
Trilepida macrolepis ( Peters 1857 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4244.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:4945BA57-0971-4F7B-B866-DAF157D25A08 |
|
persistent identifier |
https://treatment.plazi.org/id/03A987ED-FF85-FFDE-FF3D-F9C5FE26B1C7 |
|
treatment provided by |
Plazi |
|
scientific name |
Trilepida macrolepis ( Peters 1857 ) |
| status |
|
Trilepida macrolepis ( Peters 1857)
Figs. 3–4 View FIGURE 3 View FIGURE 4
Stenostoma macrolepis Peters 1857:402 .
Stenostoma (Tricheilostoma) macrolepis — Jan 1861:190. Glauconia macrolepis — Boulenger 1893:69.
Leptotyphlops macrolepis — Ruthven 1922:64.
Leptotyphlops ihlei Brongersma 1933 — Orejas-Miranda 1966:1. Stenostoma (Tricheilostoma) macrolepis — Loveridge 1957:246. Tricheilostoma macrolepis —Adalsteinsson et al. 2009:11. Trilepida macrolepis — Hedges 2011:63.
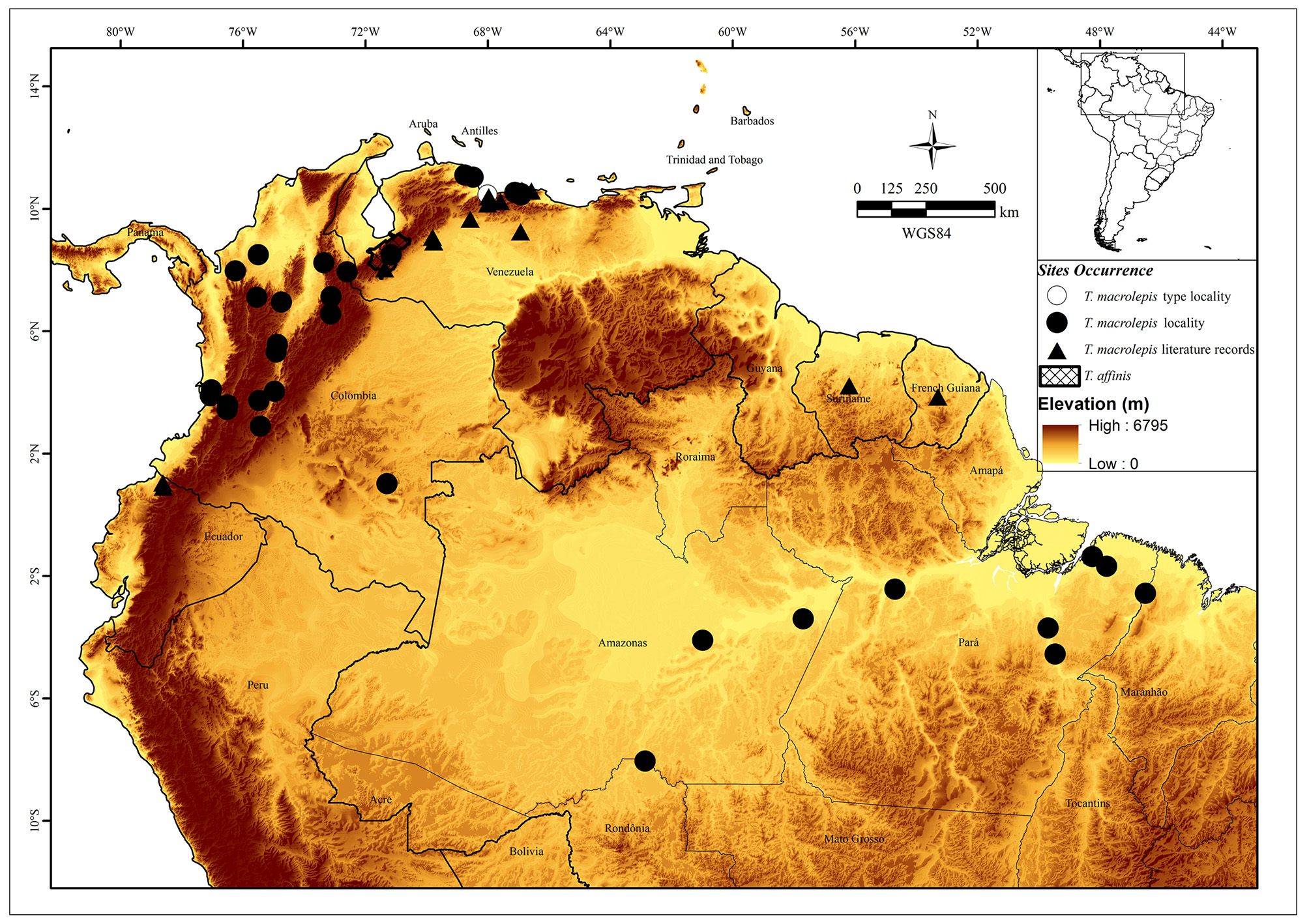
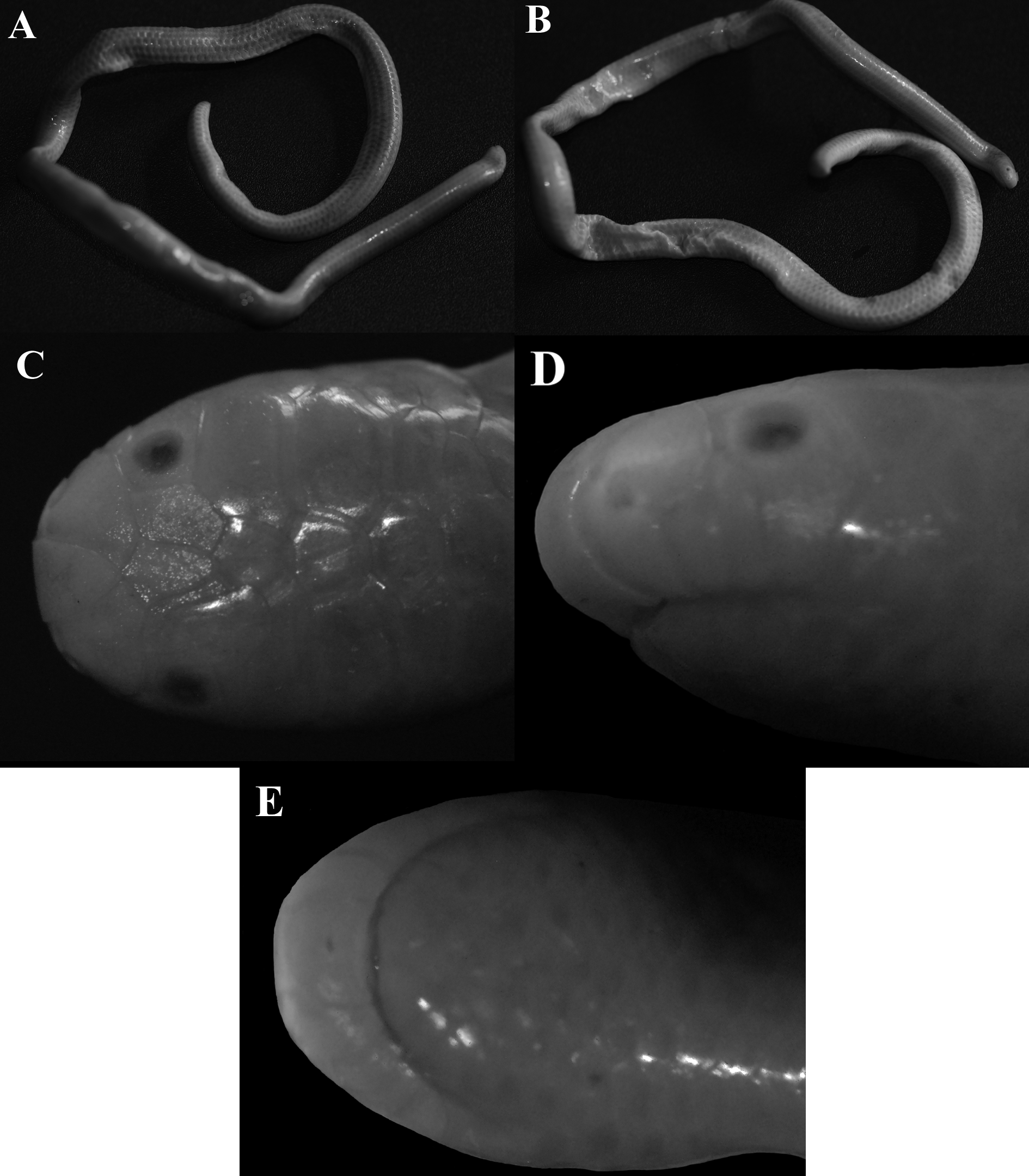
Lectotype. Adult specimen, ZMB 1434 View Materials ( Fig. 3 View FIGURE 3 ) formerly from “ Caracas and Puerto Cabello, Venezuela; restricted to Puerto Cabello (1025'23 N, 6800'45 W), Carabobo State, Venezuela by Orejas-Miranda (1967).
Paralectotypes. Adult, ZMB 5294 View Materials ( Fig. 4 View FIGURE 4 A), from Puerto Cabello , Carabobo State, Venezuela. Adult, undetermined sex , ZMB 5722 View Materials ( Fig. 4 View FIGURE 4 B), from “ S. Amerika (= South America) (see remarks).
Diagnosis. Trilepida macrolepis can be distinguished from all congeners by the following combination of characters: (1) snout truncate in dorsal and ventral views and rounded on lateral view; (2) supraocular present; (3) rostral subtriangular in dorsal view not reaching the anterior limit of ocular scale; (4) rostral similar in size as supranasals; (5) frontal scale longer than other middorsal cephalic shields, and smaller than supraoculars; (6) three supralabials (2+1); (7) four infralabials; (8) scales at midtail 10; (9) fused caudals present; (10) middorsals 211–243 in females and 218–243 in males; (11) midventrals 217–225 in females and 204–221 in males; (12) subcaudals 16– 21 in females and 18–24 in males; and (13) seven dorsal scales rows with dark brown to black pigmentation in contrast with seven pale brown ranging to brown covering the center of scales in ventral rows with lighter borders.
Comparisons. Trilepida macrolepis differs from T. brasiliensis by having supraocular scales (vs. absent of supraocular); differs from T. guayaguilensis by having fused caudals (vs. absence of fused caudals); differs from T. affinis , T. dimidiata , T. jani , and T. nicefori by having three supralabials (vs. two supralabials); differs from T. pastusa by having four, rarely six infralabials (vs. five infralabials); differs from T. joshuai by having snout rounded on lateral view (vs. truncate); differs from T. fuliginosa and T. koppesi by having snout truncate on dorsal and ventral views (vs. rounded); differs from T. macrolepis from T. anthracina by having frontal smaller than supraocular (vs. frontal longer than supraocular); differs from T. brevissima by having frontal longer than other middorsal cephalic shields (vs. with similar size); differs from T. dugandi by having rostral subtriangular in dorsal view (vs. semicircular); differs from T. salgueiroi by having rostral as long as supranasals (vs. shorter than supranasals). Refers to Table 1 for additional meristic and color differences between congeners
Coloration in preservative. Body with seven dorsal scale rows dark brown to black, darkened pigmented on the center of each scale and bordered by lighter edges (pale brown); venter with seven pale brown to brown center of scales bordered cream edges; coloration of the head follows the body pattern; lower limits of supralabials cream; cloacal shield and terminal spine brown.
Variation. Middorsal scales 221–255 (241.1 11.3, n = 16) in females and 211–247 (231.2 8.7, n = 22) in males; midventrals 214–237 (225.7 8.6, n = 13) in females and 201–228 (214.1 7.4, n = 16) in males; subcaudals 17–22 (19.4 1.3, n = 16) in females and 16–24 (20.5 2.3, n = 22) in males; TL 145–378 mm (287.7 mm 63.7, n = 16) in females and 119–321 mm (220.0 mm 59.0, n = 22) in males; TL/ TAL ratio 11.3–15.5 (13.7 1.1, n = 16) in females and 8.5–15.6 (12.5 1.9, n = 22) in males; TAL 6.4–8.9% from TL (7.4% 0.0, n = 16) in females and 6.3– 11.7% (8.2% 0.0, n = 22) in males; TL/MB ratio 32.2–57.7 (48.4 7.0, n = 16) in females and 37.1–68.3 (48.6 8.5, n = 22) in males; TAL /MT ratio 3.8–6.7 (4.9 0.7, n = 14) in females and 4.3–6.6 (5.3 0.6, n = 16) in males; relative eye diameter 1.1–1.9 (1.5 0.2, n = 13) in females and 1.3–2.1 (1.7 0.3, n = 14) in males; relative rostral width 0.3– 0.4 (0.4 0.04, n = 9) in females and 0.3–0.4 (0.4 0.03, n = 11) in males; temporal not distinct (58%, n = 14) or distinct (42%, n =10). Salazar-Valenzuela et al. (2015) reported a single specimen with six infralabials (DHMECN 11400), and other with 12 scale rows at mid-tail ( QCAZ 10247).
Sexual dimorphism. We found secondary sexual dimorphism in the number of middorsal scales (F 1, 36 = 9.3, P <0.01, n = 38), midventral scales (F 1, 27 = 15.4, P <0.001, n = 29), and TL/ TAL ratio (U = 103.0, P <0.05, n = 38) in which females have highest values. Males had the larger proportions of TAL considering the TL (U = 103.0, P <0.05, n = 38). No sexual dimorphism was found in the number of subcaudals scales (U = 119.0, P = 0.09, n = 38). Although, males have relatively larger eyes than females, this difference was not significant (F 1, 25 = 3.0, P = 0.09, n = 27).
Geographic distribution. Trilepida macrolepis is distributed in the trans-Andean portion of Magdalena Valley and Eastern Cordillera of Colombia to cis-Andean lowlands at rain and dry forests of Suriname, French Guiana, Colombia, Venezuela, and Brazil from 0–1,000 m above sea level ( Fig. 2 View FIGURE 2 ).
Remarks. Bauer et al. (2002) analyzed the original type labels in the ZMB collection, and found an error in the type locality indicated in the original description of T. macrolepis . According to W. Peters (1857), the type locality of Stenosoma macroleps was Caracas and Puerto Cabello, Venezuela. However no specimen among the syntypes is labeled as being from Caracas (see Bauer et al. 2002). On the other hand, Puerto Cabello was an important port located about 75 km west from the Caracas (today), where most collected specimens were shipped to the main museums in Europe in the beginning of the last century. Puerto Cabello maybe not corresponds to real locality of collection, as already know for other reptiles and amphibians species also described from the same locality (see Rivas & Manzanilla 1999; Rivas et al. 2012b). Rivas et al. (2012b) also questioned the possibility of some species were actually collected in the mountains surrounding Puerto Cabello, at a nearby locality such as within San Esteban National Park.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trilepida macrolepis ( Peters 1857 )
| Pinto, Roberta R. & Fernandes, Ronaldo 2017 |
Leptotyphlops ihlei
| Hedges 2011: 63 |
| Orejas-Miranda 1966: 1 |
| Loveridge 1957: 246 |
Leptotyphlops macrolepis
| Ruthven 1922: 64 |
Stenostoma (Tricheilostoma) macrolepis
| Boulenger 1893: 69 |
Stenostoma macrolepis
| Peters 1857: 402 |