
Stephanometra indica ( Smith, 1876 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.184295 |
|
DOI |
https://doi.org/10.5281/zenodo.6227271 |
|
persistent identifier |
https://treatment.plazi.org/id/03D4870E-FFE3-770B-FF6C-FB25BAA11C63 |
|
treatment provided by |
Plazi |
|
scientific name |
Stephanometra indica ( Smith, 1876 ) |
| status |
|
Stephanometra indica ( Smith, 1876) View in CoL
Comatula indica Smith, 1876:406 ; 1879:564, pl. 51, figs. 3, 3b [not 3a].
Antedon protectus Lütken, 1874:190 (nomen nudum, in Carpenter, 1879, p. 18–19).— Carpenter, 1879:19.
Antedon spicata Carpenter, 1881:190 .—AH Clark, 1912e:81.
Antedon indica: Carpenter, 1882:746 ; 1888:35, 54, 210, 225, 232, 233, 366, 379.—AH Clark, 1912d:34, 39, 40.
Antedon oxyacantha Hartlaub, 1890:178 ; 1891:6, 11, 15, 39, 55, 58, 113, pl. 3, figs. 35, 37.—AH Clark, 1912a:2.
Antedon monacantha Hartlaub, 1890:179 .
Antedon spinipinna Hartlaub, 1890:179 ; 1891:11, 39, 58, 61, 113, pl. 4, figs. 42, 44.—AH Clark, 1912d:37.
Antedon tuberculata Hartlaub, 1891:38 , 58, 59.—AH Clark, 1912c:385.
Antedon View in CoL ? spicata: Bell, 1894:396 .—AH Clark, 1912d:38.
Himerometra monocantha : AH Clark, 1907:356 (part).
Stephanometra spicata View in CoL : AH Clark, 1909a:10; 1918:94, 95, 272, pl. 7; 1941, 424–436, pl. 49, figs. 223, 224; pl. 91, fig. 447.— Gislén, 1936:3, 4, 5, 11.—AM Clark & Rowe, 1971:24.—AM Clark, 1972:108.— Chen et al., 1988:78, fig, 20. — Stevens, 1989:4 –23, 4–28, 5–8, fig. 5.3.— Messing, 1998:189, 191.— Kogo, 1998:61 –63, fig. 49.
Stephanometra tuberculata View in CoL : AH Clark, 1909a:10.— Hartmeyer, 1916:235.
Stephanometra monacantha View in CoL : AH Clark, 1909a:10 (part); 1912a:21.—HL Clark, 1921:22.
Stephanometra marginata View in CoL AH Clark, 1909c:169, 170.
Stephanometra indica View in CoL : AH Clark, 1909c:6, 8, 13, 26.— Gislén, 1934:20.—AM Clark & Rowe, 1971:23, fig. 8c —AM Clark, 1972:107 –108, fig. 10 (f–h).— Meyer & Macurda, 1980:86 –87.— Zmarzly, 1985:352 –353.— Stevens, 1989:4 –2, 4–22, pls. 8a, 8b, 5–8, fig. 5.3.— Messing, 1994:239; 1998:189 & 191.
Stephanometra oxyacantha View in CoL : AH Clark, 1911b:183; 1941:418–423, pl. 47, fig. 217, pl. 48, figs. 218–221.— Gislén, 1934:25.—AM Clark & Rowe, 1971:24.— Meyer & Macurda, 1980:87 –88, fig. 7c.— Stevens, 1989: 4 –22, 4–23, pl. 8c, fig. 5.3.— Messing, 1998:189,191.
Stephanometra spinipinna View in CoL : AH Clark, 1912d:37, 132.— Gislén, 1934:20.—AM Clark & Rowe, 1971:24.— Stevens, 1989:4 –28.— Messing, 1998:189.
Stephanometra stypacantha View in CoL HL Clark, 1915b:103.
Stephanometra indica protectus View in CoL : AH Clark, 1941:443–459, pl. 49, fig. 222, pl. 50, figs. 225–230, pl. 51, figs. 231, 232. Stephanometra indica indica View in CoL : AH Clark, 1936:88, 100; 1941:436–443, pl. 51, fig. 233, 234, pl. 91, figs. 449, 450. Stephanometra indica protecta View in CoL : AM Clark, 1975:401.
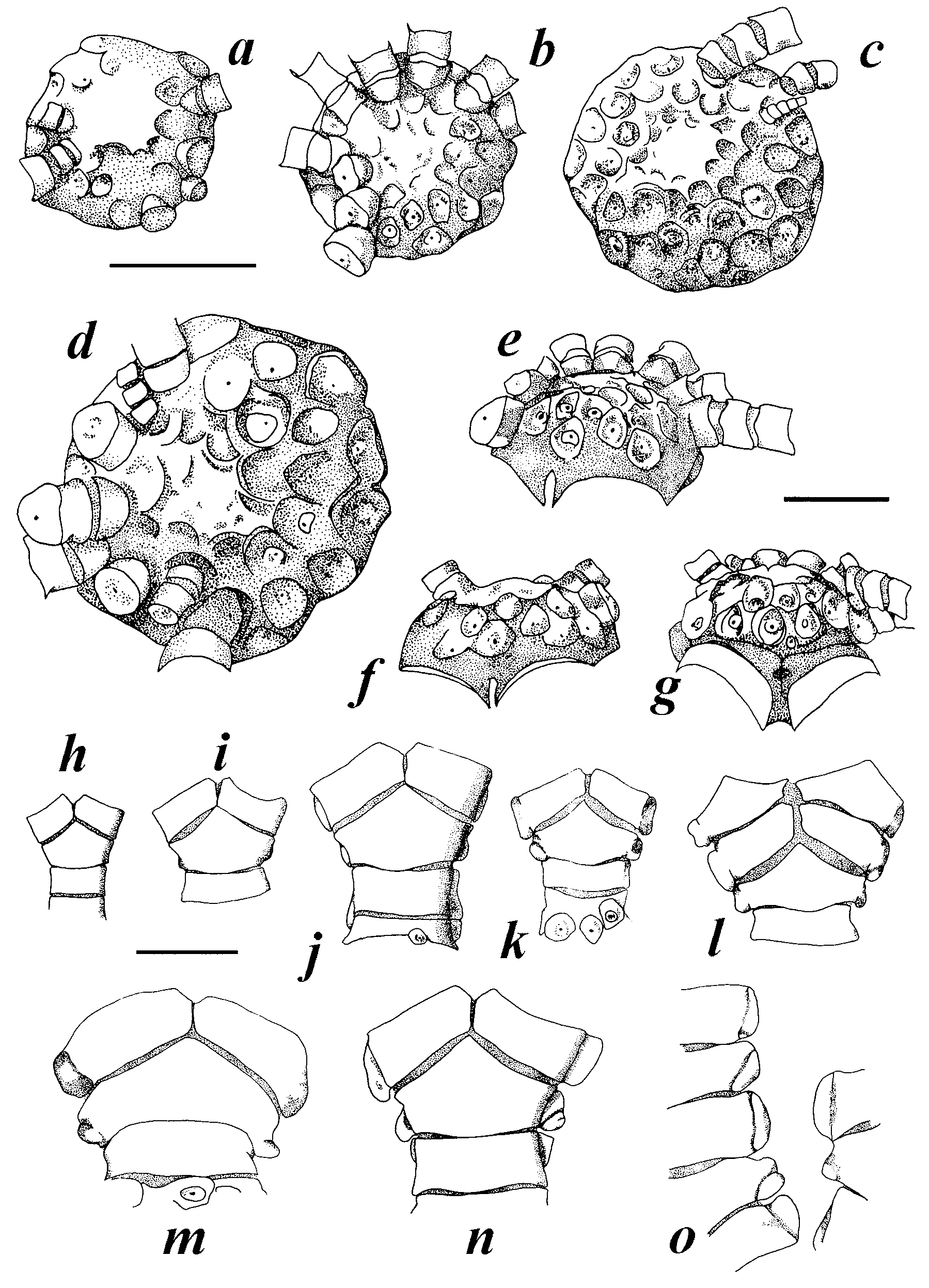
FIGURE 8. Stephanometra tenuipinna View in CoL . a–d. Cirri. a. USNM 1094104. b. NSUOC 256. c. FLMNH 6859. d. NSUOC 628. e. Distal cirral, USNM 1094104. f–i. Oral pinnules, NSUOC 312. f. P1. g. P2. h. P 3. i. P4. j–n. Oral pinnules, FLMNH 6864. j. P1. k. P2. l. P3. m. P4. n. P5. o–s. Oral pinnules, IRSNB/CRI 388. o. P1. p. P2. q. P3. r. P4. s. P5. Scale bars: upper (a– d), lower (f–s) all 2 mm; right (e) 1 mm.
Holotype. Comatula indica Smith, 1876 , NHM 76.5.5.24, Transit of Venus Expedition, 1874, Rodriguez I., Mauritius, Henry Slater, coll., no other data.
Other material examined. RED SEA: USNM E34613 View Materials (1 spec.), Gulf of Aqaba, DL Meyer coll., no additional data. SRI LANKA: USNM 35079 (1), off NE Corner, 0 8°51'30" N, 0 81°11'52"E, 51 m, no additional data. MAURITIUS: USNM E34699 View Materials (1), DL Meyer coll., no additional data. SEYCHELLES: USNM E34960 View Materials (2), Jun 1975, DL Meyer, coll. MALDIVE ISLANDS: NHM 1902.3.31.21 (1), Hulule, Male I., S Gardiner, coll.; NSUOC 633, 636 (2), Nilandu Atoll, E of Madali I., Jewellers' Is., 0 2°52'6"N, 72°50'8"E, 4.5–12 m, 21 Jan 1999, DL Rankin, coll. COCOS ISLANDS: USNM E11712 (1), 1941, CA Gibson-Hill, coll., no additional data. INDONESIA: USNM E35050 (2), NW coast Banda Besar, Banda Is., 2–3 m, 31 Jan 1975, DL Meyer, coll.; USNM E469 (2), Java I., Siboga sta.?, Mar 1899; USNM E34838 View Materials (2), E coast Marsegoe I., Ceram Sea, 0 2°59'48"S, 128°03'E, 0–15.2 m, DL Meyer, coll.; USNM E35376 (1), Rumphius II, Tapalol I., Biga Bay, Moluccas, 0 2°01'30"S, 130°19'18"E, 24 Jan 1975. SINGAPORE: USNM E34854 View Materials (4+), USNM E34549 View Materials (3), DL Meyer, coll., no additional data. MALAYSIA: NSUOC 313 (1), Dive Center, Mabul I., Sabah, Borneo, 0 4°15'N, 118°38'E, 7 m, 24 Apr 1997, CG Messing coll.; NSUOC 315 (1), E side Mabul I., 0 4°15'N, 118°38'E, 11 m, 22 Apr 1997, CG Messing, coll.; USNM 1102741, NSUOC 322 & 323 (3), E side Mabul I., 0 4°15'N, 118°38'E, 9–11 m, 21 Apr 1997, CG Messing, coll.; USNM E34546 View Materials (5), DL Meyer, coll., no additional data; USNM E34548 View Materials (3+), DL Meyer, coll., no additional data. PHILIPPINES: USNM E5269 (2), Willebrord Snellius Expedition, Bongao I., Tawi-Tawi Is., 27 m, 9 Sep 1929; NSUOC 314 (1), 316 (1), USNM 1102738 (4), N Tubbataha Reef, Sulu Sea, 0 9°49'N, 119°52'E, 8–21 m, 20–21 Apr 1995, CG Messing & L Sharron, colls.; USNM 1093368, 1094115, 1094116, NSUOC 331 (4), W end S Tubbataha Reef, 0 9°49'N, 119°52'E, 20–30 m, 21 Apr 1995, CG Messing, coll.; USNM 35221 (1), Albatross 5147, W of Lugus I., 0 5°41’40”N, 120°47’10”E, 38 m, 16 Feb 1908; NSUOC 318 (1), Pasig Reef, off Constance Shoal, 0 9°55'N, 119°30'E, 11–12 m, 23 Apr 1995, CG Messing, coll.; NSUOC 328 (1), Honda Bay, Palawan, 10°50'N, 118°45'E, 21 m, 21 Apr 1995, CG Messing, coll.; NSUOC 330, FLMNH 6862, USNM 1094114 (3), Jesse Beasley Reef, Sulu Sea, 0 9°01'N, 119°48'E, 4–8 m, 19 Apr 1995, CG Messing, coll.; USNM 35262 (1), Albatross 5109, Luzon I., 14°03'45"N, 120°16'30"E, 18–22 m, 15 Jan 1908. SOUTH CHINA SEA: NHM 1892.8.22.28 (1), Macclesfield Bank, 24 m, British Admiralty, coll. CHUUK ATOLL: USNM 1094092 (1), SE of Scheiben I., NW of Weno I., 9.1 m, 10 Jun 1993, CG Messing, coll.; USNM 1094089, NSUOC 333, 335 & 339 (4), N side NE Pass, S of Quoi I., 0 7°31'38"N, 151°58'05"E, 9–11 m, 8–11 Jun 1993, CG Messing, coll.; USNM 1094093, 1094094, LACM 1993–209.1 (3), Fringing reef, E side Yanagi I., 0 7°25'N, 151°50'E, 14–18 m, 13 Jun 1993, CG Messing, coll. KIRIBATI: USNM E18323 View Materials (1), PE Cloud sta. GOC-25, Onotoa Atoll, no additional data. PAPUA NEW GUINEA: NSUOC 321, LACM 1992–218.1, FLMNH 6822, USNM 1102761 (4), Jais Aben Reef, N side Nagada Harbor, Madang, 0 5°09'29"S, 145°49'21''E, 3–4 m, 2 Jun 1992, CG Messing, coll.; USNM 1102737, NSUOC 332 (2), Padoz Reef, Madang, 0 5°09'S, 145°50'E, <18.5 m, 5 Jul 1991, CG Messing, coll.; USNM 1094083 (1), Barrier I. outside Magic Pass, Madang, 6–8 m, 9 Jul 1991, L Harris, coll.; USNM 1094085 (1), Barrier I. outside Wongat I., Madang, 0 5°08'09"S, 145°50'51"E, 3–4.5 m, 11 Jun 1991, CG Messing, coll.; IRSNB / CRI 390–393 (4), Epave (Sisimangun), 5–18 m, 20–23 Jun 1989, MC Lahaye, coll.; IRSNB / CRI 394 (1), Laing I., Hansa Bay, sta. 13, 26 m, 26 Jun 1989, MC Lahaye, coll.; IRSNB / CRI 395, 396 (2), autour de l'Ile, Laing I., Hansa Bay, 0–5 m, 6 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 397 (1), Epave, 36 m, 7 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 398 (1), Platier, 12 m, 9 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 399–401 (3), Pointe Sud, Laing I., Hansa Bay, 12 m, 10 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 402 (2), Hansa Point, Hansa Bay, 36 m, 12 Jul 1989, MC Lahaye coll.; IRSNB / CRI 403 (1), Laing I., Hansa Bay, H1-2/12 (S/ O), 26 m, 15 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 404 (1), Hansa Bay, MC Lahaye, coll., no additional data.; IRSNB / CRI 405, 406 (2), Pointe Nord, Laing I., Hansa Bay, 23 m, 19 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 407, 408 (2), Pointe O, Laing I., Hansa Bay, 25 m, 22 Jul 1989, MC Lahaye, coll.; IRSNB / CRI 409 (1), Mandy Passage, near Hansa Bay, 41 m, 24 Jul 1989, MC Lahaye, coll. LOUISIADES ARCHIPELAGO: (1) Panaete I., Deboyne Group, 10°40'11"S, 152°21'08"E, 10 m, 31 May 1998, G Paulay, coll. AUSTRALIA: USNM E34738 View Materials (1), Heron I., DL Meyer, coll., no additional data. FIJI: USNM E34572 View Materials (2), 1976, DL Meyer, coll., no additional data; USNM E34793 View Materials (5), Suva Point, 12 Apr 1975, DL Meyer, coll. NO LOCALITY DATA: CRRF 1656K (1), no additional data.
Diagnosis. A species of Stephanometra with brachitaxis ossicles weakly swollen laterally or with thick rounded ridge-like extensions oriented obliquely to longitudinal axis of ray. Middle and distal cirrals bearing a midaboral carination, which may develop into a small spine distally. P1 slender, tapering delicately to a conical tip; P2 alone or P2 and following 1–3 pinnules composed of elongated pinnulars with reduced ambulacral groove, a conical terminal segment, and flattened articulations lacking triangular fossae.
Description. Centrodorsal discoidal or dome-shaped, 2.1–6.3 mm across, 0.7–2.4 mm high; DH 2.0–4.0. Cirri in two to four alternating marginal rows. Polar area concave, irregular in shape, with encroaching cirri, 0.7–3.1 mm across ( Fig. 10 View FIGURE 10 a–g); DP 1.7–3.9. Cirri XIII–LI, 15–30, 10– 31 mm long; c1-3 short; c4 through c9–11 longer than broad; c7-9 longest, LW 1.0–1.9; c10–12 through penultimate cirral compressed, each with an aboral keel that may be weak or develop into a small spine (Figs. 11a–e). Penultimate cirral shorter than terminal claw and with a sharp aboral spine.
Basals not visible externally; radials projecting slightly beyond edge of centrodorsal or not visible. Brachitaxis ossicles well separated, by 0.4–2.0 mm between adjacent Ibr1. Ibr1 oblong, with parallel or converging lateral margins, usually free laterally but sometimes united proximally; L to 1.2 mm, W to 3.1 mm, WL 2.0– 5.0, rarely less. Ibr2 (axil) with diverging lateral margins, bearing a weakly ( Figs. 10 View FIGURE 10 h–j) to strongly developed (Figs. 3b, 10k–o) obliquely-oriented (relative to ray axis) proximolateral elongated knob; L to 2.9 mm, W to 4.1 mm, WL 1.7–3.7, rarely less. IIBr2 series similar to IBr2. IIbr1 bearing weak to strongly developed obliquely-oriented process along entire length of lateral margin; IIbr2 (axil) with short proximolateral knob (Figs. 3b, 10k–o). IIIBr2 developed externally and, rarely, internally; IVBr2 external when present. Synarthrial tubercles weak to well developed.
Arms 11–33; R 50–160 mm. Rays most often with 3–6 arms each, less often with 2 or 7. Br1 through br6 – br8 oblong; subsequent brachials cuneate; br10 with W to 1.9 mm, L to 1.1 mm, WL 0.7–3.6. Syzygies at br3+4, br9+10, br16+ 17 and br19+20, less often at br5+6, br11+12 or br 25+26. Distal syzygial intervals 4–8, less often 3, 9 or 10.
Pinnules on outer arms generally longer and thicker than on inner arms. Specimens with P1 and P2 apparently stiffened formerly distinguished as S. spinipinnna (Figs. 11g, k); with P2 through P4 or P5 enlarged formerly distinguished as S. oxyacantha (Figs. 3c, 11j); with P2 and P3 enlarged as S. spicata (Fig. 11h), and with P2 alone enlarged as S. indica (Figs. 11f, i, l).
P1 3.6–14.1 mm long, of 10–27 pinnulars, slender, composed of elongated segments tapering to a conical tip. First few proximal pinnulars laterally compressed; subsequent pinnulars cylindrical. First and second pinnulars oblong, third slightly longer than broad; fourth and following with LW 2.0–4.0, and terminal pinnular a conical spike. P2 alone, or P2 through P3, P4 or P5, robust, stiff and spine-like, much larger than subsequent pinnules, composed of elongated cylindrical segments with both ambulacral groove and tube feet reduced.
P2 5.4–20.0 mm long, 8–18 pinnulars; pinnular 6 LW 1.5–4.0. In smaller specimens, P3-P5 tending to be small weak and flexible with well-developed ambulacral groove and resembling following pinnules (Fig. 11f, l); P3 2.1–6.0 mm long, 7–14 pinnulars; P4 2.0– 4.8 mm long, 7–14 pinnulars, and P5 2.4–5.7 mm, 10–14 pinnulars. In larger specimens, robust P3 is 3.4–15.0 mm, of 7–14 pinnulars; P4 2.3–10.2 mm, 9–14 pinnulars; P5 2.4–9.4 mm, 8–14 pinnulars (Figs. 11g, h, j, k). Subsequent pinnules gradually increasing in length. P distal 6.3– 9.4 mm long, 18–23 pinnulars; terminal pinnulars elongated and covered with small spines (Fig. 11m). A few large specimens have only P2 enlarged (Fig. 11i).
Color patterns. Widely variable. Rays usually concentrically banded with various combinations of white, tan, brown, gray, pink and orange (Fig. 3b). Other variations include mostly white or pale tan with a few brown bands; entirely dark purplish (Fig. 3c), sometimes with darker articulations; or chiefly yellow, orange or purple proximally, with purple, brown or black articulations. A few specimens are speckled, chiefly purple with tiny pale spots. Pinnules with banding continued from arms, or beaded brown and white; some with silver, yellow or orange pinnule tips (Fig. 3c). Meyer & Macurda (1980) also reported bands of red-brown, purple and yellow, a white variegated pattern, and pink or lavender with a white section midway along arms and pinnules. They noted black or brown brachial articulations consistently in S. oxyacantha but absent from S. indica . However, some typical S. indica examined here have brown or dark orange (Figs. 3a–b) articulations, and patterns of previously recognized species ( S. indica , S. spicata and S. oxyacantha ) overlap. Previous descriptions (AH Clark 1921b; Meyer & Macurda 1980; Zmarzly 1985; Stevens 1989) may be based on incorrectly identified specimens.
FIGURE 11. Stephanometra indica . a–d. Cirri. a. Paulay specimen (Panaete I.). b. USNM 1094116. c. NSUOC 333. d. USNM 1102738. e. Carinate distal cirral, USNM 1094116. f–l. Oral pinnules. f. USNM 1102738, P1-P4 ( S. indica ). g. Paulay specimen, P1-P4 ( S. spinipinna ). h. FLMNH 6822, P1-P4 ( S. spicata ). i. NSUOC 335, P1-P3 ( S. indica ). j. IRSNB/ CRI 396, P1-P4 ( S. oxyacantha ). k. NSUOC 314, P1-P5 ( S. spinipinna ). l. NSUOC 333, P1-P3 ( S. indica ). m. NSUOC 314, terminal pinnulars of distal pinnule. Scale bars: top right (a), top left (b–d), middle right (f–h, l), lower left (i, j), lower right (k), all 2 mm; middle left (e) 1 mm; bottom right (m) 0.5 mm.
FIGURE 13. Stephanometra indica with pinnules intermediate between previously recognized species. a–b. NSUOC 330, “ indica / spicata ”. a. “ indica ” pinnules, P2-P3. b. “ spicata ” pinnules, P2-P4. c-d. Uncatalogued “ spicata / oxyacantha ” specimen. c. “ spicata ” pinnules, P2-P4. d. “ oxyacantha ” pinnules, P2-P4. Scale bar: 2 mm.
Distribution. On reefs from the Red Sea to Madagascar in the west, to Guam and the Tonga Islands in the east, including tropical Australia as far south as the Capricorn Channel, Queensland, and as far north as Okinawa, Japan. Previous records, possibly as far east as Kwajalein (AH Clark 1921b; Meyer & Macurda 1980; Zmarzly 1985; Stevens 1989) may be based on incorrectly identified specimens. Depth range: shoreline possibly to 73 m (AH Clark 1941); most specimens have been collected in <30 m.
Ecology.—Cryptic during the day under rubble or slabs, or within the reef infrastructure. At night, perched on prominent coral heads, coral rock projections, and, occasionally, on massive soft corals. The typical nocturnal posture is a biplanar arcuate fan. However, individuals also form flat or shallow bowl-like radial fans with arms spread across the surface of their coral perch. Meyer & Macurda (1980) reported some individuals attributed to S. oxyacantha at Palau as being day-active, but restricted to undersides of ledges and in reef recesses where they form pendant arcuate fans, sometimes with arms not aligned as fans. They found one specimen fully expanded on an alcyonacean, but under reduced light conditions at the inner entrance of a tunnel to a marine lake. At Heron and Wistari Reefs, southern Great Barrier Reef, Australia, Stevens (1989) found one specimen each of S. spicata and S. oxyacantha apparently exposed during the day but in> 10 m depth.
FIGURE 14. Stephanometra indica . Plot of centrodorsal diameter against aboral pole diameter (a) and number of cirri (b). c. Plot of Ibr2 width against length of pinnular 6 of P2. Symbols as in Figure 12 View FIGURE 12 .
FIGURE 15. Stephanometra indica (including all forms) versus S. tenuipinna . a. Plot of ray length against number of cirri. b. Plot of Ibr2 width against length of pinnular 6 of P2.
Remarks. Nominal members of the second Stephanometra species group— S. spinipinna , S. indica , S. spicata and S. oxyacantha —lack strong aboral cirral spines and have been differentiated by features of their oral pinnules (AH Clark 1941). All have at least P2 stiff and spine-like. As noted in the diagnosis and description above, such pinnules are better characterized as having flattened articular facets (Fig. 2).
The latter three species of this second group have P1 flexible and slender and have been distinguished by the number of enlarged oral pinnules that follow (AH Clark 1941). Comments ( Gislén 1940; AH Clark 1941; AM Clark & Rowe 1971; AM Clark 1972) on the possibility that some or all three might represent a growth series were mentioned above. Meyer & Macurda (1980) maintained the species as distinct but reported some conflicting data. Their specimens with only P2 stiff and spine-like were smaller (maximum R= 80 mm) than those with more stiff pinnules, and they recorded one S. indica with some P3 “somewhat enlarged and spikelike”, approaching S. spicata (p. 86). However, they found S. indica and S. oxyacantha only at separate sites and noted distinct but undescribed differences in their cirri.
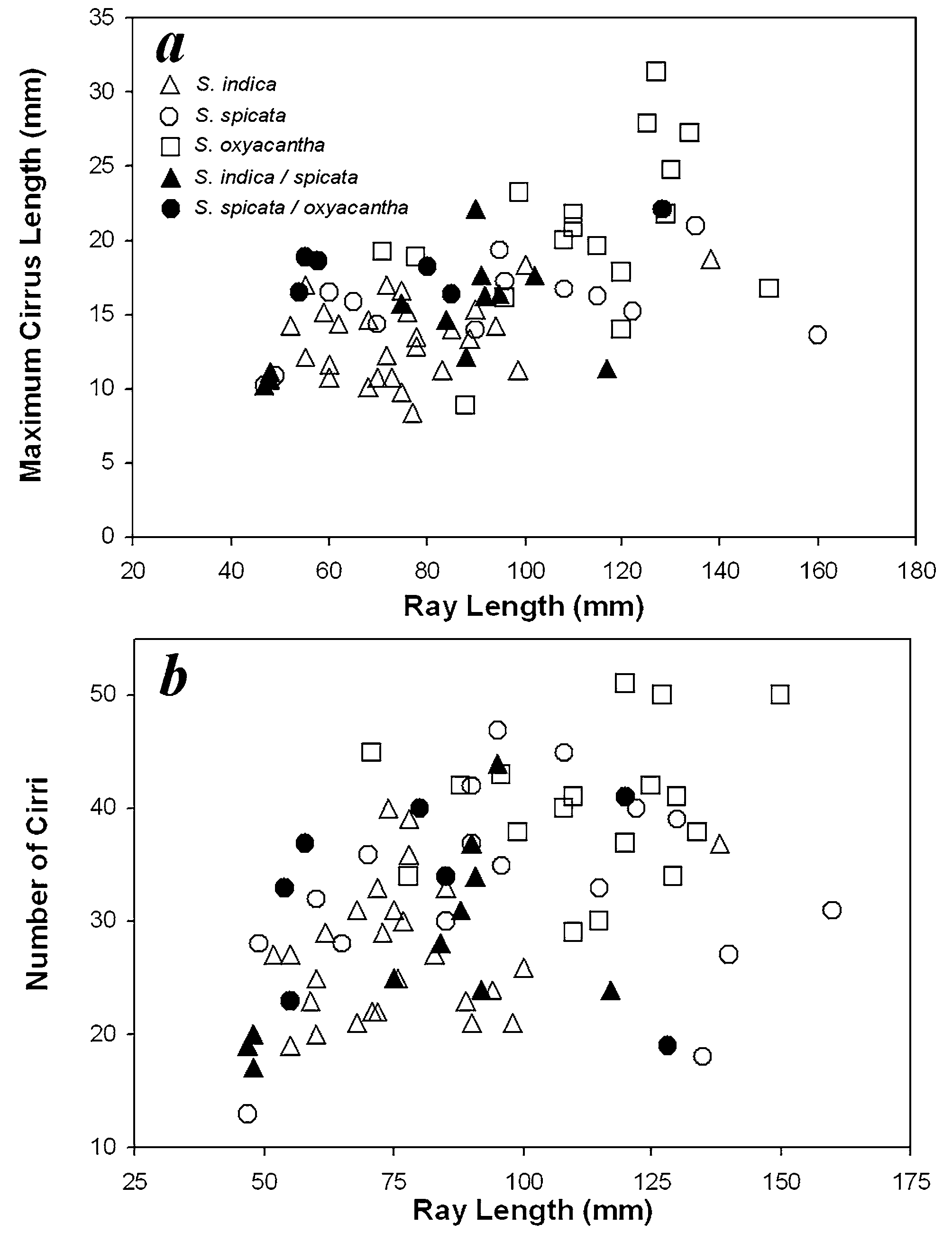
Stevens (1989) suggested that these three species represented points along continua of size, arm number, form of proximal pinnules, nocturnalism and depth, and could be considered a monospecific complex. Yet, he noted distinct color patterns, that overlap between characters was small, and that they should be kept distinct. However, he considered only a small portion of the range of these taxa and measured only two characters: ray and cirrus lengths. The current study indicates that a great deal of overlap occurs between these two characters for the indica-spicata-oxyacantha series ( Fig. 12 View FIGURE 12 ).
We recognize an apparent size continuum for the indica-spicata-oxyacantha series based on several characters. Although ray lengths of specimens examined overlap strongly— S. indica 50–140 mm (mean 77 mm), S. spicata 50–160 mm (mean 114 mm), and S. oxyacantha 55–150 mm (mean 109 mm)—only one large S. indica (NSUOC 335) has a ray length approaching that of S. oxyacantha , while there are no small specimens of the latter. The holotype of Comatula indica (NHM 76.5.5.24) is also a large specimen, with the third largest P 2 in the current study. Plots of maximum cirrus length and number of cirri against ray length ( Fig. 12 View FIGURE 12 ) show that the three overlap but that S. oxyacantha generally falls out as the largest, with S. spicata in the middle and S. indica smallest.
Approximately 20% of specimens referable to the indica-spicata-oxyacantha series in the current study are intermediates that cannot be satisfactorily assigned to species based on previous diagnoses; they either bear oral pinnules characteristic of both S. indica and S. spicata , or of both S. spicata and S. oxyacantha (Fig. 13). The former account for 18% of all S. indica , S. spicata and S. indica / spicata specimens combined, and the latter 13% of all S. spicata , S. oxyacantha and S. spicata / oxyacantha combined. Also, oral pinnules in some S. spicata and S. indica specimens exhibit traces of basal stiffening, suggesting that pinnules stiffen with age. However, the regenerating arms so far examined resemble existing arms, e.g., S. spicata specimens regenerate S. spicata arms. We have not yet found, for example, S. indica specimens with regenerating arms completely characteristic of S. spicata .
Characters that vary with overall size include centrodorsal diameter and height, aboral pole diameter, number and length of cirri, width of Ibr2, ray length, development of lateral adambulacral margins, and pinnule length. Lateral adambulacral margins are most strongly developed in S. oxyacantha , and range from weak to strong in both S. indica and S. spicata ( Figure 10 View FIGURE 10 h-o). Such variations likely reflect growth, though Messing (1994) reported that species of several genera in the comatulid family Comasteridae tended to have shorter more numerous arms in habitats subject to high hydrodynamic energy. Some ecophenotypic response or environmental partitioning of the different nominal species may exist in Stephanometra . Messing (unpublished) found numerous similarly colored S. indica at one site in Chuuk Lagoon and a single S. spicata only at another site. However, such disjunct distributions may be a function of patchiness or collecting effort. Meyer & Macurda (1980) found S. indica and S. oxyacantha at separate sites at Palau, while Messing (2007) found all three forms together there, as well as in the Philippines and Papua New Guinea (Messing, unpublished).
Bivariate plots of various characters (Fig. 14) indicate that S. oxyacantha tends to be largest and S. indica smallest, with S. indica / spicata and S. spicata / oxyacantha intermediates scattered throughout. Additional plots (not shown) of centrodorsal height against diameter and against Ibr2 width, and centrodorsal diameter against maximum cirrus length and length of pinnular 6 of P2 all show similar patterns. In all cases, too much overlap exists to distinguish the three; the evidence strongly suggests that S. indica , S. spicata , and S. oxyacantha represent a continuum of form within a single taxon.
Stephanometra spinipinna View in CoL supposedly differs from the other three in having a stiff and spine-like P1 not enlarged like the following pinnules. Hartlaub (1890: 62–63) noted that it was a small very delicately built form (“eine sehr zierlich gebaute kleine Form”), with P1 thin, but rather rigid and styliform (“dünn, aber ziemlich steif und griffelförmig) and intermediate between S. tenuipinna View in CoL on one hand and S. spicata View in CoL and S. oxyacantha View in CoL on the other. He also described it as not sexually mature. Although AH Clark (1941) used Hartlaub’s (1890) description of the type specimen, he referred to other specimens as having P1 “slender but very stiff” (p. 416) and “somewhat stiffened but not spinelike” (p. 417). AH Clark (1941) also described a large specimen (USNM E8969) but did not mention its pinnules. Re-examination of this specimen shows P1 as stiff and robust as P2, approaching P1 of S. tenuipinna View in CoL , although the cirri are typical of the S. indica group.
Stevens (1989) recorded specimens of S. indica bearing partially stiffened P1 on one or more arms. Some specimens in the current study attributable to all nominal species of the second group also bear a slender but stiffened P1. These pinnules point toward the mouth and cover the disk, which contribute to their stiffened appearance. However, their articulations are identical to those of the slender flexible P1 typical of the indica - spicata View in CoL - oxyacantha View in CoL series. Variations in apparent stiffness may at least partly result from preservational differences.
In addition, although AH Clark (1941) described S. spinipinna as easily recognized when typically developed, he acknowledged that P1 varied and suggested “possible intergradation between S. spinipinna on the one hand and S. spicata and S. oxyacantha on the other" (p. 408). Similarly, the centrodorsal, cirri, and lateral adambulacral margins are identical to those of the indica-spicata-oxyacantha series. In bivariate plots of the same characters as described above ( Fig. 12 View FIGURE 12 ), the “typical” S. spinipinna specimens all fall within the morphospace of the indica-spicata-oxyacantha series (graph not shown). We therefore treat the species as a synonym of S. indica .
AH Clark (1941) distinguished S. indica protectus as a subspecies of S. indica based only on its shorter P2 with 9–16 segments and a sharp tip versus 16–20 segments tapering to a fine point in S. i. indica . As with S. spinipinna , although he wrote that the two were very different when “typically developed” (p. 408), he also noted that some specimens possessed a P2 intermediate between the two. Eight percent of specimens examined in this study possess a P2 similar to AH Clark’s description of S. indica protectus . As this represents a negligible, variable feature among specimens that do not differ in any other recognizable way and have practically co-extensive geographic ranges, we follow AM Clark (1972) and treat S. indica protectus as a synonym of S. indica .
Meyer & Macurda (1980), who maintained S. indica , S. spicata and S. oxyacantha as distinct, noted that the cirri of S. indica were “distinctly different” from those of S. oxyacantha and resembled Figure 340 in AH Clark (1915:287). However, they did not describe the difference between the two. Cirri from specimens of indica , spicata , oxyacantha and intermediates in the current study resemble Clark’s illustration and do not differ except in size-related features. We treat both S. spicata and S. oxyacantha as junior synonyms of S. indica .
Bivariate plots show both overlap and separation of characters for S. tenuipinna versus all forms of S. indica . Distributions of ray length plotted against number of cirri overlap (Fig. 15a), but S. indica generally develops more cirri at a smaller ray length than S. tenuipinna . The same relationship holds for centrodorsal diameter versus number of cirri (not shown). On the other hand, the length of pinnular 6 of P2 is always greater in S. tenuipinna regardless of centrodorsal diameter (Fig. 15b) or Ibr2 width (not shown). Other characters that distinguish S. tenuipinna from S. indica are descriptive.
Note. Re-examination of the holotype of Comatula indica Smith (NHM 76.5.5.24) confirmed that the cirrus “found among debris at the bottom of the bottle containing the specimen” (AH Clark 1941:439) does not belong to Stephanometra .
| USNM |
Smithsonian Institution, National Museum of Natural History |
| SRI |
Serengetti Research Institute |
| FLMNH |
Florida Museum of Natural History |
| LACM |
Natural History Museum of Los Angeles County |
| NEW |
University of Newcastle |
| IRSNB |
Institut Royal des Sciences Naturelles de Belgique |
| CRI |
Universidade do Extremo Sul Catarinense, Bairro Universitário |
| FIJI |
University of the South Pacific |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Stephanometra indica ( Smith, 1876 )
| Rankin, Dana L. & Messing, Charles G. 2008 |
Stephanometra stypacantha
| Clark 1915: 103 |
Stephanometra spinipinna
| Messing 1998: 189 |
| Stevens 1989: 4 |
| Clark 1971: 24 |
| Gislen 1934: 20 |
| Clark 1912: 37 |
Stephanometra oxyacantha
| Stevens 1989: 4 |
| Meyer 1980: 87 |
| Clark 1971: 24 |
| Gislen 1934: 25 |
| Clark 1911: 183 |
Stephanometra spicata
| Messing 1998: 189 |
| Kogo 1998: 61 |
| Stevens 1989: 4 |
| Chen 1988: 78 |
| Clark 1972: 108 |
| Clark 1971: 24 |
| Gislen 1936: 3 |
| Clark 1909: 10 |
Stephanometra tuberculata
| Hartmeyer 1916: 235 |
| Clark 1909: 10 |
Stephanometra monacantha
| Clark 1909: 10 |
Stephanometra marginata
| Clark 1909: 169 |
Stephanometra indica
| Messing 1994: 239 |
| Stevens 1989: 4 |
| Zmarzly 1985: 352 |
| Meyer 1980: 86 |
| Clark 1972: 107 |
| Clark 1971: 23 |
| Gislen 1934: 20 |
| Clark 1909: 6 |
Himerometra monocantha
| Clark 1907: 356 |
Antedon
| Clark 1912: 38 |
| Bell 1894: 396 |
Antedon tuberculata
| Clark 1912: 385 |
| Hartlaub 1891: 38 |
Antedon oxyacantha
| Clark 1912: 2 |
| Hartlaub 1890: 178 |
Antedon monacantha
| Hartlaub 1890: 179 |
Antedon spinipinna
| Clark 1912: 37 |
| Hartlaub 1890: 179 |
Antedon indica:
| Clark 1912: 34 |
| Carpenter 1882: 746 |
Antedon spicata
| Clark 1912: 81 |
| Carpenter 1881: 190 |
Antedon protectus Lütken, 1874:190
| Carpenter 1879: 19 |
Comatula indica
| Smith 1876: 406 |