, cylindrical to subcylindrical, or obclavate, 32–35 µm long, basal cell 5–8 µm wide, 1–2-septate, brown, smooth-walled. Conidiogenous cells integrated, terminal, subcylindrical to lageniform, 25–45 µm long, medium brown to dark brown, smooth, transition from venter to collarette gradual; venters subcylindrical to ellipsoidal, 14–17 × 6.5–8.5 μm; collarettes cylindrical, 25–30 × 4.5–6.5 μm. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 10–18 × 3.4–4 µm, apex blunt or rounded, base truncate with distinct marginal frills, hyaline, smooth, uniseptate. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, purple brown, reverse dark brown, becoming dark soil brown towards the margin, with yellow pigment diffused into agar, sterile, up to 7 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Guangxi Province, Shangsi, Shiwandashan, on dead bark of Pinus sp., 18 December 1997, Wenping Wu, Wu1353. Living strain: CGMCC 3.23360 (= 43153, from Wu1353).
Ecology/substrate/host: Saprobe on dead bark of Pinus sp. and Selaginella rupestres.
Geographical distribution: China and USA (Nag Raj and Kendrick 1975; Bates et al. 2018).
Notes: Nagrajchalara selaginellae differs from other species in wider conidiophores, phialides and conidia (Nag Raj and Kendrick 1975).
Nagrajchalara septata W.P. Wu & Y.Z. Diao, sp. nov., Fig. 82, MycoBank MB845241.

Nagrajchalara septata (Wu16068, holotype). a–c Setae. d–p Conidiophores and conidiogenous cells. q, r Conidia. Scale bar: 10 µm for a–j, 5 µm for k–r
Etymology: Refers to its septate conidia.
Typification: China, Zhejiang Province, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Holotype HMAS 352215 (= Wu16068), ex-type strain CGMCC3.23457 (= 76452).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae directly arising from superficial hyphae, solitary or aggregated in small group, associated with conidiophores at the base, cylindrical, 167–269 μm long, 5–7 μm wide at the basal part, tapering towards the acute or obtuse apex, dark brown to black, 8–12-septate, with 1–2 percurrent proliferation; basal cell slightly flattened, up to 12 μm wide. Conidiophores formed from superficial hyphae or aggregated cells, solitary or in small group, erect, straight of slightly curved obclavate, subcylindrical, 62–73 µm long, 4–5 µm wide at the base, 1–4-septate, medium brown, smooth, terminating into a phialide. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 48–62 µm long, concolorous, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, lageniform, 19–22 µm long and 5–6 µm wide, medium brown; collarettes cylindrical, 25–33 × 2.8–3 µm, medium brown, pale brown in the upper part; ratio of mean lengths of collarette and venter = 1.4:1. Conidia endogenous, extruded in short chains, cylindrical, 15–17 × 2 µm, base truncated with small frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 8:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to light brown, reverse concolorous, sterile, up to 3 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Zhejiang Province, Huaian County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Wu16064 and Wu16159. Living strains: CGMCC3.23458 (= 76457, from 16068) and 76490 (from Wu16159).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara septata is characterized by presence of dark brown and septate setae, 1–4-septate conidiophores, lageniform or obclavate phialides with cylindrical venters, abrupt transition from venters to collarettes, and 1-septate conidia (15–17 × 2 μm). Morphologically N. septata is similar to N. jonesii and N. mutabilis, but differs from them by septation and size of conidiophores, phialides and conidia (Silva et al 2015; Nag Raj and Kendrick 1975). Compared with N. septata, N. jonesii has longer conidiophores with more septa; C. mutabilis has both fertile and sterile setae, and slightly shorter collarettes.
The three examined specimens were collected from the same locality, and were with identical morphology and ITS sequences. The ITS sequence of N. septata has 14 bp difference from the one of N. mutabilis, while it has very low identity with the one of N. jonesii. On PDA, both N. jonesii and N. septata grow slowly and form packed colony with poorly developed aerial mycelium.
Nagrajchalara setosa (Harkn.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845283.
≡ Chalara setosa Harkn., Bull. Calif. Acad. Sci. 1(no. 3): 164, 1885.
≡ Chaetochalara setosa (Harkn.) Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 155, 1975.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae solitary or aggregated in small group, associated with conidiophores at the base, erect, straight or flexuous, simple, cylindrical, 250–300 µm long, slightly inflated at the base to a width of up to 6.5 µm, gradually tapering toward the apex to a width of 2.5–3 µm, 4–6 septate, dark brown to reddish brown, smooth. Conidiophores reduced to a single stalk cell incorporated into hyphae and a phialide, lageniform, obclavate, pale to medium brown, smooth, basal cell up to 5 µm long and 5–6 µm wide. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, lageniform, obclavate, ampulliform, 30–40 µm long, pale to medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual; venters ampulliform to subcylindrical, 10–13 µm long and 5–6 µm wide; collarettes cylindrical, 12–25 µm long, 3.5–4 µm wide, concolorous with venter, smooth. Conidia endogenous, extruded in short chains, cylindrical, 13–16 × 2–2.5 µm, hyaline, 1-septate, both ends rounded or flattened. Teleomorph: Unknown.
Materials examined: China, Guangxi Province, Damingshan, on dead leaves of Cinnamomum sp., 20 December 1997, Wenping Wu, Wu1464; Guangxi Province, Shiwandashan, on dead leaves of ?Cinnamomum sp., 30 December 1997, Wenping Wu, Wu1559h; Guangxi Province, Shiwandashan, on dead leaves of unidentified plant, 30 December 1997, Wenping Wu, Wu1555a.
Ecology/substrate/host: Saprobe on dead leaves of Cinnamomum sp., Quercus sp..
Geographical distribution: China and USA (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Nagrajchalara setosa was redescribed under Chaetochalara setosa by Nag Raj and Kendrick (1975). Morphologically N. setosa resembles Chalara africana and C. bulbosa, but differs by its 1-septate conidia (Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975).
Nagrajchalara sichuanensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 83, MycoBank MB845242.

Nagrajchalara sichuanensis (Wu13251, holotype). a–l Conidiophores with cylindrical collarettes. m–o Conidia. Scale bar: 10 μm for a–h, 5 μm for i–o
Etymology: Refers to the province Sichuan in China where the fungus was originally discovered.
Typification: China, Sichuan Province, Ya An, Wanguan, Bifengxia, on dead leaves of unidentified tree, 15 December 2013, Wenping Wu, Holotype HMAS352216 (= Wu13251), ex-type strain CGMCC3.23398 (= NN57620).
Description on the natural substrate: Colonies effuse, pale brown, sparse, yellowish appearance from conidial mass. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, smooth-walled, branched hyphae of 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, reduced to a 1–3-celled stalk and a terminal phialide, erect, straight or slightly curved, simple, obclavate, lageniform, subcylindrical, 60–85 × 5–6.5 µm, brown, thick- and smooth-walled in the lower part, becoming thin-walled in the upper part, basal cell inflated. Conidiogenous cells integrated, terminal, phialidic, straight or slightly curved, obclavate, lageniform, 35–75 µm long, concolorous, medium brown to brown, smooth-walled, consisting of a venter and collarette; transition from venter to collarette gradual; venters cylindrical, subcylindrical, 20–28 × 5–6.5 µm; collarettes cylindrical, 35–45 × 3–3.5 µm; ratio of mean length of collarette and venter = 1.7:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 12–15 × 2–2.5 µm, both ends rounded or flattened, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white then pale brown, reverse dark brown, with light brown margin, with yellow pigment diffused into agar, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Sichuan Province, Ya An, Wanguan, Bifengxia, on dead leaves of unidentified tree, 15 December 2013, Wenping Wu, HMAS 352216 (= Wu13251, holotype); Sichuan Province, Ya An, Wanguan, Bifengxia, on dead leaves of Cinnamomum sp., 15 December 2013, Wenping Wu, Wu13203. Living strains: CGMCC3.23399 (= 57632, from 13203).
Ecology/substrate/host: Saprobe on dead leaves of Cinnamomum sp. and other trees.
Geographical distribution: China.
Notes: Nagrajchalara sichuanensis is similar to Chalara alabamensis, Cylindrocephalum aurea and N. agathidis in producing reduced conidiophores and 1-septate conidia, but differs in shape and size of phialide and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In C. alabamensis and C. aurea, phialides are with ellipsoidal venters, while the venters in N. sichuanensis are cylindrical or subcylindrical. In addition, conidiophores of C. aurea are more well-developed, longer and with more septa. Identical ITS sequences were generated from two studied strains, and they had relatively low identity to those from the existing species of Nagrajchalara.
Nagrajchalara sivanesanii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 84, 85, MycoBank MB845243.

Nagrajchalara sivanesanii (Wu16285, holotype). a–g Conidiophores and phialidic conidiogenous cells. Scale bar: 10 µm for a–d, 5 µm for e–g

Nagrajchalara sivanesanii (Wu16285, holotype). a–g Conidiophores and phialidic conidiogenous cells. h Basal part of conidiophores. i–k Conidia. Scale bar: 5 µm
Etymology: Named after the former IMI mycologist A. Sivanesan.
Typification: China, Guangdong Province, Guangzhou, South China Botanical Garden, on dead leaves of Acacia sp., 2 December 2018 Wenping Wu, Holotype HMAS 352217 (= Wu16285), ex-type strain CGMCC3.23424 (= NN76694).
Description on the natural substrate: Colonies effuse, pale brown, sparse. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, smooth-walled, branched hyphae of 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or aggregated irregularly, reduced to a single basal cell bearing a phialide, simple, erect, straight, obclavate, lageniform, 57–70 μm long, 5–8 µm wide at the basal part, brown, thick- and smooth-walled. Conidiogenous cells integrated, terminal, obclavate, lageniform, 54–67 µm long, concolorous or slightly darker in the lower part of collarettes, medium brown to brown, smooth-walled, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters bulbous, ellipsoidal, 15–20 μm long, 6–7 µm wide, brown; collarettes cylindrical, 39–50 × 3–3.5 µm, lower part slightly darker than the upper part and venter; ratio of mean lengths of collarette and venter = 2.5:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 14–19 × 2.5–2.8 µm, apex rounded or flattened, base truncated with frill, hyaline, uniseptate, smooth- and thin-walled, guttulate; mean conidium length/width ratio = 6.2:1.Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, with a brown circle around the margin, reverse concolorous, sterile, up to 13 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves of Acacia sp.
Geographical distribution: China.
Notes: Nagrajchalara sivanesanii is characterized by reduced conidiophores composing of one basal cell and a phialide, bulbous or ellipsoidal venters, and 1-septate conidia (14–19 × 2.6–2.8 μm). It superficially resembles Chalara rhynchophiala and N. angionacea and in morphology of conidiophores, phialide and conidia, but differs from them in shape and size of phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Compared with N. sivanesanii, N. angionacea produces similar-sized phialides (53–64 μm long), but has slightly longer and wider venters (16–23 × 7.5–9 μm) and broader conidia (13–18.5 × 3–3.5 μm); C. rhynchophiala (phialides 37–57 μm long, venters 9–15 × 7.5–10 μm, collarettes 25–42 × 2.5–3.2 μm, conidia 12–23 × 2.5–3.5 μm) produces shorter phialides and wider conidia.
Nagrajchalara strobilina (Fr.) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 86, MycoBank MB845284.

Nagrajchalara strobilina. a–o, q Conidiophores, conidiogenous cells and conidia. p Conidia. Scale bar: 10 μm for a–h; 5 μm for i–q
≡ Peziza strobilina Fr., Syst. Mycol. (Lundae) 2(1): 125, 1822.
= Antinoa strobilina (Fr.) Velen., Monogr. Discom. Bohem. (Prague): 215, 1934.
= Chalara strobilina Sacc., Nuovo G. bot. ital. 8(2): 185, 1876.
≡ Cyathicula strobilina (Fr.) Korf & J.R. Dixon, Mycotaxon 1(2): 92, 1974.
≡ Helotium strobilinum (Fr.) Fuckel, Jb. nassau. Ver. Naturk. 23–24: 313, 1870 (1869–1870).
≡ Hymenoscyphus strobilina (Fr.) W. Phills (as ‘Hymenoscypha’), Man. Brit. Discomyc. (London): 133, 1887.
≡ Ombrophila strobilina (Fr.) P. Karst., Bidr. Känn. Finl. Nat. Folk 23: 92, 1873.
≡ Phialea strobilina (Fr.) Gillet, Champignons de France, Discom.(4): 103, 1879.
= Phialea eustrobilina Korf. Mycologia 49(6): 861, 1957.
Description on the natural substrate: Anamorph: Conidiophores cylindrical, obclavate, 3–10-septate, brown to dark brown, up to 130 μm long. Conidiogenous cells phialidic, obclavate, lageniform, pale brown to brown, 18–35 μm long, transition from venter to collarette abrupt; venter subcylindrical, or rarely subellipsoidal, 5–12 × 3.5–4.5 μm; collarette cylindrical. Conidia hyaline, cylindrical, aseptate, 3–5 × 1–1.5 μm, ends blunt or slightly rounded (From Gams and Philippi 1992). Teleomorph: See Gams and Philippi 1992).
Description in pure culture: Colonies effuse, rounded, dark brown to brown, aerial, reverse side dark brown to black. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Stroma absent. Setae absent. Conidiophores solitary, reduced to a basal cell and a phialide, erect, straight or slightly curved, lageniform, obclavate, 15–18.5 × 3.5–4.5 µm, 1-septate, simple, brown to dark brown, smooth; the basal cells integrated into the hyphae. Conidiogenous cells monophialidic, lageniform, obclavate, 12.5–16.5 µm long, lower part 3.8–4.5 µm wide, upper part 2–2.5 µm wide, brown to dark brown, smooth; collarettes funnel-shaped, or inconspicuous. Conidia extruded in wet spore mass, globose, subglobose, 4–6.5 µm in diam., hyaline, aseptate, smooth. Teleomorph: Unknown.
Material examined: Denmark, Copenhagen, Deer Park, on plant litter, 23 November 1996, Wenping Wu, HMAS 352251 (= Wu9796). Living strain: NN 9796.
Ecology/substrate/host: Saprobes on cone of Picea abies and other plant material.
Geographical distribution: Denmark, Germany, Italy and Norway.
Description and illustration: Gams and Philippe (1992).
Ecology/substrate/host: Saprobe on dead plant material.
Geographical distribution: Denmark.
Notes: Morphologically Nagrajchalara strobilina is characterized by long conidiophores with multiseptate stalks and terminal phialides, abrupt transition from venter to collarette, and hyaline, aseptate, cylindrical conidia in small size (3–5 × 1–1.5 μm) (Nag Raj and Kendrick 1975; Gams and Philippi 1992). Its connection with the teleomorph Phialea strobilina was confirmed by pure culture study from single spore isolates (Gams and Philippi 1992). The pure culture studied here was originally obtained from the typical morphotype of Chalara strobilina collected in natural substrate from Denmark. However, in PDA the fungus morphologically resembled Cadophora in producing sessile conidiogenous cells and hyaline, aseptate, globose conidia in wet spore mass. The phylogenetic analyses clearly showed that it belonged to N. strobilina (Nag Raj and Kendrick 1975).
The three strains (CBS803.84, single ascospore isolate; CBS643.85, single conidial isolate) studied by Gams and Philippi (1992) had identical LSU sequences. The Danish strain from this study had identical ITS sequence with the one from the strain CBS643.85. To our surprise, the phylogenetical analyses with different datasets in this study showed that this species with aseptate conidia clustered together with Nagrajchalara species with septate conidia. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain NN 9796 included Phialea strobilina (EF596821 from the strain CBS643.85, 100% identity), Phialea sp. (MH268052, 93% identity), Mollisina uncinata (JN033404, 92% identity), Hymenoscyphus serotinus (KU204586, 91% identity), and many unidentified fungi of Leotiomycetes.
Nagrajchalara tengii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 87, 88, MycoBank MB845244.

Nagrajchalara tengii (Wu15210, holotype). a–l Conidiophores and phialidic conidiogenous cells. Scale bar: 10 µm for a–k, 5 µm for l

Nagrajchalara tengii (Wu15210, holotype). a–g Conidiophores and phialidic conidiogenous cells. h–j Conidia. Scale bar: 5 µm
Etymology: Named after the Chinese mycologist Prof. Teng Shuchun.
Typification: China, Yunnan Province, Jinghong, Xishuangbanna, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Holotype HMAS 352218 (= Wu15210), ex-type strain CGMCC3.23410 (= NN76061).
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, branched hyphae with thin and smooth walls, 2–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or aggregated irregularly, erect, straight or slightly curved, composed of a 1–3-celled stalk and a terminal phialide, obclavate, lageniform, 70–85 µm long, 6–10 µm at base, brown, smooth- and thick-walled; basal cell inflated, dark brown. Conidiogenous cells integrated, terminal, phialidic, straight or slightly curved, lageniform, obclavate, 70–80 long, concolorous, medium brown to brown, smooth-walled, consisting of a venter and collarette; venters ellipsoidal, or subcylindrical, 22–30 µm long and 9–10 µm wide; collarettes cylindrical, 50–60 µm long and 4.5–5 µm wide, concolorous, slightly paler in the apical part; ratio of mean lengths of collarette and venter = 2.1:1. Conidia endogenous, extruded in short and loose chains, 18–20 × 4–4.5 µm, hyaline, uniseptate, smooth- and thin-walled, cylindrical, apex rounded, base truncate to rounded, without basal frills; mean conidium length/width ratio = 4.5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, brown with paler colored margin, reverse concolorous, with yellow brown pigment diffused into agar, sterile, up to 13 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead leaves of unidentified plant, 2 January 1997, Wenping Wu, Wu1337b and Wu1337d. Living strains: 44417 (from Wu1337b), 44481 (from Wu1337d), 76062 and 76085 (from Wu15210).
Ecology/substrate/host: Saprobe on dead leaves of unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara tengii is characterized by its reduced conidiophores, obclavate or lageniform phialides with ellipsoidal or subcylindrical venters, and wider conidia. Nagrajchalara tengii superficially resembles Cylindrocephalum hughesii and N. angionacea, but differs in shape and size of phialides and conidia. Conidia of N. tengii are longer and broader than those of C. hughesii (12–17 × 2–2.5 µm) and N. angionacea (13–18.5 × 3–3.5 µm). In addition, C. hughesii also differs from N. tengii by shorter conidiophores (33–54 µm long).
Nagrajchalara tropicalis W.P. Wu & Y.Z. Diao, sp. nov., Figs. 89, 90, MycoBank MB845245.

Nagrajchalara tropicalis (a–c from Wu1311g, holotype; d–t from the ex-type strain on PDA in 20 days). a, b, d–i Conidiophores and phialidic conidiogenous cells. j–t Conidia. Scale bar: 10 µm for d–i; 5 µm for a–c, j–t

Nagrajchalara tropicalis (from the ex-type strain 1311 g, on PDA in 20 days). a–f Conidiophores and phialidic conidiogenous cells. g Conidia. Scale bar: 5 µm
Etymology: Refers to its occurrence in tropical area.
Typification: China, Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead leaves of palm, 2 January 1998, Wenping Wu, Holotype HMAS 352219 (= Wu1311g), ex-type strain CBS3.23357 (= NN42844).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or irregularly aggregated at the base, reduced to a single basal cell bearing a phialide, erect, straight, obclavate, lageniform, 60–75 × 7–8 μm, medium brown, 1-septate, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight, obclavate, lageniform, 60–74 µm long, medium brown, concolorous, or slightly darker in the lower part of collarettes, brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 30–34 × 7–8 µm; collarettes cylindrical, 35–40 × 3.7–4.5 μm, medium brown, slightly darker in the lower part; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded in short chains, cylindrical, 14.5–16 × 3.6–3.8 µm, base truncated and with short frills, apex rounded or flattened, hyaline, uniseptate; mean conidium length/width ratio = 3.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, lobbed, aerial mycelium poorly developed, soil brown, reverse light brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Other material examined: China, Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead leaves of Smilax sp., 2 January 1997, Wenping Wu, Wu1284g. Living strain: CGMCC3.23365 (= NN45844, from Wu1284g).
Ecology/substrate/host: Saprobe on dead leaves of Smilax sp. and unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara tropicalis superficially resembles Cylindrocephalum hughesii, N. angionacea and N. tengii, but differs in shape and size of phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Conidia of N. tropicalis differs from C. hughesii (conidia 12–17 × 2–2.5 µm) by longer phialides and broader conidia. Nagrajchalara tropicalis differs from N. angionacea (phialides 53–64 μm long; venters 16–23 × 7.5–9 μm, collarettes 35–43 × 3.5–4.5 μm, and conidia 13–18.5 × 3–3.5 µm) in longer phialides and venters.
Nagrajchalara truncata W.P. Wu and Y.Z. Diao, sp. nov., Fig. 109a, b, MycoBank MB845246.
Etymology: Refers to its conidia with truncated base.
Typification: China, Ningxia Province, Jingyuan County, Liupan Mountain, Liantianxia, on wood of unidentified plant, 25 August 1997, Wenping Wu, Holotype HMAS 352248 (= Wu1541g).
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae with thin and smooth wall, 2–3 µm diam. Anamorph: Stroma absent. Setae absent. Conidiophores arising from superficial hyphae or aggregated cells, solitary or in group, erect, straight or slightly curved, reduced to a basal cell bearing a phialide, obclavate, lageniform, brown; basal cells 5–8 × 3–4 µm. Conidiogenous cells integrated, terminal, lageniform, subcylindrical, 40–60 µm long, concolorous, medium brown, thin- and smooth-walled, consisting of a venter and a collarette; transition from venters into collarettes gradual or abrupt; venters ellipsoidal, 20–35 µm long and 4–5 µm wide; collarettes cylindrical, 25–30 µm long, 2.5–3 µm wide, smooth, concolorous with venter; ratio of mean lengths of collarette and venter = 1:1. Conidia endogenous, extruded in long and loose chains, cylindrical, 11–14 × 2–2.5 µm, apex rounded, base truncated and with short frill, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 5.6:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on rotten wood.
Geographical distribution: China.
Notes: Nagrajchalara truncata resembles Chalara alabamensis, C. gracilis, Cylindrocephalum Kendrickii, N. agathidis, and N. angionacea,, but differs in shape and size of phialides and conidia (Nag Raj and Kendrick 1975). It differs from C. alabamensis, C. kendrickii, N. agathidis, and N. angionacea in narrower venter (less than 5 µm wide at the widest part) and conidia (2–2.5 µm wide). Chalara gracilis also has narrower venter, which is less than 4.5 µm wide, but its phialides are with distinctly rough-walled basal cell and conidia are slightly longer but narrower (9–17 × 1.5–2 µm).
Nagrajchalara tsuensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 91, MycoBank MB845247.

Nagrajchalara tsuensis (Wu16932, holotype). a–h, j, k, m Conidiophores and phialidic conidiogenous cells. i, l, n Conidia. Scale bar: 10 µm for a–e, k; 5 µm for f–j, l–n
Etymology: Refers to the locality Tsu in Japan, where the fungus was originally discovered.
Typification: Japan, Mie Prefecture, Tsu, Tsukairaku Park, on dead leaves of unidentified tree, 2 October 2019, Wenping Wu, Holotype HMAS 352220 (= Wu16932a), ex-type strain CGMCC3.23435 (= NN77469).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or in small group, erect, straight or slightly curved, reduced to a single basal cell bearing a phialide, obclavate, lageniform, 60–72 µm long, 4–5 µm wide at the base, 1-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, obclavate, lageniform, 48–63 µm long, medium brown, versicolorous, slightly darker in the lower part of collarettes, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 20–22 µm long and 5–7.5 µm wide; collarettes cylindrical, 25–37 × 3–3.5 µm, slightly darker in the lower part of collarettes, medium brown; ratio of mean lengths of collarette and venter = 1.5:1. Conidia endogenous, extruded in short chains, cylindrical, 15–21 × 2.3–2.5 µm, base truncates and with short frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 7.5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, then grey, reverse soil brown to brown, becoming paler towards the margin, with brown pigment diffused into agar, sterile, up to 6 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on decaying leaves.
Geographical distribution: China.
Notes: Nagrajchalara tsuensis is similar to N. agathidis, N. angionacea and N. sichuanensis in producing reduced conidiophores, ellipsoidal or subcylindrical venters and 1-septate conidia, but differs in shape and size of phialide and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Compared with N. tsuensis, N. angionacea produces concolorous phialides and wider conidia; N. agathidis produces slightly longer and wide conidia; and N. sichuanensis produces shorter conidia (12–15 × 2–2.5 µm). Furthermore, ITS sequence from the ex-type strain has relatively lower identity with those of N. agathidis, N. angionacea and N. sichuanensis.
Nagrajchalara tsukairakuensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 92, MycoBank MB845248.

Nagrajchalara tsukairakuensis (Wu16941, holotype). a–l, p Conidiophores and phialidic conidiogenous cells. m–o Conidia. Scale bar: 10 µm for a–h; 5 µm for i–p
Etymology: Refers to the locality Tsukairaku Park in Tsu, Mie, Japan, where the fungus was originally discovered.
Typification: Japan, Mie Prefecture, Tsu, Tsukairaku Park, on dead leaves of unidentified tree, 2 October 2019, Wenping Wu, Holotype HMAS 352221 (= Wu16941), ex-type strain CGMCC3.23430 (= NN77323).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or in small group, erect, straight or slightly curved, reduced to a single basal cell bearing a phialide, obclavate, lageniform, 65–75 µm long, 4–5.5 µm wide at the base, 1-septate, medium to dark brown, smooth. Conidiogenous cells integrated, terminal, phialidic, straight or slightly curved, obclavate, lageniform, (57–)65–74 µm long, concolorous, medium to dark brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 23–29 µm long and 6.8–7.2 µm wide, brown; collarettes cylindrical, 46–48 × 3.5–4.5 µm, brown, slightly darker in the lower part; ratio of mean lengths of collarette and venter = 1.8:1. Conidia endogenous, extruded in short chains, cylindrical, 11–13 × 2.8–3 µm, base truncated and with short frills, apex rounded or flattened, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 4.1:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, reverse fresh soil brown, becoming paler towards the margin, with purple pigment diffused into agar, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Other studied living strain: 77355 (from Wu16941).
Ecology/substrate/host: Saprobe on decaying leaves.
Geographical distribution: China.
Notes: Nagrajchalara tsukairakuensis is characterized by reduced conidiophores, obclavate or lageniform phialides with ellipsoidal venters, cylindrical collarette with slightly darker basal part, and 1-septate and shorter conidia (11–13 × 2.8–3.0 μm). Morphologically N. agathidis (conidia 17–24 × 2.5–3 μm) and N. angionacea (conidia 13–18.5 × 3–3.5 μm) are similar to N. tsukairakuensis, but differs from the latter species by longer conidia. In addition, phialides of N. angionacea are unicolourous (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Nagrajchalara tsukairakuensis also resembles N. tubakii, but differs from it by shorter conidia. Pure culture of the ex-type strain of N. tsukairakuensis produces purple pigment diffused into agar plate, which is very characteristic among the known species of the genus.
Nagrajchalara tubakii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 93, 94, 95, MycoBank MB845249.

Nagrajchalara cf. tubakii (Wu16325). a–e, h–k Conidiophores and conidiogenous cells. f, g Conidia. Scale bar: 10 µm for a–e, 5 µm for f–k

Nagrajchalara tubakii (Wu16906, holotype). a–g Conidiophores and conidiogenous cells. h Basal part of conidiophores. i Basal part of conidiophore with percurrent proliferation. j, k Conidia. Scale bar: 5 µm

Nagrajchalara tubakii (Wu16947). a–h Conidiophores and conidia. i–k Conidia. Scale bar: 5 µm
Etymology: Named after the Japanese mycologist K. Tubaki.
Typification: Japan, Mie Prefecture, Tsu, Mie Center for the Arts, on dead leaves of unidentified tree, 3 October 2019, Wenping Wu, Holotype HMAS 352222 (= Wu16906), ex-type strain CGMCC3.23431 (= NN77326).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3.5 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or in small group of 2–4, erect, straight or slightly curved, reduced to a 1–2-celled basal stalk and a phialide, obclavate, 55–74 µm long, 5.5–6.5 µm wide at the base, brown to dark brown, smooth; basal cell lobbed and up to 13 μm. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, obclavate, lageniform, 59–67 µm long, pale brown to medium brown, concolorous or only slightly darker in the basal part of collarettes, smooth, composing of a venter and a collarette; transition from venter to collarette abrupt; venters ellipsoidal, 21–26 µm long and 6.5–7.5 µm wide, brown; collarettes cylindrical, 37–44 × 3.8–4.5 µm, slightly darker than venter, brown; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, extruded in short chains, cylindrical, 13–16.5 × 3–3.2 µm, base truncate with short frills, apex rounded or flattened, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 4.8:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, slightly yellow brown, soil brown, reverse concolorous, becoming paler towards the margin, sterile, up to 13 mm on PDA at 25 °C in 4 weeks. The different strains appear differently in growth rate, color and pigment on PDA.
Other materials examined: China, Guangdong Province, Guangzhou, South China Botanical Garden, on dead leaves of unidentified tree, 2 December 2018, Wenping Wu, Wu16325b; Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Wu15164; Japan, Mie Prefecture, Tsu, Mie Center for the Arts, on dead leaves of unidentified tree, 3 October 2019, Wenping Wu, Wu16891, Wu16896 and HMAS 352198 (= Wu16947). Living strains: CGMCC3.23404 (= 75999, from Wu15164), CGMCC3.23425 (= 76708, from Wu16325b), CGMCC3.23429 (= 77258, from 16896R), CGMCC3.23421 (= 77326), 77268 (from Wu16906), 77311 (from Wu16891R) and 77343 (from Wu16891), 77345 (from 16896).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: Japan.
Notes: Nagrajchalara tubakii is morphologically similar to N. agathidis (conidia 17–24 × 2.5–3 μm) and N. angionacea (conidia 13–18.5 × 3–3.5 μm), but differs from them by longer conidia. In addition, phialides of N. angionacea are unicolourous (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Nagrajchalara tubakii also resembles N. tsukairakuensis, but differs by longer conidia.
ITS sequences of N. tubakii were generated from seven studied strains. The ITS sequences from two Chinese strains (75999 and 76708) are with minor difference with those from the other five Japanese strains (77258, 77268, 77311, 77343, 77345).
Nagrajchalara unicolor (S. Hughes & Nag Raj) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 96, MycoBank MB845285.

Nagrajchalara unicolor (Wu1248b). a–d, g Conidiophores and conidiogenous cells. e, f, h–m Conidia. Scale bar: 20 µm for a, b; 10 µm for c–e, m; 5 µm for g–l
≡ Chalara unicolor S. Hughes & Nag Raj, in Nag Raj & Hughes, N.Z. Jl Bot. 12: 121, 1974.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent or present, dark brown, composed of dark brown, irregularly shaped cells. Setae absent. Conidiophores directly arising from the cells of basal stroma or superficial hyphae, solitary or aggregated, erect, straight or slightly curved, reduced to a small single basal cell incorporated into hyphae and a phialide, cylindrical, lageniform, 80–130 µm long, unicolorous, medium brown to dark brown, verruculose in the lower part, smooth in the upper part; basal cell 5–10 × 6–9 µm. Conidiogenous cells discrete or integrated, terminal, erect, straight or slightly curved, subcylindrical, lageniform, 65–130 µm long, concolorous, brown to dark brown, aspirate in the lower part; venter and collarette barely differentiated, transition from venter to collarette gradual; venters ellipsoidal, 12–16 µm wide in the widest part; collarettes cylindrical, 8.5–9 µm at the opening part, apex irregularly ruptured with untruncated and rounded apical appearance. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 25–30 × 7–8 µm, apex blunt or rounded, base truncate or slightly rounded with distinct marginal frills, hyaline, 3 septate; mean conidium length/width ratio = 3.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, grey to brown, reverse concolorous, with yellow pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks. The two studied strains are different in growth rate and appearance on PDA.
Materials examined: China, Guangdong Province, on dead branches of unidentified plant, 9 October 1998, Wenping Wu, Wu2052a; Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead branch of unidentified plant, 2 January 1997, Wenping Wu, Wu1248b, Wu1322b and Wu1322d; Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead branch of Pinus sp., 2 January 1997, Wenping Wu, Wu1257a; Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead culms of Saccharum sp., 2 January 1997, Wenping Wu, Wu1258e; Yunnan Province, Simao, on dead branches of unidentified plant, 12 October 1999, Wenping Wu & Yan Huang, Wu2670b. Living strains: CGMCC3.23364 (= NN 44014, from Wu1322b) and CGMCC4.23447 (= NN 45977, from Wu2670b).
Ecology/substrate/host: Saprobe on dead culm, branches and rotten wood of different plants including Leptospermum scoparium, Pinus sp., Saccharum sp., and other plants.
Geographical distribution: China, New Zealand and USA (Nag Raj and Kendrick 1975; Li et al. 2013a, b).
Description and illustration: Nag Raj and Hughes (1974), Nag Raj and Kendrick (1975), and Li et al. (2013a, b).
Notes: Nagrajchalara unicolor, described from New Zealand, has barely differentiated conidiophores, uniformly colored venter and collarette, and hyaline, 3-septate and cylindrical conidia bearing distinct frill (Nag Raj and Hughes 1974; Nag Raj and Kendrick 1975). Morphologically N. unicolor resembles N. pulchra in shape of conidiophores, conidiogenous cells, and conidia, but differs in its 7–8 septate and longer conidia in N. pulchra. In conidial septation, several species of Nagrajchalara and Chalara s. lat. produces 3-septate conidia, such as C. grandispora, C. magnispora, C. pteridina, N. angustata, N. inflatipes, N. paraunicolor, and N. unicolor. Nagrajchalara unicolor differs from all these species by its reduced conidiophores composed of one small basal cell and a terminal phialide, uniformly colored venter and collarettes, and broader conidia (Nag Raj and Hughes 1974; Nag Raj and Kendrick 1975; Matsushima 1993; Kowalski and Halmschlager 1996).
The studied Chinese specimens of this fungus had almost identical morphology as those from the type material, however conidia from these collections were slightly shorter in average (18–42 × 5–8 µm in the original description, Nag Raj and Hughes 1974). In the specimen Wu1248b, the fungus produced well-developed basal stroma, and the conidiophores were aggregated in cluster from the stroma; while in other specimens, no well-developed stroma were observed, and the conidiophores were solitary and directly arising from the superficial hyphae. Li et al. (2013a, b) reported occurrence of this species from China, where the conidia were with less variation in size than those in the original description (Nag Raj and Hughes 1974).
ITS sequences of this species were generated for the first time from two studied strains. The phylogenetic analyses showed that they clustered together with other Nagrajchalara species with septate conidia.
Nagrajchalara venicola W.P. Wu & Y.Z. Diao, sp. nov., Figs. 97, 98, MycoBank MB845250.

Nagrajchalara venicola (WuYN015, holotype). a–j Conidiophores and conidiogenous cells. k–m Conidia. Scale bar: 10 µm for a–j, 5 µm for k–m

Nagrajchalara venicola (WuYN015, holotype). a–d, f–h Conidiophores and conidiogenous cells. e Conidia. Scale bar: 5 µm
Etymology: Refers to its growing on leaf vein of decaying leaves.
Typification: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Y. Zhang, Holotype HMAS 352223 (= YN15), ex-type strain CGMCC3.23407 (= NN76015).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, solitary or rarely 2–3 in groups, reduced to a 1–2-celled basal stalk and a phialide, erect, straight or curved, obclavate, lageniform, 97–117 µm long, 4–5.5 µm wide at the base, 1–2-septaete, occasionally 2-septate, medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or curved, obclavate, lageniform, 89–96 µm long, medium brown, versicolorous, slightly darker in the lower part of collarettes, smooth; basal cell flattened, dark brown; composing of a venter and a collarette, transition from venter to collarette abrupt; venters subcylindrical, subellipsoidal, 40–48 µm long and 6.5–7 µm wide, medium brown; collarettes cylindrical, 48–55 × 3.3–3.6 µm, medium brown, versicolorous, slightly darker in the lower part; ratio of mean lengths of collarette and venter = 1.2:1. Conidia endogenous, extruded in short chains, cylindrical, 15–18.5 × 2.5–2.8 µm, both ends obtuse or flattened, without frill, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 6.3:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, pale brown, reverse soil brown to brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Yunnan Province, Xishuangbanna, Jinghong, on dead leaves of unidentified tree, 6 December 2018, Wenping Wu, YN08 and YN11. Living strains: 76975 (from YN08) and CGMCC3.23406 (= 76011, from YN11).
Ecology/substrate/host: Saprobe on decaying leaves.
Geographical distribution: China.
Notes: Nagrajchalara venicola is similar to N. agathidis in possessing reduced conidiophores with a single basal cell and a phialide, versicolorous phialides with ellipsoidal or subcylindrical venter, and 1-septate conidia, but differs by longer conidia (Nag Raj and Kendrick 1975). Nagrajchalara venicola also resembles N. guangcaii, N. nawawii and N. puerensis described in this study, including, but differs in shape and size of phialides and conidia. Nagrajchalara venicola is mostly similar to N. guangcaii (conidiophores 1–4-septatae and 110–135 µm long, phialides 94–101 µm long, venters 41–54 × 6.5–8.5 µm, collarettes 51–54 × 3.7–4.2 µm, conidia 16.5–21 × 2.5–3 µm), and can be distinguished by slightly shorter conidiophores with fewer septa, and shorter and narrower conidia in N. venicola. Furthermore, ITS sequence of N. guangcaii and N. venicola had 29 bp differences from each other. In N. puerensis, collarettes are 2 times longer than venter, while size in N. venicola collarettes and venters are equal in size. Nagrajchalara venicola differs from N. nawawii (phialides 65–86 µm long, venters 25–37 × 5.5–6.5 µm, collarettes 30–47 × 3.5–4 µm, conidia 14–19 × 3–3.5 µm) in its longer phialides and narrower conidia.
Nagrajchalara versicolor W.P. Wu & Y.Z. Diao, sp. nov., Fig. 99, MycoBank MB845251.

Nagrajchalara versicolor (Wu5542a, holotype). a–h Conidiophores and phialidic conidiogenous cells. i–k Conidia. Scale bar: 10 µm
Etymology: Refers to its versicolorous phialides with darker collarettes than venters.
Typification: China, Hainan Province, on dead branches of unidentified plant, 19 December 2000, Wenping Wu & Yan Huang, Holotype HMAS 352249 (= Wu5542b).
Description on the natural substrate: Colonies effuse, scattered, brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma absent or poorly developed, composed of dark brown, irregularly shaped cells. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or 2–4 in groups, reduced into a single cell incorporated into hyphae and a phialide, erect, straight or slightly curved, obclavate, lageniform, 150–190 µm long, versicolorous, brown in the lower part, dark brown in the upper part, verruculose; basal cell 5–7 × 8–10 µm, dark brown. Conidiogenous cells integrated, terminal, erect, straight or slightly curved, obclavate, lageniform, 145–180 µm long, verruculose, versicolorous, brown to dark brown; transition from venter to collarette abrupt; venters ellipsoidal, 30–38 × 14–15 µm, brown, verruculose; collarettes cylindrical, 80–95 × 9–10 µm, dark brown, significantly darker than venter, verruculose, asperate, apex irregularly ruptured with untruncated and rounded apical appearance; ratio of mean lengths of collarette and venter = 2.6:1. Conidia endogenous, extruded in readily seceding long chains, cylindrical, (18–)30–45 × 6.5–7 µm, hyaline, 3–7-septate but most 7-septate, smooth- and thin-walled, apex blunt or rounded, base truncated or flattened and with distinct marginal frills; mean conidium length/width ratio = 5.6:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead branches.
Geographical distribution: China.
Notes: Nagrajchalara versicolor resembles Chalara bicolor, C. cibotti, N. insignis, N. pulchra and N. wenyingiae in having 7-septate conidia, but can be distinguished from them by shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In C. cibotti, conidia are clavate and smaller in size (24–36 × 4.5–6.5 µm), thus can be easily distinguished from N. versicolor. In Chalara bicolor (conidia 50–66 × 5.5–6 µm) and N. insignis (conidia 18–54 × 5–7 µm), conidiophores are well-developed with a multi-septate basal stalk and a terminal phialide, and conidia are longer than those in N. versicolor. Similar as in N. versicolor, conidiophores of N. pulchra (conidia (39–56 × 8.5–11 µm) and N. wenyingiae (conidia 35–50 × 3.7–4.5 µm) are also reduced to a single basal cell and a phialide, but phialides and conidia are significantly wider (in N. pulchra) or narrower (in N. versicolor) than those in N. versicolor.
Nagrajchalara wenyingiae W.P. Wu & Y.Z. Diao, sp. nov., Fig. 100, MycoBank MB845252.

Nagrajchalara wenyingiae (Wu8010, holotype). a–i Conidiophores and conidiogenous cells. j–o Conidia. Scale bar: 10 µm for a–f; 5 µm for g–o
Etymology: Named after the Chinese mycologist Prof. Wenying Zhuang from Institute of Microbiology, The Chinese Academy of Science, Beijing.
Typification: China, Hubei Province, Shengnongjia, on dead culm of bamboo, 17 Sept. 2004, Wenping Wu, Holotype HMAS 352250 (= Wu8010).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae, solitary or 2–3 in groups, erect, straight or slightly curved, reduced to 1–5-celled basal stalk and a terminal phialide, obclavate, lageniform, 110–160 µm long, basal part 10–12.5 µm wide, dark brown, smooth at the basal part, verruculose at the upper part, 0–5 septate, often with 1–4 percurrent proliferations at the basal part. Conidiogenous cells integrated, terminal, erect, straight, lageniform, obclavate, 110–125 µm long, concolorous, or slightly darker in the collarettes, brown to dark brown, aspirate; transition from venter to collarette abrupt; venters ellipsoidal, subcylindrical, 30–37.5 µm long, 10–12.5 µm wide in the widest part, brown, smooth or slightly aspirate; collarettes cylindrical, 67.5–75 × 5–6.5 µm, slightly darker than venters, aspirate; ratio of mean lengths of collarette and venter = 2.1:1. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 35–50 × 3.7–4.5 µm, apex blunt or rounded, base truncated or flattened and with distinct marginal frills, hyaline, 3–7 but mostly 7-septate; mean conidium length/width ratio = 10.4:1. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead culm.
Geographical distribution: China.
Notes: Nagrajchalara wenyingiae also resembles Chalara bicolor, C. cibotti, N. insignis, N. pulchra and N. versicolor in producing 7-septate conidia, but differs from them in shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In C. cibotti, the conidia are clavate and smaller in size (24–36 × 4.5–6.5 µm), thus differs from N. wenyingiae. In Chalara bicolor (conidia 50–66 × 5.5–6 µm) and N. insignis (conidia 18–54 × 5–7 µm), conidiophores are well-developed and with a multi-septate basal stalk and a terminal phialide, and conidia are wider than those of N. wenyingiae. Similar as in N. wenyingiae, conidiophores of N. pulchra (conidia (39–56 × 8.5–11 µm) and N. versicolor (conidia 30–45 × 6.5–7 µm) are also reduced, but phialides and conidia are significantly wider than those of N. wenyingiae.
Nagrajchalara xiaohuiae W.P. Wu, sp. nov., Fig. 101, MycoBank MB846919.

Nagrajchalara xiaohuiae (Wu18064, holotype). a–f Conidiophores and conidiogenous cells. g Basal part of conidiophores. h–l Conidia. Scale bar: 5 μm
Etymology: Named after my friend Xiaohui Chen who was born in Hangzhou where the type specimen was collected during our trip together.
Typification: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on rotten cupules of Castanopsis tibetana Hance, 24 September 2022, Wenping Wu, Holotype HMAS 352301 (= Wu18084); ex-type strain CGMCC 3.24555 (= NN78663).
Description on the natural substrate: Colonies effuse, hairy, white to pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae with thin and smooth wall, 2–4 µm diam. Anamorph: Stroma absent. Setae absent. Conidiophores arising from superficial hyphae, solitary or 2–3 in groups, erect, straight or curved, unbranched, cylindrical, (49–)59–71 μm long, 3.2–3.8 μm wide above the basal swollen cells, composed of a cylindrical stalk and a terminal obclavate phialide, 2–4-septate, smooth, thin-walled, subhyaline to pale brown, lower part brown, paler toward the upper part; basal cell lobbed, dark brown, thick-and rough-walled, 5–7 μm high, 4–6 μm wide. Conidiogenous cells integrated, terminal, solitary, erect, straight or slightly curved, obclavate, 32–37 μm long, subhyaline to pale brown, composed of a venter and a collarettes, transition from venter to collarette abrupt; venter subcylindrical, 17–20 µm long, 4.2–4.5 μm wide at the widest part, subhyaline to pale brown, smooth- and thin-walled; collarette cylindrical, 13–16 µm long, 2.2–2.8 µm wide, smooth, concolorous or slightly with venter or basal part slightly darker than venter and upper part of collarette; ratio of mean lengths of collarette and venter = 0.78:1. Conidia endogenous, extruded in long and loose chains, cylindrical, (11.5–)13–15.5 × 2–2.3 µm, apex rounded or obtuse, base truncate and without basal frill, hyaline, aseptate, thin- and smooth-walled; mean conidium length/width ratio = 6.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white to slightly yellow colored, reverse soil brown to dark brown, becoming paler towards the margin, with brown pigment diffused into agar, sterile, up to 8 mm on PDA at 25 °C in 3 weeks.
Other materials examined: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on rotten cupules of Castanopsis tibetana, 24 September 2022, Wenping Wu, Wu18085, Wu18086, Wu18094. Living strains: 78664 (from Wu18084), 78665 and 78666 (from Wu18085), 78667 (from Wu18086) and 78670 (from Wu18094).
Ecology/substrate/host: Saprobe on rotten cupule of Castanopsis tibetana.
Geographical distribution: China.
Notes: Nagrajchalara xiaohuiae is characterized by well-developed pale brown conidiophores consisted of a 2–4-septated basal stalk and a terminal phialide, abrupt transition from venter to collarette, and hyaline, septate and cylindrical conidia bearing no basal frill. It morphologically resembles to several existing species with well-developed conidiophores and hyaline, aseptate and cylindrical conidia, such as Chalara cylindrosperma (conidiophores 32–190 μm long; phialides 16–47 μm long, venter 9.5–22 × 4.5–8 μm, collarettes 12–28 × 2–3 μm, conidia 5.5–17 × 1.5–2.5 μm), C. longiphora (conidiophores 90–155 μm long; phialides 31–35 μm long, venter 16–17.5 × 5–6 μm, collarettes 14–17 × 2.5–2.7 μm, conidia 6–8 × 2–2.5 μm), C. longipes (conidiophores 63–120 μm long; phialides 21–29 μm long, venter 15–18 × 2–5 μm, collarettes 5.5–9.5 × 1.5–2 μm, conidia 3.5–6.5 × 1–1.5 μm), C. nothofagi (conidiophores 115–168 μm long; phialides 45–58 μm long, venter 20–27 × 6–8.5 μm, collarettes 26–33 × 3.5–4 μm, conidia 13–17 × 2.5–3 μm) and C. platanicola (conidiophores 70–100 μm long; phialides 26–28 μm long, venter 11–12.5 × 5.6–7.5 μm, collarettes 17–18 × 2.8–3.3 μm, conidia 9–10.5 × 2.2–2.5 μm) (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Nagrajchalara xiaohuiae differs from all these species by producing shorter, dark brown and fewer septated conidiophores. In addition, compared with N. xiaohuiae, conidia of C. longipes and C. platanicola are shorter; collarettes and conidia of C. nothofagi are much wider; and phialide and conidia of C. cylindrosperma are with wider ranges (Nag Raj and Kendrick 1975).
The phylogenetic analyses showed that this fungus with aseptate conidia clustered together with other species of Nagrajchalara producing septate conidia, rather than with species of Chalara s. str. with aseptate conidia (Fig. 101). Among accepted species of Nagrajchalara, N. strobilina (conidia aseptate, 2.5–5 × 1.5–2.2 µm) is the only species with aseptate conidia and differs from N. xiaohuiae by smaller conidia (Gams and Philippi 1992). Identical ITS sequences of N. xiaohuiae were generated from six studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain CGMCC 3.24555 included Mollisina uncinata (JN033457, 90% identity), Calycellina fagina (OL752703, 90%), Phialina lachnobrachyoides (JN033412, 90% identity), and many unnamed fungi of Leotiomycetes. Based on a partial LSU megablast search of GenBank nucleotide database, the closest matches to the ex-type strain CGMCC 3.24555 include Leptodontidium beauverioides (MH872794, 98% identity), Phialea strobilina (EF596821, 98% identity), Tricladium caudatum (GQ477318, 98% identity), Triposporium cycadicola (NG_067285, 98% identity), T. deviatum (KJ869177, 98% identity), and many unnamed fungi of Leotiomycetes.
Nagrajchalara yinglaniae W.P. Wu & Y.Z. Diao, sp. nov., Figs. 102, 103, 104, MycoBank MB845253.

Nagrajchalara yinglanii (Wu16125, holotype). a–j, p Conidiophores and conidiogenous cells. l part of collarette and conidia. k, m, n Conidia. o Lobbed basal cell of conidiophores. Scale bar: 10 µm for a–c, 5 µm for d–p

Nagrajchalara yinglanii (Wu16182). a–c, e–h Conidiophores and conidiogenous cells. d, i Conidia. Scale bar: 10 µm for a–c, 5 µm for d–i

Nagrajchalara yinglanii (Wu16197). a–h Conidiophores and conidiogenous cells. i, j Conidia. Scale bar: 10 µm for a–b, 5 µm for c–j
Etymology: Named after the Chinese mycologist Prof. Yinglan Guo from Institute of Microbiology, The Chinese Academy of Science, Beijing, for her significant contribution to anamorphic fungi from China.
Typification: China, Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Holotype HMAS 352224 (= Wu16125), ex-type strain CGMCC3.23455 (= NN76400).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 3–4 µm wide. Anamorph: Stroma absent or poorly developed, composed of dark brown, irregular–shaped and thick-walled cells. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or 2–5 in groups, erect, straight or curved, obclavate, lageniform, subcylindrical, 60–85 µm long, 4–5 µm wide, 1–6 septate, concolorous, medium brown, smooth; basal cell dark brown, lobbed, up to 10 μm wide. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 40–60 µm long, concolorous, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, subcylindrical, 17–24 µm long and 5–6.5 µm wide; collarettes cylindrical, 21–34 × 2.5–2.8 µm, medium brown; ratio of mean length of collarette and venter = 1.3:1. Conidia endogenous, extruded in short chains, cylindrical, 11–15 × 2.3–2.5 μm, base truncated, apex obtuse or rounded, hyaline, uniseptate, thin- and smooth-walled; mean conidium length/width ratio = 5.4:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, grey brown to soil brown, reverse soil brown to brown, with brown pigment diffused into agar, sterile, up to 3 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead fruit of Fagaceae, 26 August 2022, Wenping Wu, Wu13131 and Wu13132; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of Quercus sp., 18 October 2018, Wenping Wu, Wu16191; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Wu16159, Wu16182, Wu16197 and Wu16198). Living strains: 76436 (from 16191a), 76438 (from Wu16197b), 76473 (from 16191), 76474 (from 16197), 76475 (from Wu16198), 76549 (from Wu16182) and 76552 (from Wu16159a), 78708 and 78728 (from Wu18131), and 78729 (from Wu18132).
Ecology/substrate/host: Saprobe on dead leaves of Quercus sp. and unidentified tree.
Geographical distribution: China.
Notes: Nagrajchalara yinglaniae resembles Chalara aotearoae, C. inaequalis, C. tubifera and N. agathidis in septate conidiophores, versicolorous phialides, cylindrical to subcylindrical venters, and 1-septate conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In C. aotearoae, the conidia (13–18 × 3–3.5 μm) are slightly longer and significantly wider than those of N. yinglaniae. Nagrajchalara yinglaniae differs from C. inaequalis by conidia with two cells in equal size; from C. tubifera by shorter conidiophores; and from N. agathidis in multiseptate conidiophores and shorter conidia. Morphological variations were found among different specimens, for example, in the specimen Wu16125 conidiophores were with fewer (1–3) septa and conidia were shorter (11–13 μm long), while in the specimen Wu16182 and u16197, conidiophores were 3–6-septate, and conidia were longer (up to 16 μm long). Identical ITS sequences of this species were generated from eight strains isolated from different specimens.
Nagrajchalara yongnianii W.P. Wu & Y.Z. Diao, sp. nov., Figs. 105, 106, 107, 108, MycoBank MB845254.

Nagrajchalara yongnianii (Wu11023). a–f Conidiophores and phialidic conidiogenous cell. g–j Conidia. Scale bar: 5 µm

Nagrajchalara yongnianii (Wu16916, holotype). a–h Conidiophores and phialidic conidiogenous cell. Scale bar: 5 µm

Nagrajchalara yongnianii (Wu16916, holotype). a Conidiophores and phialidic conidiogenous cell. b–i Conidia. Scale bar: 10 µm for a, 5 µm for b–i

Nagrajchalara yongnianii (Wu17588). a–h Conidiophores and phialidic conidiogenous cell. i–w Conidia. Scale bar: 10 µm for a–e, 5 µm for f–w
Etymology: Named after the former Chinese mycologist Prof. Yongnian Yu from Institute of Microbiology, The Chinese Academy of Science, Beijing.
Typification: Japan, Mie Prefecture, Tsu, Mie Center of the Arts, on dead fruit of unidentified tree, 3 October 2019, Wenping Wu, Holotype HMAS 352225 (= Wu16916), ex-type strain CGMCC3.23432 (= NN77394).
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to brown, septate, branched hyphae with thin and smooth walls, 2–3.5 µm wide. Anamorph: Stroma absent or poorly developed, composed of dark brown, irregularly shaped cells. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or 2–5 in small groups, reduced a single basal cell incorporated into hyphae and a phialide, erect, straight or slightly curved, lageniform, obclavate, subcylindrical, 70–85 µm long, 5–7 µm at base, basal cell inflated, concolorous, brown, smooth-walled. Conidiogenous cells integrated, terminal, straight or slightly curved, lageniform, obclavate, subcylindrical, 68–80 µm long, concolorous, medium brown to brown, smooth-walled, consisting of a venter and a collarette; transition from venters to collarettes abrupt; venters ellipsoidal, 25–30 µm long and 7–9 µm wide; collarettes cylindrical, 40–55 µm long and 3.5–4.5 µm wide; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, extruded in short and loose chains, cylindrical, 22–27 × 3.3–3.6 µm, apex rounded or flattened, base truncated and with distinct frills, hyaline, uniseptate, smooth- and thin-walled, guttulate; mean conidium length/width ratio = 7.1:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, grey to brown, reverse concolorous, sterile, up to 16 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Guangdong Province, Shenzhen, Yangtaishan Forestry Park, on dead seed of unidentified tree, 17 October 2020, Wenping Wu, Wu17588; Hainan Province, Sanya, Yalongwan Park, on dead leaves of unidentified tree, 28 December 2020, Wenping Wu, Wu17639; China: Hunan Province, Zhangjiajie, on dead leaves of unidentified tree, 15 October 2010, Wenping Wu, Wu11022, Wu11023 and Wu11041a; Jiangsu Province, Wuxi, Wuxi Forestry Park, on dead fruit of Cyclobalanopsis sp., 25 August 2019, Wenping Wu, Wu17878 and Wu17879; Yunnan Province, Xishuangbanna, on dead leaves of unidentified plant, 16 October 1999, Wenping Wu & Yan Huang, Wu2737b; Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead fruits of Fagaceae, 26 August 2022, Wenping Wu, Wu18042, Wu18090, Wu18129, Wu18137; Zhejiang Province, Huai An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Wu16144; Japan, Mie Prefecture, Tsu, Mie Center of the Arts, on dead fruit of unidentified tree, 3 October 2019, Wenping Wu, Wu16912. Living strains: CGMCC3.23366 (= NN 46066, from Wu2737b), CGMCC3.23427 (= 77190, from Wu16879a), 53374 (from Wu11041a), 53376 (from Wu11022), 53436 (from Wu11023), 76336 (from Wu16144a), 76337 (from Wu16144b), 77191 (from Wu16879b), 77192 (from Wu16879c), 77212 (from Wu16878a), 77213 (from Wu16878b), 77214 (from Wu16878c), 77391 (from Wu16912), 78397, 78398 and 78399 (from Wu17588), 78462 and 78463 (from Wu17639), 78645 and 78646 (from Wu18042), 78668 and 78669 (from Wu18090), 78706 and 78727 (from Wu18129), 78714 and 78730 (from Wu18137).
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Nagrajchalara yongnianii resembles Chalara alabamensis, N. angionacea and N. aspera in the reduced conidiophores composed of one basal cell and a terminal phialide, but differs from them by the unique combination of different morphological characters such as setae, conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In C. alabamensis and N. angionacea, phialides are shorter, and conidia are shorter and narrower than those of N. yongnianii. Nagrajchalara aspera produces sterile setae among the conidiophores, the phialides are verruculose in upper part, and conidia are shorter and narrower than those of N. yongnianii.
The conidia size of this species varied among different specimens, and usually between 22 to 27 µm long. However, in the four specimens (Wu18042, Wu18090, Wu18129 and Wu18137) recently collected on decaying fruit of Fagaceae from Hangzhou, the conidia were smaller (16.5–22 × 3.5–3.8 µm). Eight living strains were obtained from these four specimens, and their ITS sequences were identical to those from many other strains. ITS sequences were generated from 26 strains isolated from eleven different specimens collected in China and Japan. The ITS sequences from the strains 46066, 78462 and 78463 are identical and have 11 bp differences from those generated from all other strains. On PDA, these strains appeared with white to brown colony, relatively fast growing, and without pigment diffused into agar plate.
Nagrajchalara yunnanensis (W.P. Wu) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 109c, d, MycoBank MB845286.

Nagrajchalara truncata (a, b, Wu1541g, holotype) and N. yunnanensis (c, d Wu952a, holotype). a, c Conidiophores and conidiogenous cells. b, d Conidia. Scale bar: 20 μm
≡ Chalara yunnanensis W.P. Wu, Mycosystema 32: 321, 2004.
Description on the natural substrate: Colonies effuse, pale brown, superficial. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae with thin and smooth wall, 2–3.5 µm diam. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, solitary or 2–4 in small groups, reduced to sessile phialides or occasionally composed of a basal cell incorporated into hyphae and a phialide, erect, straight. Conidiogenous cells discrete, determinate, ampulliform or occasionally lageniform, 17.5–20 µm long, concolorous, medium brown to brown, thin- and smooth-walled, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters subglobose, globose, bulbous, 6–8 µm long and 7–8.5 µm wide; collarettes cylindrical, 12.5–15 µm long and 2.5–3 µm wide, smooth, concolorous with venter; ratio of mean lengths of collarette and venter = 2:1. Conidia endogenous, extruded in short or long and loose chains, cylindrical, 11–15 × 2.5 µm, both ends truncated except for the apical conidium which is obtuse at the apex, hyaline, 1-septate, smooth- and thin-walled; mean conidial length/width ratio = 5.2:1. Teleomorph: Unknown.
Material examined: China, Kunming, Yunnan Province, on dead leaves of undetermined host, 24 November 1995, Wenping Wu, Holotype Wu952.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China (Wu 2004).
Description and illustration: Wu (2004).
Notes: Of the many species published under Chalara s. lat., C. curvata, C. dennisii, C. fusidioides and C. rhynchophiala resemble N. yunnanensis in producing sessile phialides arising from superficial hyphae, globose venter and cylindrical collarettes, and cylindrical conidia (Nag Raj and Kendrick 1975; Kirk 1986; McKenzie et al. 2002; Wu 2004). Compared with N. yunnanensis, Chalara dennisii (conidia 4–6 × 0.8–1.2 µm) and C. fusidioides (4.5–12 × 1.5–3.5 µm) produce aseptate conidia in smaller sizes; C. curvata has small-sized conidiogenous cells and uniseptate conidia in smaller size; C. rhynchophiala has longer conidiogenous cells and conidia (12–23 × 2.5–3.5 µm). In addition, the conidiogenous cells (35–50 µm long), collarette 28–42 × 3–3.5 µm) and conidia (22–30 × 2.5–3 µm) of C. curvata are longer. In (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976).
Other known Chalara species with septate conidia
Except for the above accepted species, the following species under Chalara s. lat. are also with septate conidia and most likely also belong to the genus Nagrajchalara (Nag Raj and Kendrick 1975; Gadgil and Dick 1999; McKenzie et al. 2002). However, no living strain or DNA sequence was available for molecular phylogenetic analysis, thus their phylogenetic relationship with other chalara-like fungi within Leotiomycetes remains to be studied in future.
Chalara alabamensis Morgan-Jones & E.G. Ingram, Mycotaxon 4(2): 489, 1976.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri.
Geographical distribution: USA (Morgan-Jones and Ingram 1976).
Description and illustration: Morgan-Jones and Ingram (1976).
Chalara aotearoae Nag Raj & S. Hughes, N.Z. Jl Bot. 12(1): 120, 1974.
Ecology/substrate/host: Saprobe on dead bark of Aristotelia serrata.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Hughes (1974) and Nag Raj and Kendrick (1975).
Notes: Chalara aotearoae is characterized by 2- to many-septate conidiophores (65–280 × 6.5–8 µm) with a main stipe and a terminal phialide (59–69 µm long), subcylindrical venter (22–33 × 8–10 µm), cylindrical collarettes (31–42 × 5–6.5 µm), and hyaline, uniseptate and cylindrical conidia (13–18 × 3.5–5 µm) with rounded apex and truncated base, bearing minute marginal frills.
Chalara bicolor S. Hughes, in Nag Raj & Hughes, N.Z. Jl Bot. 12: 122, 1974.
Ecology/substrate/host: Saprobe on decaying wood of Podocarpus spicatus.
Geographical distribution: New Zealand (Nag Raj and Hughes 1974; Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Hughes (1974) and Nag Raj and Kendrick (1975).
Notes: Chalara bicolor is characterized by up to 12-septate conidiophores (up to 170 µm long) with a main stipe and a terminal phialide (132–165 µm long) with rough wall, ellipsoidal to subcylindrical venter (50–55 × 14.4–16.2 µm), cylindrical and rough-walled collarettes (7–9 µm wide), and hyaline, 7-septate and cylindrical conidia (50–66 × 5.9–6.6 µm) with rounded apex and truncated base. It is similar to C. inflatipes and C. insignis but differs in septation and size of conidia.
Chalara cibotii (Plunkett) Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 101, 1975.
≡ Excioconidium cibotti Plunkett (as ‘cibotti’), in Stevens, Bulletin Of the Bernice P. Bishop Museum, Honolulu, Hawaii 19: 156, 1925.
Ecology/substrate/host: Saprobe on Cibotium chamissoi.
Geographical distribution: USA (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara cibotti is special in processing septate conidiophores with subcylindrical phialides, gradual transition from venter to collarette, and hyaline, 7-septate, cuneiform or obovoid conidia with broad, rounded apex, and a narrow, truncate base bearing marginal frill. It can easily be distinguished from other species by its 7-septate conidia in cuneiform or obovoid shape.
Chalara cladii M.B. Ellis, Mycol. Pap. 79: 20, 1961.
≡ Chaetochalara cladii B. Sutton & Piroz., Trans. Br. mycol. Soc. 48: 352, 1965.
Ecology/substrate/host: Saprobe on decaying leaves of Cladium mariscus.
Geographical distribution: UK (Ellis 1961; Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara cladii is characterized by forming septate and dark brown setae, sessile, subhyaline to pale brown and subcylindrical conidiogenous cells (30–45 × 4.5–9 µm), and hyaline, 1-septate cylindrical conidia (10–18 × 4–5 µm) with rounded apex and truncated base, bearing marginal frills.
Chalara curvata Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 105, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Dracophyllum traversii.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara curvata differs from other species by sessile conidiogenous cells with very shorter and rough-walled venter and longer collarettes (ratio of collarette and venter = 5.3:1), and 1-septate, cylindrical and slightly curved conidia.
Chalara connari (Bat. & Peres) Santo & Dianese, not published name.
≡ Sporoschisma connari Bat. & Peres, Publicações Inst. Micol Recife 298: 33, 1960.
Ecology/substrate/host: Saprobe on decaying leaves of Salacia crassifolia.
Geographical distribution: Brazil (Batista et al. 1960).
Description and illustration: Batista et al. (1960).
Chalara dictyoseptata Nag Raj & S. Hughes, N.Z. Jl Bot. 12: 128, 1974.
Ecology/substrate/host: Saprobe on decaying stem of Rhipogonum scandens.
Geographical distribution: New Zealand (Nag Raj and Hughes 1974; Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Hughes (1974) and Nag Raj and Kendrick (1975).
Notes: Chalara dictyochaeta is the only known Chalara species with dictyospore.
Chalara distans McKenzie, Mycotaxon 46: 292, 1993.
Ecology/substrate/host: Saprobe on decaying leaves of Dracophyllum arboreum.
Geographical distribution: New Zealand (McKenzie 1993).
Description and illustration: McKenzie (1993).
Chalara dracophylli McKenzie, Mycotaxon 61: 306, 1997.
≡ Chalara australis McKenzie, Mycotaxon 46: 291 (1993).
Ecology/substrate/host: Saprobe on decaying leaves of Dracophyllum arboreum.
Geographical distribution: New Zealand (McKenzie 1993, 1997).
Description and illustration: McKenzie (1993, 1997).
Notes: Chalara dracophylli resembles C. alabamensis and Cylindroephalum hughesii but differs by larger conidiophores and conidia.
Chalara emodensis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera: 115, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Quercus incana.
Geographical distribution: Pakistan (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara emodensis is characterized by sessile conidiogenous cells with rough wall and in smaller size (phialides 16–28 µm long, venters 6.5–11 × 7–11 µm, collarettes 15–18 × 3–4 µm), and hyaline, uniseptate and cylindrical conidia (11–15 × 2.5–3 µm) with rounded or blunt ends and no basal frill.
Chalara germanica Nag Raj & W.B. Kendr., Can. J. Bot. 49(2): 2121, 1971.
Ecology/substrate/host: Saprobe on decaying twigs of unidentified tree.
Geographical distribution: Germany (Nag Raj and Kendrick 1971, 1975).
Description and illustration: Nag Raj and Kendrick (1971, 1975).
Notes: Both aseptate and uniseptate conidia were reported from the holotype specimen of this species.
Chalara ginkgonis Ferd. & Winge, Bot. Tidsskr. 28: 256, 1907.
Ecology/substrate/host: Saprobe on decaying leaves of Ginkgo biloba.
Geographical distribution: Denmark (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Both aseptate and uniseptate conidia were reported in the original description.
Chalara gracilis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 120, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Knightia excelsa.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara grandispora Matsush., Matsush. Mycol. Mem. 7: 46, 1993.
Ecology/substrate/host: Saprobe on decaying material of palm.
Geographical distribution: Peru (Matsushima 1993).
Description and illustration: Matsushima (1993).
Notes: Chalara grandispora (conidia 3-septate, 30–50 × 10–12.5 µm) is similar to C. unicolor (conidia 3-septate, 18–42 × 5–8 µm), but differs by longer and wider conidia.
Chalara inaequalis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 122, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara inaequalis differs from other existing species by 1-septate conidia (9.5–18 × 2–2.5 µm with a longer basal cell and a shorter upper cell.
Chalara laevis (B. Sutton & Hodges) P.M. Kirk, in Kirk & Spooner, Kew Bull. 38(4): 580, 1984.
≡ Chaetochalara laevis B. Sutton & Hodges, Nova Hedwigia 27(1–2): 343, 1976.
Ecology/substrate/host: Saprobe on decaying leaves of Astrocaryum and Eucalyptus.
Geographical distribution: Brazil and Malaysia (Sutton and Hodges 1976; Monteiro et al. 2019).
Description and illustration: Sutton and Hodges (1976) and Monteiro et al. (2019).
Chalara magnispora Matsush., Matsush. Mycol. Mem. 7: 46, 1993.
Ecology/substrate/host: Saprobe on decaying petiole of palm.
Geographical distribution: Peru (Matsushima 1993).
Description and illustration: Matsushima (1993).
Notes: Chalara magnispora differs from other known species by its 3–4-septate conidia in larger size (25–56 × 7.5–10.5 µm).
Chalara nigricollis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 125, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Cyperus longus.
Geographical distribution: UK (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara paramontellica McKenzie, Fungal Diversity 11: 135, 2002.
Ecology/substrate/host: Saprobe on decaying twigs of Thea sp.
Geographical distribution: Mauritius (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara prolifera Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 129, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara prolifera is characterized by the short conidiophores with multiple percurrent proliferations, and uniseptate and broader conidia (10–16 × 3–4.5 µm).
Chalara pteridina Syd., Annls. Mycol. 10(5): 450, 1912.
Ecology/substrate/host: Saprobe on Bidens, Ptridium, Senecio, Tanacetum, etc.
Geographical distribution: Austria, Germany, Luxembourg, New Zealand and UK (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara ramosa (Nag Raj & W.B. Kendr.) P.M. Kirk, in Kirk & Spooner, Kew Bull. 38(4): 580, 1984.
≡ Chaetochalara ramosa Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 154, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Quercus densiflora.
Geographical distribution: USA (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara rhynchophialis Nag Raj & W.B. Kendr., Monograph Chalara Allied Genera (Waterloo): 132, 1975.
Ecology/substrate/host: Saprobe on Aesculus and Dracophyllum.
Geographical distribution: New Zealand and UK (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara rhynchophialis differs from other species by sessile conidiogenous cells with bulbous venter and a longer collarettes, and uniseptate conidia.
Chalara rostrata Nag Raj & W.B. Kendr., Monogr Chalara Allied Genera (Waterloo): 132, 1975.
Ecology/substrate/host: Saprobe on Geostachys rupestris.
Geographical distribution: Malaysia and North America (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara scabrida Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 133, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara scabrida (conidia 15–19 × 2.5–3 µm) is similar to C. curvata (conidia 22–30 × 2.5–3 µm) in sessile conidiogenous cells with shorter and rough-walled venter, and 1-septate conidia, but differs in straight and smaller conidia.
Chalara spiralis Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 135, 1975.
Ecology/substrate/host: Saprobe on Fagus sp.
Geographical distribution: UK (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara stipitata Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 135, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Agathis australis and Podocarpus thallii.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara transkelensis Morgan-Jones, J.B. Sinclair & Eicker, S. Afr. J. Bot. 58(3): 147, 1992.
Ecology/substrate/host: Saprobe on decaying leaves of Nothofagus solandri.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara transkelensis is similar to C. alabamensis, Cylindrocephalum hughesii and N. angionacea in producing reduced conidiophores with a few basal cells and a long terminal phialides, and hyaline, uniseptate and cylindrical conidia (12–17 × 2.5–3 µm), but differs from them in size of phialides and conidia.
Chalara tubifera Nag Raj & W.B. Kendr., Monograph Chalara Allied Genera (Waterloo): 137, 1975.
Ecology/substrate/host: Saprobe on decaying leaves of Knightia excelsa.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Chalara urecolata Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 139, 1975.
Ecology/substrate/host: Saprobe on Rhopalostylis sapida, Rumex sp., and other plants.
Geographical distribution: Ireland, New Zealand, North America and UK (Nag Raj and Kendrick 1975; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975).
Chaetochalara proteae Joanne E. Taylor & M.E. Palm, Mycotaxon 78: 451, 2001.
Ecology/substrate/host: Saprobe on decaying leaves of Protea.
Geographical distribution: USA (Taylor et al. 2001).
Description and illustration: Taylor et al. (2001).
Notes: Chaetochalara proteae differs from other species by broader conidia (10.5–12 × 6.5–7 µm).
Cylindrocephalum Bonord., Handb. Allgem. mykol. (Stuttgart): 103, 1851.
Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, smooth, septate and branched hyphae. Anamorph: Stroma absent or present, composing of dark brown and irregular cells. Setae absent or present, cylindrical to clavate, septate, brown, fertile or sterile. Conidiophores arising from superficial hyphae or aggregated cells, scattered or aggregated at the base, erect, straight or slightly curved, simple, reduced or well-developed, composed of a basal stalk and a terminal phialide, obclavate, lageniform, subcylindrical, pale brown to dark brown, septate, smooth or verruculose, terminating in a phialidic conidiogenous cell. Conidiogenous cells lageniform, obclavate, urceolate or subcylindrical, pale brown to dark brown, smooth or verruculose, composed of a venter and a collarette; transition from venter to collarette gradual or abrupt. Conidia endogenous, extruded in in short or long and loosing chains, cylindrical, straight, hyaline, septate, smooth, with rounded or truncated bases, often provided with basal marginal frill or rarely fringes of wall material, slightly rounded at the apex. Teleomorph: Unknown.
Type species: Cylindrocephalum aureum (Corda) Bonord.
Ecology/substrate/host: Saprobe on dead branches and rotten wood.
Geographical distribution: Widely distributed.
Notes: In the phylogenetical analyses, five chalara-like fungi with septate conidia, similar to species of Nagrajchalara (Crous et al. 2021), were clustered together as a strongly supported monophyletic clade distinct from Nagrajchalara clade (Figs. 1, 2, 3, 8, 40). A new generic name was needed to accommodate them. Based on literature review and supported by molecular phylogeny and pure culture study, the generic name Cylindrocephalum was reinstated for these five chalara-like fungi. Under the genus Chalara s. lat., six generic synonyms, Cylindrocephalum, Thielaviopsis, Stilbochalara, Chalaropsis, Excioconidium and Hugehsiella were included by Nag Raj and Kendrick (1975). Among them, the type species of Thielaviopsis, Stilbochalara, Chalaropsis, and Hughesiella were congeneric and now reclassified as Thielaviopsis in Ceratocystidaceae (Microascales). This made Cylindrocephalum (1851) and Excioconidium (1925) as the only potential choices. Both genera were with typical chalara-like phialides consisting of a venter and a collarettes with deeply seated conidiogenous loci, and septate conidia (Hughes 1958; Nag Raj and Kendrick 1975). Excioconidium was known with only the type species E. cibotti, and no living strain or molecular sequence data were available, thus its phylogenetic relationship with other chalara-like fungi remains to be studied. Morphologically Excioconidium cibotti produces 7-septate conidia with thick cell wall, which differs from those in those five species. The type species of Cylindrocephalum, C. aureum, was represented with two strains in the Chalara aurea clade. Based on these analyses, the genus Cylindrocephalum is reinstated as the generic name for these six species.
Morphologically these five species were hardly distinguishable from Neochalara and Nagrajchalara. However, the living cultures of all these five species of Cylindrocephalum produced orange-colored pigment diffused into agar plate on PDA (Figs. 110, 111), which could be used to distinguish the genus from Nagrajchalara. The living cultures of many species of Nagrajchalara were also studied and they usually did not produce orange-colored pigment on PDA (Figs. 41, 42, 43, 44, 45, 46, 47).

Colony of Cylindrocephalum species on PDA in 4 weeks. a, b Cylindrocephalum clavatisetosum (ex-type strain 53375). c–h C. aureum (c, d 50498; e, f 77468; g, h 42820). i, j C. hughesii (i, j 54360; k, l 53396). m–p C. kendrickii (m, n 54252; o, p 43817)

Colony of Cylindrocephalum species on PDA in 4 weeks at 25 degrees. a–f Cylindrocephalum aureum (a, b CBS729.69; c, d 42820; e, f 44387). g, h C. zhejiangense (76401). i–l C. hughesii (43813). m–p C. kendrickii (m, n 43817; o, p CBS 490.77)
Based on the phylogenetic analyses (Figs. 2, 3, 8), a total of five species were accepted in the genus. Morphologically they were variable in setae, conidiophores, phialides and conidia. Conidiophores in this genus are well-developed with multiseptate basal stalk or reduced to one-celled basal stalk with a terminal phialide. Conidia of these species are always cylindrical or short-cylindrical, with obtuse apex and truncated or obtuse base. Basal frills of conidia are often seen from these species (Nag Raj and Kendrick 1975).
Cylindrocephalum aureum (Corda) Bonord., Handb. Allgem. Mykol. (Stuttgart): 103, 1851. Figure 112.

Cylindrocephalum aureum (Wu16169). a–p, t–y Conidiophores and phialidic conidiogenous cells. q–s Conidia. Scale bar: 10 µm for a–p; 5 µm for q–y
≡ Menispora aurea Corda, Icon. fung. (Prague) 2: 12, 1838.
≡ Chalara aurea (Corda) S. Hughes, Can. J. Bot. 36: 747, 1958.
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin– or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Stroma absent or poorly developed, composing of brown and irregular cells. Setae absent. Conidiophores arising from superficial hyphae or aggregated cells, scattered or 2–3 aggregated at the base, erect, straight or slightly curved, composed of a basal stalk and a terminal phialide, simple, obclavate, subcylindrical, 60–90 µm long, 4–6 µm wide, medium to dark brown, 2–6-septate, smooth; basal cells slightly swollen, dark brown; terminating in a phialide. Conidiogenous cells integrated, terminal, phialidic, obclavate, lageniform, 45–55 µm longer, versicolorous, medium brown, darker in the lower part of collarettes, smooth-walled, composed of a venter and a collarette; transition from venter to collarette abrupt; venters cylindrical, 18–23 µm long, 5.5–6.5 µm at widest part, brown; collarettes cylindrical, 30–38 µm long, 3.5–4 µm wide, brown, slightly darker in the lower part; ratio of mean lengths of collartte and venter = 1.7:1. Conidia endogenous, extruded in short or long chains, cylindrical, 9–14 × 2–2.5 µm, straight, base truncated and without basal marginal frill, apex rounded or flattened, hyaline, aseptate, smooth; mean conidium length/width ratio = 5.1:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, grey, pale brown, reverse fresh yellow brown, soil brown to brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 16 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangxi Province, Damingshan, on dead branches of unidentified plant, 19 December 1997, Wenping Wu, Wu1414b; Guangxi Province, Damingshan, on dead bark of unidentified plant, 19 December 1997, Wenping Wu, Wu1442b; Guangxi Province, Damingshan, on rotten wood of unidentified plant, 18 December 1997, Wenping Wu, Wu1363c; Zhejiang Province, Huai An County, Qiandaohu, on dead fruit of unidentified tree, 18 October 2018, Wenping Wu, Wu16169 and Wu16031. Living strains: CGMCC3.23355 (= 42820, from Wu1363c), CGMCC3.23434 (= 77468, from Wu16169), 44387 (from Wu1414b), 77476 (from Wu16031b), 77404 (from Wu16031a), 50498 (= CBS729.69).
Ecology/substrate/host: Saprobe on dead leaves of Aesculus, Betula, Carpinus, Ilex and Quercus.
Geographical distribution: Austria, China, Czechoslovakia, North America and UK (Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Ellis and Ellis 1987; Bates et al. 2018).
Description and illustration: Hughes (1958), Nag Raj and Kendrick (1975), Holubová-Jechová (1984), and Ellis and Ellis (1987).
Notes: Cylindrocephalum aureum, the type species, is characterized by absence of setae, well-developed conidiophores consisting of a 1–6-septate basal stalk and a terminal phialide, lageniform or obclavate phialides, cylindrical to subellipsoidal venter, and 1-septate and cylindrical conidia with truncated base and rounded or obtuse apex. It superficially resembles Chalara aotearoae, C. inaequalis, C. rostrata, C. tubifera, C. urceolata and Nagrajchalara acuaria but differs in shorter and narrower conidia (Nag Raj and Kendrick 1975). In addition, venters of C. inaequalis, C. tubifera and C. urceolata are urceolate and with the widest right below transition from venter to collarette; collarettes of C. aotearoae, C. rostrata, and C. urceolata and N. acuaria are concolorous; and conidiophores of C. stipitata and C. urceolata are much longer than those of C. aureum (Nag Raj and Kendrick 1975). Compared with the description given by Nag Raj and Kendrick (1975), conidiophores and phialides from the Chinese collections were less variable in length and septation, and conidia are slightly shorter.
ITS sequences generated from 5 studied strains were almost identical (with 1 bp difference). In the CBS fungal collection database, three strains identified as Chalara aurea were not conspecific and their ITS sequences were with low similarity (< 93%). The ITS sequence from the strain CBS 729.69, isolated from Polyporus picipes besides Hypomyces aurantius from Netherlands by W. Gams, was identical to those from the Chinese specimens. The ITS sequence from the CBS633.75 was with 23 bp differences from the CBS 729.69, which might represent another species.
Cylindrocephalum clavatisetosum W.P. Wu & Y.Z. Diao, sp. nov., Figs. 113, 114, MycoBank MB845255.

Cylindrocephalum clavatisetosum (Wu11011, holotype). a–e Conidiophores in two layers with both longer sterile or fertile setae and short conidiophores. f Sterile setae with pale brown and multi-septate upper part. g Phialidic conidiogenous cell developed endogenously inside the setiform conidiophore. h Upper part of fertile setiform conidiophore bearing phialide. g–l Conidia. Scale bar: 20 µm for a–c, 10 µm for d–f, 5 µm for g, h

Cylindrocephalum clavatisetosum (Wu11011, holotype). a–d Conidiophores and phialidic conidiogenous cells. e–g Terminal phialidic conidiogenous cells at the apex of fertile setae. h–j Conidia. Scale bar: 5 µm
Etymology: Refers to its clavate shaped setae.
Typification: China, Hunan province, Zhangjiajie, on dead leaves of unidentified tree, 15 October 2010, Wenping Wu, Holotype HMAS 352184 (= Wu11011), ex-type strain CGMCC3.23445 (= NN53375).
Description on the natural substrate: Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or thick-walled, 2.5–5 µm wide. Anamorph: Stroma absent or poorly developed, composed of dark brown and irregularly shaped cells. Setae solitary, scattered among conidiophores, erect, straight or flexuous, long clavate, cylindrical, 150–300 μm long, 4–5.5 μm wide at the basal part, 6–8 μm wide at the upper part, smooth-walled, dark brown at the basal part, becoming paler towards the upper part, 10–13-septate; basal cells flattened, up to 15 μm wide; apex rounded, sterile, or fertile with a phialide. Conidiophores arising from superficial hyphae or aggregated cells, scattered or 2–4 aggregated at the base, erect, straight or flexuous, simple, lageniform, obclavate, (65–)95–122 µm long, 4–5.5 µm wide at the lower part, concolorous, medium to dark brown, 3–6 septate, smooth-walled; basal cells slightly swollen and up to 15 µm wide; terminated into a phialide. Conidiogenous cells integrated, terminal, phialidic, urceolate, lageniform, 60–68 µm long, concolorous, medium to dark brown, composed of a venter and a collarette; transition from venter to collarette abrupt; venters urceolate, narrowly obconic, subcylindrical, 15–22 × 6.5–8.5 µm, widest at the transition from venter to collarette; collarettes cylindrical, 30–47 × 3–4.5 µm, concolorous with venter; ratio of mean lengths of collarette and venter = 2.1:1. Conidia endogenous, extruded in short or long chains, cylindrical, (12–)14–16 × 2–3 µm, straight, flattened or obtuse at both ends, without basal frill, hyaline, 1-septate, smooth; mean conidium length/width ratio = 6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, grey to light brown, reverse dark brown with a yellow brown margin, with yellow pigment diffused into agar, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Guangdong Province, Guangzhou, Baiyunshan, on dead leaves of Ficus sp., 5 March 2012, Wenping Wu, Wu12051; Guangdong Province, Zhaoqing, Dinghushan, on dead leaves of unidentified tree, 3 March 2015, Wenping Wu, Wu12083 and 12084; Sichuan Province, Qingchengshan, on unidentified plant, 24 August 2002, Wenping Wu, Wu5550c. Living strains: 54210 (from Wu12083) and 54219 (Wu12084).
Ecology/substrate/host: Saprobe on dead leaves of Ficus sp., Rhopalostylis sapida, Rumex sp. and other plants.
Geographical distribution: China.
Notes: Among the existing Chalara s. lat. species with 1-septate conidia, Chalara paramontellica, C. tubifera and C. urceolata are morphologically similar with Cylindrocephalum clavatisetosum, but differs in lacking setae (Matsushima 1971; Nag Raj and Kendrick 1975; McKenzie et al. 2002). In addition, compared with C. clavatisetosum, Chalara tubifera has versicolored phialides with darker collarette than venter, and its conidia are with basal frill; Chalara urceolata has wider venter (8–13 μm wide). Cylindrocephalum clavatisetosum also resembles Chaetochalara mutabilis in producing sterile or fertile setae and 1-septate conidia without basal frill, but differs in producing clavate-shaped setae and multiseptate conidiophores in the former species (Silva et al. 2015).
Cylindrocephalum hughesii (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 115, MycoBank MB845287.

Cylindrocephalum hughesii (Wu16061). a–d Conidiophores and conidiogenous cells. e–j Conidia. Scale bar: 10 µm for a, 5 µm for b–j
≡ Chalara hughesii Nag Raj & W.B. Kendr., N.Z. Jl Bot. 12(1): 118, 1974.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores arising from superficial hyphae, solitary or 2–3 in groups, reduced to a 1–2-celled basal stalk and a phialide, erect, straight or slightly curved, lageniform, obclavate, 35–55 µm, 4–5 µm wide in the basal stalk, pale to medium brown, smooth. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 37–48 µm long, concolorous, medium brown, smooth, consisted of a venter and a collarette; transition from venter to collarette abrupt; venters conical to subcylindrical, 12–15 µm long and 5.5–7.5 µm wide; collarettes cylindrical, 20–25 µm long, 2.5–3 µm wide, concolorous; ratio of mean lengths of collarette and venter = 1.7:1. Conidia endogenous, extruded in short chains, cylindrical, 14–16 × 2–2.5 µm, both ends rounded or flattened, without basal marginal frill, hyaline, aseptate, smooth- and thin-walled; mean conidium length/width ratio = 6.7:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, grey to brown, reverse soil brown to brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 15 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangdong Province, Guangzhou, South China Botanical Garden, On dead leaves of unidentified tree, 4 March 2015, Wenping Wu, Wu12263; Guangxi Province, Nanning, on dead leaves of Eucalyptus sp., 3 January 1998, Wenping Wu, Wu1612c; Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 13 September 2004, Wenping Wu, Wu8162b; Hunan Province, Zhangjiajie, on dead leaves of unidentified tree, 15 October 2010, Wenping Wu, Wu11019; Ningxia Province, Shiwandashan, on dead branches of ?Eucalyptus sp., 31 December 1997, Wenping Wu, Wu1593b; Zhejiang Province, Hua An County, Qiandaohu, on dead leaves of unidentified tree, 18 October 2018, Wenping Wu, Wu16061 and Wu16062. Living strains: CGMCC3.23388 (= 53396, from Wu11019), CGMCC3.23393 (= 54360, from Wu12263), 43813 (from Wu1612c), 76351 (from Wu16061), 76352 (from Wu16062a), 76314 (from Wu16061), 76315 (from 16062).
Ecology/substrate/host: Saprobe on dead leaves of Arundinaria, Dipterocarpus, Eucalyptus, Hoheria, Ilex, Laurus, Ocotea, Pinus, Quercus, Viburnum. and other plants.
Geographical distribution: Argentina, Brazil, Canada, China, Japan, Kenya, Malaysia, New Zealand, Sri Lanka, UK and USA (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; Gamundi et al. 1977; Matsushima 1980; Holubová-Jechová 1984).
Description and illustration: Nag Raj and Kendrick (1975), Morgan-Jones and Ingram (1976), Gamundi et al. (1977), Holubová-Jechová (1984), and Wu (2004).
Notes: Cylindrocephalum hughesii resembles Chalara alabamensis, but has abrupt transition from venter to collarette, shorter conidiogenous cells and slightly narrower conidia (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976). Cylindrocephalum hughesii also resembles C. fungorum, but the latter species has aseptate conidia (Nag Raj and Kendrick 1975).
ITS sequences of Cylindrocephalum hughesii were generated from seven studied strains, and they are almost identical (1 bp difference). Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 54360 included Chalara hughesii (KY853434, 98.5% identity), C. aurea (MH860959, 95% identity) and many unnamed fungi of Leotiomycetes.
Cylindrocephalum kendrickii (Nag Raj) W.P. Wu & Y.Z. Diao, comb. nov., Fig. 116, MycoBank MB845288.

Cylindrocephalum kendrickii (Wu12166). a Conidiophores in clusters on basal stroma. b–g Conidiophores and conidiogenous cells. h, i Conidia. Scale bar: 10 µm for a, 5 µm for b–i
≡ Chalara kendrickii Nag Raj, in Nag Raj & Kendrick, Monogr. Chalara Allied Genera (Waterloo): 123, 1975.
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2.5–3 µm wide. Anamorph: Stroma present, dark brown, composed of dark brown and irregularly shaped cells. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, up to 11 in groups, reduced to a 1–2-celled basal stalk and a terminal phialide, obclavate, lageniform, 50–70 µm long, basal cell 5–8 × 4–6 µm, versicolorous, medium brown to brown, darker in collartte, smooth, terminating into a phialide. Conidiogenous cells integrated, terminal, phialidic, erect, straight or slightly curved, lageniform, obclavate, 45–60 µm long, medium brown to dark brown, smooth; transition from venter to collarette abrupt; venters cylindrical, subellipsoidal, 15–23 × 5–7 µm, medium brown; collarettes cylindrical, 23–32 × 3–3.7 µm, dark brown; ratio of mean lengths of collarette and venter = 1.6:1. Conidia endogenous, extruded in readily seceding short chains, cylindrical, 13–15 × 2–2.5 µm, blunt or flattened at both ends, without basal marginal frills, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 6.2:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded, aerial mycelium poorly developed, white, reverse fresh yellow brown, becoming paler towards the margin, with yellow pigment diffused into agar, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Guangdong Province, Zhaoqing, Dinghushan, on dead leaves of unidentified tree, 3 March 2012, Wenping Wu, Wu12166; Guangdong Province, Guangzhou, South China Botanical Garden, on dead leaves of Eucalyptus sp., 4 March 2012, Wenping Wu, Wu12302; Guangxi Province, Damingshan, on rotten wood, 20 December 1997, Wenping Wu, Wu1463; Ningxia Province, Jingyuan County, Liupan Mountain, Liantianxia, on dead bark of unidentified plant, 25 August 1997, Wenping Wu, Wu1024c; Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead fruit of Fagaceae, 26 August 2022, Wenping Wu (Wu18138, Wu18139). Living strains: CGMCC3.23363 (= NN43817, from Wu1024c), CGMCC3.23392 (= NN54252, from Wu12166), 12302, 78715 and 78716 (from Wu18138), 78717 and 78718 (from Wu18139), and CBS490.77.
Ecology/substrate/host: Saprobe on dead leaves of Baissea, Eucalyptus and Rubus.
Geographical distribution: China, Sierra Leone and UK (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Cylindrocephalum kendrickii was originally described as Chalara kendrickii on Baissea and Rubus from Sierra Leone and UK. It was characterized by barely differentiated conidiophores and hyaline, 1-septate conidia with distinct frills at base. Cylindrocephalum kendrickii superficially resembles N. angionacea and N. agathidis. However, N. agathidis has much longer and slightly narrower conidia (17–24 × 2.5–3 µm); while N. angionacea differs from N. kendrickii by its more bulbous venter, narrower collarette and longer conidia (13–18.5 × 3–3.5 µm). Two specimens (Wu18138 and Wu18139) were recently collected on decaying fruit of Fagaceae from Hangzhou, and four living strains were successfully obtained from them by using single spore isolation method. The obtained ITS sequences from these strains were identical to C. kendrickii from other strains generated in this study. However, the fungus from these two specimens were with shorter conidiophores (40–43 µm long) and conidiogenous cells (32–37 µm long, venter 10–12 × 6.3–6.8 µm, collarette 21–25 × 2.5–2.6 µm), but longer conidia (15.5–17.5 × 2–2.3 µm).
ITS sequences generated from eight studied strains of Cylindrocephalum kendrickii were identical, and they were Also identical to CBS490.77 under Chalara kendrickii. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 43817 includes Cylindrocephalum hughesii (KY853434, 100% identity), Chalara sp. (FR667232, 98% identity), Chalara sp. (KX096663, 98% identity), Chalara sp. (MK432782, 99% identity), and many unnamed fungi of Leotiomycetes.
Cylindrocephalum zhejiangense W.P. Wu & Y.Z. Diao, sp. nov., Figs. 117, 118a–e, k–n, u–w, MycoBank MB845256.

Cylindrocephalum zhejiangense (Wu16134, holotype). a–p Conidiophores and conidia. q–u Conidia. Scale bar: 10 µm for a–k, 5 µm for l–u

Cylindrocephalum zhejiangense (a–e, k–n, u–w from Wu16134) and C. hughesii (f–j, o–t, x–ac from Wu12263). a–t Conidiophores and conidiogenous cells. u–ac Conidia. Scale bar: 10 µm for a–j, 5 µm for k–ac
Etymology: Refers to the locality Huai An County, Zhejiang Province, where the fungus was discovered.
Typification: China, Zhejiang Province, Hangzhou, Hangzhou Botanical Garden, on dead fronds of fern, 18 October 2018, Wenping Wu, Holotype HMAS 352185 (= Wu16134); ex-type strain CGMCC3.23422 (= NN76401).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3.5 µm wide. Anamorph: Stroma absent or poorly developed, composed of dark brown, irregular-shaped and thick-walled cells. Setae absent. Conidiophores formed from superficial hyphae or aggregated cells, erect, straight of slightly curved, composing of a 2–3-celled basal stalk and a terminal phialide, obclavate, lageniform, 74–90 µm long, 4–4.5 µm wide at the basal part, 1–2 septate, medium brown, becoming paler towards the upper part, smooth. Conidiogenous cells integrated, terminate, phialidic, erect, straight or slightly curved, lageniform, obclavate, 60–74 µm long, concolorous, medium brown, becoming slightly paler towards the apex, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters lageniform, urceolate, subellipsoidal, 24–26 µm long, 6–7 µm wide at the widest part right below the transition, 4–4.5 μm wide at the base, medium brown; collarettes cylindrical, 44–47 × 3.5–3.7 µm, concolorous, becoming slightly paler towards the opening, widest at the upper part; ratio of mean lengths of collarette and venter = 1.8:1. Conidia endogenous, extruded in short chains, cylindrical, 15–18 × 3–3.6 µm, base truncated or flattened and without frills, apex rounded or flattened, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 5:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, brown, reverse concolorous, with brown pigment diffused into agar, sterile, up to 4 mm on PDA at 25 °C in 4 weeks.
Other living strain: 76402 (from holotype Wu16134).
Ecology/substrate/host: Saprobe on dead leaves of undetermined tree.
Geographical distribution: China.
Notes: Cylindrocephalum zhejiangense morphologically resembles C. aurea, Chalara inaequalis (conidiophores 1–5-septate, 70–100 μm long; phialides 45–65 μm long, venter subcylindrical to ellipsoidal and 17–25 × 5.5–8 μm, collarette 29–47 × 2.5–3.5 μm; conidia 9.5–18 × 2–2.5 μm) and C. tubifera (conidiophores few to many septate, 65–220 μm long; phialides 49–65 μm long, venter narrowly obconic and 19–26 × 6–7 μm, collarette 28–40 × 2.5–3 μm; conidia 11–19 × 2–2.5 μm), all producing short but well-differentiated conidiophores and 1-septate conidia. Cylindrocephalum zhejiangense differs from them by longer collarettes. In addition, the phialides of C. inaequalis and C. tubifera are versicolorous and with darker collarettes (Nag Raj and Kendrick 1975).
Neochalara Crous, in Crous et al., Persoonia 47: 217, 2021.
Anamorph: Conidiophores arising from bulbous base, or as extension of hyphae, frequently aggregated, cylindrical, brown, erect, thick-walled, unbranched, septate, with terminal conidiogenous cells. Conidiogenous cells phialidic, brown, smooth-walled, consisting of a venter and a collarette; transition from venter to collarette gradual; venter subcylindrical brown; collarette cylindrical, pale brown. Conidia endogenous, extruded in long and unbranched chains, cylindrical, both ends truncated and with marginal frill, medially 1-septate, hyaline, guttulate, smooth-walled (Adapted from Crous et al. 2021). Teleomorph: Unknown.
Type species. Neochalara spiraeae Crous.
Ecology/substrate/host: Saprobes on Spiraea japonica (Rosaceae).
Geographical distribution: Netherland.
Description and illustration: Crous et al. (2021).
Notes: The recently established genus Neochalara is morphologically indistinguishable from Cylindrocephalum and Nagrajchalara, but phylogenetically distinct (Crous et al. 2021, 2022; Nag Raj and Kendrick 1975; McKenzie et al. 2002). In the phylogenetic analyses, the two known species N. lolae and N. spiraeae clustered together with the Chalara s. str. species producing aseptate conidia, but as a distinct lineage.
Neochalara lolae Crous, in Crous et al., Persoonia 48: 287, 2022.
Description on the natural substrate: Anamorph: Conidiophores solitary, cylindrical, 50–70 μm tall, 1–2-septate, medium brown, smooth-walled. Conidiogenous cells phialidic, medium brown, transition from venter to collarette gradual; venter 20–30 μm long, 4–6 μm wide; collarette cylindrical, 10–25 μm long, 3–4 μm wide, paler towards apex. Conidia endogenous, extruded in long and unbranched chains, cylindrical, (9–)12–16(–28) × (2.5–)3 μm, 1–3(–4)-septate, hyaline, guttulate, smooth-walled, guttulate, ends bluntly rounded (Adapted from Crous et al. 2022). Teleomorph: Unknown.
Ecology/substrate/host: Saprobes on stems of Pteridium aquilinum.
Geographical distribution: Netherland (Crous et al. 2022).
Description and illustration: Crous et al. (2022).
Notes: Neochalara lolae is a recently described species commonly found on the fronds of Pteridium aquilinum in the Netherlands (Crous et al. 2022). It was compared with Chalara pteridina, that also has septate conidia and occurs on the same host in Europe (Nag Raj and Kendrick 1975). It differs from the latter that has single or short conidial chains, conidiophores with percurrent rejuvenation, a wider venter (5–9.2 µm), longer collarettes (12–44(–50) × 3–4.5 µm), and mostly 3-septate conidia, (8–)12(–18) × (2–)2.5(–3) µm (Nag Raj and Kendrick 1975). Neochalara lolae is also similar to N. spiraeae, but is distinct in that the latter species has smaller, 1-septate conidia (Crous et al. 2021, 2022).
Neochalara spiraeae Crous, in Crous et al., Persoonia 47: 217, 2021.
Description on the natural substrate: Anamorph: Conidiophores cylindrical, 100–170 × 6–8 μm, 1–4-septate, brown, smooth-walled. Conidiogenous cells phialidic, brown, transition from venter to collarette gradual; venter 30–40 μm long, 7–10 μm wide, brown; collarette cylindrical, 30–50 μm long, 4–5 μm wide, pale brown. Conidia endogenous, extruded in long and unbranched chains, cylindrical, (11–)12–14(–16) × (3–)4(–4.5) μm, both ends truncate and with marginal frill, medially 1-septate, hyaline, guttulate, smooth-walled (Adapted from Crous et al. 2021). Teleomorph: Unknown.
Ecology/substrate/host: Saprobes on Spiraea japonica (Rosaceae).
Geographical distribution: Netherland (Crous et al. 2021).
Description and illustration: Crous et al. (2021).
Notes: Morphologically Neochalara spiraeae is distinct from other known Nagrajchalara and Chalara s. lat. species in having conidia with both ends being truncated (Crous et al. 2021).
Phaeoscypha Spooner, in Kirk & Spooner, Kew Bull. 38(4): 574, 1984.
Type species: Phaeoscypha cladii (Nag Raj & W.B. Kendr.) Spooner,
Ecology/substrate/host: Saprobe on dead plants.
Geographical distribution: South Africa and UK (Nag Raj and Kendrick 1975; Kirk and Spooner 1984).
Description and illustration: Ellis (1961, 1976), Sutton and Pirozynski (1965), Nag Raj and Kendrick (1975), and Kirk and Spooner (1984).
Notes: The genus Phaeoscypha was established with P. cladii as the type species, a fungus with both discomycete teleomorph and Chaetochalara anamorph (Nag Raj and Kendrick 1975; Kirk and Spooner 1984). The second species P. pteridiicola was described on fern in South Africa, but without chalara-like anamorph (Raitviír and Schneller 2007). Its assignment into Pezizellaceae was not based on the type species P. cladii, but the recently describe species P. pteridiicola (Ekanayaka et al. 2019).
Phaeoscypha cladii (Nag Raj & W.B. Kendr.) Spooner, in Kirk & Spooner, Kew Bull. 38(4): 574, 1984.
≡ Hyaloscypha cladii Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 183, 1975.
≡ Chaetochalara cladii B. Sutton & Piroz., Trans. Br. mycol. Soc. 48: 352, 1965.
≡ Chalara cladii M.B. Ellis, Mycol. Pap. 79: 21, 1984.
Ecology/substrate/host: Saprobe on dead plants including Cladium mariscus, Cortaderia sp. and Juncus spp.
Geographical distribution: UK (Ellis 1961, 1976; Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975; Kirk and Spooner 1984).
Description and illustration: Ellis (1961, 1976), Sutton and Pirozynski (1965), Nag Raj and Kendrick (1975), and Kirk and Spooner (1984).
Notes: The fungus was described with both anamorphs and teleomorph in literatures. No living strain or DNA sequence was available for molecular phylogenetic study, thus its affinity to other chalara-like fungi remains to be studied.
Rodwayella Spooner, in Dennis, Fungi Hebrid: 383, 1986.
Type species: Rodwayella sessilis (Rodway) Spooner, Biblthca Mycol. 116: 652, 1987.
Ecology/substrate/host: Saprobe on plants.
Geographical distribution: Australia (Spooner 1987).
Description and illustration: Spooner (1987).
Notes: The genus Rodwayella was established with R. sessilis as the type species (Spooner 1987). The other known species are R. citrinula, R. haematoidea and R. myricae (Dennis 1986; Spooner 1987). Among of them, the type species R. sessilis was reported to be associated with Chalara aff. aotearoae, but this observation was not from the type material (Baral 2002). The phylogenetic analyses based on the available ITS sequences showed that one specimen of the type species R. sessilis was closely related to Pezizellaceae, while the other species R. citrinula belonged to Hyaloscyphaceae (Clade 9 in Han et al. 2014).
Rodwayella sessilis (Rodway) Spooner, Biblthca Mycol. 116: 652, 1987.
≡ Helotium sessile Rodway, Pap. Proc. R. Soc. Tasm.: 103, 1925 (1924).
Ecology/substrate/host: Saprobe on plant.
Geographical distribution: Australia (Spooner 1987).
Description and illustration: Spooner (1987).
Tapesina Lambotte, Mém. Soc. roy. Sci. Liége, série. 2 14: 305 (prepr.), 1887 (1988).
Type species: Tapesina griseovitellina (Fuckel) Höhn.
Ecology/substrate/host: Saprobe or minor pathogens of Tsuda heterophylla.
Geographical distribution: Canada (Nag Raj and Kendrick 1975; Verkley 1999).
Description and illustration: Funk (1966b, 1975) and Nag Raj and Kendrick (1975).
Notes: The fungus was fully described with both anamorphs and teleomorph in literatures.
Tapesina griseovitellina (Fuckel) Höhn., Ber. dt. bot. Ges. 37: 108, 1919.
≡ Peziza griseovitellina Fuckel, Fungi rhenani exsic., suppl. fasc. 4: no. 1873, 1866 (1867).
= Arachnopeziza ruborum (Cooke & W. Phillips) Rehm, Rabenh. Krypt.-Fl., Edn 2 (Leipzig) 1.3(lief. 38): 701, 1892 (1896).
≡ Belonidium griseovitellina (Fuckel) Rehm, Rabenh. Krypt.-Fl., Edn 2 (Leipzig) 1(3): 562, 1891.
= Belonidium ruborum (Cooke & W. Phillips) Sacc., Syll. Fung. (Abellini) 8: 501, 1889.
= Chalara rubi Sacc., Rev. Mycol. Toulouse 8: 24, 1886.
= Peziza ruborum Cooke & W. Phillips, Grevillea 9(no. 51): 105, 1881.
= Tapesina ruborum (Cooke & W. Phillips) Lambotte, Mém. Soc. roy. Sci. Liège. série. 2, 14: 305 (prepr.), 1887 (1888).
≡ Trichobelonium griseovitellinum (Fuckel) Rehm, Ber. bayer. bot. Ges. 14: 106, 1914.
≡ Velutaria griseovitellina (Fuckel) Fuckel, Jb. nassau. Ver. Naturk. 23–24: 300, 1870 (1869–1870).
Ecology/substrate/host: Saprobe on decaying stem of Rubus spp.
Geographical distribution: France, North America ad UK (Höhnel 1919, 1923; Dennis 1949; Nag Raj and Kendrick 1975; Baral 2002; Bates et al. 2018).
Description and illustration: Höhnel (1919, 1923), Dennis (1949), Nag Raj and Kendrick (1975), and Baral (2002).
Notes: Tapesina griseovitellina was redescribed in detail on the basis of rich fresh collections using vital characters, and was known with both anamorph and teleomorph (Baral 2002). The connection to the associated hyphomycete Chalara rubi was established by the observation that the large phialoconidia of the Chalara germinated with the very small phialides which also occurred on the overmature ascospores of the teleomorph. Three different synnamorphs were described by Bara (2002). The 1-septate conidia with frayed fringes of wall material at both ends were special among the known species of Chalara s. lat. (Nag Raj and Kendrick 1975; Baral 2002). Baral (2002) considered it more closely related to Calycellina due to presence of yellow, refractive vacuoles in the living paraphyses, a violet stain of the ascospore wall in cresyl blue, and the chalara-like anamorph. No living culture or DNA sequence was available from the public database, its phylogenetic relationship with other chalara-like fungi remains to be studied in future.
Arachnopezizeae Nannf. ex Korf, Lloydia 14(3): 139, 1952.
Type genus: Arachnopeziza Fuckel.
Ecology/substrate/host: Saprobe on dead plant materials.
Geographical distribution: Widely distributed.
Description and illustration: Korf (1952) and Ekanayaka et al. (2019).
Accepted chalara-like genus: Leochalara.
Notes: The family Arachnopezizeae (Helotiales) is characterized by apothecia covered by hairs; ectal excipulum textura angularis to prismatica cells, medullary excipulum textura prismatica to textura oblita cells; hyaline and cylindrical paraphyses; 8-spored, cylindric-clavate, amyloid asci arising from croziers, and ellipsoid to fusoid, 0–7-septate ascospores (Ekanayaka et al. 2019). Five genera (Arachnopeziza, Arachnoscypha, Austropezia, Eriopezia and Durella) were included in the family (Korf 1952; Ekanayaka et al. 2019; Johnston et al. 2019; Kosonen et al. 2021). None of them were reported with chalara-like anamorph. One specimen collected in China had a hyaline chalara-like fungus with hyaline conidiophores, and cylindrical, hyaline and aseptate conidia. The phylogenetic analyses showed it represented a new genus of Arachnopezizeae (Fig. 119), and Leochalara is established for it.

Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for Arachnopezizeae. Bootstrap support values ≥ 60% are shown at the nodes. Neotainosphaeria microspora NN44779 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Leochalara W.P. Wu, gen. nov., MycoBank MB845261.
Etymology: Refers to its morphological similarity to Chalara in conidiophores, phialides and conidiogenesis, and phylogenetically also belong to Leotiales.
Type species: Leochalara danxiashanensis W.P. Wu & Y.Z. Diao.
Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall. Anamorph: Stroma absent. Setae absent. Conidiophores arising from the superficial mycelium, solitary or rarely in groups, hyaline to subhyaline, smooth or slightly rough-walled, 0–2 septate, straight or slightly curved, composed of a 1(–2)-celled basal stalk and a terminal phialide. Conidiogenous cells integrated, determinate, phialidic, erect, straight, lageniform, hyaline to subhyaline, smooth- and thin-walled, consisting of a venter and a collarette, transition from venter to collarette gradual, venter cylindrical, collarette cylindrical, concolorous with venter, smooth. Conidia endogenous, extruded in long chains, cylindrical, both apex and base truncated or flattened, hyaline, aseptate, thin- and smooth-walled. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: The new genus Leochalara is established for a microfungus with hyaline to very pale brown conidiophores, chalara-like phialides, and hyaline, aseptate and cylindrical conidia. Morphologically hyaline phialides and hyaline, aseptate conidia in this genus resemble those of Pyxidiophora, but phylogenetically it belonged to Leotiomycetes.
Leochalara danxiashanensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 120, MycoBank MB845269.

Leochalara danxiashanensis (Wu17421, holotype). a–l conidiophores and phialides. m–u conidia. Scale bar 5 µm
Etymology: Refers to the locality where the fungus was originally discovered.
Typification: China, Guangdong Province, Shaoguan, Danxiashan, on dead leaves of unidentified tree, 25 December 2012, Wenping Wu, Holotype HMAS 352189 (= Wu12471), ex-type strain CGMCC3.23397 (= NN55374).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Stroma absent. Anamorph: Stroma absent. Setae absent. Conidiophores arising from the superficial mycelium, solitary or rarely 2–3 in clusters, erect, straight or slightly curved, obclavate, lageniform, subcylindrical, 16–32 × 2.5–3 µm, hyaline to subhyaline, smooth or slightly rough-walled, 0–2 septate, composed of a 1(–2)-celled basal stalk and a terminal phialide. Conidiogenous cells integrated, terminal, determinate, phialidic, erect, straight, obclavate, lageniform, 16–18 µm long, hyaline to subhyaline, smooth- and thin-walled, consisting of a venter and a collarette; transition from venter to collarette gradual; venters cylindrical, 9–10 µm long and 3–3.5 µm wide; collarettes cylindrical, 6–8 µm long, 1.5–2 µm wide; ratio of mean lengths of collarette and venter = 0.7:1. Conidia endogenous, extruded in long chains, cylindrical, 4–5 × 1–1.5 µm, both ends truncated or flattened, hyaline, aseptate, smooth- and thin-walled; mean conidium length/width = 3.6:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to pale brown, reverse concolorous, sterile, up to 4 mm on PDA at 25 °C in 4 weeks.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: China.
Notes: Leochalara danxiashanensis is characterized by its hyaline to subhyaline conidiophores, chalara-like phialides, and hyaline, aseptate and cylindrical conidia in chains. Four chalara-like species, including Chalara hyalina, C. schoenoplecti, C. siamense and C. sibika, produced hyaline conidiophores, and hyaline, aseptate conidia (Nag Raj and Kendrick 1975; Morgan-Jones and Ingram 1976; Morgan-Jones 1984; Subramanian and Sudha 1986; McKenzie 2002). The new species differs from them by having smaller conidia (4–5 × 1–1.4 µm) and the unique ITS sequence.
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 55374 included Dwayaangam colodena (MH863042, 84% identity), Eriopezia caesia (MZ322849, 85% identity), Durella macrospora (KY462813, 89%). Based on LSU blast in GenBank, the closest matches to the ex-type strain 55374 include Durella macrospora (KY462813, 98%), Eriopezia caesia (MZ322849, 97% identity) and Dwayaangam colodena (NG_064195, 96% identity).
Hamatocanthoscyphaceae Ekanayaka & KD. Hyde, in Ekanayaka et al., Mycosphere 10(1): 347, 2019.
Type genus: Hamatocanthoscypha Svrček.
Ecology/substrate/host: Saprobe on dead plant materials.
Geographical distribution: Widely distributed.
Description and illustration: Ekanayaka et al. (2019).
Accepted chalara-like fungal genera: Brachyalara, Catenulifera, Constrictochalara, Cylindrochalara, Infundichalara, Stipitochalara and Xenochalara.
Notes: Hamatocanthoscyphaceae was a recently established family for those genera included in Han’s Clade 1 and the included genera are Brachyalara, Chalara, Ciliolarina, Curviclavula, Gremmenia, Hamatocanthoscypha, Hyalodendriella, Infundichalara, Microscypha, Pseudohelotium, Xenochalara and Xenopolyscytalum (Han et al. 2014; Ekanayaka et al. 2019). In the phylogenetic analyses based on 15 concatenated sequences of Leotiomycetes, separation of Hamatocanthoscyphaceae as a different family from Pezizellaceae was not supported, thus were treated as members of Pezizellaceae (Johnston et al. 2019). However, the family Pezizellaceae was split into three clades in the ITS tree, and one of these clades was equal to Hamatocanthoscyphaceae (Johnston et al. 2019).
Phylogenetic analyses with 4 different datasets in this study supported separation of Hamatocanthoscyphaceae from Pezizellaceae as two different families. In the phylogenetic trees (Figs. 1, 2, 3, 121), thirteen chalara-like fungi clustered in the strongly supported Hamatocanthoscyphaceae clade. These species were known as Brachyalara straiminea, Chalara hyalocuspica, C. constricta, C. ellisii, C. holubovae, C. longipes, C. piceae-abietis, C. recta, Hamatocanthoscypha podocarpi, Infundichalara microchona, I. minuta, Xenochalara juniperi, and one undescribed chalara-like species (Coetsee et al. 2000; Réblová and Gams 2011; Ekanayaka et al. 2019; Johnston et al. 2019; Crous et al. 2020). Based on morphological and phylogenetic analyses in this study, five monophyletic genera were proposed to accommodate these chalara-like fungi (Fig. 121). Constrictochalara W.P. Wu & Y.Z. Diao was established for Chalara constricta, C. ellisii, C. holubovae, Hamatocanthoscypha podocarpi and one undescribed species. The new genus is characterized by short conidiophores consisting of 1–2-celled basal stalk and a terminal phialide with sharp transition from venter to collarette, obvious constriction and darker between venters and collarettes, and hyaline, aseptate, clavate, subcylindrical or cylindrical conidia (Nag Raj and Kendrick 1975; Koukol 2011). Cylindrochalara W.P. Wu & Y.Z. Diao was introduced for C. hyalocuspica. In the phylogenetic tree, C. hyalocuspica and Infundichalara microchona (the type species of the genus) clustered together as a distinct clade. The two genera can be distinguished by shape of collarette and conidia. In I. microchona, the collarettes are typical funnel-shaped, and the conidia are aseptate and clavate; while in C. hyalocuspica, the collarettes and the conidia are cylindrical. Stipitochalara W.P. Wu & Y.Z. Diao was established as new genus for Chalara longipes, C. piceae-abietis and C. recta. The genus is characterized by multi-septate conidiophores, terminal phialides with abrupt transition from venter to collarette, and hyaline, aseptate, cylindrical conidia without basal frill. Two existing chalara-like genera were accepted, Infundichalara for I. microchona and I. minuta; and Xenochalara for X. juniperi, a monotypic genus. The two species of Infundichalara were morphologically similar, but phylogenetically did not cluster together as one clade.

Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the chalara-like anamorphic fungi in Hamatocanthoscyphaceae (Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.90 are shown at the nodes. Hymenoscyphus albidus CBS126533 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
The phylogenetic analyses in this study also showed that some genera in family Hamatocanthoscyphaceae were polyphyletic and generic delimitation were problematic (Fig. 121). Four species of Hamatocanthoscypha scattered in four different clades: the type species H. laricionis was clustered together with Xenochalara juniperi as a strongly supported clade; the chalara-like species H. podocarpi was clustered together with three other chalara-like species (Constrictochalara clavatospora, C. constricta, C. ellisii and C. holubovae) H. straminella and H. uncipila); H. straminella and Xenopolyscytalum pinea clustered as one strongly supported clade; and H. uncipila and Infundichalara minuta formed a distinct clade but without strong support. For the genus Microscypha, in the strongly supported clade with two Microscypha species, two other species Gremmenia infestans and Pezizella chrysostigma (the type species) were also included; while Microscypha cajaniensis formed a separate branch distinct from other two species. Future revisions for those genera are needed to clarify their phylogenetic relationship towards establishing the monophyletic generic concepts.
Constrictochalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845257.
Etymology: Refers to its phialides with significant constriction between venters and collarettes.
Type species: Constrictochalara clavatospora W.P. Wu & Y.Z. Diao.
Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae, scattered, cylindrical, brown, septate, smooth, with or without percurrent and/or sympodial proliferations. Conidiogenous cells integrated, determinate, phialidic, subcylindrical, lageniform, ampulliform, erect, straight or slightly curved, brown, smooth, consisting of a venter and a collarette, transition from venter to collarette abrupt and marked by a pronounced constriction; venter ellipsoidal, subcylindrical, brown; collarette cylindrical. Conidia enteroblastic, endogenous, hyaline, aseptate, clavate, cylindrical, apex rounded, base truncate, extruded in short chains. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on decaying material of plants or from soil.
Geographical distribution: Europe and China.
Notes: In the phylogenetic analyses from this study (Figs. 2, 3, 121), four species of chalara-like fungi, known as Chalara constricta, C. ellisii, C. holubovae and Hamatocanthoscypha podocarpi, and an undescribed species, were grouped together as a strongly supported monophyletic clade distinct from other chalara-like fungi. Morphologically all the five species shared some similarities such as solitary, simple and short conidiophores, abrupt transition from venter to collarette and marked by a pronounced and dark colored constriction, and hyaline, aseptate conidia with obtuse to truncate ends, which were different from other chalara-like fungal genera. The new genus Constrictochalara is established for these species, which are morphologically similar to Chalara s. str., but differs in transition from venter to collarettes marked by a constriction and darker area in the cell wall.
Phylogenetically Constrictochalara is also closely related to Infundichalara, but morphologically differs from it by tubular-shaped collarettes and endogenous conidia, while collarettes in Infundichalara are more or less funnel-shaped, and conidia are ellipsoidal and not endogenous (Réblová et al. 2011). Five species were accepted in this new genus (Figs. 122, 123). However, several other known species of Chalara s. lat. with similar morphology, such as Chalara novae-zelandiae, C. verruculosa, Chalara state of Ceratocystis autographa, and Chalara state of Cryptendoxyla hypophloia, might also belong to the genus (Nag Raj and Kendrick 1975; Sutton 1993; McKenzie et al. 2002).

Colony of Nagrajchalara, Constrictochalara and Lareunionomyces species on PDA in 4 weeks. a–d Nagrajchalara morganjonesii (a, b 43682, c, d 44060). e, f Constrictochalara constricta (CBS248.76). g–j C. clavatospora (g, h 78182; i, j ex-type strain 76089). k, l Lareunionomyces loeiensis (76001). m, n L. minimus (ex-type strain 77983). o, p L. syzygii (77239)

Colony of Neolauriomyces, Minichalara, Hymenoscyphus, Pyxidiophora and Leochalara on PDA in 4 weeks. a, b Neolauriomyces beijingensis (ex-type strain 78013). c, d Neolauriomyces crousii (ex-type strain 50673). e, f Minichalara aseptata (ex-type strain 48017). g, h Hymenoscyphus globus (78159). i, j Pyxidiophora schoenoplecti (47773). k, l Leochalara danxiashanensis (ex-type strain 55374)
Constrictochalara clavatospora W.P. Wu & Y.Z. Diao, sp. nov., Fig. 124, MycoBank MB845264.

Constrictochalara clavatospora (Wu15217, holotype). a–l, o–q conidiophores and phialides. m, n, r–t conidia. Scale bar 5 µm
Etymology: Refers to its clavate-shaped conidia.
Diagnosis: Morphologically similar to Chalara constricta, C. ellisii and C. holubovae, but differs in size of conidiophores, phialides and conidia.
Typification: China, Beijing, Mentougou, Baihuashan, on dead fruit of Pinus sp., 4 August 2018, Wenping Wu, Holotype HMAS 352183 (= Wu15217a), ex-type strain CGMCC3.23411 (= NN76089).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores formed from superficial hyphae, scattered or aggregated in small groups, cylindrical, obclavate, 30–40 µm long, 3.5–4 µm wide, versicolorous, medium brown, daker in the lower part of collarette, 1–4-septate, smooth, with 1–3 percurrent proliferations and 1–2 sympodial proliferations. Conidiogenous cells integrated, terminal or becoming lateral due to sympodial proliferation, determinate, phialidic, erect, straight or slightly curved, ampulliform, lageniform, obclavate, 13–17 µm long, medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt, often marked by a darker and pronounced constriction; venters cylindrical, conical, subellipsoidal, 8–11 × 2.5–3.5 µm, medium brown; collarettes cylindrical, 6–8 × 1.5–1.7 µm, versicolorous, darker brown in the lower part; ratio of mean lengths of collarette and venter = 0.7:1. Conidia endogenous, extruded in short chains, slightly obclavate, subcylindrical, 4.5–6 × 1.2–1.3 µm, tapering gradually towards ends, usually wider in one end, both ends obtuse or rounded, aseptate, hyaline, thin- and smooth-walled; mean conidium length/width = 4.2:1. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, grey to pale brown, reverse white to slightly soil brown, sterile, up to 10 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Hebei Province, Zhangjiakou, Chicheng, Haituoshan, on dead fruit of Pinus sp., 25 September 2020, Wenping Wu, Wu7418a, Wu17419 and Wu17420. Living strains: CGMCC3.23442 (= 78182, from Wu17419), 76014 (from Wu15217b), 76090 (from Wu15217a), 78179 (from Wu17418a), 78180 and 78181 (from Wu17419), 78183, 78184 and 78185 (from Wu17420).
Ecology/substrate/host: Saprobe on dead cone of pine tree.
Geographical distribution: China.
Notes: Constrictochalara clavatospora is morphologically similar to other three species in the genus, but can be distinguished from them by shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). Constrictochalara clavatospora differs from C. ellisii and C. holubovae by its clavate conidia with rounded apex and truncate base, while the conidia in the other two species are cylindrical with rounded or truncate ends. Constrictochalara constricta also has clavate conidia, but has much longer (17–26 μm) phialides than those in C. clavatospora.
Ten studied strains of this species have almost identical ITS (1 bp difference) and partial LSU sequences. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 76068 includes Infundichalara microchona (HM036588, 98% identity), Hamatocanthoscypha podocarpi (NR_170825, 97% identity), Chalara holubovae (ON261244, 96% identity) and many unnamed fungi of Leotiomycetes.
Constrictochalara constricta (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845289.
≡ Chalara constricta Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 103, 1975.
Material examined: CBS 248.76, isolated from decaying wood from Belgium.
Ecology/substrate/host: Saprobe on dead cone of pine tree.
Geographical distribution: Belgium and New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara constricta was originally described from dead leaves of Agathis australis (Nag Raj and Kendrick 1975). Morphologically it is similar to Constrictochalara clavatospora, but differs in longer phialides (17–26 μm) and conidia (3.5–8 × 1–2 µm). Furthermore, their ITS sequences are with 10 bp differences from each other. Constrictochalara constricta also resembles also Hamatocanthoscypha podocarpi, but differs in shorter and narrower conidia, and their ITS sequences are with 4 bp differences from each other.
ITS and partial LSU sequences of this species were generated from the living strain CBS 248.76. Based on ITS blast in GenBank, the closest match to this strain includes Infundichalara microchona (HM036588, 100% identity), Hamatocanthoscypha podocarpi (NR_170825, 99% identity), Chalara holubovae (FR667222, 97% identity) and many unnamed fungi of Leotiomycetes.
Constrictochalara ellisii (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845290.
≡ Chalara ellisii Nag Raj & B.W.B. Kendr., Monogr. Chalara Allied Genera: 113, 1975.
Description on the natural substrate: Anamorph: Conidiophores formed from superficial hyphae, multiseptate, cylindrical, dark brown and verrucose at the base, becoming paler above, terminating in a phialide, 32–185 µm long, 2.5–4.5 µm wide at the base. Conidiogenous cells integrated, determinate, phialidic, monoblastic, erect, straight or slightly curved, pale brown to medium brown, smooth, subcylindrical to lageniform, 20–38 µm long, consisting of a venter and a collarette, transition from venter to collarette abrupt and marked by a constriction in the wall; venters cylindrical to subcylindrical, pale brown, 13–29 × 3–4.5 µm long; collarettes cylindrical, pale brown, 7–10 × 2–2.5 µm. Conidia enteroblastic, endogenous, hyaline, aseptate, cylindrical with truncate or obtuse ends, 3–10 × 1.5 –2 µm, extruded singly or in short chains. Teleomorph: Unknown.
Material examined: CBS 928.97, isolated from human material from Slovenia.
Ecology/substrate/host: Saprobe in soil.
Geographical distribution: Canada and Slovenia (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara ellisii was originally described from soil. The living strain CBS 928.97 isolated from human material was studied by us. Morphologically it differs from other species in the genus by multiseptate conidiophores and phialides with longer venter (Nag Raj and Kendrick 1975).
ITS and partial LSU sequences of this species were generated from the living strain CBS 928.97. Based on a megablast search of GenBank nucleotide database, the closest matches to this strain include Hamatocanthoscypha sp. (MH921853, 100% identity), Hamatocanthoscypha podocarpi (NR_170825, 97% identity), Chalara holubovae (FR667222, 96% identity) and many unnamed fungi of Leotiomycetes.
Constrictochalara holubovae (Koukol) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845291.
≡ Chalara holubovae Koukol, Fungal Diversity 49(1): 81, 2011.
Ecology/substrate/host: Saprobe in decaying needled of Picea abies.
Geographical distribution: Czech Republic and Lithuania (Koukol 2011).
Description and illustration: Koukol (2011).
Notes: Chalara holubovae is a recently described species, which fits well to the concept of the genus in both morphology and phylogeny (Koukol 2011).
Constrictochalara podocarpi (Crous) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845292.
≡ Hamatocanthoscypha podocarpi Crous, in Crous et al., Persoonia 44: 327, 2020.
Anamorph: Conidiophores subcylindrical, branched below, 0–4-septate, 12–60 × 3–5 µm. Conidiogenous cells 13–40 × 3–4 µm, integrated, terminal and intercalary, subcylindrical, pale brown, smooth, base tapering to long cylindrical, apical venter, 3–9 µm long, slightly flared or not, 2–3 µm diam. Conidia in long unbranched chains, aseptate, hyaline, smooth, guttulate, subcylindrical with truncate ends, (6–)7–8(–9) × (1.5–)2 µm (Adapted from Crous et al. 2020). Teleomorph: Unknown.
Ecology/substrate/host: Pathogenic and causing leaf spot on Podocarpus latifolius.
Geographical distribution: South Africa (Crous et al. 2020).
Description and illustration: Crous et al. (2020).
Notes: Hamatocanthoscypha podocarpi was known only with chalara-like anamorph and closely related to other species of Constrictochalara (Nag Raj and Kendrick 1975; Crous et al. 2020). The phylogenetic analyses clearly showed that H. podocarpi was congeneric with other species of Constrictochalara, and distinct from the type species of Hamatocanthoscypha, H. laricionis. Morphologically it also fits well to the new genus Constrictochalara in producing short conidiophores, terminal phialides with abrupt transition from venters to collarettes, slightly constriction and darker between venter and collarettes, and hyaline, aseptate, subcylindrical to clavate conidia (Nag Raj and Kendrick 1975; Koukol 2011; Crous et al. 2020).
Other relevant existing chalara-like species:
The following two species are with similar morphology as the type species of Constrictochalara, but their phylogenetic relationship with the genus needs to be confirmed by molecular phylogenetic study in future (Nag Raj and Kendrick 1975; Sutton 1993; McKenzie et al. 2002).
Chalara novae-zelandiae Nag Raj & W.B. Kendr., Monogr. Chalara Allied Genera (Waterloo): 126, 1975.
Ecology/substrate/host: Saprobe in decaying leaves of Nothofagus menziesii and Podocarpus dacrydioides.
Geographical distribution: New Zealand (Nag Raj and Kendrick 1975).
Description and illustration: Nag Raj and Kendrick (1975).
Notes: Chalara novae-zealandiae fits well to the concept of Constrictochalara in proliferation of conidiogenous cells, sharp transition from venter to collarette, clear constriction between the collarette and venter, and cylindrical, small-sized conidia.
Chalara verruculosa B. Sutton, Mycol. Pap. 167: 16, 1993.
Ecology/substrate/host: Saprobe in dead stem of Rubus ellipticus.
Geographical distribution: Malawi (Sutton 1993).
Description and illustration: Sutton (1993).
Notes: Chalara verruculosa also fits well to the concept of Constrictochalara, and differs from other species by rough-walled conidiophores and phialides, and size of conidia.
Cylindrochalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845258.
Etymology: Refers to its cylindrical venter and similarity with Chalara.
Anamorph: Conidiophores solitary, erect, cylindrical, consisting of 1–2-celled basal stalk and a terminal phialide, light greenish gray, smooth. Conidiogenous cells intergrade and terminal, or formed directly on vegetative hyphae, phialidic, lageniform, straight or slightly bent, light brown, smooth, gradually tapering towards the collarette, venter cylindrical, collarette cylindrical, transition from venter to collarette abrupt. Conidia cylindrical with rounded ends, rarely clavate, hyaline, aseptate, formed in long chains and sometimes connected with connectives (adapted from Koukol 2011). Teleomorph: unknown.
Type species: Cylindrochalara hyalocuspica (Koukol) W.P. Wu & Y.Z. Diao.
Ecology/substrate/host: Saprobe on needle of Picea abies.
Geographical distribution: Czech Republic and Lithuania.
Notes: Chalara hyalocuspica is a recently described species with short conidiophores consisting of 1–2-celled basal stalk and a terminal phialide, abrupt transition from venter to collarette, and hyaline, aseptate cylindrical conidia without basal frill (Koukol 2011). The phylogenetic analyses in this study showed that C. hyalocuspica was phylogenetically related to Stipitochalara, Infundichalara and Xenochalara in Hamatocanthoscyphaceae, and distinct from other chalara-like fungi. Morphologically it is hardly distinguished from Chalara s. str., except for its conidia sometimes with connectives.
Cylindrochalara hyalocuspica (Koukol) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845293.
≡ Chalara hyalocuspica Koukol, Fungal Diversity 49(1): 83, 2011.
Anamorph: Conidiophores cylindrical, consisting of a 1–2-celled basal stalk and a terminal phialide, light greenish gray, smooth, (25–)28.5–39 (–48) × 3.5–5 μm. Conidiogenous cells terminal on conidiophores or formed directly on vegetative hyphae, lageniform, straight or slightly bent, light brown, smooth, gradually tapering towards the collarette, (14.5–)18.5–30(–33) × 3.5–5 μm, transition from venter to collarette abrupt; venter cylindrical, (14.5–)17–24.5(27.5) × 3–5 μm; collarette cylindrical, 7.5–10 × 2–2.5 μm. Conidia cylindrical with rounded ends, rarely clavate, 4.5–7.5(–9.5) × 1.5–2.5 μm, one-celled, hyaline, formed in long chains and sometimes connected with connective 0.5–1 μm long (adapted from Koukol 2011). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on needle of Picea abies.
Geographical distribution: Czech Republic and Lithuania (Koukol 2011).
Description and illustration: Koukol (2011).
Notes: Cylindrochalara hyalocuspica is monotypic and found on decaying needles of pine tree from Europe.
Infundichalara Réblová & W. Gams, Fungal Diversity. 46(1): 78, 2011.
Anamorph: Conidiophores brown, often arising from dark fascicles of vegetative hyphae, either as simple phialides or with additional septa. Conidiogenous cells phialidic, pigmented mainly in the lower part, subhyaline towards the apex, often sympodially proliferating, tapered slightly below the collarette, collarette funnel-shaped. Conidia hyaline, smooth-walled, clavate, with a truncate base, rounded apically, cohering in long chains. Chlamydospores absent (Adapted from Réblová et al. 2011). Teleomorph: Unknown.
Typus: Infundichalara microchona (W. Gams) Réblová & W. Gams.
Ecology/substrate/host: Saprobe on leaves, dead wood and cone of coniferous tree, and basidiomata of polyporales.
Geographical distribution: Austria, Canada, China, Denmark, Netherland and UK.
Description and illustration: Réblová et al. (2011).
Notes: Infundichalara was created for a single species Chalara microchona W. Gams, which was different from other known chalara-like genera in morphology and phylogeny (Nag Raj and Kendrick 1975; McKenzie et al. 2002; Réblová et al. 2011). Morphologically it differs from other chalara-like genera in producing funnel-shaped collarettes, and non-endogenous, clavate conidia. The genus was known with two species, the type species and I. minuta (Réblová et al. 2011; Koukol 2012). Our phylogenetic analyses showed that these two species were closely related to three known teleomorphic species, Ciliolarina ligniseda, C. pinicola, and Hamatocanthoscypha unicipila.
Infundichalara microchona (W. Gams) Réblová & W. Gams, Fungal Divers. 46: 80, 2011.
≡ Chalara microchona W. Gams, in Gams & Holubová-Jechová, Stud. Mycol. 13:73. 1976.
Ecology/substrate/host: Saprobe on wood and cone of coniferous tree, and basidiomata of polyporales.
Geographical distribution: Austria, Canada, China, Czechslovakia, Denmark, Germany, Netherland and UK (Gams and Holubová-Jechová 1976; Holubová-Jechová 1984; Réblová et al. 2011).
Description and illustration: Gams and Holubová-Jechová (1976) and Réblová et al. (2011).
Notes: Infundichalara microchona was fully documented by Gams and Holubová-Jechová (1976), Holubová-Jechová (1984), and Réblová et al. (2011).
Infundichalara minuta Koukol, Mycotaxon 120: 346, 2012.
Ecology/substrate/host: Saprobe on needles of Pinus spp.
Geographical distribution: Czech Republic (Koukol 2012).
Description and illustration: Koukol (2012).
Notes: Infundichalara minuta produces both anamorphic morphology of Infundichalara and Xenopolyscytalum. In our phylogenetic analyses, I. minuta clustered together with I. microchona, Ciliolarina ligniseda, C. pinicola, and Hamatocanthoscypha unicipila as a strongly supported group.
Stipitochalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845259.
Etymology: Refers to its conidiophores with a multiseptate basal stipe and terminal phialides.
Type species: Stipitochalara longipes (Preuss) Wenping Wu & Y.Z. Diao (≡ Chalara longipes (Preuss) Cooke).
Saprobe on decaying plant material or in soil. Anamorph: Conidiophores solitary, erect, straight or slightly curved, cylindrical, multiseptate, brown, smooth, consisting of a basal multiseptate stipe and a terminated phialide, proliferation percurrently and/or sympodially. Conidiogenous cells integrated, terminal, phialidic, lageniform, brown, consisting of a venter and a collarette; transition from venter to collarette gradual or abrupt; venters subcylindrical; collarettes cylindrical. Conidia endogenous, hyaline, aseptate or uniseptate, cylindrical, with obtuse or truncate ends, extruded singly or in easily dispersible chains. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe from soil and on decaying plant material, including needles of Pinus and Picea, rhizosphere, decaying grass.
Geographical distribution: Australia, Czech Republic, Finland, France, Germany, Poland, Sweden, UK and USA.
Notes: In the phylogenetic tree (Fig. 121), two species of Chalara s. lat., Chalara longipes and C. picea-abietis, clustered together as a strongly supported clade distinct from other known chalara-like fungi. Morphologically these two fungi are characterized by the well-developed conidiophores composed of a multiseptated main stalk and a terminal phialide, and hyaline, aseptate or septate conidia. The new genus Stipitochalara is created for them. These two species are morphologically related to Infundichalara, Constrictochalara and Cylindrochalara in forming well-developed conidiophores with multi-septate stipes and terminal phialides, cylindrical collarettes, and hyaline, aseptate or septate conidia. The new genus is indistinguishable from some known Chalara s. lat. species such as Chalara cylindrosperma, C. nothofagi and C. parvispora (Nag Raj and Kendrick 1975), but they are phylogenetically distinct. Three species are accepted in the genus.
Stipitochalara longipes (Preuss) W.P. Wu & Y.Z. Diao comb. nov., MycoBank MB845294.
≡ Cylindrosporium longipes Preuss, Linnaea 24: 106, 1851.
≡ Chalara longipes (Preuss) Cooke, Grevillea 10(no. 54): 50, 1881.
Anamorph: Conidiophores erect, straight or slightly curved, cylindrical, multiseptate, brown, smooth, 63–120 µm long, terminated with a phialide. Conidiogenous cells phialidic, lageniform, brown, 21–29 µm long, consisting of a venter and a collarette, transition from venter to collarette gradual; venters subcylindrical, 15–18 µm long and 3–5 µm wide; collarettes cylindrical, 5.5–9.5 µm. Conidia enteroblastic, endogenous, hyaline, aseptate, cylindrical, with obtuse or truncate ends, 3.5–6.5 × 1–1.5 µm, extruded singly or in easily dispersible chains. Teleomorph: Unknown.
Material examined: Finland, isolated from Hordeum vulgare, T. Tuomi, CBS264.94; Sweden, isolated from forest soil, B.E. Söderström, CBS411.76.
Ecology/substrate/host: Saprobe from soil and on decaying plant material, including needles of Pinus and Picea, rhizosphere, decaying grass etc.
Geographical distribution: Australia, Czech Republic, Finland, France, Germany, Poland, Sweden, UK and USA (Nag Raj and Kendrick 1975; Holubová-Jechová 1984; Koukol 2011; Bates et al. 2018).
Description and illustration: Nag Raj and Kendrick (1975), Holubová-Jechová (1984), and Koukol (2011).
Notes: Morphologically Stipitochalara longipes resembles Chalara cylindrosperma, C. nothofagi, and C. parvispora, but differs from them by shape and size of conidiophores, phialides and conidia (Nag Raj and Kendrick 1975; Koukol 2011). Compared with S. longipes, Chalara cylindrosperma (conidiophores 32–190 µm long, conidia 5.5–17 × 1.5–2.5 µm), C. nothofagi (conidiophores 115–168 µm long, conidia 13–17 × 2.5–3 µm) and C. parvispora (conidiophores 95–190 µm long, conidia 3.5–6 × 1.5–2 µm) have longer conidiophores, and longer and wider conidia. Stipitochalara longipes was recently documented by Koukol (2011) who also described the great morphological variation in colony, conidiophores, phialides and percurrent proliferations among different strains. In Koukol’s phylogenetic analyses, the studied strains were divided into two moderately supported groups (S. longipes and S. recta), but morphologically these two species were hardly distinguished.
Stipitochalara piceae-abietis (Hol.-Jech.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845295.
≡ Chalara piceae-abietis Hol.-Jech., Folia geobot. Phytotaxon. 19(4): 412, 1984.
Typification: Czech Republic, Horní Lomná, Natural reservation Mionší, litter needles of Picea abies. 28 Jul 1982, V. Holubová-Jechová (holotype, PRM829859).
Ecology/substrate/host: Saprobe on dead plant material.
Geographical distribution: Czech Republic (Holubová-Jechová 1984; Koukol 2011).
Description and illustration: Holubová-Jechová (1984) and Koukol (2011).
Notes: The phylogenetic analyses in this study further confirmed that Chalara piceae-abietis represented a closely related but distinct species from S. longipes. It differs from C. longipes by 1-septate conidia (Koukol 2011).
Stipitochalara recta (Koukol) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845296.
≡ Chalara recta Koukol, Fungal Diversity 49: 87, 2011.
Anamorph: Conidiophores septate, unbranched, erect, straight or slightly bent, moderate to dark yellowish green when fresh, light brown when dry, thick-walled (up to 1.5 μm thick), not constricted at septa, 106–176 μm long (including the phialide), 3.5–5.5 μm wide and up to 10 μm at the base. Conidiogenous cells phialidic, lageniform, hyaline to light brown at the base, smooth, 24–29 μm long; venter subcylindrical, 15–19.5 μm long and 3.5–4.5 μm wide; collarette cylindrical, slightly darker at the basal part, 7.5–11.5 μm long and 1.5–2 μm wide; transition from venter to collarette abrupt, ratio of mean lengths of collarette and venter is 0.6:1. Conidia endogenous, extruded in long chains, cylindrical with obtuse ends, one-celled, hyaline, 5–7 μm × 1–1.5 μm; the mean conidium length/width ratio is 4.5:1 (Adapted from Koukol 2011). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe from soil and on decaying plant material, including needles of Pinus and Picea, rhizosphere, decaying grass etc.
Geographical distribution: Sweden (Koukol 2011).
Description and illustration: Koukol (2011).
Notes: Morphologically Stipitochalara recta resembles S. longipes, but differs from the latter species by longer conidiophores, slightly wider phialides and shorter conidia (Koukol 2011).
Xenochalara M.J. Wingf. & Crous, S. Afr. J. Bot. 66(2): 101, 2000.
Mycelium consisting of branched, septata hyphae; hyphae hyaline and smooth, becoming brown and rough, occurring singly or in strands. Chlamydospores absent. Anamorph: Conidiophores micronematous, arising from aerial mycelium or submerged hyphae, erect, simple, frequently reduced to conidiogenous cells, or 1-septate, subcylindrical, straight to slightly curved, light brown, smooth. Conidiogenous cells phialidic, terminal, subcylindrical or lageniform, smooth, pale to light brown; venter conical to ellipsoid; cellarette narrowly obconical to subcylindrical. Conidia hyaline, smooth, aseptate, occurring in false chains; short clavate, apex rounded, base truncate (Adapted from Coetsee 2000). Teleomorph: Unknown.
Type species: Xenochalara juniperi M.J. Wingf. & Crous.
Ecology/substrate/host: Saprobe on decaying needle of Juniperus communis.
Geographical distribution: Netherland.
Description and illustration: Coetsee et al. (2000).
Notes: Xenochalara morphologically resembles Chalara s. str., but differs in producing conidia through apical wall building, and being tolerant of cycloheximide (Coetsee et al. 2000). Phylogenetically these two genera are also distinct. The genus remained to be monotypic.
Xenochalara juniperi M.J. Wingf. & Crous, S. Afr. J. Bot. 66(2): 102, 2000.
Ecology/substrate/host: Saprobe on decaying needle of Juniperus communis.
Geographical distribution: Netherland (Coetsee et al. 2000).
Description and illustration: Coetsee et al. (2000).
Notes: Xenochalara juniperi is morphologically similar to Chalara state of Ceratocystis autographa in producing sessile conidiogenous cells, transition from venter to collarette abrupt to gradual, and hyaline, aseptate, short-clavate, and small-sized conidia in false chains (Coetsee et al. 2000).
Helotiaceae Rehm (as ‘Helotieae'), Rabenh. Krypt.-Flora, Edn 2 (Leipzig) 1, 3: 647, 1892 (1896).
Type genus: Helotium Pers.
Ecology/substrate/host: Pathogenic or saprobe on plants.
Geographical distribution: Widely distributed.
Description and illustration: Ekanayaka et al. (2019).
Accepted chalara-like genera: Hymenoscyphus.
Notes: In this family, Hymenoscyphus is the only genus known to produce chalara-like anamorphs (Figs. 2, 3, 125). Five species of Hymenoscyphus were known with chalara-like anamorphs, such as H. albidus, A. albidoides, H. fraxineus, H. globus and H. koreanus. The chalara-like anamorphs of Hymenoscyphus are characterized by brown and reduced conidiophores consisting of 1–2 basal cells and a terminal phialide, gradual or disrupt transition from venter to collarette, and hyaline, aseptate, subglobose, short cylindrical to cylindrical conidia. They differs from chalara-like fungal genera in processing reduced and small-sized conidiogenous cells, and short cylindrical or globose and aseptate conidia (Gross and Han 2015; Kowalski and Bilański 2018). All these species are well-connected with their Hymenoscyphus teleomorphs. The anamorph and teleomorph connection was established for H. globus in this study by the phylogenetic analysis. In this species, phialides had obvious constriction between venter and collarette, and conidia were slightly clavate-cylindrical.

Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the chalara-like anamorphic fungi in Helotiaceae (Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Catenulifera lururians CBS647.75 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Hymenoscyphus Gray, Natural Arr. Brit. Pl. 1: 673, 1821.
= Articulospora Ingold, Trans. Br. Mycol. Soc. 25(4): 376, 1942 (1941).
= Belospora Clem., Genera Fung.: 87, 1909.
= Ciboriella Seaver, North American Cup-fungi, (Inoperculates) (New York): 107, 1951.
= Helicodendron Peronel., G. bot. ital., n.s. 25: 460, 1918.
= Helicodesmus Linder, Am. J. Bot. 12: 267, 1925.
= Hymenoscypha (Fr.) W. Phillips, Man. Brit. Discomyc. (London): 111, 1887.
= Lambertellinia Korf. & Lizoñ, Mycotaxon 50: 168, 1994.
= Lambertellinia Korf. & Lizoñ, Inoculum, Newsletter of the Mycological Society of America 44(2): 43, 1993.
= Malotium Velen., Monogr. Discom. Bohem. (Prague): 210, 1934.
= Peziza trib. Hymenoscypha Fr., Syst. Mycol. (Lundae) 2(1): 117, 1822.
= Septatium Velen., Monogr. Discom. Bohem.: 211, 1934.
Type species: Hymenoscyphus fructigenus (Bull.) Gray.
Ecology/substrate/host: Saprobe on dead leaves.
Geographical distribution: widely distributed in Europe, America and Asia.
Notes: The genus Hymenoscyphus (Helotiaceae) has more than 150 existing species with broad distribution in Asia, America and Europe. Most of them are saprophytic on plant debris such as twigs, fruit, seeds, leaves and herbaceous stems, but a few species are plant pathogens, such as the causing agent of ash dieback pathogens H. fraxineus (Baral et al. 2014; Gross et al. 2014a, b; Zheng and Zhuang 2013, 2014; Kowalski and Bilański 2018). Their affinity with Helotiaceae were confirmed by the phylogenetic analyses (Baral et al. 2014; Gross et al. 2014a, b; Zheng and Zhuang 2013, 2014; Kowalski and Bilański 2018; Figs. 1, 2, 3, 125). The chalara-like anamorphs were described for six species of Hymenoscyphus, including H. albidus, H. fraxineus, H. koreanus, H. linearis, H. occultus and H. pusillus (Kowalski 2006; Zhao et al. 2013; Baral et al. 2014; Gross and Han 2015; Gross et al. 2014a, b, 2015). In the phylogenetic analyses several subgroups were recognized for existing species of Hymenoscyphus, and species with chalara-like anamorphs formed a strongly supported subgroup distinct from other species (Zhang and Zhuang 2004; Zheng and Zhuang 2014, 2015; Kowalski and Bilański 2018).
Among the known species under Chalara s. lat. species, C. ampullula is morphologically similar to the anamorphs of Hymenoscyphus in producing sessile conidiogenous cells with an ellipsoidal venter and a shorter collarette (less than length of one conidium), abrupt transition from venter to collarette, and hyaline, aseptate and cylindrical conidia (Nag Raj and Kendrick 1975). Future molecular phylogenetic study is needed to clarify its phylogenetic relationship with Chalara s. lat. and Hymenoscyphus.
Hymenoscyphus globus W.Y. Zhuang & Yan H. Zhang, in Zhang & Zhuang, Nova Hedwigia 78(3–4): 480, 2004, Fig. 126.

Hymenoscyphus globus (a–o from Wu15217; p–ae from Wu17400). a–n, p–s, x, y conidiophores and phialides. o, t–w, z–ae conidia. Scale bar 10 µm for a, b; 5 µm for all others
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores directly arising from superficial hyphae or aggregated cells, scattered or aggregated, erect, straight, consisting of basal supporting cell incorporated into hyphae and a phialide, ampulliform, lageniform, 16–24 µm long, 2–3 µm wide at the base, medium brown, 0–1 septate, smooth- and thin-walled. Conidiogenous cells phialidic, erect, straight or slightly curved, ampulliform, lageniform, 11–23 µm long, versicolorous, medium brown, darker in the transition region from venter to collarette, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt, often marked by a pronounced constriction; venters ellipsoidal, subglobose, 11–15 µm long and 4–6 µm wide, brown; collarettes cylindrical, 4–9 × 2.4–2.7(–3.5) µm, significantly darker than venter, medium brown; ratio of mean lengths of collarette and venter = 0.5:1. Conidia endogenous, extruded in short chains, cylindrical, 6–9 × 2.3–2.5 (–2.8) µm, usually wider in one end, both ends truncated of flattened, hyaline, uniseptate, smooth- and thin-walled; mean conidium length/width ratio = 3.1:1. Teleomorph: Not observed.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, green to dark green, reverse dark green to brown, becoming paler towards the margin, sterile, up to 9 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Hebei Province, Zhangjiakou, Chicheng, Haituoshan, on dead leaves of Quercus dentata, 25 September 2020, Wenping Wu, Wu17398, Wu17399 and Wu17400; Jilin Province, Yanbian, Changbaishan, on dead branches of unidentified plant, 5 Sept. 1998, Wenping Wu, Wu1809b. Living strains: CGMCC3.23463 (= 78159) and 78132 (from Wu17400), 78157, 78131 (from Wu17398), 78158 (from Wu17399).
Ecology/substrate/host: Saprobe on dead leaves of Quercus dentata.
Geographical distribution: China (Zhang and Zhuang 2004).
Description and illustration: Zhang and Zhuang (2004).
Notes: Morphologically the fungus is similar to other chalara-like anamorphs of Hymenoscyphus in producing reduced conidiophores, short phialides, and hyaline, aseptate conidia (Kowalski 2006; Zhao et al. 2012; Gross and Han 2015; Gross et al. 2014a, b, 2015; Kowalski and Bilański 2018). ITS sequences from the studied strains were identical to those from the holotype of H. globus, thus we assigned our fungus into H. globus (Fig. 125). Hymenoscyphus globus was originally described with the teleomorph only (Zhuang and Zhang 2004). The anamorph discovered in this study is morphologically very similar to Chalara ampullula and other known anamorphs of Hymenoscyphus such as H. albidus, H. fraxineus, H. koreanus, H. linearis and H. occultus, but differs in shape and size of phialides and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002; Kowalski 2006; Zhao et al. 2012; Gross and Han 2015; Gross et al. 2014a, b, 2015). In the phylogenetic tree, Hymenoscyphus globus clustered together with non-pathogenic species such as H. crataegi and H. emmutabilis and distinct from the pathogenic species with the chalara-like anamorph (Zheng and Zhuang 2014, 2015; Figs. 3, 122).
Other Hymenoscyphus species known with chalara-like anamorphs:
Hymenoscyphus albidus (Gillet) W. Phillips, Man. Brit. Discomyc. (London): 138, 1887.
≡ Phialea albida Gillet, Champignons de France. Discom (4): 105, 1881.
≡ Helotium albidum (Gillet) Pat., Tab. analyt. Fung. (Paris) (4): 173, 1885.
= Helotium robergei Dennis, Mycol. Pap. 62: 93, 1956.
= Helotium scutula var. albidum P. Karst., Bidr. Känn. Finl. Nat. Folk 19: 112, 1871.
≡ Lambertella albida (Gillet) Korf, Mycotaxon 14(1): 2, 1982.
≡ Lanzia albida (Gillet) S.E. Carp., Mem. N. Y. bot. Gdn. 33: 187, 1981.
= Peziza albida Roberge, in Desmazières, Pl. Crypt. Nord France, Edn 1: no. 2004, 1851.
Ecology/substrate/host: Saprobes on dead leaf of ash trees.
Geographical distribution: Europe (Baral and Bemmann 2014).
Description and illustration: Kowalski (2006), Kowalski and Holdenrieder (2009), Kiristis et al. (2013), Baral et al. (2014), and Gross et al. (2014).
Notes: Kiristis et al. (2013) studied many strains of H. albidus and found no anamorph associated in pure culture.
Hymenoscyphus fraxineus (T. Kowalski) Baral, Queloz & Hosoya, IMA Fungus 5(1): 79, 2014.
≡ Chalara fraxinea T. Kowalski, For. Pathol. 36(4): 264, 2006.
= Hymenoscyphus pseudoalbidus V. Queloz, C.R. Grünig, R. Berndt, T. Kowalski, T.N. Sieber & O. Holdenrieder, For. Path. 41(2): 140, 2011.
Ecology/substrate/host: Pathogens and causes necrotic lesions on lead, twigs and stems of Fraxinus angustifolia and F. excelsior.
Geographical distribution: Asia and Europe, including China, Russia, Korea, Poland, Russia, Switzerland, etc. (Baral et al. 2014; Gross et al. 2014).
Description and illustration: Kowalski (2006), Kowalski and Holdenrieder (2009), Baral et al. (2014), and Gross et al. (2014).
Notes: Hymenoscyphus fraxineus was described with both anamorph and teleomorph from leaf petioles of Fraxinus chinensis subsp. Rhynchophylla (Kowalski (2006; Kowalski and Holdenrieder 2009; Baral et al. 2014; Gross et al. 2014). In this species, the phialides are subcylindrical, obclavate or occasionally lageniform, 16–24 μm long, with a short-cylindrical or ellipsoidal venter (11–15 × 4–5 μm) and a cylindrical collarettes (5–7(–9) × 2.2–2.7 μm), and the conidia are short-cylindrical, 3.2–4 × 2–2.5 μm, ends rounded or blunt, sometimes with a truncate base bearing small marginal frills, aseptate, hyaline, with 1–2 oil droplets, extruded in short chain or slimy droplets (Kowalski 2006; Kowalski and Holdenrieder 2009).
Hymenoscyphus koreanus A. Gross & J.G. Han, Mycol. Progr. 14(1): 7, 2015.
Ecology/substrate/host: Pathogenic on shoot of Fraxinus excelsioris.
Geographical distribution: South Korea (Gross and Han 2015).
Description and illustration: Gross and Han (2015).
Notes: Hymenoscyphus koreanus was described with both anamorph and teleomorph from leaf petioles of Fraxinus chinensis subsp. rhynchophylla (Gross and Han 2015). The anamorph was also similar to chalara-like appearance of Hymenoscyphus fraineus, but only non-sporulating phialidic conidiogenous structures were observed. The subcylindrical to obclavate (20.9–29.5 μm long) consisted of a short-cylindrical to ellipsoidal venter (11–22 × 3.5–4.5 μm) and a cylindrical collarette (7.5–14.5 × 2.5–3 μm), and no conidia was observed by Gross and Han (2015).
Hymenoscyphus linearis Hosoya, Andr. Gross & Baral, in Gross, Hosoya, Zhao & Bara, Mycol. Progr. 14(1): 8, 2015.
Ecology/substrate/host: Saprobe on leaf petioles of Fraxinus platypoda.
Geographical distribution: Japan (Gross et al. 2015).
Description and illustration: Gross et al. (2015).
Notes: Hymenoscyphus linearis was described with both anamorph and teleomorph from leaf petioles of Fraxinus platypoda. Two different anamorphs, chalara-like and Sporotrichum-like, were described from pure culture. The chalara-like anamorph was similar to chalara-like appearance of Hymenoscyphus fraineus, the phialide (16.7–36.7 μm long) consisted of an ellipsoidal or rarely cylindrical venter (8.5–21 × 3.6–5.2 μm) and a cylindrical collarette (7.5–16.5 × 2.5–3 μm), and the conidia were aseptate, cylindrical, slightly clavate to almost globose, 2.1–5.9 × 1.8–2.8 μm, extruded in slimy droplets or rarely in short chains (Gross et al. 2015).
Hymenoscyphus occultus Andr. Gross & J.G. Han, Mycol. Progr. 14(1): 9, 2015.
Ecology/substrate/host: Saprobe on leaf petioles of Fraxinus chinensis subsp. rhynchophylla.
Geographical distribution: Korea.
Description and illustration: Gross and Han (2015).
Notes: Hymenoscyphus linearis was described with both anamorph and teleomorph from leaf petioles of Fraxinus chinensis subsp. Rhynchophylla (Groass and Han 2015). The anamorph was also similar to chalara-like appearance of Hymenoscyphus fraineus, the subcylindrical to obclavate phialide (16–22 μm long) consisted of a short–cylindrical to ellipsoidal venter (8–13 × 3.75–4.75 μm) and a cylindrical collarette (7.5–10.75 × 2–2.75 μm), and the conidia are aseptate, short cylindrical to short-clavate, ends rounded or truncated, 3–5 × 2–2.5 μm, extruded in slimy droplets.
Neolauriomycetaceae Crous, in Crous et al., Persoonia 40: 359, 2018.
Anamorph: Conidiophores solitary, erect, subcylindrical, straight, slightly flexuous, unbranched, medium brown, smooth, septate, terminating in a phialide, or in a penicillate head: primary branches subcylindrical to doliform, medium brown, smooth; secondary branches doliform to subcylindrical, medium brown, smooth, giving rise to phialides. Conidiogenous cells phialidic, ampulliform, medium brown, smooth, including the apical collarette, cylindrical, medium brown. Conidia occurring in chains, unbranched, hyaline, smooth-walled, cylindrical, aseptate, ends truncate (Adapted from Crous et al. 2018a, b). Teleomorph: Unknown.
Type genus: Neolauriomyces Crous.
Ecology/substrate/host: Saprobe on dead leaves of trees.
Geographical distribution: Australia, China, Colombia, France and Thailand.
Description and illustration: Crous et al. (2018a, b).
Accepted chalara-like genera: Exochalara, Lareunionomyces, Neolauriomyces and Minichalara.
Notes: The family Neolauriomycetaceae was created to accommodate three hyphomycetes genera, namely Exochalara, Lareunionomyces and Neolauriomyces (Crous et al. 2018a, b). None of these fungi was known with teleomorph (Crous et al. 2016a, b, 2018a, b, 2019). Based on this study (Fig. 3, 127), the family was further expanded with a new genus Minichalara and 5 additional species, including one new combination and four new species. In the phylogenetic trees generated from two different datasets of integrated LSU and ITS sequences, separation of Exochalara, Lareunionomyces and Neolauriomyces was strongly supported, and species of each genus formed a distinct lineage with strong support in at least one of the dataset. Two species of Minichalara, M. aseptata and M. microspora did not cluster into one strongly supported group. However, these two species were morphologically very similar, and treated as one genus for now. Roseodiscus sinicus also clustered together with other members of Neolauriomycetaceae. Roseodiscus sinicus was clearly genetically distinct from the type species of Roseodiscus, R. rhodoleucus (Johnston et al. 2014a, b). Future study with involvement of more species and marker genes might support a better resolution of the family.

Maximum likelihood (ML) tree based on 28S rDNA and ITS sequence data for the chalara-like anamorphic fungi in Neolauriomyceteae (Leotiomycetes). Bootstrap support values ≥ 60%, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Cylindrocephalum kendrickii 43800 was chosen as the outgroup. Species names given in bold are the new sequences generated in this study. Species names given in bold and marked in blue color are the new sequences generated from the ex-type material in this study. Ex-type strains are indicated with “T” in the end of the taxa labels
Key to genera of Neolauriomycetaceae
-
1.
Conidiophores with branched apical fertile region………2
-
1.
Conidiophores with single terminal phialides………3
-
2.
Apical conidiogenous apparatus intricate, with numerous tightly aggregated branches and phialides; conidia produced in wet spore cylindrical mass………Lareunionomyces
-
2.
Apical conidiogenous apparatus relatively simple and with less secondary branches and less packed phialides; conidia produced in dry chain………Neolauriomyces
-
3.
Phialide with chalara-like cylindrical collarette; conidia endogenous and cylindrical………Minichalara
-
3.
Phialide with simple funnel-shaped collarette; conidia fusiform………Exochalara
Exochalara W. Gams & Hol.-Jech., Stud. Mycol. 13: 56, 1976.
Colonies effuse, hairy. Anamorph: Conidiophores upright, septate, brown, becoming gradually paler towards the tip, simple or rarely branched, often percurrently proliferating. Conidiogenous cells terminal, integrated, monophialidic; collarette short, funnel–shaped. Conidia hyaline, 1-celled or 1-septate, cohering end-to-end in basipetal chains. Chlamydospores absent. Teleomorph: Unknown.
Typus: Exochalara longissima (Grove) W. Gams & Hol.-Jech.
Ecology/substrate/host: Saprobe on decaying wood and herbaceous stems.
Geographical distribution: Europe (Gams and Holubová-Jechová 1976; Réblová et al. 2011).
Description and illustration: Gams and Holubová-Jechová (1976) and Réblová et al. (2011).
Notes: Exochalara is a monotypic genus and characterized by cylindrical conidiophore with a few stalk cells and a terminal phialide bearing a funnel-shaped collarette, and hyaline, aseptate and fusiform to slightly obclavate conidia in chains (Réblová et al. 2011). In the phylogenetic tree generated from the integrated LSU and ITS dataset (Fig. 3), two strains of Exochalara formed a strongly supported clade within the Neolauriomycetaceae. Morphologically it resembles Minichalara in cylindrical conidiophores with a septate stalk and a terminal phialide, but differs in shape of collarettes and conidia. In Exochalara, collarettes are funnel-shaped and less than 1/3 of length of conidium, and conidia are ellipsoidal with truncated ends (Réblová et al. 2011); while in Minichalara, collarettes are cylindrical and longer than length of one conidium, and they are cylindrical with truncated ends.
Exochalara longissima (Grove) W. Gams & Hol.-Jech., Stud. Mycol. 13: 56, 1976.
Ecology/substrate/host: Saprobe on decaying wood and herbaceous stems.
Geographical distribution: Czech Republic and Netherlands (Gams and Holubová-Jechová 1976; Réblová et al. 2011).
Description and illustration: Gams and Holubová-Jechová (1976) and Réblová et al. (2011).
Notes: The species was fully documented by Réblová et al. (2011).
Lareunionomyces Crous & M.J. Wingf., in Crous et al., Persoonia 36: 387, 2016.
Mycelium consisting of hyaline, smooth, branched hyphae. Anamorph: Conidiophores solitary, erect, subcylindrical, straight, slightly flexuous, unbranched, dark brown, smooth, septate, thick-walled, basal cell slightly swollen, lacking rhizoid, terminating in a pale brown penicillate conidiogenous apparatus. Penicillate conidiogenous apparatus pale brown, smooth; primary branches brown, smooth, subcylindrical to clavate, giving rise to up to one to several secondary branches, pale brown, the upper layers giving rise to several phialides. Conidiogenous cells phialidic, pale brown to medium brown, smooth, flexuous, ampulliform to lageniform, with prominent cylindrical collarette, venter cylindrical to lageniform. Conidia hyaline, smooth, subcylindrical, aseptate, apex bluntly rounded, base truncate, in short chains that form slimy spore masses (Based on Crous et al. 2016b). Teleomorph: Unknown.
Type species: Lareunionomyces syzygii Crous & M.J. Wingf.
Ecology/substrate/host: Saprobe on dead leaves of trees.
Geographical distribution: Australia, China, Columbia, France, Kenya, Malawi and Thailand.
Description and illustration: Crous et al. (2016b).
Notes: The genus Lareunionomyces was created for L. syzygii, which morphologically resembled Neolauriomyces, Sporendocladia and Phialocephala in conidiogenous apparatus and conidial morphology, but was phylogenetically distinct (Wingfield et al. 1987; Jacobs et al. 2003; Crous et al. 2016b, 2018a, b). In Lareunionomyces the conidiogenous apparatus are more intricate, with numerous tightly aggregated branches and phialides, while in other three genera mentioned above, the conidiogenous apparatus are relatively simple and with less secondary branches and less packed phialides (Crous et al. 2016b). Four species, L. eucalypti, L. eucalypticola, S. loeiensis and L. syzygii, are known in the genus and they can be distinguished by conidiogenous apparatus and conidial morphology (Crous et al. 2016b, 2018a, b, 2019). In the phylogenetic trees generated by the integrated LSU and ITS dataset, five accepted species of Lareunionomyces clustered together as one strongly supported monophyletic clade (Figs. 3, 127). In this study, three species including one new species were discovered from China.
Lareunionomyces eucalypti Crous, in Crous et al., Persoonia 40: 357, 2018.
Ecology/substrate/host: Saprobe on dead leaves of Eucalyptus sp.
Geographical distribution: Australia (Crous et al. 2018a, b).
Description and illustration: Crous et al. (2018a, b).
Lareunionomyces eucalypticola Crous, in Crous et al., Persoonia 43: 269, 2019.
Ecology/substrate/host: Saprobe on leaves of Eucalyptus grandis.
Geographical distribution: Columbia (Crous et al. 2019).
Description and illustration: Crous et al. (2019).
Lareunionomyces follicola (P.M. Kirk) W.P. Wu & Y.Z. Diao, Fungal Diversity 116: 334, 2022.
≡ Phialocephala follicola P.M. Kirk, Mycotaxon 23: 337, 1985.
≡ Sporendocladia follicola (P.M. Kirk) M.J. Wingf., Trans. Br. mycol. Soc. 89: 515, 1987.
Geographical distribution: Kenya (Kirk et al. 1985).
Description and illustration: Kirk (1985) and Wu and Diao (2022).
Lareunionomyces kionochaetoides (B. Sutton) W.P. Wu & Y.Z. Diao, Fungal Diversity 116: 334, 2022.
≡ Sporendocladia kionochaetoides B. Sutton, Mycol. Pap. 167: 58, 1993.
Typification: On dead stem of Rubus ellipticus (Rosaceae), Malawi: Mt. Mulanje, Chambe hut track, south-east on track to Lichenya, S1550, E03450, 2000 m a.s.l., 13 Apr. 1991, B.C. Sutton MM28 (IMI347108b, holotype).
Geographical distribution: Malawi (Sutton 1993).
Description and illustration: Sutton (1993) and Wu and Diao (2022).
Lareunionomyces loeiensis Pinruan, Nuankaew & P. Khamsuntorn, in Crous et al., Persoonia 41: 377 2018. Figure 128.

Lareunionomyces loeiensis (Wu15169). a, b Conidiophores and apical fertile region with phialides. c–e Apical fertile region with branches and phialides. f, g Conidiogenous cells. h–m Conidia. Scale bar: 10 µm for a, b; 5 µm for c–m
Description on the natural substrate: Colonies effuse, sparse, brown to reddish brown. Mycelium partly superficial, partly immersed in the substratum, composed of branched, septate, smooth, pale brown to brown hyphae 3–4.5 μm wide. Stroma present, composed of brown, thick- and smooth-walled, and irregular cells. Anamorph: Conidiophores arising from upper layer of the basal stroma, solitary, erect, straight, 150–200 × 12–15 μm, composed of a basal stalk and a branched fertile region; the basal stalk cylindrical, 3–6-septate, smooth- and thick-walled, blackish brown, tapered slightly towards the apex of 8–9 μm; the terminal penicillate conidiogenous apparatus obpyriform, 50–55 × 35–40 μm, consisting of 2–3 orders of branches, the short branches arising in compact closely appressed whorls situated immediately below the septa on the main axis, progressively paler; first order branches dark brown, cylindrical, 6–8 × 3–5 μm; second orders 5–7 × 3–4 μm; third orders 5–6.5 × 2.5–3 μm. Conidiogenous cells formed at the apices of branches, lageniform, subcylindrical, consisting of a basal venter and a tubular collarette within which conidia are formed, 12–14 μm long, 3–3.5 μm wide at the base, tapering to 2 μm wide at the apex, venter and collarette hardly distinguished, pale brown, smooth. Conidia formed in a pale brownish red droplet or white cylindrical mass completely covering the fertile region, cylindrical, 4–5.5 × 1.5–1.6 μm, truncated at both ends, straight, hyaline, aseptate, thin- and smooth-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse slightly soil brown, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Materials examined: China, Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 16 September 2004, Wenping Wu, Wu8109a, Wu8256 and Wu8268b; Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 16 September 2004, Wenping Wu, Wu8131a; Yunnan Province, Baoshan, Lujiang, Bawan, Gaoligongshan, on dead leaves of Quercus sp., 15 October 2003, Wenping Wu, Wu7291d; Yunnan Province, Xishuangbanna, on dead leaves of unidentified tree, 6 December 2018, Wenping Wu, Wu15169. Strains studied: CGMCC3.23405 (= 76001, from Wu15169), 47769 (from Wu7291d), 50576 (from 8109a), 48038 (from Wu8131), and 50629 (from Wu8268b).
Ecology/substrate/host: Saprobe on dead leaves of trees.
Geographical distribution: China and Thailand.
Description and illustration: Crous et al. (2018a, b).
Notes: Lareunionomyces loeiensis differs from other known species in the genus by a combination of morphological characters and ITS sequence (Crous et al. 2016a, b, 2018a, b). Identical ITS sequences of this species were generated from six studied strains, and they were identical to the one from the ex-type strain of L. loeiensis. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 76001 included Lareunionomyces loeiensis (NR_161149, 100% identity), L. eucalypticola (NR_166318, 98% identity), L. eucalypti (NR_10352, 97% identity), L. syzygii (NR_145315, 96% identity), Chalara microspora (FR667228, 93% identity) and many unnamed fungi of Leotiomycetes.
Lareunionomyces minimus W.P. Wu & Y.Z. Diao, sp. nov., Fig. 129, MycoBank MB845265.

Lareunionomyces minimus (Wu17291, holotype). a–d Conidiophores and apical fertile region with phialides. e, f Apical fertile region with branches and phialides. g–m Conidia. Scale bar: 10 µm for a–d, 5 µm for e–m
Etymology: Refers to its small conidia.
Typification: China, Beijing, Huairou, Hongluosi, on dead fruit of Quercus sp., 29 July 2020, Wenping Wu, Holotype HMAS 352188 (= Wu17291), ex-type strain CGMCC3.23440 (= NN77983).
Description on the natural substrate: Colonies effuse, sparse, brown to reddish brown. Mycelium partly superficial, partly immersed in the substratum, composed of branched, septate, smooth, pale brown to brown hyphae 3–4 μm wide. Stroma present, composed of brown, thick- and smooth-walled, and irregular cells. Anamorph: Conidiophores macronematous, mononematous, arising from upper layer of the basal stroma, erect, straight, 110–150 μm long, composed of a basal stalk and a branched fertile region; the basal stalk cylindrical, 2–4-septate, smooth- and thick-walled, blackish brown, tapered slightly towards the upper part, terminating in a pale brown penicillate conidiogenous apparatus; the penicillate conidiogenous apparatus apical, subglobose, obpyriform, 35–46 μm high, 24–40 μm wide, consisting of 2–3 orders of branches, the short branches arising in compact closely appressed whorls situated immediately below the septa on the main axis, first order branches dark brown, second and third orders progressively paler. Conidiogenous cells formed at the apices of branches, lageniform, subcylindrical, consisting of a basal venter and a tubular collarette within which conidia are formed, 7–12 μm long, 2–3 μm wide at the base, tapering to 1.2–1.8 μm wide at the apex, venter and collarette hardly distinguished, pale brown, smooth. Conidia formed in a pale brownish red droplet or white cylindrical mass completely covering the fertile region, short-cylindrical, 3–3.5 × 1–1.2 μm, truncated at both ends, straight, hyaline, aseptate, smooth- and thin-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to dark brown, reverse pale soil brown to brown, sterile, up to 12 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Beijing, Huairou, Hongluosi, on dead fruit of Quercus sp., 29 July 2020, Wenping Wu, Wu17296, Wu17306 and Wu17359; Beijing, Huairou, Beigoucun, on dead fruit of Quercus sp., 9 October 2022, Wenping Wu, Wu18142; Yunnan Province, Kunming, Heilongtan Park, on dead fruit of ?Quercus sp., 19 October 2003, Wenping Wu, Wu7237a. Living strains: CGMCC3.23372 (= NN 47774, from Wu7237a), 77957 (from Wu17291), 77958 (from Wu17296), 78039 (from Wu17359a), 78015 (from Wu17359b), and 78733 (from Wu18142).
Ecology/substrate/host: Saprobe on dead fruit of Quercus spp.
Geographical distribution: China.
Notes: Lareunionomyces minimus differs from other three known species in the genus by smaller conidia (Crous et al. 2016b, 2018a, b, 2019). Compared with L. minimus, L. eucalypti and L. loeiensis have longer conidia. Similar to L. minimus, L. eucalypticola and L. syzygy also produce the short-cylindrical conidia in smaller size, but their conidiogenous cells are not incurved and closely packed together; furthermore, the conidiogenous cells in L. eucalypticola are hyaline or subhyaline.
Lareunionomyces syzgii has similar-sized conidia, but fertile region in this species are incurved and densely packed, and conidiogenous cells are colored (Crous et al. 2016b, 2018a, b). In L. eucalypti and L. loeiensis, conidia are longer than those of L. minimus (Crous et al. 2016b, 2018a, b, 2019). Identical ITS sequences of this species were generated from seven studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 77957 included Lareunionomyces loeiensis (NR_161149, 98% identity), L. eucalypticola (NR_166318, 97% identity), L. eucalypti (NR_10352, 97% identity), L. syzygii (NR_145315, 96% identity), Neolauriomyces eucalypti (NR_160353, 94% identity), Chalara microspora (FR667228, 93% identity) and many unnamed fungi of Leotiomycetes.
Lareunionomyces syzygii Crous & M.J. Wingf., in Crous et al., Persoonia 36: 386, 2016. Figure 130.

Lareunionomyces syzygii (Wu16875). a–d Conidiophores and apical fertile region with phialides. e Basal stroma. f, g, i, j Apical fertile region with branches and phialides. h, k–m Conidia. Scale bar: 20 µm for a, b; 10 µm for b–d; 5 µm for e–m
Description on the natural substrate: Mycelium partly immersed and partly superficial, consisting of hyaline, smooth, branched, 2–3 μm diam hyphae. Anamorph: Stroma present, composed of dark brown, thick-walled, and irregular cells. Setae absent. Conidiophores solitary or 2–3 in groups from the basal stroma, erect, straight or slightly curved, (85–)140–170 × 5–5 μm, composed of a basal stalk and a apical penicillate conidiogenous apparatus; the basal stalk cylindrical, (50–)90–135 × 5–7 μm, unbranched, subcylindrical, dark brown, smooth, 3–4-septate, thick-walled, basal cell slightly swollen and up to 10 μm wide; the apical penicillate conidiogenous apparatus pale to medium brown, globose, subglobose, 32–36 μm high, 33–28 μm wide, consisting of 2–3 orders of branches, the short branches arising in compact closely appressed whorls situated immediately below the septa on the main axis; primary branches brown, smooth, subcylindrical to clavate, 7–10 × 5–6 μm, giving rise to up to 8 secondary branches, pale brown, subcylindrical to clavate, 5–6.5 × 2.5–5 μm; tertiary branches 5–6 × 2–3 μm, pale brown, giving rise to several phialides. Conidiogenous cells phialidic, subulate, 10–14 × 2–2.5 μm, pale brown, flexuous, venter cylindrical, with prominent collarette, apex 1.2–1.5 μm diam. Conidia formed in a pale brownish red droplet or white cylindrical mass completely covering the fertile region, subcylindrical, 3–4 × 1–1.5 μm, apex bluntly rounded, base truncated, hyaline, aseptate, smooth- and thin-walled. Teleomorph: Unknown.
Culture characteristics: Colony effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse pale soil brown, sterile, up to 11 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Jiangsu Province, Wuxi, Wuxi Forestry Park, on dead fruit of Cyclobalanopsis sp., 25 Aug. 2019, Wenping Wu, Wu16875. Strains studied: CGMCC3.23428 (= 77239, from 16875), 77210 (from Wu16875).
Ecology/substrate/host: Saprobe on dead fruit of Quercus spp.
Geographical distribution: China and France (Crous et al. 2016b).
Notes: Lareunionomyces syzygii differs from other four known species in the genus, L. eucalypti, L. eucalypticola, L. loeiensis and L. sinensis, by a combination of morphological characters and ITS sequence (Crous et al. 2016b, 2018a, b, 2019). It differs from L. eucalypti and E. loeiensis by smaller conidia. Similar to L. syzygy, L. eucalypticola and L. minimus also produce short-cylindrical conidia in smaller size, but conidiogenous cells of L. eucalypticola are hyaline or subhyaline in the two latter species; branches and conidiogenous cells of L. minimus are closely packed together.
Identical ITS sequences were generated from two studied strains. Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 77210 included Lareunionomyces syzygii (NR_145315, 100% identity), L. eucalypti (NR_10352, 97% identity), L. loeiensis (NR_161149, 97% identity), L. eucalypticola (NR_166318, 96% identity), Neolauriomyces eucalypti (NR_160353, 96% identity), Chalara microspora (FR667228, 94% identity) and many unnamed fungi of Leotiomycetes.
Minichalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845260.
Etymology: Refers to its similarity to Chalara in shape of conidiophores, phialide and conidia.
Type species: Minichalara aseptata W.P. Wu & Y.Z. Diao.
Colonies effuse, pale brown, superficial, hairy. Mycelium partly immersed and partly superficial, composed of pale to medium brown, septate and branched hyphae with smooth and thin walls. Anamorph: Stroma absent. Setae absent. Conidiophores arising from superficial hyphae or aggregated cells, solitary or in small group, erect, straight or slightly curved, simple, consisting of a short basal stalk and a terminal phialide, obclavate, subcylindrical, pale to medium brown, smooth. Conidiogenous cells integrated, terminate, determinate, phialidic, erect, straight, obclavate, lageniform, pale to medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt, often marked by a darker and pronounced constriction; venters subcylindrical, long ellipsoidal, lageniform, pale brown; collarettes cylindrical, medium brown, significantly darker in the lower part, smooth. Conidia endogenous, extruded in short chains, cylindrical, with truncated of flattened ends, hyaline, aseptate. Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves of plant, and fruitbody of other fungus.
Geographical distribution: China, Czechoslovakia, Netherland and USA.
Notes: Morphologically Minichalara is characterized by reduced conidiophores composed of a short basal stalk and a terminal phialide in pale brown color, obclavate to lageniform phialides, abrupt transition from a long venter to a short collarette, and hyaline, aseptate and cylindrical conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). It resembles Chalara s. str., Constrictochalara, Cylindrochalara and Stipitochalara, but differs from them in reduced conidiophores, obclavate to lageniform phialides and narrower conidia (< 1.5 µm). In addition, these genera are phylogenetically distinct and scattered among different clades in the phylogenetic trees generated from different datasets (Figs. 2, 3). Two species, M. aseptata and M. microspora, are accepted under the genus. However, in the phylogenetic trees generated from two integrated LSU and ITS datasets (Figs. 3, 127), these two species were not resolved as a monophyletic clade. Furthermore, their ITS sequences had rather low identity (89%). There were treated as one genus for now, however future phylogenetic analysis with involvement of additional gene markers might show they represent two different genera.
Minichalara aseptata W.P. Wu & Y.Z. Diao, sp. nov., Fig. 131, MycoBank MB845266.

Minichalara aseptata (Wu8110, holotype). a–l conidiophores and phialides. m conidia. Scale bar 5 µm
Etymology: Refers to its aseptate conidia.
Typification: China: Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 18 September 2004, Wenping Wu, Holotype HMAS352190 (= Wu8110), ex-type strain CGMCC3.23377 (= NN48017).
Description on the natural substrate: Colonies effuse, scattered, pale brown, minute. Mycelium partly immersed and partly superficial, composed of pale brown, septate and branched hyphae with smooth and thin wall, 2–3 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores macronematous, micronematous, arising from superficial hyphae or aggregated cells, solitary or 2–6 in small groups, erect, straight or slightly curved, simple, composed of a 1–3-celled basal stalk and a terminal phialides, obclavate, subcylindrical, 20–38 µm long, versicolorous, pale brown, medium brown in the transition area from venter to collarette, smooth- and thin-walled. Conidiogenous cells integrated, terminal, determinate, phialidic, erect, straight, obclavate to lageniform, 22–28 µm long, versicolorous, pale to medium brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt, often marked by a darker and pronounced constriction; venters subcylindrical to obclavate, 12–15 µm long and 2–3 µm wide, pale brown; collarettes cylindrical, 5–7 × 1.5 µm, pale brown to medium brown, smooth; ratio of mean lengths of collarette and venter = 0.4:1. Conidia endogenous, extruded in short chains cylindrical, 5–7.5 × 1–1.2 µm, both ends truncated or flattened, hyaline, aseptate, thin- and smooth-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white to brown, reverse soil brown to brown, sterile, up to 5 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Hubei Province, Shengnongjia, on dead leaves of unidentified broad leaf tree, 18 September 2004, Wenping Wu, Wu8271b and Wu8299. Living strain: CGMCC3.23450 (= 50633, from Wu8271b).
Ecology/substrate/host: Saprobe on dead leaves of broad leaf tree.
Geographical distribution: China.
Notes: Minichalara aseptata differs from C. microspora, the only known species in the genus in its versicolorous phialides with a pronounced constriction in the lower part of collarettes (Nag Raj and Kendrick 1975). In addition, collarettes in M. aseptata (5–7 × 1.5 μm) are much shorter in average than those from M. microspora (6–17 × 1–2.5 μm).
ITS and partial LSU sequences of this species were generated from two studied strains, and they were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 48017 included Neolauriomyces eucalypti (NR_160353, 90% identity), Lareunionomyces eucalypti (NR_10352, 90% identity), L. syzygii (NR_145315, 90% identity), L. loeiensis (NR_161149, 88% identity), L. eucalypticola (NR_166318, 88% identity), Chalara microspora (FR667228, 89% identity) and many unnamed fungi of Leotiomycetes.
Minichalara microspora (Coda.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845297.
≡ Fusidium clandestimum var. microsporum Corda, Icon. Fung. (Prague) 2: 43, 1838.
≡ Chalara microspora (Corda.) S. Hughes, Can. J. Bot. 36: 747, 1958.
≡ Cylindrium clandestimum var. microsporum (Corda.) Sacc., Syll. Fung. (Abellini) 4: 37, 1886.
≡ Fusidium microsporum (Corda) Mussat, in Saccardo, Syll. Fung. (Abellini) 15: 145, 1901).
Description on the natural substrate: Stroma absent. Setae absent. Anamorph: Conidiophore reduced, composed of a 2(–4)-celled basal stalk and a terminal, phialide, obclavate, lageniform, 18–36 μm long, pale brown. Conidiogenous cells phialidic, obclavate to lageniform, 18–36 µm long, concolorous, pale brown, smooth, consisting of a venter and a collarette; transition from venter to collarette abrupt; venters subcylindrical, 10–16 × 2.5–6.5 µm; collarettes cylindrical, 6–17 × 1–2.5 µm. Conidia endogenous, extruded in short chains, cylindrical, 3–8.5 × 1–1.5 µm, ends truncated, hyaline, aseptate (Adapted from Nag Raj and Kendrick 1995). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead wood and bark of Fraxinus, Quercus, Picea etc., or needle of Pinus sylvestris, or fruitbody of other fungi such as Niesslia exosporioides on Elymus arenarius, or from soil under Picea abies.
Geographical distribution: Czechoslovakia, Netherland, Poland, Sweden, UK and USA (Nag Raj and Kendrick 1975; Holubová-Jechová 1984).
Description and illustration: Hughes (1958), Nag Raj and Kendrick (1975), and Holubová-Jechová (1984).
Notes: ITS sequences of M. microspora were known from three different strains occurring on coniferous needles, and these sequences were nearly identical (1 bp difference). Morphologically this fungus is similar to Minichalara aseptata in conidiophores and phialidic, but differs in concolorous phialides, no constriction at the lower part of collarettes, and different ITS sequence. No specimen was examined in this study, and the above description was adapted from Nag Raj and Kendrick (1975).
Neolauriomyces Crous, in Crous et al., Persoonia 40: 359, 2018.
Mycelium consisting of hyaline, smooth, branched hyphae. Anamorph: Conidiophores solitary, erect, subcylindrical, straight, slightly flexuous, unbranched, dark brown, smooth, septate, thick-walled, basal cell slightly swollen, lacking rhizoid, terminating in a pale brown penicillate conidiogenous apparatus. Penicillate conidiogenous apparatus pale brown, smooth; primary branches subcylindrical to doliform, medium brown, smooth; secondary branches doliform to subcylindrical, medium brown, smooth, giving rise to 1–2 phialides. Conidiogenous cells phialidic, ampulliform, medium brown, smooth, with apical cylindrical collarette. Conidia endogenous, extruded in short and dry chains, hyaline, smooth, subcylindrical, aseptate, ends truncate (Adapted from Crous et al. 2018b). Teleomorph: Unknown.
Type species: Neolauriomyces eucalypti Crous.
Ecology/substrate/host: Saprobe on dead leaves of trees.
Geographical distribution: Australia and China.
Description and illustration: Crous et al. (2018b).
Notes: The genus Neolauriomyces was created for N. eucalypti, which morphologically resembled Lareunionomyces in conidiogenous apparatus and conidial morphology, but differed phylogenetically (Wingfield et al. 1987; Crous and Wingfield 1994; Jacobs et al. 2003; Crous et al. 2018b). In Lareunionomyces conidiogenous apparatus are more intricate, with numerous tightly aggregated branches and phialides, and conidia are formed in slimy droplet or cylindrical slimy mass; while in Neolauriomyces, conidiogenous apparatus are much simple with 1 primary branch and 1–2 secondary branches, each bearing 1–3 ampulliform-shaped conidiogenous cells, and conidia are formed in short and dry chains (Crous et al. 2016b, 2018b). Three species, including two new species, are accepted under the genus (Fig. 127).
Neolauriomyces beijingensis W.P. Wu & Y.Z. Diao, sp. nov., Fig. 132, MycoBank MB845267.

Neolauriomyces beijingensis (Wu17316, holotype). a–d Conidiophores and apical fertile region with phialides. e–i Apical fertile regions with branches and terminal phialides. j, k Conidia. Scale bar: 10 µm for a–d, 5 µm for e–k
Etymology: Refer to the location Beijing, China, where the type specimen was collected.
Typification: China, Beijing, Huairou, Hongluosi, on dead cupule of Quercus sp., 29 July 2020, Wenping Wu, Holotype HMAS 352228 (= Wu17316a), ex-type strain CGMCC3.23462 (= NN78013).
Description on the natural substrate: Colonies effuse, sparse, brown to black. Mycelium partly superficial, partly immersed in the substratum, composed of branched, septate, smooth, pale brown to brown hyphae 2–4 μm wide. Anamorph: Stroma absent, or poorly developed if present and composed of a few brown, thick- and smooth-walled, irregular cells. Conidiophores macronematous, mononematous, arising from upper layer of the basal stroma or directly from superficial hyphae, solitary, simple, erect, straight or slightly flexuous, composed of a basal stalk and a penicillate conidiogenous apparatus, 115–170 × 5–7 μm, smooth, dark brown to blackish brown at the base, medium brown at the upper part; the basal stalk cylindrical, 90–133 μm long, with a swollen basal cell up to 8 μm wide, 5–7-septate, dark brown, becoming paler towards the upper part; the fertile region apical, 20–25 μm high, 20–35 μm wide, consisting of branches with terminal phialides in loose arrangement with a mono- to bi-verticillate branching pattern; primary branches cylindrical, 4–6 × 3–3.7 μm, medium to dark brown, smooth, thick-walled; secondary and third branches cylindrical, 4–5 × 3–3.5 μm, medium brown, smooth, thin- or thick-walled, bearing 1–2 conidiogenous cells. Conidiogenous cells formed at the apices of branches, ampulliform, 8–13 μm long, pale brown to medium brown, smooth, consisting of a basal venter and a tubular collarette within which conidia are formed; transition from venter to collarette abrupt; venters cylindrical, 4–7 × 2.7–3.2 μm; collarettes cylindrical, 2–4 × 1.2–1.3 μm. Conidia endogenous, formed in short and dry chains, cylindrical, 3–4 × 1 μm, both ends obtuse, rounded or flattened, straight, hyaline, smooth, aseptate, thin- and smooth-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, dark brown, reverse brown, sterile, up to 9 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Beijing, Huairou, Hongluosi, on dead cupule of Quercus sp., 11 June 2020, Wenping Wu, WuBJ05, WuBJ06, WuBJ07, WuBJ08, WuBJ17 and WuBJ20; Beijing, Huairou, Beigoucun, on dead cupule of Quercus sp., 9 October 2022, Wenping Wu, Wu18122. Living strains: ex-type strain CGMCC3.23462 (= 78013, from Wu17316) and 77425 (from WuNJ17), 78696 and 78697 (from Wu18122).
Ecology/substrate/host: Saprobe on decaying cupule of Quercus sp.
Geographical distribution: China.
Notes: Neolauriomyces beijingensis is morphemically similar to the other two species of the genus, N. eucalypti and N. crousii, but differs on morphology of conidiogenous apparatus, shape of phialides, and size of conidia (Crous et al. 2018b). In N. eucalypti and N. crousii, conidiogenous apparatus are dark brown, with fewer phialides, and phialides are with a subglobose or ellipsoidal venters, and conidia are cylindrical and larger in size (Crous et al. 2018b). While in N. beijingensis, conidiogenous apparatus are with many radiantly arranged conidiogenous cells, venters are cylindrical or conical, collarettes are much shorter, and conidia are shorter and narrower.
ITS and partial LSU sequences of this species were generated from two studied strains, and they were identical. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 78013 included Neolauriomyces eucalypti (NR_160353, 95% identity), Lareunionomyces eucalypti (NR_10352, 93% identity), L. eucalypticola (NR_166318, 93% identity), L. loeiensis (NR_161149, 93% identity), L. syzygii (NR_145315, 93% identity), Pseudofabraea citrcicarpa (NR_154319, 91% identity) and many unnamed fungi of Leotiomycetes.
Neolauriomyces crousii W.P. Wu & Y.Z. Diao, sp. nov., Fig. 133, MycoBank MB845268.

Neolauriomyces crousii (Wu8268a, holotype). a–d Conidiophores and apical fertile region with phialides. e Basal stroma from which the conidiophore is formed. f–m Conidia. Scale bar: 5 µm
Etymology: Named after the CBS mycologist Pedro Crous, who established the genus Neolauriomyces.
Typification: China, Hubei Province, Shengnongjia, on wood of undetermined plant, 16 September 2004, Wenping Wu, Holotype HMAS 352229 (= Wu8268a), ex-type strain CGMCC3.23386 (= NN50673).
Description on the natural substrate: Colonies effuse, sparse, brown to black. Mycelium partly superficial, partly immersed in the substratum, composed of branched, septate, smooth, pale brown to brown hyphae 2–4 μm wide. Anamorph: Stroma present, composed of brown, thick- and smooth-walled, irregular cells. Conidiophores macronematous, mononematous, arising from upper layer of the basal stroma, simple, erect, straight, composed of a basal stalk and an apical penicillate conidiogenous apparatus, 85–150. The basal stalk cylindrical, 65–120 × 4.5–5.5 μm, 3–6-septate, dark brown to blackish brown at the base, medium brown at the upper part, smooth. The fertile penicillate conidiogenous apparatus apical, consisting of branches with terminal phialides in loose arrangement with a mono- to bi-verticillate branching pattern; primary branches cylindrical, medium to dark brown, smooth, thick-walled; secondary branches cylindrical, medium brown, smooth, thin- or thick-walled, bearing 1–2 conidiogenous cells. Conidiogenous cells formed at the apices of branches, ampulliform, 10–14 μm long, concolorous, pale brown to medium brown, smooth, consisting of a basal venter and a tubular collarette within which conidia are formed; transition from venter to collarette abrupt; venters subglobose, 6–8 × 3–4 μm; collarettes cylindrical, 5–6 × 1.5 μm. Conidia endogenous, extruded in short and dry chains, cylindrical, 4.5–5.5 × 1–1.2 μm, both ends truncated or flattened, straight, hyaline, aseptate, thin- and smooth-walled. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, grey to brown, reverse pale brown to dark brown, sterile, up to 4 mm on PDA at 25 °C in 4 weeks.
Other materials examined: China, Hubei Province, Shengnongjia, on wood of undetermined plant, 16 September 2004, Wenping Wu, Wu8262; Yunnan Province, Kunming, on rotten wood, 12 September 2002, Wenping Wu, Wu7233. Living strain: ex-type strain CGMCC3.23386 (= NN50673).
Ecology/substrate/host: Saprobe on rotten wood.
Geographical distribution: China.
Notes: Neolauriomyces crousii is morphemically similar to the type species N. eucalypti on appearance of conidiophores and conidiogenous cells, and shape and size of conidia, but differs by relatively longer conidia and different ITS sequence (Crous et al. 2018b).
Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain 50673 included Neolauriomyces eucalypti (NR_160353, 95% identity), Lareunionomyces eucalypti (NR_10352, 93% identity), L. eucalypticola (NR_166318, 93% identity), L. loeiensis (NR_161149, 93% identity), L. syzygii (NR_145315, 93% identity), Chalara microspora (FR667226, 92% identity) and many unnamed fungi of Leotiomycetes.
Neolauriomyces eucalypti Crous, in Crous et al., Persoonia 40: 359, 2018.
Ecology/substrate/host: Saprobe on Eucalyptus leaf litter.
Geographical distribution: Australia.
Description and illustration: Crous et al. (2018a, b).
Notes: Neolauriomyces eucalypti is the type species of the genus and was well documented by Crous et al. (2018a, b).
Leotiomycetes genera incertae sedis
Several other discomycete genera were known with chalara-like anamorphs, such as Ascoconidium, Bioscypha, Chalarodendron, Didonia, Phaeoscypha, Rodwayella and Tapesina. However, none of these genera was with living strain or DNA sequence available for phylogenetic analyses, thus their phylogenetic relationships with other chalara-like fungi of Leotiomycetes remain to be studied in future.
Ascoconidium Seaver, Mycologia 34(4): 414, 1942.
= Sageria A. Funk, Can. J. Bot. 53: 1196, 1974.
Colonies black, erumpent. Anamorph: Conidiophores arising from a pseudoparenchamytous basal stroma and becoming erumpent through the periderm, simple, short, sparsely septate, terminating in a phialidic conidiogenous cell. Conidiogenous cells sessile, aggregated in densely packed sori, clavate to subcylindrical, brown, thick-walled. Conidia cylindrical, septate, hyaline, formed single and successively. Teleomorph: described under Sageria, see Funk 1975).
Type species: Ascoconidium purpurascens (Ellis & Everh.) Rossman.
Ecology/substrate/host: Saprobe or minor pathogens on dead branches of trees such as Castanea dentata, Tsuga heterophylla etc.
Geographical distribution: Canada and USA (Nag Raj and Kendrick 1975; Verkley 1999).
Description and illustration: Seaver (1942), Funk (1966a, b, 1975), and Nag Raj and Kendrick (1975).
Notes: Ascoconidium differs from Chalara s. lat. and Sporoschisma in several aspects, such as its phialides lack a well-differentiated venter and collarette, conidiogenous loci near the base of the phialide, thick-walled phialide ruptured at maturity by a vertical split at the apex (Funk 1966a, b; Nag Raj and Kendrick 1975). Two species, A. purpurascens and A. tsugae, are known for the genus and can be distinguished by septation and size of conidia. Both species are known with the Ascoconidium anamorphs and helotiaceous discomycete teleomorphs (Seaver 1942; Funk 1975; Verkley 1999; Johnston et al. 2014a, b). The teleomorphs are helotiaceous discomycetes. It was recommended to protect the asexually typified Ascoconidium (1942) over sexually typified Sageria (1975) by Johnston et al. (2014a, b). No DNA sequence was available for molecular phylogenetic analyses, thus its phylogenetic relationship with other chalara-like fungi remains to be studied in future.
Key to species of Ascoconidium:
-
1.
Conidia with transverse septa only, consistently 3-septate, 30–35 × 8–10 µm………A. purpurascens
-
1.
Conidia with both transverse and vertical septa, 3–7-septate, 40–60 × 10–14 µm………A. tsugae
Ascoconidium purpurascens (Ellis & Everh.) Rossman, in Johnston, Seifert, Stone, Rossman & Marvonová, IMA Fungus 5: 94, 2014.
≡ Dermatea purpurascens Ellis & Everh., J. Mycol. 4(10): 100, 1888.
= Ascoconidium castaneae Seaver, Mycologia 34(4): 414, 1942.
≡ Pezicula purpurascens (Ellis & Everh.) Seaver, Mycologia 34(4): 414, 1942.
≡ Sageria purpurascens (Ellis & Everh.) Verkley, Stud. Mycol. 44: 150, 1999.
Ecology/substrate/host: Saprobe or minor pathogens on dead branches of Castanea dentata.
Geographical distribution: USA (Nag Raj and Kendrick 1975).
Description and illustration: Seaver (1942), Funk (1966a, 1975), and Nag Raj and Kendrick (1975).
Notes: The fungus was described with both anamorphs and teleomorph in literatures. It differs from the other known species in the genus, A. tsugae (conidia 3–7-septate, 40–60 × 10–14 µm) by possessing smaller phialides, shorter conidia that are only 3-septa and not muriform (Nag Raj and Kendrick 1975).
Ascoconidium tsugae A. Funk, Can. J. Bot. 44: 219, 1966.
≡ Sageria tsugae A. Funk, Can. J. Bot. 53(12): 1196, 1975.
Ecology/substrate/host: Saprobe or minor pathogens of Tsuga heterophylla.
Geographical distribution: Canada (Nag Raj and Kendrick 1975; Verkley 1999).
Description and illustration: Funk (1966b, 1975) and Nag Raj and Kendrick (1975).
Notes: The fungus was fully described with both anamorphs and teleomorph in literatures.
Bioscypha Syd., Annls Mycol. 25(1/2): 102, 1927.
Type species: Bioscypha cyatheae Syd.
Ecology/substrate/host: Pathogens on pinnae of ferns such as Cyathea sp.
Geographical distribution: Costa Rica and Columbia (Samuels and Rogerson 1990).
Description and illustration: Sydow (1927) and Samuels and Rogerson (1990).
Notes: Bioscypha was known with two species, B. cyatheae and B. pteridicola, both of which occurred on pinnae of ferns in the American tropics. The chalara-like anamorphs were connected for both species. No living strain or DNA sequence was available for phylogenetic analyses. However, Samuels and Rogerson (1990) stated that ‘the Chalara found associated with B. cyatheae is undoubtedly the anamorph of this discomycete because the conidiophores arise directly from hyphae of the outer excipulum, and because the apothecia of the second species, B. pteridicola, are associated with a similar Chalara’. Carpenter (1981) discussed the genus Bioscypha and its relationship to Crocicreas and Bisporella, in which chalara-like anamorphs were also connected.
Zymochalara, a recently described chalara-like genus with two known species as pathogens of ferns, is closely related to the anamorphs of Bioscypha (Guatimosim et al. 2016). No teleomorph was discovered for Zymochalara, but its affinity with Pezizellaceae was confirmed by the phylogenetic analyses. The anamorphs from both known species, Z. cyatheae and Z. lygodii, produced very similar morphology as those described in Bioscypha and occurred on the same host genera. It is most likely that these two genera, Bioscypha and Zymochalara, are congeneric, but this needs to be confirmed by future phylogenetic analyses when the DNA sequence will be available for species of Bioscypha.
Bioscypha cyatheae Syd., Annls Mycol. 25(1/2): 103, 1927.
Ecology/substrate/host: Pathogens on pinnae of Cyathea sp.
Geographical distribution: Costa Rica (Samuels and Rogerson 1990).
Description and illustration: Sydow (1927) and Samuels and Rogerson (1990).
Notes: Samuels and Rogerson (1990) described the anamorph as ‘Conidiophores are 50–55 µm long and brown; conidia are minute, rectangular, about 3.5 × 1.5 µm, colorless. Conidiophores also form in abundance on the surface of the pinna in crustose areas formed of densely intertwined hyphae’. The conidia in this species are much smaller in size than those in B. pteridicola 7–8.8 × 1.7–2.5 µm, Zymochalara cyatheae (6–10 × 1.5–3 μm) and Z. lygodii (6.5–12 × 1.5–3 μm).
Bioscypha pteridicola Samuels & Rogerson, Brittonia 42(2): 110, 1990.
Ecology/substrate/host: Pathogens on pinnae of ferns such as Cyathea sp.
Geographical distribution: Costa Rica and Columbia (Samuels and Rogerson 1990).
Description and illustration: Sydow (1927) and Samuels and Rogerson (1990).
Notes: Samuels and Rogerson (1990) described the anamorph of Bioscypha pteridicola as ‘Chalara conidiophores arise from hyphae of the outer ectal excipulum and are identical to those formed by B. cyatheae. The conidia are rectangular, ( 3.5)7–8.8 × 1.7–2.5(3.5) µm.
Chalarodendron C.J.K. Wang & B. Sutton, Mycologia 76: 569, 1984.
Anamorph: Conidiomata synnematous, unbranched, brown to black. Conidiophores of the Chalara-type formed along the whole length of the synnemata, unbranched with terminal phialidic conidiogenous cells. Phialides with long, tubular collarettes. Conidia endogenously produced in the collarettes, rectangular, hyaline, aseptate, in short chain. Teleomorph unknown.
Type species: Chalarodendron fuscum C.J.K. Wang & B. Sutton.
Ecology/substrate/host: Saprobes on decaying wood.
Geographical distribution: USA.
Description and illustration: Wang and Sutton (1984).
Notes: The genus Chalarodendron was established for the synnematous fungus C. fuscum which is similar to Chalara s. lat., but differs in producing synnemata. The genus remains to be monotypic. No living culture was available for phylogenetic analyses, and its relationship with other chalara-like fungi is unclear. The synnematous conidiomata was also described for Chalara longipes in pure culture (Koukol 2001).
Chalarodendron fuscum C.J.K. Wang & B. Sutton, Mycologia 76: 569, 1984.
Description on the natural substrate: Anamorph: Conidiomata synnematous, unbranched, brown to black, up to 800 μm high, 22–42 μm wide. Conidiophores of the chalara-type formed along the whole length of the synnemata, unbranched, septate, light brown, smooth, with terminal phialidic conidiogenous cells. Phialides pale brown, consisting of a venter and a long, tubular collarettes; venter 15–20 × 3–4 μm; collarettes cylindrical 4–7.5 × 1.25–1.5 μm. Conidia endogenously produced in the collarettes, rectangular, hyaline, aseptate, 3.5–6.5 × 1 μm in short chain (Adapted from Wang and Sutton 1984). Teleomorph: Unknown.
Ecology/substrate/host: Saprobes on decaying wood.
Geographical distribution: USA (Wang and Sutton 1984).
Description and illustration: Wang and Sutton (1984).
Didonia Velen., Monogr. Discom. Bohem. (Prague): 296, 1934.
Type species: Didonia picea Velen.
Ecology/substrate/host: Saprobe on dead plant material.
Geographical distribution: Europe (Svrček 1992b).
Description and illustration: Velenovský (1934) and Svrček (1992b).
Notes: Seven species were known for the genus Didonia (Svrček 1992a, b). Two of these species were associated with chalara-like anamorph, including the type species D. picea and D. betulina (Svrček 1992a, b). The chalara-like anamorphs of these two species were with sessile conidiogenous cells on apothecium, however no conidia were described. Phylogenetic relationships of these two species with other chalara-like species having aseptate conidia remains to be studied in future.
Excluded species from Leotiomycetes
Four described species of Chalara s. lat. such as C. breviclavata, C. hyalina, C. schoenoplecti and C. vaccinii were phylogenetically distinct from other chalara-like fungi, and should be excluded from Leotiomycetes and reclassified. In addition, the recently described fungus Pseudosclerococcum golindoi with apothecial ascomata and chalara-like anamorph is also presented for comparison with Chalara phaeospora, both species produce pale colored conidia from cylindrical collarettes.
Chaetosphaeriaceae Réblová, M.E. Barr & Samuels, Sydowia 51(1): 56, 1999.
(Chaetosphaeriales, Sordariomycetidae)
Type genus: Chaetosphaeria Tul. & C. Tul.
Ecology/Substrate/host: Saprobes on decaying leaves, fruits, branches, stems, barks and wood of herbaceous or woody plants, terrestrial or submersed.
Geographical distribution: Cosmopolitan, very frequent in temperate, subtropical, and tropical climates.
Description and illustration: Réblová et al. (1999) and Wu and Diao (2022).
Chalarosphaeria W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845262.
Etymology: Refers to its chalara-like anamorph.
Type species: Chalarosphaeria breviclavata (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao
Colonies gregarious, effuse, brown to dark brown. Anamorph: Conidiophores solitary, scattered or a few aggregated at the base, erect, straight or slightly flexuous, simple, cylindrical to subcylindrical, smooth, dark brown, becoming pale brown to medium brown towards the apex, septate, terminating in a phialide. Conidiogenous cells integrated, terminal, phialidic, subcylindrical, smooth, pale brown to almost subhyaline at the apex; transition from venter to collarette abrupt, strongly constricted between venter and collarette; venter subcylindrical; collarette subcylindrical to obconical. Conidia endogenous, extruded in short chains, ellipsoidal to shortly clavate, rounded at the apex, truncate at the base with minute marginal frill, hyaline, aseptate, smooth, guttulate. Teleomorph: Ascomata perithecia, superficial, attached to the substrate, without subiculum, gregarious, subglobose, black, smooth; ostiole region slightly conical; wall composed of rather thick-walled, dark pigmented cells in textura epidermoidea or angularis, loosely covered with phialophores of chalara-like anamorph. Paraphyses hyaline, septate, slightly branched at basal part. Asci thin-walled, cylindrical to clavate, with thin apical part. Ascospore obliquely uniseriate, fusiform to elongate fusiform, with rounded ends, hyaline, at maturity 1-sepate, not constricted at septa, guttulate (Adapted from Holubová-Jechová 1984).
Ecology/substrate/host: Saprobe on dead plant material.
Geographical distribution: China, Czechoslovakia, Canada, Poland and Romania.
Chalarosphaeria breviclavata (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845298, Fig. 134.

Chalarosphaeria chalaroides (Wu1760c). a Conidia. b Conidiophores and conidiogenous cells. Scale bar: 20 µm
≡ Chalara breviclavata Nag Raj & W. Kendr., Monogr. Chalara Allied Genera: 95, 1975.
= Chaetosphaeria chalaroides Hol.-Jech., Folia geobot. Phytotax. 19(4): 396, 1984.
Colonies effuse, brown to dark brown, hairy. Mycelium partly immersed and partly superficial, composed of pale brown to medium brown, septate and branched hyphae, smooth, thin- or medium thick-walled, 2.5–3.5 µm wide. Anamorph: Conidiophores solitary, scattered or a few aggregated at the base, erect, straight or slightly flexuous, simple, cylindrical to subcylindrical, 100–150 µm long, 5–6 µm wide, smooth, dark brown, becoming pale brown to medium brown towards the apex, 3–7-septate, terminating in a phialide. Conidiogenous cells integrated, terminal, phialidic, subcylindrical, 40–50 µm long, smooth, pale brown to almost subhyaline at the apex; transition from venter to collarette abrupt, strongly constricted between venter and collarette; venter subcylindrical, 18–22 µm long, 5–7 µm wide; collarette subcylindrical to obconical, 25–40 µm long, 4–6 µm wide; ratio of mean lengths of collarette and venter = 1.6. Conidia endogenous, extruded in short chains, ellipsoidal to shortly clavate, 10–20 × 3.5–4.5 µm, rounded at the apex, truncate at the base with minute marginal frill, hyaline, aseptate, smooth. Teleomorph: Unobserved.
Materials examined: China, Guangxi Province, Shangsi, Shiwandashan, Wangle, on dead branches of unidentified plant, 2 January. 1997, Wenping Wu, Wu1322b; Jilin Province, Changbaishan, on rotten wood of Betula sp., 5 September 1998, Wenping Wu, Wu1760c.
Ecology/substrate/host: Saprobe on dead branches of different trees, including Alnus glutinosa, Betula verrucosa, Carpinus betulus, Fagus sylvatica, Fraxinus excelsior, Quercus petraea, Q. robur, Tilia cordata, Betula sp., ?Fraxinus sp. and other plants
Geographical distribution: Canada, China, Costa Rica, Czechoslovakia, Poland, Puerto Rico and Romania (Nag Raj and Kendrick 1975; Holubová-Jechová 1984).
Description and illustration: Nag Raj and Kendrick (1975) and Holubová-Jechová (1984).
Notes: The new genus Chalarosphaeria is created for Chaetosphaeria chalaroides (anamorph Chalara breviclavata), which is phylogenetically distinct from Chaetosphaeria and Paragaeumannomyces, two closely related genera. The anamorphic fungus Chalara breviclavata was originally described on wood of ?Fraxinus collected from Canada (Nag Raj and Kendrick 1975). Holubová-Jechová (1984) described its teleomorph as Chaetosphaeria chalaroides Hol.-Jech., which was based on an observation of their occurrence on the same substrate. Fernández et al. (2006) confirmed the anamorph and teleomorph connection by molecular phylogenetic analyses. Phylogenetically Chaetosphaeria chalaroides was distinct from the type species of Chaetosphaeria, but closely related to members of Paragaeumannomyces (Fernández et al. 2006; Wu and Diao 2022). The type species of Chaetosphaeria s. str. also produces 1-septate ascospore, but has Chloridium anamorph with wet spore mass, and differs from the chalara-like anamorph in Chalarosphaeria. The ascospores of Paragaeumannomyces are filiform or cylindrical and their anamorphs are Obeliospora-like with funnel-shaped collarettes and globose to subglobose conidia, thus they can easily be distinguished from Chalarosphaeria (Réblová et al. 2020; Wu and Diao 2022).
The anamorph of this fungus (Chalara breviclavata) differs from the typical Chalara fungi in producing subcylindrical phialides with strong constriction between venter and collarettes, and its conidia are aseptate, ellipsoidal to shortly clavate, and with rounded apex and truncate base. Holubová-Jechová (1984) found this species commonly occurred on dead wood and bark of various broad-leaved trees.
Phaeodischloridium W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB846973.
= Phaeodischloridium W.P. Wu & YZ. Diao, Fung. Diversity 116: 347, 2022 (nom. inval.)
Etymology: Refers to its similarity to Dischloridium in monophialidic conidiogenous cells with broad opening and cylindrical conidia, but has colored and septated conidia.
Diagnosis: Similar to Dischloridium but differs in producing colored and septate conidia.
Type species: Phaeodischloridium aquaticum (Z.L. Luo, K.D. Hyde & H.Y. Su) W.P. Wu (≡Chaetosphaeria aquatica Z.L. Luo, K.D. Hyde & H.Y. Su).
Colonies effuse, hairy, velvety, dark brown. Stroma absent. Anamorph: Conidiophores solitary, erect, unbranched, straight, or slightly flexuous, septate, smooth or verruculose, dark brown, percurrently regenerating, paler towards the apex, with a swollen base. Conidiogenous cells integrated, terminal, cylindrical, lageniform, collarette inconspicuous. Conidia holoblastic, solitary, accumulating in slimy brown masses at the apices or sometimes sliding down along the sides of conidiogenous cells after secession, cylindrical to cylindric-clavate, obtuse at both ends, medium to dark brown except the pale brown basal cells, 3-septate, apical cell longer than others, thin-walled, smooth. Teleomorph: Unknown.
Ecology/Substrate/Host: Saprobe on dead material of plants.
Geographical distribution: China and Japan.
Description and illustration: Wu and Diao (2022).
Notes: The generic name Phaeodischloridium is invalid due to citation of the invalid basinym (Endophragmia inaequiseptatum Matsush.) for the type species (P. inaequiseptatum) (Wu and Diao 2022). This unique fungus, E. inaequiseptatum was originally described as Endophragmia inaequiseptata (Matsushima 1975; Holotype Matsushima Fungus Collection 2086). Since Endophragmia was not used as the generic name after Hughes’s relocation of its species, and its inclusion in Endophragmiella was not accepted by Hughes (1979), Holubová-Jechová (1987) invalidly transferred it to Dischloridium, which was highly questioned due to its brown and 3-septate conidia. Senwanna (2019) validly published the name Dischloridium inaequiseptatum Matsush. ex P.M. Kirk (effectively published on 28 November 2019). Morphologically it is identical to the recently described Chaetosphaeria aquatica Z.L. Luo, K.D. Hyde & H.Y. Su (published online on 11 November 2019) on conidiophores (181–271 × 5–7 μm), conidiogenous cells and conidia (1–3-septate, 20–24 × 6–8 μm). Phylogenetically this fungus belongs to Chaetosphaeriaceae, but is clearly separated from other known Chaetosphaeria species including the type species (Luo et al. 2019; Wu and Diao 2022). Thus, the new genus Phaeodischloridium W.P. Wu & Y.Z. Diao was created to accommodate this fungus by Wu and Diao (2022). Unfortunately, this new generic name is invalid due to the wrong citation of the invalid basinym (Endophragmia inaequiseptata Matsush.) for the type species of the genus. Both names, Dischloridium inaequiseptatum Matsush. ex P.M. Kirk and Chaetosphaeria aquatica are the valid names for this unique fungus, however Chaetosphaeria aquatica is the earlier name. Here the genus Phaeodischloridium is validly published with the type species P. aquatica (Z.L. Luo, K.D. Hyde & H.Y. Su) W.P. Wu, comb. nov. (basinym Chaetosphaeria aquatica Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 582, 2019).
Phaeodischloridium aquaticum (Z.L. Luo, K.D. Hyde & H.Y. Su) W.P. Wu, comb. nov., MycoBank MB846974.
≡ Chaetosphaeria aquatica Z.L. Luo, K.D. Hyde & H.Y. Su, Fungal Divers. 99: 582, 2019 (published 11 November 2019).
= Dischloridium inaequiseptatum Matsush. ex. Hol.-Jech., Česká Mykol. 41(2): 111, 1987 (nom. inval., Art. 40.1, Shenzhen).
= Dischloridium inaequiseptatum Matsush. ex P.M. Kirk, Index Fungorum 421: 1, 2019 (published 28 November 2019).
= Endophragmia inaequiseptata Matsush., Icon. microfung. Matsush. lect. (Kobe): 69, 1975 (nom. inval., Art. 35.1, Shenzhen).
= Phaeodischloridium inaequiseptatum (Matsush.) W.P. Wu & Y.Z. Diao, Fung. Diversity 116: 347, 2022 (nom. inval., Art. 40.1, Shenzhen).
Typification: China: Yunnan Province, Nujiang River, saprobic on submerged decaying wood, May 2015,
Z.L. Luo, S-752 (MFLU 18–1618, holotype), ex-type living culture MFLUCC 18–1341.
Materials examined: China, Hainan Province, Haikou, on rotten wood of unidentified plant, 20 December 2000, W.P. Wu (Wu5523); Guangdong Province, Guangzhou, South China Botanical Garden, on dead branches of unidentified plant, 9 October 1998, W.P. Wu (Wu2221).
Ecology/Substrate/Host: Saprobe on dead material of plants.
Geographical distribution: China and Japan (Matsushima 1975; Luo et al. 2019).
Description and illustration: Matsushima (1975), Luo et al. (2019), and Wu and Diao (2022).
Lasiosphaeriaceae Nannf., Nova Acta R. Soc. Scient. Upsal., Ser. 4 8(no 2): 50, 1932.
(Sordariales, Sordariomycetidae)
Type genus: Lasiosphaeria Ces. & De Not.
Ecology/Substrate/host: Saprobes on decaying materials.
Geographical distribution: Cosmopolitan.
Description and illustration: Marin-Felix et al. (2020).
Notes: The family was recently emended with a narrower concept by Marin-Felix et al (2020). It is characterized by the production of (mostly) ostiolate ascomata with a tomentose ascomatal wall or bearing septate hairs on or below the neck, and producing one- and two-celled ascospores. The phylogenetic analysis showed that Chalara vaccinii represented a new of this family.
Sordariochalara W.P. Wu & Y.Z. Diao, gen. nov., MycoBank MB845263.
Etymology: Refers to its morphological similarity with Chalara, but phylogenetically belongs to Sordariomycetes.
Type species: Sordariochalara vaccinii (Carris) W.P. Wu & Y.Z. Diao.
Endophytic in leaf and stem. Colonies effuse, olivaceous, velvety. Mycelium composed of hyaline or pigmented hyphae. Anamorph: Stroma absent. Setae absent. Conidiophores cylindrical, septate, medium to dark brown, smooth or verrucose, terminating in a phialide, often with sympodial proliferation. Phialides lageniform, pale brown, smooth, consisting of a venter and collarette, transition from venter to collarette abrupt; venter conical, subcylindrical; collarettes obconical, funnel-shaped. Conidia extruded in long chain, short clavate with apex rounded and base truncate, aseptate, hyaline, smooth-walled. Phragmospores produced holoblastically, arising from aerial and submerged hyphae, cylindrical, apex rounded, septate, medium to dark brown, solitary (Adapted from Carrie 1988). Teleomorph: Unknown.
Ecology/Substrate/Host: Endophytic fungi in leaf and stem.
Geographical distribution: USA.
Notes: The phylogenetic analyses of this study showed that Chalara vaccinii was not a member of Leotiomycetes. Supported by both morphology and very low homology with any other known member of Sordariomycetes, the new genus is created for this fungus. Morphologically it differs from other Chalara s. lat. species by its phialides with very short collarettes and clavate-shaped conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In a megablast search of GenBank nucleotide database, the closest hits (94% identity) using the partial LSU sequence from the ex-type strain were members of Arcoilus, Anopodium, Arnium, Bombardia, Bombardioidea, Naviculispora in Lasiosphaeriaceae (Sordariales). Morphologically none of these genera is similar with Sordariochalara.
Sordariochalara vaccinii (Carris) W.P. Wu & Y.Z. Diao, comb. nov., MycoBank MB845299.
≡ Chalara vaccinii Carris, Mycologia 80(6): 875, 1989 (1988).
Endophytic in leaf and stem. Colonies effuse, olivaceous, velvety. Mycelium composed of hyaline or pigmented hyphae. Anamorph: Conidiophores cylindrical, 2–13-septate, medium to dark brown, smooth or verrucose, terminating in a phialide, 25.5–110 × 3.4–8.5 μm, often with sympodial proliferation. Conidiogenous cells phialidic, lageniform, pale brown, smooth, 14–38 μm long, consisting of a venter and collarette, transition from venter to collarette abrupt; venter conical, subcylindrical, 10–16 × 3.4–5.4 μm; collarettes obconical, funnel-shaped, 3.7–6.7 μm deep, 2–2.7 μm wide at margin. Conidia extruded in long chain, short clavate with apex rounded and base truncate, aseptate, hyaline, smooth-walled, 4–6 × 1–1.3 μm. Phragmospores produced holoblastically, arising from aerial and submerged hyphae, cylindrical, apex rounded, septate, medium to dark brown, solitary, 30–652 × 4.3–8.6 μm (Adapted from Carris 1988). Teleomorph: Unknown.
Ecology/Substrate/Host: Endophyte of Vaccinum macrocarpon.
Geographical distribution: USA (Carris 1988).
Description and illustration: Carris (1988).
Notes: Based on a megablast search of GenBank, the closest hits using the ITS sequence from the ex-type strain were Epicoccum layuense, Podospora and Zopfiella. The above description was based on the original documentation by Carris (1988).
Pyxidiophoraceae G.R.W. Arnold, Z. Pilzk. 37: 191, 1971.
(Pyxidiophorales, Laboulbeniomycetes)
Type genus: Pyxidiophora Bref. & Travel.
Ecology/Substrate/host: Coprophilous or saprophytic on other substrates, rarely mycoparasitic (Marchael 1885; Lundqvist 1980; Haelewaters et al. 2021a, b).
Geographical distribution: Cosmopolitan.
Description and illustration: Arnold (1971) and Haelewaters et al. (2021a, b).
Notes: Gliocephalis, Pyxidiophora and Mycorhynchidium are the three representatives of the Pyxidiophoraceae in Pyxidiophorales, an order related to Laboulbeniales (Lundqvist 1980; Blackwell et al. 1988, 2020; Blackwell & Malloch 1989a, b, 1990; Blackwell 1994; Weir and Beakes 1995; Gams and Arnold 2007). Morphologically fungi in the order Pyxidiophorales produce perithecial ascomata and chalara-like or Thaxteriola anamorphs. The closely phylogenetic relationship of Pyxidiophora and Gliocephalis with Laboulbeniales was recently established by molecular phylogenetic analyses (Jacobs et al. 2005; Tedersool et al. 2017; Haelewaters et al. 2021a, b, 2022).
The member of this family is less than 30 species, mostly saprotrophic, growing on dung or decaying vegetable material, but a few (Gliocephalis hyalina, P. asterophora, P. lundqvistii and P. spinulorostrata) are mycoparasites (Richardson 1972; Barrasa and Moreno 1982; Corlett 1986; Doveri and Coué 2006; Haelewaters et al. 2021a, b). They probably are more common and diverse in nature, but their rapid development in hidden habitats and difficulty in cultivation make them easily overlooked (Blackwell and Malloch 1989a, b; Malloch and Blackwell 1990; Jacobs et al. 2005; Gams and Arnold 2007; Haelewaters et al. 2021a, b). The group is crucial, however, because of its taxonomic position within the Laboulbeniomycetes to provide a morphological link between the thallus-forming Herpomycetales and Laboulbeniales and perithecial ascomycetes (Haelewaters et al. 2021a, b).
A number of different conidial states such as chalara-like or Gabarnaudia-like were described in connection with Pyxidiophora (Spegazzini 1909; Lundqvist 1980; Gams and Arnold 2007). The holoblastic conidia of Pyxidiophora were blunt-ended or bullet-shaped conidia, often in chains produced in phialides on the mycelium. Most were described as chalara-like (e.g., in P. arvernensis, P. asterophora, P. grovei, P. nyctalidis, P. spinuliformis, P. trisporus) or Gabarnaudia-like (e.g., in P. corallisetosa, P. cuniculicola). Pleurocatena produced Gabardnaudia-like anamorph in pure culture (Arnaud 1952, 1953; Arambarri et al. 1981; Gams and Arnold 2007). Other hyphal forms, probably also with phialidic conidia were described, including Gliocephalis hyalina for which no perithecial state was known (Jacobs et al. 2005). In this study, we confirmed that the two hyaline Chalara-like species, Chalara hyalina and Chalara schoenoplecti belonged to Pyxidiophoraceae (Haelewaters et al. 2021a, b), thus, they were excluded from chalara-like fungi of Leotiomycetes and reclassified as two new combinations of Pyxidiophora.
Pyxidiophora Bref. & Tavel., in Brefeld, Unters. Gesammtgeb. Mykol. 10: 189, 1891.
= Acariniola T. Majewski & J. Wiśn., Acta Mycologica, Warszawa 14: 7, 1978.
= Amphoropsis Speg., Anal. Soc. cient. argent. 85(3): 312, 1918.
= Ascolanthanus Cailleux, C. r. hebd. Séanc. Acad. Sci., Paris, sér. III. 265: 1473, 1967.
= Copranophilus Speg., Anal. Mus. nac. B. Aires, Ser. 3, 12: 410, 1909.
= Eleutherosphaera Grove, J. Bot., Lond. 45: 171, 1907 = Endosporella Thaxt., Bot. Gaz. 69: 16, 1920.
= Entomocosma Speg., Anal. Soc. cient. argent. 85(3): 315, 1918.
= Mycorhynchus Sacc. & D. Sacc., Syll. Fung. (Abellini) 18: 418, 1906.
= Myriapodophila Speg., Anal. Soc. cient. argent. 85(3): 313, 1918.
= Rhynchomyces Sacc. & Marchal, in Marchal, Bull. Soc. R. Bot. Belg. 24(1): 60, 1885.
= Rhynchonectria Höhn., Sber. Akad. Wiss. Wien, Math.-naturw. Kl., Abt. 1 111: 1023, 1902.
= Thaxteriola Speg., Anal. Soc. cient. argent. 85(3): 314, 1918.
= Treleasia Speg., Revista Fac. Agron. Univ. nac. La Plata 2(19): 235, 1896.
Coprophilous or saprophytic on other substrates, rarely mycoparasitic. Anamorph: Stroma absent. Setae absent. Conidiophores macronematous, mononematous, arising laterally from aerial hyphae, scattered, solitary or aggregated, hyaline, septate, rarely branched, smooth. Conidiogenous cells integrated, terminal, determinate, phialidic, erect, straight or slightly curved, hyaline to very pale brown, smooth, transition from venter to collarette gradual, venter subcylindrical, collarette cylindrica. Conidia endogenous, catenate, hyaline, cylindrical, smooth-walled, truncate at each end, aseptate, first formed conidia subglobose or ellipsoid. Teleomorph: See Lundqvist (1980).
Type species: Pyxidiophora nyctalidis Bref. & Tavel.
Ecology/substrate/host: Mostly saprotropic, growing on dung or decaying vegetable material, associated with insect, mites and nematode, but a few (P. asterophora, P. lundqvistii, P. spinulorostrata) are mycoparasites (Lundqvist 1980; Haelewaters et al. 2021a, b; Doveri and Coué 2006).
Geographical distribution: Widely distributed (Marchael 1885; Lundqvist 1980; Haelewaters et al. 2021a, b).
Description and illustration: Lundqvist (1980).
Notes: Taxonomy of Pyxidiophora were discussed by Breton and Faurel (1967), Hawksworth and Webster (1977), Lundqvist (1980), Blackwell et al. (1986a, b, 1989) and Haelewaters et al. (2021a, b, 2022). Lundqvist (1980) considered Mycorhynchus, Treleasia, Copranophilus, Ascolanthanus, and Acariniola to be congeneric with Pyxidiophora and recognized about twenty species in the genus. Since then, a number of new species have been added to the genus (Hawksworth and Webster 1977, 1986; Barrasa and Moreno 1982; Blackwell et al. 1986a, b; Kirschner 2003; Haelewaaters et al. 2021b). Pyxidiophora differs from the only other genus Mycorhynchidium (sexually typified genus, without known anamorph) of the family in having long beaked perithecioid rather than cleistothecioid ascomata.
Pyxidiophora was placed under Hypocreales (Breton and Faurel 1967; Rogerson 1970; Müller and von Arx 1973; Hawksworth and Webster 1977; Webster and Hawksworth 1986). Arnold (1971), however, considered the genus distinct from other hypocrealean genera and erected the monotypic family Pyxidiophoraceae. Lundqvist (1980) followed this arrangement and emended the family to include the cleistothecial genus Mycorhynchidium (Malloch and Cain 1971). Lundqvist (1980) was the first to recognize the similarity between ascospores of Pyxidiophora and the thalli of Acariniola spp. and Thaxteriola spp. on mites associated with bark beetles and placed these species in Pyxidiophora. The hypophoretic spores of Pyxidiophora often possess an attachment apparatus (a dark, adhesive, apical or subapical area) which allows them to be carried by the mites linked to these insects (Lundqvist 1980; Blackwell et al. 1988; Blackwell & Malloch 1989a, b; Blackwell 1994; Weir and Beakes 1995; Gams and Arnold 2007; Haelewaters et al 2021a, b, 2022). The closely phylogenetic relationship of Pyxidiophora with Laboulbeniales was recently confirmed by molecular phylogenetic analyses (Blackwell et al. 2015; Tedersool et al. 2017; Haelewaters et al. 2019; Haelewaters et al. 2022).
Pyxidiophora includes about 30 species, mostly saprotrophic, growing on dung or decaying vegetable material, but a few (P. asterophora, P. lundqvistii, P. spinulorostrata) are mycoparasites (Kirschner 2003; Jacobs et al. 2005; Doveri and Coué 2006; Haelewaters et al. 2021a, b; 2022). As discussed by Haelewaters et al. (2021b), the life cycle of most species of Pyxidiophora is complicated, consisting of three different morphs: (1) a dispersal morph (Thaxteriola state) derived from an ascospore that delivers conidia to a fresh substrate, (2) a conidial morph developed on hyphae, and (3) an ascospore-producing perithecial morph to come full circle.
The conidiophores of Chalara s. lat. in Leotiomycetes are usually pigmented, being pale to dark brown, but those of a few species including C. schoenoplecti, C. sibika, C. siamense, C. hyalina and Chalara state of Cryptendoxyla hypophloia were reported to have hyaline to subhyaline conidiophores (Nag Raj and Kendrick 1975; McKenzie et al. 2002). In the phylogenetic tree (Fig. 1), the two species, C. hyalina and C. schoenoplecti, clustered together as a strongly supported clade distinct from other Chalara s. lat. species. The other phylogenetic analyses showed that Chalara hyalina and C. schoenoplecti belonged to Pyxidiophoraceae (Blackwell et al. 2015; Tedersool et al. 2017; Haelewaters et al. 2019, 2021a, b, 2022). The first formed conidia among these species with hyaline conidiophores and conidiogenous cells were described with different morphology from the normal cylindrical conidia, for example, the turbinate in C. sibika, and the subglobose or ellipsoid in C. hyalina, C. schoenoplecti and C. siamense (Subramanian and Sudha 1986; Morgan-Jones et al. 1984; Traue and Arnold 1991; McKenzie et al. 2002).
Pyxidiophora hyalina (Morgan-Jones & Gintis) W.P. Wu, comb. nov., MycoBank MB845300.
≡ Chalara hyalina Morgan-Jones & Gintis, in Morgan-Jones, Gintis & Rodriguez-Kabana, Mycologia 76(2): 211, 1984.
Anamorph: Conidiophores mononematous, semi-macronematous, ascending, arising laterally from aerial hyphae, simple or branches, hyaline. Conidiogenous cells phialidic, integrated, determinate, cylindrical to lageniform, 22–23 × 4–5 μm, apical part 2.5 μm wide. Conidia formed in chain, cylindrical, hyaline, aseptate, truncate at both ends, 10–11 × 3–4 μm. Teleomorph: Unknown.
Ecology/substrate/host: Isolated from cyst of Heteroderae glycines.
Geographical distribution: USA (Morgan-Jones et al. 1984).
Description and illustration: Morgan-Jones et al. (1984).
Notes: Pyxidiophora hyalina was originally described from cyst of nematode. Morphologically it produced hyaline conidiophores and aseptate conidia. In the phylogenetic analyses, the ex-type strain CBS558.92 of Chalara hyalina grouped together with P. schoenoplecti. Morphologically P. hyalina differs from P. schoenoplecti and P. siamense by short conidiophores and longer conidia. It differs from P. arvernensis (conidia 12–15 × 3 µm), P. asterophora (phialide 6–11 µm long; venter 5.5–8 µm wide; cylindrical collarette 2–3.4 µm wide; conidia 6–19 × 3–3.5 µm), P. grovei (conidia similar to P. arvernensis and P. asterophora), P. spinuliformis (conidia 12–15 × 3 µm) and P. trisporus (conidia 11–17 × 4–5.5 µm) by relatively shorter conidia (10–11 × 3–4 μm) (Ludnqvist 1980). Phylogenetically it is also distinct from other species in the genus Pyxidiophora (Haelewaters et al. 2021b).
Pyxidiophora schoenoplecti (M.K.M Wong) W.P. Wu, comb. nov., Fig. 135, MycoBank MB845301.

Pyxidiophora schoenoplecti (Wu7235). a–i conidiophores and phialides. j–q conidia. Scale bar 5 µm
≡ Chalara schoenoplecti M.K.M Wong, in McKenzie, Pinnoi, Wong, Hyde & Jones, Fungal Diversity 11: 131, 2002.
Description on the natural substrate: Colonies effuse, scattered, white, minute. Mycelium partly immersed and partly superficial, composed of hyaline, septate and branched hyphae with smooth and thin wall, 2.5–4 µm wide. Anamorph: Stroma absent. Setae absent. Conidiophores arising from the superficial hyphae, erect, straight or curved, obclavate, subcylindrical, 40–50 × 4–6 μm, 1–2-septate, hyaline to very pale brown, smooth- and thin-walled. Conidiogenous cells integrated, terminal, determinate, phialidic, erect, straight or slightly curved, lageniform, 30–35 µm long, hyaline, smooth, consisting of a venter and a collarette; transition from venter to collarette gradual; venter subcylindrical, 10–15 µm long and 4.5–5.5 (–6) µm wide; collarette cylindrical, 15–19 × 3–3.5 µm. Conidia endogenous, extruded in short or long chains, cylindrical, 7–11 × 2.8–3.2 µm, both ends truncated or flattened, hyaline, aseptate; the first formed conidium turbinate with a truncate base and rounded apex, 6–11 × 2.8–3.2 µm. Teleomorph: Unknown.
Culture characteristics: Colonies effuse, rounded or lobbed, aerial mycelium poorly developed, white, reverse pale soil brown sterile, up to 18 mm on PDA at 25 °C in 4 weeks.
Material examined: China, Yunnan Province, Baoshan, on dead fruit of ?Quercus sp., 19 October 2003, Wenping Wu, Wu7235. Living strain: CGMCC3.23371 (= 47773, from Wu7235).
Ecology/substrate/host: Saprobe on dead leaves of Quercus sp. and Schoenoplectus litoralis.
Geographical distribution: China (McKenzie et al. 2002).
Description and illustration: McKenzie et al. (2002).
Notes: Chalara schoenoplecti was described from Hongkong by McKenzie et al. (2002). It differs from other hyaline chalara-like species, including Chalara. sibika, C. siamense and C. hyalina by a combination of conidiophores, conidiogenous cells and conidia (Nag Raj and Kendrick 1975; McKenzie et al. 2002). It differs from Pyxidiophora arvernensis (conidia 12–15 × 3 µm), P. asterophora (phialide 6–11 µm long; venter 5.5–8 µm wide; cylindrical collarette 2–3.4 µm wide; conidia 6–19 × 3–3.5 µm), P. grovei (conidia similar to P. arvernensis and P. asterophora), P. spinuliformis (conidia 12–15 × 3 µm) and P. trisporus (conidia 11–17 × 4–5.5 µm) by relatively shorter conidia (10–11 × 3–4 μm) (Ludnqvist 1980). Phylogenetically it is also distinct from other species of the genus Pyxidiophora (Haelewaters et al. 2021b).
Based on a megablast search of GenBank nucleotide database, the closest matches to the strain 47773 includes Pyxidiophora microspora (MG438314, 83% identity) and Chalara sp. (JN604461, 92% identity). Based on LSU blast in GenBank, the closest matches to the ex-type strain 47773 include P. siamense (FJ165257, 100%), P. hyalina (MH874039, 95% identity), P. microspora (MG438362, 95% identity), P. arvemensis (MH873278, 91% identity) and Hermatomyces sphaericus (MK348002, 89% identity).
Pyxidiophora siamensis (Pinnoi) W.P. Wu, comb. nov., MycoBank MB845302.
≡ Chalara siamense Pinnoi, in McKenzie, Pinnoi, Wong, Hyde & Jones, Fungal Diversity 11: 131, 2002.
Description on the natural substrate: Anamorph: Conidiophores subcylindrical, obclavate, 41–57 × 5–6 μm hyaline, 1-septate. Conidiogenous cells 35–42 μm long, transition from venter to collarette gradual; venter subcylindrical, 10–20 × 4.56 μm; collarette cylindrical, 20–25 × 2.5–3.5 μm. Conidia endogenous, catenate, hyaline, cylindrical, 7–12 × 2.5–3 μm, rounded at both ends, smooth, aseptate; first formed conidia ellipsoid, 5–6 × 2.5–3 μm, truncate at base, rounded at apex (Adapted from McKenzie et al. 2002). Teleomorph: Unknown.
Ecology/substrate/host: Saprobe on dead leaves of Schoenoplectus litoralis.
Geographical distribution: China (McKenzie et al. 2002).
Description and illustration: McKenzie et al. (2002).
Notes: Chalara siamense was described from Hongkong (McKenzie et al. 2002) and differed from other relevant species by shape and size of conidiogenous cells and conidia.
Sclerococcaceae Réblová, Unter. & W. Gams, Mycol. Progr. 16: 35, 2016.
(Sclerococcales, Sclerococcomycetidiae)
Type genus: Sclerococcum Fr.
Ecology/Substrate/host: Fungicolous, lichenicolous, lignicolous or associated with beetle.
Geographical distribution: Widely distributed.
Description and illustration: Réblová et al. (2016).
Rhopalophora Réblová, Unter. & W. Gams, Mycol. Progr. 16(1): 35, 2016 (2017).
Lignicolous. Anamorph: Conidiophores pale brown, unbranched, macronematous, often reduced to phialides arising directly from undifferentiated hyphae, sometimes regenerating percurrently. Conidiogenous cells phialidic, light brown and paler toward the tip, integrated, subcylindrical, sometimes proliferating sympodially, tapering slightly toward the collarette; collarettes slightly divergent. Conidia hyaline, aseptate, clavate, truncate at the base, arranged in chains or heads. Teleomorph: Unknown.
Type species. Rhopalophora clavispora (W. Gams) Réblová, Unter. & W. Gams.
Ecology/Substrate/host: Saprobes and lignicolous.
Geographical distribution: Widely distributed.
Description and illustration: Réblová et al. (2016).
Rhopalophora hainanensis W.P. Wu & Y.Z. Diao, sp. nov. Figures 136, 137, MycoBank MB841750.

Rhopalophora hainanensis (Wu17643, holotype). a Conidiophores in small groups. b Mycelium and basal part of conidiophores. c–l Conidiophores and conidiogenous cells with collarettes. m–s Conidia. Scale bar: a 10 µm, b–l 5 µm, m–s 2.5 µm

Rhopalophora hainanensis (ex-type strain NN78512 on PDA). a Conidiophores from superficial hyphae. b–i Conidiophores and conidiogenous cells with collarettes. j–t Conidia. Scale bar: a 10 µm, b–t 5 µm
Etymology: refers to the locality, Hainan Province in China, where this fungus was originally discovered.
Typification: China, Hainan Province, Sanya, Yalongwan Park, on decaying seed pod of Bauhinia purpurea, 28 Dec. 2020, Wenping Wu, Holotype Wu17643, ex-type strain CGMCC 3.20807 (= NN78512).
Description on the natural substrate: Colonies effuse, hairy. Anamorph: Conidiophores macronematous, cylindrical, single or 2–5 aggregated, erect, straight or slightly flexuous, 2–5-septate, brown to dark brown, thick- and smooth-walled, with 1–2 percurrent proliferations, terminated with a phialide, 40–52 × 2–2.5 μm. Conidiogenous cells terminal, integrated, lageniform, subcylindrical, pale to medium brown, thin– and smooth-walled, proliferating sympodially with up to 3 phialidic openings, 13–17 × 2–2.5 μm, slightly tapering to 1.0 − 1.5 μm below the collarette; collarettes slightly divergent, slightly darker than other part of the conidiogenous cells, 2–3 μm deep, 1.5 − 2 μm wide; collarettes after sympodial proliferation up to 5 μm deep. Conidia in short chains, fusiform, slightly clavate, aseptate, with an obtuse apex, and a truncate base, guttulate, (3.7)4–5.7 × 1.2–1.5 μm. Teleomorph: Not observed.
Culture characteristics: On PDA, colonies up to 20 mm in 14 days, flat, aerial mycelium sparse, cottony, grey to pale brown, reverse greyish–orange. Conidiophores well developed or reduced to single phialides, directly formed from the superficial hyphae, cylindrical, yellow brown, 0–3 septate, thin– and smooth-walled, 12–40 × 2–3 μm, terminated with a conidiogenous cells. Conidiogenous cells terminal or directly from basal cell in the hyphae, monophialidic, lageniform, subcylindrical, pale brown to yellow brown, 20–25 × 2–2.3 μm; collarette slightly darker than other part of the conidiogenous cells, funnel-shaped, 1.5–2.5 μm wide, 2–3 μm deep. Conidia in short chain or aggregated into dry head, fusiform, ellipsoidal, slightly tapering towards both ends, apex rounded, base truncate, guttulate, 4–5 × 1.2–1.5 μm.
Living strains: 78513, 78524, 78525, 78526 and 78527 (all from the type specimen Wu17643).
Ecology/Substrate/Host: Saprobe on dead material of plant.
Geographical distribution: China.
Notes: Rhopalophora hainanensis differs from the type species of the genus R. clavispora (conidia (4.5–)5–5.5(–6) × 1.5–2 μm on PCA; 4–5 × 1.5–2 μm on MLA) in slightly shorter and narrower conidia. In addition, their ITS sequences have 17 bp differences from each other.
In the phylogenetic analysis, the genus Cylindroconidiis, typified by C. aquaticus, with holoblastic conidiogenous cells and septate and dark brown conidia, clustered together with two species of Rhopalophora. In the original description of this species, the Rhopalophora-like conidiogenous cells in Cylindroconidiis aquaticus were also illustrated from the pure culture on PDA (Yu et al. 2018). In ITS sequences comparison, C. aquaticus has 9 bp differences from R. hainanensis. Based on a megablast search of GenBank nucleotide database, the closest matches to the ex-type strain were members of Rhopalophora, Cylindroconidiis in Sclerococcaceae, including Rhopalophora clavispora (NR152542, 96% identity).
Pseudosclerococcum Olariaga, Teres, J.M. Martín, M. Prieto & Baral, in Olariaga, Teres, Martin, Prieto & Baral, Mycol. Progr. 18(7): 898, 2019.
Type species: Pseudosclerococcum golindoi Olariaga, Teres, J.M. Martín, M. Prieto & Baral
Ecology/Substrate/Host: Saprobe on rotten, decorticated log of Platanus hispanica.
Geographical distribution: Spain (Olariaga et al. 2019).
Description and illustration: Olariaga et al. (2019).
Notes: Pseudosclerococcum with apothecial ascomata is similar to Sclerococcum, but differs in having cylindrical asci embedded in an overall hemiamyloid gelatin with a fissitunicate dehiscence. Unlike Sclerococcum, Pseudosclerococcum golindoi produces a chalara-like asexual morph. This fungus brought into our consideration due to its morphologically similarity with Chalara phaeospora, both fungi produce cylindrical phialides, and aseptate, short-cylindrical or short-clavate, and pale brown conidia with truncated base and rounded apex (Kirk 1985; Olariaga et al. 2019). No living strain or DNA sequence was available for C. phaeospora, its phylogenetic relationship with Pseudosclerococcum and other chalara-like fungi remains to be studied in future.
Pseudosclerococcum golindoi Olariaga, Teres, J.M. Martín, M. Prieto & Baral, in Olariaga, Teres, Martin, Prieto & Baral, Mycol. Progr. 18(7): 898, 2019.
Description in culture: Anamorph: Conidiophores scattered, simple, brown, erect, straight, rarely with a sinuous wall, cylindrical, septate, smooth, sometimes with brown resinous exudates, thin-walled to slightly thick-walled, 59–132 × 3–5.5 μm. Conidiogenous cells cylindrical, brown, smooth- and thin-walled. Conidia in short chains, ellipsoid to shortly clavate, truncate at the basal end with minute inconspicuous marginal frill, non-septate, thin-walled, smooth, pale brown, with a few inconspicuous minute guttulates, 6.5–8(–8.5) × 3–4(–4.5) μm (Adapted from Olariaga et al. 2019). Teleomorph: See Olariaga et al. 2019).
Ecology/Substrate/Host: Saprobe on rotten, decorticated log of Platanus hispanica.
Geographical distribution: Spain (Olariaga et al. 2019).
Description and illustration: Olariaga et al. (2019).
Notes: The anamorph of Pseudosclerococcum golindoi is similar to Chalara phaeospora (phialides 95–125 × 7–10 μm, 14–18 μm wide at the base; conidia (7–)10–14 × (5–)6–8 μm), but differs in narrower phialides and small-sized conidia in the former species (Kirk 1985).
Discussion
Phylogenetic relationship of chalara-like fungi in Leotiomycetes
The chalara-like anamorphs with cylindrical collarettes and deeply seated conidiogenous loci are paraphyletic and phylogenetically connected to several ascomycetous orders such as Chaetosphaeriales, Dothideales, Helotiales, Laboulbeniales, Microascales, Mytilinidiales, Sclerococcales, Sordariales and Trichosphaeriales (Holubová-Jechová 1973, 1993; Matsushima 1975; Nag Raj and Kendrick 1975; Upadhyay 1981; Gams and Philippi 1992; Paulin and Harrington 2000; Seifert et al. 2011; Johnston et al. 2019; Crous et al. 2021). Within Leotiomycetes, the chalara-like fungi have been known in many genera of Pezizellaceae, such as the asexually typified Bloxamia, Chalara s. lat., Chalarodendron, Lareunionomyces, Neochalara, Neolauriomyces, Xenochalara and Zymochalara, and sexually typified Ascoconidium, Calycellina, Calycina, Bioscypha, Bisporella, Crocicreas, Hymenoscyphus, Mollisina, Phaeoscypha, Rodwayella, Sageria and Tapesina. For those known with both asexual and sexual stages, most of these connections had been built through observation of their co-existing on natural substrate, and seldomly confirmed by experiments (Funk 1966a, b, 1975; Nag Raj and Kendrick 1975; Wang and Sutton 1984; Samuels and Rogerson 1990; Gams and Phlippi 1992; Seifert et al. 2011).
Only in last 20 years, more and more connections were confirmed or established by phylogenetic analysis (Paulin and Harrington 2000; McKenzie et al. 2002; Wu 2004; Pratibha et al. 2005; Kowalski 2006; Shabunin 2007; Cai et al. 2009; Koukol 2011; Réblová et al 2011; Silva et al. 2015; Crous et al. 2016a, b, 2018a, b, 2019, 2021, 2022; Guatimosim et al. 2016; Hosoya and Zhao 2016; Suija and Motiejünaité 2016; Ekanayaka et al. 2019; Johnston et al. 2019; Etayo and Silanes 2020; Hosoya 2021; Mitchell et al. 2022). Crous et al. 2016a, b, 2019, 2021). For example, Chalara s. lat. as the largest genus of chalara-like fungi with 146 legitimate names, were showed to be highly diverse in phylogeny and many species were relocated to other genera, such as those associated with Ceratocystidaceae (Microascales) to Thielaviopsis (Blackwell and Gilbertson 1985; Réblová 1999; Paulin and Harrington 2000; Crous et al 2021). While the remnant Chalara species were still phylogenetically heterogeneous in Leotiomycetes, supported by recent molecular phylogenetic studies (Coetsee et al. 2000, Paulin and Harrington 2000; Baral 2002; Paulin et al. 2002; Kowalski 2006; Cai et al. 2009; Réblová et al. 2011; Gross et al. 2015; Baral and Rama 2015; Olariaga et al. 2019; Crous et al. 2021; Karunarathna et al. 2021).
The phylogenetic analyses with inclusion of broad diversity of chalara-like fungi in this study (Figs. 1, 2, 3, 8) clearly demonstrated that the chalara-like fungi in Leotiomycetes were paraphyletic. The 116 analyzed chalara-like fungal species scattered in 20 genera belonging to 5 families such as Arachnopezizaceae, Hamatocanthoscyphaceae, Helotiaceae, Neolauriomycetaceae and Pezizellaceae. The polyphyletic genus Chalara s. lat. was revised with monophyletic generic concepts by redelimitation of Chalara s. str. in a narrow concept, adaption of the emended Calycina to accommodate asexually typified chalara-like fungi, reinstatement of Cylindrocephalum, and introduction of six new genera: Constrictochalara, Leochalara, Minichalara, Nagrajchalara, Parachalara and Stipitochalara. The above results further indicated that the morphological characters used in the classification of anamorphic fungi were often not good predictors of the phylogenetic relationships (Cai et al. 2009). The phenotypes of the chalara-like species in Leotiomycetes had a multiple origin in the evolution. Similar results were also reported from other asexually defined genera such as Codinaea, Dictyochaeta, Dictyosporium, Menisporopsis, Penicillium, Phialocephala, Philalophora and Sporidesmium (Cai et al. 2009; Réblová et al. 2021a, b, c, d; Wu and Diao 2022).
Apart from those studied genera with living strain or DNA sequence available, there are also eight other genera reported with the chalara-like anamorphs, such as Allophylaria, Ascoconidium, Bioscypha, Chalarodendron, Didonia, Phaeoscypha, ?Rodwayella, and Tapesina (Baral 1989, 2002; Höhnel 1919, 1923; Dennis 1949; Martínez-Gil and Baral 2018). However, the phylogenetic relationship of these genera with other chalara-like fungi of Leotiomycetes still remains unknown and to be studied in future, mainly due to lacking living strain and/or DNA sequence for molecular phylogenetic analysis.
Anamorph and teleomorph connection for chalara-like fungi
Although many genera and species of chalara-like fungi were known, only few species were reported with teleomorphs, and number with confirmation by pure culture study or DNA sequencing was even limited (Nag Raj and Kendrick 1975; Seifert 1989; Gams and Philippi 1992; Hosoya and Otani 1997; Paulin and Harrington 2000; Baral 2002; Raitviír 2004; Wang et al. 2007; Friggens et al. 2017). Despite this, the phylogenetic analyses of this study showed closely phylogenetic relationship of some asexually typified genera with the sexually typified genera in Leotiomycetes, such as Bloxamia and Chalara s. str. with Calycina and Calycellina, Chalara-like anamorphs and Hymenoscyphus, Rodwayella, Mollisia, and Tapesina, etc. This result further demonstrated that molecular phylogeny as a powerful tool could be used to build up solid connection of anamorphs and teleomorphs, and explore their evolutional relationship, in particular for those with only anamorph found in nature or pure culture.
Chalara-like fungi with Hymenoscyphus: The chalara-like anamorphs were described for five species of Hymenoscyphus, including H. albidus, H. fraxineus, H. koreanus, H. linearis and H. occultus (Zhang and Zhuang 2004; Kowalski 2006; Zhao et al. 2013; Gross and Han 2015; Gross et al. 2014a, b, 2015). In this study the chalara-like anamorph was described for H. globus and confirmed by the molecular phylogenetic analyses. Morphologically these anamorphs are similar to Chalara s. str. in conidiophores, phialides and conidia, but always have reduced conidiophores consisting of a 1–2-celled basal stalk and a terminal phialide, and hyaline, aseptate, short-cylindrical, almost globose to clavate conidia. Phylogenetically these fungi are distinct from other chalara-like fungi and belong to Helotiaceae.
Bloxamia, Bisporella and Calycina: There were a few reports of the sporodochial Bloxamia anamorphs associated with the genus Bisporella, including the three confirmed connection in Bisporella cyatheicola, B. discedens and B. sulphurina, by pure culture study or molecular phylogeny (Berthet 1964; Carpenter 1975; Nag Raj and Kendrick 1975; Johnston 1988; Paulin and Harrington 2000; Hosoya and Zhao 2016; Guatimosim et al. 2016; Mitchell et al. 2022). Berthet (1964) reported a Bloxamia anamorph in culture from single ascospore isolations of B. sulphurina. Carpenter (1975, 1981) found the Bloxamia-like anamorphs developing on the receptacle of the teleomorph apothecia of B. discedens and B. polygoni, and the links were not proven experimentally. Johnston (1988) reported the Bloxamia anamorph of B. discedens from New Zealand, both from host and culture following single ascospore isolation. Hosoya and Zhao (2016) phylogenetically connected the Bloxamia anamorph with B. discedens by comparing the DNA sequences from both anamorph and teleomorph. Guatimosim et al. (2016) described the new species Bloxamia cyatheicola as pathogen of fern, and both anamorph and teleomorph were found on natural substrate and the connection was proven experimentally by pure culture study and DNA sequence analyses.
The genus Bisporella was recently revised and became to be a later synonym of the monotypic genus Bispora in Helotiaceae. Most of the published species were transferred to Calycina in Pezizellaceae (Mitchell et al. 2022). Our phylogenetic analyses supported that the genus ‘Bisporella’ is polyphyletic, and most species were with Calycina affinity. However, Bloxamia cyatheicola was morphologically similar, but phylogenetically distinct from other genera and belonged to a different genus, thus the new genus Bloxamiella was created. In the ITS tree the three Bloxamia species including the type species B. truncata formed a strongly supported lineage within the Calycina clade. The genus Calycina is a large genus with 235 legitimate names in MycoBank (accessed on July 13th, 2022), and future revision might show the genus is heterogenous. At present no more than 20 species were with the DNA sequences available in GenBank for molecular phylogenetic analysis (GenBank, accessed on July 13th, 2022).
Chalara s. str. and Calycina: Among the existing Chalara species with aseptate conidia and reported teleomorphs, only few species were confirmed by the pure culture study and/or phylogenetic analysis, these were Chalara strobilina with Cyathicula strobilina, Chalara spp. with Calycina parilis and Calycina lactea, etc. (Gams and Philippi 1992; Hosoya and Otani 1997; Paulin and Harrington 2000; Baral 2002; Friggens et al. 2017). Our phylogenetic analyses with different datasets showed that except for two species, all the analyzed species of Chalara s. str. with aseptate conidia clustered together with members of Calycina (including the type species C. herbarum) as a strongly supported group distinct from other genera in Pezizelaceae., a strong indication of closer phylogenetic relationship between Chalara s. str. and Calycina. However, these chalara-like fungi were further separated into at least two groups, one group clustered together with the type species C. herbarium thus congeneric with Calycina; the other group with only chalara-like anamorphs formed a distinct branch in the phylogenetic tree and was kept under Chalara, which needs to be further validated in future study with the type material of Chalara fusidioides, the type species of Chalara. As previously mentioned, the genus Calycina was a large genus with many species known, however less than 20 species were known with living strains and/or DNA sequence data for phylogenetic analysis. Future revision for Calycellina, Calycina and Chalara needs to be made after more species including the type species are included for phylogenetic study.
Nagrajchalara and Cylindrocephalum: These two genera were created or reinstated to accommodate chalara-like fungi which are morphologically similar to Chalara s. str. but with septate conidia and phylogenetically distinct. Although potentially with more than 100 known species of chalara-like fungi with septate conidia, only three species were described with teleomorphs, including Chalara rubi with Tapesina griseovitellina, Nagrajchalara aspera with Phaeoscypha carolinensis, and ‘Chaetochalara’ cladii with Hyaloscypha cladii, and none of these connections was proven experimentally (Nag Raj and Kendrick 1975; Baral 2002). We studied many specimens and isolates of N. aspera (≡ Chaetochalara aspera) from China, and their affinity with Nagrajchalara was confirmed by the phylogenetic analyses (Figs. 2, 3, 8, 15, 40). Unfortunately, no living strain or DNA sequence data was available from other species for phylogenetic study, thus, the phylogenetic relationship of these three fungi with other Leotiomycetes remains to be studied in future.
Phylogenetic significance of morphological characters in delimitation of genera
Presence of sterile or fertile setae among the conidiophores as the diagnostic character for Chaetochalara was not supported in the phylogenetic analyses, thus the genus was disassembled, and the known species were reclassified to Chalara s. str. and Nagrajchalara. Chaetochalara was established to accommodate the fungi morphologically similar to Chalara species but possessing sterile or fertile setae (Sutton and Pirozynski 1965; Nag Raj and Kendrick 1975). Kendrick (1980) was in doubt of the difference between the two closely related genera Chalara and Chaetochalara, differentiated by a single character of absence or presence of sterile setae. Kirk and Spooner (1984) merged the two genera and transferred all Chaetochalara species known by that time into Chalara. Cai et al. (2009) in their phylogenetic analyses further confirmed the congeneric of the two genera, and concluded presence or absence of sterile setae was not a reliable character in delimiting these two genera. However, some other authors did not agree with this merging and continued to use the generic name Chaetochalara as originally defined, even for new species such as Chaetochalara mutabilis and C. proteae (Taylor et al. 2001; Seifert et al. 2011; Silva et al. 2015; Monteiro et al. 2019). In our phylogenetic analyses, seven Chalara species with setae were included and they clustered into three different clades, each represents a different genus, i.e., Chalara africana and C. qinlingensis with aseptate conidia in Chalara s. str., Nagrajchalara aspera, N. jonesii, N. mutabilis and N. septata with septate conidia in Nagrajchalara, and Cylindrocephalum clavatisetosum with clavate-shaped setae and septate conidia in Cylindrocephalum. This firmly demonstrated that members of Chaetochalara were paraphylogenic and the presence of setae was not a reliable character to define the genus (Cai et al. 2009). The type species of Chaetochalara, typified by C. bulbosa with aseptate conidia (Sutton and Pirozynski 1965), was treated as a later synonym of Chalara s. str., and the existing species were reclassified under Chalara s. str. and Nagrajchalara.
Phylogenetic significance of conidial morphologies such as conidial septation and width were further confirmed and used to distinguish different genera in this study. Cai et al. (2009) reported that the conidial width and conidia septation were, to some extent, phylogenetically indicative. Although overall no clearly cut phylogenetic relationships between the species with aseptate and septate conidia, and between species wider and thinner than 2.5 μm, it appeared that species possessing a similar conidial width and conidial septation appeared closely related in the phylogenetic trees. In our phylogenetic analyses with inclusion of broad diversity and different datasets, the species with septate conidia were in most cases well separated from those species with aseptate conidia, and the species with septate conidia were usually with wide conidia. In the phylogenetic tree, the species with septate conidia scattered in 4 distinct clades and each represents a different genus: Neochalara spiraeae in the monotypic genus Neochalara; Cylindrocephalum aureum, C. clavatisetosum, C. huaianensis, C. hughesii and C. kendrickii in the reinstated genus Cylindrocephalum; Chalara eucalypticola in Chalara s. str., and all other species with septate conidia in Nagrajchalara. The species with aseptate conidia were also paraphyletic in Leotiomycetes, and they were recognized in many different genera, distinguished by a combination of morphology and phylogeny: Calycina, Constrictochalara, Hymenoscyphus, Leochalara, Minichalara, Mollisina, Stipitochalara, Xenochalara, and Zymochalara. However, two exceptions were also found for Chalara eucalypticola, N. strobilina and Nagrajchalara xiaohuiae Chalara eucalypticola with septate conidia, was clustered together with all other species of Chalara s. str. producing aseptate conidia; while N. strobilina and N. xiaohuiae with aseptate conidia clustered together in the same clade with all other species of Nagrajchalara with septate conidia. Significance of conidial septation in fungal phylogeny were confirmed in other fungi and used for separation of genera, the recent example is in the revision of Codinaea and Dictyochaeta (Réblová et al. 2021a, b, c, d; Wu and Diao 2022).
With inclusion of more sampled species and diversity, other morphologies such as conidiomata, conidiophores, phialides, conidial length, conidial ornamentation were further examined against the phylogenetic relationships. Aligned with the result from Cai et al. (2009), little phylogenetic significance was found for morphology of phialide, conidial length, conidial ornamentation, as species sharing a similar type of above morphologies scattered in the tree and did not constitute monophyletic clade. However, some of these morphological characters, such as type of conidiomata, branched conidiophores, constriction between venter and collarette, were also phylogenetically indicative for delimitation of relevant genera. For example, three species with significant constriction between collarettes and venters in phialides, Chalara constricta, C. ellisii and an undescribed species, clustered together as a separate genus Constrictochalara; the phylogenetically well-supported genera Lareunionomyces and Neolauriomyces were characterized by apically branched conidiophores; the two sporodochial forming genera, Bloxamia and Bloxamiella, are well-defined monophyletic genera.
Three Chalara species having hyaline conidiophore and conidiogenous cells, i.e., Chalara hyalina, C. schoenoplecti, and Leochalara danxiashanensis, were not congeneric with other chalara-like fungi producing colored conidiophores. In the phylogenetic tree, C. hyalina and C. schoenoplecti clustered together in a strongly supported clade distinct from others. Similar results for two known species were also obtained in previous studies (Paulin and Harrington 2000; Cai et al. 2009). Both species were excluded from Leotiomycetes and reclassified as members of Pyxidiophora in Pyxidiophoraceae (Laboulbeniomycetes) (Haelewaters et al. 2021a, b). Most Chalara species with hyaline conidiophores and conidia were associated with Ceratocystis in Sordariomycetes (Paulin et al. 2000), these two species were the only exceptions with affinity to Pyxidiophoraceae in Laboulbeniomycetes (Haelewaters et al. 2021a, b). Other morphologically similar species wee Chalara heteroderae, C. siamense, C. sibika, Chalara state of Cryptendoxyla hypophloia and Chalara state of Ceratocystis autographa (Nag Raj and Kendrick 1975; Carris & Glawe 1984; Morgan-Jones 1984; Subramanian and Sudha 1986; McKenzie 2002). Among them Chalara state of Cryptendoxyla hypophloia and Chalara state of Ceratocystis autographa were confirmed to belong to Cephalothecaceae and Certosystidaceae respectively (Vu et al. 2019), while no DNA sequence was available for C. siamense and C. sibika. Chalara heterpderae, C. siamense and C. sibika are most likely members of Pyxidiophora, but this needs to be further validated by molecular phylogenetic analyses when the living strain and DNA sequences will be available. The other studied species with hyaline conidiophores and conidiogenous cells formed a completely distinct lineage in Leotiomycetes, and the new genus Leochalara was created.
Chalara phaeospora was described to have cylindrical phialides without clear differentiation of venter and collarette, and aseptate, brown, ellipsoidal conidia (Kirk 1985; Grunig et al. 2009). This was the only species with colored conidia in the genus Chalara s. lat. No living strains was available for molecular phylogenetic analysis, its relationship with other chalara-like fungi remains to be studied in future. Among the numerous anamorphic fungi, only two other fungi, Discomycetoidea aequatorialis Matsush. and Pseudosclerococcum golindoi, were reported to have the chalara-like phialides producing dark brown conidia (Matsushima 1993; Olariaga et al. 2019). Pseudosclerococcum golindoi is the member of Sclerococcales in Eurotiomycetes. No DNA sequence is available for Discomycetoidea aequatoriales.
Unlike other species of Chalara s. lat. with cylindrical collarettes and cylindrical conidia, Chalara antarctica, C. breviclavata and C. caribensis are a small group of species with funnel-shaped collarettes and ellipsoidal to obclavate conidia with rounded apex and truncate base, the typical conidiogenous apparatus and conidia of Phialophora but with the chained conidia. One of them, C. breviclavata was known with Chaetosphaeria teleomorph, thus, was excluded from Leotiomycetes (Réblová 2004).
Future research for chalara-like fungi in Leotiomycetes
With the integrated approach of literature study, morphological observation and phylogenetic analyses, the chalara-like fungi in Leotiomycetes was revised and a framework was provided in generic level for future study. The number of genera was expanded to 26 genera which were asexually or sexually typified. It is expected that the number of species will be further expanded with exploration on diversity from subtropical and tropical areas. For future study of the chalara-like fungi of Leotiomycetes, several potential interesting areas could be explored: (a). It is critical to determine the taxonomic relationship of asexually typified genera (Ascoconidium, Bloxamia, Chalara s. str., Zymochalara) and sexually typified genera such as Bioscypha, Calycina, Calycellina, Didonia, Phaeoscypha, Sageria, Tapesina, etc.; inclusion of the type species for all the genera in future study is necessary; (b). molecular phylogenetic analyses and revision of the polyphyletic genus Bloxamia, Chaetochalara and Chalara s. lat.; (c). biodiversity exploration, especially in subtropical and subtropical areas, could lead to further expansion of these fascinating fungi and also help to obtain more living strains for molecular phylogenetic study.
A list of new names proposed in this study
Bloxamia discedens (P. Karst.) W.P. Wu, comb. nov. (≡ Helotium discedens P. Karst.)
Bloxamia elegans W.P. Wu & Y.Z. Diao, sp. nov.
Bloxamia elongata W.P. Wu & Y.Z. Diao, sp. nov.
Bloxamiella cyatheicola (Guatimosim, R.W. Barreto & Crous) W.P. Wu & Y.Z. Diao, comb. nov.
Calycina affinis (Sacc. & Berl.) W.P. Wu, comb. nov. (≡ Chalara affinis Sacc. & Berl.)
Calycina brevipes (Nag Raj & W.B. Kendr.) W.P. Wu, comb. nov. (≡ Chalara brevipes Nag Raj & W.B. Kendr.)
Calycina brevispora (Nag Raj & W.B. Kendr.) W.P. Wu, comb. nov. (≡ Chalara brevispora Nag Raj & W.B. Kendr.)
Calycina crassipes (Preuss) W.P. Wu, comb. nov. (≡ Cylindrosporium crassipes Preuss)
Calycina dualis (Aramb. & Gamundí) W.P. Wu, comb. nov. (≡ Chalara dualis Aramb. & Gamundí)
Calycina eucalypticola (Crous) W.P. Wu, comb. nov. (≡ Chalara eucalypticola Crous)
Calycina fungorum (Sacc.) W.P. Wu, comb. nov. (≡ Cylindrium fungorum Sacc.)
Calycina oxenbolliae W.P. Wu & Y.Z. Diao, sp. nov.
Calycina parvispora (Nag Raj & S. Hughes) W.P. Wu, comb. nov. (≡ Chalara parvispora Nag Raj & S. Hughes)
Calycina pseudoaffinis (Koukol) W.P. Wu, comb. nov. (≡ Chalara pseudoaffinis Koukol)
Calycina riisgaardii W.P. Wu, sp. nov.
Chalara bacillaris W.P. Wu & Y.Z. Diao, sp. nov.
Chalara bambusicola W.P. Wu & Y.Z. Diao, sp. nov.
Chalara clavatophora W.P. Wu and Y.Z. Diao, sp. nov.
Chalara cylindrophora W.P. Wu & Y.Z. Diao, nomen. nov.
Chalara kirkii W.P. Wu and Y.Z. Diao, sp. nov.
Chalara longiphora W.P. Wu & Y.Z. Diao, sp. nov.
Chalara pengii W.P. Wu & Y.Z. Diao, sp. nov.
Chalara platanicola W.P. Wu & Y.Z. Diao, sp. nov.
Chalara qinlingensis W.P. Wu & Y.Z. Diao, sp. nov.
Chalara sporendocladioides W.P. Wu & Y.Z. Diao, sp. nov.
Chalara versicolor W.P. Wu & Y.Z. Diao, sp. nov.
Chalarosphaeria W.P. Wu & Y.Z. Diao, gen. nov.
Chalarosphaeria breviclavata (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara breviclavata Nag Raj & WB. Kendr.)
Constrictochalara W.P. Wu & Y.Z. Diao, gen. nov.
Constrictochalara clavatospora W.P. Wu & Y.Z. Diao, sp. nov.
Constrictochalara constricta (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara constricta Nag Raj & W.B. Kendr.)
Constrictochalara ellisii (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao (≡ Chalara ellisii Nag Raj & W.B. Kendr.)
Constrictochalara holubovae (Koukol) W.P. Wu & Y.Z. Diao (≡ Chalara holubovae Koukol)
Constrictochalara podocarpi (Crous) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Hamatocanthoscypha podocarpi Crous)
Cylindrocephalum clavatisetosum W.P. Wu & Y.Z. Diao, sp. nov.
Cylindrocephalum hughesii (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara hughesii Nag Raj & W.B. Kendr.)
Cylindrocephalum kendrickii (Nag Raj) W.P. Wu & Y.Z. Diao, comb. nov. (= Chalara kendrickii Nag Raj & W.B. Kendr.)
Cylindrocephalum zhejiangense W.P. Wu & Y.Z. Diao, sp. nov.
Cylindrochalara W.P. Wu & Y.Z. Diao, gen. nov.
Cylindrochalara hyalocuspica (O. Koukol) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara hyalocuspica Koukol)
Lareunionomyces minimus W.P. Wu & Y.Z. Diao, sp. nov.
Leochalara W.P. Wu, gen. nov.
Leochalara danxiashanensis W.P. Wu & Y.Z. Diao, sp. nov.
Minichalara W.P. Wu & Y.Z. Diao, gen. nov.
Minichalara aseptata W.P. Wu & Y.Z. Diao, sp. nov.
Minichalara microspora (Coda.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Cylindrium clandestimum var. microsporum (Corda) Sacc.)
Nagrajchalara W.P. Wu & Y.Z. Diao, gen. nov.
Nagrajchalara acuaria (Cooke & Ellis) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara acuaria Cooke & Ellis)
Nagrajchalara acuariella W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara agathidis (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara agathidis Nag Raj & W.B. Kendr.)
Nagrajchalara angionacea (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara angionacea Nag Raj & W.B. Kendr.)
Nagrajchalara angustata (T. Kowalski & Halmschl.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara angustata T. Kowalski & Halmschl.)
Nagrajchalara aspera (Pirozy. & Hodges) W.P. Wu & Y.Z. Diao, comb. nov. (≡Chaetochalara aspera Pirozy. & Hodges)
Nagrajchalara aunstrupii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara cannonii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara conifericola W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara curviphora W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara ejneri W.P. Wu, sp. nov.
Nagrajchalara ellipsoidea W.P. Wu, sp. nov.
Nagrajchalara guangcaii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara haituoshanensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara hangzhouensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara inflatipes (Pr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Cylindrosporium inflatipes Preuss)
Nagrajchalara insignis (Sacc., Rouss. & Bomm.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Sporoschisma insigne Sacc.)
Nagrajchalara intermedia (W.P. Wu) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara intermedia W.P. Wu)
Nagrajchalara japonica W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara jonesii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara keqinii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara knudsonii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara morganjonesii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara mutabilis (C.R. Silva, S.S. Silva, Gusmão & R.F. Castañeda) W.P. Wu & Y.Z. Diao comb. nov. (≡ Chaetochalara mutabilis C.R. Silva, S.S. Silva, Gusmão & R.F. Castañeda)
Nagrajchalara nawawii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara neonawawii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara novozymia W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara ohmanii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara panamensis (Koukol, T.A. Hofm. & M. Piepenbr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara panamensis Koukol, T.A. Hofm. & M. Piepenbr.)
Nagrajchalara paraunicolor W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara pseudoaurea W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara puerensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara pulchra (Nag Raj & W.B. Kendr.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara pulchra Nag Raj & S. Hughes)
Nagrajchalara qingchengshanensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara selaginellae (M.L. Farr) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara selaginellae M.L. Farr in Farr & Horner)
Nagrajchalara septata W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara septata W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara setosa (Harkn.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara setosa Harkn.)
Nagrajchalara sichuanensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara sivanesanii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara strobilina (Fries) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Peziza strobilina Fr.)
Nagrajchalara tengii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara tropicalis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara truncata W.P. Wu and Y.Z. Diao, sp. nov.
Nagrajchalara tsuensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara tsukairakuensis W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara tubakii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara unicolor (S. Hughes apud Nag Raj & S. Hughes) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara unicolor S. Hughes & Nag Raj)
Nagrajchalara venicola W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara versicolor W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara wenyingiae W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara xiaohuiae W.P. Wu, sp. nov.
Nagrajchalara yinglaniae W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara yongnianii W.P. Wu & Y.Z. Diao, sp. nov.
Nagrajchalara yunnanensis (W.P. Wu) W.P. Wu & Y.Z. Diao, comb. nov. (≡Chalara yunnanensis W.P. Wu)
Neolauriomyces beijingensis W.P. Wu & Y.Z. Diao, sp. nov.
Neolauriomyces crousii W.P. Wu & Y.Z. Diao, sp. nov.
Parachalara W.P. Wu & Y.Z. Diao, gen. nov.
Parachalara olekirkii W.P. Wu & Y.Z. Diao, sp. nov.
Phaeodischloridium W.P. Wu & Y.Z. Diao, gen. nov.
Phaeodischloridium aquaticum (Z.L. Luo, K.D. Hyde & H.Y. Su) W.P. Wu, comb. nov. (≡ Chaetosphaeria aquatica Z.L. Luo, K.D. Hyde & H.Y. Su)
Pyxidiophora hyalina (Morgan-Jones & Gintis) W.P. Wu, comb. nov. (≡ Chalara hyalina Morgan-Jones & Gintis)
Pyxidiophora schoenoplecti (M.K.M Wong) W.P. Wu, comb. nov. (≡Chalara schoenoplecti M.K.M Wong, in McKenzie)
Pyxidiophora siamensis (Pinnoi) W.P. Wu, comb. nov. (≡Chalara siamense Pinnoi)
Rhopalophora hainanensis W.P. Wu & Y.Z. Diao, sp. nov.
Sordariochalara W.P. Wu & Y.Z. Diao, gen. nov.
Sordariochalara vaccinii (Carris) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara vaccinii Carris)
Stipitochalara W.P. Wu & Y.Z. Diao, gen. nov.
Stipitochalara longipes (Preuss) W.P. Wu & Y.Z. Diao comb. nov. (≡ Chalara vaccinii Carris)
Cylindrosporium longipes Preuss)
Stipitochalara piceae-abietis (Hol.-Jech.) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara piceae-abietis Hol.-Jech.)
Stipitochalara recta (O. Koukol) W.P. Wu & Y.Z. Diao, comb. nov. (≡ Chalara recta Koukol)
Data availability
The datasets generated and/or analyzed during the current study are available in the MycoBank repository (included in the manuscript), GenBank (included in Tables 1, 2 and Supplementary Table 1) and TreeBASE (Submission Number: 29633). Supplementary table (STable 1–8) and figures (SFigs. 1–24) were also provided to support the results and discussion. And also, the datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Aime MC, Miller AN, Aoki T et al (2021) How to publish a new fungal species, or name, version 3.0. IMA Fungus 12:11
Aramarri A, Cabello M, Cazau C (1992) A new hyphomycetes from Santiago River. V. Bloxamia cremea. Mycotaxon 63:327–330
Arambarri AM, Gamundí IJ, Bucsinszky AM (1981) Micoflora de la hojarasca de Nothofagus dombeyi. III. Darwiniana 23:327–348
Arambarri AM, Minter TJ, Cabello MN, Minter DW (2007) Spegazzini's drawings of fungi: a digitized library. Treleasia sacchari Speg. http://www.cybertruffle.org.uk/spegazzini/pics/001036aa.jpg. Accessed 18 Aug 2020
Arendholz WR, Sharma R (1984) Observations on some eastern Himalayan Helotiales. Mycotaxon 20:633–680
Arnaud G (1952) Mycologie concrète: genera. Bull Trim Soc Mycol Fr 68:181–223
Arnaud G (1953) Mycologie concrète: genera II. Bull Trim Soc Mycol Fr 69:265–306
Arnold GRW (1971) Über einige neue Taxa und Kombinationen der Sphaeriales. Zeitschrift Für Pilzkunde 37:187–198
Baral HO (1989) Beiträge zur Taxonomie der Discomyceten II. Die Calycellina-Arten mit 4-sporigen Asci. Beitr Kenntn Pilze Mitteleuropas 5:209–236
Baral HO (2002) Tapesina griseovitellina, ein selten berichteter Discomyzet, und seine Nebenfruchtform Chalara rubi. Z Mykol 68:117–133
Baral HO et al (2005) Over 10000 images of fungi and plants (microscopical drawings, water colour plates, photomacro- & micrographs), with material on vital taxonomy and xerotolerance. DVD: In vivo veritas. 3rd edition
Baral HO (2016) Inoperculate discomycetes, pp 157–205. In: Jaklitsch W, Baral HO, Lücking R, Lumbsch HT, Frey W (eds) Syllabus of plant families: A. Engler’s syllabus der Pflanzenfamilien part 1/2. Borntraeger, Stuttgart, p 322
Baral HO, Bemmann M (2014) Hymenoscyphus fraxineus vs. Hymenoscyphus albidus—a comparative light microscopic study on the causal agent of European ash dieback and related foliicolous, stroma-forming species. Mycology 5:228–290
Baral HO, Rämä T (2015) Morphological update on Calycina marina (Pezizellaceae, Helotiales, Leotiomycetes), a new combination for Laetinaevia marina. Bot Mar 58:523–534
Baral HO, Queloz V, Hosoya T (2014) Hymenoscyphus fraxineus, the correct scientific name for the fungus causing ash dieback in Europe. IMA Fungus 5:79–80
Barrasa JM, Moreno G (1982) Pyxidiophora badiorostris Lundq. y Pyxidiophora fimbriata sp. nov. en España (Pyrenomycetes). Cryptog Mycol 3:41–49
Baschien C, Manz W, Neu TR, Marvanová L, Szewzyk U (2008) In situ detection of freshwater fungi in an alpine stream by new taxon-specific fluorescence in situ hybridization probes. Appl Environ Microbiol 74:6427–6436
Bates ST, Miller AN, Collections TM, Consortia MC (2018) The protochecklist of North American nonlichenized Fungi. Mycologia 110:1222–1348
Batista AC, Bezerra JL, Peres GEP (1960) Singera n. gen. e outros fungos Moniliales. Publ Inst Micol Univ Recife 298:1–33
Berkeley MJ, Broome CE (1854) Notices of British fungi (730–784). Ann Mag Nat Hist 13:458–469
Berthet P (1964) Formes conidiennes de divers discomycetes. Bull Trimest Soc Mycol Fr 80:125–149
Blackwell M (1994) Minute mycological mysteries: the influence of arthropods on the lives of fungi. Mycologia 86:1–17
Blackwell M, Gilbertson RL (1985) Quesiconcha reticulata and its anamorph from conifer roots. Mycologia 77:50–54
Blackwell M, Malloch D (1989a) Pyxidiophora: a link between the Laboulbeniales and hyphal ascomycetes. Mem N Y Bot Gard 49:23–32
Blackwell M, Malloch D (1989b) Pyxidiophora: life histories and arthropod associations of two species. Can J Bot 67:2552–2562
Blackwell M, Malloch D (1990) Discovery of a Pyxidiophora with Acariniola-type ascospores. Mycol Res 94:415–417
Blackwell M, Bridges JR, Moser JC, Perry TJ (1986a) Hyperphoretic dispersal of a Pyxidiophora anamorph. Science 232:993–995
Blackwell M, Perry TJ, Bridges JR, Moser JC (1986b) A new species of Pyxidiophora and its Thaxteriola anamorph. Mycologia 78:605–612
Blackwell M, Moser JC, Wiśniewski J (1989) Ascospores of Pyxidiophora on mites associated with beetles in trees and wood. Mycol Res 92:397–403
Blackwell M, Haelewaters D, Pfister DH (2020) Laboulbeniomycetes: evolution, natural history, and Thaxter’s final word. Mycologia 112:1048–1059
Breton A, Faurel L (1967) Etude des affinites du genre Mycorhyncus Sacc. et description de plusieurs especes nouvelles. Rev Mycol 32:229–258
Cabello MN (1989) Deuteromycotina from Antarctica. New species of hyphomycetes from Danco Coast, Antarctic Peninsula. Mycotaxon 36:91–94
Cai L, Wu WP, Hyde KD (2009) Phylogenetic relationships of Chalara and allied species inferred from ribosomal DNA sequences. Mycol Prog 8:133–143
Carpenter SE (1975) Bisporella discedens and its Cystodendron state. Mycotaxon 2:123–126
Carpenter SE (1981) Monograph of Crocicreas (Ascomycetes, Helotiales, Leotiaceae). Mem N Y Bot Gardens 33:1–290
Carris LM (1988) Chalara vaccinii sp. nov., a Vaccinium endophyte. Mycologia 80:875–879
Carris LM, Glawe DA (1984) Chalara heteroderae, a new species isolated from cysts of Heterodera glycines in Illinois. Mycotaxon 21:441–448
Catania M, Romero A (2009) Micromicetes asociados con corteza y/o madera de Podocarpus parlatorei Pilg. en la Argentina. V. Ascomycota anamórficos. Lilloa 46 43–51
Centro de Estudios Micologicos Asturianos (CEMAS) Inicio. Asociación “Centro de Estudios Micológicos Asturianos” (centrodeestudiosmicologicosasturianos.org)
Christiansen MS (1993) Chalara lichenicola n. sp. (Deuteromycotina), a lichenicolous hyphomyete from Svalbard. Nord J Bot 13:309–312
Coetsee C, Wingfield MJ, Crous PW, Wingfield BD (2000) Xenochalara, a new genus of dematiaceous hyphomycetes for Chalara-like fungi with apical wall building conidial development. S Afr J Bot 66:99–103
Cole GT, Kendrick B (1973) Taxonomic studies of Phialophora. Mycologia 65:661–688
Coppins BJ, Minter DW (1981) A new hyphomycete from Northhumberland. Notes RBG Edinb 38:363–365
Corlett M (1986) Pyxidiophora lundqvistii n. sp. (Hypomycetales, Ascomycetes). Can J Bot 64:805–807
Crous PW, Wingfield MJ, Burgess TI, Hardy GJ, Crane C, Barrett S et al (2016a) Fungal planet description sheets: 469–557. Persoonia 37:218–403
Crous PW, Wingfield MJ, Richardson DM, Le Roux JJ, Strasberg D et al (2016b) Fungal planet description sheets: 400–468. Persoonia 36:316–458
Crous PW, Luangsaard JJ, Wingfield MJ, Carnegie AJ, Hernández-Restrepo M et al (2018a) Fungal planet description sheets: 785–867. Persoonia 41:238–417
Crous PW, Wingfield MJ, Burgess TI, Hardy GESJ, Gene J et al (2018b) Fungal planet description sheets: 716–784. Persoonia 40:240–393
Crous PW, Wingfield MJ, Lombard L, Roets F, Swart WJ et al (2019) Fungal planet description sheets: 951–1041. Persoonia 43:223–425
Crous PW, Osieck ER, Jurjevic Z, Boers J, van Iperen AL, Starink-Willemse M et al (2021) Fungal planet description sheets: 1284–1382. Persoonia 47:178–374
Crous PW, Boers J, Holdom D, Osieck ER, Steinrucken TV, Tan YP et al (2022) Fungal planet description sheets: 1383–1435. Persoonia 48:261–371
Cyberliber, an Electronic Library for Mycology www.cybertruffle.org.uk/cyberliber. Accessed 25 July 2022
Dennis RWG (1949) A revision of the British Hyaloscyphaceae. Mycol Pap 32:1–97
Dennis RWG (1956) A revision of the British Helotiaceae in the Herbarium of the Royal Botanic Garden, Kew with notes on related European species. Mycol Pap 62:1–216
Dennis RWG (1962) A reassessment of Belonidium Mont. & Dur. Persoonia 2:171–191
Dennis RWG (1968) British Ascomycetes. 3310 Lehre Verlag von J. Cramer
Dennis RWG (1986) Fungi of the Hebrides 1–383
Dixon JR (1974) Chlorosplenium and its segregates. I. Introduction and the genus Chlorosplenium. Mycotaxon 1:65–104
Doveri F, Coue B (2006) First recrd of Pyxidiophora badiorostris from France. Documents Mycologigues XXXIV 133–134:33–41
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull 19:11–15
Ekanayaka AH, Hyde KD, Gentekaki E, McKenzie EHC, Zhao Q, Bulgakov TS, Camporesi L (2019) Preliminary classification of Leotiomycetes. Mycosphere 10:310–489
Ellis MB (1961) Dematiaceous hyphomycetes. II. Mycol Pap 79:1–23
Ellis MB (1971) Dematiaceous hyphomycetes. Commonwealth Mycological Institute, Kew
Ellis MB and Ellis JP (1987) Microfungi on land plants an identification handbook. Croom Helm
Etayo J, Direderich P (1996) Lichenicolous fungi from the Western Pyrenes, France and Spain. III. Species on Lobaria pulmonaria. Bull Soc Nat Luxemb 97:93–118
Etayo J, Sancho LG (2008) Hongos liquenícolas del sur de Sudamérica, especialmente de Isla Navarino (Chile). Bibl Lichenol 98:1–302
Etayo J, López de Silanes ME (2020) Hongos liquenícolas del norte de Portugal, especialmente del Parque Natural Montesinho Lichenicolous fungi from Northern Portugal, specially from Montesinho Natural Park Lichenicolous fungi from Northern Portugal. Nova Acta Ci Compostelana 27:35–50
Finnish Biodiversity Information Facility (laji.fi). Accessed 10 July 2022
Friggens NL, Taylor JE, Koukol O (2017) Diversity and community composition of aquatic ascomycetes varies between freshwater, estuarine and marine habitats in western Scotland. Mycosphere 8:1267–1287
Funk A (1966a) The type species of Ascoconidium. Can J Bot 44:39–41
Funk A (1966b) Ascoconidium tsugae n. sp. associated with bark diseases of Western Hemlock in British Columbia. Can J Bot 44:219–222
Funk A (1975) Sageria, a new genus of Helotiales. Can J Bot 53:1196–1199
Gadgil PD, Dick M (1999) Fungi silvicolae Novazelandiae: 2. N Z J for Sci 29:440–458
Galán R (1993) Helotiella maireana Rehm, a forgotten cupuliferous species. Rivista Di Micologia 36:149–154
Gams W (2000) Phialophora and some morphologically little differentiated anamorphs of divergent ascomycetes. Stud Mycol 45:187–199
Gams W, Arnold GRW (2007) The hyphomycete genus Pleurocatena anamorphs of Pyxidiophora. Nova Hedwigia 84:381–393
Gams W, Holubová-Jechová V (1976) Chloridium and some other dematiaceous hyphomycetes growing on decaying wood. Stud Mycol 13:1–99
Gams W, Philippi S (1992) A study of Cyathicula strobilina and its Chalara anamorph in vitro. Persoonia 14:547–552
Gamundí IJ, Giaiotti A (1998) Nota sorbe discomycetes Andino-Patagónicos II. Novedades taxonómicas. Darwiniana 35:49–160
Gamundí IJ, Arambarri AM, Giaiotti A (1977) Micoflora de la Hojarasca de Nothofagus dombeyi. Darwiniana 21:81–114
Gernandt DS, Platt JL, Stone JK, Spatafora JW, Holst-Jensen A, Hamelin RC, Kohn LM (2001) Phylogenetics of Helotiales and Rhytismatales based on partial small subunit nuclear ribosomal DNA sequences. Mycologia 93:915–933
Glawe DA (1984) Bloxamia truncata in artificial culture. Mycologia 76:741–745
Graddon WD (1980) Some new Discomycete species: 5. Trans Br Mycol Soc 74:265–269
Gray SF (1821) A natural arrangement of British plants. 1:1–824
Gross A, Han JG (2015) Hymenoscyphus fraxineus and two new Hymenoscyphus species identified in Korea. Mycol Prog 14:1–13
Gross A, Holderueder O, Pautasso M, Queloz V, Sieber TN (2014) Hymenoscyphus pseudoalbidus, the causal agent of European ash dieback. Mol Plant Pathol 15:5–21
Gross A, Hosoya T, Zhao Y, Baral HO (2015) Hymenoscyphus linearis sp. nov.—another close relative of the ash dieback pathogen H. fraxineus. Mycol Prog 14:1–15
Grunig C, Queloz V, Duo A, Sieber TN (2009) Phylogeny of Phaeomollisia piceae gen. sp. nov.: a dark, septate, confer–needle endophyte and its relationships to Phialocephala and Acephala. Mycol Res 113:207–221
Guatimosim E, Schwartsburd PB, Crous PW, Barreto RW (2016) Novel fungi from an ancient niche: lachnoid and chalara-like fungi on ferns. Mycol Progress 15:1239–1267
Guindon S, Dufayard JF, Lefort V, Anisimova M (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 59:307–321
Guo YL (1989) Folicolous hyphomycetes from Shengnongjia. In Anon.: Fungi and Lichens of Shengnongjia, pp 341–385 (In Chinese)
Guo YL (1997) Imperfect fungi of the Qinling Mountains. In Mao XL, Zhuang JY (eds) Fungi of the Qinling Mountain, pp 150–181
Haelewaters D, Pfliegler WP, Gorczak M, Pfister DH (2019) Birth of an order: comprehensive molecular phylogenetic study excludes Herpomyces (Fungi, Laboulbeniomycetes) from Laboulbeniales. Mol Phylogenet Evol 133:286–301
Haelewaters D, Blackwell M, Pfister DH (2021a) Laboulbeniomycetes: intimate fungal associates of arthropods. Ann Rev Entomol 66:257–276
Haelewaters D, Gorczak M, Kaishian P, De Kesel A, Blackwell M (2021b) Laboulbeniomycetes, enigmatic fungi with a turbulent taxonomic history. In: Encyclopedia of mycology, vol 1, pp 263–283
Haelewaters D, Lubbers M, Kesel AD (2022) The haustorium as a driving force for speciation in thallus-forming Laboulbeniomycetes. IMA Fungus 13:1. https://doi.org/10.1186/s43008-021-00087-7
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Han JG, Hosoya T, Sung GH, Shin HD (2014) Phylogenetic reassessment of Hyaloscyphaceae sensu lato (Helotiales, Leotiomycetes) based on multigene analyses. Fungal Biol 118:150–167
Hawksworth DL, Webster J (1977) Studies on mycorhynchus in Britain. Trans Br Mycol Soc 68:329–340
Hennebert GL, Bellemere A (1979) Les formes conidiennes des discomycètes: essai taxonomique. Rev Mycol 43:259–315
Hernández-Restrepo M, Gené J, Castañeda-Ruiz RF, Mena-Portales J, Crous PW, Guarro J (2017) Phylogeny of saprobic microfungi from Southern Europe. Stud Mycol 86:53–97
Holubová-Jechová V (1973) Lignicolous hyphomycetes from Czechoslovakia. 3. Sporoschisma, Sporoschismopsis and Cutenularia. Folia Geobot Phytotaxon 19:387–438
Holubová-Jechová V (1982) New or interesting phialidic hyphomycetes from Cuba. Mycotaxon 15:277–292
Holubová-Jechová V (1984) Lignicolous hyphomycetes from Czechoslovakia. 7. Chalara, Exochalara Fusichalara and Dictyochaeta. Folia Geobot Phytotaxon 8:209–218
Holubová-Jechová V (1987) Studies on hyphomycetes from Cuba V Six new species of dematiaceous hyphomycetes from Havana Province. Ceská Mykol 41:29–36
Hosoya T (2021) Systematics, ecology, and application of Helotiales: recent progress and future perspectives for research with special emphasis on activities within Japan. Mycoscience 62:1–9
Hosoya T, Otanl Y (1997) Hyaloscyphaceae in Japan (2): glassy-haired members of the tribe Hyaloscypheae. Mycoscience 38:187–205
Hosoya T, Zhao YJ (2016) Enumeration of remarkable Japanese Discomycetes (10): two Helotiales and a Helotialean anamorph new to Japan. Bull Nat Mus Nat Sci Ser B 4:113–119
Hosoya T, Han JG, Saito Y, Zhao YJ, Kakishima M, Shin HD (2011) Enumeration of remarkable Japanese discomycetes (5): first record of one operculate and two inoperculate discomycetes in Japan. Bull Nat Mus Nat Sci Ser B 37:147–154
Huelsenbeck JP, Ronquist F (2001) MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755
Hughes SJ (1958) Revisiones Hyphomycetum aliquot cum appendice de nominibus rejiciendis. Can J Bot 36:727–836
Hughes SJ (1979) Relocation of species of Endophragmia auct. with notes on relevant generic names. N Z J Bot 17:139–188
Husson C, Scala B, Caël O, Frey P, Feau N, Ioos R, Marçais B (2011) Chalara fraxinea is an invasive pathogen in France. Eur J Plant Pathol 130:311–324
Jacobs A, Coetzee MPA, Wingfield BD, Jacobs K, Wingfield MJ (2003) Phylogenetic relationships among Phialocephala species and other ascomycetes. Mycologia 95:637–645
Jacobs K, Holtzman K, Seifert KA (2005) Morphology, phylogeny and biology of Gliocephalis hyalina, a biotrophic contact mycoparasite of Fusarium species. Mycologia 97:111–120
Johnston P (1988) The Bloxamia anamorph of Bisporella discedens. Mycotaxon 31:345–350
Johnston PR, Park D, Baral HO, Galán R, Platas G, Tena R (2014a) The phylogenetic relationships of Torrendiella and Hymenotorrendiella gen. nov. within the Leotiomycetes. Phytotaxa 177:1–25
Johnston P, Seifert KA, Stone JK, Rossman AY, Marvanová L (2014b) Recommendations on generic names competing for use in Leotiomycetes (Ascomycota). IMA Fungus 5:91–120
Johnston PR, Quijada L, Smith CA, Baral HO, Hosoya T, Baschien C, Pärtel K, Zhuang WY, Haelewaters D, Park D, Carls S, López-Giráldez F, Wang Z, Townsend JP (2019) A multigene phylogeny toward a new phylogenetic classification of Leotiomycetes. IMA Fungus 10:1–22
Karunarathna A, Działak P, Jayawardena RS, Karunarathna SC, Kuo CH, Suwannarach N, Tibpromma S, Lumyong S (2021) A novel addition to the pezizellaceae (rhytismatales, ascomycota). Phytotaxa 480:251–261
Katoh K, Standley DM (2016) A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016:1–10
Kendrick WB (1961) The Leptographium complex. Phialocephala gen. nov. Can J Bot 39:1079–1085
Kendrick WB (1980) The generic concept in Hyphomycetes—a reappraisal. Mycotaxon 11:339–364
Kiffer E, Delon R (1983) Chalara elegans (Thielaviopsis basicola) and allied species II. Validation of two taxa. Mycotaxon 18:165–174
Kile GA, Walker J (1987) Chalara australis sp. nov. (Hyphomycetes), a vascular pathogen of Nothofagus cunninghamii (Fagaceae) in Australia and its relationship to other Chalara species. Aust J Bot 35:1–32
Kile GA, Harrington TC, Yuan ZQ, Dudzinski MJ, Old KM (1996) Ceratocystis eucalypti sp. nov., a vascular stain fungus from eucalypts in Australia. Mycol Res 100:571–579
Kirisits L, Dämpfle D, Kräutler K (2013) Hymenoscyphus albidus is not associated with an anamorphic stage and displays slower growth than Hymenoscyphus pseudoalbidus on agar media. Forest Pathol 43(5):386–389
Kirk PM (1981) New or interesting microfungi: III. A preliminary account of microfungi colonizing Laurus nobilis leaf litter. Trans Br Mycol Soc 77:457–473
Kirk PM (1984) New or interesting microfungi XIII. Ascomycetes on Laurus nobilis leaf litter. Mycotaxon 19:307–322
Kirk PM (1985) New or interesting microfungi. XIV Dematiaceous hyphomycetes from Mt Kenya. Mycotaxon 23:305–352
Kirk PM (1986) New or interesting microfungi. XV. Miscellaneous hyphomycetes from the British Isles. Trans Br Mycol Soc 86:409–428
Kirk PM, Ansell AE (1992) Authors of fungal names: a list of authors of scientific names of fungi. IMI, Kew
Kirk PM, Spooner BM (1984) An account of the fungi of Arran, Gigha and Kintyre. Kew Bull 38:503–597
Kirk PM, Cannon PF, David JC, Stalpers JA (2001) Dictionary of the fungi, 9th edn. CAB International, Wallingford
Kirschner R (2003) Two new species of Pyxidiophora associated with bark beetles in Europe. Mycol Progr 2:209–218
Korf RP (1952) A monograph of the Arachnopezizeae. Lloydia 14:129–186
Kosonen T, Huhtinen S, Hansen K (2021) Taxonomy and systematics of Hyaloscyphaceae and Arachnopezizaceae. Personnia 46:26–62
Koukol O (2011) New species of Chalara occupying coniferous needles. Fungal Divers 49:75–91
Koukol O (2012) A new species of Infundichalara from pine litter. Mycotaxon 120:343–352
Koukol O, Kolařik M, Kolářová Z (2012) Diversity of foliar endophytes in wind-fallen Picea abies trees. Fungal Divers 54:69–77
Koukol O, Halasů V, Janošík L, Mlčoch P, Polhorský A, Šandová M, Zíbarová L (2020) Citizen science facilitates phylogenetic placement of old species of non-lichenised Pezizomycotina based on newly collected material. Czech Mycol 72:263–280
Kowalski T (2006) Chalara fraxinea sp. nov. associated with dieback of ash (Fraxinus excelsior) in Poland. For Path 36:264–270
Kowalski T, Bilański P (2018) Hymenoscyphus pusillus, a new species on leaves of Fraxinus pennsylvanica in Poland. Forest Pathol 49:e12482
Kowalski T, Halmschlager E (1996) Chalara angustata sp. nov. from roots of Quercus petraea and Quercus vobur. Mycol Res 100:1112–1116
Kowalski T, Holdenrieder O (2009) The teleomorph of Chalara fraxinea, the causal agent of ash dieback. For Path 39:304–308
Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular evolutionary genetics analysis Version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874
Lambotte (1887). La Flore Mycologique de la Belgique. Premier Supplement comprenant les Hyménomycètes, Pyrénomycètes, Discomycètes. Addition de 1070 especes a la flore de 1880. Mém Soc Roy Sci. Liège. ser. 2, 14: 1–350
Li DW, Chen JY, Wang YX (2013a) Hyphomycetes: Linodochium sinense sp. nov. and new records from fallen Sycopsis sinensis leaves in China. Mycotaxon 126:15–22
Li XX, Ma LG, Xia JW, Zhang XG (2013b) Three newly recorded species of Parasympodiella and Chalara from China. Mycotaxon 126:121–126
Liu YL, Zhang ZY (1998) A new species of the genus Bloxamia. Mycosystema 17:7–10
Lizon P, Korf RP (1995) Taxonomy and nomenclature of Bisporella claroflava (Leotiaceae). Mycotaxon 64:471–478
Locquin, M. 1984. Mycologie générale et structurale. 1–551
Lowen R, Dumont KP (1984) Taxonomy and nomenclature in the genus Calycellina (Hyaloscyphaceae). Mycologia 76:1003–1023
Lu BS, Hyde KD, Ho WH, Tsui KM, Taylor JE, Wong KM, Yanna, Zhou DQ (2000) Checklist of Hong Kong Fungi. Fungal Divers Press, 1–207
Lundqvist N (1980) On the genus Pyxidiophora sensu lato (Pyrenomycetes). Bot Not 133:121–144
Luo ZL, Hyde KH, Liu JK, Maharachchikumbrua SSN, Jeewon R, Bao DF, Bhat DJ, Lin CG, Li WL, Yang J, Liu NG, Jayawardena RS, Li JF, Su HY (2019) Freshwater sordariomycetes. Fungal Divers 99:451–660
Malloch D, Blackwell M (1990) Kathistes, a new genus of pleomorphic ascomycetes. Can J Bot 68:1712–1721
Malloch D, Cain RF (1971) Four new genera of cleistothecial ascomycetes with hyaline ascospores. Can J Bot 49:847–854
Marchal E (1885) Champignons coprophiles de Belgique IV. Bull Soc Roy Bot Belg 24:57–77
Marin-Felix Y, Miller AN, Cano-Lira GJ, Garcia D, Stadler M, Huhndorf SM, Stchigel AM (2020) Re-evaluation of the order sordariales: delimitation of Lasiosphaeriaceae s. str., and Introduction of the New Families Diplogelasinosporaceae, Naviculisporaceae, and Schizotheciaceae. Microorganisms 8(1430):1–41
Martínez-Gil R, Baral H (2018) Allophylaria subliciformis (Pezizellaceae, Helotiales), a rare species rediscovered in Europe and North America. Ascomycetes.org 10(2):67–75
Matsushima T (1971) Microfungi of the Solomon Islands and Papua-New Guinea. Published by author, Kobe, Japan
Matsushima T (1975) Icones Microfungorum a Matsushima Lectorum. Published by author, Kobe
Matsushima T (1980) Matsushima Mycological Memoirs. No.1, Published by the Author, Kobe
Matsushima T (1993) Matsushima Mycological Memoirs 7:1–141. Published by the Author, Kobe
McKenzie EHC, Pinnoi A, Wong MKM, Hyde KD, Jones EBG (2002) Two new hyaline Chalara species and a key to species described since 1975. Fungal Divers 11:129–139
McKenzie EHC (1982) New hyphomycetes on monocotyledons. NZ J Bot 20:245–252
McKenzie EHC (1993) New hyphomycete species from litter in the Chatham Islands, New Zealand. Mycotaxon 46:291–297
McKenzie EHC (1997) Bahusutrabeeja bunyensis sp. nov. (Hyphomycetes) from Queensland, Australia, and a new name for Chalara australis McKenzie. Mycotaxon 61:303–306
Minter DW, Holubová-Jechová V (1981) New or interesting Hyphomycetes on decaying pine litter from Czechoslovakia. Folia Geobot Phytotaxon 16:195–217
Minter DW, Kirk PM, Sutton BC (1982) Holoblastic phialides. Trans Br Mycol Soc 79:75–93
Minter DW, Kirk PM, Sutton BC (1983) Thallic phialides. Trans Br Mycol Soc 80:39–66
Mitchell JK, Quijada L, Johnston PR, Pfister DH (2022) Species of the common discomycete genus Bisporella reassigned to at least four genera. Mycologia. https://doi.org/10.1080/00275514.2022.2058853
Monteiro JS, Samento PSM, Satao HMP (2019) Saprobic conidial fungi associated with palm leaf litter in eastern Amazon, Brazil. Acad Bras Cienc 91(3):e20180545
Morgan-Jones G, Ingram EG (1976) Notes on Hyphomycetes. XII. A new species of Chalara. Mycotaxon 4:489–492
Morgan-Jones G, Gintis BO, Rodriguez-Kabana R (1984) New species of Chalara and Scytalidium isolated from cysts of Heterodera glycines. Mycologia 76:211–217
Morgan-Jones G, Sinclair RC, Eicker A (1992) Dematiaceous hyphomycetes from South Africa: II. Some phialidic species, including Chalara transkeiensis, new species, and Zanclospora brevispora var. transvaalensis, new variety, two new taxa. S Afr J Bot 58:147–154
Morozova II (2014) New records of discomycetous fungi from Ukraine. Turk J Bot 38:398–405
Mouton M, Wingfield MJ (1993) Comparison between conidial development in Sporendocladia bactrospora and Phialocephala virens. Can J Bot 71:985–991
Nag Raj TR (1993) Coelomycetous anamorphs with appendage-bearing conidia. Mycologue Publications, Waterloo
Nag Raj TR, Hughes SJ (1974) New Zealand Fungi. 21. Chalara (Corda) Rabenhorst. N Z J Bot 12:115–129
Nag Raj TR, Kendrick WB (1971) On the identity of three species of Cylindrosporium described by Preuss. Can J Bot 49:2119–2122
Nag Raj TR, Kendrick B (1975) A monograph of Chalara and allied genera. Wilfrid Laurier University Press, Waterloo
Nag Raj TR, Kendrick B (1993) The anamorph as generic determinant in the holomorph: the Chalara connection in the ascomycetes, with special reference to the ophiostomatoid fungi. In: Wingfield MJ, Seifert KA, Webber JF (eds) Ceratocystis and ophiostoma. American Phytopathological Society, St Paul, pp 61–70
Newsham KK, Cox F, Sands CJ, Garnett MH, Magan N, Horrocks CA, Dungait JAJ, Robinson CH (2020) A previously undescribed helotialean fungus that is superabundant in soil under maritime antarctic higher plants. Front Microbiol 11:615608. https://doi.org/10.3389/fmicb.2020.615608
Nylander JAA (2004) MrModeltest Version 2. Program Distributed by the Author Evolutionary Biology Centre, Uppsala University, Uppsala
Olariaga I, Teres J, Martin J, Prieto M, Baral HO (2019) Pseudosclerococcum golindoi gen. et sp. nov., a new taxon with apothecial ascomata and a Chalara-like anamorph within the Sclerococcales (Eurotiomycetes). Mycol Prog 18:895–905
Onofri S, Zucconi L (1984) Two new species of the genus Phialocephala. Mycotaxon 20:185–195
Paulin AE, Harrington TC (2000) Phylogenetic placement of anamorphic species of Chalara among Ceratocystis species and other ascomycetes. Stud Mycol 45:209–222
Paulin AE, Harrington TC, McNew DL (2002) Phylogenetic and taxonomic evaluation of Chalara, Chalaropsis, and Thielaviopsis anamorphs associated with Ceratocystis. Mycologia 94:62–72
Pirozynski KA, Hodges CS (1973) New hyphomycetes from South Carolina. Can J Bot 51:157–173
Pirozynski KA, Morgan-Jones G (1968) Notes on microfungi III. Trans Br Mycol Soc 51:185–206
Pratibha SJ, Gawas P, Hyde KD, Bhat DJ (2005) Chalara indica sp. nov. and Sorocybe indicus sp. nov. from India. Cryptogam Mycol 26:97–103
Raitviír A (2004) Revised synopsis of the Hyaloscyphaceae. Scr Mycol Tartu 20:1–133
Raitviír A, Schneller J (2007) Some new Helotiales on ferns from South Africa. Sydowia 59:255–265
Rannala B, Yang Z (1996) Probability distribution of molecular evolutionary trees: a new method of phylogenetic inference. J Mol Evol 43:304–311
Rao PR (1970) Studies on hyphomycetes from the pacific north-west – IV. Mycopathol Mycol Appl 41:3–4
Rayner RW (1970) A mycological colour chart. CMI, Kew
Réblová M (1999) Teleomorph-anamorph connections in Ascomycetes 2. Ascochalara gabretae gen. et ap. nov. and its Chalara-like anamorph. Sydowia 51:210–222
Réblová M (2004) Four new species of Chaetosphaeria from New Zealand and redescription of Dictyochaeta fuegiana. Stud Mycol 50:171–186
Réblová M, Winka K (2000) Phylogeny of Chaetosphaeria and its anamorphs based on morphological and molecular data. Mycologia 92:939–954
Réblová M, Gams W, Stepanek V (2011) The new hyphomycete genera Brachyalara and Infundichalara, the similar Exochalara and species of ‘Phialophora sect. Catenulatae’ (Leotiomycetes). Fungal Divers 46:67–86
Réblová M, Nekvindová J, Fournier J, Miller AN (2020) Delimitation, new species and teleomorph-anamorph relationships in Codinaea, Dendrophoma, Paragaeumannomyces and Striatosphaeria (Chaetosphaeriaceae). MycoKeys 74:17–74
Réblová M, Kolařík M, Nekvindová J, Réblová J, Sklenář F, Miller AN, Hernández-Restrepo M (2021a) Phylogenetic reassessment, taxonomy, and biogeography of Codinaea and similar fungi. J Fungi 7(12):1097
Réblová M, Nekvindová J, Hernández-Restrepo M (2021b) Reflections on Menisporopsis, Multiguttulispora and Tainosphaeria using molecular and morphological data. J Fungi 7(6, no 438):1–36
Réblová M, Nekvindová J, Kolařik M, Hernández-Restrepo M (2021c) Delimination and phylogeny of Dictyochaeta, and introduction of Archochaeta and Tubulicolla, genera nova. Mycologia 113:390–433
Réblová M, Nekvindová J, Miller AN (2021d) Phylogeny and taxonomy of Catenularia and similar fungi with catenate conidia. MycoKeys 81:1–44
Richardson MJ (1972) Coprophilous ascomycetes on different dung types. Trans Br Mycol Soc 58:37–48
Ronquist F, Teslenko M, van der Mark P, Ayres DL (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61:539–542
Samuels GJ, Rogerson CT (1990) Some ascomycetes (fungi) occurring on tropical ferns. Brittonia 42:105–115
Sánchez Márquez S, Bills GF, Zabalgogeazcoa I (2007) The endophytic mycobiota of the grass Dactylis glomerata. Fungal Divers 27:171–195
Savic D (2020) Addition of data to the checklists of fungi of the classes leotiomycetes and pezizomycetes of serbi. Bull Nat Hist Mus 13:61–85
Seifert KA (1989) Coryne trichophora comb. nov, and the implications of its conidial anatomy. Stud Mycol 31:157–164
Seifert KA, Morgan-Jones G, Gams W, Kendrick WB (2011) The genera of hyphomycetes. CBS Biodiversity Series, 9, 1–997 CBS-KNAW Fungal Biodiversity Center, Utrecht
Seaver FJ (1934) Photographs and descriptions of cup-fungi-XXI. The genus Calycina. Mycologia 36(4):344–347
Seaver FJ (1942) Photographs and descriptions of cup-fungi. XXXVII. Peziza purpurescens. Mycologia 34:412–415
Senwanna C (2019) Nomenclatural novelties. Index Fungorum 421:1–1
Shabunin DA (2007) Two new anamorphic fungi on alder from Novgorod and Pskov regions. Mikol Fitopatol 41:337–341
Silva SS, Silva CR, Gusmao LFP, Castañeda-Ruiz RF (2015) A new species of Chaetochalara on decaying leaves from Brazil. Mycotaxon 130:505–509
Spegazzini C (1909) Mycetes Argentinenses, ser. IV. Anales Mus Nac Buenos Aires, ser. III, Tomo XII:257–458
Spooner BM (1987) Helotiales of Australasia: Geoglossaceae, Orbiliaceae, Sclerotiniaceae, Hyaloscyphaceae. Bibl Mycol 116:1–711
Spooren M (2014) A new species of Bloxamia from freshwater in the Netherlands. Mycosphere 5:346–349
Subramanian CV (1958) Hyphomycetes – V. J Indian Bot Soc 37:47–64
Subramanian CV, Sudha K (1986) Hyphomycetes from leaf litter-II. Kavaka 14:37–40
Suija A, Motiejünaité J (2016) Calycina alstrupii sp. nov. (Pezizellaceae, Helotiales), a new lichenicolous fungus from Norway. Phytotaxa 307:113–122
Sutton BC (1975) Hyphomycetes on cupules of Castanea sativa. Trans Br Mycol Soc 64:405–426
Sutton BC (1980) The Coelomycetes. CMI, Kew
Sutton BC (1989) Notes on Deuteromycetes II. Sydowia 41:330–343
Sutton BC (1993) Mitosporic fungi from Malawi. Mycol Pap 167:1–93
Sutton BC, Hodges CS (1976) Eucalyptus microfungi: some setose hyphomycetes with phialides. Nova Hedwigia 27:343–352
Sutton BC, Pirozynski KA (1965) Notes on microfungi. II. Trans Br Mycol Soc 48:349–366
Svrček M (1982) Mykofióra chráněného naleziště “Na ostrově” a Nemize. Sbor Vlast. Prací Podblan Benešov 23:75–98
Svrček M (1984) A taxonomic revision of inoperculate discomycetes described by J. Velenovský in the genus Helotium, preserved in National Museum, Prague. Acta Musei Nationalis Pragae 40B:129–215
Svrček M (1992a) New or less known Discomycetes. XXII. Ceská Mykol 46:33–40
Svrček M (1992b) On the genus Didonia Vel. (Helotiales). Ceská Mykol 46:41–49
Sydow H (1927) Fungi in itinere costaricensi collecti. Ann Mycol 25:1–160
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 60. Mol Biol Evol 30:2725–2729
Taylor JE, Crous PW, Palm ME (2001) Folia and stem fungal pathogens of Proteaceae in Hawaii. Mycotaxon 78:449–490
Tedersoo L, Bahram M, Puusepp R, Nilsson RH, James TY (2017) Novel soil-inhabiting clades fill gaps in the fungal tree of life. Microbiome. https://doi.org/10.1186/s40168-017-0259-5
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The CLUSTAL-X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Traue H, Arnold GRW (1991) Chalara ovoidea Nag Raj & Kendrick – eine weitere Pilzart im Krankheitsgeschehen der Trauben und Stieleichen. Feddes Repert 102:133–136
Tsui KM, Hyde KD, Hodgkiss IJ (2001) Longitudinal and temporal distribution of freshwater ascomycetes and dematiaceous hyphomycetes on submerged wood in the Lam Tsuen River, Hong Kong. J N Am Benthol Soc 20:533–549
Upadhyay HP (1981) A monograph of Ceratocystis and Ceratocystiopsis. University of Georgia Press, Athens
Velenovský J (1934) Monographia Discomycetum Bohemiae. 1–436
Verkley GJM (1999) A monograph of the genus Pezicula and its anamorphs. Stud Mycol 44:1–180
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J Bacteriol 172:4239–4246
von Bonsdorff T, Niskanen T, Liimatainen K, Kytövuori I, Huhtinen S, Vauras J, Höijer P, Kekki T, Lahti M, Puolasmaa A, Purhonen J, Halme P, Jakobsson S, Toivonen M, Söderholm U (2016) New national and regional biological records for Finland 8. Contributions to agaricoid, gastroid and ascomycetoid taxa of fungi 5. Memoranda Societatis pro Fauna Et Flora Fennica 92:120–128
von Höhnel F (1919) Vierte vorläufige Mitteilung mykologische Ergebnisse (Nr. 305–398). Ber Deutsch Bot Ges 37:107–115
Von Höhnel F (1923) Fragmente zur Mykologie. XXV. Mitteilung, Nr. 1215 bis 1225. Sitz.-ber. Akad. Wiss. Wien, Math.-nat. Kl. Abt I 132:89–118
Vu D, Groenewald M, de Vries M, Gehrmann T, Stielow B, Eberhardt U, Al-Hatmi A, Groenewald JZ, Cardinali G, Houbraken J, Boekhout T, Crous PW, Robert V, Verkley GJM (2019) Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud Mycol 92:135–154
Wang CJK, Sutton BC (1984) Two new synnematous genera: Chalarodendron and Paradidymobotryum. Mycologia 76:569–572
Wang Z, Johnston PR, Takamatsu S, Spatafora JW, Hibbett DS (2007) Toward a phylogenetic classification of the Leotiomycetes based on rDNA data. Mycologia 98:1065–1075
Webster J, Hawksworth DL (1986) Pyxidiophora spinulo-rostrata, a new species with denticulate conidiophores from submerged twigs in South-West England. Trans Br Mycol Soc 87:77–79
Weir A, Beakes G (1995) An introduction to the Laboulbeniales: a fascinating group of entomogenous fungi. Mycoligist 9:6–10
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic Press, California, pp 315–322
Willoughby LG, Archer JF (1973) The fungal spore of a freshwater stream and its colonization pattern on wood. Freshw Biol 3:219–239
Wingfield MJ, Van Wyk PS, Wingfield BD (1987) Reclassification of Phialocephala based on conidial development. Trans Br Mycol Soc 89:509–520
Wingfield MJ, Benade E, Van Wyk PS, Visser C (1995) Conidium development in Ceratocystis autographa. Mycol Res 99:1289–1294
Wu WP (2004) Chalara and allied genera from China: new species and new records of Chalara from China. Mycosystema 23:313–323
Wu WP, Diao YZ (2022) The anamorphic Chaetosphaeriaceous fungi from China. Fungal Divers 116:1–546
Xu JJ, Zhang TY (2012) A preliminary report of dematiaceous hyphomycetes from soil in tropical Yunnan, China. Mycosystem 31:17–33
Yu XD, Dong W, Bhat DJ, Boonmee S, Zhang D, Zhang H (2018) Cylindroconidiis aquaticus gen. et sp. nov., a new lineage of aquatic hyphomycetes in Sclerococcaceae (Eurotiomycetes). Phytotaxa 372:79–87
Zhang YH, Zhuang WY (2004) Phylogenetic relationships of some members in the genus Hymenoscyphus (Ascomycetes, Helotiales). Nova Hedwigia 78:3–4
Zhao YJ, Hosoya T, Baral HO, Hosaka K, Kakishima M (2013) Hymenoscyphus pseudoalbidus, the correct name for Lambertella albida reported from Japan. Mycotaxon 122:25–41
Zhaxybayer O, Gogarten JP (2002) Bootstrap, Bayesian probability and maximum likelihood mapping: exploring new tools for comparative genome analyses. BMC Genomics 3:1–15
Zheng HD, Zhuang WY (2013) Four new species of the genus Hymenoscyphus (fungi) based on morphology and molecular data. Sci China Life Sci 56:90–100
Zheng HD, Zhuang WY (2014) Hymenoscyphus albidoides sp. nov. and H. pseudoalbidus from China. Mycol Prog 13:625–638
Zhuang WY, Hyde KD (2001) Discomycetes of tropical China. V. Species new to Hong Kong. Fungal Divers 6:181–188
Acknowledgements
This project was partly supported by the Chinese Natural Science Foundation (31093440). This work could not have been accomplished without the courtesy, cooperation, and support of many friends during the field trips. All laboratory work was carried out in in the R&D Center, Novozymes China, Beijing, from which both authors received tremendous encouragement and support in time and funding for this work. Special thanks to Zhang Yan who did most of lab work of strain preservation, cultivation and molecular biology work, to Yan Huang, Shiying Shang, Yu Zhang, Jie Zheng for their great support in field trip and lab work. Special thanks to Dr. Konstanze Bensch in the MycoBank team (CBS) for reviewing all Latin names of new taxa. Finally, we are grateful to the three reviewers for very constructive comments and valuable suggests, which have helped to improve the quality of this paper.
Funding
This work was supported by The Chinese Natural Science Foundation [31093440]. Author Wenping Wu and Yongzhao Diao have received research support from Novozymes (China) Investment Co. Ltd.
Author information
Authors and Affiliations
Contributions
Both authors contributed equally to the publication. All classical mycological work was conducted by WW. The phylogenetic analyses were conducted by YD. Both authors contributed to the study conception and design. Material preparation, data collection and analyses were performed by WW and YD. The first draft of the manuscript was written by WW and both authors commented on previous versions of the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Handling Editor: Jian-Kui Liu.
Supplementary Information
Below is the link to the electronic supplementary material.
13225_2023_515_MOESM1_ESM.tif
Supplementary file1 (TIF 14969 kb) SFig. 1 Chalara affinis (Wu16147). a–g Conidiophores and phialidic conidiogenous cells. h, i Conidia. Scale bars: 5 µm
13225_2023_515_MOESM2_ESM.tif
Supplementary file2 (TIF 10883 kb) SFig. 2 Chalara parilis (Wu7569). a–i Conidiophores and phialidic conidiogenous cells. Scale bar: 10 µm for i; 5 µm for a–h
13225_2023_515_MOESM3_ESM.tif
Supplementary file3 (TIF 15934 kb) SFig. 3 Chalara platanicola (Wu8177). a–j, o, p Conidiophores and phialidic conidiogenous cells. k–m Conidia. Scale bar: 10 µm for a–h, 5 µm for i–p
13225_2023_515_MOESM4_ESM.tif
Supplementary file4 (TIF 18629 kb) SFig. 4 Chalara longiphora (Wu16192). a–j Conidiophores and conidiogenous cells. k–o Conidia. Scale bar: 5 µm
13225_2023_515_MOESM5_ESM.tif
Supplementary file5 (TIF 17834 kb) SFig. 5 Nagrajchalara acuaria (Wu1382f). a, c–f Conidiophores and conidiogenous cells. b, g–i Conidia. Scale bar: 10 µm for a, 5 µm b–i
13225_2023_515_MOESM6_ESM.tif
Supplementary file6 (TIF 14549 kb) SFig. 6 Nagrajchalara aspera (Wu6052). a–c Setae. d–g Conidiophores and conidiogenous cells. h–k Conidia. Scale bar: 10 µm for a–c, 5 µm for d–k
13225_2023_515_MOESM7_ESM.tif
Supplementary file7 (TIF 13799 kb) SFig. 7 Nagrajchalara aspera (Wu11022). a–e Conidiophores and conidiogenous cells. f–i Conidia. Scale bar: 5 µm
13225_2023_515_MOESM8_ESM.tif
Supplementary file8 (TIF 11916 kb) SFig. 8 Nagrajchalara cannonii (Wu16066). a mycelium and conidiophores on substrate. b–f Conidiophores and conidiogenous cells. g–k Conidia. Scale bar: 10 µm for a, g–k; 5 µm for b–f, h, i
13225_2023_515_MOESM9_ESM.tif
Supplementary file9 (TIF 15434 kb) SFig. 9 Nagrajchalara cannonii (Wu8283b). a–e, h–o Conidiophores and conidiogenous cells. f, g Conidia. Scale bar: 10 µm for h–k, 5 µm for a–e, f, g, l–o
13225_2023_515_MOESM10_ESM.tif
Supplementary file10 (TIF 14672 kb) SFig. 10 Nagrajchalara curviphora (Wu15021, Holotype). a–e Conidiophores and conidiogenous cells. Scale bar: 10 µm
13225_2023_515_MOESM11_ESM.tif
Supplementary file11 (TIF 11571 kb) SFig. 11 Nagrajchalara inflatipes (Wu12532). a–g Conidiophores and conidiogenous cells. n Conidia. Scale bar: 10 µm
13225_2023_515_MOESM12_ESM.tif
Supplementary file12 (TIF 17672 kb) SFig. 12 Nagrajchalara paraunicolor (Wu8194b, holotype). a–d Conidiophores and conidiogenous cells. e superficial mycelium. Scale bar: 5 µm
13225_2023_515_MOESM13_ESM.tif
Supplementary file13 (TIF 22192 kb) SFig. 13 Nagrajchalara puerensis (Wu15179). a–h, l–n Conidiophores and phialidic conidiogenous cells. i–k Conidia. Scale bar: 10 µm for a–e, 5 µm for f–n
13225_2023_515_MOESM14_ESM.tif
Supplementary file14 (TIF 21908 kb) SFig. 14 Nagrajchalara pulchra (Wu1354b). a–i Conidiophores and conidiogenous cells. Scale bar: 10 µm for a–f, 5 µm for g–i
13225_2023_515_MOESM15_ESM.tif
Supplementary file15 (TIF 13418 kb) SFig. 15 Nagrajchalara pulchra (Wu6107). a–c, e–i Conidiophores and conidiogenous cells. d Conidia. Scale bar: 5 µm
13225_2023_515_MOESM16_ESM.tif
Supplementary file16 (TIF 20166 kb) SFig. 16 Nagrajchalara sichuanensis (Wu13251, holotype). a–j Conidiophores with cylindrical collarettes. k–n Conidia. Scale bar: 5 μm
13225_2023_515_MOESM17_ESM.tif
Supplementary file17 (TIF 9754 kb) SFig. 17 Nagrajchalara sichuanensis (Wu13203). a–e, g Conidiophores and phialidic conidiogenous cells. f, h–l Conidia. Scale bar: 10 µm for a–d, 5 µm for e–l
13225_2023_515_MOESM18_ESM.tif
Supplementary file18 (TIF 22283 kb) SFig. 18 Nagrajchalara unicolor (a, b, r, s from Wu1322b; c–q from Wu2052a). a–i Conidiophores and conidiogenous cells. j Superficial mycelium. k–r Conidia. s Conidiophores bearing collarette with constriction at the base. Scale bar: 10 µm for a–f; p, q; 5 µm for g–o, r, s
13225_2023_515_MOESM19_ESM.tif
Supplementary file19 (TIF 12966 kb) SFig. 19 Nagrajchalara versicolor (Wu5542a, holotype). a–b Conidiophores and phialidic conidiogenous cells. c–g Conidia. Scale bar: 10 µm for a–d; 5 µm for e–g
13225_2023_515_MOESM20_ESM.tif
Supplementary file20 (TIF 19265 kb) SFig. 20 Nagrajchalara wenyingiae (Wu8010, holotype). a–f Conidiophores and conidiogenous cells. Scale bar: 5 µm
13225_2023_515_MOESM21_ESM.tif
Supplementary file21 (TIF 14957 kb) SFig. 21 Cylindrocephalum clavatisetosum (Wu11011, holotype). a–f Conidiophores and phialidic conidiogenous cell. Scale bar: 5 µm
13225_2023_515_MOESM22_ESM.tif
Supplementary file22 (TIF 15195 kb) SFig. 22 Cylindrocephalum clavatisetosum (Wu5550c). a, b, h–k Conidiophores and phialidic conidiogenous cells. c–g, l Conidia. Scale bar: 10 µm for a–b, h, 5 µm for c–g, i–l
13225_2023_515_MOESM23_ESM.tif
Supplementary file23 (TIF 17990 kb) SFig. 23 Nagrajchalara folicola and related species. a–c Conidiophores and phialidic conidiogenous cells. d–p Conidia. Scale bar: 5 µm
13225_2023_515_MOESM24_ESM.tif
Supplementary file24 (TIF 1832 kb) SFig. 24 Maximum likelihood (ML) tree based on 18S rDNA and 28S rDNA sequence data for the chalara-like anamorphic fungi in Leotiomycetes. Bootstrap support values ≥ 60 %, Bayesian posterior probability values ≥ 0.80 are shown at the nodes. Fuscocatenula variegata Wu12520 was chosen as the outgroup. Highlighted generic names are members of chalara-like fungi
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

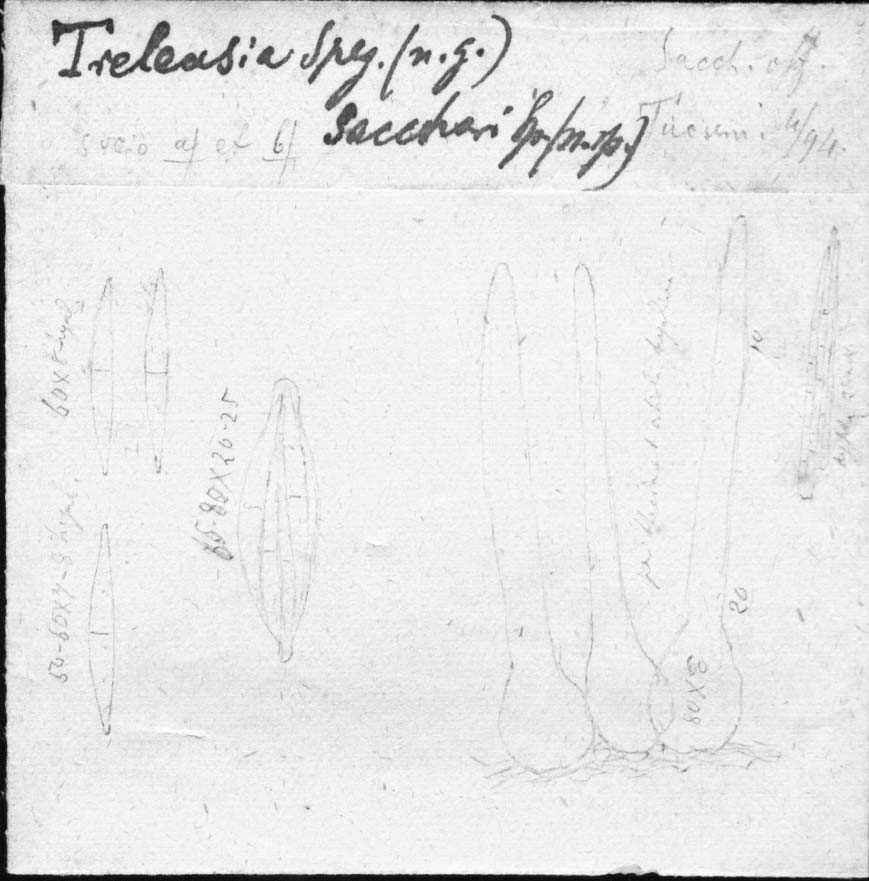{kind=link}
Cite this article
Wu, W., Diao, Y. The chalara-like anamorphs of Leotiomycetes. Fungal Diversity 119, 213–490 (2023). https://doi.org/10.1007/s13225-023-00515-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13225-023-00515-6