
Zygothuria thomsoni (Théel, 1886)
|
publication ID |
https://doi.org/ 10.1080/00222933.2011.638423 |
|
persistent identifier |
https://treatment.plazi.org/id/6931320B-FFBD-FF98-FE43-134FF03FFAE9 |
|
treatment provided by |
Felipe |
|
scientific name |
Zygothuria thomsoni (Théel, 1886) |
| status |
|
Zygothuria thomsoni (Théel, 1886)
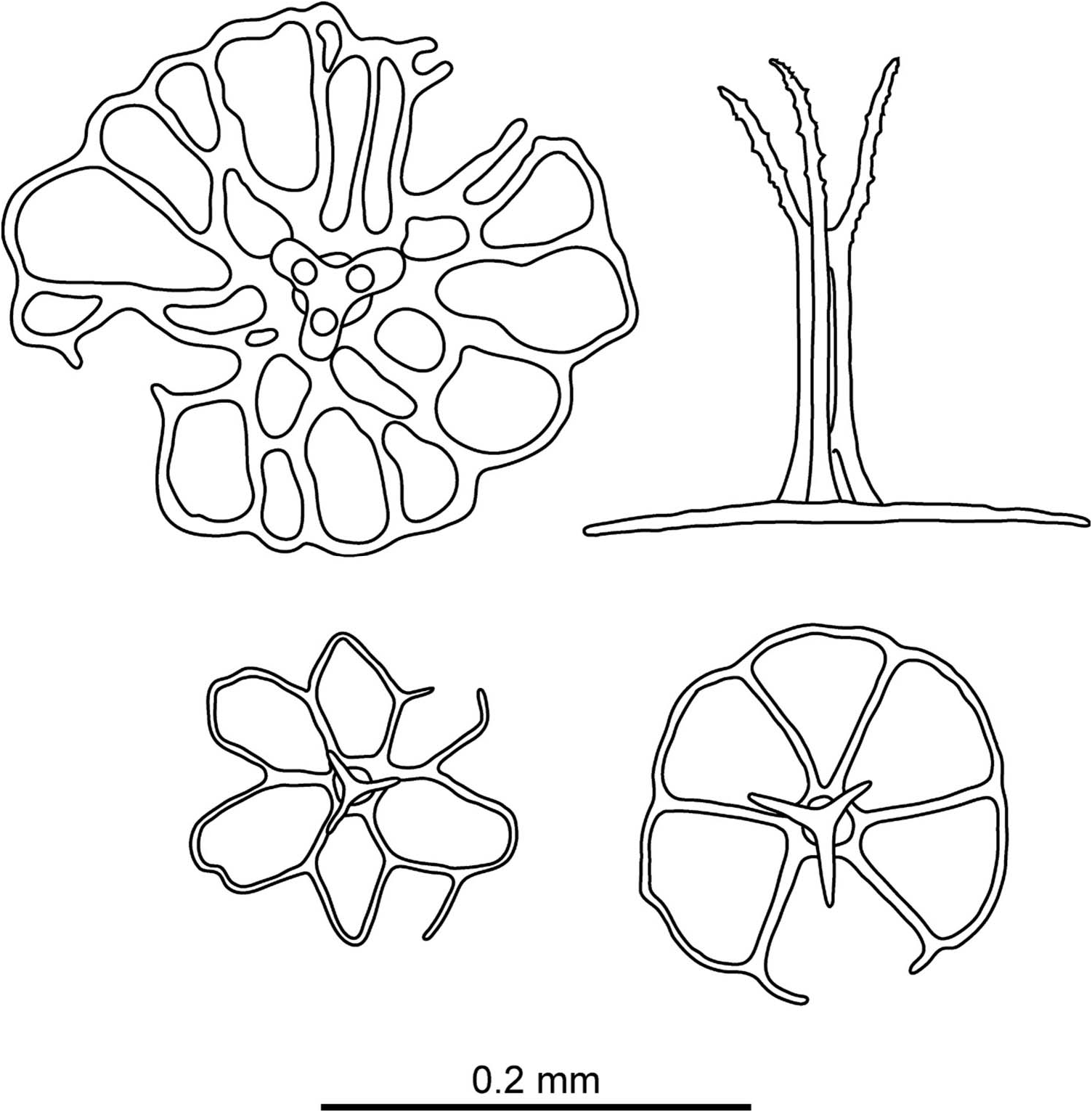
( Figure 20 View Figure 20 )
Holothuria thomsoni – Théel, 1886a: 184–185, pl. 10, figs. 8, 11.
Holothuria thomsoni var. hyalina – Théel, 1886a: 185.
Mesothuria thomsoni (Théel, 1886) – Östergren, 1896: 350–351.
Material examined
See Table 15.
Type material
Syntypes, NHM (labelled as co-types), NHM 1886.10.2.182, HMS Challenger, St. 237, 17 June 1875, 34 ◦ 37 ′ N, 140 ◦ 32 ′ E, 3375 m, single incomplete specimen; NHM 1886.10.2.183 and NHM 1886.10.2.184, HMS Challenger, 28 June 1875, St. 244, 35 ◦ 22 ′ N, 169 ◦ 53 ′ E, 5307 m, three specimens.
Type locality
North-west and mid-north Pacific, 3375–5307 m.
Diagnosis
Body oval. Mouth ventral. Anus terminal. Twelve tentacles. Simple alternating row of pedicels along each side of ventral surface, and some small papillae scattered on
) ventral surface in neighbourhood of these rows. No other ambulacral appendages visible. Calcareous ring narrow, fragile, without posterior processes. Three Polian vesicles.
Ossicles: body wall unusually rough, caused by abundance of tables with large, irregularly perforated disks with central holes usually smaller than peripheral ones; stem terminates in three arms, shorter than stem, arms slender and spinous ( Figure 20 View Figure 20 ).
Remarks
See remarks for Z. candelabri .
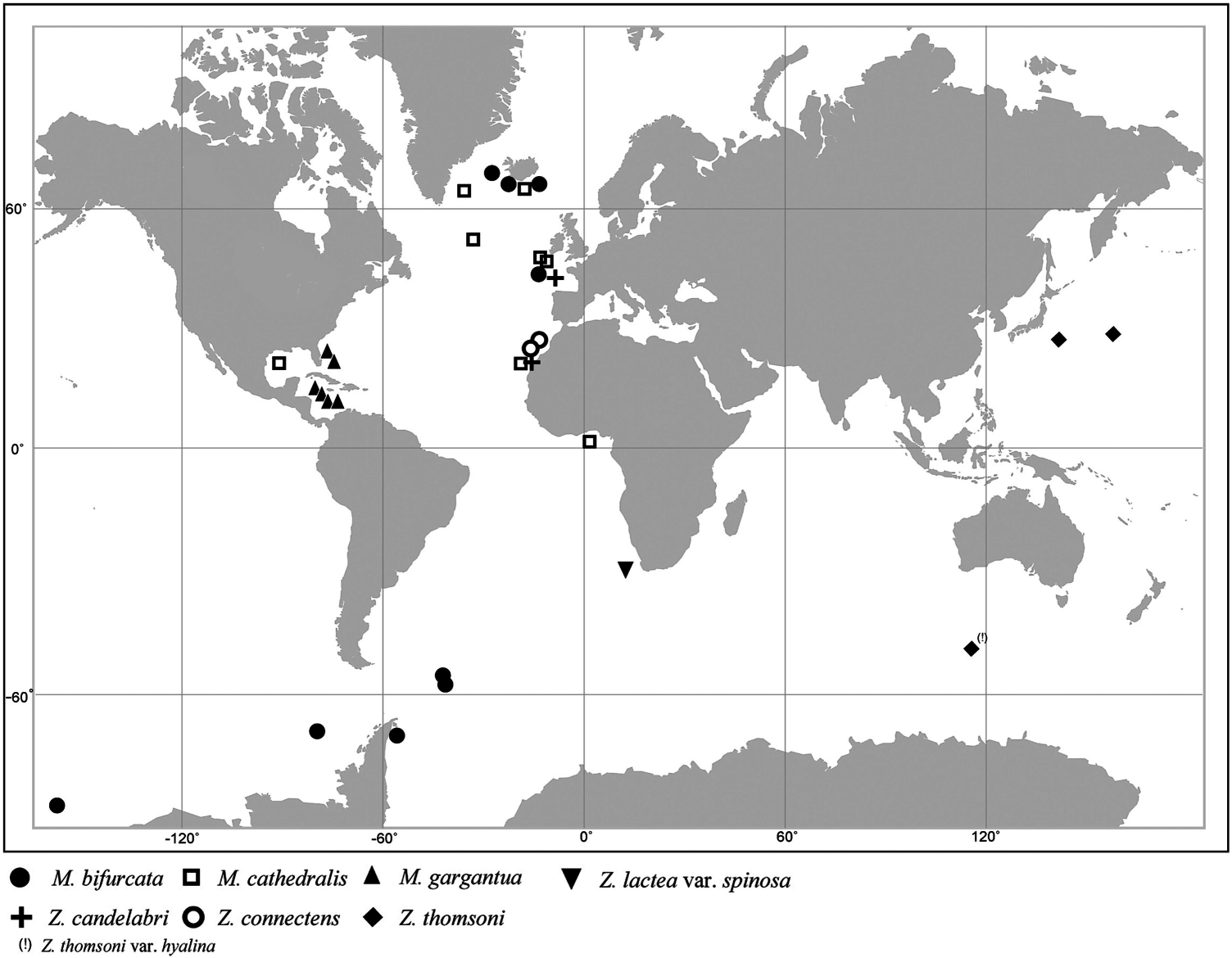
Distribution
Only known from type locality ( Figure 6 View Figure 6 ).
Bathymetric range
Depth 3431–5307 m.
Population structure and biomass of Mesothuria and Zygothuria species in the north-east Atlantic Ocean
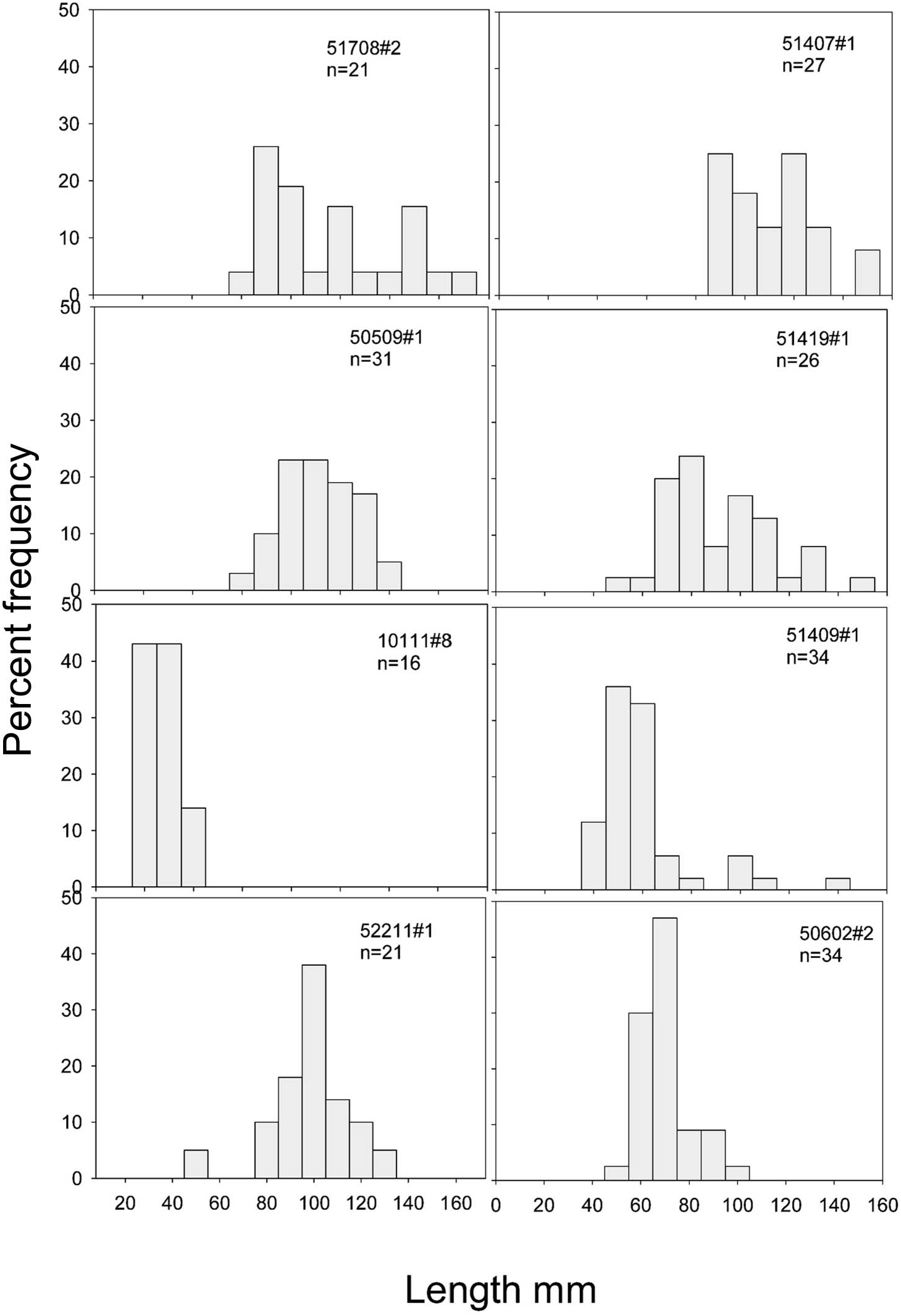
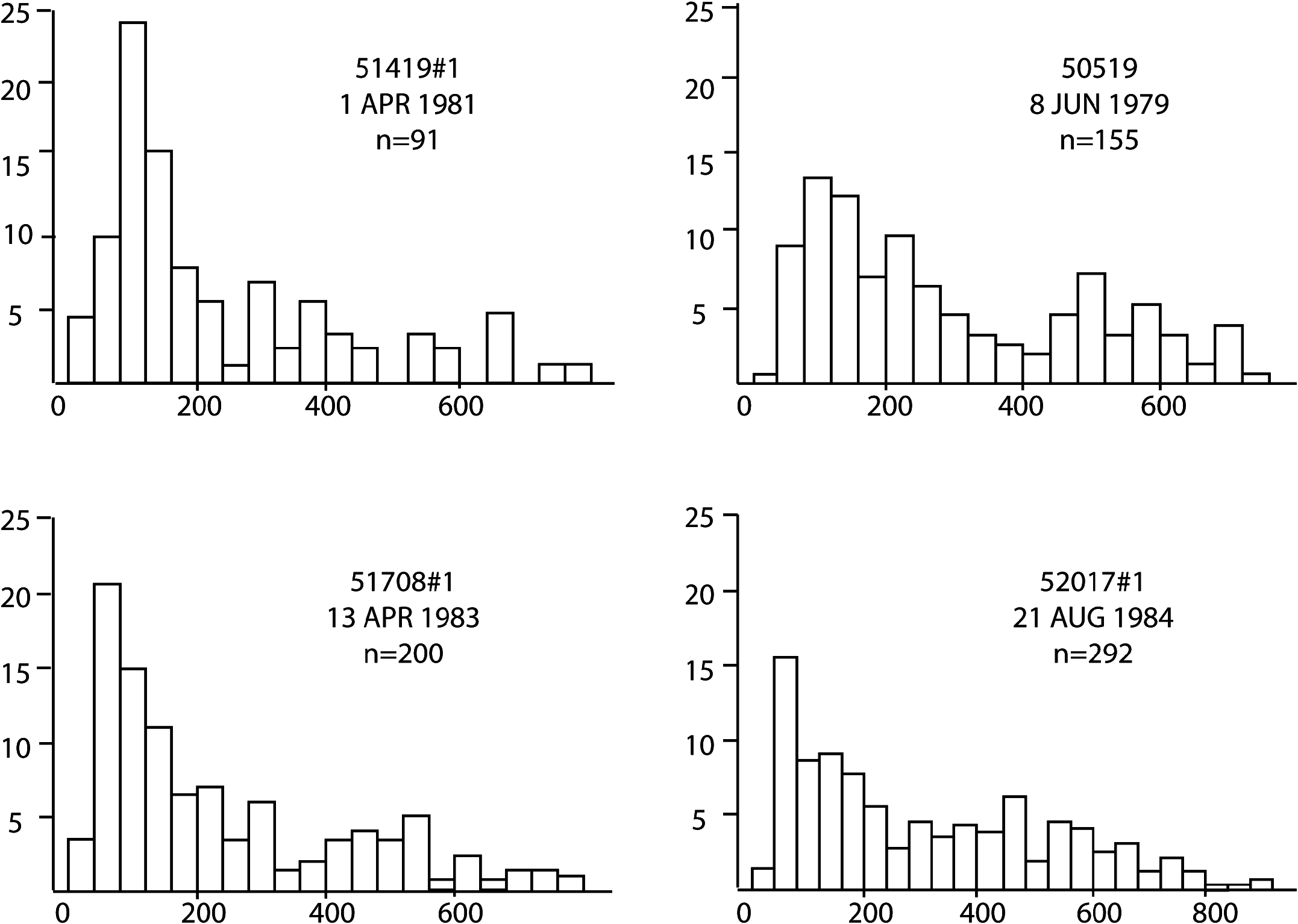
The only species that occurred in sufficient numbers to permit the construction of size / frequency histograms was Zygothuria lactea ( Figure 21 View Figure 21 ). Most of the samples were dominated by large (> 70 mm) specimens although two samples were composed mainly of small (<60 mm) specimens. There was no relationship of depth or time of year with the population size distribution. However, the occurrence of small Z. lactea was concomitant with the appearance of small specimens of the elasipodid Benthogone rosea and over 2000 specimens of the minute holothurian Elpidia , which taken together may indicate patchy and periodic (but not necessarily seasonal) recruitment to the adult population.
The biomass of Z. lactea , in terms of wet and dry weight, ash-free dry weight and calorific content, is shown in Table 16. A maximum ash-free dry weight of 230 g hectare−1 was found at 1500 m, although nearby stations had values <53 g hectare−1, again emphasizing the patchy nature of the distribution of this species. These data correspond closely to those of Walker et al. (1987) who reported a caloric value for this species of 26.24 J mg−1 giving a biomass of 6085 KJ hectare−1.
Reproduction
Gonad morphology and gametogenesis
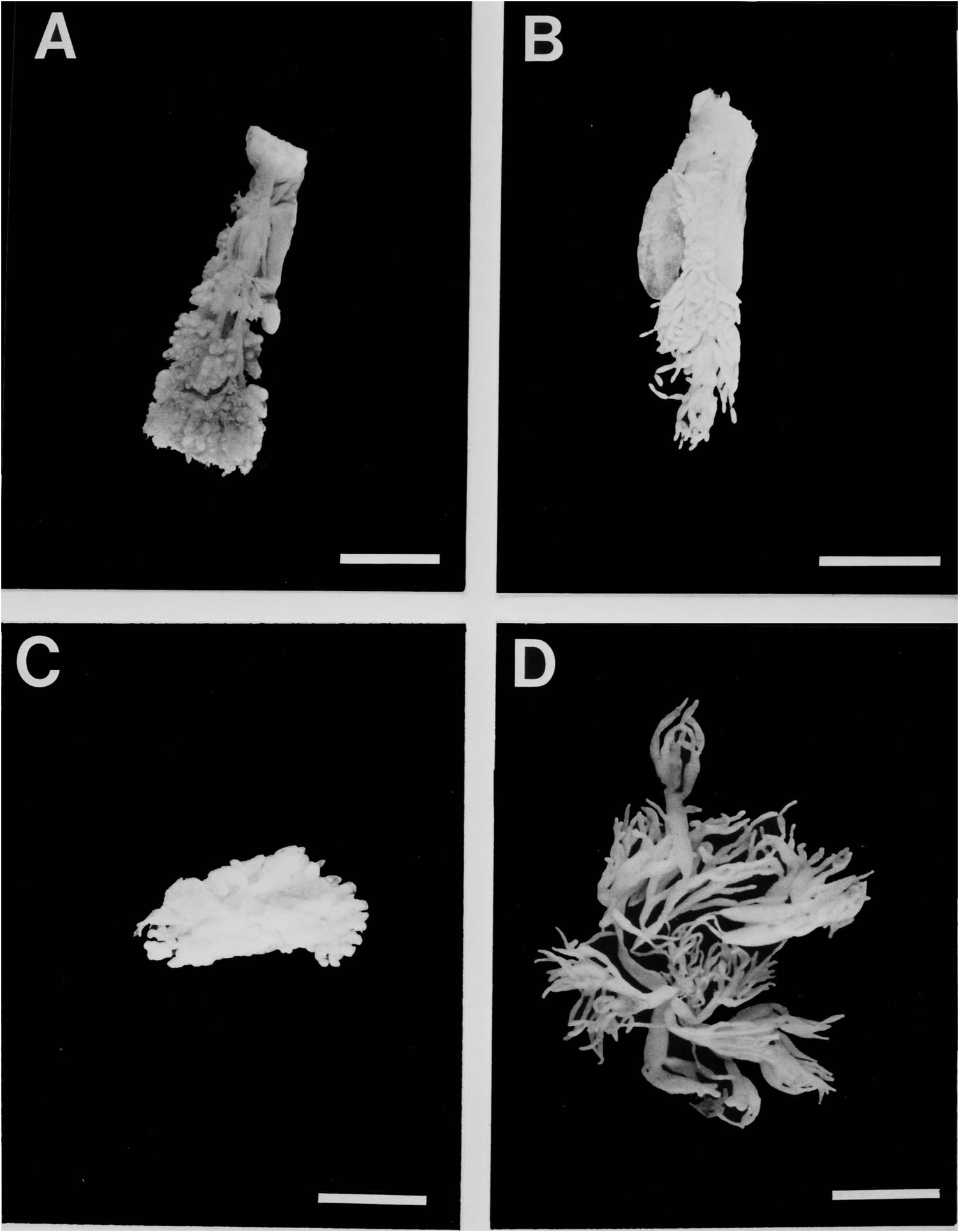
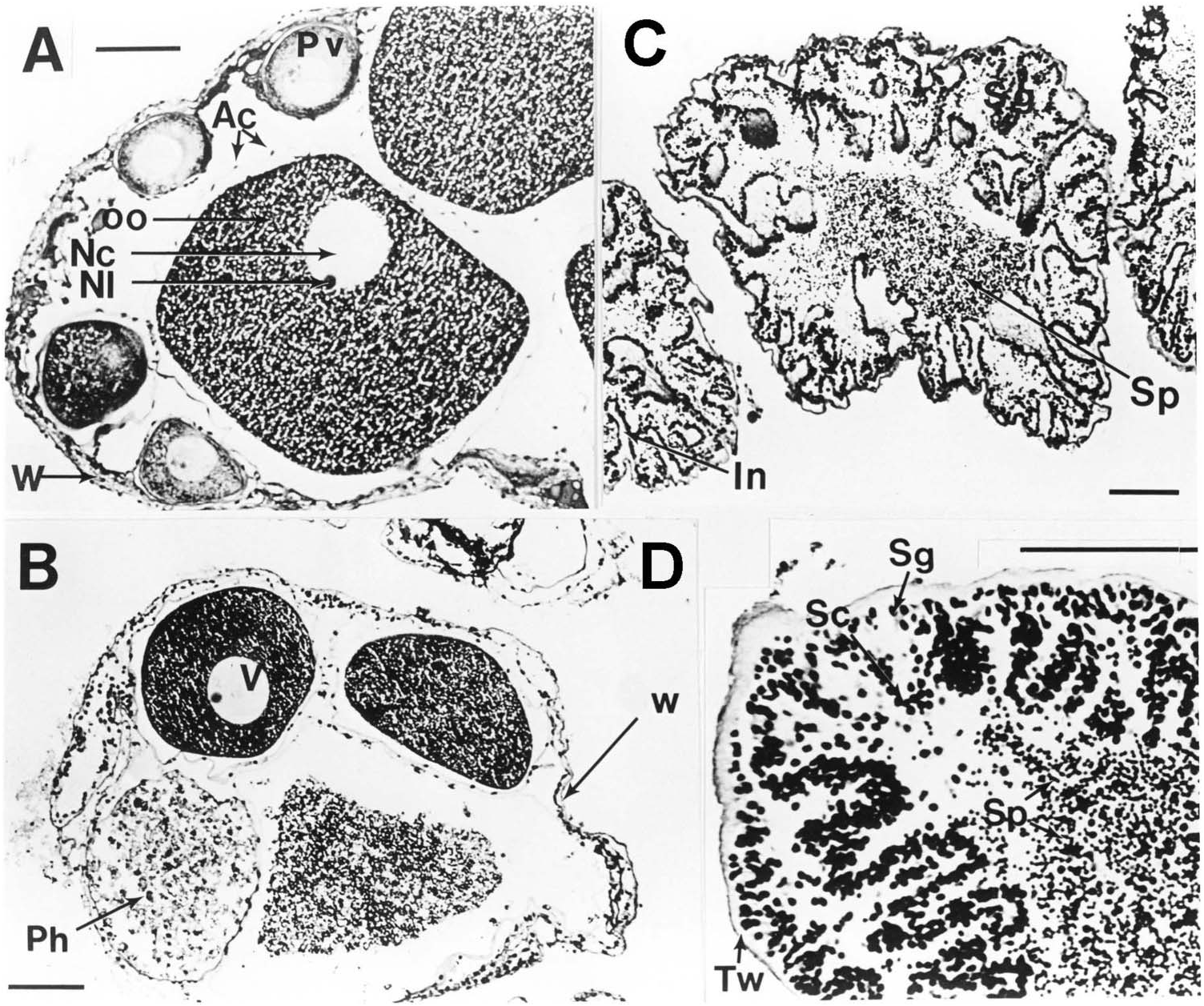
The ovary of specimens of Mesothuria and Zygothuria [except M. intestinalis which is a sequential hermaphrodite ( Mortensen 1927)] consists of many short tubules, which are arranged tightly about the central branching system of ducts, which arise from the gonoduct ( Figure 22 View Figure 22 ). On dissection of live material Z. lactea was found to have large bright pink yolky oocytes. In all species examined the testes consisted of numerous uniformly shaped tubules of similar length and diameter.
Oogenesis in Mesothuria and Zygothuria species
In the newly developing tubules the wall is c. 40 µm thick and consists of three layers; an outer coelomic epithelium, a muscle layer and a connective tissue layer. The inner part of the wall is the germinal epithelium. As the oocytes develop the wall becomes progressively thinner with the muscle layer becoming unrecognizable and the connective tissue layer decreasing in thickness. Newly developed previtellogenic oocytes occur peripherally on the internal surface of the ovarian tubule and are often covered with a layer of follicle cells contiguous with the germinal epithelium ( Figure 23A View Figure 23 ). As with oogenesis in most echinoderms, these previtellogenic oocytes are basophilic and are surrounded by a strong β -metachromatic band. Beyond this stage there appears to be subtle differences between species in the progress of oogenesis.
In Z. lactea the previtellogenic oocytes continue to grow to c. 200 µm diameter at which size the basophilic material becomes peripheral in the ooplasm while the rest of the ooplasm becomes filled with periodic acid Schiff-positive material that increases until the oocyte size attains its maximum of 650 µm diameter ( Figure 23A View Figure 23 ). During this development, however, there are two periods of oocyte breakdown. The first occurs at a diameter of c. 150 µm where some oocytes break down and become filled with globular breakdown products. There are no obvious phagocytes. This process is reminiscent of the ‘nurse cell’ activity seen in Bathybiaster vexillifer ( Tyler et al. 1982) . The second phase of oocyte breakdown is the more typical phagocytosis of unspawned oocytes, as seen in other deep-sea echinoderm species ( Tyler et al. 1985). In this case the large oocytes become filled with phagocytes followed by internal degeneration of the ooplasm ( Figure 23A View Figure 23 ). Two hermaphrodites of Z. lactea were found. One showed different tubules of different sexes and the other showed intratubular hermaphroditism. This gives an incidence of hermaphroditism in this species of <2%.
Oogenesis in M. milleri sp. nov. ( Figure 23B View Figure 23 ) is very similar to that in Z. lactea except that there is no ‘nurse cell’ development. Maximum oocyte size would appear to be similar to that of Z. lactea . In some specimens of M. milleri sp. nov. the proportion of phagocytosed oocytes is very high. In some cases the entire tubule is filled with this material suggesting that the mature oocytes can only be maintained for a certain period before they have to be resorbed.
Examination of the oocytes size / frequency data for these two species ( Figures 24 View Figure 24 and 25 View Figure 25 ) shows that most oocytes are less than 200 µm in diameter. About 10% appear to be undergoing development to the maximum oocyte size. There is no evidence of any reproductive seasonality in either species.
Mesothuria intestinalis is a hermaphrodite, although ‘male and female products are not found ripe at the same time’ ( Mortensen 1927). Examination of the limited number of specimens from the Porcupine Seabight support the observations of Mortensen. Male development occurs first. At a body length of c. 80–110 mm female development occurs and all specimens> 110 mm, up to a maximum size observed of 155 mm, are developing, developed or spent females. Only one specimen was observed to contain both eggs and sperm. These data suggest protandry. Oogenic development is very similar to that of Z. lactea (see above) including the two periods of oocyte breakdown. In the limited number of specimens of M. maroccana available, the smallest specimen is female, whereas larger specimens are either male or have both eggs and spermatogonia. The data for this species are too limited to speculate about its reproductive processes. In Z. candelabri only three specimens were examined of which one was a female. This specimen was poorly preserved and so we are unable to comment on the production of oocytes.
In all species of Mesothuria examined, spermatogenesis would appear to be similar. In early development the testis tubules have a highly convoluted inner surface ( Figure 23D View Figure 23 ). As development proceeds a distinct band of spermatogonia and spermatocytes can be distinguished from mature spermatozoa. In well-developed testes the lumen is packed with spermatozoa ( Figure 23D View Figure 23 ). The only obvious difference between the species is the thickness of the connective tissue layer of the testis wall during early development, which is thin in Z. lactea , M. milleri sp. nov. and Z. candelabri and thick in M. intestinalis and M. maroccana .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Zygothuria thomsoni (Théel, 1886)
| Gebruk, Andrey V., Solis-Marin, Francisco A., Billett, David S. M., Rogacheva, Antonina V. & Tyler, Paul A. 2012 |
Mesothuria thomsoni (Théel, 1886)
| Ostergren H 1896: 350 |
Holothuria thomsoni
| Theel H 1886: 184 |
Holothuria thomsoni var. hyalina
| Theel H 1886: 185 |