
Abstract
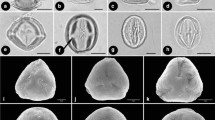
The neotropical subtribe Cuspariinae (Rutaceae) comprises as many as 26 genera and over 125 species. Pollen grains from 111 collections representing 71 species and 24 genera were examined by LM, SEM, and TEM. The pollen morphology of this subtribe is very diverse. Grains are mostly 3–6-aperturate and colporate, rarely porate (Spiranthera) or pantocolporate (Almeidea). Exine sculpturing is most commonly reticulate, sometimes perforate, foveolate-perforate, foveolate, foveolate-reticulate, reticulate, striate-reticulate, echinate, clavate, or baculate. The exine structure is columellate and tectate-perforate, columellate and semitectate, or intectate and is stratified into ektexine and endexine. The exine ofLeptothyrsa is distinctive in that the ektexine of the mesocolpium is longitudinally deeply ridged. The pollen ofHortia, characterized by a psilate exine with rare perforations, a very thick foot-layer, and reduced columellae, is unlike that of any member of the Cuspariinae and offers no support for the transfer of this genus from the Toddalioideae. The pollen data correlate with macromorphological characters and are taxonomically useful.
Similar content being viewed by others

Literature Cited
Barth, O. M. 1982. Variaçoes polínicas em espécies brasileiras da família Rutaceae. Bol. Inst. Geoci. Univ. São Paulo 13: 129–134.
Barth, O. M. 1983. Pollen morphology of Brazilian Rutaceae: suggestions for a more natural taxonomy 409–419.
— 1985. Pollen morphology of Brazilian Rutaceae:Pilocarpus. Pollen et Spores 27: 145–154.
Emmerich, M. 1978. Contribuição ao conhecimento da tribo Cusparineae [sic] (Rutaceae). Nova conceituação deRaputia e gêneros derivados. Rodriguesia 30, 45: 223–307.
Engler, A. 1931. Rutaceae. Pages 187–359.In: A. Engler & K. Prantl. Natürl. Pflanzenfam, ed. 2. 19a.
Erdtman, G. 1969. Handbook of palynology. Munksgaard Co., Copenhagen.
— 1971. Pollen morphology and plant taxonomy: angiosperms. Hafner Publishing Co., New York.
Fernandes da Silva, M. F. das G., O. R. Gottlieb &F. Ehrendorfer. 1988. Chemosystematics of the Rutaceae: suggestions for a more natural taxonomy and evolutionary interpretation of the family. Plant Syst. Evol. 161: 97–134.
Gereau, R. 1990.Achuaria, nuevo género de Rutaceae, con una sinopsis de las Cuspariinae peruanas. Contribución al estudio de la flora y de la vegetación de la Amazonía peruana. XVIII. Candollea 45: 363–372.
Heusser, C. J. 1971. Pollen and spores of Chile. University of Arizona Press, Tucson, Arizona.
Kallunki, J. A. 1990. An emended description of and new combinations inRaputia Aublet (Cuspariinae, Rutaceae). Brittonia 42: 175–177.
— 1992. A revision ofErythrochiton sensu lato (Cuspariinae, Rutaceae). Brittonia 44: 107–139.
Selling, O. 1947. Studies in Hawaiian pollen statistics, Part II. The pollens of the Hawaiian phanerogams. Spec. Publ. Bishop Mus. 38. Gothenburg.
Steyermark, J. A. 1984. Flora of the Venezuelan Guyana—I. Ann. Missouri Bot. Gard. 71: 297–340.
van der Ham, R. W. J. M. 1990. Nephelieae pollen (Sapindaceae): form, function, and evolution. Rijksherbarium/Hortus Botanicus. Leiden.
Walker, J. W. &J. A. Doyle. 1975. The bases of angiosperm phylogeny: palynology. Ann. Missouri Bot. Gard. 62: 664–732.
Ybert, J.-P. 1979. Atlas de pollens de Côte D'Ivoire. O.R.S.T.O.M., Paris.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Morton, C.M., Kallunki, J.A. Pollen morphology of the subtribe Cuspariinae (Rutaceae). Brittonia 45, 286–314 (1993). https://doi.org/10.2307/2807604
Issue Date:
DOI: https://doi.org/10.2307/2807604