

Biological Properties, Phenolic Profile, and Botanical Aspect of Nigella sativa L. and Nigella damascena L. Seeds: A Comparative Study

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Results
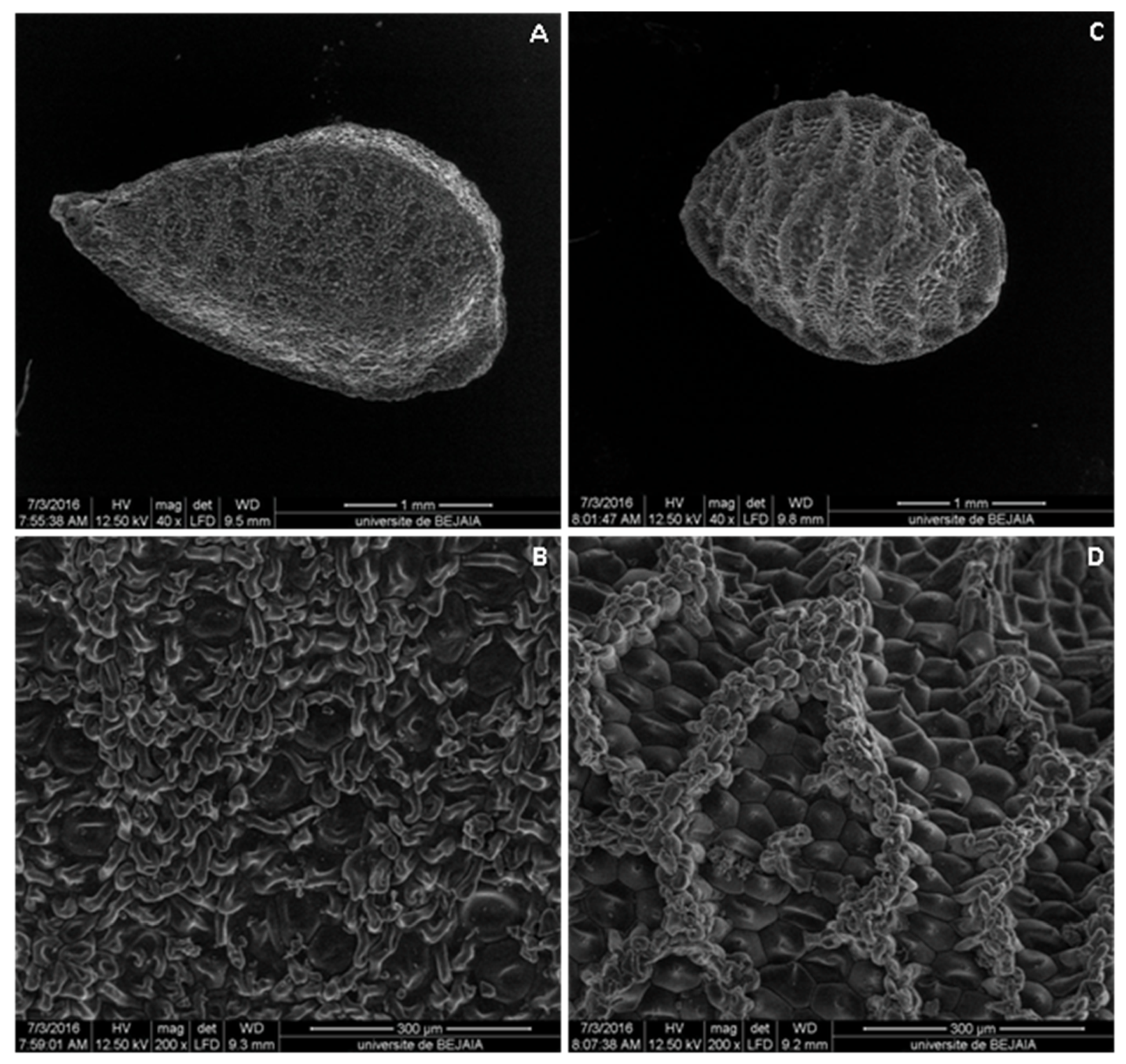
2.1. Botanical Study
2.2. Chemical Composition
2.2.1. Nutrient Contents
2.2.2. Phenolic Compound Contents
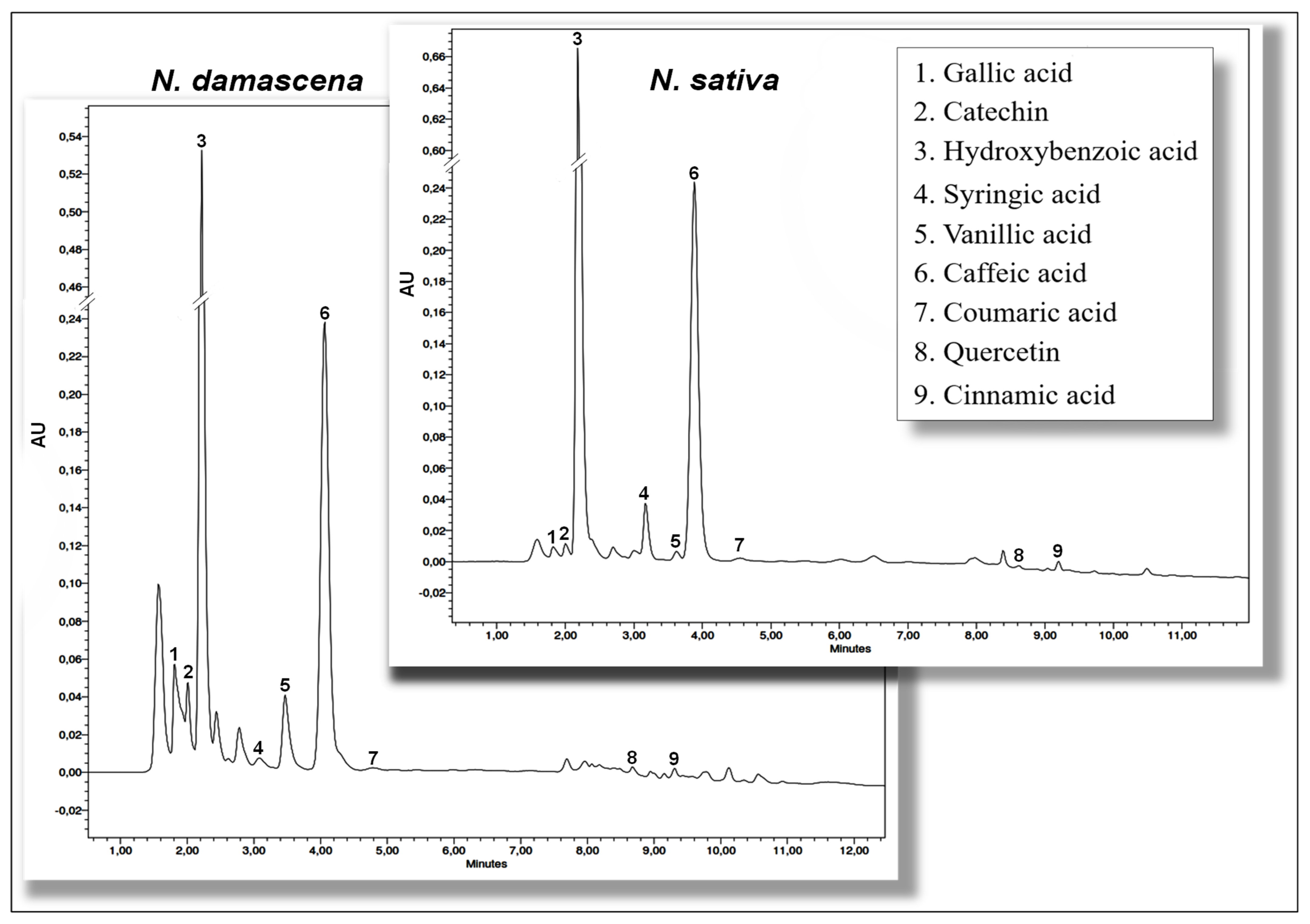
2.2.3. Identification and Quantification of Phenolic Compounds Using HPLC-DAD
2.3. Biological Activities
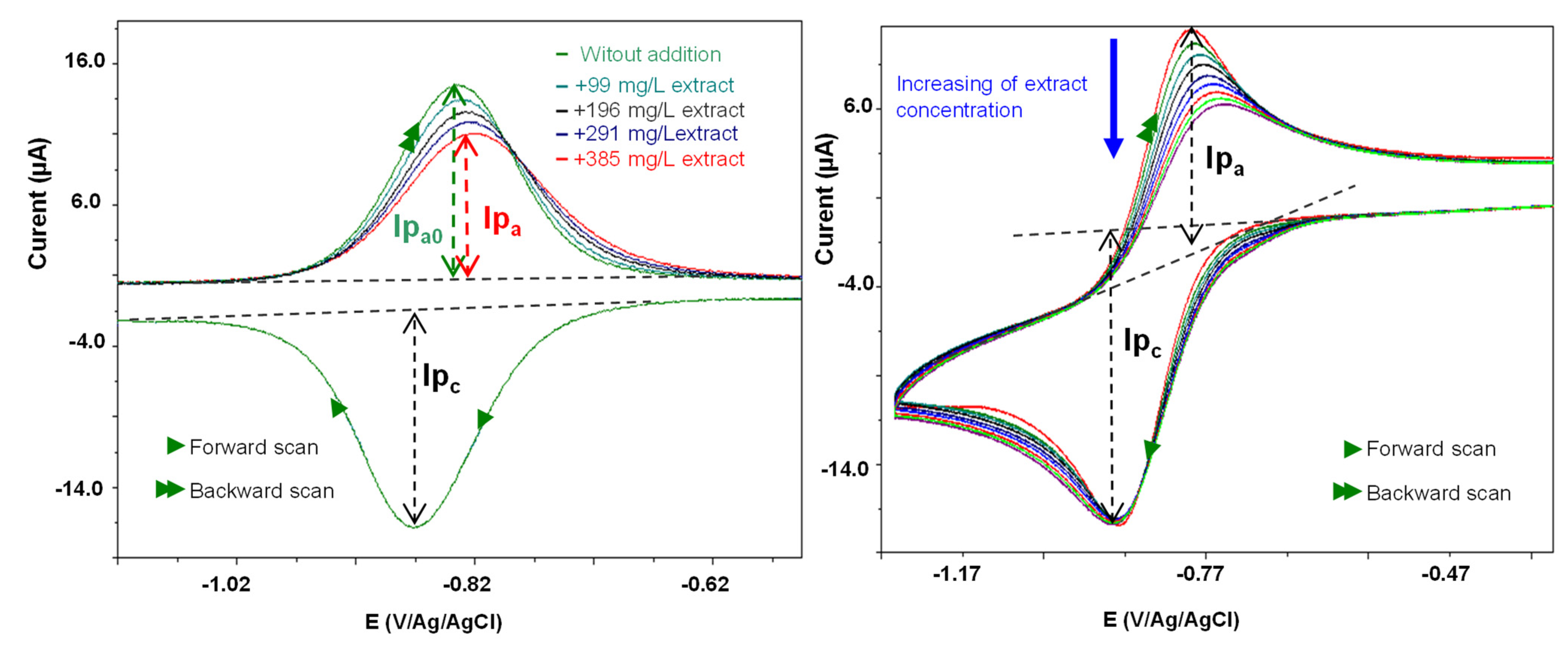
2.3.1. In Vitro Antioxidant Activities
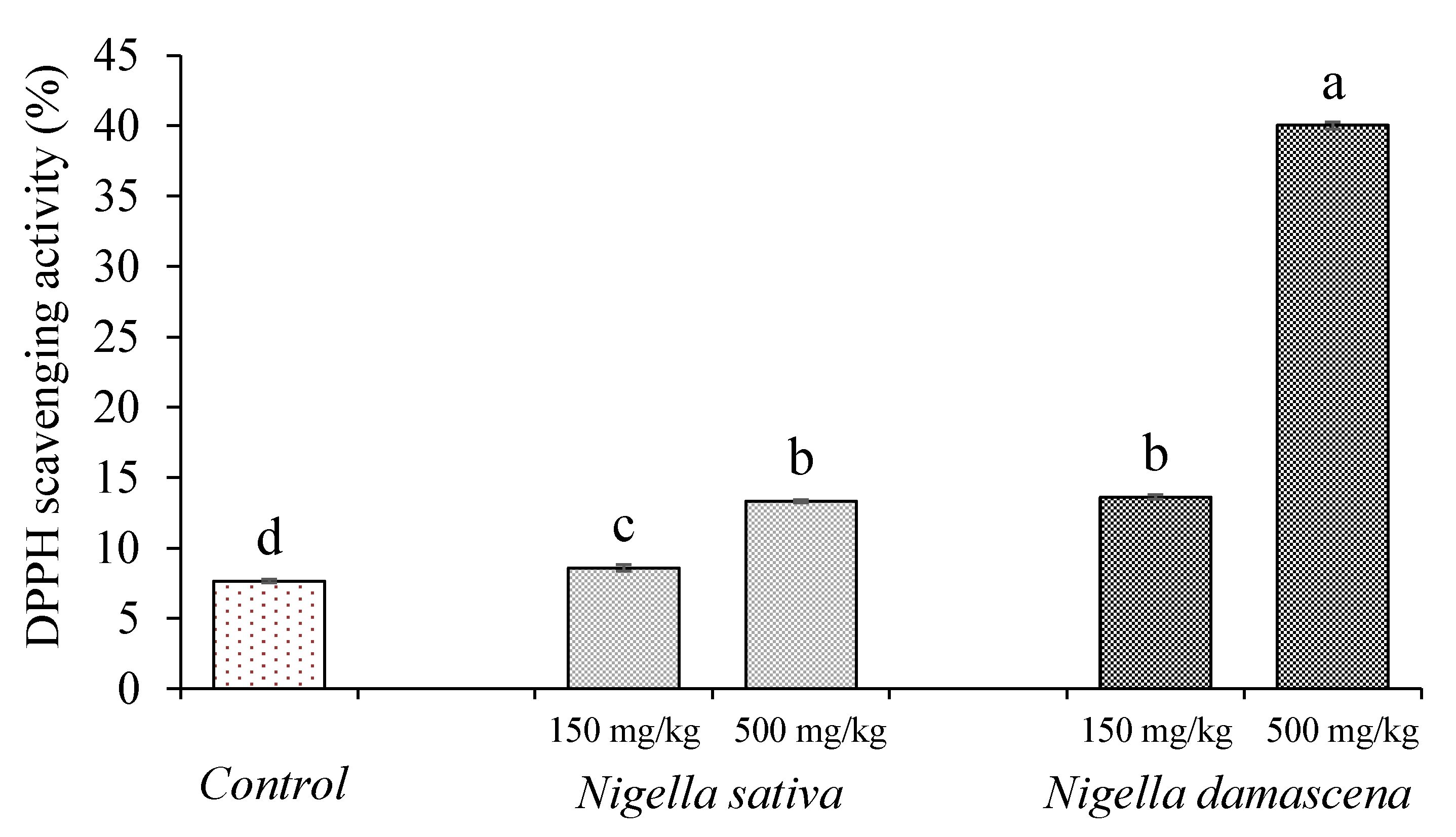
2.3.2. In Vivo Antioxidant Activity
2.3.3. Antimicrobial Activity
3. Materials and Methods
3.1. Plants Material
3.2. Proximal Analysis
3.3. Phenolic Compounds Analysis
3.4. Bioactive Properties
3.4.1. In Vitro Antioxidant Activities
3.4.2. In Vivo Antioxidant Properties
3.4.3. Antimicrobial Activity
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Helvacıoğlu, S.; Charehsaz, M.; Güzelmeriç, E.; Oçkun, M.A.; Ayran, İ.; Kırmızıbekmez, H.; Kan, Y.; Aydın, A.; Yeşilada, E. protective effect of Nigella sativa and Nigella damascena fixed oils against aflatoxin induced mutagenicity in the classical and modified ames test. Chem. Biodivers. 2021, 18, e2000936. [Google Scholar] [CrossRef] [PubMed]
- Kokoska, L. Chemistry and Biological Activity of Nigella Genus: The Antimicrobial and Anti-Inflammatory Effects of Seed Extracts, Essential Oils and Compounds of Six Nigella Species; LAP LAMBERT Academic Publishing: Berlin, Germany, 2011; p. 90. [Google Scholar]
- Margout, D.; Kelly, M.T.; Meunier, S.; Auinger, D.; Pelissier, Y.; Larroque, M. Morphological, microscopic and chemical comparison between Nigella sativa L. cv (black cumin) and Nigella damascena L. cv. J. Agric. Food Environ. 2013, 11, 165–171. [Google Scholar]
- Salehi, B.; Quispe, C.; Imran, M.; Ul-Haq, I.; Živković, J.; Abu-Reidah, I.M.; Sen, S.; Taheri, Y.; Acharya, K.; Azadi, H. Nigella plants–Traditional uses, bioactive phytoconstituents, preclinical and clinical studies. Front. Pharmacol. 2021, 12, 625386. [Google Scholar] [CrossRef] [PubMed]
- Topcagic, A.; Zeljkovic, S.C.; Karalija, E.; Galijasevic, S.; Sofic, E. Evaluation of phenolic profile, enzyme inhibitory and antimicrobial activities of Nigella sativa L. seed extracts. Bosn. J. Basic Med. Sci. 2017, 17, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Zhou, L.; Meng, L.; Chen, S.; Ma, C.; Liu, Z.; Kang, W. Recent progress on chemical constituents and pharmacological effects of the genus Nigella. Evid. Based Complement. Altern. Med. 2020, 2020, 6756835. [Google Scholar] [CrossRef]
- Shahbazi, E.; Safipor, B.; Golkar, P. Responses of Nigella damascena L. and Nigella sativa L. to drought stress: Yield, fatty acid composition and antioxidant activity. J. Agric. Sci. Technol. 2022, 24, 693–705. [Google Scholar]
- Sharma, R.; Sahu, P.; Jain, A.; Kumar, V.; Khokhar, D.; Geda, A.K.; Gupta, B. Nigella sativa. In Nutraceuticals in Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2019; pp. 91–101. [Google Scholar]
- Gilani, A.-u.H.; Jabeen, Q.; Khan, M.A.U. A review of medicinal uses and pharmacological activities of Nigella sativa. Pak. J. Biol. Sci. 2004, 7, 441–451. [Google Scholar]
- Khare, C.P. Indian Herbal Remedies: Rational Western Therapy, Ayurvedic, and Other Traditional Usage, Botany; Springer Science & Business Media: New Delhi, India, 2004. [Google Scholar]
- Silva, A.F.C.; Haris, P.I.; Serralheiro, M.L.; Pacheco, R. Mechanism of action and the biological activities of Nigella sativa oil components. Food Biosci. 2020, 38, 100783. [Google Scholar] [CrossRef]
- AI-Jassir, M.S. Chemical composition and microflora of black cumin (Nigella sativa L.) seeds growing in Saudi Arabia. Food Chem. 1992, 45, 239–242. [Google Scholar] [CrossRef]
- Iqbal, M.J.; Butt, M.S.; Qayyum, M.M.N.; Suleria, H.A.R. Anti-hypercholesterolemic and anti-hyperglycaemic effects of conventional and supercritical extracts of black cumin (Nigella sativa). Asian Pac. J. Trop. Biomed. 2017, 7, 1014–1022. [Google Scholar] [CrossRef]
- Gholamnezhad, Z.; Shakeri, F.; Saadat, S.; Ghorani, V.; Boskabady, M.H. Clinical and experimental effects of Nigella sativa and its constituents on respiratory and allergic disorders. Avicenna J. Phytomed. 2019, 9, 195. [Google Scholar] [PubMed]
- Butt, A.S.; Nisar, N.; Ghani, N.; Altaf, I.; Mughal, T.A. Isolation of thymoquinone from Nigella sativa L. and Thymus vulgaris L., and its anti-proliferative effect on HeLa cancer cell lines. Trop. J. Pharm. Res. 2019, 18, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Majdalawieh, A.F.; Fayyad, M.W. Recent advances on the anti-cancer properties of Nigella sativa, a widely used food additive. J. Ayurveda Integr. Med. 2016, 7, 173–180. [Google Scholar] [CrossRef]
- Noor, N.A.; Ezz, H.S.A.; Faraag, A.R.; Khadrawy, Y.A. Evaluation of the antiepileptic effect of curcumin and Nigella sativa oil in the pilocarpine model of epilepsy in comparison with valproate. Epilepsy Behav. 2012, 24, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kökdil, G.; Tamer, L.; Ercan, B.; İlcim, A.; Aras, N.; Atik, U. Effects of Nigella unguicularis fixed oil on blood biochemistry and oxidant/antioxidant balance in rats. J. Ethnopharmacol. 2005, 99, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Rani, R.; Dahiya, S.; Dhingra, D.; Dilbaghi, N.; Kim, K.-H.; Kumar, S. Improvement of antihyperglycemic activity of nano-thymoquinone in rat model of type-2 diabetes. Chem.-Biol. Interact. 2018, 295, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Toma, C.-C.; Olah, N.-K.; Vlase, L.; Mogoșan, C.; Mocan, A. Comparative studies on polyphenolic composition, antioxidant and diuretic effects of Nigella sativa L. (black cumin) and Nigella damascena L. (lady-in-a-mist) seeds. Molecules 2015, 20, 9560–9574. [Google Scholar] [CrossRef] [Green Version]
- D’Antuono, L.F.; Moretti, A.; Lovato, A.F. Seed yield, yield components, oil content and essential oil content and composition of Nigella sativa L. and Nigella damascena L. Ind. Crops Prod. 2002, 15, 59–69. [Google Scholar] [CrossRef]
- Moretti, A.; D’Antuono, L.F.; Elementi, S. Essential oils of Nigella sativa L. and Nigella damascena L. seed. J. Essent. Oil Res. 2004, 16, 182–183. [Google Scholar] [CrossRef]
- Benkaci-Ali, F.; Baaliouamer, A.; Wathelet, J.P.; Marlier, M. Chemical composition of volatile oils from Algerian Nigella sativa L. seeds. J Essent. Oil Res 2010, 22, 318–322. [Google Scholar] [CrossRef]
- Kabir, Y.; Shirakawa, H.; Komai, M. Nutritional composition of the indigenous cultivar of black cumin seeds from Bangladesh. Prog. Nutr. 2019, 21, 428–434. [Google Scholar]
- Nergiz, C.; Ötleş, S. Chemical composition of Nigella sativa L. seeds. Food Chem. 1993, 48, 259–261. [Google Scholar] [CrossRef]
- Takruri, H.R.; Dameh, M.A. Study of the nutritional value of black cumin seeds (Nigella sativa L.). J. Sci. Food Agric. 1998, 76, 404–410. [Google Scholar] [CrossRef]
- Fico, G.; Panizzi, L.; Flamini, G.; Braca, A.; Morelli, I.; Tomè, F.; Cioni, P. Biological screening of Nigella damascena for antimicrobial and molluscicidal activities. Phytother. Res. 2004, 18, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Modica, A.; Bazan, G.; Marino, P.; Bruno, M. The ethnobotany, phytochemistry, and biological properties of Nigella damascena–A review. Phytochemistry 2022, 198, 113165. [Google Scholar] [CrossRef]
- Fico, G.; Braca, A.; Tomè, F.; Morelli, I. A new phenolic compound from Nigella damascena seeds. Fitoterapia 2001, 72, 462–463. [Google Scholar] [CrossRef]
- Sajfrtova, M.; Sovova, H.; Karban, J. Enrichment of Nigella damascena extract with volatile compounds using supercritical fluid extraction. J. Supercrit. Fluids 2014, 94, 160–164. [Google Scholar] [CrossRef]
- Fico, G.; Bader, A.; Flamini, G.; Cioni, P.L.; Morelli, I. Essential oil of Nigella damascena L. (Ranunculaceae) seeds. J. Essent. Oil Res. 2003, 15, 57–58. [Google Scholar] [CrossRef]
- Ogawa, K.; Nakamura, S.; Hosokawa, K.; Ishimaru, H.; Saito, N.; Ryu, K.; Fujimuro, M.; Nakashima, S.; Matsuda, H. New diterpenes from Nigella damascena seeds and their antiviral activities against herpes simplex virus type-1. J. Nat. Med. 2018, 72, 439–447. [Google Scholar] [CrossRef]
- Piras, A.; Rosa, A.; Marongiu, B.; Porcedda, S.; Falconieri, D.; Dessi, M.A.; Ozcelik, B.; Koca, U. Chemical composition and in vitro bioactivity of the volatile and fixed oils of Nigella sativa L. extracted by supercritical carbon dioxide. Ind. Crops Prod. 2013, 46, 317–323. [Google Scholar] [CrossRef]
- Rchid, H.; Chevassus, H.; Nmila, R.; Guiral, C.; Petit, P.; Chokaïri, M.; Sauvaire, Y. Nigella sativa seed extracts enhance glucose-induced insulin release from rat-isolated Langerhans islets. Fundam. Clin. Pharmacol. 2004, 18, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Sieniawska, E.; Michel, P.; Mroczek, T.; Granica, S.; Skalicka-Woźniak, K. Nigella damascena L. essential oil and its main constituents, damascenine and β-elemene modulate inflammatory response of human neutrophils ex vivo. Food Chem. Toxicol. 2019, 125, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zaitoun, S.; Al-Ghzawi, A.A.-M.; Samarah, N.; Alqudah, A. Comparative study in seed yield and flowers attractivity to bee visitors between Nigella sativa L. and Nigella damascena L. (Ranunculaceae) grown under semiarid conditions. Ital. J. Agron. 2008, 3, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Kökdil, G.; Ilçim, A.; Özbilgin, B.; Uygun, C. Morphology and stem anatomy of some species of genus Nigella L. in Turkey: Türkiye’deki Bazi Nigella Türlerinin Morfoloji ve Gövde Anatomileri. J. Fac. Pharm. Ankara 2006, 35, 19–41. [Google Scholar] [CrossRef]
- Datta, A.K.; Saha, A. Cytomorphological studies and seed protein characterization of Nigella sativa L. and Nigella damascena L. Cytologia 2003, 68, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Dadandi, M.; Kökdil, G.; İlçim, A.; Özbilgin, B. Seed macro and micro morphology of the selected Nigella (Ranunculaceae) taxa from Turkey and their systematic significance. Biologia 2009, 64, 261–270. [Google Scholar] [CrossRef]
- Barkah, N.N.; Wiryawan, K.G.; Retnani, Y.; Wibawan, W.T.; Wina, E. Physicochemical properties of products and waste of black seed produced by cold press method. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2021; p. 012025. [Google Scholar]
- Javorková, V.; Václavík, J.; Kubínová, R.; Muselík, J. Comparison of antioxidant activity and polyphenolic content in different extracts of Nigella sativa, Nigella orientalis and Nigella damascena. Sci. J. Phytotech. Zootechn. 2011, 14, 1–4. [Google Scholar]
- Thippeswamy, N.; Naidu, K.A. Antioxidant potency of cumin varieties—Cumin, black cumin and bitter cumin—On antioxidant systems. Eur. Food Res. Technol. 2005, 220, 472–476. [Google Scholar] [CrossRef]
- Khattak, K.F.; Simpson, T.J. Effect of gamma irradiation on the extraction yield, total phenolic content and free radical-scavenging activity of Nigella staiva seed. Food Chem. 2008, 110, 967–972. [Google Scholar] [CrossRef]
- Al-Bishri, W.; Danial, E.N. Comparative study on the antioxidant, antimicrobial activities and total phenolic content of selected seeds from Saudi Arabia. J. Agric. Food Environ. 2013, 11, 202–207. [Google Scholar]
- Dalli, M.; Azizi, S.-E.; Kandsi, F.; Gseyra, N. Evaluation of the in vitro antioxidant activity of different extracts of Nigella sativa L. seeds, and the quantification of their bioactive compounds. Mater. Today Proc. 2021, 45, 7259–7263. [Google Scholar] [CrossRef]
- Miho, H.; Moral, J.; Barranco, D.; Ledesma-Escobar, C.; Priego-Capote, F.; Díez, C. Influence of genetic and interannual factors on the phenolic profiles of virgin olive oils. Food Chem. 2021, 342, 128357. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Külheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef]
- Navarro, J.M.; Flores, P.; Garrido, C.; Martinez, V. Changes in the contents of antioxidant compounds in pepper fruits at different ripening stages, as affected by salinity. Food Chem. 2006, 96, 66–73. [Google Scholar] [CrossRef]
- Moghimi, M.; Farzaneh, V.; Bakhshabadi, H. The effect of ultrasound pretreatment on some selected physicochemical properties of black cumin (Nigella sativa). Nutrire 2018, 43, 18. [Google Scholar] [CrossRef] [Green Version]
- Pinelo, M.; Rubilar, M.; Jerez, M.; Sineiro, J.; Núñez, M.J. Effect of solvent, temperature, and solvent-to-solid ratio on the total phenolic content and antiradical activity of extracts from different components of grape pomace. J. Agric. Food Chem. 2005, 53, 2111–2117. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, F.; Lounis, N.; Mebarakou, S.; Guendouze, N.; Yalaoui-Guellal, D.; Madani, K.; Boulekbache-Makhlouf, L.; Duez, P. Impact of growth sites on the phenolic contents and antioxidant activities of three algerian mentha species (M. pulegium L., M. rotundifolia (L.) Huds., and M. spicata L.). Front. Pharmacol. 2022, 13, 886337. [Google Scholar] [CrossRef]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Hernández-Carlos, B.; Villanueva-Cañongo, C. Shikimic acid pathway in biosynthesis of phenolic compounds. In Plant Physiological Aspects of Phenolic Compounds; Soto-Hernández, M., García-Mateos, R., Palma-Tenango, M., Eds.; Intechopen: London, UK, 2019; Volume 1, pp. 35–50. [Google Scholar]
- Merfort, I.; Wray, V.; Barakat, H.; Hussein, S.; Nawwar, M.; Willuhn, G. Flavonol triglycosides from seeds of Nigella sativa. Phytochem 1997, 46, 359–363. [Google Scholar] [CrossRef]
- Mariod, A.A.; Ibrahim, R.M.; Ismail, M.; Ismail, N. Antioxidant activity and phenolic content of phenolic rich fractions obtained from black cumin (Nigella sativa) seedcake. Food Chem. 2009, 116, 306–312. [Google Scholar] [CrossRef]
- Mechraoui, O.; Ladjel, S.; Nedjimi, M.S.; Belfar, M.L.; Moussaoui, Y. Determination of polyphenols content, antioxidant and antibacterial activity of Nigella sativa L. seed phenolic extracts. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2018, 19, 411–421. [Google Scholar]
- Tiji, S.; Benayad, O.; Berrabah, M.; El Mounsi, I.; Mimouni, M. Phytochemical profile and antioxidant activity of Nigella sativa L. growing in Morocco. Sci. World J. 2021, 2021, 6623609. [Google Scholar] [CrossRef] [PubMed]
- Ansary, J.; Regolo, L.; Machì, M.; Salinari, A.; Cianciosi, D. Evaluation of the in vitro bioaccessibility of phenolic compounds of black cumin (BARI-1cumin) methanolic extract. eFood 2022, 3, e15. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Biosynthesis of flavonoids and effects of stress. Curr. Opin. Plant Biol. 2002, 5, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Tiji, S.; Rokni, Y.; Benayad, O.; Laaraj, N.; Asehraou, A.; Mimouni, M. Chemical Composition related to antimicrobial activity of Moroccan Nigella sativa L. extracts and isolated fractions. J. Evid. Based Complement. Altern. Med. 2021, 2021, 8308050. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, F.; Achat, S.; Guendouze-Bouchefa, N.; Benazzouz-Smail, L.; Elsebai, M.F.; Madani, K. Recent advances in the identification and the study of composition and activities of medicinal plants. J. Coast Life Med. 2016, 4, 983–999. [Google Scholar]
- Achat, S.; Tomao, V.; Madani, K.; Chibane, M.; Elmaataoui, M.; Dangles, O.; Chemat, F. Direct enrichment of olive oil in oleuropein by ultrasound-assisted maceration at laboratory and pilot plant scale. Ultrason. Sonochem. 2012, 19, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Audibert, L.; Fauchon, M.; Blanc, N.; Hauchard, D.; Ar Gall, E. Phenolic compounds in the brown seaweed Ascophyllum nodosum: Distribution and radical-scavenging activities. Phytochem. Anal. 2010, 21, 399–405. [Google Scholar] [CrossRef]
- Le Bourvellec, C.; Hauchard, D.; Darchen, A.; Burgot, J.-L.; Abasq, M.-L. Validation of a new method using the reactivity of electrogenerated superoxide radical in the antioxidant capacity determination of flavonoids. Talanta 2008, 75, 1098–1103. [Google Scholar] [CrossRef]
- Blanc, N.; Hauchard, D.; Audibert, L.; Gall, E.A. Radical-scavenging capacity of phenol fractions in the brown seaweed Ascophyllum nodosum: An electrochemical approach. Talanta 2011, 84, 513–518. [Google Scholar] [CrossRef]
- Brahmi, F.; Hauchard, D.; Guendouze, N.; Madani, K.; Kiendrebeogo, M.; Kamagaju, L.; Stévigny, C.; Chibane, M.; Duez, P. Phenolic composition, in vitro antioxidant effects and tyrosinase inhibitory activity of three Algerian Mentha species: M. spicata (L.), M. pulegium (L.) and M. rotundifolia (L.) Huds (Lamiaceae). Ind. Crops Prod. 2015, 74, 722–730. [Google Scholar]
- Guendouze-Bouchefa, N.; Madani, K.; Chibane, M.; Boulekbache-Makhlouf, L.; Hauchard, D.; Kiendrebeogo, M.; Stévigny, C.; Okusa, P.N.; Duez, P. Phenolic compounds, antioxidant and antibacterial activities of three Ericaceae from Algeria. Ind. Crops Prod. 2015, 70, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Oldham, K.B. Diffusive transport to planar, cylindrical and spherical electrodes. Electroanal. Chem. Interfacial Electrochem. 1973, 41, 351–358. [Google Scholar] [CrossRef]
- Dangles, O. Antioxidant activity of plant phenols: Chemical mechanisms and biological significance. Curr. Org. Chem. 2012, 16, 692–714. [Google Scholar] [CrossRef]
- Merkl, R.; HRádkoVá, I.; FIlIp, V.; ŠMIdRkal, J. Antimicrobial and antioxidant properties of phenolic acids alkyl esters. Czech J. Food Sci. 2010, 28, 275–279. [Google Scholar] [CrossRef] [Green Version]
- Achat, S.; Rakotomanomana, N.; Madani, K.; Dangles, O. Antioxidant activity of olive phenols and other dietary phenols in model gastric conditions: Scavenging of the free radical DPPH and inhibition of the haem-induced peroxidation of linoleic acid. Food Chem. 2016, 213, 135–142. [Google Scholar] [CrossRef]
- Meziti, A.; Meziti, H.; Boudiaf, K.; Mustapha, B.; Bouriche, H. Polyphenolic profile and antioxidant activities of Nigella sativa seed extracts in vitro and in vivo. Int. J. Botechnol. Bioeng. 2012, 6, 109–117. [Google Scholar]
- El Rabey, H.A.; Al-Seeni, M.N.; Bakhashwain, A.S. The antidiabetic activity of Nigella sativa and propolis on streptozotocin-induced diabetes and diabetic nephropathy in male rats. Evid.-Based Complement. Altern. Med. 2017, 2017, 5439645. [Google Scholar] [CrossRef] [Green Version]
- Kaleem, M.; Kirmani, D.; Asif, M.; Ahmed, Q.; Bano, B. Biochemical effects of Nigella sativa L seeds in diabetic rats. Indian J. Exp. Biol. 2006, 44, 745–748. [Google Scholar]
- Asgary, S.; Ghannadi, A.; Dashti, G.; Helalat, A.; Sahebkar, A.; Najafi, S. Nigella sativa L. improves lipid profile and prevents atherosclerosis: Evidence from an experimental study on hypercholesterolemic rabbits. J. Funct. Foods 2013, 5, 228–234. [Google Scholar] [CrossRef]
- Mahmoudi, A.; Samani, K.G.; Farrokhi, E.; Heidarian, E. Effects of Nigella sativa extracts on the lipid profile and uncoupling protein-1 gene expression in Brown adipose tissue of mice. Adv. Biomed. Res. 2018, 29, 121. [Google Scholar]
- Abbasnezhad, A.; Niazmand, S.; Mahmoudabady, M.; Rezaee, S.A.; Soukhtanloo, M.; Mosallanejad, R.; Hayatdavoudi, P. Nigella sativa L. seed regulated eNOS, VCAM-1 and LOX-1 genes expression and improved vasoreactivity in aorta of diabetic rat. J. Ethnopharmacol. 2019, 228, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Kanter, M.; Coskun, O.; Korkmaz, A.; Oter, S. Effects of Nigella sativa on oxidative stress and β-cell damage in streptozotocin-induced diabetic rats. Anat. Rec. 2004, 279, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Benhaddou-Andaloussi, A.; Martineau, L.C.; Spoor, D.; Vuong, T.; Leduc, C.; Joly, E.; Burt, A.; Meddah, B.; Settaf, A.; Arnason, J.T. Antidiabetic activity of Nigella sativa. seed extract in cultured pancreatic β-cells, skeletal muscle cells, and adipocytes. Pharm. Biol. 2008, 46, 96–104. [Google Scholar] [CrossRef]
- Benhaddou-Andaloussi, A.; Martineau, L.; Vallerand, D.; Haddad, Y.; Afshar, A.; Settaf, A.; Haddad, P. Multiple molecular targets underlie the antidiabetic effect of Nigella sativa seed extract in skeletal muscle, adipocyte and liver cells. Diabetes Obes. Metab. 2010, 12, 148–157. [Google Scholar] [CrossRef]
- Meddah, B.; Ducroc, R.; Faouzi, M.E.A.; Eto, B.; Mahraoui, L.; Benhaddou-Andaloussi, A.; Martineau, L.C.; Cherrah, Y.; Haddad, P.S. Nigella sativa inhibits intestinal glucose absorption and improves glucose tolerance in rats. J. Ethnopharmacol. 2009, 121, 419–424. [Google Scholar] [CrossRef]
- Boskabady, M.; Shafei, M.; Parsaee, H. Effects of aqueous and macerated extracts from Nigella sativa on guinea pig isolated heart activity. Die Pharmazie-An Int. J. Pharm. Sci. 2005, 60, 943–948. [Google Scholar]
- Rasoli, M.S.; Khalili, M.; Mohammadi, R.; Soleimani, A.; Kohzadi, R.; Ilkhanipour, M.; Heidari, R.; Golkari, S. The chemical composition of Nigella sativa L. and its extract effects on lipid peroxidation levels, total antioxidant capacity and catalase activity of the liver and kidney in rats under stress. Gene Cell Tissue 2018, 5, e61323. [Google Scholar] [CrossRef]
- Burtis, C.A.; Ashwood, E.R. Tietz Textbook of Clinical Chemistry, 2nd ed.; Amer Assn for Clinical Chemistry: St. Louis, MO, USA, 1994. [Google Scholar]
- Ugwah-Oguejiofor, C.J.; Okoli, C.O.; Ugwah, M.O.; Umaru, M.L.; Ogbulie, C.S.; Mshelia, H.E.; Umar, M.; Njan, A.A. Acute and sub-acute toxicity of aqueous extract of aerial parts of Caralluma dalzielii NE Brown in mice and rats. Heliyon 2019, 5, e01179. [Google Scholar] [CrossRef] [Green Version]
- Lum, G.; Leal-Khouri, S. Significance of low serum urea nitrogen concentrations. Clin. Chem. 1989, 35, 639–640. [Google Scholar] [CrossRef]
- Yacine, B.; Noureddine, B.; Mustapha, T.; Samira, A.; Bachra, K. Acute and subchronic toxicity study of Nigella damascena methanolic seed extract in mice. Int. J. Pharm. Biol. Sci. 2013, 4, 413–419. [Google Scholar]
- Hanafy, M.; Hatem, M. Studies on the antimicrobial activity of Nigella sativa seed (black cumin). J. Ethnopharmacol. 1991, 34, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Hannan, A.; Saleem, S.; Chaudhary, S.; Barkaat, M.; Arshad, M.U. Anti bacterial activity of Nigella sativa against clinical isolates of methicillin resistant Staphylococcus aureus. J. Ayub. Med. Coll. 2008, 20, 72–74. [Google Scholar]
- Salem, E.M.; Yar, T.; Bamosa, A.O.; Al-Quorain, A.; Yasawy, M.I.; Alsulaiman, R.M.; Randhawa, M.A. Comparative study of Nigella Sativa and triple therapy in eradication of Helicobacter Pylori in patients with non-ulcer dyspepsia. Saudi J. Gastroenterol. 2010, 16, 207. [Google Scholar] [PubMed]
- Nadaf, N.; Gawade, S.; Muniv, A.; Waghmare, S.; Jadhav, D.; Sonawane, K. Exploring anti-yeast activity of Nigella sativa seed extracts. Ind. Crops Prod. 2015, 77, 624–630. [Google Scholar] [CrossRef]
- Tanis, H.; Aygan, A.; Digrak, M. Antimicrobial activity of four nigella species grown in southern turkey. Int. J. Agric. Biol. 2009, 11, 771–774. [Google Scholar]
- Riaz, M.; Syed, M.; Chaudhary, F. Chemistry of the medicinal plants of the genus Nigella. Hamdard Med. 1996, 39, 40–45. [Google Scholar]
- Hasan, N.A.; Nawahwi, M.Z.; Ab Malek, H. Antimicrobial activity of Nigella sativa seed extract. Sains Malays 2013, 42, 143–147. [Google Scholar]
- Bakathir, H.; Abbas, N. Detection of the antibacterial effect of Nigella Sativa ground seeds with water. J. Tradit Complement. Altern. Med. 2011, 8, 159–164. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Mahmoudvand, H.; Sharififar, F.; Rahmat, S.; Tavakoli, R.; Saedi Dezaki, E.; Jahanbakhsh, S.; Sharifi, I. Evaluation of antileishmanial activity and cytotoxicity of the extracts of Berberis vulgaris and Nigella sativa against Leishmania tropica. Vector Borne Dis. 2014, 51, 294–299. [Google Scholar]
- Sheik Noor, M.; Jaikumar, K.; Babu, A.; Anand, D.; Saravanan, P. A study on the in vitro antifungal activity of Nigella sativa (Linn.) seed extract and its phytochemical screening using GC-MS analysis. World J. Pharm. Pharm. Sci. 2015, 4, 1003–1011. [Google Scholar]
- Ariamanesh, H.; Tamizi, N.; Yazdinezhad, A.; Salah, S.; Motamed, N.; Amanloo, S. The effectiveness of Nigella sativa alcoholic extract on the inhibition of Candida albicans colonization and formation of plaque on acrylic denture plates: An in vitro study. J. Dent. 2019, 20, 171. [Google Scholar]
- Stangarlin, J.; Schwan-Estrada, K.; Cruz, M.; Nozaki, M. Medicinal plants and alternative control of phytopathogens. Biotecnolog. Cienc. Desenvolv. 1999, 11, 16–21. [Google Scholar]
- Simões, C.; Amoros, M.; Girre, L. Mechanism of antiviral activity of triterpenoid saponins. Phytother. Res. 1999, 13, 323–328. [Google Scholar] [CrossRef]
- Halawani, E. Antibacterial activity of thymoquinone and thymohydroquinone of Nigella sativa L. and their interaction with some antibiotics. Adv. Biol. Res. 2009, 3, 148–152. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; AOAC: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Swain, T.; Hillis, W. The phenolic constituents of Prunus domestica. I.—The quantitative analysis of phenolic constituents. J. Sci. Food Agric. 1959, 10, 63–68. [Google Scholar] [CrossRef]
- Bahorun, T.; Gressier, B.; Trotin, F.; Brunet, C.; Dine, T.; Luyckx, M.; Vasseur, J.; Cazin, M.; Cazin, J.; Pinkas, M. Oxygen species scavenging activity of phenolic extracts from hawthorn fresh plant organs and pharmaceutical preparations. Arzneimittel-Forschung 1996, 46, 1086–1089. [Google Scholar]
- Hagerman, A.E.; Butler, L.G. Protein precipitation method for the quantitative determination of tannins. J. Agric. Food Chem. 1978, 26, 809–812. [Google Scholar] [CrossRef]
- Bourgou, S.; Ksouri, R.; Bellila, A.; Skandrani, I.; Falleh, H.; Marzouk, B. Phenolic composition and biological activities of Tunisian Nigella sativa L. shoots and roots. C R Biol. 2008, 331, 48–55. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Hasani, P.; Yasa, N.; Vosough-Ghanbari, S.; Mohammadirad, A.; Dehghan, G.; Abdollahi, M. In vivo antioxidant potential of Teucrium polium, as compared to α-tocopherol. Acta Pharm. 2007, 57, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansouri, A.; Embarek, G.; Kokkalou, E.; Kefalas, P. Phenolic profile and antioxidant activity of the Algerian ripe date palm fruit (Phoenix dactylifera). Food Chem. 2005, 89, 411–420. [Google Scholar] [CrossRef]
- Voidarou, C.; Alexopoulos, A.; Plessas, S.; Karapanou, A.; Mantzourani, I.; Stavropoulou, E.; Fotou, K.; Tzora, A.; Skoufos, I.; Bezirtzoglou, E. Antibacterial activity of different honeys against pathogenic bacteria. Anaerobe 2011, 17, 375–379. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parts | Nigella sativa L. | Nigella damascena L. |
|---|---|---|
| Stem | Erect, ribbed, and profusely branched. Plant height is 27.80–60 cm tall (50.08 ± 6.45 cm). | Erect, simple, or branched, 30–70 cm tall (60.67 ± 5.90 cm) |
| Leaves | Alternate, pinnatisect divided into three to nine lobes. | Alternate, two to three pinnate with very thin segments. |
| Inflorescence | Not involucrate. | Involucrate. |
| Flower | Terminal, solitary, star-shaped, 2–2.5 cm in diameter. | Terminal, solitary, 3.5–4.5 cm in diameter, surrounded by five involucral leaves forming involucres of bracts. |
| Sepal | Five petalloid sepals, lanceolate, light blue or white. | 5–25 petaloid sepals, clawed, blue or white in color. |
| Petal | Eight reduced petals; each petal is formed of two bases enclosing nectar-pocket. | Eight petals, smaller than sepals, which are located at the base of the stamens. |
| Fruit | Follicles (capsule), green, dry, and brown at maturity, longer than wide. Formed from 3–10 carpals; each carpel ends with elongated style that persists after ripening of the fruit. Seeds are inserted one on the other in the fruit. | Follicles, green, dry, and brown in late summer, large and inflated, composed of several follicles welded to the top, containing seeds, with a horizontally spreading persistent style. |
| Seed | Small, obpyramidal form with a broad base and has a rough appearance; 3.16 ± 0.03 mm length and 1.77 ± 0.02 mm wide, black at maturity, with hard teguments and transversely ribbed. | Small, domed, 2.45 ± 0.03 mm length and 1.73 ± 0.01 mm wide, black at maturity and striated, transversely ribbed, with hard teguments. |
| Root | Well-developed yellow taproot. | Well-developed yellow taproot. |
| Parameters | Nigella sativa L. Seeds | Nigella Damascena L. Seeds |
|---|---|---|
| Moisture | 4.71 ± 0.09 a | 3.41 ± 0.03 b |
| Crude fat | 42.3 ± 0.67 a | 34.3 ± 0.35 b |
| Crude protein | 21.60 ± 0.35 b | 25.10 ± 0.74 a |
| Total carbohydrates | 27.23 ± 0.41 b | 32.47 ± 0.39 a |
| Reducing sugars | 6.90 ± 0.39 a | 5.62 ± 0.14 b |
| Ash | 4.16 ± 0.12 b | 4.72 ± 0.09 a |
| Magnesium | 0.01 ± 0.002 a | 0.01 ± 0.001 a |
| Sodium | 0.33 ± 0.03 b | 0.56 ± 0.02 a |
| Parameters | NS Seeds | ND Seeds | Reference Standards |
|---|---|---|---|
| Phenolic contents | |||
| TEC (%) | 20.64 ± 0.08 b | 29.94 ± 0.11 a | - |
| TPC (mg GAE/100 g DW) | 628.98 ± 4.38 b | 1841.46 ± 9.60 a | - |
| TFC (mg QE/100 g DW) | 59.87 ± 0.22 b | 138.45 ± 0.32 a | - |
| Tannin (mg TAE/100 g DW) | 44.54 ± 0.82 b | 69.58 ± 3.14 a | - |
| Antioxidant activity | |||
| Quenching of DPPH• (IC50, mg/L) | 617.52 ± 0.01 a | 318.84 ± 0.02 b | 75.65 ± 2.10 α, 2.9 ± 0.02 β, 70.6 ± 6 γ |
| Quenching of ABTS•+ (IC50, mg/L) | 82.92 ± 0.02 a | 44.15 ± 0.02 b | 2.465 ± 0.003 δ, 1.244 ± 0.002 β, 0.502 ± 0.001 ε |
| Quenching of O2•− (IC50, mg/L) | 994.73 ± 0.02 a | 854.30 ± 0.01 b | 472 ± 10 δ, 139 ± 6 β, 84 ± 2GA ε |
| Reducing power (A0.5, mg/L) | 254.75 ± 0.03 a | 116.30 ± 0.01 b | 3.21 ± 0.001 β, 1.79 ± 0.01 ε |
| Phenolic Compound (mg/100 g Dry Weight) | Nigella sativa L. | Nigella damascena L. |
|---|---|---|
| Phenolic acids | ||
| 1. Gallic acid | 1.69 ± 0.02 b | 17.86 ± 0.06 a |
| 2. Hydroxybenzoic acid | 374.12 ± 0.37 a | 1010.71 ± 0.60 b |
| 3. Syringic acid | 7.40 ± 0.05 a | 3.44 ± 0.06 b |
| 4. Vanillic acid | 3.17 ± 0.01 b | 20.54 ± 0.08 a |
| 5. Caffeic acid | 111.06 ± 0.21 a | 66.40 ± 0.11 b |
| 6. Coumaric acid | 9.94 ± 0.09 a | 4.83 ± 0.06 b |
| 7. Cinnamic acid | 1.55 ± 0.03 a | 1.52 ± 0.06 a |
| Flavonoids | ||
| 8. Quercetin | 0.15 ± 0.03 b | 1.42 ± 0.07 a |
| 9. Catechin | 6.94 ± 0.05 b | 32.23 ± 0.06 a |
| Mice Group | Extract Dose (mg/kg) | Protein | Albumin | Glycemia | Urea | Cholesterol | Triglycerides |
|---|---|---|---|---|---|---|---|
| CTL | 0 | 46.19 ± 0.05 a | 27.24 ± 0.01 a | 2.50 ± 0.03 a | 0.77 ± 0.03 a | 1.70 ± 0.01 a | 1.92 ± 0.04 a |
| NS1 | 150 | 40.97 ± 0.01 b | 26.18 ± 0.04 b | 2.15 ± 0.06 b | 0.65 ± 0.02 b | 1.37 ± 0.02 bc | 1.81 ± 0.01 b |
| NS2 | 500 | 40.59 ± 0.04 c | 25.27 ± 0.04 c | 2.00 ± 0.05 c | 0.56 ± 0.05 c | 1.32 ± 0.02 cd | 1.41 ± 0.03 c |
| ND1 | 150 | 40.27 ± 0.03 d | 25.10 ± 0.06 d | 1.88 ± 0.02 d | 0.63 ± 0.06 bc | 1.39 ± 0.08 b | 1.80 ± 0.05 b |
| ND2 | 500 | 40.04 ± 0.02 e | 24.78 ± 0.02 e | 1.58 ± 0.02 e | 0.57 ± 0.01 c | 1.30 ± 0.05 d | 1.45 ± 0.07 c |
| Strains | NS Extract | ND Extract | ||
|---|---|---|---|---|
| 5 mg/mL | 195 mg/mL | 5 mg/mL | 195 mg/mL | |
| Gram-negative bacteria | ||||
| Escherichia coli | n.a. | n.a. | n.a. | n.a. |
| Pseudomonas aeruginosa | n.a. | n.a. | n.a. | 18 ± 0.50 |
| Gram-positive bacteria | ||||
| Methicillin-resistant Staphylococcus aureus | n.a. | 15 ± 0.30 a | n.a. | 14 ± 0.20 b |
| Methicillin-sensitive Staphylococcus aureus | n.a. | 14 ± 0.50 b | n.a. | 22 ± 0.50 a |
| Molds | ||||
| Aspergillus niger | n.a. | n.a. | n.a. | n.a. |
| Aspergillus flavus | n.a. | 16 ± 0.40 b | n.a | 20 ± 0.50 a |
| Mucor rammanianus | n.a. | 17 ± 0.40 a | 12 ± 0.20 | 14 ± 0.30 b |
| Aspergillus ochraceus | n.a. | n.a. | n.a. | 12 ± 0.50 |
| Aspergillus parasiticus | n.a. | 17 ± 0.30 b | 19 ± 0.40 | 22 ± 0.50 a |
| Yeast | ||||
| Candida albicans | n.a. | 13 ± 0.50 | n.a. | n.a. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benazzouz-Smail, L.; Achat, S.; Brahmi, F.; Bachir-Bey, M.; Arab, R.; Lorenzo, J.M.; Benbouriche, A.; Boudiab, K.; Hauchard, D.; Boulekbache, L.; et al. Biological Properties, Phenolic Profile, and Botanical Aspect of Nigella sativa L. and Nigella damascena L. Seeds: A Comparative Study. Molecules 2023, 28, 571. https://doi.org/10.3390/molecules28020571
Benazzouz-Smail L, Achat S, Brahmi F, Bachir-Bey M, Arab R, Lorenzo JM, Benbouriche A, Boudiab K, Hauchard D, Boulekbache L, et al. Biological Properties, Phenolic Profile, and Botanical Aspect of Nigella sativa L. and Nigella damascena L. Seeds: A Comparative Study. Molecules. 2023; 28(2):571. https://doi.org/10.3390/molecules28020571
Chicago/Turabian StyleBenazzouz-Smail, Leila, Sabiha Achat, Fatiha Brahmi, Mostapha Bachir-Bey, Radia Arab, José Manuel Lorenzo, Aicha Benbouriche, Kahina Boudiab, Didier Hauchard, Lila Boulekbache, and et al. 2023. "Biological Properties, Phenolic Profile, and Botanical Aspect of Nigella sativa L. and Nigella damascena L. Seeds: A Comparative Study" Molecules 28, no. 2: 571. https://doi.org/10.3390/molecules28020571