1. 서론
2. 재료 및 방법
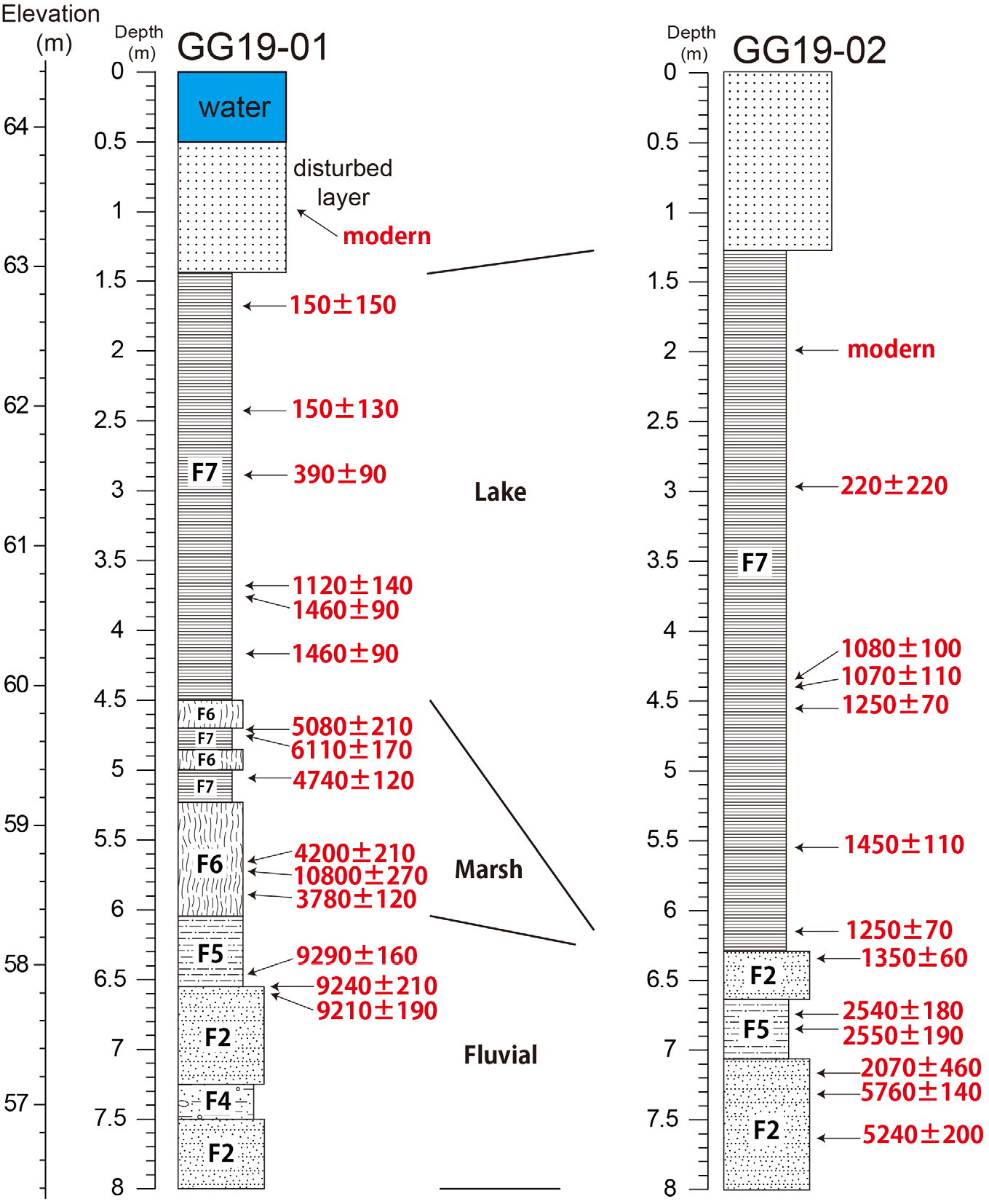
퇴적토 채취 및 연대분석
돌말류 전처리 및 형태 동정
3. 결과
3.1 Cosmioneis hawaiiensis Lowe & Sherwood 2010 (Fig. 3)
3.2 Cymbopleura apiculata Krammer 2003 (Fig. 4)
3.3 Gomphonema contraturris Lange-Bertalot & Reichardt 1993 (Fig. 5)
3.4 Hantzschia vivacior Lange-Bertalot 1993 (Figs 6-8)
3.5 Haslea nautica (Cholnoky) Giffen 1980 (Figs 9, 10)
3.6 Navicula neomundana (Lange-Bertalot & Rumrich) Lange-Bertalot, Jarlman & van de Vijver 2009 (Fig. 11)
3.7 Neidium krasskei Metzeltin & Lange-Bertalot 2007 (Fig. 12)
3.8 Pinnularia interruptiformis Krammer 2000 (Fig. 13)
3.9 Pinnularia platycephala (Ehrenberg) Cleve 1891 (Fig. 14)
3.10 Pinnularia rexlowei Potapova, Kersey & Aycock 2023 (Figs 15-17)
3.11 Placoneis scharfii Lange-Bertalot & Rumrich 2000 (Fig. 18)
3.12 Stauroneis acidoclinatopsis Van de Vijver & Lange-Bertalot 2004 (Fig. 19)
3.13 Stauroneis fuegiana Casa & Van de Vijver 2017 (Figs 20, 21)
3.14 Stauroneis kootenai Bahls 2010 (Figs 22-24)
3.15 Stauroneis pikuni Bahls 2010 (Figs 25, 26)
3.16 Tabellaria flocculosa var. linearis Koppen 1975 (Fig. 27)
4. 고찰
1. 서론
홀로세(Holocene) 시기는 신생대 제4기에 속하는 시기로, 충적세 또는 현세라고도 부르며, 약 10,000년 전부터 현재에 이르는 지질시대이다(Kim et al. 2011). 이 시기는 최후 빙하기가 끝나고 범지구적인 기온상승과 함께 해수면이 상승한 시기로, 현재 국제적으로 직면한 온난화 및 기후변화를 효과적으로 이해하고, 미래의 기후변화를 예측하는데 있어 매우 중요한 시기이다(Chernicoff and Venkatakrishnan 1995; Jung et al. 2010). 이러한 홀로세 시기의 지구환경 변화에 대한 연구는 전세계적으로 주로 해양 퇴적층을 통해서 활발히 이루어져 왔으며, 이에 반해 강, 호수 등의 육상 퇴적층을 이용한 연구는 일부만이 수행되어 왔다(Williams 1995; Sintsov 2003; Cheong and Kim 2008). 그 이유는 과거의 환경 변화 기록을 잘 보존하고 있는 육상 퇴적층의 분포가 전세계적으로 매우 제한적이며, 기술적으로도 양호한 심부의 퇴적층을 채취하기가 쉽지 않았기 때문이다(Cheong and Kim 2008).
고환경을 규명하고 미래의 환경 변화를 예측하기 위해서는 과거부터 퇴적층에 분포하고 있는 돌말류, 화분, 유공충 등 미화석을 이용한 방법이 흔히 사용되어 왔다(Cheong 1989; Paik et al. 2016; Lee et al. 2018; Lee et al. 2020a; Lee et al. 2020b). 이 중 돌말류는 규조류라고도 불리는 미세조류의 한 분류군으로 수 μm~수 백 μm의 매우 작은 크기이며, 모든 수서환경(해수, 기수 및 담수)에서 저서성 또는 부유성으로 서식하면서 광합성을 통해 에너지를 생산하는 일차생산자로 알려져 있다(Ryu et al. 2005). 이들은 염분농도, 수온, 각종 무기염류 등의 변화에 민감하며, 이러한 환경 요인들의 변화에 따라 이들의 군집이 크게 변하므로 오랜 기간 동안의 환경변화를 규명하는데 유용한 생물이다(Ryu et al. 2005). 특히, 돌말류가 가진 규산질(SiO2) 성분의 매우 단단한 세포벽은 오래된 퇴적층 속에서도 화석 상태로 잘 보존되기 때문에, 과거 퇴적층 속에 포함된 돌말류 화석의 생태적 특성을 이용해 고기후 및 고환경을 규명할 수 있다(Sato et al. 1983; Vos and Wolf 1993; Hwang 1998; Ryu et al. 2005). 우리나라에서는 Skvortzow(1929)가 수원의 서호에서 자생하는 마름풀로부터 58종의 돌말류 보고를 시작으로, 현재는 약 1680여종의 돌말류가 기록되었다(Lee and Joh 2015). 그러나 최근 연구 결과에 따르면 우리나라에는 훨씬 더 많은 수의 돌말류 종들이 서식하고 있을 것으로 예측되었다(Park et al. 2014).
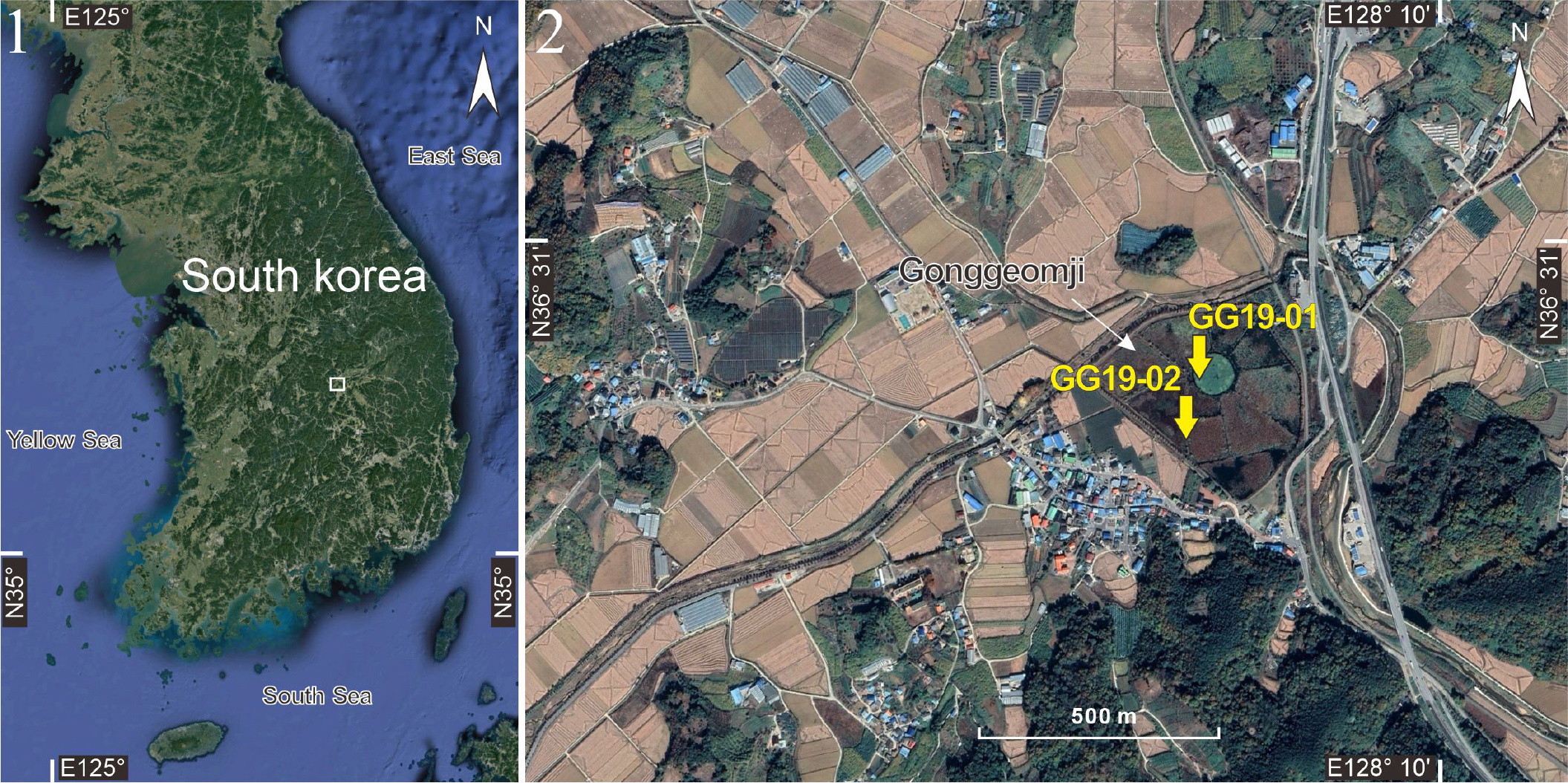
상주 공검지는 삼한시대에 축조된 것으로 전해지며, 제천의 의림지, 밀양의 수산제, 김제의 벽골제 등과 함께 역사적으로 가장 오래된 고대 저수지 중의 하나로 알려져 왔다. 그 동안 공검지에 대한 연구들은 주로 역사학적 측면에서의 연구(Kwon et al. 1995; Park 2007), 생태관광 자원화 측면에서의 연구가 주를 이루었으며(Gwak et al. 2008; Roh et al. 2010; Lim et al. 2011), 돌말류를 대상으로 한 연구는 미비한 실정이었다(Lee et al. 2017). 일부 연구에서 공검지 일대의 퇴적층 시료를 대상으로 돌말류를 분석하여 고환경 및 고기후 변화에 대한 해석이 시도된 바 있지만(Hwang et al. 2014), 퇴적층에 깊이 별 분포하고 있는 돌말류의 종 다양성을 규명하기 위한 연구는 전무했다. Lee et al. (2020b)은 최초로 상주 공검지에서 과거부터 살아 온 돌말류의 종 다양성을 파악하기 위해 4개 지점의 퇴적층에서 깊이별로 출현하는 돌말류의 다양성을 분석하였으며, 화석 돌말류 미기록종 14종(Caloneis wardii, Cocconeis klamathensis, Encyonema vulgare, Eunotia myrmica, Fallacia californica, Gomphonema asiaticum, Gomphonema insigniturris, Gomphonema neoapiculatum, Gomphonema tumens, Pinnularia fusana, Pinnularia parvulissima, Punctastriata lancettula, Sellaphora capitata, and Staurosira dimorpha)을 보고하였다. 우리나라에서 과거에 살았던 미기록 돌말류를 보고한 최초의 사례였다. 이후 공검지의 다른 지점의 퇴적층에서 6종의 미기록 돌말류(Gomphonema lacusrankala, Pinnularia diandae, P. gibba var. hyaline, P. lacunarum, Sellaphora labda var. nipponica, Stauroneis angustilancea)가 추가로 보고되어(Kwon et al. 2023), 현재까지 공검지에서는 20종의 미기록 화석 돌말류가 보고되었다.
본 연구에서는 우리나라 국가생물종목록(NIBR 2022)의 확대를 위해, 공검지에서 과거에 살았던 화석 돌말류 16종을 추가로 보고하고자 한다. 이를 위해 광학현미경(LM) 또는 전계방사형주사전자현미경(FE-SEM)을 통해 종을 동정할 수 있는 정확한 형태적 특징을 확보하기 위해 노력하였으며, 최신의 문헌을 참고하여, 종을 동정하였다. 더불어, 기존에 보고되어 있던 전 세계의 분포에 대한 정보를 제시하고, 공검지에서 각각의 종이 발견된 퇴적층의 연대 및 고환경에 대해 고찰하고자 한다.
2. 재료 및 방법
퇴적토 채취 및 연대분석
퇴적토 채취는 현재 공검지에 물이 채워져 있는 지점(GG19-01)과 산책로 지점(GG19-02)을 대상으로 타격식 소형 시추장비(Percussion drilling set, Royal Eijkelkamp, Netherlands)를 이용해 수행되었다(Table 1, Figs 1, 2). 시추관의 내부에는 퇴적토의 회수를 용이하게 해줄 수 있도록 PVC관을 삽입하여 무수타격방식으로 시추하였다. 시추 코어는 1 m 단위로 채취되었으며, 퇴적토의 건조 또는 산화를 방지하기 위해 현장에서 바로 비닐백으로 진공 포장하여 실험실로 운반되었다. 실험실에서 시추 코어의 비닐백을 제거한 후 나일론 줄을 이용하여 세로로 절반을 절단하였으며, 절반은 퇴적상, 연대, 돌말류 분석 등을 위해 사용하였으며, 나머지 절반은 다시 진공 포장을 하여 향후 추가 분석을 위해 보관하였다. 분석용 코어를 깊이 순으로 나열 후 연대분석을 위해 유기물(식물편, 목편 등)을 확보하였고, 유기물이 없는 경우 퇴적토 시료를 확보하였다(Table 2). 식물편과 목편은 산-염기-산(AAA) 화학처리를 통해 오염물을 제거한 후 분석용 흑연을 추출하였으며, 퇴적토 시료는 휴믹산(Humic acid)을 처리하여 흑연을 추출하였다. 이후 가속질량분석기(Accelerator mass spectrometer, AMS, 한국지질자원연구원)를 이용하여 방사성탄소(14C)를 분석하여 연대측정을 실시하였다. 본 연구에 적용된 모든 탄소 연대의 보정은 Oxcal v4.3 프로그램을 이용하였고 2σ 오차 범위(신뢰한계 95.4%)를 활용하였다.
Table 1.
Information of sampling sites.
| Core | Coordinate (N/E) | Elevation (m) | Length of recovered core (m) |
| GG19-01 | 36°30′50.92″/128°09′39.98″ | 63.8 | 8.5 |
| GG19-02 | 36°30′48.53″/128°09′38.89″ | 64.4 | 9.0 |
돌말류 전처리 및 형태 동정
시추 시료에 분포하고 있는 돌말류의 종 동정을 위해 분석용 코어를 깊이 순으로 나열 후 10 cm 간격으로 부시료를 채취하여 유리병에 보관하였다. 유리병에 보관된 부시료를 건조 후 1 g을 35.5% 과산화수소(Jonsei Chemical Co., Ltd., Japan)와 증류수 15 mL가 들어 있는 50 mL 비이커에 넣고 핫플레이트(SMHS-6, Daihan Scientific Co. Ltd., Republic of Korea)에서 3~4시간 동안 가열하였다. 시료의 반응이 일어나고 침전물이 정체되었을 때 충분한 증류수를 첨가하여 과산화수소를 제거하기 위한 세정 과정을 3회 반복하였다. 세정이 완료된 시료는 100 μm sieve에 옮겨 담아 퇴적층을 제거한 후 시료는 유리병에 보관하였다. 이후 광학현미경 관찰용 영구 슬라이드표본을 만들기 위해 슬라이드 워머(XH-2001, Premiere, USA)에 커버슬립을 놓고, 그 위에 전처리 된 시료 50 μL와 적당량의 증류수를 희석한 후 건조하였다. 봉입제(Mountmedia, Wako, Japan)를 건조된 시료 위에 1~2방울을 떨어뜨린 후 슬라이드로 덮고 알코올 램프로 가열하여 봉입제가 세포 내부에 침투할 수 있도록 하였다. 완성된 영구 슬라이드표본은 건조하고 시원한 곳에 보관하여 경화시켰다. 완전히 경화된 영구 슬라이드는 미분 간섭 광학 현미경(Nikon, Eclipse Ni-U)을 사용하여 정밀하게 관찰되었고, 현미경에 부착된 디지털 카메라(DS-Ri2, Nikon, Japan)를 사용하여 고배율로 촬영되었다. 일부 전처리 시료는 전자현미경을 통한 초미세구조의 관찰을 위해 공극 크기가 2.0 μm인 폴리카보네이트 필터(Whatman plc, UK)를 사용하여 여과되었다. 세포의 초미세구조는 세포가 여과된 필터를 금으로 코팅 후 전계방사형주사전자현미경(FE-SEM, MIRA 3, TESCAN, Czech republic)을 활용하여 종당 최소 5개체 이상을 분석하였다. 돌말류의 크기는 이미지 v1.32 소프트웨어(Schneider et al. 2012)를 사용하여 측정되었으며, 종 당 평균 수치를 기재하였다. 분류학적인 용어는 Ross and Sims (1972), Round et al. (1990), Lee et al. (1995)를 참고하였으며, 형태적 동정은 분류군 별 원기재문 및 최신의 참고문헌을 참고하였다. 분류체계는 ‘AlgaeBase’ (AlgaeBase 2023, https://www.algaebase.org)를 따랐다. 동정된 종은 ‘국가생물종목록’(NIBR 2022, https://species.nibr.go.kr)과 비교하여, 국내 미기록종 여부를 판단하였으며, 속에 대한 국명은 ‘국가 생물종 국명 부여(균류∙조류(藻類) 분야)’(NIBR 2017)을 참고했다.
3. 결과
Genus Cosmioneis Mann & Stickle 1990
앞뒤혹쪽배돌말속
3.1 Cosmioneis hawaiiensis Lowe & Sherwood 2010 (Fig. 3)
형태적 특징: 뚜껑면(valve)은 세로축과 가로축을 중심으로 좌우대칭으로 거의 타원형이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 양 끝은 머리형(capitate)이다. 뚜껑면의 세로축역(axial area, sternum)은 좁으며, 중앙에 가까워질수록 둥글게 확대되고, 등줄(raphe)을 포함한다. 등줄은 세포의 윗뚜껑면(epivalve)과 아래뚜껑면(hypovalve)에 모두 존재하며, 뚜껑면 중앙부의 등줄 끝(proximal raphe ends)은 물방울 모양이며, 뚜껑면 양 끝의 등줄 끝(distal raphe ends)은 같은 방향으로 구부러져 있다. 점무늬열(striae)은 점문(areola, punctum)으로 이루어져 있으며, 가장자리에서 뚜껑 중앙을 향해 방사형(radiate)으로 곡선을 이룬다. 점무늬열의 수는 뚜껑면의 중앙에서는 10 μm 당 11개이며, 끝으로 갈수록 10 μm 당 20개로 촘촘해진다. 점문의 수는 10 μm당 18개로, 뚜껑면의 끝으로 갈수록 작아지고, 중앙부로 갈수록 비교적 커진다. 뚜껑면의 길이는 48.7 µm이며, 폭은 15.1 µm이다.
주요 참고문헌: Lowe and Sherwood 2010.
분포 및 서식환경: Cosmioneis hawaiiensis는 담수에 서식하는 돌말류로, 미국 하와이의 마우이(Maui) 섬에서 최초 발견되었으며, 담수의 습기가 있는 지면(subaerial)에서 서식한다고 알려져 왔다(Lowe and Sherwood 2010). 전세계적으로 출현이 거의 보고되지 않은 종으로(AlgaeBase 2023), 국내에서는 상주 공검지의 퇴적층의 GG19-01 지점 2.4 m, 3.8 m, 4.0 m 깊이 와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m, 3.7 m, 3.9 m 깊이에서 처음 보고되었다.
Genus Cymbopleura (Krammer) Karmmer 1999
바른꼴반달돌말속
3.2 Cymbopleura apiculata Krammer 2003 (Fig. 4)
형태적 특징: 뚜껑면은 좌우비대칭으로 등면과 배면이 구분되는 형태(dosiventral)로, 등면은 볼록하게 올라와 있으며, 배면은 완만하다. 뚜껑면의 양 끝은 좁은 부리형으로 약간 배면쪽을 향해 있다. 뚜껑면의 세로축역은 세포의 말단에서 중앙에 가까워질수록 점점 넓어지고, 중앙에서는 약간 비대칭인 원형 모양이 된다. 등줄은 세로축역에서 뚜껑면의 한 쪽으로 약간 치우쳐 있으며, 뚜껑면의 중앙부와 양 끝의 등줄 끝은 모두 얇다. 뚜껑면 양 끝의 등줄은 등면 쪽으로 구부러진다. 점무늬열은 점문으로 이루어져 있으며, 직선 또는 약간의 방사형이며, 세포의 가로축을 중심으로 사선을 이룬다. 점무늬열의 수는 뚜껑면의 중앙에서는 10 μm 당 9개이며, 끝으로 갈수록 10 μm 당 13개로 촘촘해 진다. 뚜껑면의 길이는 59.9 µm이며, 폭은 20.5 µm이다.
주요 참고문헌: Krammer 2003; Bahls 2019; Buczkó 2016; John 2018; Kulikovskiy et al. 2016; Le Cohu et al. 2011; Shi 2013; Tanaka 2014.
분포 및 서식환경: Cymbopleura apiculata은 담수 돌말류이자 화석 돌말류로 보고되었다(AlgaeBase 2023). 미국의 호수 퇴적층에서 보고되었으며(Krammer 2003), 루마니아의 호수(Buczkó 2016)와, 러시아 자그자(Zagza) 강 내 식물에서 채집된 바 있다(Kulikovskiy et al. 2016). 중국의 타이후(Taihu) 호수(Shi 2013), 일본 세타나타운(Setana town) 개울에서도 본 종이 보고되었다(Tanaka 2014). 이 종은 유기 기질 및 풍부한 수생 식물과 관련이 있는 캐나다 로키산맥(Rocky Mountains)의 작은 연못과 소택지에서 드물게 기록되었으며, 미국 글라셔 국립공원(Glacier National Park)의 컷뱅크 연못(Cut Bank Pond)에서의 pH는 7.77였으며, 전기전도도는 53 µS/cm로 측정되었다(Schweiger et al. 2018). 미국 스키너 연못(Skinner Pond)에서는 pH는 7.82로 측정되었고, 전기전도도는 49 µS/cm로 측정되었다(Bahls et al. 2018). 국내에서는 상주 공검지의 퇴적층의 GG19-01 지점 3.7 m, 3.8 m, 4.0 m 깊이 와 GG19-02 지점 3.7 m, 3.9 m 깊이에서 처음 보고되었다.
Genus Gomphonema Ehrenberg 1832
쐐기돌말속
3.3 Gomphonema contraturris Lange-Bertalot & Reichardt 1993 (Fig. 5)
형태적 특징: 뚜껑면은 윗부분이 넓고 아래부분이 좁은 곤봉형태로 세로축을 중심으로 대칭이며, 가로축을 중심으로 비대칭이다. 가장자리는 약간 각이 져 있고 중앙 부분이 가장 부풀어 있으며, 뚜껑면의 말단까지 거의 선형이다. 뚜껑면의 가장 윗부분(Apice)은 약간 부리모양(rostrate)이며, 가장 아래 부분은 좁고 둥글다. 뚜껑면의 세로축역은 약간 선형으로 좁으며, 등줄은 세로축역에서 뚜껑면의 한 쪽으로 약간 치우쳐 있고 굴곡이 있다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 뚜껑면 중앙부의 등줄 끝은 작은 물방울 모양이며, 뚜껑면 양 끝의 등줄 끝은 갈고리 모양의 같은 방향으로 구부러져 있다. 점무늬열은 점문으로 이루어져 있으며, 약간의 방사형 또는 직선형태이다. 뚜껑면의 가로축 중앙에 위치한 점무늬열은 유리점(stigma)을 지닌다. 점무늬열의 수는 10 μm 당 11개이며, 점문의 수는 2 μm당 11개로, 초승달 또는 길게 찢어진 모양 등을 가진다. 뚜껑면의 길이는 49.8 µm이며, 폭은 9.9 µm이다.
주요 참고문헌: Lange-Bertalot 1993.
분포 및 서식 환경: Gomphonema contraturris는 담수에 서식하는 돌말류로, 독일 프랑켄(franken) 호수에서 발견되어 보고되었으며(Rumrich et al. 2000), 미국의 강, 호수, 저수지, 습지, 연못, 하수, 석호, 퇴적층 등에서도 발견되었고(Bahls 2009), 콜롬비아의 르베니르(Río Porvenir) 강, 아야펠 습지(complejo cenagoso de Ayapel) (Montoya-Moreno et al. 2013), 이라크 샤트 알아랍(Shatt Al-Arab)강(Al-Saedy et al. 2020), 일본 나카이케미 습지(Nakaikemi Wetland)에서도 본 종이 보고되었다(Kihara et al. 2015). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 3.7 m, 3.8 m 깊이와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m, 3.7 m, 3.9 m 깊이에서 처음 보고되었다.
Genus Hantzschia Grunow 1877
마름돌말속
3.4 Hantzschia vivacior Lange-Bertalot 1993 (Figs 6-8)
형태적 특징: 뚜껑면은 배면의 중앙 가장자리가 약간 구부러지고 수축되어 있어, 세로축을 중심으로 비대칭형이다. 반면, 가로축을 중심으로는 대칭형이다. 세포는 활처럼 약간 휘어져 있지만, 거의 직선에 가깝다. 양 끝은 약간 볼록하게 튀어나와 있으며, 등면 쪽으로 약간 구부러져 있다. 등줄은 뚜껑면의 한 쪽 가장자리에 운하(canal)가 길게 이어져 있으며, 이 속에 등줄이 위치한다. 등줄은 양 뚜껑면(상각과 하각)에서 서로 같은 쪽에 위치한다. 점무늬열은 점문으로 이루어져 있으며, 중앙부위부터 세포의 끝까지 가로축과 거의 평행한 직선형이다. 뚜껑면의 한쪽 가장자리에 등줄 안다리(fibulae)가 존재하며, 10 μm 당 7-8개이고, 점무늬열의 수는 뚜껑면의 중앙에서는 10 μm 당 17개이다. 뚜껑면의 길이는 138.3 µm며, 폭은 10.85 µm이다.
주요 참고문헌: Lange-Bertalot 1993; Wang 2018; Wolf 2004; Zelazna-Wieczorek 2011.
전세계 분포 및 서식 환경: Hantzschia vivacior는 담수에 서식하는 돌말류로, 폴란드 도랑에서 보고되었으며(Zelazna-Wieczorek 2011), 캐나다 담수 및 기수, 정수 및 유수 서식지(Bahls et al. 2018), 콜롬비아의 르베니르 강(Río Porvenir), 아야펠 습지 단지(complejo cenagoso de Ayapel)에서 본 종을 보고하였다(Montoya-Moreno et al. 2013). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 1.9 m 깊이에서 처음 보고되었다.
Genus Haslea Simonsen 1974
고래무늬쪽배돌말속
3.5 Haslea nautica (Cholnoky) Giffen 1980 (Figs 9, 10)
Basionym: Navicula nautica Cholnoky 1963
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 대칭이며, 피침형(lanceolate)의 모양을 가진다. 뚜껑면의 양 끝은 약간 날카롭다. 뚜껑면의 세로축역은 매우 좁으며, 중앙에 가까워질수록 약간 확대되고, 등줄을 포함한다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 세로축역을 따라 거의 직선이지만, 중앙 근처에서 한 방향으로 구부러진다. 중앙부의 등줄 끝은 물방울 모양이다. 점늬열(striae)은 뚜껑면 위에 격자 무늬를 이루며, 가로열과 세로열 모두 10 μm당 13-14개이다. 점문의 수 또한 가로 점무늬열, 세로 점무늬열 모두 10 μm당 13-14개이다. 뚜껑면의 길이는 156.9 µm이며, 폭은 34.7 µm이다.
주요 참고문헌: Giffen 1980 (p. 146, Fig. 21); Cholnoky 1963; Giffen 1967.
전세계 분포 및 서식 환경: Haslea nautica는 담수와 해수에서 서식하는 돌말류로, 남아프리카의 키드해(Kidd’s Beach) (Giffen 1967), 멕시코(López-Fuerte and Siqueiros-Beltrones 2016), 터키의 흑해(Black Sea), 마르마라해(Sea of Marmara), 에게해(Aegean Sea), 지중해(Mediterranean Sea) 지역에서 발견되어 보고하였다(Kaleli and Akçaalan 2020). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m 깊이에서 처음 보고되었다.
Genus Navicula Bory 1822
쪽배돌말속
3.6 Navicula neomundana (Lange-Bertalot & Rumrich) Lange-Bertalot, Jarlman & van de Vijver 2009 (Fig. 11)
Basionym: Navicula viridulacalcis subsp. neomundana Lange-Bertalot & Rumrich 2000
Synonyms: Navicula viridulacalcis subsp. neomundana Lange-Bertalot & Rumrich 2000
형태적 특징: 뚜껑면은 좌우대칭으로 거의 타원형이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 양 끝은 약간 쐐기형(wedge-shaped)이다. 뚜껑면의 세로축역은 좁으며, 거의 직선형이고, 중앙에 가까워질수록 둥글게 확대된다. 세로축역의 중앙은 비대칭으로 세로축을 기준으로 한 쪽은 약간 사다리꼴이고, 다른 한 쪽은 반원 모양이다. 세로축역의 등줄은 거의 직선이며, 세포의 윗뚜껑면(epivalve)과 아래뚜껑면(hypovalve)에 모두 존재한다. 뚜껑면 중앙부의 등줄 끝은 한 방향으로 약간 구부러진다. 점무늬열은 점문으로 이루어져 있으며, 뚜껑면의 중앙으로 갈수록 방사형이 두드러지며, 세포의 끝으로 갈수록 약간 직선형이다. 점무늬열의 수는 중앙 부위에서는 10 μm 당 6-7개이며, 말단 부위에서는 10 μm 당 9개이다. 점문의 수는 10 μm당 13-14개다. 뚜껑면의 길이는 57.4 µm이며, 폭은 14.2 µm이다.
주요 참고문헌: Van de Vijver & Lange-Bertalot 2009; Rumrich et al. 2000.
전세계 분포 및 서식 환경: Navicula neomundana는 담수에 서식하는 돌말류로, 스웨덴의 베르군다카날(Bergundakanal), 스크레번(Scraven), 바슬란(Baslan) 하천에서 발견되어 보고되었다(Van de Vijver et al. 2010). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m 깊이에서 처음 보고되었다.
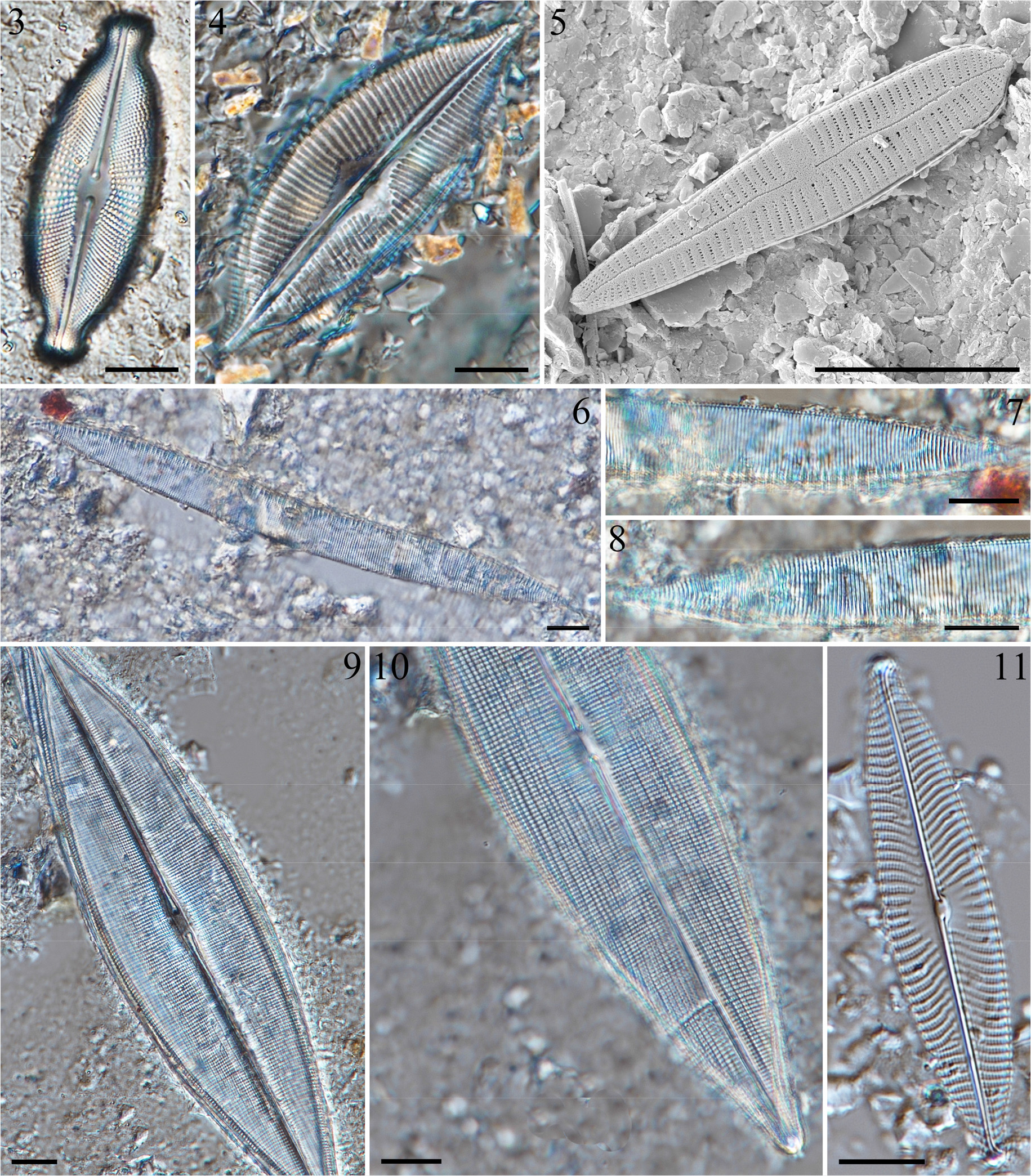
Figs. 3-11.
Light microscope (LM) and Scanning electron microscope (SEM) microphotographs. Figs 3. Cosmioneis hawaiiensis, Fig. 4. Cymbopleura apiculata, Fig. 5. Gomphonema contraturris, Figs 6-8. Hantzschia vivacior, Figs 9, 10. Haslea nautica, Fig. 11. Navicula neomundana. Scale bars: Figs 3, 4, 6-11=10 µm, Fig. 5=20 µm.
Genus Neidium Pfitzer 1871
베틀북돌말속
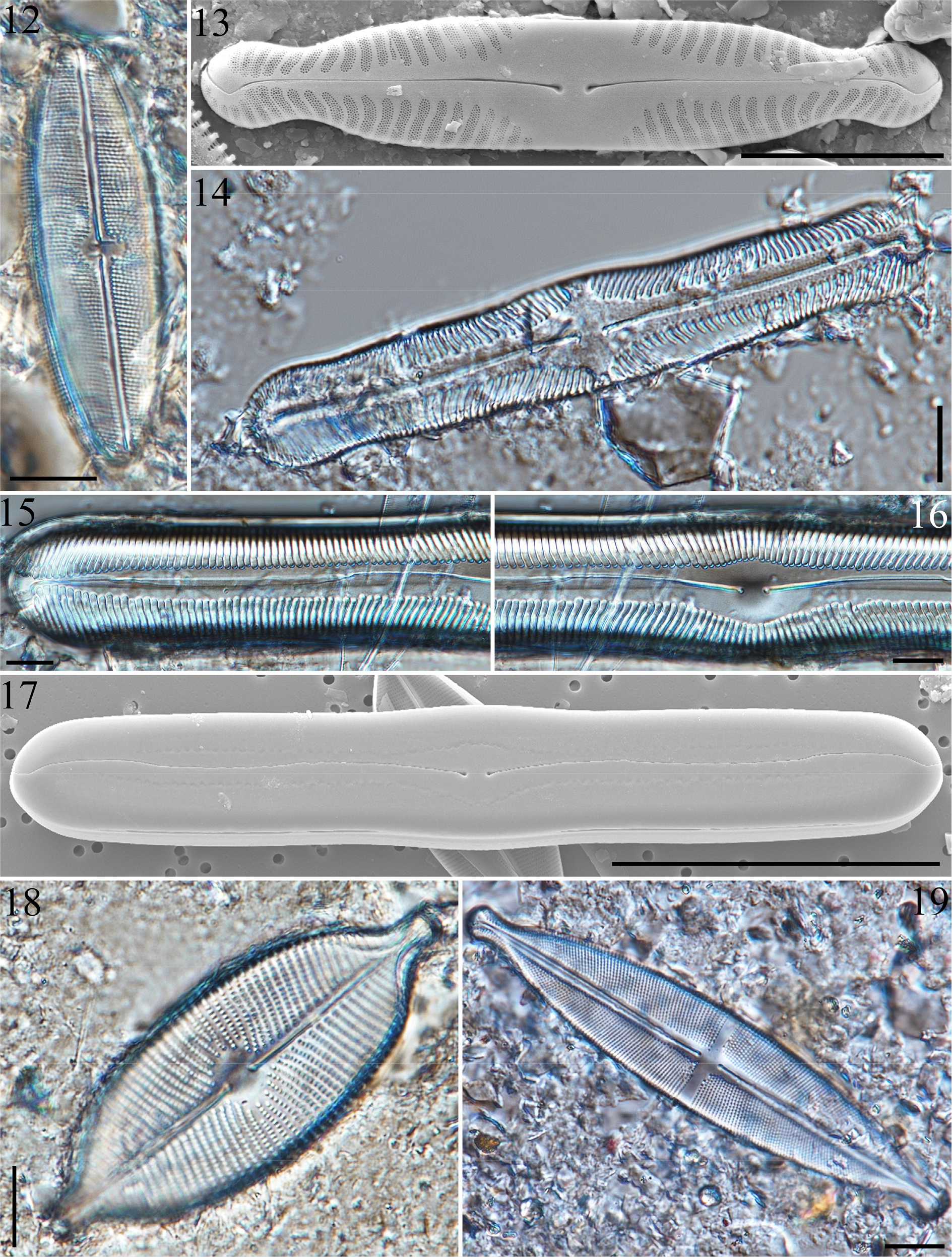
3.7 Neidium krasskei Metzeltin & Lange-Bertalot 2007 (Fig. 12)
형태적 특징: 뚜껑면은 좌우대칭으로 다소 넓은 타원형이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 뚜껑면의 가장자리는 약간 볼록하며, 양 끝은 뾰족하게 둥글다. 세로축역은 좁으며, 중앙에 가까워질수록 둥글게 확대되고, 등줄을 포함한다. 세로축역의 중앙은 세로축을 기준으로 양쪽 모두 반원 모양이지만, 대칭적이지는 않다. 등줄은 약간 비스듬하게 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 뚜껑면 중앙부의 양 등줄 끝은 세로축을 기준으로 각기 반대 방향으로 직각으로 구부러진다. 뚜껑면 양 끝의 등줄 끝은 선형이다. 점무늬열(striae)은 점문(areola, punctum)으로 이루어져 있으며, 가장자리에서 뚜껑 중앙을 향해 약간 비스듬하다. 점무늬열의 수는 10 μm 당 19-20개이며, 점문의 수는 10 μm당 11-12개이다. 뚜껑면의 길이는 48.1 µm이며, 폭은 13.8 µm이다.
주요 참고문헌: Metzeltin and Lange-Bertalot 2007.
전세계 분포 및 서식 환경: Neidium krasskei는 담수에 서식하는 돌말류로 콜롬비아의 브룩샌드(brook-sand)에서 처음 발견되었다(Metzeltin and Lange-Bertalot 2007). 그 후, 콜롬비아의 르베니르(Río Porvenir) 강, 아야펠 습지(complejo cenagoso de Ayapel) (Montoya-Moreno et al. 2013), 일본 나카이케미(Nakaikemi) 습지에서도 본 종을 보고하였다(Kihara et al. 2015). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.7 m, 2.8 m, 3.1 m 깊이와 GG19-02 지점 2.9 m, 3.0 m 깊이에서 처음 보고되었다.
Genus Pinnularia Ehrenberg 1843
빗살돌말속
3.8 Pinnularia interruptiformis Krammer 2000 (Fig. 13)
Synonyms: Pinnularia interrupta f. minor Petersen 1928
형태적 특징: 뚜껑면은 좌우대칭으로 거의 선형이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 뚜껑면의 중앙부 가장자리는 거의 선형이며, 양 끝은 머리형으로 거의 둥글다. 뚜껑면의 세로축역은 중앙으로 갈수록 매우 넓어지며, 중앙부위에서는 뚜껑면의 가장자리까지 확대되어 마치 매우 넓은 육각형 모양을 띈다. 세로축역은 중앙에 등줄을 포함하며, 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재한다. 등줄은 사상형(filiform)으로 전체적으로 거의 직선이지만, 중앙부에서 같은 방향으로 약간 구부러진다. 중앙부의 등줄 끝 모양은 물방울 모양이다. 뚜껑면 양 끝의 등줄은 갈고리 또는 물음표 모양으로 휘어져 있다. 점무늬열은 벌집열(alveolus striae) 구조로, 벌집구멍과 같은 다공으로 구성된 다발의 형태(alveolus)이다. 다발 형태의 점무늬열은 뚜껑면의 중앙에 가까울수록 거의 직선형이고, 끝 부분으로 갈수록 휘어지며, 머리형 부분에서는 다시 직선형으로 변한다. 벌집열의 수는 10 μm 당 11개이며, 뚜껑면의 길이는 48 µm이며, 폭은 7.7 µm이다.
주요 참고문헌: Krammer 2000; Noga et al. 2014.
전 세계 분포 및 서식 환경: Pinnularia interruptiformis 는 담수에 서식하는 돌말류로, 캐나다 로렌시아 오대호(Laurentian Great Lakes)의 합류 지점에 있는 습지 및 저수지에서 보고되었으며(Stoermer et al. 1999), 캐나다 알래스카 툰드라(tundra) 습지와 호수(Sheath and Steinman 1982), 미국 노스포크 니넨스카(North Fork Ninnescah) 강(Patrick and Reimer 1966), 러시아 에르비나이긴(Ervynaigytgyn) 호수, 마타칭아이긴(Matachingaigytgyn) 호수, 에키티키(Ekitiki) 호수(Kharitonov 2010), 미국 우드퍼드(Wooford) 습지(Wołowski et al. 2011), 케냐의 나이바샤(Naivasha) 호수(Cocquyt and Verschuren 2023), 인도 솔트레이크(Salt Lake)에서 발견되었다(Gupta and Das 2020). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m 깊이에서 처음 보고되었다.
3.9 Pinnularia platycephala (Ehrenberg) Cleve 1891 (Fig. 14)
Basionym: Stauroptera platycephala Ehrenberg 1854
Synonyms: Stauroptera platycephala Ehrenberg 1854, Navicula platycephala (Ehrenberg) Cleve & Möller 1882, Schizonema platycephalum (Ehrenberg) Kuntze 1898
형태적 특징: 뚜껑면은 좌우대칭으로 거의 선형이며, 중앙부는 약간 부풀어 있으며, 끝으로 갈수록 약간 좁아지다 다시 부풀어진다. 뚜껑면의 양 끝은 쐐기형(Cuneate)이다. 세로축역은 약간 좁으며, 중앙부로 갈수록 넓어져 마름모꼴의 모양을 띈다. 마름모꼴의 중앙부는 보통 뚜껑면의 가로축 가장자리까지 연장되어 있다. 세로축역은 중앙에 등줄을 포함하며, 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재한다. 등줄은 사상형으로 전체적으로 약간 구부러져 있다. 뚜껑면 양 끝의 등줄은 갈고리 모양으로 강하게 휘어져 있다. 점무늬열은 벌집열(alveolus striae) 구조로, 벌집구멍과 같은 모양(alveolus)의 다공으로 구성된 다발의 형태이다. 벌집열의 수는 10 μm 당 10-11개이며, 뚜껑면의 길이는 93.1 µm이며, 폭은 15.6 µm이다.
주요 참고문헌: Cleve 1891; Hustedt and Pascher 1930; Krammer 2000; Kulikovskiy et al. 2016; Hartley 1996.
전세계 분포 및 서식 환경: Pinnularia platycephala는 담수에 서식하는 돌말류로 러시아 자그자(Zagza) 강에서 식물에 부착된 종을 채집하여 보고하였다(Kulikovskiy et al. 2016). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m 깊이에서 처음 보고되었다.
3.10 Pinnularia rexlowei Potapova, Kersey & Aycock 2023 (Figs 15-17)
형태적 특징: 뚜껑면은 좌우대칭으로 거의 선형이며, 중앙부는 약간 부풀어 있으며, 끝으로 갈수록 약간 좁아지다가 다시 부풀어진다. 뚜껑면의 양 끝은 쐐기형이다. 세로축역은 선형으로 약간 좁으며, 뚜껑면 폭의 약 1/4–1/3 정도의 두께이다. 중앙부로 갈수록 넓어져 타원형의 모양을 띄며, 세로축을 기준으로 한 쪽이 약간 더 부풀어 있는 비대칭형이다. 뚜껑면의 양 끝으로 갈수록 세로축역은 가늘고 날카로워진다. 세로축역은 중앙에 등줄을 포함하며, 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재한다. 등줄은 전체적으로 약간 물결 무늬와 같이 휘고, 중앙부의 등줄 끝 모양은 물방울 모양이며, 같은 방향으로 약간 휜다. 뚜껑면 양 끝의 등줄은 약간 갈고리 모양으로 같은 방향으로 구부러진다. 점무늬열은 벌집열(alveolus striae) 구조로, 벌집구멍과 같은 다공으로 구성된 다발의 형태(alveolus)이다. 벌집열의 수는 10 μm 당 9-10개이며, 뚜껑면의 길이는 183 µm이며, 폭은 24 µm이다.
주요 참고문헌: Potapova et al. 2023.
전세계 분포 및 서식 환경: Pinnularia rexlowei는 담수에 서식하는 돌말류로, 미국에서 채취한 규조토, 스텀프(Stump) 연못과 빅무스(BigMoose) 호수 등 두 호수의 퇴적층 코어 샘플에서 발견되었으며(Potapova et al. 2023), 뉴잉글랜드(New englinad) 호수의 퇴적층에서 발견되어 보고되었다(Hartley 1996; Pentecost and Haworth 2021). 표면 퇴적층에서 발견된 수역은 전기전도도 74 μS/cm, 약산성(pH 6.2) 및 낮은 영양분 농도(TP 7 μm/L, TN 369 μm/L)를 가진 얕은 연못이었다(Potapova et al. 2023). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m 깊이에서 처음 보고되었다.
Genus Placoneis Mereschkowsky 1903
참외쪽배돌말속
3.11 Placoneis scharfii Lange-Bertalot & Rumrich 2000 (Fig. 18)
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 좌우대칭으로 완만한 타원형이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 양 끝은 부리형(rostrate)로 둥글게 돌출되어 있다. 뚜껑면의 세로축역은 좁은 선형으로, 중앙에서 약간 비대칭으로 둥글게 확대되고, 등줄을 포함한다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 전체적으로 거의 선형이지만, 한쪽으로 치우쳐 있다. 뚜껑면 중앙부의 등줄 끝은 물방울 모양으로 거의 직선이다. 점무늬열은 점문으로 이루어져 있으며, 가장자리에서 뚜껑 중앙을 향해 방사형으로 곡선을 이룬다. 뚜껑면의 중앙으로 갈수록 방사형이 두드러지며, 세포의 끝으로 갈수록 거의 직선형이다. 점무늬열의 수는 뚜껑면의 중앙을 중심으로 10 μm 당 10개이며, 점문의 수는 10 μm당 9개이다. 뚜껑면의 길이는 58.5 µm이며, 폭은 22 µm이다.
주요 참고문헌: Rumrich et al. 2000.
전 세계 분포 및 서식 환경: Placoneis scharfii는 담수에 서식하는 돌말류로 베네수엘라 안데스산(Andes Mountains)의 하천에서 본 종을 발견하여 보고하였다(Rumrich et al. 2000). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m 깊이에서 처음 보고되었다.
Genus Stauroneis Ehrenberg 1843
십자돌말속
3.12 Stauroneis acidoclinatopsis Van de Vijver & Lange-Bertalot 2004 (Fig. 19)
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 좌우대칭으로 완만한 창모양(lanceolate)이며, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 양 끝은 부리형으로 돌출되어 있다. 뚜껑면의 세로축역은 좁은 선형으로 등줄을 포함한다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 전체적으로 거의 선형이지만, 약간 한쪽으로 치우쳐 있다. 뚜껑면 중앙부의 등줄 끝은 작은 물방울 모양으로 거의 직선이다. 뚜껑면의 중앙부에는 가로축 방향으로 두꺼운 직사각형의 투명대가 존재하여, 세로축역과 함께 보면 마치 십자가 모양(stauros)처럼 보인다. 이 직사각형의 투명대는 뚜껑면의 세로축을 중심으로 거의 대칭이다. 뚜껑면 양 끝의 등줄 끝은 같은 방향으로 약간 구부러져 있다. 점무늬열은 점문으로 이루어져 있으며, 뚜껑면의 가로축을 중심으로 평행이 아닌 사선을 이룬다. 점무늬열의 수는 뚜껑면의 중앙에서는 10 μm 당 17-18개이며, 점문의 수는 10 μm당 20개이다. 뚜껑면의 길이는 93 µm이며, 폭은 17.5 µm이다.
주요 참고문헌: Van de Vijver et al. 2004.
전세계 분포 및 서식 환경: Stauroneis acidoclinatopsis는 담수에 서식하는 돌말류로, 캐니다 로키산맥(Rocky Mountains)의 5개의 호수와 습지에서 발견되어 보고하였으며, 전기전도도가 낮은 중성 수역(pH 범위는 5.5~8.5, 전기전도도는 10~257 µS/cm)을 선호한다고 알려져 있다(Van de Vijver et al. 2004). 덴마크 그린란드(Greenland) 케케타르수아크(Qeqertarsuaq)에서 수집된 자료에서 기술되었으며, 약산성(pH 6.0), 낮은 전기전도도(<50 µS/cm), 그리고 매우 낮은 영양염을 가진 얕은 수역에서 발견되었다고 보고하였다(Bahls 2010). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m, 3.1 m, 3.4 m 깊이와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m 깊이에서 처음 보고되었다.
3.13 Stauroneis fuegiana Casa & Van de Vijver 2017 (Figs 20, 21)
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 좌우대칭으로 완만하고 넓은 창모양이고, 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 좁아진다. 뚜껑면의 양 끝은 약간 부리형이다. 뚜껑면의 세로축역은 좁은 선형으로 등줄을 포함한다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 전체적으로 거의 선형이지만, 약간 한쪽으로 치우쳐 있다. 뚜껑면 중앙부의 등줄 끝은 작은 물방울 모양이며, 같은 방향으로 약간 휘어진다. 뚜껑면 양 끝의 등줄 끝은 갈고리 모양으로 같은 방향으로 구부러져 있다. 뚜껑면의 중앙부에는 가로축 방향으로 두꺼운 약간 직사각형 모양의 투명대가 존재하여, 세로축역과 함께 보면 마치 십자가 모양처럼 보인다. 이 직사각형의 투명대는 뚜껑면의 가장자리로 갈수록 넓어지며, 중앙으로 갈수록 좁아지고, 세로축을 중심으로 거의 대칭이다. 뚜껑면의 가장자리를 따라 길게 이어진 좁고 투명한 선(marginal crest)이 뚜렷이 보인다. 점무늬열은 점문으로 이루어져 있으며, 뚜껑면의 가로축을 중심으로 평행이 아닌 사선 또는 약간의 방사형을 이룬다. 점무늬열의 수는 뚜껑면의 중앙에서는 10 μm 당 17-18개이며, 끝으로 갈수록 10 μm 당 18-19개이다. 점문의 수는 10 μm당 16-17개이다. 뚜껑면의 길이는 105.4 µm이며, 폭은 22.4 µm이다.
주요 참고문헌: Casa et al. 2017.
전세계 분포 및 서식 환경: Stauroneis fuegiana는 담수에 서식하는 돌말류로, 아르헨티나 이탄 늪지, 안도라 계곡 상류(Upper Andorra Valley) 이탄습지 퇴적층에서 채집하여 발견되었다(Casa et al. 2017). 이 종은 과거에 S. phoenicenteron 또는 S. gracilis로 동정되었을 가능성이 있으며 일반적으로 빈영양, 전기전도도가 낮고 산성에서 약산성(pH 3,6-6,8) 수체의 퇴적층과 이끼 샘플에서 발견된다고 알려져 있다(Casa et al. 2017). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m, 3.1 m, 3.4 m 깊이와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m 깊이에서 처음 보고되었다.
3.14 Stauroneis kootenai Bahls 2010 (Figs 22-24)
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 좌우대칭이며, 완만한 창모양으로 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 매우 좁아진다. 양 끝은 부리형으로 매우 돌출되어 있다. 뚜껑면의 세로축역은 좁은 선형으로 등줄을 포함하며 약간 융기되어 있다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 전체적으로 거의 선형이지만, 약간 한쪽으로 치우쳐 있다. 뚜껑면 중앙부의 등줄 끝은 아주 작은 물방울 모양이며, 약간 완만하게 휘어진다. 뚜껑면 양 끝의 등줄 끝은 갈고리 모양으로 같은 방향으로 구부러져 있다. 뚜껑면의 중앙부에는 가로축 방향으로 두꺼운 약간 직사각형 모양의 투명대가 존재하여, 세로축역과 함께 보면 마치 십자가 모양처럼 보인다. 이 직사각형의 투명대는 뚜껑면의 가장자리로 갈수록 좁아지는 경향을 보이며, 세로축을 중심으로 약간 비대칭이다. 점무늬열은 점문으로 이루어져 있으며, 뚜껑면의 가로축을 중심으로 평행이 아닌 사선을 이룬다. 점문은 둥근 모양 또는 타원형으로 다소 불규칙적인 열을 형성하고 있으며, 뚜껑면 전체에 산재되어 있다. 세로축열의 가장자리에는 점문이 세로축을 따라 길게 연결되어 점무늬열을 이루고 있으며, 10 μm당 20-22개의 점문으로 구성되어 있다. 점무늬열의 수는 10 μm 당 14-15개이다. 뚜껑면의 길이는 103 µm이며, 폭은 25 µm이다. 뚜껑면의 안쪽에서 세포의 양 끝에 등줄끝말림(helictoglossa)이 뚜렷히 관찰된다.
주요 참고문헌: Bahls 2010.
전세계 분포 및 서식 환경: Stauroneis kootenai는 담수에 서식하는 돌말류로, 캐나다 워터튼 레이크 국립공원(Waterton Lakes national park), 하이다과이(Haida Gwaii), 클리어워터(Clearwater) 강, 코퍼마인(Coppermine) 강, 배일리-백(Baillie-Back) 강 및 후드(Hood) 강에서 발견되었다(Bahls et al. 2018). 또한 본 종은 미국의 여러 호수, 연못, 늪 및 경사도가 낮은 하천에서 발견되었으며 pH 범위는 6.7~7.8이고 특정 전도도 범위는 22~169 μS/cm를 선호한다고 보고되었다(Bahls 2010). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m, 3.1 m, 3.4 m 깊이와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m 깊이에서 처음 보고되었다.
3.15 Stauroneis pikuni Bahls 2010 (Figs 25, 26)
형태적 특징: 뚜껑면은 세로축과 가로축을 중심으로 좌우대칭이며, 좁은 창모양으로 중앙부의 폭이 가장 넓고 양 끝으로 갈수록 급격히 좁아진다. 양 끝은 부리형으로 매우 돌출되어 있다. 뚜껑면의 세로축역은 좁은 선형으로 등줄을 포함하며 약간 융기되어 있다. 등줄은 세포의 윗뚜껑면과 아래뚜껑면에 모두 존재하며, 전체적으로 거의 선형이고, 중앙에 위치한다. 뚜껑면 중앙부의 등줄 끝은 물방울 모양이며, 직선형이다. 뚜껑면 양 끝의 등줄 끝은 갈고리 모양으로 같은 방향으로 구부러져 있다. 뚜껑면의 중앙부에는 가로축 방향으로 두꺼운 약간 직사각형 모양의 투명대가 존재하여, 세로축역과 함께 보면 마치 십자가 모양처럼 보인다. 이 직사각형의 투명대는 세로축을 중심으로 한 쪽에서는 뚜껑면의 가장자리로 갈수록 약간 좁아지는 경향을 보이며, 다른 한 쪽은 가장자리로 갈수록 다소 넓어지는 비대칭형이다. 점무늬열은 점문으로 이루어져 있으며, 뚜껑면의 가로축을 중심으로 평행이 아닌 약간의 사선을 이룬다. 점문은 둥근 모양 또는 타원형으로 다소 불규칙적인 배열로 뚜껑면 전체에 산재되어 있으나, 중앙의 십자형 투명대(hyaline)에 인접한 뚜껑면에는 거의 분포하지 않는다. 세로축열의 가장자리에는 점문이 세로축을 따라 길게 연결되어 점무늬열을 이루고 있으며, 10 μm당 26개의 점문으로 구성되어 있다. 점무늬열의 수는 10 μm 당 25-27개이다. 뚜껑면의 길이는 76 µm이며, 폭은 13 µm이다.
주요 참고문헌: Bahls 2010.
전세계 분포 및 서식 환경: Stauroneis pikuni는 담수에 서식하는 돌말류로, 캐나다 로키산맥(Rocky Mountains) 전역의 호수와 연못에 널리 분포되어 있지만 많이 발견되는 경우는 거의 없다. Stauroneis pikuni가 발생하는 수역의 pH 범위는 6.6~7.3이고 특정 전도도 범위는 8~71 μS/cm로 알려져 있으며(Van de Vijver et al. 2004), 덴마크 그린란드(Greendland)의 자켄버그(Zackenberg) 지역에서 이 분류군(Stauroneis gracilior) 을 보고했다(Van de Vijver et al. 2004). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 2.4 m, 2.6 m, 2.7 m, 2.8 m, 3.1 m, 3.4 m 깊이와 GG19-02 지점 2.9 m, 3.0 m, 3.3 m 깊이에서 처음 보고되었다.
Genus Tabellaria Ehrenberg ex Kützing, 1844
볼록뼈돌말속
3.16 Tabellaria flocculosa var. linearis Koppen 1975 (Fig. 27)
형태적 특징: 뚜껑면은 가로축과 세로축을 중심으로 좌우 대칭으로, 일반적으로 가운데가 볼록하게 부풀려진 직선형이며 점점 가늘어 지다가 양 끝부분은 크게 둥근 머리모양으로 돌출되어 있다. 뚜껑면의 세로축역은 거의 존재하지 않을 만큼 매우 좁다. 점무늬열은 점문으로 이루어져 있으며, 10 μm 당 19-20개가 관찰되었다. 뚜껑면의 길이는 71 µm이며, 폭은 뚜껑면 중앙의 넓은 부분은 6.7 µm이며, 그 외 부분은 3.7 µm이다.
주요 참고문헌: Koppen 1975.
전세계 분포 및 서식 환경: Tabellaria flocculosa var. linearis 는 담수에 서식하는 돌말류로 스웨덴 스칸디나비아(Scandinavia), 오스트리아 알프스(Alpls), 미국(Koppen 1975; Camburn and Charles 2000; Siver et al. 2005) 및 캐나다(Poulin et al. 1984; Lange-Bertalot 1988; Zimmermann et al. 2010)에서 보고되었다. 스웨덴의 라탄(Rat-tån) 강에서 발견된 Tabellaria flocculosa var. linearis는 수온 18.5°C, pH 6.48, 전도도 19 μS/cm, 총질소 9 μg/L 및 총인 8 μg/L를 나타내는 등, 낮은 알칼리도, 중간 정도의 산성, 낮은 전기전도도를 선호하였다(Heudre et al. 2021). 한편 Tabellaria flocculosa var. linearis 세포는 지그재그 모양의 군집을 형성하며 다른 기질에 부착해 무한정 길어지는 것으로 보고되었다(Heudre et al. 2021). 국내에서는 상주 공검지 퇴적층의 GG19-01 지점 1.9 m, 2.4 m 깊이에서 처음 보고되었다.

4. 고찰
본 연구에서는 오래된 담수 퇴적층에서 그 동안 국내에 보고되지 않은 16종의 돌말류(Cosmioneis hawaiiensis, Cymbopleura apiculata, Gomphonema contraturris, Hantzschia vivacior, Haslea nautica, Navicula neomundana, Neidium krasskei, Pinnularia interuptiformis, Pinnularia platycephala, Pinnularia rexlowei, Placoneis scharfii, Stauroneis acidoclinatopsis, Stauroneis fuegiana, Stauroneis kootenai, Stauroneis pikuni, Tabellaria flocculosa var. linearis)의 분류학적 특징을 처음으로 기재하였다. 퇴적층의 연대분석 결과와 미기록 돌말류의 출현 깊이를 비교해 보면, 보고된 종들이 발견된 퇴적층 구간은 지금으로부터 약 1460±90년 전~150±130년 전 시기로, Cosmioneis hawaiiensis와 Gomphonema contraturris는 1460±90년 전~150±150년 전 시기의 퇴적층에서 발견되었으며, Cymbopleura apiculata는 1460±90년 전~220±220 년 전, Hantzschia vivacior와 Tabellaria flocculosa var. linearis 는 150±150년 전, Haslea nautica, Navicula neomundana, Pinnularia interruptiformis, Pinnularia platycephala, Pinnularia rexlowei와 Placoneis scharfii 는390±90년 전~150±130년 전, Neidium krasskei, Stauroneis acidoclinatopsis, Stauroneis fuegiana, Stauroneis Kootenai와 Stauroneis pikuni는 1120±140년 전~150±150년 전 시기에 발견된 것으로 판단된다(Fig. 28, Table 2). 기존 연구결과에 의하면, 이 종들이 출현한 퇴적층의 구간(F7 구간)은 미약한 판상의 엽리층을 가지는 점토층으로 당시 호소의 환경이었을 것으로 추정되는 구간이다(Lee et al. 2017). 전세계적인 분포를 보면, 이 종들은 대부분 현생의 담수 환경에서만 서식한다고 보고되어 왔지만(Cymbopleura apiculata, Gomphonema contraturris, Pinnularia rexlowei, Stauroneis fuegiana는 퇴적층에서도 보고), 본 연구를 통해 우리나라의 경우 이들이 홀로세 시기 담수환경에서 오래 전에 살았다는 것을 알 수 있었다. 향후 이 종들이 우리나라의 현생 담수 환경에서도 서식하고 있는지에 대한 추가 조사가 필요하지만, 아직까지는 현생에서 발견된 적이 없다는 점으로 보아, 과거의 환경변화 또는 기후변화에 민감한 종들이라 판단된다. 본 연구에서 발견된 16종을 포함하여 이제껏 공검지의 오래된 퇴적층에서 보고된 미기록 돌말류는 총 36종으로, 이는 현재까지 우리나라에서 기록되지 않았던 다양한 돌말류가 과거 담수 퇴적층에서 발견되었다는데 큰 의의를 지닌다.