Patch reefs as sources of long and continuous skeletal records of horizontal reef expansion in Dongsha Atoll, South China Sea
 Yi Wei
Yi Wei Chuan-Chou Shen
Chuan-Chou Shen Keryea Soong
Keryea Soong- 1Department of Oceanography, National Sun Yat-sen University, Kaohsiung, Taiwan
- 2Institute of Oceanography, National Taiwan University, Taipei, Taiwan
- 3High-Precision Mass Spectrometry and Environment Change Laboratory (HISPEC), Department of Geosciences, National Taiwan University, Taipei, Taiwan
- 4Research Center for Future Earth, National Taiwan University, Taipei, Taiwan
Atolls are annular midocean reefs with various topographical features. The reef rim of an atoll encloses a central lagoon and lagoonal patch reefs therein. Two hypotheses explaining the origins of patch reefs in atoll lagoons have been developed: the karst hypothesis and the growth hypothesis, which emphasize the role of uneven erosion and uneven timing of reef accretion (especially horizontal expansion), respectively, in the formation and growth of patch reefs. In this study, we investigated trends in the locations, depths, ages, sizes, and other features of flat-top and pinnacle-top reefs in the Dongsha Atoll in the South China Sea by analyzing bathymetric LiDAR images and performing U-Th dating on samples we collected from various reefs in the atoll. Three of our main findings were consistent with the growth hypothesis: (1) the differences in ages of the centers of different flat-top patch reefs reflected the different times at which each patch reef reached the surface and started to expand horizontally; (2) the top surfaces of individual flat-top patch reefs had old centers and young peripheries, reflecting horizontal growth; and (3) all the pinnacle-top reefs we examined had more live corals on their tips than on the surrounding bases. We observed no positive correlation between the ages of the centers and the diameters of flat-top patch reefs; that is, larger reefs did not necessarily have older centers, indicating that various complex factors affect the expansion and age patterns of patch reefs. Overall, the findings of this study provide support for the growth hypothesis and for the use of flat-top patch reefs as sources of easily accessible long-term coral skeletal records. They may therefore serve as a reference for future studies on asymmetric reef expansion and other relevant topics.
Introduction
Atolls are annular mid-ocean reefs. They typically comprise various topographical features, such as spurs and groves, reef crests, and backreef flats. The reef rim of an atoll encloses a central lagoon and lagoonal patch reefs therein (Goldberg, 2016). Stony corals grow in these habitats, and reefs are shaped by different environmental factors in their respective habitats. For example, mechanical or physical erosion is an obvious force shaping spurs and grooves in shallow forereefs (Duce et al., 2014); however, such a force is substantially diminished in atoll lagoons. Charles Darwin’s (Darwin, 1842) subsidence hypothesis of atoll formation; that is, an atoll is formed by a volcanic eruption to the sea surface and the later subsidence and immersion of the resulting volcanic island, was supported by the discovery of old reef strata >1000 m in depth at the base of a Pacific atoll (Davis, 1928). However, the geological history of atolls is more complex than that envisioned by Darwin (Dickinson, 2009; Droxler and Jorry, 2021). The fluctuation of seawater levels since the last glacial maximum has presumably shaped atoll reefs (Grigg et al., 2002; Siddall et al., 2003; Woodroffe and Horton, 2005), too. During the Holocene, as sea levels rose to near current levels, environmental factors such as wind, waves, water temperature, sea level, and current have been highly variable within an atoll. The dynamics of reef formation can thus differ substantially at the local scale (Mazzullo et al., 1992; Purdy and Gischler, 2005). Because forereefs are exposed to strong waves, they may be highly dynamic; the processes of skeletal deposition and erosion compete with each other within forereefs (Hennige et al., 2017). By contrast, reefs may easily grow and accumulate in lagoons, which are protected from waves by reef crests and reef rims (Brown and Dunne, 1980; Wang and Hung, 2009). Atolls developed on a continental shelf, edge, or slope are commonly considered to differ considerable from other types of atolls (Woodroffe and Biribo, 2011). Three types of atolls have been identified in the South China Sea: ocean, shelf, and slope atolls (Guozhong, 1998). Dongsha Atoll (Figure 1) is a shelf atoll in the northern South China Sea (Tomascik et al., 1997).
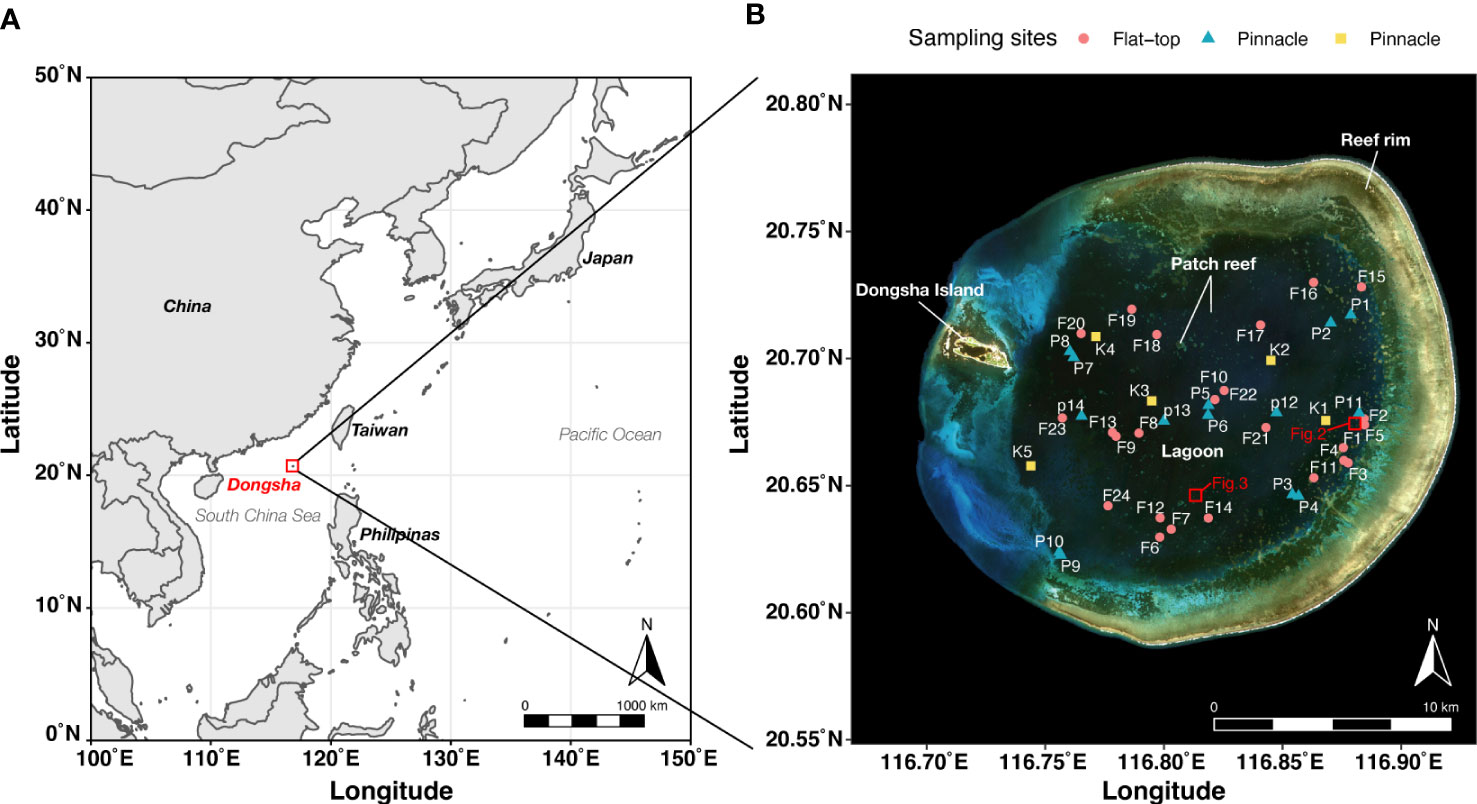
Figure 1 Dongsha Atoll sampling sites. (A) Dongsha Atoll (red square) is located in the northern South China Sea at 20.70°N and 116.80°E. (B) Coral core samples were collected from the surfaces of 24 flat-top patch reefs (red circles, F1–F24) and 14 pinnacle-top patch reefs (blue triangles, P1–P14) in the Dongsha Atoll lagoon. Coral cover data of five pinnacle-top patch reefs were obtained using transect photographs (photographs along 25 m line transect; yellow squares, K1–K5).
Patch reefs (Figure 2), also known as lagoonal reefs, are typically cone- or truncated cone-shaped and can be divided into three sections: the reef top, reef base, and reef slope (James, 1983). Two hypotheses explaining the origins of patch reefs in atoll lagoons have been developed: the karst hypothesis and the growth hypothesis. The karst hypothesis (Purdy, 1970), which posits erosion as the principal force during reef exposure in air, e.g., during the last glacial maximum; when the eustatic sea level dropped to 120 m below the current sea level (Woodroffe and Horton, 2005), may explain the mechanisms underlying patch reef formation in lagoons. The geomorphology of a lagoon and the patch reefs therein may be affected by the variable rates of erosion caused by freshwater (Purdy and Winterer, 2001). On the other hand, when the sea level rose during the Late Pleistocene, more vertical and horizontal spaces in the water column were available for reef development. During this stage, patch reefs often grew and were scattered throughout lagoons (James, 1983). Horizontal growth of reef expansion represents the dominant lateral pattern of patch reef accumulation in static sea-level settings in shallow water environments (Mazzullo et al., 1992). The two hypotheses may be two alternating processes with the erosion mechanism working principally when the reefs are exposed, whereas the growth mechanism dominates when the reefs are underwater and alive. A number of studies have shown that patch reefs are formed by the thick growth of Holocene reefs on Pleistocene limestone of the basement (Halley et al., 1977; Aronson et al., 1998; Gischler and Hudson, 2004). Coral growth can also form considerably thick reef on karst landforms. e.g., in the submerged karst landforms of the Ryukyu Islands, ~34 m of postglacial reef complex overlies the Pleistocene limestone was founded by drilling survey (Kan et al., 2015).

Figure 2 Aerial view of flat-top patch reefs in the Dongsha Atoll lagoon. Brown areas are living coral and white areas are sand; all are submerged. The patch reefs are 114 m in length; the research vessel, Atoll 2, in the right middle is 11 m in length, right middle is 11 m in length.
Using light detection and ranging (LiDAR), researchers have obtained bathymetric details, which have enabled the categorization of patch reefs into two types according to their morphological characteristics, i.e., pinnacle-top and flat-top patch reefs [Figure 3; (James, 1983; Brock et al., 2008)]. There has been no direct evidence of the mechanisms causing these geomorphological differences. The lagoon in Dongsha Atoll contains thousands of patch reefs (Figure 2) with diameters up to 600 m, as discovered using satellite images and LiDAR. Although patch reefs are discernible in high-quality satellite images and LiDAR, their origins have rarely been investigated (Brock et al., 2008; Shih et al., 2011). Both the aforementioned hypotheses can be used to explain the formation of the rugged bottom of the Dongsha Atoll lagoon (Figure 3). The karst and the growth mechanism both exists but each dominates in different geological periods, they emphasize the effects of uneven erosion and uneven timing of reef accretion, respectively.

Figure 3 Bathymetric LiDAR image of Dongsha Atoll. Oblique view revealing flat-top and pinnacle-top patch reefs in the Dongsha Atoll lagoon. Vertical relief is not proportional in scale to the planar surface, as indicated by colors. The maximum depth of the Dongsha Atoll lagoon is approximately −20 m.
In this study, we investigated how and where to locate long coral skeletal records in an atoll and tested hypotheses related to mechanisms underlying the formation of patch reefs. Our study focused on three main areas: first, the age patterns of the surfaces of flat-top patch reefs; second, trends in the ages of different flat-top patch reefs within a lagoon; and third, the depth distributions of flat-top and pinnacle-top patch reefs.
Materials and methods
Bathymetry
Most of the flat-top patch reefs in the Dongsha Atoll were visible in the high-resolution satellite images, whereas the pinnacle-top patch reefs were not visible because of their smaller horizontal dimensions and deeper distributions.
We investigated the bathymetry of the Dongsha Atoll by using records from the LiDAR database compiled (Shih et al., 2014). Because water clarity in the Dongsha Atoll is high, the maximum water depth of the laser scanner is <50 m, and the entire atoll was successfully mapped. The horizontal resolution of the collected images was 3.5 m, and the water depth deviation was <0.15 m (Shih et al., 2011; Shih et al., 2014; Lin et al., 2016).
The working definition of the distinction between flat-top and pinnacle-top patch reefs was that the surfaces of flat-top patch reefs must have diameters of >5 m (Brock et al., 2008).
Patch reef sampling
We collected coral core samples from the surface of patch reefs by using two types of drills, namely hydraulic and electric drills (Nemo Diver DD18-3Li-50); the samples had diameters of 10 and 3 cm and core lengths of 30 and 15 cm, respectively. We avoided live corals and removed unconsolidated materials. Before coring, in some cases, the surfaces of reefs were covered by thick layers (0.5–1 m) of loose coral fragments, which required substantial effort for their removal. After returning to the laboratory, we collected 5 g skeletal samples from the sides of the cores at 5 cm from the surface to avoid potential contamination from the environment.
We selected and drilled a flat-top patch reef of approximately 100 m in diameter at a depth of approximately −5 m on the lagoon’s southwest side (Figure 1, F12) along a horizontal transect from the presumed center to the periphery. Two batches of core samples were collected. The first batch was collected using a hydraulic drill at 8 m intervals along the selected transect in 2016. The second batch was collected using an electric drill at 1–4 m intervals along the same transect line in 2017.
We used electric and hydraulic drills to collect core samples from the centers and peripheries of the flat tops of 13 additional patch reefs (F1–F11, F13, F14), ranging in depth from −1 to −6 m. We measured the lengths of the long axes of 10 additional flat-top patch reefs (F15–F24) by using LiDAR images; we collected cores samples of the reefs using an electric drill to investigate possible correlations between the diameters and ages of various patch reefs (Figure 1).
To assess the ages of pinnacle-top patch reefs, we randomly selected 14 patch reefs (Figure 1), ranging in depth from −4 to −14 m, and sampled the surfaces of the pinnacles of the reefs, again focusing on consolidated reefs and avoiding live corals.
U-Th dating
Chipped subsamples (0.4–1.0 g each) were cut from the bulk of the core samples. X-ray diffractometric techniques showed only aragonitic composition and the nondetectable post-depositional calcite (<3%). Each subsample was gently crushed into 0.5 to 1 mm3 segments and cleaned using ultrasonic methods (Shen et al., 2008). Approximately 100–200 mg was used for U-Th dating. A triple-spike (229Th-233U-236U) isotope dilution method was used to correct mass bias and determine the U-Th concentrations and isotopic compositions (Shen et al., 2002; Shen et al., 2008) on a multicollector inductively coupled plasma mass spectrometer (MC-ICPMS), Thermo Neptune, at the High-Precision Mass Spectrometry and Environment Change Laboratory of the Department of Geosciences, National Taiwan University (Shen et al., 2012). The uncertainties of the isotopic compositions and contents were calculated at the 2σ level. U-Th half-lives used are given in Cheng et al. (2013). Conditions of 146 ± 4 for initial δ234U values (Cutler et al., 2003) and U contents of 1.9-4.0 ppm (Shen and Dunbar, 1995; Shen et al., 2008) were applied to evaluate reliable dates of pristine corals.
Spatial distributions of flat-top and pinnacle-top patch reefs
We drew 16 equally spaced gridlines at 1 km intervals in the horizontal (E–W) and vertical (N–S) directions on the bathymetric LiDAR images. We then paired and systematically sampled individual patch reefs (both pinnacle-tops and flat-top reefs) closest to each intersection point, obtaining a total of 256 pairs. We measured the depths of the surfaces of the reefs according to the LiDAR images.
Coral cover on pinnacle-top patch reefs
The pinnacle-top patch reefs were extruded 6.5 ± 2.6 m from the bottom. We surveyed the coral cover on the pinnacle and the surrounding base of each of the five pinnacle-top patch reefs (Figure 1, K1–K5) along a 25 m line transect. Coral Point Count with Excel extensions (Kohler and Gill, 2006), a software program for determining coral cover using transect photographs, was used to analyze the images. We sampled 25 random points on each square image, identified the substrate type underlying the points, and calculated coverage ratios accordingly.
Results
Age patterns on surfaces of flat-top patch reefs
The age patterns of the first batch of samples collected followed a significant trend; that is, the samples collected near the center of the reef were old, whereas those collected near the periphery of the reef were young. The regression was as follows: Age (years) = −62.4 (years/m) + 3624.7 (years) (R² = 0.90, n = 7, p < 0.001), where the presumed center was at 0 m and the periphery of the surface of this patch reef was 48 m from the center.
Given the small sample size of the preliminary data set, we collected an additional batch of samples by using electric drills with diameters of 3 cm. The small bits of the electric drills could easily fit into depressions on reefs, in which the large bits of the hydraulic drills could not fit into.
After preliminary data examination, we elected to ignore the samples younger than 250 years (n = 7) when performing regression analysis because they were likely collected from small corals growing in the crevices of older reefs. After this adjustment, the regression—Age (years) = −54.2 (year/m) + 3792.7 (years) (R² = 0.67, n = 14, p < 0.001)—was similar to that obtained using the first batch of samples based only on large cores (the 10 cm diameter samples). We performed analysis of covariance and discovered no significant differences in the slopes or intercepts of the two regressions (p = 0.51 for the two intercepts).
The oldest skeletal sample in our transect, which was 4256 ± 19 years old, was not at the presumed center of the flat top of the patch reef but was >15 m from the center. Nevertheless, none of the 10 samples collected close to the center were <2000 years old (Figure 4 and Supplementary Table 1).
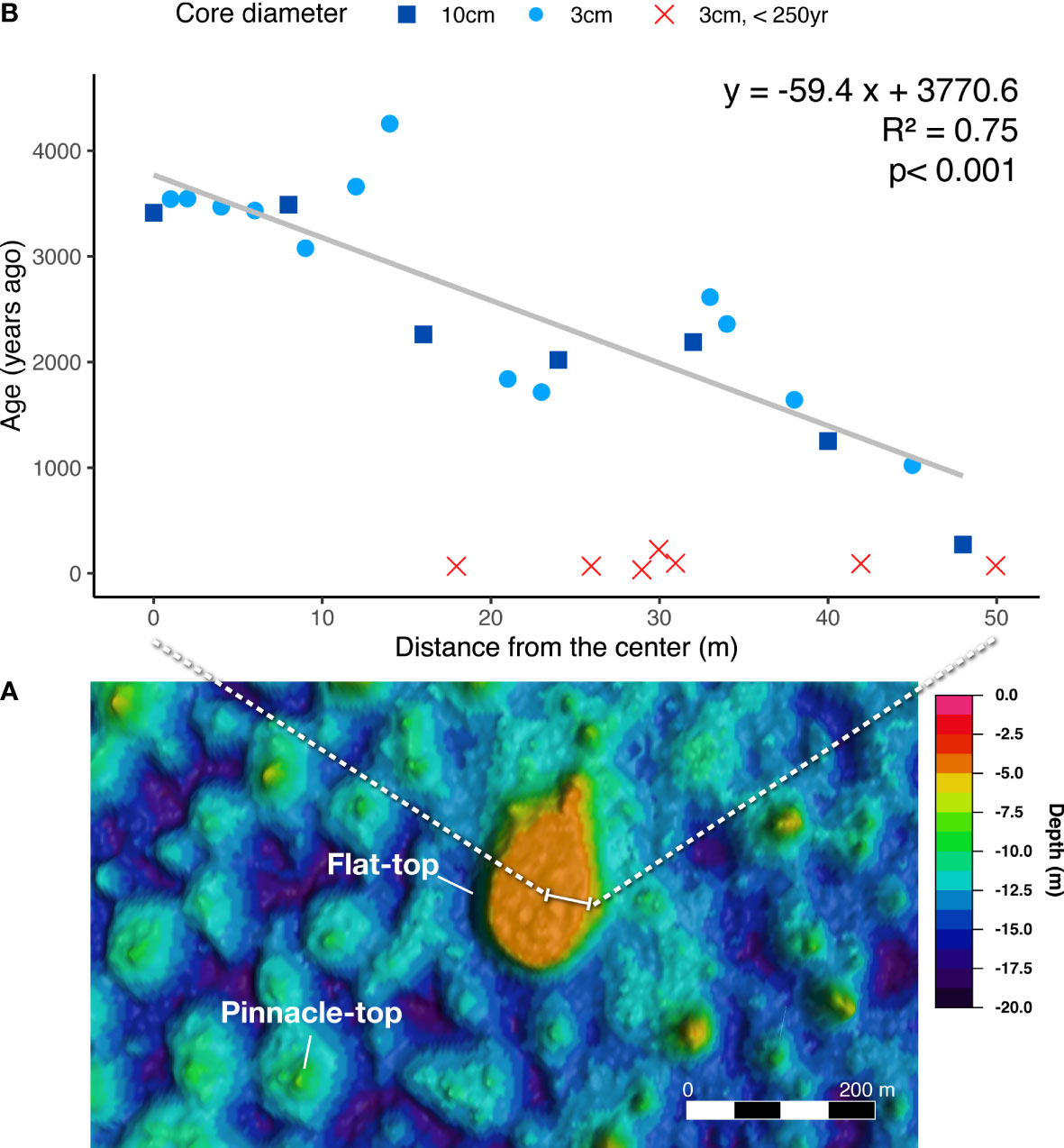
Figure 4 Age pattern on flat top of a patch reef. (A) LiDAR image of the actual sampled flat-top patch reef (F12) in the Dongsha Atoll lagoon. The top of the patch reef is −5 m deep, and the diameter of the surface is 100 m. (B) Ages of corals collected from reef surfaces from the center to the periphery of the surface of a patch reef. The linear regression of the blue spots has a slope of 59.4 years/m, equivalent to an average rate of 1.68 cm/year. The red crosses were modern or recent corals attached on the surface and were not included in the regression analysis.
Age comparison of patch reefs
Among the different flat-top patch reefs sampled, all the shallow reefs (those with surfaces at depths of 0 to −2 m) had young centers; by contrast, the ages of those with surfaces at depths of −3 to −6 m varied widely, from a few hundred to >3000 years (Figure 5 and Supplementary Table 1).

Figure 5 Comparison of the ages of the centers of surfaces of flat-top patch reefs with different depths. The 24 core samples collected from the centers of the surfaces of the flat-top patch reefs (F1–F24) were divided into two groups: 0 to −2 m (n = 9) and −3 to −6 m (n = 15). A nonparametric Kruskal–Wallis test revealed a significant difference in center age between the two groups.
The peripheries of both the deep and shallow flat-top patch reefs were all young (<300 years old). Therefore, the tree stump–like concentric circular age pattern in which the corals closer to the center of the reef were older than those at the periphery occurred only in the relatively deep flat-top patch reefs (Figure 5).
Spatial distributions of patch reefs
The tops of the flat-top patch reefs were no deeper than approximately −6 m and tended to be deeper toward the southwest end of the lagoon (Figure 6) and away from the reef rim. The maximum depth of the pinnacle-top patch reefs was −16 m close to the bottom of the lagoon. The distribution of the depths of the pinnacle tops was heterogeneous; most of the reefs with deeper tops were located toward the northeast end of the lagoon. When we compared the flat-top and pinnacle-top reefs paired by proximity (Figure 3), the surfaces of the flat-top reefs were consistently shallower than those of their neighboring pinnacle-top patch reefs (Figure 7). The shallow patch reefs (both flat-top and pinnacle-top) were distributed in the lagoon’s periphery or just behind the reef rim (Figure 6).
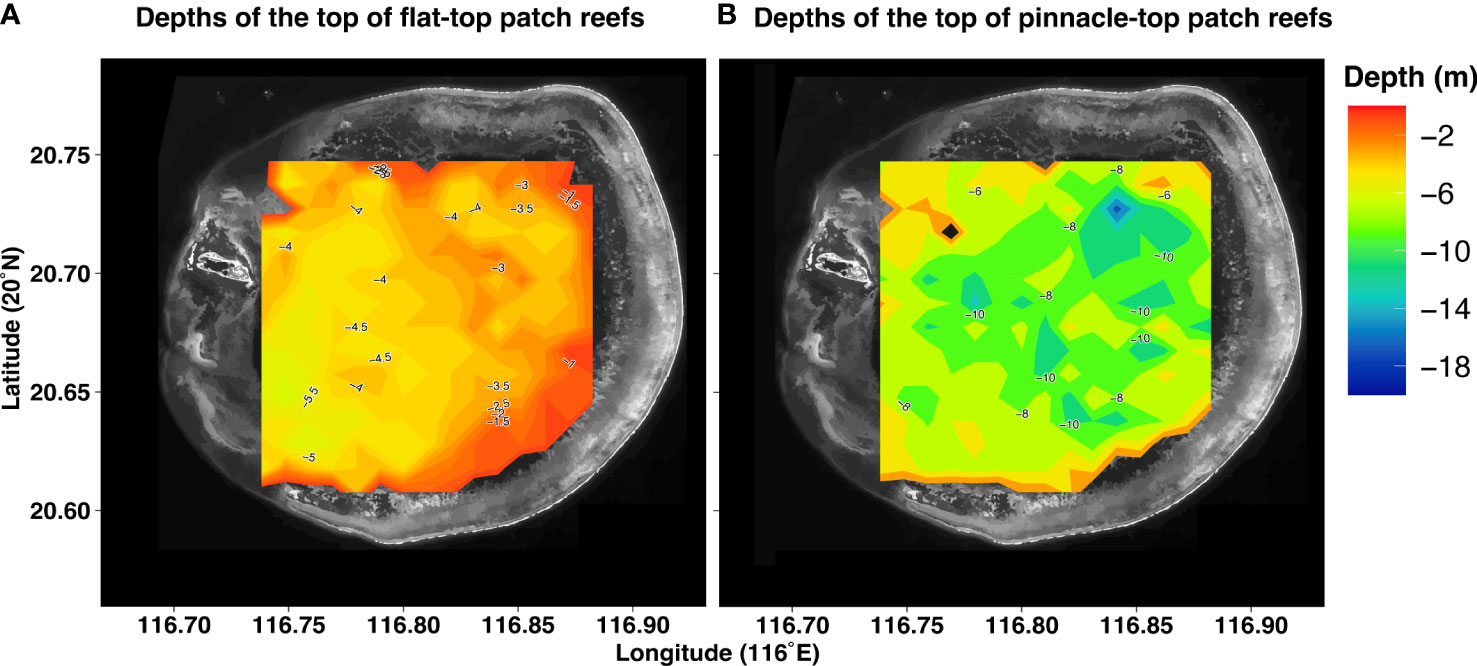
Figure 6 Contour maps of the surfaces of patch reefs in the Dongsha Atoll lagoon. Both maps are based on LiDAR images of systematically selected samples from the Dongsha Atoll lagoon. (A) Contour map of the surfaces of flat-top patch reefs (n = 208). (B) Contour map of the surfaces of pinnacle-top patch reefs (n = 206). The dark part is in grayscale of the satellite photo.
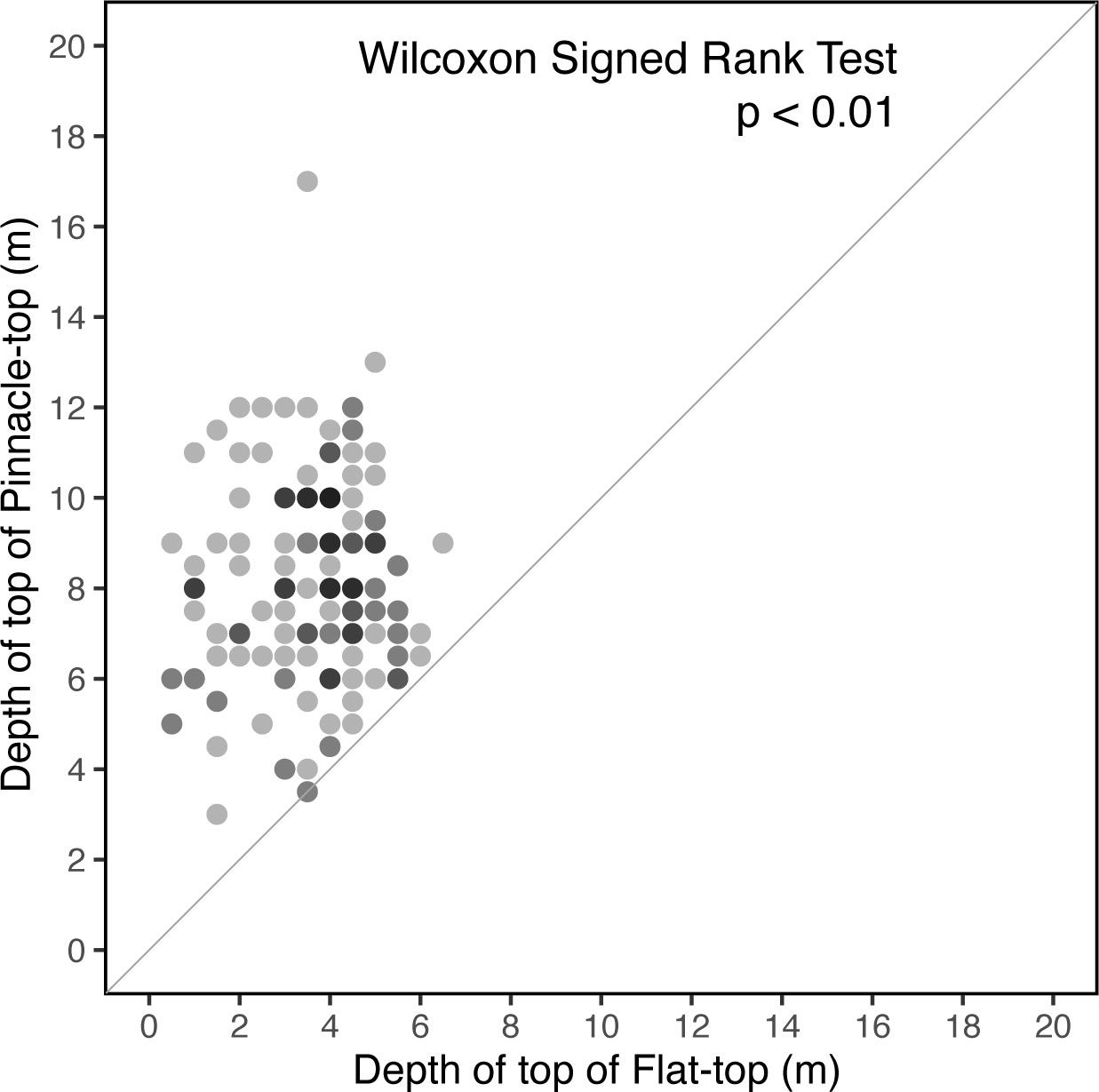
Figure 7 Paired comparison of neighboring flat-top and pinnacle-top patch reefs. None of the surfaces of the pinnacle-top reefs were shallower than those of their neighboring flat-top reefs. Darker dots indicate multiple observations.
Discussion
Growth patterns of patch reefs
Three of our main observations to explain the present form of patch reef landform in the lagoon of Dongsha atoll are consistent with the growth hypothesis. First, the differences in ages of the centers of the different flat-top patch reefs reflected the different times at which each patch reef reached the surface and started to expand horizontally, which is consistent with the growth hypothesis. Second, the age patterns within patch reefs, that is, the old centers and young peripheries on the surfaces of individual flat-top patch reefs, can readily be explained by the growth hypothesis, given that the new reef growth was distributed in the peripheries of the patch reefs. The karst hypothesis, which emphasizes the role of erosion, cannot explain the age variability among the patch reefs or the concentric age patterns within individual patch reefs (Figures 4, 8). Third, all the pinnacle-top reefs we examined had more live corals on their tips than on the surrounding bases. This phenomenon is compatible with the growth hypothesis, but not with the karst hypothesis. According to the karst hypothesis, live coral cover is expected to be similar among different parts of pinnacle-top reefs (Figure 9). However, this explanation is only discussed in terms of the present form of patch reef formation, and it is not known whether the patch reef is constructed on the Pleistocene limestone. The species compositions on the surfaces of flat-top reefs, especially on the edges of the surfaces (Figure 10), were dominated by massive poritids (Soong et al., 2009). Living poritids often form rings surrounding the peripheries of the surfaces of patch reefs and are often incorporated into the patch reefs thereafter. Most of the reef core samples examined in this study consisted of massive poritid skeletons (Figure 11). The concentric circular growth patterns of patch reefs, similar to those in the Dongsha Atoll lagoon, are believed to result from a formation process that has been ongoing for thousands of years since the stabilization of the sea level (Grigg et al., 2002; Siddall et al., 2003; Woodroffe and Horton, 2005). e.g., for reef F12, it reached its present level 4256 years ago and began to expand horizontally.
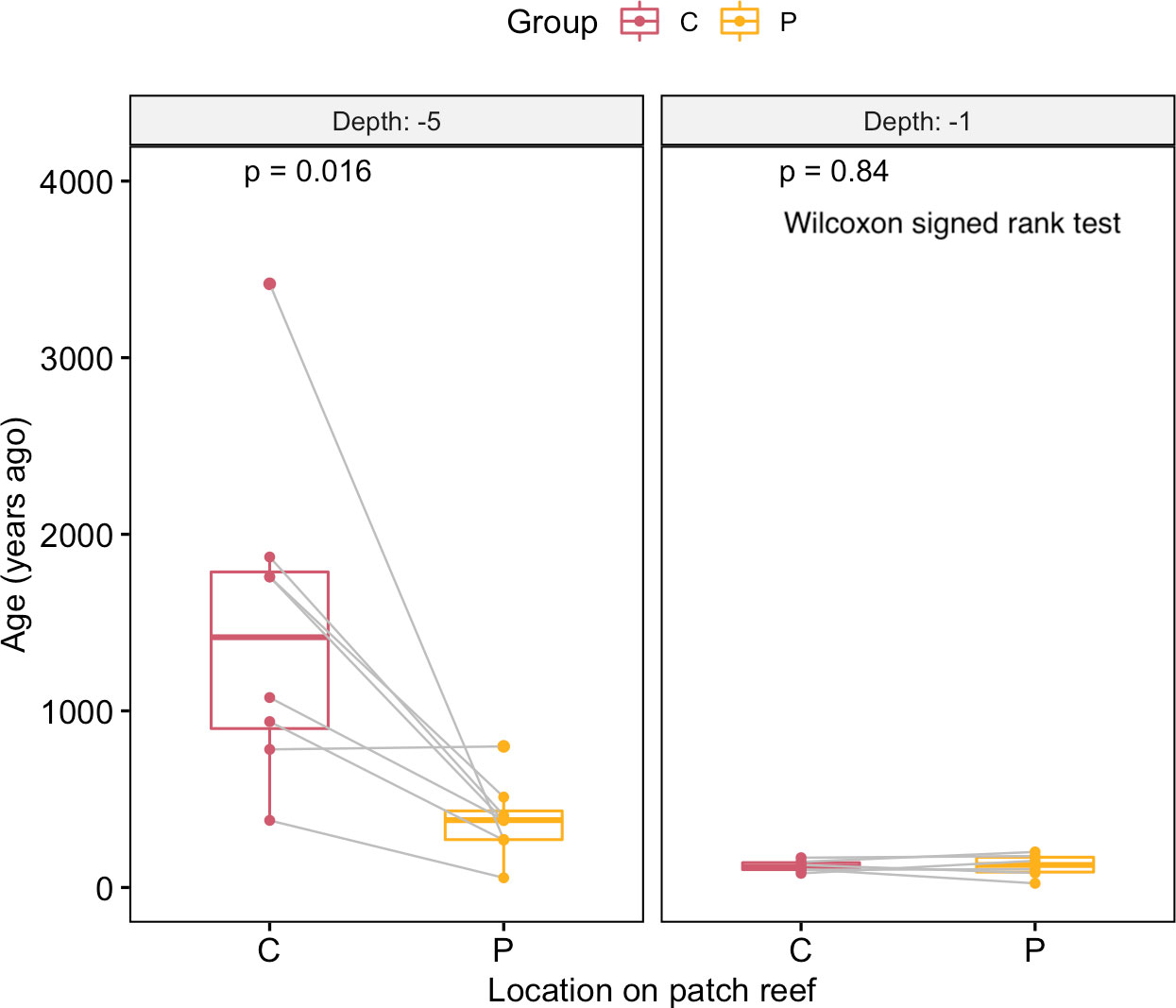
Figure 8 Age comparison of the centers and peripheries of flat-top patch reefs. The centers of the surfaces of the flat-top patch reefs at −5 m were older than their peripheries; this trend was not observed among the reefs at −1 m. A significant difference was identified between the reefs at depths of −5 m and −1 m. Paired comparisons of the ages in different parts of patch reefs in two depths, respectively. C: Center, P: Periphery, of a patch reef.
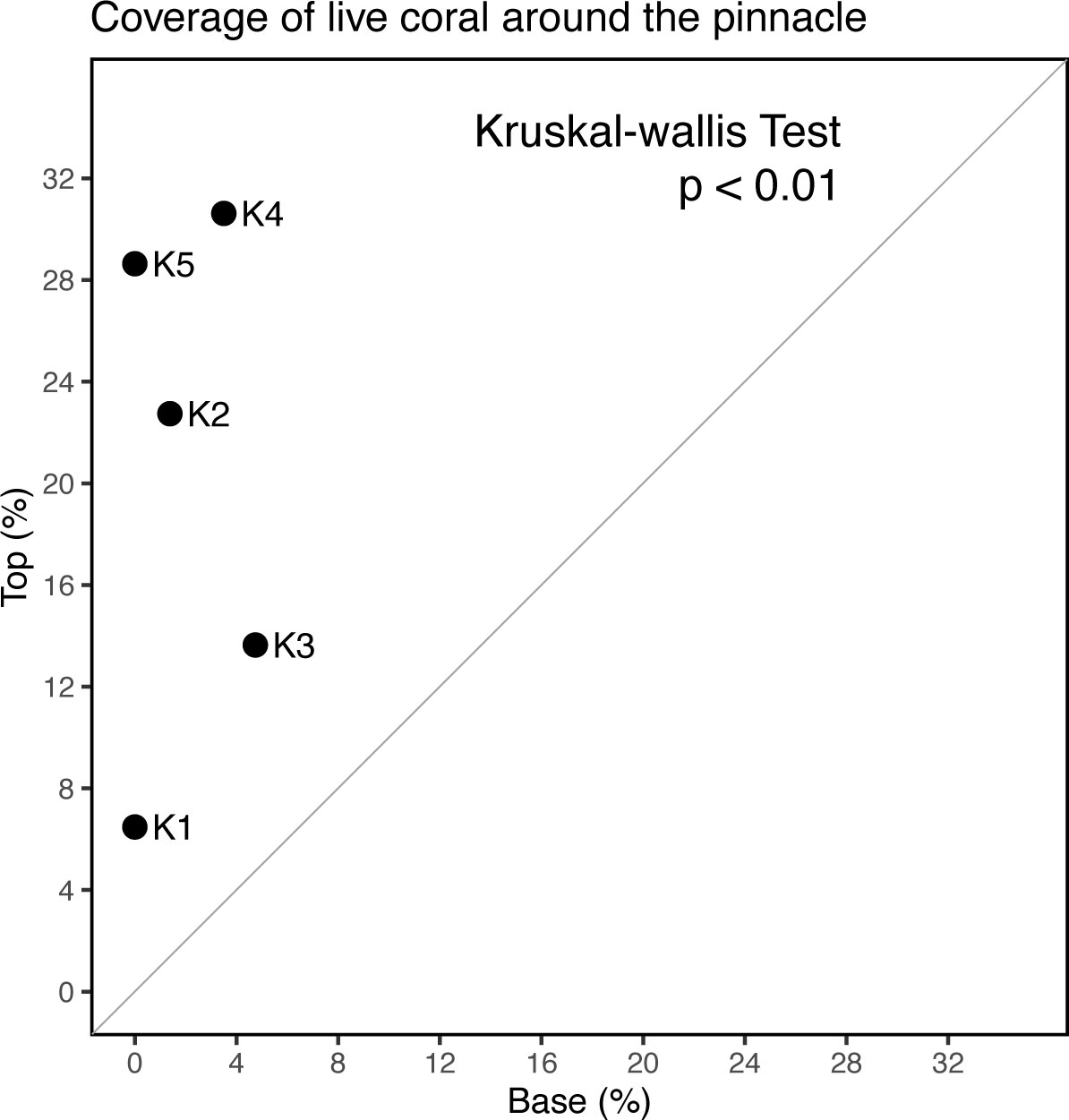
Figure 9 Comparison of live coral cover on the tops and bases of pinnacle-top patch reefs in the Dongsha Atoll lagoon. The living coral cover on the tops of the pinnacle-top patch reefs was significantly higher than that on the bases.

Figure 10 Images of flat-top patch reefs. (A) Colonies of massive coral Porites on the edges of flat-top patch reefs in the Dongsha Atoll lagoon. (B) Lack of corals in the central part of a flat-top patch reef.

Figure 11 X-ray slice image of a core sample. Core sample was from the surface of the center of a flat-top patch reef (F12-2). It was collected using a hydraulic drill with a diameter of 10 cm and a core length of 30 cm. The average age of the specimens collected from the sides of the core approximately 5 cm from the surface was 3489.2 ± 16.6 years.
Neighboring core samples along a reef transect, separated by a few meters on the surfaces of flat-top patch reefs, could differ in age by 500–1000 years. In some cases, the age pattern was even reversed; that is, the samples collected near the peripheries were older than those collected near the center (Figure 4). A few factors may have contributed to this phenomenon. First, in contrast to individual coral colonies, patch reefs do not necessarily expand homogeneously (at a similar rate) at different points around their circumferences. The spaces among large colonies may take a long time to fill because of the growth of new colonies on the reef. The bareness of some flat-top surfaces suggests that some new colonies that had been growing on these surfaces must have been translocated toward the peripheries of the patch reefs (Figure 10). A mixture of colonies, presumably of different ages, might occur because of major forces that dislodge and move coral colonies on shallow flat tops. For example, the translocation of colonies of massive poritids is reflected by the presence of loose and disoriented colonies at the peripheries and slopes of patch reefs (Hennige et al., 2017).
Second, the horizontal growth rate in the patch reef radius (1.68 cm/year, flat-top patch reef F12) over the period of >4000 years recorded in this study is faster than previously recorded vertical reef accretion rates (0.3–0.5 cm/year) (Mazzullo et al., 1992). But such patterns can be affected by the growth rate of the species and the locations of reefs (Supplementary Table 1). It is also faster than the recorded growth rate of individual corals (0.90 ± 0.04 cm/year) on the surfaces of patch reefs in the Dongsha Atoll (Soong et al., 2009). We hypothesize that both colony translocation and on-site growth may contribute to this phenomenon. Although the entire surfaces of patch reefs are suitable for coral growth, at least temporarily, coral skeletons can accumulate only at the peripheries of patch reefs. i.e., the center portion of the upper surface of patch reefs that reach sea level (low tide level) are not always suitable for coral accumulation. Because of disruptive physical events, coral colonies of various ages pushed to the peripheries of the surfaces of the reefs either fell to the slopes of the patch reefs or filled the gaps among other coral colonies. Both processes occurred as the flat-top patch reefs increased in diameter over time. Moreover, because the rate at which the area of a flat top increases is proportional to the square of its diameter, whereas the area of the periphery increases linearly with diameter, a greater supply of coral colonies is available, and these colonies can be pushed to the peripheries of the flat tops of the patch reefs. Therefore, the average expansion rate of larger flat-top patch reefs is expected to be higher than that of smaller flat-top patch reefs, which can be investigated through high-resolution sampling.
Third, when the pinnacle-top patch reefs ceased vertical expansion and began to expand horizontally, they were transformed into flat-top patch reefs (Figure 4). Because of the asymmetric expansion of flat-top patch reefs, the oldest parts of such reefs are usually not found in the center of the reefs and are therefore difficult to locate. Just as the asymmetry of tree trunks is caused by the angle of sunlight, that of patch reefs is caused by the prevailing wave and current direction. However, the merging of two pinnacle-top patch reefs in proximity as their respective diameters expanded could also have resulted in the oldest parts of the patch reefs being off-center. Distinguishing between these two potential mechanisms, namely asymmetric growth and merging, may be a valuable direction for future research. The former predicts that there is only one center of the oldest parts in a patch reef, whereas the latter predicts that there could be more than one. Presumably, asymmetric growth may occur in every flat-top patch reef under the influence of water motion (Brown and Dunne, 1980; Mazzullo et al., 1992; Huang et al., 2022), but merging is more likely the cause of the large and irregularly shapes of flat-top patch reefs. Some patch-reefs, e.g., the ones near the reef rim, or the periphery of the lagoon, have centers and peripheries of similarly young ages (Figure 12). These patch reefs have much less potential to provide long records of core. Flat-top patch reefs at different depths or locations can have different sediments on top, and coral composition. These young patch reefs along lagoon periphery were also characterized by growing closer to sea surface (Figure 12).
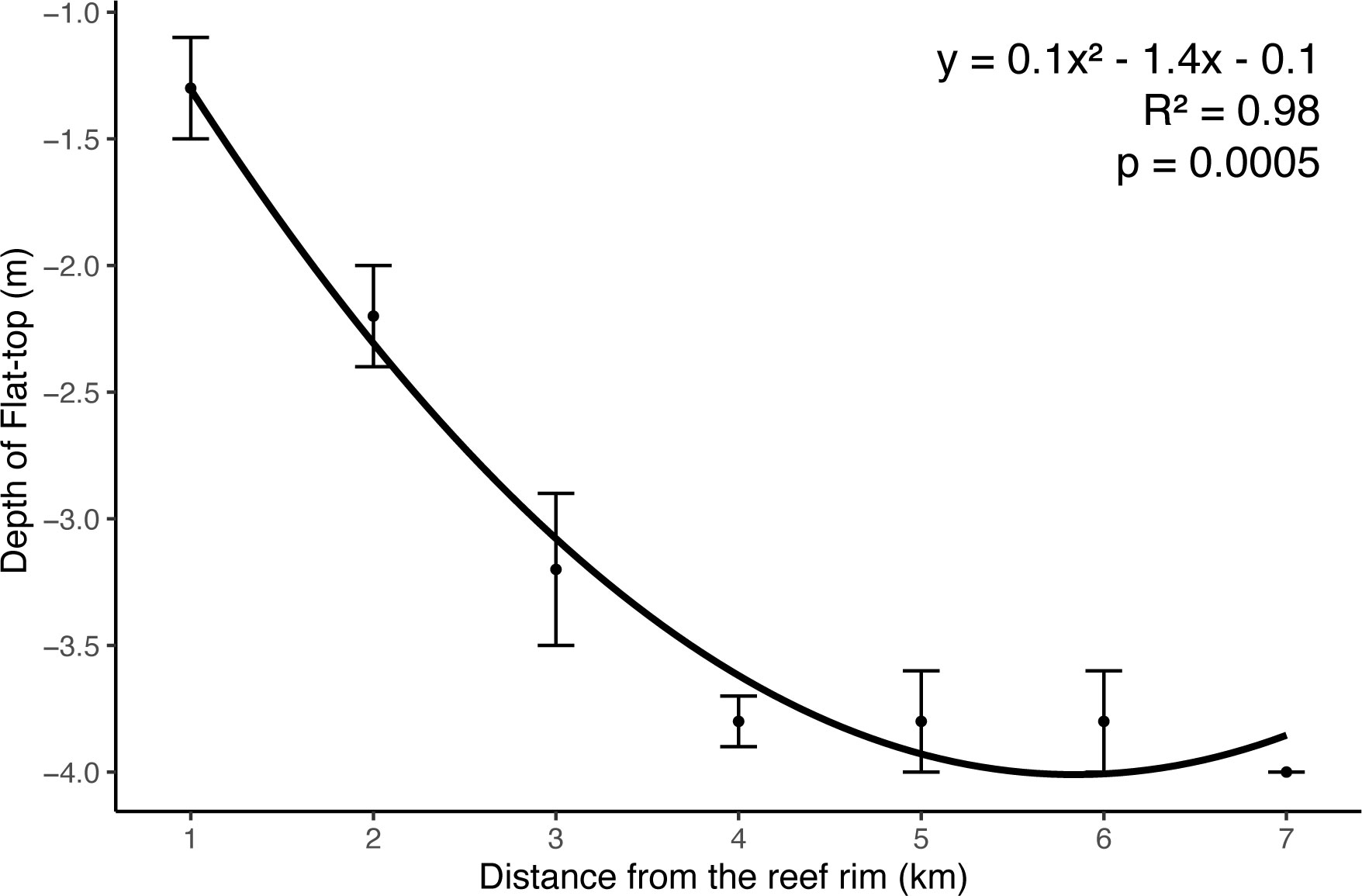
Figure 12 The quadratic regression showed the relationship between the depth of the top of the flat-top patch reefs and the distance from the reef rim.
Despite the aforementioned trends, patch reefs vary widely, as indicated by the findings of the present study. Some patch reefs, such as those near the reef rim or peripheries of the Dongsha Atoll lagoon, have centers and peripheries of similar young ages (Figure 8). These patch reefs are unlikely to serve as sources of long skeletal records. Flat-top patch reefs at different depths or locations can have different sediments on their surfaces and distinct coral compositions. The young patch reefs along the periphery of the Dongsha Atoll lagoon were distributed close to the sea surface (Figure 6). Because of the shallowness of the water near the reef rim, reefs may easily accumulate and approach the surface in the lagoon, which is protected by the reef rim from wave-induced erosion. Because of the protection provided by the reef rim, fragments of branching corals in various habitats can disperse more easily. We hypothesize that small fragments of these branching corals homogenize the coral compositions of the centers and peripheries of reefs.
We failed to identify a positive correlation between the ages of the centers and the diameters of flat-top patch reefs; that is, larger reefs did not necessarily have older centers. This unexpected result suggests that various complex factors affect the expansion and age patterns of flat-top patch reefs. First, the concentric age rings on the surface of flat-top patch reefs may not be radially symmetric. For a better chance of identifying the oldest core on a flat-top patch reef, researchers have more chance to collect complete continuous expansion changes along the long axis of the flat tops of the reefs. Alternatively, researchers could investigate the mechanisms underlying asymmetric patch reef expansion. For example, the prevailing wave direction may determine where dislodged massive colonies were relocated as well as the orientation of the long axes of flat-top patch reefs. As previously mentioned, the large sizes of the flat tops may be attributable to the merging of patch reefs rather than to reef accretion over time, as we had assumed. Methods for distinguishing between the two mechanisms with reasonable effort have yet to be developed. Some relatively small flat-top reefs were old; for example, some patch reefs with diameters of approximately 30 m were dated to almost 2000 years ago. This may be an example of “give-up” or the cessation of carbonate accumulation for a long period (Neumann and Macintyre, 1985). We could not explain how some of the patch reefs “gave up” for approximately 2000 years but others in the same lagoon did not; these observations warrant further investigation.
The topographical distinction between flat-top and pinnacle-top reefs is attributable to forces at the water surface; that is, in contrast to the vertical accretion of flat-top reefs, that of pinnacle-top reefs has yet to be constrained by surface forces. The evidence of this phenomenon is twofold. First, none of the surfaces of the pinnacle-top reefs were shallower than those of their neighboring flat-top reefs (Figure 7). This strongly suggests that an ongoing mechanism causes growing pinnacle-top patch reefs to become flat-topped as they approach the sea surface. The reef expanded horizontally after being restricted. Second, according to a previous report, waves near the entrance were greater than those in the inner lagoon (Wang and Hung, 2009). Additionally, a study in Okinawa revealed that the energy of most waves dissipates in the outer forereefs during typhoons (Vila-Concejo et al., 2017). This may explain why the flat tops near the entrance of the atoll lagoon were deep, whereas those near the reef rim were shallower (Figure 6). We speculate that protection of the reef rim resulted in a lower average wave height, allowing more vertical space for patch reef growth.
Patch reefs as sources of long and continuous skeletal records of horizontal reef expansion
The old skeletons on the surfaces of flat-top patch reefs are easily accessible with handheld electric drills; more importantly, their ages follow a pattern. In this study, we determined that the location, size, shape, depth, and other characteristics of coral reefs may be related to the ages of the reefs. When seeking out long, horizontally-oriented coral skeletal records, researchers must seek out patch reefs that started expanding horizontally a long time ago, which may have occurred for two reasons. First, the reefs started horizontal expansion early, for example, by starting in shallow water but far from the reef rim. Alternatively, the reefs were restricted and forced to expand horizontally in deeper waters, for example, at −5 to −6 m.
According to our analysis, the patch reefs in the Dongsha Atoll were forced to expand horizontally, given that the centers of surfaces of the patch reefs near the reef rim, in relatively shallow water, which average was only 133 ± 48 years old. In the Dongsha Atoll, the depths of the surfaces of the flat-top reefs ranged from −1 to −6 m, and a difference of 5 m in vertical reef accretion could correspond to an age difference of 500–5000 years if reefs grow vertically at 1–10 mm/year. Researchers seeking out flat-top reefs in a lagoon should focus their efforts on deep waters.
In summary, flat-top patch reefs may serve as adequate sources of easily accessible long coral skeletal records. Sections of corals in such reefs can easily be subsampled for high-resolution analysis of temporal changes within each year of skeletal growth (Figure 11). After U-Th dating, various cores can be arranged chronologically, with each core sample available for high-resolution, within-decade, and within-year studies, similar to skeletal cores collected from live coral colonies (Ren et al., 2017). Determining where to drill to obtain skeletal samples of intermediate age is crucial.
Conclusion
Three of our main observations to explain the present form of patch reef landform in the lagoon of Dongsha atoll are consistent with the growth hypothesis. Pinnacle-top reefs expand horizontally after being constrained by sea surface action and are thus transformed into flat-top reefs. Moreover, flat-top patch reefs may serve as adequate sources of easily accessible long coral skeletal records.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
YW designed the study, performed fieldwork, contributed to data collection and analysis, and wrote the manuscript. KS designed the study and contributed to the data interpretation. C-CS was responsible for U-Th data analysis. All authors contributed to the article and approved the submitted version.
Funding
Funding was provided by the Ministry of Science and Technology of Taiwan grants #107-21-2119-M-110-009 awarded to KS, the Science Vanguard Research Program of the Ministry of Science and Technology #110-2123-M-002-009 awarded to C-CS, the National Taiwan University #109L8926 awarded to C-CS, and the Higher Education Sprout Project of the Ministry of Education #110L901001 and 110L8907 awarded to C-CS.
Acknowledgments
We are deeply indebted to the Dongsha Atoll Research Station, Marine National Park, the Coast Guard Administration of Dongsha, Kuo-Fang Huang in Academia Sinica, who provided core X-ray slice images, and field workers who were essential in collection, and the assistants who analyzed the U-Th dating data in the High-precision mass spectrometry and environment change laboratory (HISPEC), National Taiwan University. Without their effort, this study would not have been possible. We also wish to express our sincere gratitude to Tian-Yuan Shih of National Yang Ming Chiao Tung University for assistance in LiDAR images, and Shou-Yeh Gong of the National Museum of Natural Science for comments on an early draft of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2022.994966/full#supplementary-material
References
Aronson R. B., Precht W. F., Macintyre I. G. (1998). Extrinsic control of species replacement on a Holocene reef in Belize: The role of coral disease. Coral Reefs 17 (3), 223–230. doi: 10.1007/s003380050122
Brock J. C., Palaseanu-Lovejoy M., Wright C. W., Nayegandhi A. (2008). Patch-reef morphology as a proxy for Holocene sea-level variability, northern Florida keys, USA. Coral Reefs 27 (3), 555–568. doi: 10.1007/s00338-008-0370-y
Brown B. E., Dunne R. P. (1980). Environmental controls of patch-reef growth and development. Mar. Biol. 56 (1), 85–96. doi: 10.1007/BF00390598
Cheng H., Edwards R. L., Shen C.-C., Polyak V. J., Asmerom Y., Woodhead J., et al. (2013). Improvements in 230Th dating, 230Th and 234U half-life values, and U–Th isotopic measurements by multi-collector inductively coupled plasma mass spectrometry. Earth Planetary Sci. Lett. 371, 82–91. doi: 10.1016/j.epsl.2013.04.006
Cutler K. B., Edwards R. L., Taylor F. W., Cheng H., Adkins J., Gallup C. D., et al. (2003). Rapid sea-level fall and deep-ocean temperature change since the last interglacial period. Earth Planetary Sci. Lett. 206 (3-4), 253–271. doi: 10.1016/S0012-821X(02)01107-X
Davis W. M. (1928). “The coral reef problem,” in Special publication no. 9 (New York: American Geographical Society), 596.
Dickinson W. R. (2009). Pacific atoll living: How long already and until when. GSA Today 19 (3), 4–10. doi: 10.1130/GSATG35A.1
Droxler A. W., Jorry S. J. (2021). The origin of modern atolls: Challenging darwin's deeply ingrained theory. Annu. Rev. Mar. Sci. 13, 537–573. doi: 10.1146/annurev-marine-122414-034137
Duce S., Vila-Concejo A., Hamylton S., Bruce E., Webster J. M. (2014). Spur and groove distribution, morphology and relationship to relative wave exposure, southern great barrier reef, Australia. J. Coast. Res. 70 (10070), 115–120. doi: 10.2112/SI70-020.1
Gischler E., Hudson J. H. (2004). Holocene Development of the Belize barrier reef. Sedimentary Geology 164 (3-4), 223–236. doi: 10.1016/j.sedgeo.2003.10.006
Goldberg W. M. (2016). Atolls of the world: Revisiting the original checklist. Atoll Res. Bull. 610, 1–47. doi: 10.5479/si.0077-5630.610
Grigg R. W., Grossman E. E., Earle S. A., Gittings S. R., Lott D., McDonough J. (2002). Drowned reefs and antecedent karst topography, au'au channel, S.E. Hawaiian islands. Coral Reefs 21 (1), 73–82. doi: 10.1007/s00338-001-0203-8
Guozhong W. (1998). “Tectonic and monsoonal controls on coral atolls in the south China Sea,” in Reefs and carbonate platforms in the pacific and Indian oceans. Eds. Camoin G. F., Davies P. J. (United Kingdom: Blackwell Science), 237–248.
Halley R. B., Shinn E. A., Hudson J. H., Lidz B. (1977). Recent and relict topography of boo bee patch reef, Belize. Proc. Third Int. Coral Reef Symposium 2, 29–35.
Hennige S. J., Burdett H. L., Perna G., Tudhope A. W., Kamenos N. A. (2017). The potential for coral reef establishment through free-living stabilization. Sci. Rep. 7 (1), 1–7. doi: 10.1038/s41598-017-13668-7
Huang Y., Hu X., Zhang Y. (2022). Distribution and morphological features of the patch reefs at xiaonanxun reef, zhenghe atoll of the nansha islands, south China Sea. Mar. Geology Quaternary Geology (Chinese) 42 (3), 25–35. doi: 10.16562/j.cnki.0256-1492.2021082201
James N. P. (1983). “Reef environment,” in Carbonate depositional environments. Eds. Scholle P. A., Bebout D. G., Moore C. H. (USA: American Association of Petroleum Geologists), 345–462.
Kan H., Urata K., Nagao M., Hori N., Fujita K., Yokoyama Y., et al. (2015). Submerged karst landforms observed by multibeam bathymetric survey in nagura bay, ishigaki island, southwestern Japan. Geomorphology 229, 112–124. doi: 10.1016/j.geomorph.2014.07.032
Kohler K. E., Gill S. M. (2006). Coral point count with excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. geosciences 32 (9), 1259–1269. doi: 10.1016/j.cageo.2005.11.009
Lin W.-T., Shih P. T.-Y., Jie-Chung C., Liao C.-J. (2016). Bathymetric LiDAR green channel derived reflectance: An experiment from the dongsha 2010 mission. Terrestrial Atmospheric Oceanic Sci. 27 (4), 565. doi: 10.3319/TAO.2016.02.19.02(ISRS
Mazzullo S. J., Anderson-Underwood K. E., Burke C. D., Bischoff W. D. (1992). Holocene Coral patch reef ecology and sedimentary architecture, northern Belize, central America. Palaios 7:591–601. doi: 10.2307/3514871
Neumann A. C., Macintyre I. G. (1985). “Reef response to sea-level rise: keep-up, catch-up, or give-up,” in Proceedings of the 5th international coral reef congress (Tahiti: Antenne Museum-EPHE), 105–110.
Purdy E. G. (1970). Reef configurations: Some causes and effects. Am. Assoc. Petroleum Geologists Bull. 54 (5), 865–866. doi: 10.1306/5D25CB29-16C1-11D7-8645000102C1865D
Purdy E. G., Gischler E. (2005). The transient nature of the empty bucket model of reef sedimentation. Sedimentary Geology 175 (1-4), 35–47. doi: 10.1016/j.sedgeo.2005.01.007
Purdy E. G., Winterer E. L. (2001). Origin of atoll lagoons. Geological Soc. America Bull. 113 (7), 837–854. doi: 10.1130/0016-7606(2001)113<0837:Ooal>2.0.Co;2
Ren H. J., Chen Y. C., Wang X. T., Wong G. T. F., Cohen A. L., DeCarlo T. M., et al. (2017). 21st-century rise in anthropogenic nitrogen deposition on a remote coral reef. Science 356 (6339), 749–752. doi: 10.1126/science.aal3869
Shen G. T., Dunbar R. B. (1995). Environmental controls on uranium in reef corals. Geochimica Cosmochimica Acta 59 (10), 2009–2024. doi: 10.1016/0016-7037(95)00123-9
Shen C.-C., Edwards R. L., Cheng H., Dorale J. A., Thomas R. B., Moran S. B., et al. (2002). Uranium and thorium isotopic and concentration measurements by magnetic sector inductively coupled plasma mass spectrometry. Chem. Geology 185 (3-4), 165–178. doi: 10.1016/S0009-2541(01)00404-1
Shen C.-C., Li K.-S., Sieh K., Natawidjaja D., Cheng H., Wang X., et al. (2008). Variation of initial 230Th/232Th and limits of high precision U–Th dating of shallow-water corals. Geochimica Cosmochimica Acta 72 (17), 4201–4223. doi: 10.1016/j.gca.2008.06.011
Shen C.-C., Wu C.-C., Cheng H., Edwards R. L., Hsieh Y.-T., Gallet S., et al. (2012). High-precision and high-resolution carbonate 230Th dating by MC-ICP-MS with SEM protocols. Geochimica Cosmochimica Acta 99, 71–86. doi: 10.1016/j.gca.2012.09.018
Shih P. T.-Y., Arumugam D., Shyue S. W. (2011). Bathymetric lidar survey of penghu islands and dongsha atoll. Sea Technol. 52 (11), 42–45.
Shih P. T.-Y., Chen Y. H., Chen J. C. (2014). Historic shipwreck study in dongsha atoll with bathymetric LiDAR. Archaeological Prospection 21 (2), 139–146. doi: 10.1002/arp.1466
Siddall M., Rohling E. J., Almogi-Labin A., Hemleben C., Meischner D., Schmelzer I., et al. (2003). Sea-Level fluctuations during the last glacial cycle. Nat. (London) 423 (6942), 853–858. doi: 10.1038/nature01690
Soong K., Wang J.-T., Chen C. A., Nozawa Y. (2009). Investigative analysis and restorative tactic on coral reef of dongsha atoll (I) (Taiwan: Marine National Park Headquarters).
Tomascik T., Mah J. A., Nontji A., Moosa K. M. (1997). The ecology of the Indonesian seas part 2. Hong Kong: Periplus Editions.
Vila-Concejo A., Duce S., Nagao M., Nakashima Y., Ito M., Fujita K., et al. (2017). Typhoon waves on coral reefs. Coast. Dynamics 2017 263, 697–701. doi: 10.1038/ncomms8854
Wang Y.-H., Hung J.-J. (2009). Long-term investigation of the marine environment of dongsha atoll national park (I) (Taiwan: Marine National Park Headquarters).
Woodroffe C. D., Biribo N. (2011). “Atolls,” in Encyclopedia of modern coral reefs. encyclopedia of earth sciences series. Ed. Hopley D. (Dordrecht, Netherlands: Springer).
Keywords: patch reef, atoll, horizontal expansion, long skeletal records, U-Th dating, sea-level rise
Citation: Wei Y, Shen C-C and Soong K (2022) Patch reefs as sources of long and continuous skeletal records of horizontal reef expansion in Dongsha Atoll, South China Sea. Front. Mar. Sci. 9:994966. doi: 10.3389/fmars.2022.994966
Received: 15 July 2022; Accepted: 05 October 2022;
Published: 27 October 2022.
Edited by:
Wei Jiang, Guangxi University, ChinaCopyright © 2022 Wei, Shen and Soong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Keryea Soong, keryea@g-mail.nsysu.edu.tw