
Chemistry and Biological Activity of Ramalina Lichenized Fungi

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
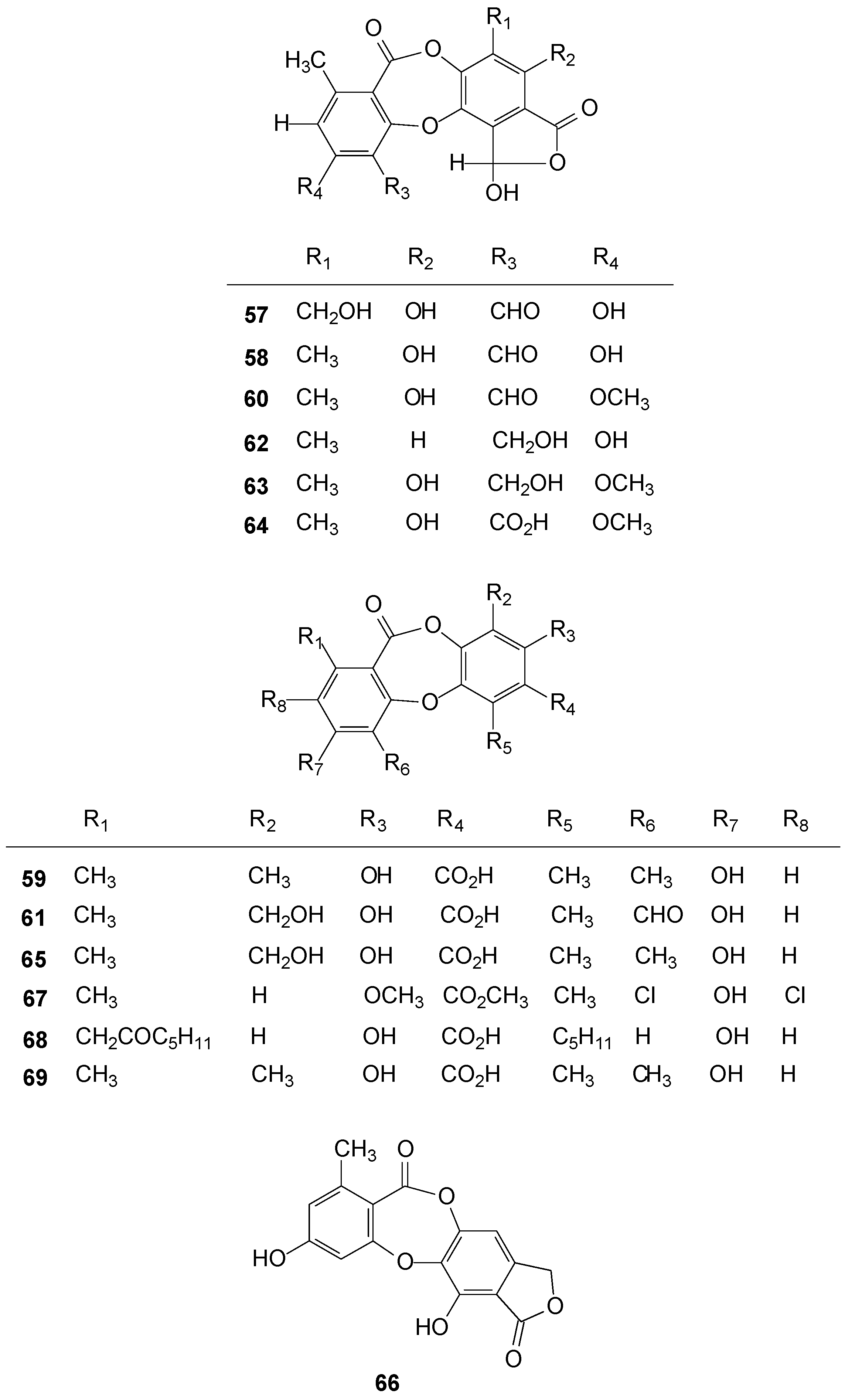{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Chemical Constituents
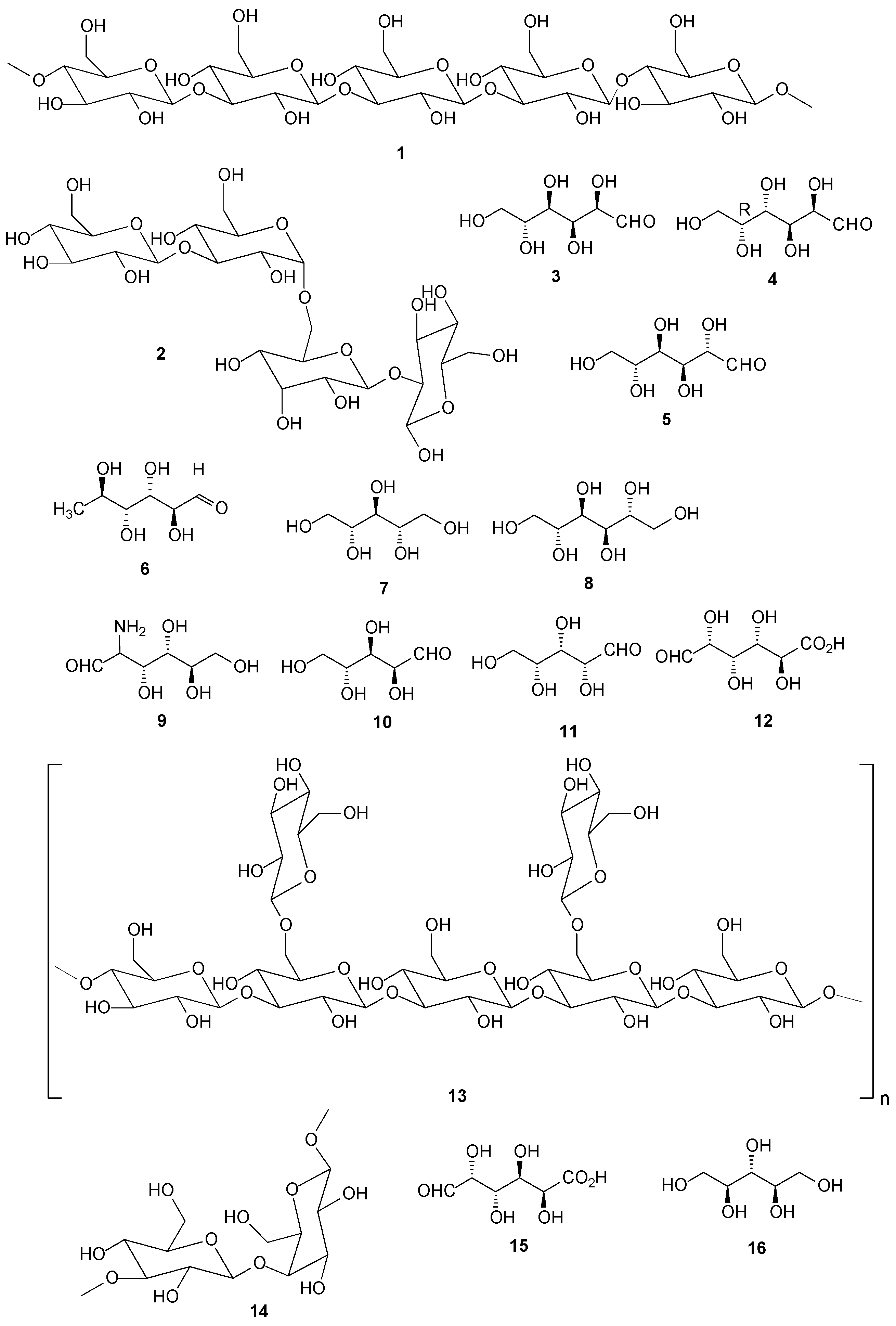
2.1. Carbohydrates

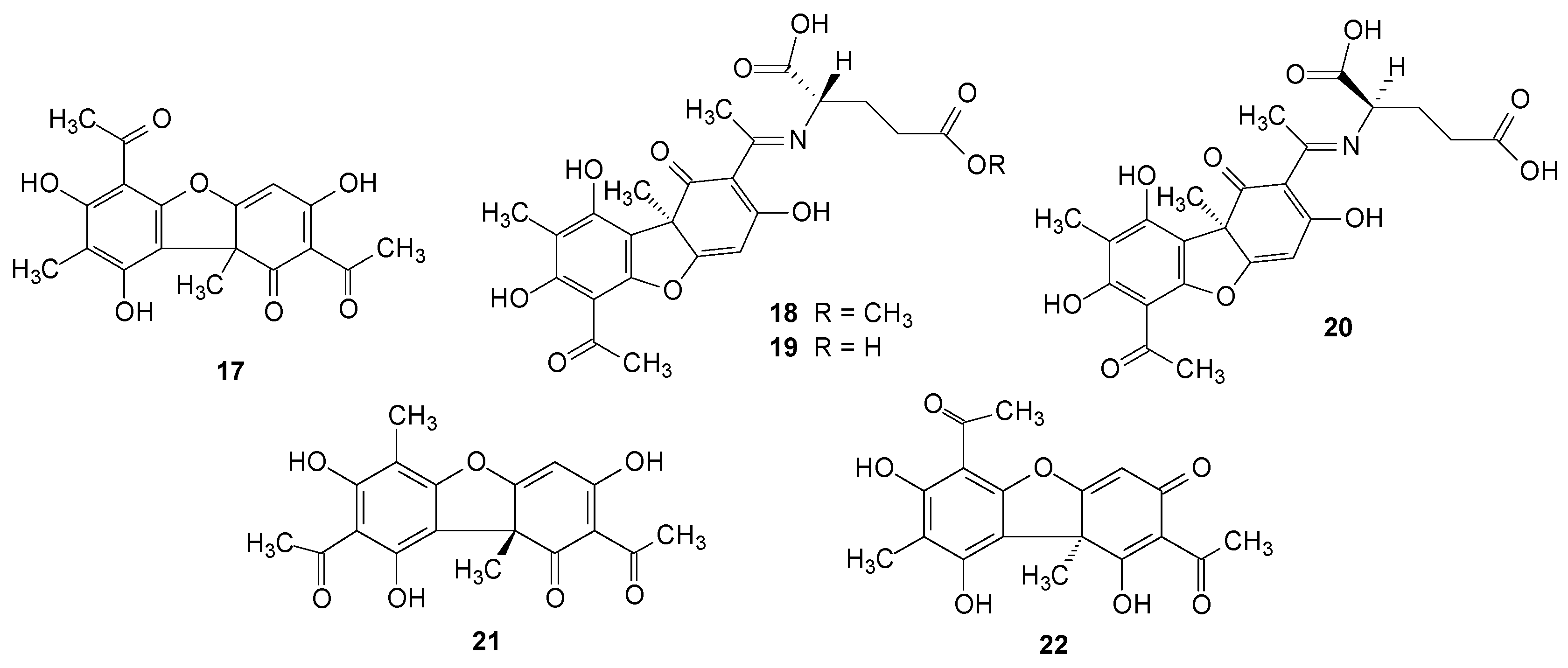
2.2. Usnic Acid and Derivatives

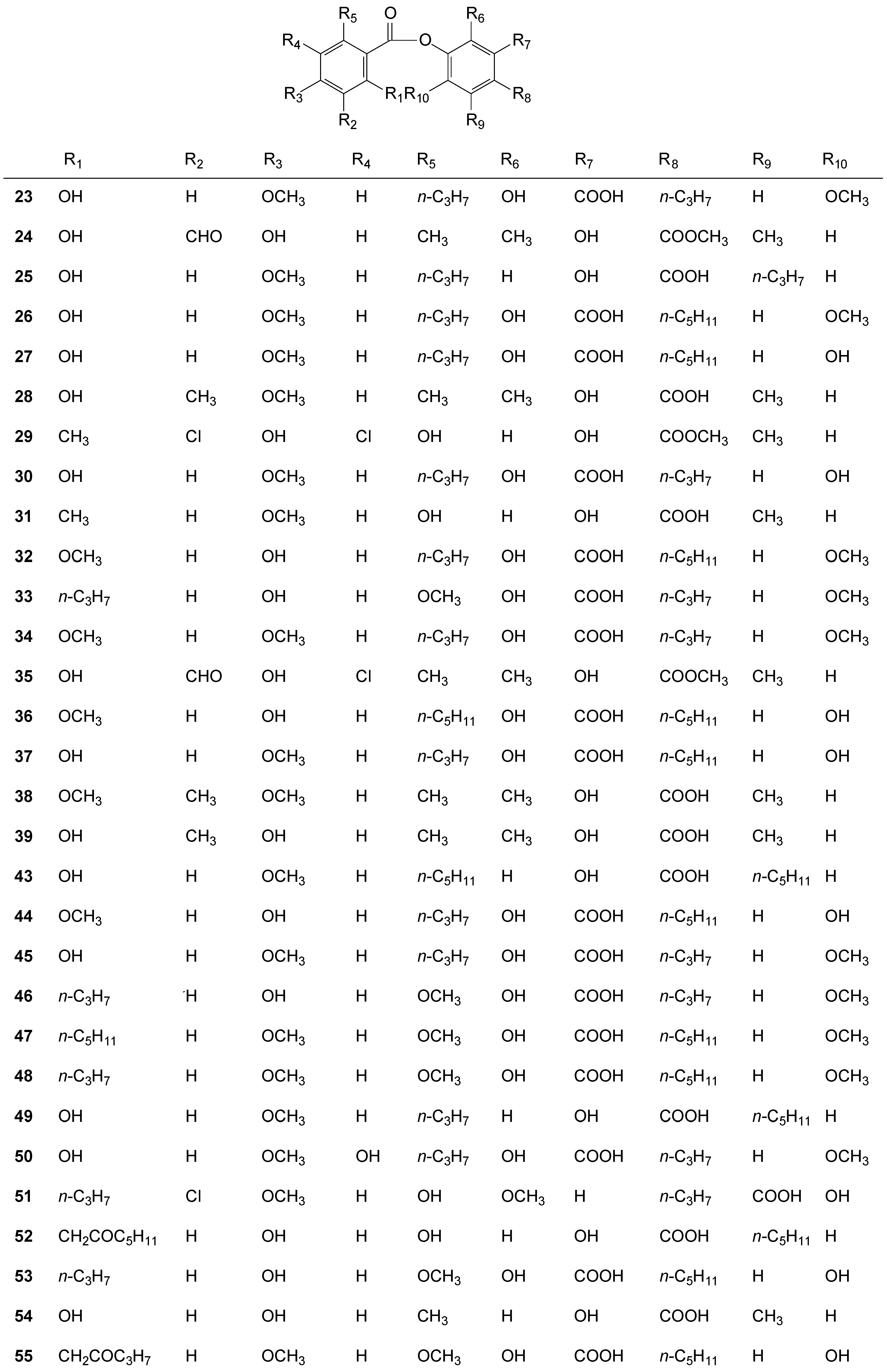
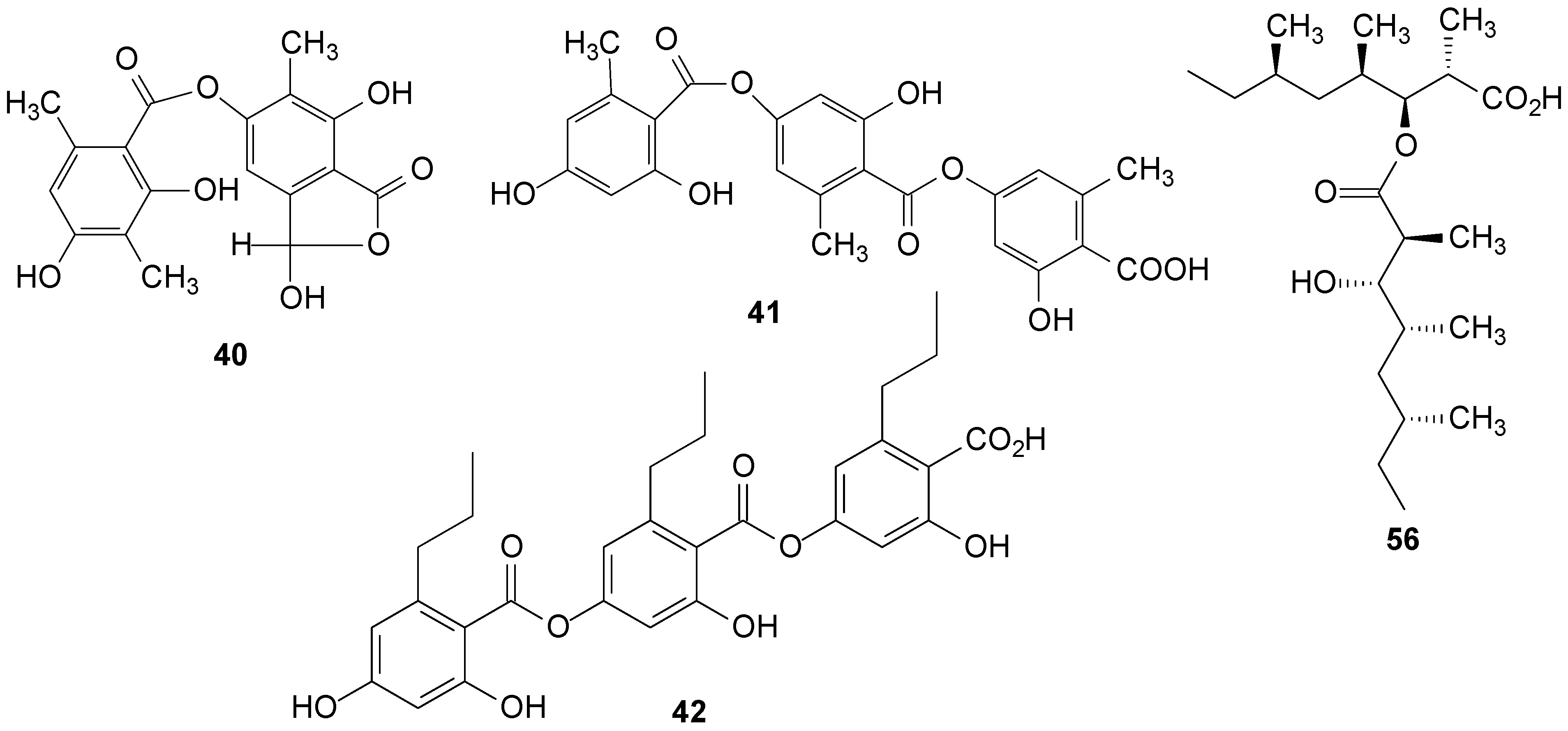
2.3. Depsides


2.4. Depsidones
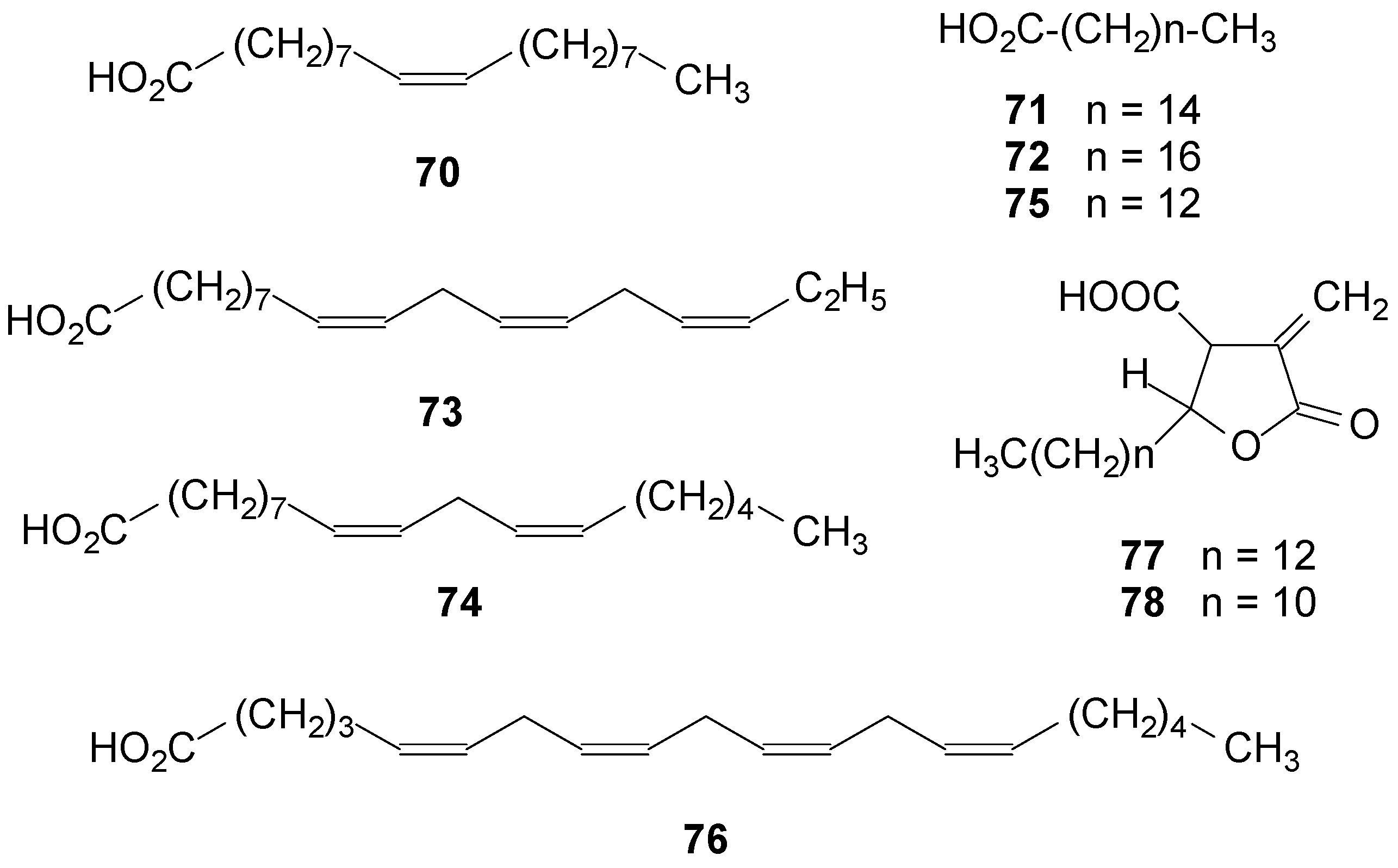
2.5. Fatty Acids


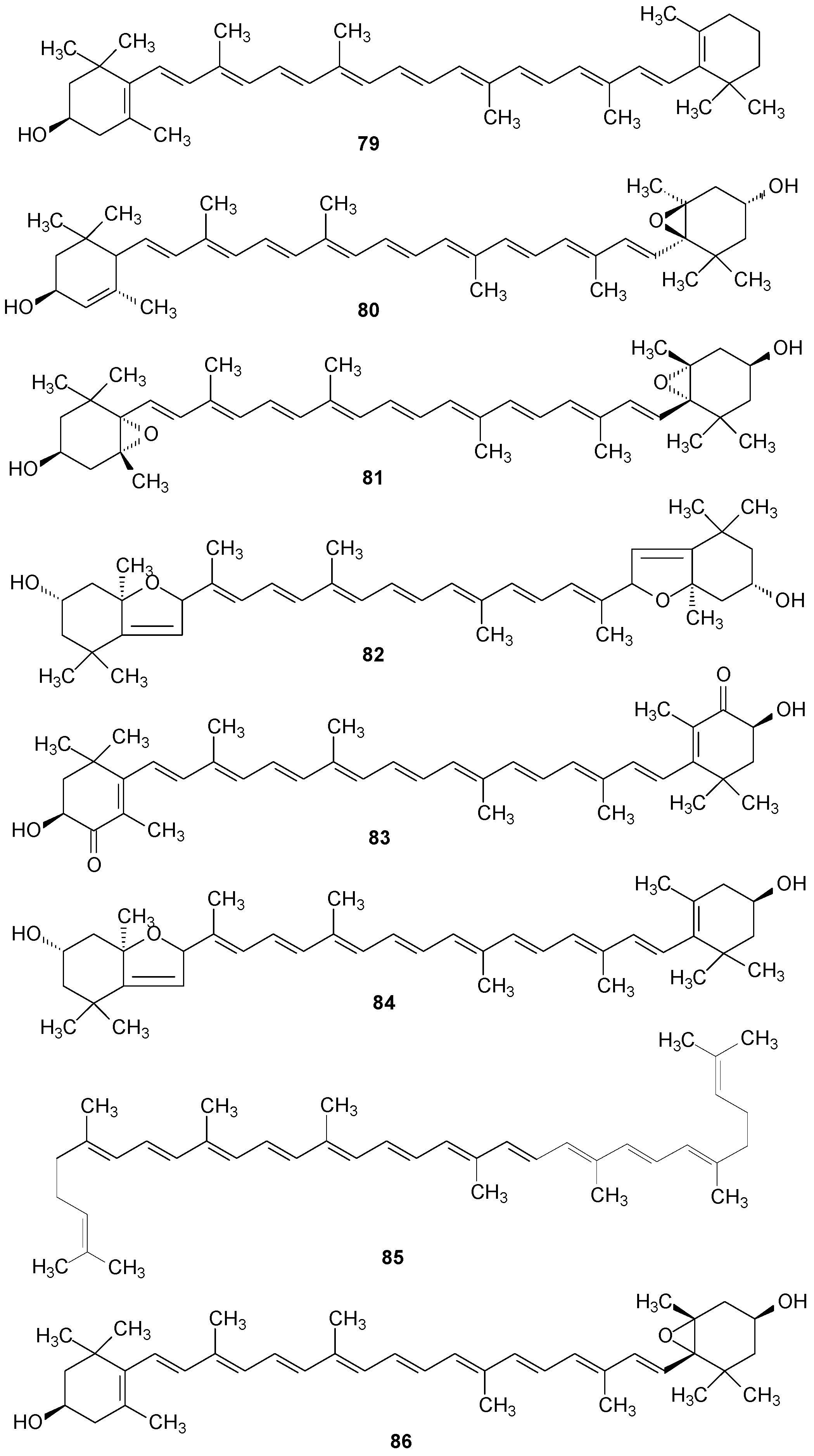
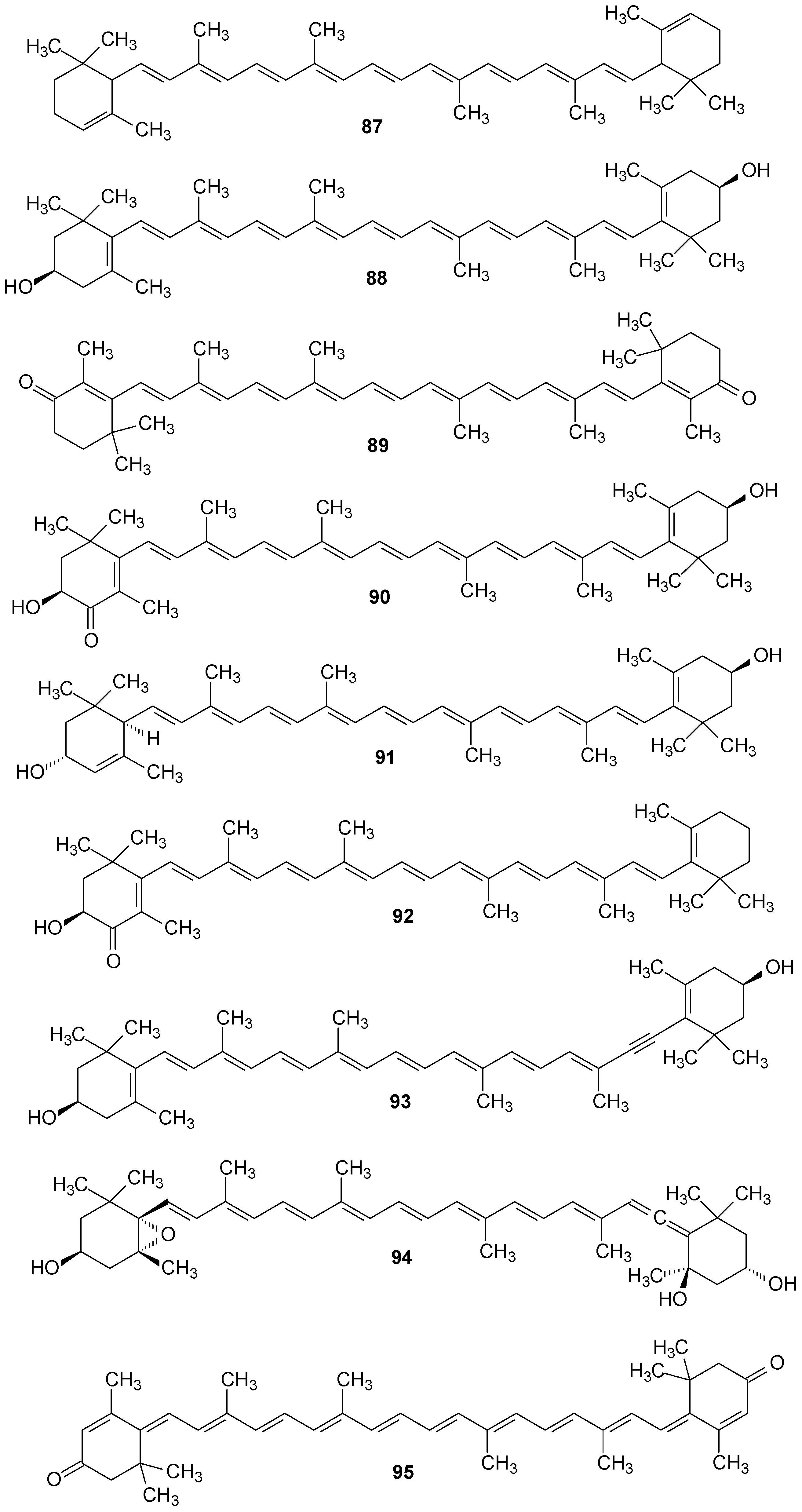
2.6. Other Compounds






3. Biological Activity
3.1. Antimicrobial Activity
3.2. Antioxidant Activity
3.4. Antitumor and Cytotoxic Activity
3.5. Anti-Inflammatory Activity
3.6. Other Activities
4. Concluding Remarks
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Huneck, S.; Yoshimura, I. Identification of Lichen Substances; Springer-Verlag: Berlin, Germany, 1996. [Google Scholar]
- Boustie, J.; Grube, M. Lichens—A promising source of bioactive secondary metabolites. Plant Genet. Resour. 2005, 3, 273–287. [Google Scholar] [CrossRef]
- Honda, N.K.; Vilegas, W. A química dos liquens. Quim. Nova 1998, 21, 110–125. [Google Scholar]
- Feige, G.B.; Lumbsch, H.T. Identification of lichen substances by a standardized high-performance liquid chromatographic method. J. Chromatogr. 1993, 647, 417–427. [Google Scholar] [CrossRef]
- Parrot, D.; Jan, S.; Baert, N.; Guyot, S.; Tomasi, S. Comparative metabolite profiling and chemical study of Ramalina siliquosa complex using LC-ESI-MS/MS approach. Phytochemistry 2013, 89, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, I. Ecological and biotechnological aspects of lichens. Appl. Microbiol. Biotechnol. 2006, 73, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi; CABI—Europe-UK, Cromwell Press: Trowbridge, UK, 2008. [Google Scholar]
- Lee, S.G.; Koh, H.Y.; Oh, H.; Han, S.J.; Kim, I.C.; Lee, H.K.; Yim, J.H. Human dermal fibroblast proliferation activity of usimine-C from Antarctic lichen Ramalina terebrata. Biotechnol. Lett. 2010, 32, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Paudel, B.; Bhattarai, H.D.; Koh, H.Y.; Lee, S.G.; Han, S.J.; Lee, H.K.; Oh, H.; Shin, H.W.; Yim, J.H. Ramalin, a novel nontoxic antioxidant compound from the Antarctic lichen Ramalina terebrata. Phytomedicine 2011, 18, 1285–1290. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.H.; Lee, H.K.; Bhattarai, H.D.; Babita, P.; Kim, I.C.; Hong, S.G.; Kim, D.K.; Lee, Y.K.; Gu, S.; Oh, H.C. Antioxidant Activities of Ramalin Isolated from Ramalina terebrata Extracts. WO 2,010,053,327 A2, 14 May 2010. [Google Scholar]
- Yim, J.H.; Kim, I.C.; Lee, S.G.; Kim, D.K.; Han, S.J.; Lee, H.S.; Seok, K.H.; Kim, S.J.; Kim, T.K.; Kang, P.S.; et al. Pharmaceutical Composition for the Treatment of Inflammatory Disease or Immune Disease Containing Ramalin. WO 2,011,152,671 A2, 8 December 2011. [Google Scholar]
- Gonzalez, A.G.; Barrera, J.B.; Rodriguez, E.M.P.; Padron, C.E.H. Chemical constituents of the lichen Ramalina hierrensis. Planta Med. 1992, 58, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Asahina, Y.; Fukuziro, F. Lichen substances.III. Constitution of obtusatic acid. Ber. Dtsch. Chem. Ges. B 1932, 65B, 580–583. [Google Scholar] [CrossRef]
- Miceno, A.M.; Gorin, P.A.J.; Iacomini, M. Galactomannan and isolichenan componentes of the carbohydrate-rich lichen Ramalina ecklonii (Spreng.) Mey. & Flot. Agric. Biol. Chem. 1991, 55, 1391–1392. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, A.Z.A.; Iacomini, M.; Gorin, A.J. An unusual glucomannan from Tornabenia intricata. Phytochemistry 1992, 31, 3467–3470. [Google Scholar] [CrossRef] [PubMed]
- Stuelp, P.M.; Carneiro Leão, A.M.A.; Gorin, P.A.J.; Iacomini, M. The glucans of Ramalina celastri; relation with chemotypes of other lichens. Carbohydr. Polym. 1999, 40, 101–106. [Google Scholar] [CrossRef]
- Cordeiro, L.M.C.; Reis, R.A.; Tisher, C.A.; Gorin, P.A.J.; Ferreira, J.C.; Iacomini, M. Linear β-mannose containig polysaccharide, β-xylan, and amylose from the cultured photobiont Trebouxia sp. of the ascolichen Ramalina celastri. FEMS Microbiol. Lett. 2003, 220, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, L.M.C.; Stocker-Wörgötter, E.; Gorin, P.A.J.; Iacomini, M. Comparative studies of the polysaccharides from species of the genus Ramalina-lichenized fungi-of three distinct habitats. Phytochemistry 2003, 63, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, L.M.C.; Stocker-Wörgötter, E.; Gorin, P.A.J.; Iacomini, M. Elucidation of polysaccharide origin in Ramalina peruviana symbiosis. FEMS Microbiol. Lett. 2004, 238, 79–84. [Google Scholar] [PubMed]
- Cordeiro, L.M.C.; Carbonero, E.R.; Sassaki, G.L.; Reis, R.A.; Stocker-Wörgötter, E.; Gorin, P.A.J.; Iacomini, M. A fungus-type β-galactofuranan in the cultivated Trebouxia photobiont of the lichen Ramalina gracilis. FEMS Microbiol. Lett. 2005, 244, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Gorin, P.A.J.; Iacomini, M. Polysaccharides of the lichens Cetraria islandica and Ramalina usnea. Carbohydr. Res. 1984, 128, 119–132. [Google Scholar] [CrossRef]
- Takahashi, K.; Takeda, T.; Shibata, S. Polysaccharides of lichen symbionts. Chem. Pharm. Bull. 1979, 27, 238–241. [Google Scholar] [CrossRef]
- Mamut, R.; Tursun, T.; Abbas, A. Analysis monosaccharide of the polysaccharide in Ramalina sinensis with HPLC. Shipin Gongye 2013, 34, 209–211. [Google Scholar]
- Kosugi, M.; Miyake, H.; Yamakawa, H.; Shibata, Y.; Miyazawa, A.; Sugimura, T.; Satoh, K.; Itoh, S.; Kashino, Y. Arabitol provided by lichenous fungi enhances ability to dissipate excess light energy in a symbiotic green alga under desiccation. Plant Cell Physiol. 2013, 54, 1316–1325. [Google Scholar] [CrossRef] [PubMed]
- Krzacek, T.; Smolarz, H. Chemical composition of Ramalina fraxinea (L.) Ach. Thallus. Acta Soc. Bot. Pol. 1979, 48, 27–34. [Google Scholar] [CrossRef]
- Stark, J.B.; Walter, E.D.; Owens, H.S. Method of isolation of usnic acid from Ramalina reticulata. J. Am. Chem. Soc. 1950, 72, 1819–1820. [Google Scholar] [CrossRef]
- Mittal, O.P.; Neelakantan, S.; Seshaadri, T.R. Chemical investigation of Indian lichens. XIV. Chemical components of Ramalina calicaris and Ramalina sinensis. J. Sci. Ind. Res. India 1952, 11B, 386–387. [Google Scholar]
- Culberson, C.F. Some constituents of the lichen Ramalina siliquosa. Phytochemistry 1965, 4, 951–961. [Google Scholar] [CrossRef]
- Seshadri, T.R.; Subramanian, S.S. Indian lichens. VIII. Lichens growing on sandal trees. Proc.-Indian Acad. Sci. Sect. A 1949, 30A, 15–22. [Google Scholar]
- Ashina, Y.; Yanagita, M. Lichen substances. XXXIX. A new lichen acid, norstictinic acid, and the occurrence of d-arabitol in lichens. Ber. Dtsch. Chem. Ges. B 1934, 67B, 799–803. [Google Scholar] [CrossRef]
- Cordeiro, L.M.C.; de Oliveira, S.M.; Buchi, D.F.; Iacomini, M. Galactofuranose-rich heteropolysaccharide from Trebouxia sp., photobiont of the lichen Ramalina gracilis and its effect on macrophage activation. Int. J. Biol. Macromol. 2008, 42, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Gorin, P.A.J.; Iacomini, M. Structural diversity of d-galacto-d-mannan components isolated from lichens having ascomycetous mycobionts. Carbohydr. Res. 1985, 142, 253–267. [Google Scholar]
- Teixeira, A.Z.A.; Iacomini, M.; Gorin, P.A.J. Chemotypes of mannose-containing polysaccharides of lichens mycobionts: A possible aid in classification and identification. Carbohydr. Res. 1995, 266, 309–314. [Google Scholar] [CrossRef]
- Stuelp-Campelo, P.Ma.; de Oliveira, M.B.M.; Leão, A.M.A.C.; Carbonero, E.R.; Gorin, P.A.J.; Iacomini, M. Effect of a soluble α-d-glucan from the lichenized fungus Ramalina celastri on macrophage activity. Int. Immunopharmacol. 2002, 2, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, L.M.C.; Messias, D.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Does apossymbiotically cultivated fungus Ramalina produce isolichenan? FEMS Microbiol. Lett. 2011, 321, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Peveling, E. Occurrence of starch in Chlorophyceae phycobionts. Planta 1970, 93, 82–85. [Google Scholar] [CrossRef] [PubMed]
- McAvoy, K.; Igartuburu, J.M.; Romgni-Colvin, J. Polysaccharide composition of Ramalina terebrata. Abstr. Pap. Am. Chem. Soc. 2012, 1, 25–29. [Google Scholar]
- Komiya, T.; Shibata, S. Polyols produced by the cultured phyco- and mycobionts of some Ramalina species. Phytochemistry 1971, 10, 695–699. [Google Scholar] [CrossRef]
- Culberson, W.L. Analysis of chemical and morphological variation in the Ramalina siliquosa species complex. Brittonia 1967, 19, 333–352. [Google Scholar] [CrossRef]
- Molho, L.; Bodo, B.; Molho, D. L’acide O-méthyl-4′ norsékikaïque, nouveau metadepside isole d’um lichen du genre Ramalina. Phytochemistry 1979, 18, 2049–2051. [Google Scholar] [CrossRef]
- Curd, F.H.; Robertson, A. Lichen acids. Part VI. Constituensts of Ramalina scopulorum. J. Chem. Soc. 1935, 329, 1379–1381. [Google Scholar] [CrossRef]
- Tay, T.; Türk, A.Ö.; Yilmaz, M.; Türk, H.; Kivanç, M. Evaluation of the antimicrobial activity of the acetone extract of the lichen Ramalina farinacea and its (+)-usnic acid, norstitic acid, and protocetraric acid constituents. Z. Naturforschung C 2004, 59, 384–388. [Google Scholar]
- Imshaug, H.A. Typification of Ramalina usnea (L.) R. H. Howe. Lichenologist 1972, 5, 317–318. [Google Scholar] [CrossRef]
- Shukla, V.; Negi, S.; Rawat, M.S.M.; Pant, G.; Nagatsu, A. Chemical study of Ramalina africana (Ramalinaceae) from the Garhwal Himalayas. Biochem. Syst. Ecol. 2004, 32, 449–453. [Google Scholar] [CrossRef]
- Esimone, C.O.; Adikwu, M.U. Antimicrobial activity and cytotoxicity of Ramalina farinacea. Fitoterapia 1999, 70, 428–431. [Google Scholar] [CrossRef]
- Hanus, L.O.; Temina, M.; Dembitsky, V. Biodiversity of the chemical constituents in the epiphytic lichenized ascomycete Ramalina lacera grown on difference substrates Crataegus sinaicus, Pinus halepensis, and Quercus calliprinos. Biomed. Pap. 2008, 152, 203–208. [Google Scholar] [CrossRef]
- Bézivin, C.; Tomasi, S.; Lohézic-Le, F.; Boustie, J. Cytotoxic activity of some lichen extracts on murine and human cancer cell lines. Phytomedicine 2003, 10, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Fazio, A.T.; Adler, M.T.; Bertoni, M.D.; Sepúlveda, C.S.; Damonte, E.B.; Maier, M.S. Lichen secondary metabolites from the cultured lichen mycobionts of Teloschistes chrysophthalmus and Ramalina celastri and their antiviral activities. Z. Naturforschung C 2007, 62, 543–549. [Google Scholar]
- Hauck, M.; Willenbruch, K.; Leuschner, C. Lichen substances prevent lichens from nutrient deficiency. J. Chem. Ecol. 2009, 35, 71–73. [Google Scholar] [CrossRef] [PubMed]
- Solberg, Y.J. Dyeing of wool with lichens and lichen substances. Acta Chem. Scand. 1956, 10, 1116–1123. [Google Scholar] [CrossRef]
- Verma, N.; Behera, B.C.; Joshi, A. Studies on nutritional requirement for the culture of lichen Ramalina nervulosa and Ramalina pacifica to enhance the production of antioxidant metabolites. Folia Microbiol. 2012, 57, 107–114. [Google Scholar] [CrossRef]
- Cansaran, D.; Atakol, O.; Halici, M.G.; Aksoy, A. HPLC analysis of usnic acid in some Ramalina species from Anatolia and investigation of their antimicrobial activities. Pharm. Biol. 2007, 45, 77–81. [Google Scholar] [CrossRef]
- Ingólfsdóttir, K. Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Mizuguchi, R.; Yamada, Y. Tissue cultures of Usnea rubescens and Ramalina yasudae and production of usnic acid in their cultures. Agric. Biol. Chem. 1985, 49, 3347–3348. [Google Scholar] [CrossRef]
- Culberson, C.F. Stenosporic acid, a new depside in Ramalina stenospora. Phytochemistry 1970, 9, 841–844. [Google Scholar] [CrossRef]
- Komiya, T.; Shibata, S. Formation of lichen substances by mycobionts of lichens. Isolation of (+)-usnic acid and salazinic acid from mybobionts of Ramalina sp. Chem. Pharm. Bull. 1969, 17, 1305–1306. [Google Scholar] [CrossRef]
- Bendz, G.; Santesson, J.; Wachtmeister, C.A. Studies on the chemistry of lichens. Acta Chem. Scand. 1965, 19, 1185–1187. [Google Scholar] [CrossRef]
- Urdapilleta, A.A.A. Atividade de Substâncias Isoladas de Líquens Sobre Formas Promastigotas de Leishmania (L.) amazonensis e Leishmania (V.) braziliensis. Master’s Thesis, Universidade de Brasília, Brasilia-DF, 3 March 2006. [Google Scholar]
- Aptroot, A.; Bungartz, F. The lichen genus Ramalina on the Galapagos. Lichenologist 2007, 39, 519–542. [Google Scholar]
- Kashiwadani, H.; Kalb, K. The genus Ramalina in Brazil. Lichenologist 1993, 25, 1–31. [Google Scholar] [CrossRef]
- Rundel, P.W. Evolution relationships in the Ramalina usnea complex. Lichenologist 1978, 10, 141–156. [Google Scholar]
- Lai, D.; Odimewu, D.; Esimone, C.; Grunwald, T.; Proksch, P. Phenolic compounds with in vitro activity against respiratory syncytial from the Nigerian lichen Ramalina farinacea. Planta Med. 2013, 79, 1440–1446. [Google Scholar] [CrossRef] [PubMed]
- Huneck, S.; Troted, G. Lichen constituents. XXXIV. Structure and composition of Ramalina boulhautiana. Z. Naturforschung B 1966, 21, 904–904. [Google Scholar]
- Huneck, S.; Troted, G. Lichen constituents. XX. Chemistry of Chilean lichens. 8. Componets of Ramalina chilensis. Z. Naturforschung B 1966, 21, 90–91. [Google Scholar]
- Asahina, Y.; Nonomura, S. Lichen substances. XVI. Constituents of Ramalina species with special reference to sekikaic acid. Ber. Dtsch. Chem. Ges. B 1933, 66B, 30–35. [Google Scholar] [CrossRef]
- Kumar, K.C.S.; Banskota, A.H.; Manandhar, M.D. Chemical constituents and antibacterial activity of Ramalina subcomplanata. J. Nepal Chem. Soc. 1995, 14, 28–35. [Google Scholar]
- Morozova, O.V.; Dargaeva, T.D.; Brutko, L.I. Components of Ramalina pollinaria. Khim. Prirod. Soed. 1984, 20, 252–253. [Google Scholar]
- Rao, P.S.; Shripathy, V. Chemical constituents of Umbilicaria indica Frey and Ramalina farinacea L. Ach. Curr. Sci. 1976, 45, 517–518. [Google Scholar]
- Gonzalez, A.G.; Martin, J.D.; Martin, M.M. Chemical components of lichens. II. Ramalina tingitana. Anal. Quim. 1973, 69, 807–809. [Google Scholar]
- Hawksworth, D.L. Typification of lichen farinaceous. Bryologist 1969, 72, 254–255. [Google Scholar] [CrossRef]
- Popiolek, Z. Content of lichen substances in some species of Ramalina. Ann. Univ. Mariae Curie–Sklodowska Sect. C 1967, 22, 165–168. [Google Scholar]
- Culberson, C.F. Chemical constituents of Ramalina paludosa. Bryologist 1967, 70, 397–405. [Google Scholar] [CrossRef]
- Huneck, S.; Follmann, G. Lichen constituents. XXXVI. Chemistry of Chilean lichens. 15. Constituents of Ramalina cactacearum, var. ambigua, and Medusulina chilena. Z. Naturforschung B 1967, 22, 110–111. [Google Scholar]
- Huneck, S.; Follmann, G. The presence of usnic acid in Lecanora melanophthalma and Ramalina terebrata. Naturwissenschaften 1964, 51, 291–292. [Google Scholar] [CrossRef]
- Briner, G.P.; Gream, G.E.; Riggs, N.V. Chemistry of Australian lichens. I. Some constituents of Parmelia conspersa, Ramalina fraxinea, Usnea barbata, and Usnea ceratina from the New England region. Aust. J. Chem. 1960, 13, 277–284. [Google Scholar] [CrossRef]
- Asahina, Y.; Kusaka, T. Lichen substances. LXXXIII. A new depside, boninic acid; synthesis of boninic acid and of homosekikaic acid. Ber. Dtsch. Chem. Ges. B 1937, 70B, 1815–1821. [Google Scholar] [CrossRef]
- Nakao, M. Chemical constituents of a Chinese drug “shi-hoa” and their constitutions. Yakugaku Zasshi 1923, 496, 423–497. [Google Scholar]
- Ryan, H.; O’Riordan, W.M. Tinctorial constituents of some lichens, which are used as dyes in Irland. Proc. R. Irish Acad. B 1917, 33, 91–104. [Google Scholar]
- Salkowski, H. Circular polarization of usninic acids and other lichen substances. Liebigs Ann. Chem. 1911, 377, 123–126. [Google Scholar] [CrossRef]
- Zopf, W. A contribution to the knowledge of the lichen constituents. Liebigs Ann. Chem. 1907, 352, 1–44. [Google Scholar] [CrossRef]
- Zopf, W. To the knowledge of the lichen material. Liebigs Ann. 1897, 297, 271–312. [Google Scholar] [CrossRef]
- Tugunay, T.; Reyim, M.; Adilijiang, A.A. Content determination and antioxidant activity evaluation of usnic acid in Ramalina sinensis Jatta. Shipin Kexue 2010, 31, 150–152. [Google Scholar]
- Reyes, Q.A.; Vicuna, L.P.; Almonacid, V.J. Usnic acid and atranorin in some regional lichens. Rev. Latinoam. Quim. 1981, 12, 130–131. [Google Scholar]
- Wade, A.E. The Genus Ramalina in the British Isles. Lichenologist 1961, 5, 226–241. [Google Scholar] [CrossRef]
- Marshak, A.G. Extracting Usnic Acid. US 2,557,987 A, 26 June 1951. [Google Scholar]
- Zopf, W. To the knowledge of the lichen substances. Liebigs Ann. Chem. 1905, 340, 276–309. [Google Scholar] [CrossRef]
- Cordeiro, L.M.C.; Iacomini, M.; Stocker-Wörgötter, E. Culture studies and secondary compounds of six Ramalina species. Mycol. Res. 2004, 108, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Asahina, Y.; Shibata, S. Chemistry of Lichen Substances, A; Asher & Co. Ltd: Vaals–Amsterdam, The Netherlands, 1971. [Google Scholar]
- Hamada, N. Individual variation in the contents of lichen substances in Ramalina nervulosa and Ramalina subcomplanata. Bryologist 1988, 91, 95–97. [Google Scholar] [CrossRef]
- Molho, D.; Bodo, B.; Culberson, W.L. A chemically distinctive new Ramalina from Fiji. Bryologist 1981, 84, 396–398. [Google Scholar] [CrossRef]
- Rangaswami, S.; Rao, V.S. Chemical components of Ramalina farinacea. Indian J. Pharm. 1954, 16, 197–199. [Google Scholar]
- Sisodia, R.; Geol, M.; Verma, S.; Rani, A.; Dureja, P. Antibacterial and antioxidante activity of lichen species Ramalina roesleri. Nat. Prod. Res. 2013, 27, 2235–2239. [Google Scholar] [CrossRef] [PubMed]
- Vinayaka, K.S.; Kekuda, T.R.P.; Swathi, D.; Praveen Kumar, S.V. Studies on chemical composition and in vitro antibacterial activity of solvent extracts of the lichen Ramalina hossei Vain. BioTechnol. Indian J. 2009, 3, 309–311. [Google Scholar]
- Praven Kumar, S.V.; Kekuda, T.R.P.; Vinayaka, K.S.; Sudharshan, S.J. Anthelmintic and antioxidant efficacy of two macrolichens of Ramalinaceae. Pharmacogn. J. 2009, 1, 238–242. [Google Scholar]
- Dias, D.A.; Urban, S. Phytochemical investigation of the Australian lichens Ramalina glaucescens and Xanthoria parietina. Nat. Prod. Commun. 2009, 4, 959–964. [Google Scholar] [PubMed]
- Luo, H.; Wei, X.; Yamamoto, Y.; Liu, Y.; Wang, L.; Jung, J.S.; Koh, Y.J.; Hur, J.-S. Antioxidant activities of edible lichen Ramalina conduplicans and its free radical scavenging constituents. Mycoscience 2010, 51, 391–395. [Google Scholar] [CrossRef]
- Ramesh, P.; Baig, E.S.A. Chemical investigation of south Indian lichens: Ramalina species. Indian Drugs 1994, 31, 445–446. [Google Scholar]
- Hamada, N. The effect of temperature on lichen substances in Ramalina subbreviuscula (Lichens). Bot. Mag. 1983, 96, 121–126. [Google Scholar] [CrossRef]
- Asahina, Y.; Kusaka, T. Lichen substances. LXV. A new depside, ramalinolic acid. Ber. Dtsch. Chem. Ges. B 1936, 69B, 450–455. [Google Scholar] [CrossRef]
- Huneck, S. Lichen constituents. LXXXIX. Secondary substances of some lichens. Phytochemistry 1972, 11, 1493–1495. [Google Scholar] [CrossRef]
- Huneck, S. Lichen constituents. XXXIV. Structure of tumidulin, a new chlorine containing depside. Chem. Ber. 1966, 99, 1106–1110. [Google Scholar] [CrossRef]
- Bendz, G.; Santesson, J.; Wachtmeister, C.A. Chemistry of lichens. XXI. Isolation and synthesis of methyl 3,5-dichlorolecanorate, a new depside from Ramalina spp. Acta Chem. Scand. 1965, 19, 1188–1190. [Google Scholar] [CrossRef]
- Koller, G. Ramalic acid. Monatsh. Chem. 1932, 61, 286–292. [Google Scholar] [CrossRef]
- Chester, D.O.; Elix, J.A. The identification of four new meta-depsides in the lichen Ramalina asahinae. Aust. J. Chem. 1978, 31, 2745–2749. [Google Scholar] [CrossRef]
- Islas, N.V.; Zambrano, A.; Rojas, J.L. Ozone Reactivity and Free Radical Scavenging Behavior of Phenolic Secondary Metabolites in Lichens Exposed to Chronic Oxidant Air Pollution from Mexico City. J. Chem. Ecol. 2007, 33, 1619–1634. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.; Ueno, T. Depside from an isolated lichen mycobiont. Agric. Biol. Chem. 1987, 51, 1705–1706. [Google Scholar] [CrossRef]
- Huneck, S.; Follmann, G.; Santesson, J. Lichen components. XLIX. 4-O-demethylbarbatinic acid, a new depside from Ramalina subdecipiens. Z. Naturforschung B 1968, 23, 856–860. [Google Scholar]
- Hesse, O. Contribution for the knowledge of the lichens and their characteristic components. J. Prakt. Chem. 1903, 68, 1–71. [Google Scholar] [CrossRef]
- Culberson, C.F.; LaGreca, S.; Johnson, A.; Culberson, W.L. Trivaric acid, a new tridepside in the Ramalina americana chemotype complex (lichenized Ascomycota: Ramalinaceae). Bryologist 2000, 102, 595–601. [Google Scholar] [CrossRef]
- Elix, J.A.; Peculiarities of the Australiasian lichen flora: Accessory metabolites, chemical and hybrid strains. J. Hattori Bot. Lab. 1982, 52, 407–415. [Google Scholar]
- Culberson, C.F.; Culberson, W.L.; Johnson, A. A novel meta-depside from the Ramalina subfraxinea complex (Ascomycotina, Ramalinaceae). Bryologist 1990, 93, 193–196. [Google Scholar] [CrossRef]
- Bodo, B.; Hebrard, P.; Molho, L.; Molho, D. Desmaziera evernioides and Ramalina bourgeana. Tetraedron Lett. 1973, 18, 1631–1634. [Google Scholar] [CrossRef]
- Bodo, B. Bourgeanic acid: A new metabolite of lichens. Structure, synthesis and biosynthesis. Bull. Mus. Natl. Hist. Nat. Sci. Phys.-Chim. 1975, 3, 23–97. [Google Scholar]
- Marshak, A.; Barry, G.T.; Craig, L.C. Antibiotic compound isolated from the lichen Ramalina reticulate. Science 1947, 24, 394–395. [Google Scholar] [CrossRef]
- Stocker-Worgotter, E.; Elix, J.A.; Grube, M. Secondary chemistry of lichen forming fungi: Chemosyndromic variation and DNA analyses of cultures and chemotypes in the Ramalina farinacea. Bryologist 2004, 107, 152–162. [Google Scholar] [CrossRef]
- Hawksworth, D.L. Chemical strains of the lichen Ramalina subfariancacea. Bot. Notiser 1968, 121, 317–320. [Google Scholar]
- Culberson, C.F. Microchemical tests for the lichen depsidone hypoprotocetraric acid. Bryologist 1965, 68, 301–304. [Google Scholar] [CrossRef]
- Huneck, S.; Lehn, J.M. Lichen constituents. XXVII. Identity between coquimboic acid and hypoprotocetraric acid. Z. Naturforschung B 1966, 21, 299–299. [Google Scholar]
- Yamamoto, Y.; Watanabe, A. Fatty acid composition of lichens and their phyco and mycobionts. J. Gen. Appl. Microbiol. 1974, 20, 83–86. [Google Scholar] [CrossRef]
- Blunt, J.W.; Munro, M.H.G. Dictionary of Marine Natural Products; Chapman & Hall–Taylor & Francis Group: New York, NY, USA, 2008. [Google Scholar]
- Fazio, A.T.; Adler, M.; Maier, M.S. Usnic acid and triacylglycerides production by the cultured lichen mycobiont of Ramalina celastri. Nat. Prod. Commun. 2014, 9, 213–214. [Google Scholar] [PubMed]
- Hirayama, T.; Fujikawa, F.; Kasahara, T.; Otsuka, M.; Nishida, N.; Mizuno, D. Anti-tumor activities of some lichen products and their degradation products. Yakugaku Zasshi 1980, 100, 755–759. [Google Scholar] [PubMed]
- Czeczuga, B.; de Corona, L.F. Carotenoids in lichens from Argentina. Biochem. Syst. Ecol. 1987, 15, 519–522. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Lipids of lichens. Prog. Lipid Res. 1992, 31, 373–397. [Google Scholar] [CrossRef] [PubMed]
- Huneck, S.; Follmann, G. Lichen constituents. XXVIII. Chemistry of Chilean lichen. Compounds from Ramalina tigrina, and Ramalina inanis. Z. Naturforschung B 1966, 21, 713–714. [Google Scholar]
- Lehn, J.M.; Huneck, S. Lichen constituents. XVIII. First separation of the diterpene (−)-16-α-hydroxykaurane from a lichen. Z. Naturforschung B 1965, 20, 1013–1013. [Google Scholar]
- Sassaki, G.L.; Cruz, L.M.; Gorin, P.A.J.; Iacomini, M. Fatty acid composition of lipids present in selected lichenized fungi: A chemotyping study. Lipids 2001, 36, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Machado, M.J.; Guerrini, M.; Gorin, P.A.J.; Torri, G.; Iacomoni, M. A galactosphingolipid from the lichen, Ramalina celastri. Phytochemistry 1997, 45, 651–653. [Google Scholar] [CrossRef]
- Unal, D.; Senkardesler, A. Abscisic acid and polyamine contents in the lichens Pseudevernia furfuracea and Ramalina farinacea. A. Sukatar. Russ. J. Plant Physiol. 2008, 55, 115–118. [Google Scholar] [CrossRef]
- Mateos, J.L.; Legaz, M.E. Presence and Identification of Polyamines and their Conjugation to Phenolics in Some Epiphytic Lichens. In Eukaryotism and Symbiosis; Springer-Verlag: Berlin, Germany, 1997; pp. 491–498. [Google Scholar]
- Solberg, Y.J. Studies on the chemistry of lichens. IX Quantitative determination of monosaccharides and amino acids in hydrolysates of several Norwegian lichen species. Lichenologist 1970, 4, 283–288. [Google Scholar] [CrossRef]
- Ramakrishnan, S.; Subrmanian, S.S. Amino acids of Lobaria subisidiosa, Umbilicaria pustulata, Parmelia nepalensis, and Ramalina sinensis. Curr. Sci. Indian 1966, 35, 124–125. [Google Scholar]
- Fujikawa, F.; Hirai, K.; Hirayama, T.; Toyota, T.; Urasaki, M.; Takagawa, M.; Fukuda, M.; Moritani, K.; Sanae, T. Free amino acids in lichens of Japan. Yakugaku Zasshi 1972, 92, 823–827. [Google Scholar] [PubMed]
- Paudel, B.; Bhattarai, H.D.; Lee, H.K.; Oh, H.; Shin, H.W.; Han, J.; Yim, J.H. Antibacterial activities of ramalin, usnic acid and its tree derivatives isolated from the Antarctic lichen Ramalina terebrata. Z. Naturforschung C 2010, 65, 34–38. [Google Scholar]
- Yim, J.H.; Kim, I.C.; Han, S.J.; Lee, H.S.; Battarai, H.D.; Yang, S-G.; Lee, D.H. Ramalin for Preveting or Treating Hepatic Fibrosis and Liver Cirrhosis. WO 2,013,129,714 A1, 6 September 2013. [Google Scholar]
- Yim, J.H.; Kim, I.C.; Lee, S.G.; Kim, D.G.; Han, S.J.; Lee, H.S.; Kim, S.J.; Kim, T.G.; Kang, P.S.; Park, H.Y. Ramalin for Preveting and Treating Inflammatory Diseases and Immune Diseases. KR 1290745 B1, 29 September 2013. [Google Scholar]
- Seo, C.; Yim, J.H.; Lee, H.K.; Park, S.M.; Sohn, J.H.; Oh, H. Stereocalpin A, a bioactive cyclic depsipeptide from the Antarctic lichen Stereocaulon alpinum. Tetrahedron Lett. 2008, 49, 29–31. [Google Scholar] [CrossRef]
- Byeon, H.E.; Park, B.K.; Yim, J.H.; Lee, H.K.; Moon, E.Y.; Rhee, D.K.; Pyo, S. Stereocalpin A inhibits the expression of adhesion molecules in activated vascular smooth muscle cells. Int. Immunopharmacol. 2012, 12, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.H.; Kim, I.C.; Kim, D.G.; Han, S.J.; Lee, H.S.; Oh, H.C.; Pyo, S.N. Stereoclapin-A for Preveting or Treating Arteriosclerosis. KR 20,130,043,995 A, 2 May 2013. [Google Scholar]
- Pfau, A.S. Constituents of lichens. II. Constitution of barbatinic acid. Helv. Chim Acta 1928, 11, 864–876. [Google Scholar] [CrossRef]
- Huang, W.; Dai, W. The constituents of absolute from Ramalina fastigiata (Pors) Ach. The main ethyl ester of fatty acids. Nat. Prod. Res. Dev. 1989, 1, 27–28. [Google Scholar]
- Zygadlo, J.A.; Pignata, M.L.; Gonzalez, C.M.; Levin, A. Alkanes in lichens. Phytochemistry 1993, 32, 1453–1456. [Google Scholar] [CrossRef]
- Rundel, P.W. The ecological role of secondary lichen substances. Biochem. Syst. Ecol. 1978, 6, 157–170. [Google Scholar] [CrossRef]
- Gulluce, M.; Aslan, A.; Sokmen, M.; Sahin, F.; Adiguzel, A.; Agar, G.; Sokmen, A. Screening the antioxidant and antimicrobial properties of the lichens Parmelia saxatilis, Platismatia glauca, Ramalina pollinaria, Ramalina polymorpha and Umbilicaria nylanderiana. Phytomedicine 2006, 13, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Paudel, B.; Bhattarai, H.D.; Lee, J.S.; Hong, S.G.; Shin, H.W.; Yim, J.H. Antibacterial potential of Antarctic lichens against human pathogenic Gram-positive bacteria. Phytother. Res. 2008, 22, 1269–1271. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, H.D.; Lee, Y.K.; Cho, K.H.; Shin, H.V. The study of antagonistic interactions among pelagic bacteria: A promising way to coin environmentally friendly antiflouling compounds. Hydrobiologia 2006, 568, 417–423. [Google Scholar] [CrossRef]
- Swenson, J.M.; Thornsberry, C.; Silcox, V.A. Rapidly growing mycobacteria: Testing of susceptibility to 34 antimicrobial agentes by microdilution. Antimiob Agent Chemother. 1982, 22, 186–192. [Google Scholar] [CrossRef]
- Paudel, B.; Bhattarai, H.D.; Pandey, D.P.; Hur, J.S.; Hong, S.G.; Kim, I.-C.; Yim, J.H. Antioxidant, antibacterial activity and brine shrimp toxicity test of some mountainous lichens from Nepal. Biol. Res. 2012, 45, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 26, 1199–1200. [Google Scholar] [CrossRef]
- Mitsuda, H.; Yuasumoto, K.; Iwami, K. Antioxidation action of indole compounds during the autoxidation of linoleic acid. Nihon Eiyo Shokuryo Gakkai Shi 1996, 19, 210–214. [Google Scholar] [CrossRef]
- Luo, H.; Yamamota, Y.; Kim, J.A.; Jung, J.S.; Koh, Y.J.; Hur, J.S. Lecanoric acid, a secondary lichen substance with antioxidant properties from Umbilicaria Antarctica in maritime Antarctica (King George Island). Polar Biol. 2009, 32, 1033–1040. [Google Scholar] [CrossRef]
- Halici, M.; Kufrevioglu, O.I.; Odabasoglu, F.; Halici, Z.; Cakir, A.; Aslan, A. The ethanol-water extract of Ramalina capitata has gastroprotective and antioxidative properties: An experimental study in rats with indomethacin-induced gastric injuries. J. Food Biochem. 2011, 35, 11–26. [Google Scholar] [CrossRef]
- Stanly, C.; Hag, D.M.; Keng, C.L.; Boey, P.-L.; Bhatt, A. Comparative evaluation of antioxidant activity and total phenolic content of selected species from Malaysia. J. Pharm. Res. 2011, 4, 2824–2827. [Google Scholar]
- García, C.C.; Rosso, M.L.; Bertoni, M.D.; Maier, M.S.; Damonte, E.B. Evaluation of the antiviral activity against Junin vírus of macrocyclic trichothecenes produced by the hypocrealean epibiont of Baccharis corififolia. Planta Med. 2002, 68, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Esimone, C.O.; Grunwald, T.; Nworu, C.S.; Kuate, S.; Proksch, P.; Ueberla, K. Broad spectrum antiviral fractions from the lichen Ramalina farinacea (L.) Ach. Chemotherapy 2009, 55, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Brandão, L.F.G.; Alcantara, G.B.; Matos, M.F.C.; Bogo, D.; Freitas, D.S.; Oyama, N.M.; Honda, N.K. Cytotoxic evaluation of phenolic compounds from lichens against melanona cells. Chem. Pharm. Bull. 2013, 61, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Nambiar, D.; Kale, R.K.; Singh, R.P. Usnic acid inhibits and induces cell cycle arrest and apoptosis in human lung carcinoma A549 cells. Nutr. Cancer 2013, 65, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Carneiro-Leão, A.M.A.; Buchi, D.F.; Iacomini, M.; Gorin, P.A.J.; Oliveira, M.B.M. Cytotoxic effect againts HeLa cells of polysaccharides from the lichen Ramalina celastri. J. Submicrosc. Cytol. Pathol. 1997, 29, 503–509. [Google Scholar] [PubMed]
- Haraldsdóttir, S.; Guölaugsdóttir, E.; Ingólfsdóttir, K.; Ögmundsdóttir, H.M. Anti-proliferative effects of lichen-derived lipoxygenase inhibitors on twelve human cancer cell lines of different tissue origin in vitro. Planta Med. 2004, 70, 1098–1100. [Google Scholar] [CrossRef] [PubMed]
- Koparal, A.T.; Tuylu, B.A.; Turk, H. In vitro cytotoxic activities of (+)-usnic acid and (−)-usnic acid on V79, A549, and human lymphocyte cells and their non-genotoxicity on human lymphocytes. Nat. Prod. Res. 2006, 20, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Einarsdóttir, E.; Groeneweg, J.; Björnsdóttir, G.G.; Haröardottir, G.; Omarsdóttir, S.; Ingólfsdóttir, K.; Ögmundsdóttir, H.M. Cellular mechanisms of the anticancer effects of the lichen compound usnic acid. Planta Med. 2010, 76, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Bačkorová, M.; Bačkor, M.; Mikeš, J.; Jendželovský, R.; Fedoročko, P. Variable responses of different human cancer cells to the lichen compounds parietin, atranorin, usnic acid and gyrophoric acid. Toxicol. In Vitro 2011, 25, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Bačkorová, M.; Jendželovský, R.; Kello, M.; Bačkor, M.; Mikeš, J.; Fedoročko, P. Lichen secondary metabolites are responsible for induction of apoptosis in HT-29 and A2780 human cancer cell lines. Toxicol. In Vitro 2012, 26, 462–468. [Google Scholar] [CrossRef] [PubMed]
- Holbeck, S.L. Update on NCI in vitro drug screen utilities. Eur. J. Cancer 2004, 40, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Honda, N.K.; Brum, R.L.; Marques, M.R.; Marcelli, M.P.; Fleig, M.; Vilegas, W. Fungitoxic activity of compounds isolated from lichens. South. Braz. J. Chem. 1999, 7, 61–66. [Google Scholar]
- Wei, X.; Xinli, J.; Jeon, H.S.; Han, K.S.; Koh, Y.J.; Hur, J.S. Antifungal activity of lichen forming fungi against Colletotrichum acutatum on hot pepper. Plant Pathol. J. 2008, 24, 202–206. [Google Scholar] [CrossRef]
- Honda, N.K.; Pavan, F.R.; Coelho, R.G.; Andrade Leite, S.R.; Micheletti, A.C.; Lopes, T.I.B.; Misutsu, M.Y.; Beatriz, A.; Brum, R.L.; Leite, C.Q.F. Antimycobacterial activity of lichen substances. Phytomedicine 2010, 17, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Lee, H.G.; Kim, I.C.; Hong, S.G.; Kim, D.G.; Lee, Y.G.; Lee, S.G.; Han, S.J.; Lee, H.S.; Lee, Y.M. Cosmetics Containing Usimine-C for Promoting Collagen Production. KR 20,110,084,753 A, 26 July 2011. [Google Scholar]
- Araojo, R.V.S.; Melo-Junior, M.R.; Beltrão, E.I.C.; Mello, L.A.; Iacomini, M.; Carneiro-Leao, A.M.A.; Carvalho, L.B.; Santos-Magalhães, N.S. Evaluation of the antischistosomal activity of sulfated α-d-glucan from the lichen Ramalina celastri free and encapsulated into liposomes. Braz. J. Med. Biol. Res. 2011, 44, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Cetin, H.; Tufan-Cetin, O.; Turk, A.O.; Tay, T.; Candan, M.; Yanikoglu, A.; Sumbul, H. Insecticidal activity of major lichen compounds, (−)- and (+)-usnic acid, against the larvae of house mosquito, Culex pipiens L. Parasitol. Res. 2008, 102, 1277–1279. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, A.S.N.; Braz-Filho, R.; Mussi-Dias, V.; Vieira, I.J.C. Chemistry and Biological Activity of Ramalina Lichenized Fungi. Molecules 2015, 20, 8952-8987. https://doi.org/10.3390/molecules20058952
Moreira ASN, Braz-Filho R, Mussi-Dias V, Vieira IJC. Chemistry and Biological Activity of Ramalina Lichenized Fungi. Molecules. 2015; 20(5):8952-8987. https://doi.org/10.3390/molecules20058952
Chicago/Turabian StyleMoreira, Antônio Sérgio Nascimento, Raimundo Braz-Filho, Vicente Mussi-Dias, and Ivo José Curcino Vieira. 2015. "Chemistry and Biological Activity of Ramalina Lichenized Fungi" Molecules 20, no. 5: 8952-8987. https://doi.org/10.3390/molecules20058952