
Phylogenomics and classification of Notropis and related shiners (Cypriniformes: Leuciscidae) and the utility of exon capture on lower taxonomic groups
- Published
- Accepted
- Received
- Academic Editor
- Dany Garant
- Subject Areas
- Genomics, Molecular Biology, Taxonomy, Zoology, Freshwater Biology
- Keywords
- Phylogenomics, Cypriniformes, Leuciscidae, Notropis, Systematics, North America, Shiners
- Copyright
- © 2022 Stout et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2022. Phylogenomics and classification of Notropis and related shiners (Cypriniformes: Leuciscidae) and the utility of exon capture on lower taxonomic groups. PeerJ 10:e14072 https://doi.org/10.7717/peerj.14072
Abstract
North American minnows of the Shiner Clade, within the family Leuciscidae, represent one of the most taxonomically complex clades of the order Cypriniformes due to the large number of taxa coupled with conserved morphologies. Species within this clade were moved between genera and subgenera until the community decided to lump many of the unclassified taxa with similar morphologies into one genus, Notropis, which has held up to 325 species. Despite phylogentic studies that began to re-elevate some genera merged into Notropis, such as Cyprinella, Luxilus, Lythrurus, and Pteronotropis, the large genus Notropis remained as a taxonomic repository for many shiners of uncertain placement. Recent molecular advances in sequencing technologies have provided the opportunity to re-examine the Shiner Clade using phylogenomic markers. Using a fish probe kit, we sequenced 90 specimens in 87 species representing 16 genera included in the Shiner Clade, with a resulting dataset of 1,004 loci and 286,455 base pairs. Despite the large dataset, only 32,349 bp (11.29%) were phylogenetically informative. In our maximum likelihood tree, 78% of nodes are 100% bootstrap supported demonstrating the utility of the phylogenomic markers at lower taxonomic levels. Unsurprisingly, species within Notropis as well as Hudsonius, Luxilus, and Alburnops are not resolved as monophyletic groups. Cyprinella is monophyletic if Cyprinella callistia is excluded, and Pteronotropis is monophyletic if it includes Hudsonius cummingsae. Taxonomic changes we propose are: restriction of species included in Alburnops and Notropis, elevation of the subgenus Hydrophlox, expansion of species included in Miniellus, movement of Hudsonius cummingsae to Pteronotropis, and resurrection of the genera Coccotis and Paranotropis. We additionally had two specimens of three species, Notropis atherinoides, Ericymba amplamala, and Pimephales vigilax and found signficant differences between the localities (1,086, 1,424, and 845 nucleotides respectively).
Introduction
Among North American cyprinoids, the shiners and related minnows have been among the most taxonomically complex groups of fishes. The group is currently placed in the Leuciscidae, subfamily Pogonichthyinae after the elevation of subfamilies to family rank in cyprinoids (Schönhuth et al., 2018; Tan & Armbruster, 2018). Ichthyologists tasked with assembling species into meaningful genera initially described a dizzying array of genera and subgenera of minnows. Species were moved between various categories (genera, subgenera, tribes) based primarily on phenetic similarity; many leuciscid genera remained relatively stable but one, Notropis (and many taxa formally placed in this genus), has continued to be difficult. A search of the Catalog of Fishes (Fricke, Eschmeyer & van der Laan, 2022) for ‘Notropis’ results in 325 species, representing a considerable bulk of nominal species of North American freshwater fishes.
Starting with Mayden (1989) comprehensive morphological phylogenetic analysis of Cyprinella and other North American taxa, a large monophyletic group was recognized as the Open Posterior Myodome clade (OPM). This clade includes most of the eastern North American leuciscids including Notropis and related genera. Mayden (1989) concluded that Notropis is an artificial group due to convergence of morphological characters and classification by phenetic similarity, and enacted nomenclatural changes elevating some subgenera in Notropis, such as Cyprinella, Luxilus, Lythrurus, and Pteronotropis, recognized other genera and reallocated species. He moved some species from Notropis to Hybopsis (including N. boucardi, N. calientis, N. dorsalis, N. longirostris, N. sabinae, N. alborus, N. bifrenatus), and differentiated six species of Notropis (i.e. N. topeka, N. mekistocolas, N. atrocaudalis, N. stramineus, N. chihuahua and N. procne) from the large clade that included all of the genera previously included in Notropis. A few now recognized genera were then not recognized (i.e. Codoma was within Cyprinella; Opsopoedus was within Notropis). Still, Notropis remained as a “taxonomic repository for small, silvery fishes of unknown relationship” (Gidmark & Simons, 2014: 379; see also Mayden et al., 2006) with approximately 91 currently recognized species loosely organized into subgenera (Jordan, 1885). Primarily because of the large number of taxa, coupled with conserved morphologies, few studies have attempted to tackle the remaining species allocated to the genus or other orphaned taxa of unknown taxonomic placement (Mayden et al., 2006; Hollingsworth et al., 2013; Schönhuth et al., 2018). However, even when necessary taxonomic decisions for species within the genus Notropis were made (sensu Mayden et al., 2006; Gidmark & Simons, 2014), they have not been widely accepted, and traditional taxonomic groups have been preferred by some for nomenclatural stability until a stronger consensus is reached on proposed nomenclatorial changes (Fricke, Eschmeyer & van der Laan, 2022).
Phylogenetically, most previous studies on shiners and relatives have focused on resolving relationships within purported monophyletic subgenera (for example Snelson, 1972; Buth, 1979; Raley, Wood & McEachran, 2001; Cashner, Piller & Bart, 2011) with varied results, and without investigation into relationships among the subgenera or to genera that have been segregated from Notropis. Mayden et al. (2006) attempted a comprehensive study of the so-called Notropin clade (the clade name does not refer to a taxonomic rank) to resolve relationships using cytb (mitochondrial marker) and revealed a nonmonophyletic Notropis as reported in prior studies either based on morphological or molecular data (Mayden, 1989; Simons, Berendzen & Mayden, 2003). The former study, in addition to corroborating monophyly of genera synonymized with Notropis as Cyprinella, Erycymba, Hybognathus, Hybopsis, Lythrurus, also identified a more restricted Notropis by recognizing the additional genera Alburnops, Aztecula, Graodus, Hudsonius, Miniellus, and Yuriria. These authors could not resolve Hydrophlox, Luxilus and Pteronotropis as monophyletic, and identified a ‘Notropis’ longirostris clade that included seven species of Notropis. Despite all these monophyletic groups having available names, their compositions were not always as initially proposed, and relocation of some species were made by Mayden (1989), Coburn & Cavender (1992), and Mayden et al. (2006). However, following this alternative phylogenetic classification of Notropin shiners, many species remained relegated to ‘Notropis’ (in single quotes) because of their uncertain placement due to weak support in analyses, and relationships among the genera listed above remained unclear. While these additional genera from within a nonmonophyletic Notropis were recognized and elevated, most subsequent studies reverted back to a larger encompassing Notropis, with perhaps recognition of some of these genera as subgenera or species groups (Bird & Hernandez, 2007; Rüber et al., 2007; Zhang et al., 2008; Chen & Mayden, 2009; Fang et al., 2009; Gaubert, Denys & Oberdorff, 2009; Scott et al., 2009; Bufalino & Mayden, 2010; Houston, Shiozawa & Riddle, 2010; Cashner, Piller & Bart, 2011; Wang et al., 2012; Hollingsworth et al., 2013; Imoto et al., 2013; Fricke, Eschmeyer & van der Laan, 2022).
Hollingsworth et al. (2013) expanded upon the cytb study by adding the RAG1 (nuclear) molecular marker for a phylogenetic reconstruction to test for a correlation between a shift from benthic to pelagic lifestyles and increased diversification rates. Unsurprisingly, this analysis also resulted in a relatively poorly resolved overall phylogeny with moderate support for non-monophyly of Notropis, but again illustrated the importance of understanding relationships to better inform our understanding of ecological and evolutionary processes.
To promote further study into this group, Gidmark & Simons (2014) amassed much of the knowledge reported for the shiners (distributions, histories, ecologies, etc.) and proposed using the designations made by Mayden et al. (2006) with the understanding that the relationships among them still remain unclear, despite support for the Shiner Clade as a whole (Simons, Berendzen & Mayden, 2003; Mayden et al., 2006; Schönhuth et al., 2008). Recently, a classification of the Holarctic family Leuciscidae based on nuclear and mitochondrial genes was proposed, where the Shiner Clade was a well-supported group within the Pogonichthyinae (Schönhuth et al., 2018). Despite the increase in both taxon and character sampling, Notropis was not resolved as monophyletic, as species of this genus were found in different parts of the Shiner Clade, and some of the genera formerly included in Notropis were not supported as monophyletic (Alburnops, Notropis s.s., Hudsonius), or their composition differed slightly (i.e. Miniellus, Hydrophlox) from that previously proposed (Coburn, 1982; Mayden et al., 2006; Cashner, Piller & Bart, 2011).
Recent advances in sequencing technologies have provided the opportunity to re-examine the shiner clade using phylogenomic markers. Most phylogenomic-scale studies thus far have focused on higher taxonomic levels (Lemmon, Emme & Lemmon, 2012; Bond et al., 2014; Eytan et al., 2015; Prum et al., 2015; Hamilton et al., 2016), but decreases in costs and the establishment of universal loci specific to fishes (Betancur-R et al., 2013; Arcila et al., 2017; Hughes et al., 2018, 2021) have helped overcome the hurdles associated with applying a phylogenomic approach to the shiner clade. In this study, we employ the probes developed by Arcila et al. (2017) in an attempt to tackle the systematic problems of Notropis and related genera, and follow the taxonomy discussed in Gidmark & Simons (2014). We also examined two specimens of three species from different geographic locations to test the utility of the markers on a smaller geographic scale. Previous research (Rincon-Sandoval, Betancur-R & Maldonado-Ocampo, 2019) had shown good utility at population-level scales, but wih the shiners, we wanted to determine if the markers could lead to a stong phylogenetic hypothesis for a group with very rapid divergence (Hollingsworth et al., 2013; Burress et al., 2017).
Materials and Methods
Taxon selection, tissue preparation, and sequencing
Every effort was made to acquire broad representation across the shiner genera. For the present study we included 88 ingroup taxa from 16 genera currently included in the Shiner Clade as proposed by Schönhuth et al. (2018) as well as outgroup taxa from five different genera (Notemigonus, Chrosomus, Erimystax, Phoxinus and Semolitus; Table S1 shows genera with number of recognized species, type species, and species sampled). Six genera from the Shiner Clade were not available for this study including Tampichthys (six species), Yuriria (three species), Algansea (seven species), Aztecula (two species), Agosia (one species), and Erimonax (one species). Except for Agosia and Erimonax all other unsampled genera are endemic to Mexico. To test the utility of the markers at an even smaller taxonomic scale, we include two specimens each of Notropis atherinoides, Ericymba amplamala, and Pimephales vigilax.
DNA was extracted from specimens using the Omegabiotek E.Z.N.A. animal tissue extraction kit (product #D3396-02) following manufacturer protocols. Extracted DNA was checked for quality using electrophoresis and quantity using nanodrop. After ensuring high molecular weight and a minimum of 2 µg total DNA, samples were sent for library preparation and Illumina sequencing to MYcroarray (now Arbor Biosciences, arborbiosci.com). Probes developed by Arcila et al. (2017) were used to target 1,060 loci. GenBank project number is PRJNA842507; aligment (File S1) and partition file (File S2) are provided.
Bioinformatics and tree reconstruction
FASTQ files were uploaded to the Alabama Supercomputer Center (ASC) for preliminary quality control processing. Trimmomatic (Bolger, Lohse & Usadel, 2014) was used to remove adapters and remove leading and trailing low quality bases in the paired end reads, as well as to remove reads with a length less than 36 base pairs. Resulting reads were then imported into Geneious v 6.1.8 (www.geneious.com), set as paired reads, and assembled using the zebrafish (Danio rerio) reference for the concatenated loci using five iterations and trimmed to each reference locus. The loci for each species were then concatenated and all concatenations were aligned in Geneious v 6.1.8 with the native alignment tool (www.geneious.com). Tree reconstruction was performed on the Center for Advanced Science Innovation and Commerce (CASIC) computer cluster at Auburn University, Auburn, AL, USA. RAxML (Randomized Axelerated Maximum Likelihood, v. 8.0.24, Stamatakis, 2014) was implemented using GTR + G model of evolution on the partitioned loci (partitioned per Arcila et al., 2017) and the resulting tree then subjected to 500 bootstrap replicates (BS is percent of trees showing this result). Species tree reconstruction was conducted using ASTRAL-II (Mirarab & Warnow, 2015) on individual RAxML gene trees that were subjected to 100 bootstrap replicates. Approximately-unbiased (AU) tests were conducted using CONSEL v.0.20 (Shimodaira, 2002) to specifically test the unconstrained maximum likelihood best tree topology against trees that were constrained to force monophyly for three genera: Cyprinella, Hudsonius, and Luxilus. Number of phylogenetically informative sites was calculated in R (v. 4.0.2; R Core Team, 2020) using the pis command in the ips package (Interfaces to Phylogenetic Software in R; Heibl, Cusimano & Krah, 2014).
Results
The final alignment yielded 1,004 loci, 286,455 base pairs, and only 0.42% missing data. Of those sites, 32,349 (11.29%) were phylogenetically informative. The range of locus size was 196–1,748 bp, with an average bp length of 285 (Fig. 1). In the resulting concatenated ML tree, 78% of nodes are 100% bootstrap supported with only six nodes collapsing below the 70% bootstrap threshold (Fig. 2). Species tree analysis produced highly congruent results, particularly at the genus level (Fig. 3). At deeper nodes there is less support in the species tree for the placement of a few clades (i.e. Hudsonius hudsonius + H. altipinnis; ‘Notropis’ atrocaudalis + ‘N.’ bifrenatus + ‘N.’ heterolepis), resulting in remaining uncertainty as to the relationships among the genera. Nevertheless, with our focus on resolving within-genera relationships, both concatenation and species tree approaches resolve the same patterns with strong support.
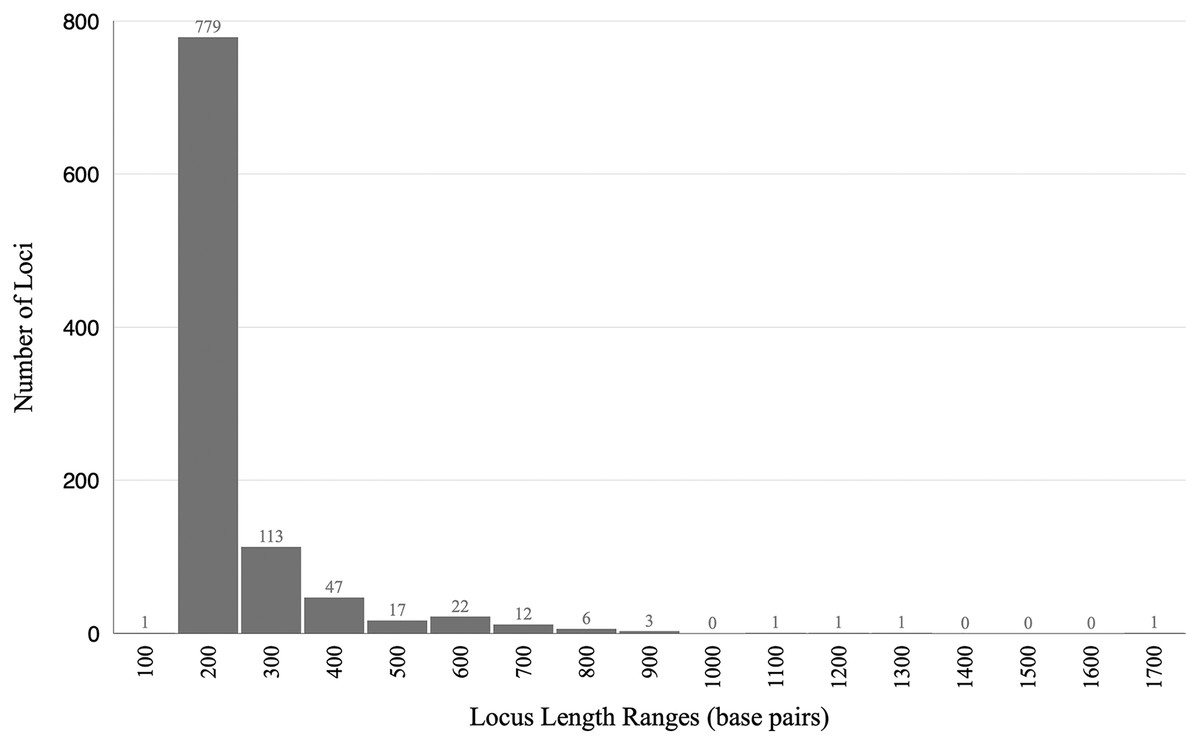
Figure 1: Histogram showing lengths of loci in base pairs.
The range of locus size was 196–1,748 bp, with an average bp length of 285.{kind=link}
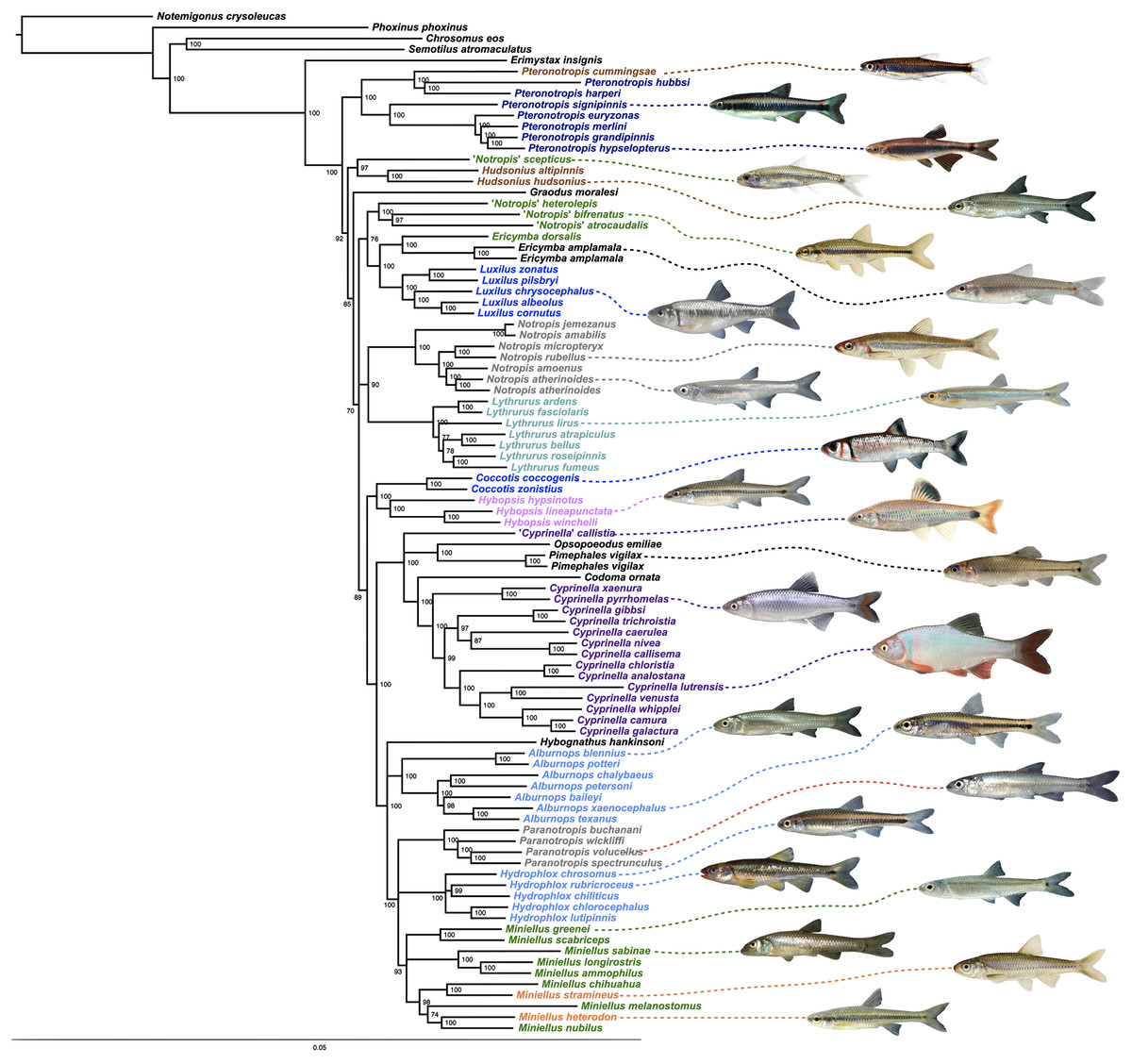
Figure 2: ML tree based on concatenated alignment.
Numbers of nodes represent bootstrap support, with nodes less than 70% supported collapsed. Scale bar represents number of substitutions per site.{kind=link}
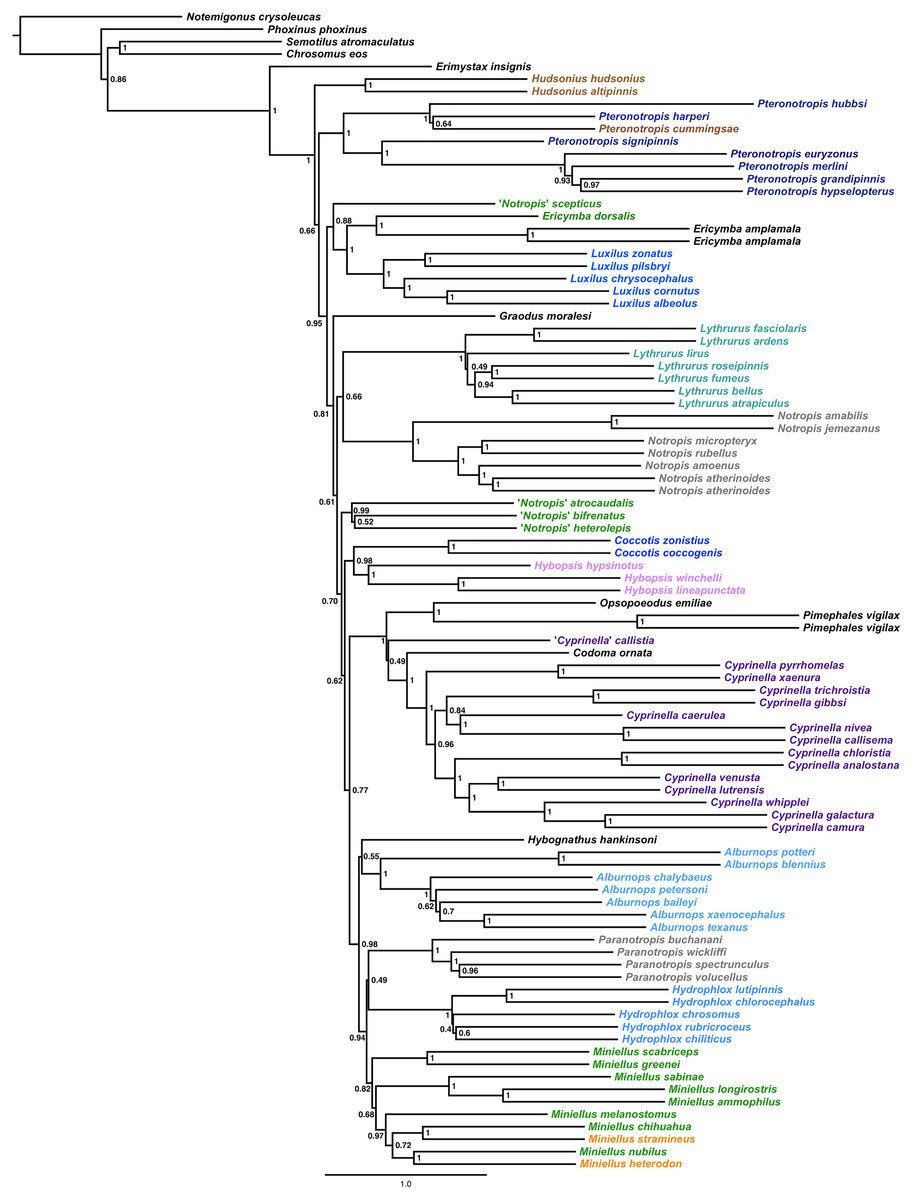
Figure 3: Species tree using ASTRAL-II.
Internal branch lengths are in coalescent units and branches that lead to tips are not calculated by ASTRAL-II but instead arbitrarily displayed. Branch support values indicate the support for a quadripartition (instead of bipartitions).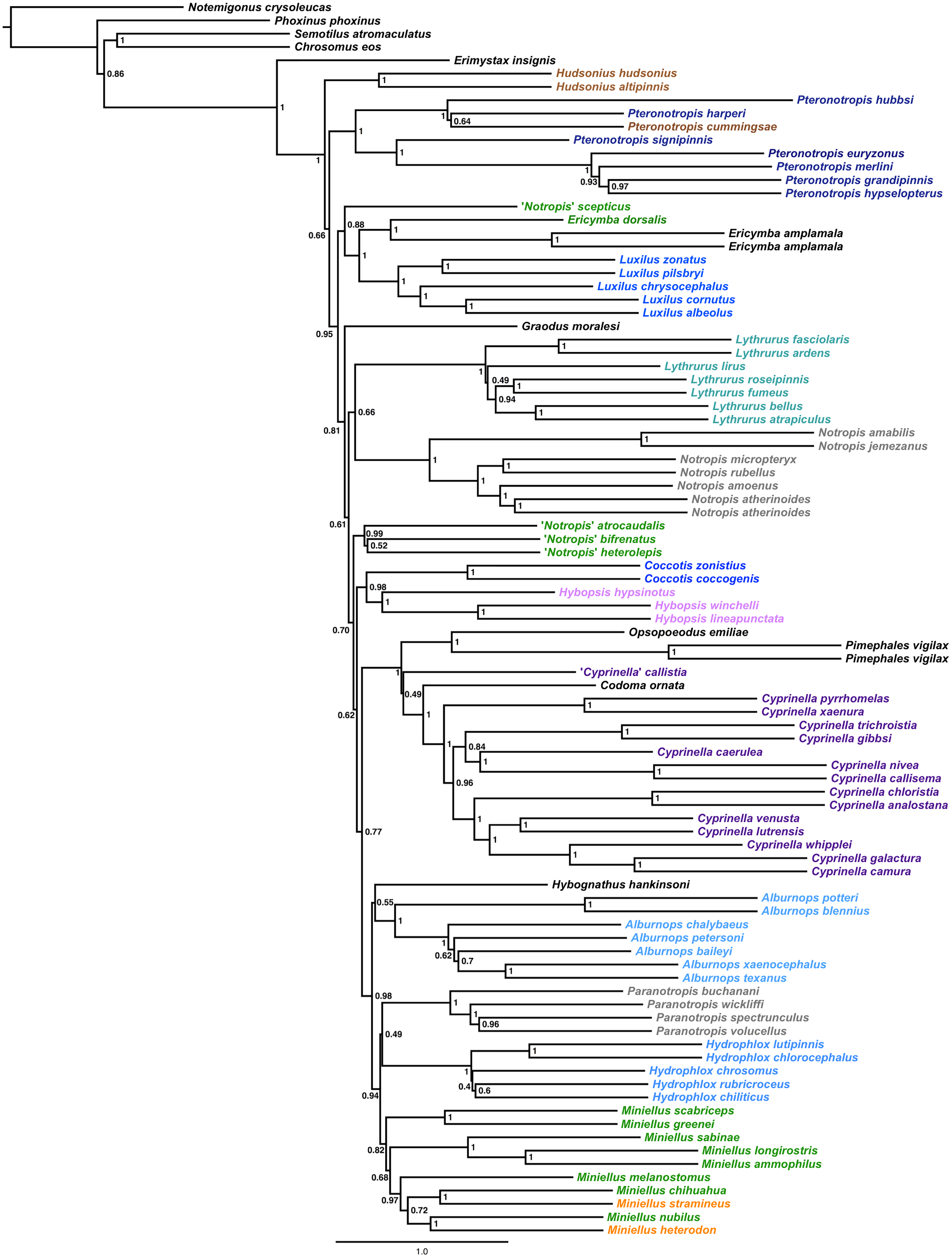{kind=link}
Species that are currently placed in Notropis are not resolved as monophyletic (Figs. 2 and 3). Notropis jemezanus, N. amabilis, N. micropteryx, N. rubellus, and N. amoenus form a clade with the type species, N. atherinoides, and a second clade included N. buchanani, N. wickliffi, N. volucellus, and N. spectrunculus. Species designated as ‘Notropis’ by Mayden et al. (2006) are found throughout the tree. The genera Hudsonius, Luxilus, and Alburnops were not monophyletic. Cyprinella forms a monophyletic group that excludes C. callistia, which forms a trichotomy with Opsopoeodus + Pimephales and the clade containing the remaining members of Cyprinella + Codoma. The results of the AU test were all significant (Cyprinella constrained, p = 3e−06; Hudsonius constrained, p = 6e−08; Luxilus constrained, p = 2e−18), indicating that all of the constrained topologies can be rejected as alternative tree hypotheses. For comparison, a summary of the four-gene phylogeny of Schönhuth et al. (2018) for the shiner clade is presented including the taxonomic changes proposed herein (Fig. 4). A list of all species, genera, and proposed taxonomic changes discussed below are given in Table S2.
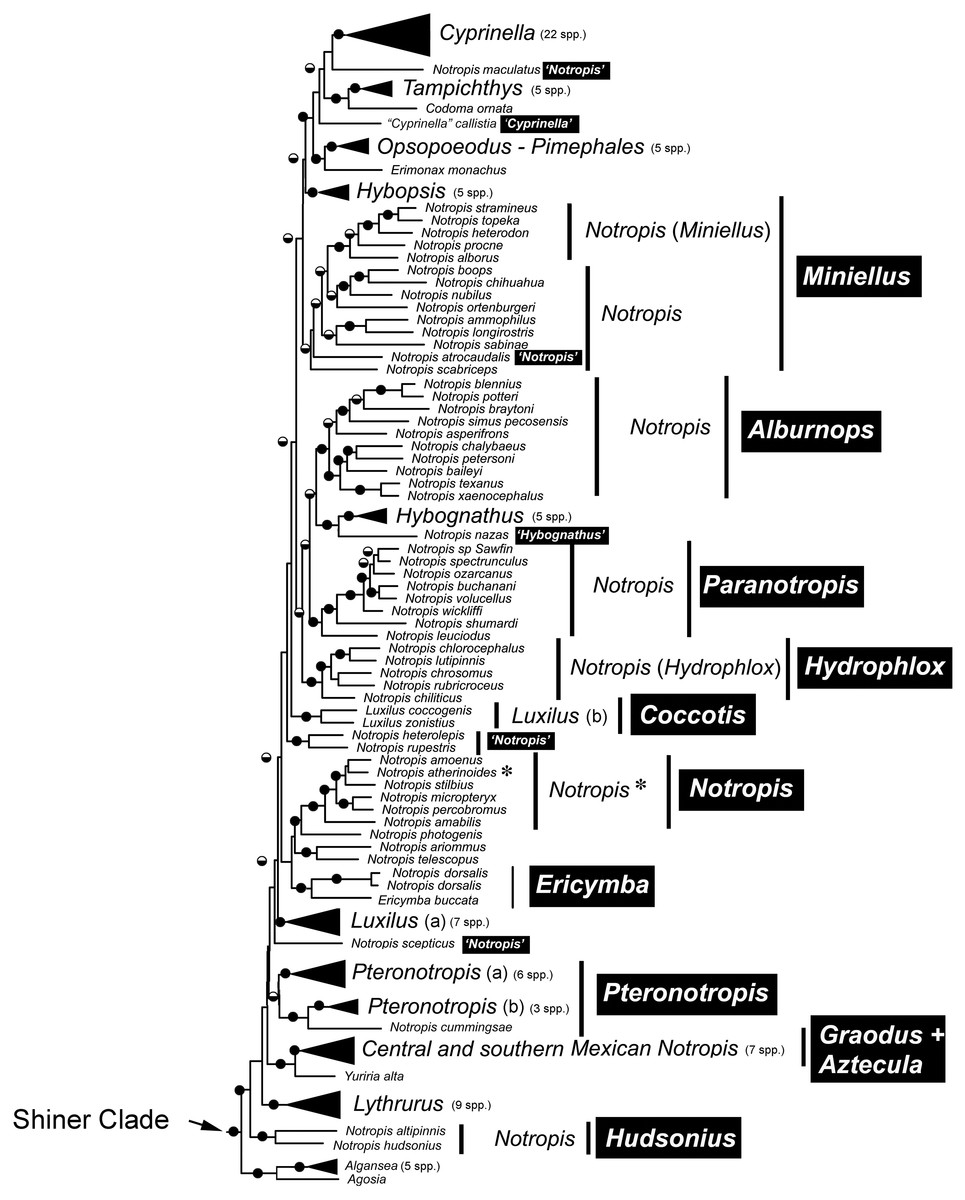
Figure 4: Major monophyletic groups and genera within the Shiner Clade based on two nuclear and two mitochondrial genes (modified from Schönhuth et al., 2018).
Black boxes with white text represent name changes. Circles at nodes represent >75% bootstrap values (BS, black top) and >95% Baysian posterior probability (PP, black bottom).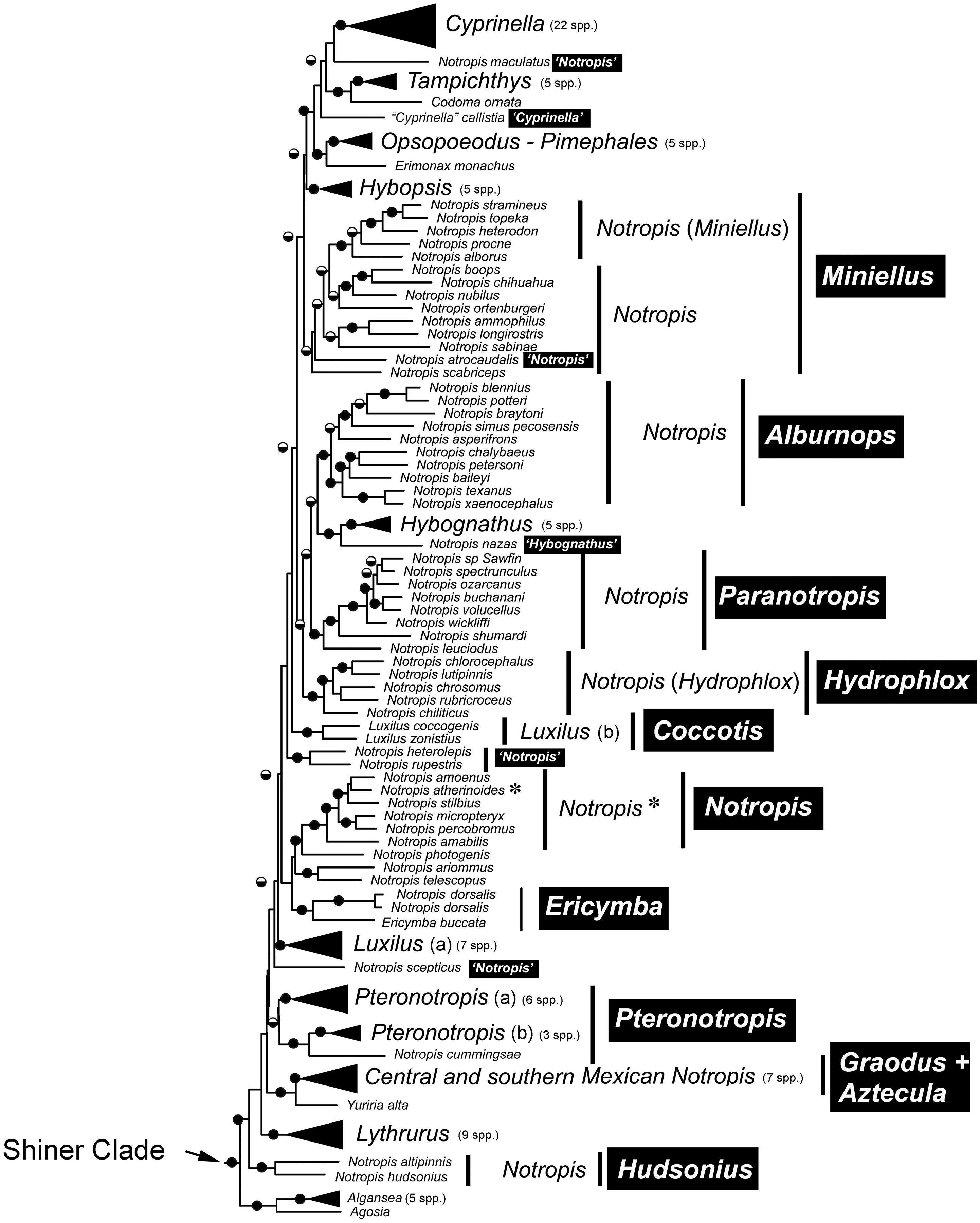{kind=link}
Discussion
Alternative taxonomic and systematic classifications have been proposed for species included within the controversial genus Notropis and various purported relatives in several studies based on different characters (Swift, 1970; Coburn, 1982; Mayden, 1989; Bielawski & Gold, 2001; Mayden et al., 2006; Gidmark & Simons, 2014; Schönhuth et al., 2018). However, while several genera have been elevated from synonymy with Notropis and are monophyletic groups, consensus regarding number and composition of some of the different clades and groups of the genus Notropis has remained elusive, and classifications were considered provisional until a more comprehensive study including sufficient taxon and character sampling can produce a well supported analysis of relationships (Cashner, Piller & Bart, 2011; Gidmark & Simons, 2014; Schönhuth et al., 2018). This study provides the results on which to make some, but not all of the remaining taxonomic considerations (Table S2). In general, we made taxonomic changes if this study and that of Schönhuth et al. (2018) concur and if there was high support values in Figs. 2 and 3 (>93% bootstrap and/or a local posterior probability of 1 in the ASTRAL-II tree). Taxonomic discussion roughly follows Fig. 2. Boostrap support (BS, Fig. 2) and local posterior probability (PP, Fig. 3) are indicated in the discussion of clades.
All phylogenomic analyses (based on concatenation and species tree approaches) resolved the same patterns with strong support. The 42 species analyzed previously included in Notropis were not resolved as monophyletic; however, analyses resolved these species in several well-supported clades, and nine other well-recognized genera, together providing an alternative classification for this controversial group of shiners. This revisionary classification for Notropis supported the recognition of the five genera previously synonymized with Notropis (Alburnops, Hudsonius, Hydrophlox, Miniellus and Paranotropis) and re-allocated some species within these groups with other genera within the Shiner Clade that differed from prior studies (Mayden, 1989; Mayden et al., 2006; Gidmark & Simons, 2014). This study also highlights some interesting relationships with other closely related genera within the Shiner Clade.
Pteronotropis and Hudsonius
All three species of Hudsonius were included in the analysis but were not recovered as monophyletic, with H. cummingsae grouping within Pteronotropis. The range for all three species of Hudsonius overlaps with that of Pteronotropis across the southeastern states of North and South Carolina, Georgia, Alabama, and Florida, but only H. hudsonius extends northward up through the Great Lakes and across much of Canada. Mayden et al. (2006) found support for a monophyletic Hudsonius, despite individuals of H. altipinnis not forming a lineage, suggesting cryptic speciation. Hollingsworth et al. (2013) found Hudsonius as monophyletic in concatenated analyses and with cytochrome b, but within Pteronotropis with just the nuclear gene Rag1. In our analysis, Hudsonius cummingsae is part of Pteronotropis (BS 100, PP 1), while H. altipinnis (collected in South Carolina) and H. hudsonius (collected in Wisconsin) were found as sister species (BS 100, PP 1), congruent with Schönhuth et al. (2018; Fig. 4). Given that mitochondrial markers are suggesting the monophyly of Hudsonius and nuclear markers are suggesting that H. cummingsae is within Pteronotropis, it is possible that there has been mitochondrial introgression between H. cummingsae and H. altipinnis; however, this will need further examination. Because H. hudsonius is the type species, we propose removing Hudsonius cummingsae from Hudsonius and transferring it to Pteronotropis to maintain monophyly of both genera Pteronotropis and Hudsonius. Pteronotropis is, in part, diagnosed by a wide, dark stripe on the body, a character that P. cummingsae shares.
Mayden (1989) and Simons, Knott & Mayden (2000) did not find Pteronotropis to be monophyletic; however the genus was monophyletic here (BS 100, PP 1) as well as in Mayden & Allen (2015) and Schönhuth et al. (2018). There are two deeply divergent clades, that of P. harperi + P. hubbsi + P. cummingsae + P. welaka, and that of P. euryzonus + P. grandipinnis + P. hypelopterus + P. merlini + P. metallicus + P. signipinnis + P. stonei, and it could be argued that these two clades deserve separate genus status. This deep divergence may have led to nonmonophyly in earlier studies without either enough characters and/or taxa in the analyses. In Schönhuth et al. (2018), the monophyly of Pteronotropis was supported only under Bayesian inference and not Maximum Likelihood (Fig. 4).
Luxilus
Our analysis includes seven of the nine recognized species of Luxilus and recovers two distinct clades. Luxilus chrysocephalus (type species) forms a clade (BS 100, PP 1) with L. zonatus, L. pilsbryi, L. albeolus, and L. cornutus that is sister to Ericymba + ‘Notropis’ dorsalis. Two other species, Luxilus coccogenis and L. zonistius, are resolved as a clade (BS 100, PP 1) distant to other members of Luxilus and instead sister to Hybopsis. Mayden (1989) removed Luxilus from Notropis, considering it sister to Cyprinella (including Codoma ornata) and monophyletic based on three morphological characters, while Coburn & Cavender (1992) considered Luxilus sister to a clade comprised of Lythrurus, Cyprinella, Pimephales, and Opsopoeodus, and they noted Luxilus could be an unnatural assemblage. Morphological and molecular studies primarily focused on members within Luxilus have assumed the monophyly of the genus, rather than including other genera, and have consistently found a sister relationship between L. coccogenis + L. zonistius, supporting our findings (Gilbert, 1964; Menzel, 1976; Dowling et al., 1992; Dowling & Naylor, 1997; Mayden et al., 2006); although an allozyme analysis found L. coccogenis and L. zonistius to be sister to L. cerasinus (Buth, 1979). Other studies that included Luxilus and a variety of other shiner taxa have argued that Luxilus is not monophyletic (Simons, Berendzen & Mayden, 2003; Mayden et al., 2006; Schönhuth & Mayden, 2010; Hollingsworth et al., 2013). We support that L. coccogenis + L. zonistius should no longer be included in Luxilus, and propose elevating the genus Coccotis Jordan 1882 (Coccotis coccogenis Jordan 1882 as the type species) for these taxa. These results are also in agreement with Schönhuth et al. (2018; Fig. 4) where Luxilus was resolved as nonmonophyletic.
Lythrurus
Lythrurus has long been considered monophyletic (Snelson, 1972; Schmidt, Bielawski & Gold, 1998; Mayden et al., 2006; Pramuk et al., 2007), and our study also supports the monophyly of this group (BS 100, PP 1). What has been more problematic, however, is determining the clade’s relationship to other genera. Formerly considered sister to a Luxilus + Cyprinella clade (Mayden, 1989), it was later poorly resolved by Mayden et al. (2006) in a clade with various ‘Notropis’ species. Coburn & Cavender (1992) determined Lythrurus was sister to a clade comprised of Cyprinella, Pimephales, and Opsopoeodus, and more recently it has been weakly resolved as the sister group to all other species of the Shiner Clade excluding Algansea-Agosia, and Hudsonius (Schönhuth et al., 2018). We find moderate support (BS 90, PP 0.66) for Lythrurus as sister to true Notropis (the clade containing Notropis atherinoides, the type species of Notropis; more discussion on Notropis below).
Cyprinella
Although Gibbs (1957) and Mayden (1989) found Cyprinella callistia as nested within Cyprinella, Broughton & Gold (2000) found the species to be sister to the remaining species of Cyprinella (Codoma and Tampichthys were not included). One of the most extensive and recent molecular studies of Cyprinella and relatives (Schönhuth & Mayden, 2010) failed to resolve the genus as monophyletic, relative to C. callistia. The remainder of Cyprinella was monophyletic, and was sister to Codoma + Tampichthys; however, Cyprinella callistia was either sister to the clade of Cyprinella + Codoma + Tampichthys or as sister to that clade plus Pimephales + Opsopoeodus. We did not include Tampichthys, but we also resolved Codoma more closely related to all other representatives of Cyprinella (BS 100, PP 1) than Cyprinella callistia. As in previous analyses, we could not resolve the node leading to Cyprinella callistia, Opsopoeodus + Pimephales, and Codoma + Cyprinella (BS < 70, PP 0.49), but we clearly show (based on topology and the AU test) that Cyprinella callistia should not be included in Cyprinella (Schönhuth & Mayden, 2010; Schönhuth et al., 2018). Cyprinella callistia was originally described as Photogenis callistius (Jordan 1877), but this genus does not apply to C. callistia. We did not include the type of the genus, Photogenis photogenis (Notropis photogenis), in our analysis, but N. photogenis has been resolved within the clade including the true Notropis, and was not closely related to C. callistia (Schönhuth et al., 2018). With no name available for the species, we refer to it as ‘Cyprinella’ callistia until such time that a broader analysis can be completed. This name will reflect that this species is clearly divergent from other Cyprinella, both morphologically (Mayden, 1989) and genetically (Schönhuth & Mayden, 2010; Schönhuth et al., 2018; this study).
We did not examine Notropis maculatus; however, Schönhuth et al. (2018) found it to be sister to Cyprinella. Notropis maculatus is found in muddy coastal streams, backwaters, and oxbows along the Gulf and Atlantic coasts. Like Cyprinella, it has instense pigmentation in nuptial males and has broad scales outlined in black. In Schönhuth et al. (2018), its position in the phylogeny was well supported only under Bayesian analysis, and further research is needed to determine its position, and we recognize it as ‘Notropis’ maculatus.
Alburnops and Hydrophlox
Gidmark & Simons (2014) resurrected Alburnops based on the monophyly recovered by Mayden et al. (2006). We do not recover monophyly of the Alburnops species here analyzed, and instead find primarily two non-sister clades. The type species, Alburnops blennius, is recovered in a clade with A. baileyi, A. chalybaeus, A. petersoni, A. potteri, A. texanus, and A. xaenocephalus (BS 100, PP 1), and thus these should retain the genus name. Schönhuth et al. (2018; Fig. 4) also recovered this well supported clade including these seven species plus A. asperifrons, A. braytoni, and A. simus pecosesis. Additionally, A. bairdi, A. buccula, A. candidus, A. edwardraneyi, A. girardi, A. hypsilepis, and A. shumardi should be included in Alburnops (Mayden, 1989; Mayden et al., 2006; Gidmark & Simons, 2014). Notropis aguirrepequenoi was described from out of A. braytoni and is included here in Alburnops (Miller, Minckley & Norris, 2005). Notropis orca is likely extinct, but similar to A. simus and is included in Alburnops (Chernoff & Miller, 1986; Mayden, 1989), but this will need to be confirmed.
The other clade (BS 100, PP 1) composed of species included within Alburnops by Gidmark & Simons (2014) includes A. chiliticus, A. chlorocephalus, A. chrosomus, A. lutipinnis, and A. rubricroceus, the five species recognized as Hydrophlox by Cashner, Piller & Bart (2011; type species Hybopsis rubricroceus Cope). This clade is here more closely related to species currently recognized under Notropis, ‘Notropis’, and Miniellus than to the Alburnops clade. Our results agreed with recent phylogenetic analyses by Schönhuth et al. (2018; Fig. 4) that also differentiated this clade of five species, and we recognize Hydrophlox as valid.
Miniellus + some ‘Notropis’
Miniellus is currently recognized as containing four species: Miniellus procne (type species), M. heterodon, M. stramineus, and M. topeka (Mayden et al., 2006; Gidmark & Simons, 2014). While our analyses did not include M. procne or M. topeka, M. stramineus was not resolved sister to M. heterodon. Instead, several ‘Notropis’ species were found to be more closely related to these two species. Five of these ‘Notropis’ were considered by Mayden et al. (2006) as belonging to a ‘Notropis’ longirostris clade. Schönhuth et al. (2018; Fig. 4) found strong support for a monophyletic Miniellus including the four initial species (Miniellus procne, M. heterodon, M. stramineus, and M. topeka) sister to ‘N.’ alborus (not analyzed here). Both studies also included in this clade ‘N.’ sabinae, ‘N.’ longirostris, ‘N.’ ammophilus, ‘N.’ chihuahua, ‘N.’ nubilus and ‘N.’ scabriceps with Schönhuth et al. (2018) also including ‘N.’ ortenburgeri, and ‘N.’ boops, and the present study also including ‘N.’ greenei and ‘N.’ melanostomus. However, while both studies analyzed ‘N.’ atrocaudalis, in Schönhuth et al. (2018) this species was collapsed with ‘N.’ scabriceps as sister to the rest of species in this clade; and in the present study ‘N.’ atrocaudalis was not resolved within this clade but in a clade with ‘N.’ bifrenatus and ‘N.’ heterolepis. Given strong support in the present study for the clade (BS 93, PP 0.82) that includes Miniellus, the ‘N.’ longirostris clade, and these other species of ‘Notropis’, we expand the genus Miniellus to include ‘Notropis’ greenei, ‘N.’ scabriceps, ‘N.’ sabinae, ‘N.’ longirostris, ‘N.’ ammophilus, ‘N.’ chihuahua, ‘N.’ melanostomus, and ‘N.’ nubilus, plus three species not included in the present analysis (‘N.’ alborus, ‘N.’ ortenburgeri, and ‘N.’ boops) that were also resolved within this clade (Schönhuth et al., 2018). Additionally, ‘N.’ mekistocholas and ‘N.’ rafinesquei, should be considered within this extended Miniellus, based on original descriptions (Snelson, 1971; Suttkus, 1991) and their strongly supported position as part of the ‘Notropis’ longirostris clade (Suttkus & Boschung, 1990; Mayden et al., 2006). ‘Notropis’ albizonatus and ‘N.’ uranoscopus are included in Miniellus per Warren, Burr & Grady (1994). Notropis perpallidus was found in Hollingsworth et al. (2013) to be sister to Miniellus sensu stricto and N. anogenus was sister to N. ortenburgeri, so both are included in Miniellus here. We note that further investigation needs to be done to resolve relationships within this clade.
‘Notropis’
Besides the species listed above that group with Miniellus, several other ‘Notropis’ are found throughout our phylogeny. ‘Notropis’ scepticus is found sister to Hudsonius in the concatenated analysis (BS 97) but sister to Ericymba + Luxilus in the species tree (PP 0.88). Sister relationship of this divergent species was also unresolved within the Shiner Clade in Schönhuth et al. (2018). The phylogenetic position of ‘N.’ scepticus varies in different studies, and despite the fact that we retain it under ‘Notropis’, the best solution nay be to describe a separate genus for this species.
We do not recover the ‘Notropis’ dorsalis group (Mayden, 1989; Raley, Wood & McEachran, 2001) as monophyletic. This group was composed of ‘Notropis’ dorsalis, ‘N.’ ammophilus, ‘N.’ longirostris, ‘N.’ rafinesquei, and ‘N.’ sabinae. Instead we find ‘N.’ ammophilus, ‘N.’ longirostris, and ‘N.’ sabinae grouping with Miniellus (see above), while ‘N.’ dorsalis was strongly supported as sister to Ericymba (BS 100, PP 1), as in Schönhuth et al. (2018). Currently, Ericymba is diagnosed by the presence of enlarged infraorbital canal scales (Pera & Armbruster, 2006), which are not found in ‘N.’ dorsalis; however, ‘N.’ dorsalis is otherwise very similar in morphology to the species of Ericymba, having a large mouth and ventrally flattened body. Given that ‘N.’ dorsalis is strongly resolved sister to Ericymba we are recognizing the species as Ericymba dorsalis; however, the species should be examined in greater detail to detemine if it requires a separate genus.
A clade of ‘Notropis’ heterolepis, ‘N.’ bifrenatus, and ‘N.’ atrocaudalis was strongly supported (BS 100, PP 0.99) with the clade sister to Ericymba + Luxilus in the concatenated analysis (BS 76) and Coccotis, Hybopsis, Opsopoeodus, Pimephales, ‘Cyprinella’ callistia, Codoma, Cyprinella, Hybognathus, Alburnops, Paranotropis, Hydrophlox, and Miniellus in the species tree (PP 0.70). These relationships differ signficantly with those of Schönhuth et al. (2018) where ‘N.’ heterolepis was resolved sister to ‘N.’ rupestris (a morphologically similar species not analyzed here) in a well supported and divergent clade with undefined relationships, ‘N.’ atrocaudalis was sister to the remaining extended Miniellus clade minus ‘N.’ scabriceps, and ‘N.’ bifrenatus was not examined. Recognizing the differences between the studies, we retain the following species as ‘Notropis’: ‘N.’ atrocaudalis, ‘N.’ bifrenatus, ‘N.’ heterolepis, and ‘N.’ rupestris.
The relationships of two small species from northeastern Mexico, ‘Notropis’ saladonis ‘N.’ tropicus have not been examined. ‘Notropis’ saladonis is currently considered to be extinct (Mercado Silva, 2019).
Notropis sensu stricto and Paranotropis
We find two clades that include species regarded as true Notropis (Mayden et al., 2006; Gidmark & Simons, 2014). The type species, Notropis atherinoides, is found in a clade (BS 100, PP 1) that is sister to Lythrurus (BS 90, PP 0.66) and contains N. amabilis, N. amoenus, N. jemezanus, N. micropteryx, and N. rubellus. Whithin this clade, Schönhuth et al. (2018) also included N. percobromus, N. photogenis, and N. stilbius and Hollingsworth et al. (2013) also included N. oxyrhynchus and N. suttkusi. Notropis should be limited to just these species. Schönhuth et al. (2018) weakly supported the clade of N. ariommus and N. telescopus to Notropis sensu stricto, and these relationships should be further studied, thus we refer to the species as ‘N.’ ariommus and ‘N.’ telescopus as well as the likely relative ‘N.’ semperasper (Coburn, 1982).
The other clade includes N. buchanani, N. wickliffi, N. volucellus, and N. spectrunculus (BS 100, PP 1), and here this clade forms a polytomy with the Hydrophlox and Miniellus clades in the concatenated analysis (BS 100, PP 1) and poorly supported as the sister to Hydrophlox in the species tree analysis (pp 0.49). Schönhuth et al. (2018) also included N. leuciodus, N. ozarcanus and N. shumardi, within this well supported clade. These species were either originally described as Notropis, or have been moved to Notropis from Alburnops, Hybognathus, or Hybopsis. Notropis leucidous is the type species of Paranotropis Fowler 1904, and its placement within the clade is well supported (Simons, Berendzen & Mayden, 2003; Schönhuth et al., 2018); thus, we propose these species be considered as Paranotropis. Also included is N. cahabae per the original description (Mayden & Kuhajda, 1989).
Central and southern Mexican Notropis
There were several species, mostly from Mexico, that were not examined in this study that were recognized within a clade called “Central and southern Mexican Notropis” (CSMN clade) in Schönhuth et al. (2018: 788). The CSMN clade contains species that have been placed into three genera: Aztecula, Graodus, and Yuriria. Of the species in the CSMN clade, we only examined G. moralesi.
Schönhuth et al. (2018) examined four species of Aztecula (listed as N. sallaei - type species, N. calientis, N. grandis, and N. marhabatiensis), three Graodus (listed as N. boucardi, N. imeldae, and N. moralesi), and one Yuriria (Y. alta). Aztecula was not monophyletic with A. sallaei sister to the species of Graodus; however, in previous studies (Schönhuth & Doadrio, 2003; Schönhuth, Doadrio & Mayden, 2006; Schönhuth et al., 2008; Hollingsworth et al., 2013), Aztecula and Graodus were both monophyletic.
Aztecula sallaei has a complex taxonomic history that includes many synonyms, movement between many genera, and a temporary change in spelling of its specific epithet to sallei (Chernoff & Miller, 1981). Aztecula additionally includes species that were described from out of N. calientis: N. amecae, N aulidion, N. calabazas, N. grandis, and N. marhabatiensis (Chernoff & Miller, 1986; Lyons & Mercado-Silva, 2004; Domínguez-Domínguez et al., 2009). The type species of Graodus is G. nigrotaeniatus Günther, 1868, which is believed to be a synonym of G. boucardi (Miller, Minckley & Norris, 2005). Graodus additionally includes N. cumingii, which is considered by some to be a senior synonym of G. imeldae (Gilbert, 1998; Miller, Minckley & Norris, 2005; Page et al., 2013) and an undescribed species in Oaxaca (referred as Notropis sp. 1 in Schönhuth, Doadrio & Mayden, 2006; Schönhuth et al., 2008). The species of Graodus and Aztecula (with the exception of A. sallaei) were considered to be in the genus Hybopsis (Miller, Minckley & Norris, 2005); however, they are not closely related to Hybopsis in any phylogenetic study. Yuriria was considered valid with the additional species Y. chaplalae (Miller, Minckley & Norris, 2005). A third species, Y. amatlana, has been described (Domínguez-Domínguez, Pompa-Domínguez & Doadrio, 2007). The species and genera of the CSMN clade will need further review and morphological examination, but given the strong support for these clades in previous studies (Schönhuth, Doadrio & Mayden, 2006; Schönhuth et al., 2008, 2018), we retain the taxonomy per Gidmark & Simons (2014) in three genera (Graodus, Aztecula and Yuriria) with some additional species placed as above.
Differences with Schönhuth et al. (2018)
Certain species/clades within the Shiner Clade have inconsistent phylogenetic placement between the present and Schönhuth et al. (2018) analyses, or were not resolved within any of these groups in either of these studies. These species include ‘Cyprinella’ callistia, ‘Notropis’ scepticus, ‘N.’ heterolepis + ‘N.’ rupestris, ‘N.’ bifrenatus, ‘N. ariomus’, ‘N.’ telescopus, ‘N.’ nazas, and ‘N.’ maculatus. In Schönhuth et al. (2018) Notropis nazas was strongly resolved sister to Hybognathus, and we suggest it to be considered as ‘Hybognathus’ until it can be further validated as a species of Hybognathus or as a separate genus. All these other taxa should be futher examined with genomic and morphological data with taxon sampling equal to or greater than that of Schönhuth et al. (2018); until that time descriptions of new genera or elevations of old genera for these species is premature.
Intraspecies utility of Arcila et al. (2017) markers
This study included two specimens of three species: Notropis atherinoides, Ericymba amplamala, and Pimephales vigilax. Notropis atherinoides specimens, one from Wisconsin and the other from Arkansas (both Mississippi River drainage), exhibited 99.6% sequence similarity with a pairwise distance of 0.003 and a total of 1,086 nucleotide differences across the entire 286,455 bp alignment. The specimens of E. amplamala were from Alabama (Mobile River Drainage) and Mississippi (Pascagoula River drainage), populations that were not found to be morphologically distinguishable in a detailed analysis (Pera & Armbruster, 2006), and had 99.5% sequence similarity, a pairwise distance of 0.005, and 1,424 differences. Our samples of Pimephales vigilax were collected from Paint Rock River (Tennessee River Drainage) and Uphapee Creek (Mobile River Drainage) and had 99.7% sequence similarity, a pairwise distance of 0.002, and 845 nucleotide differences. These results suggest two things: there may be cryptic diversity within shiner clade species, and the Arcila et al. (2017) markers are likely of utility at the population level, despite their initial development for use across a very broad taxonomic scale. Rincon-Sandoval, Betancur-R & Maldonado-Ocampo (2019) also found the markers useful for elucidating phylogeographic patterns within species in the neotropics and that they may have better utility than nuclear markers developed from a RADseq approach.
One of the targeted sequences was COI, a popular mitochondrial marker that is often used to delineate fish species and that can provide a comparison with the phylogenomic markers as a whole. We find a wide range of infraspecific differences in the 703 bp of the partial COI sequences examined. Notropis atherinoides, despite distant collection sites, has only a 2 bp difference (0.4% divergence). Pimephales vigilax from the neighboring Tennessee and Mobile River systems had a 16 bp difference (2.3% divergence). Ericymba amplamala, however, had a 54 bp difference (7.7% divergence), a degree of difference often associated with species-level differentiation, and there needs to be further investigation into the genetic structure of the species. COI alone may be suitable for identification of cryptic diversity for shiners, but the full phylogenomic dataset adds a considerable number of characters for elucidating population structure.
Conclusions
This study provides an important first step in using phylogenomics to resolve the relationships and taxonomy of the problematic leuciscid minnows. By employing a publicly available probe set (Arcila et al., 2017), future research can include more specimens that were not sampled in this study and easily be combined with our dataset. Our phylogenies help in understanding why this group has been difficult to resolve. A phylogenomic approach provides far more characters that can break the polytomies at the base of the shiner clade that are the likely result of rapid divergence. Not only has the group been described as morphologically conserved (Gidmark & Simons, 2014), thus hampering morphological interpretations of relationships, but we would argue that the same is true genetically. We find over 88% similarity (or uninformativeness) in a dataset comprised of over 288,000 base pairs. However, we find strong agreement between this study and the four gene phylogeny provided by Schönhuth et al. (2018), indicating that dense taxonomic sampling, as done in the latter study, is also a key in resolving closely related taxa. Problems with elucidating shiner relationships have been exacerbated by studies focusing only on subsets of the shiner clade due to sampling or cost restrictions. We demonstrate the utility of the exon capture method of Arcila et al. (2017) to elucidate relationships of rapidly evolving clades, and demonstrate that the markers may be of use at the population level as well. This is important as studies utilizing the Arcila et al. (2017) markers have the potential of resolving deep and shallow relationships within a single analaysis (Hughes et al., 2021). With the continuing decrease in cost of phylogenomic methods, the demonstrable utility of the Arcila et al. (2017) markers at many phylogenetic levels, and the soon to be large number of fish taxa sampled using the Arcila et al. (2017) markers, we would encourage researchers to add to this dataset. Numerous issues remain in the taxonomy and systematics of North American leuciscids, and we will continue to add species to the analysis. This study continues the trend at subtending the shiner clade into genera, but several important clades still need to be resolved and described.
Supplemental Information
Tissues used in this study.
Taxonomy follows Gidmark & Simons (2014). AUFT = Auburn University Fish Tissue Collection; UAIC = University of Alabama Ichthyological Collection; SLUM = St. Louis University Museum; SELU = Southeastern Louisiana University. Type species for each genus is indicated with an asterisk. Code number is from the alignment file. Basic localities provided with latitude and longitude when available.
Current taxonomy of species in the Shiner Clade alpahbetized by specific epithet and proposed revisions.
Notropis is limited to those species closely related to the type species, N. atherinoides, and is indicated in bold. Types of genera indicated by *.