Sialic Acid-Siglec Axis as Molecular Checkpoints Targeting of Immune System: Smart Players in Pathology and Conventional Therapy


Abstract
:1. Introduction
2. Sialic Acid and Immune Recognition
3. Sialic Acid-Siglec Checkpoint in Human Pathology
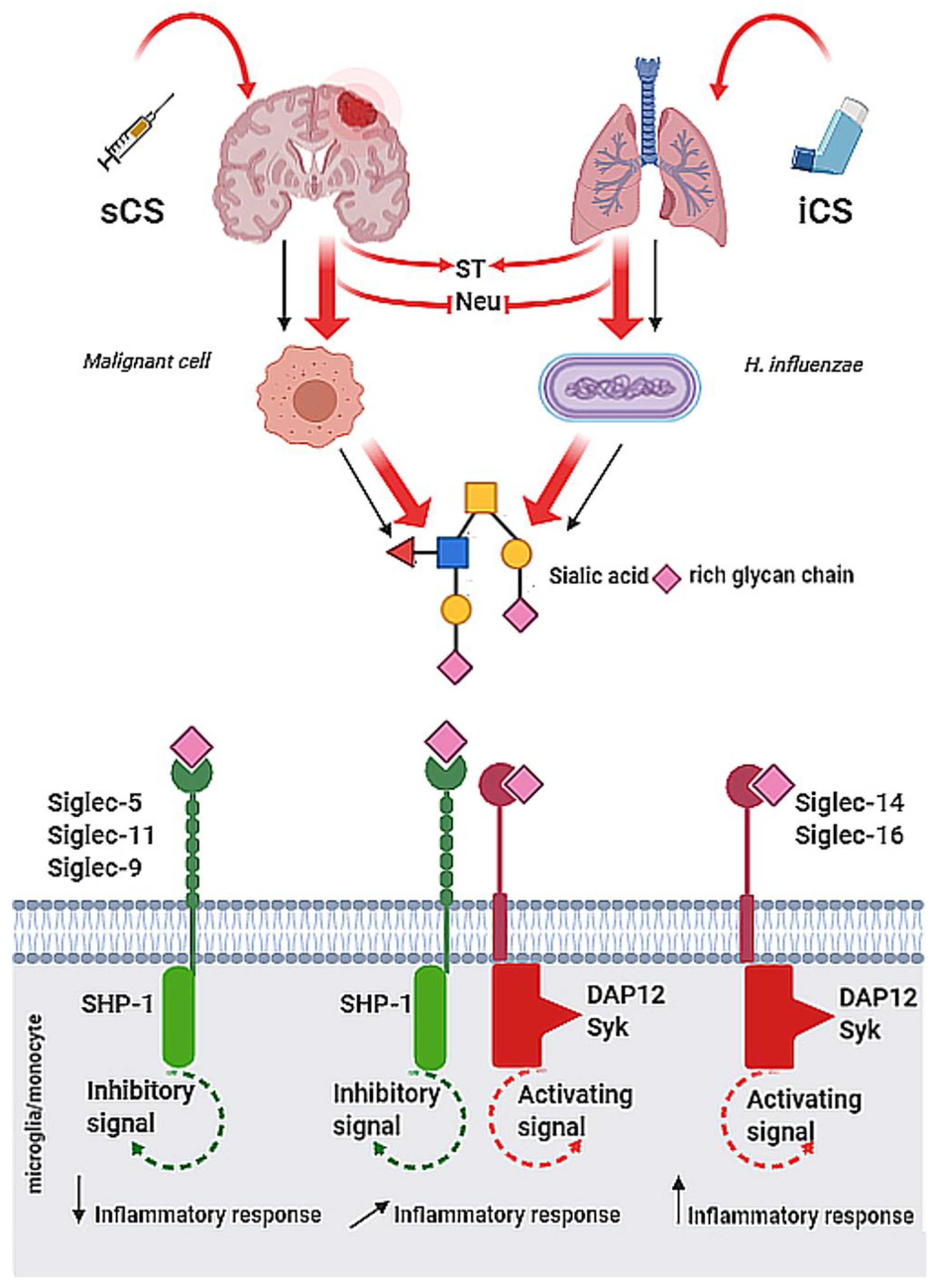
3.1. CNS Diseases
3.2. Respiratory System Disorders
3.3. Pathogen Invasion
3.4. Cancer Progression
3.5. Cardiovascular System Dysfunction
4. Sialic Acid-Siglec Checkpoint and Conventional Therapy
4.1. Sialidase Inhibitors—Not Only in Influenza Virus Infections
4.2. Sialic Acid-Siglec Axis and Standard Respiratory Obstruction Therapy
4.3. Corticosteroids—Benefits and Pitfalls in the Cancer Management
4.4. Anti-Inflammatory Management
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Crimeen-Irwin, B.; Scalzo, K.; Gloster, S.; Mottram, P.L.; Plebanski, M. Failure of immune homeostasis—The consequences of under and over reactivity. Curr. Drug Targets Immune. Endocr. Metabol. Disord. 2005, 5, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Huntington, N.D.; Gray, D.H. Immune homeostasis in health and disease. Immunol. Cell. Biol. 2018, 96, 451–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlin, H.; Piccinini, A.M. A complex interplay between the extracellular matrix and the innate immune response to microbial pathogens. Immunology 2018, 155, 186–201. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Zhang, Y.; Chen, Y.; Zhu, J.; Yang, Y.; Zhang, H.L. Role of microglia in neurological disorders and their potentials as a therapeutic target. Mol. Neurobiol. 2017, 54, 7567–7584. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Plüddemann, A.; Mukhopadhyay, S. Plasma membrane receptors of tissue macrophages: Functions and role in pathology. J. Pathol. 2020, 250, 656–666. [Google Scholar] [CrossRef]
- Huang, C.; Zhu, H.X.; Yao, Y.; Bian, Z.H.; Zheng, Y.J.; Li, L.; Moutsopoulos, H.M.; Gershwin, M.E.; Lian, Z.X. Immune checkpoint molecules. Possible future therapeutic implications in autoimmune diseases. J. Autoimmun. 2019, 104, 102333. [Google Scholar] [CrossRef]
- Rozin, A.P. From molecular mimicry to cross-reactivity or pathogen expansion? A hypothesis. Clin. Rheumatol. 2007, 26, 285–288. [Google Scholar] [CrossRef]
- Cusick, M.F.; Libbey, J.E.; Fujinami, R.S. Molecular mimicry as a mechanism of autoimmune disease. Clin. Rev. Allergy Immunol. 2012, 42, 102–111. [Google Scholar] [CrossRef]
- Guven-Maiorov, E.; Tsai, C.J.; Nussinov, R. Pathogen mimicry of host protein-protein interfaces modulates immunity. Semin. Cell. Dev. Biol. 2016, 58, 136–145. [Google Scholar] [CrossRef]
- Buchbinder, E.I.; Desai, A. CTLA-4 and PD-1 pathways: Similarities, differences, and implications of their inhibition. Am. J. Clin. Oncol. 2016, 39, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Goodman, A.; Patel, S.P.; Kurzrock, R. PD-1-PD-L1 immune-checkpoint blockade in B-cell lymphomas. Nat. Rev. Clin. Oncol. 2017, 14, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kang, H.; Lee, H.H.; Kim, C.W. Programmed cell death 1 (PD-1) and cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) in Viral Hepatitis. Int. J. Mol. Sci. 2017, 18, 1517. [Google Scholar] [CrossRef] [PubMed]
- Sperk, M.; Domselaar, R.V.; Neogi, U. Immune checkpoints as the immune system regulators and potential biomarkers in HIV-1 infection. Int. J. Mol. Sci. 2018, 19, 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotte, A. Combination of CTLA-4 and PD-1 blockers for treatment of cancer. J. Exp. Clin. Cancer Res. 2019, 38, 255. [Google Scholar] [CrossRef]
- Mahajan, V.S.; Pillai, S. Sialic acids and autoimmune disease. Immunol. Rev. 2016, 269, 145–161. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.Y.; Oswald, D.M.; Oliva, K.D.; Kreisman, L.S.C.; Cobb, B.A. The Glycoscience of immunity. Trends Immunol. 2018, 39, 523–535. [Google Scholar] [CrossRef]
- Varki, A. Sialic acids in human health and disease. Trends Mol. Med. 2008, 14, 351–360. [Google Scholar] [CrossRef] [Green Version]
- McMillan, S.J.; Crocker, P.R. CD33-related sialic-acid-binding immunoglobulin-like lectins in health and disease. Carbohydr. Res. 2008, 343, 2050–2056. [Google Scholar] [CrossRef]
- Läubli, H.; Varki, A. Sialic acid-binding immunoglobulin-like lectins (Siglecs) detect self-associated molecular patterns to regulate immune responses. Cell. Mol. Life Sci. 2020, 77, 593–605. [Google Scholar] [CrossRef]
- Schnaar, R.L. The Biology of gangliosides. Adv. Carbohydr. Chem. Biochem. 2019, 76, 113–148. [Google Scholar]
- Medus, M.L.; Gomez, G.E.; Zacchi, L.F.; Couto, P.M.; Labriola, C.A.; Labanda, M.S.; Bielsa, R.C.; Clérico, E.M.; Schulz, B.L.; Caramelo, J.J. N-glycosylation triggers a dual selection pressure in eukaryotic secretory proteins. Sci. Rep. 2017, 7, 8788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cragg, S.J.; Wagstaff, M.; Worwood, M. Sialic acid and the microheterogeneity of human serum ferritin. Clin. Sci. (Lond) 1980, 58, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Keel, B.A. Thyroid-stimulating hormone microheterogeneity. In Microheterogeneity of Glycoprotein Hormones; Keel, B.A., Grotjan, H.E., Eds.; CRC Press: Roca Raton, FL, USA; Taylor & Francis Group: Roca Raton, FL, USA, 2018; pp. 204–216. [Google Scholar]
- Miyagi, T.; Takahashi, K.; Hata, K.; Shiozaki, K.; Yamaguchi, K. Sialidase significance for cancer progression. Glycoconj. J. 2012, 29, 567–577. [Google Scholar] [CrossRef]
- Pearce, O.M.; Läubli, H. Sialic acids in cancer biology and immunity. Glycobiology 2016, 26, 111–128. [Google Scholar] [CrossRef] [Green Version]
- Vajaria, B.N.; Patel, K.R.; Begum, R.; Patel, P.S. Sialylation: An avenue to target cancer cells. Pathol. Oncol. Res. 2016, 22, 443–447. [Google Scholar] [CrossRef]
- Chiodelli, P.; Urbinati, C.; Paiardi, G.; Monti, E.; Rusnati, M. Sialic acid as a target for the development of novel antiangiogenic strategies. Future Med. Chem. 2018, 10, 2835–2854. [Google Scholar] [CrossRef]
- Bassagañas, S.; Pérez-Garay, M.; Peracaula, R. Cell surface sialic acid modulates extracellular matrix adhesion and migration in pancreatic adenocarcinoma cells. Pancreas 2014, 43, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Läubli, H.; Borsig, L. Altered cell adhesion and glycosylation promote cancer immune suppression and metastasis. Front. Immunol. 2019, 10, 2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, M.; Gottschalk, M. Streptococcus suis interactions with the murine macrophage cell line J774: Adhesion and cytotoxicity. Infect. Immun. 2002, 70, 4312–4322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakarya, S.; Ertem, G.T.; Oncu, S.; Kocak, I.; Erol, N.; Oncu, S. Escherichia coli bind to urinary bladder epithelium through nonspecific sialic acid mediated adherence. FEMS Immunol. Med. Microbiol. 2003, 39, 45–50. [Google Scholar] [CrossRef]
- Gaymard, A.; Le Briand, N.; Frobert, E.; Lina, B.; Escuret, V. Functional balance between neuraminidase and haemagglutinin in influenza viruses. Clin. Microbiol. Infect. 2016, 22, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, J.; Zheng, M.; Liu, Z.; Li, W.; Fu, X.; Lin, Y.; Yuan, J.; Zhao, J.; Shen, Q.; et al. A novel cardiovirus in wild rats. Virol. J. 2018, 15, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villar, E.; Barroso, I.M. Role of sialic acid-containing molecules in paramyxovirus entry into the host cell: A minireview. Glycoconj. J. 2006, 23, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Jaskiewicz, E.; Jodłowska, M.; Kaczmarek, R.; Zerka, A. Erythrocyte glycophorins as receptors for Plasmodium merozoites. Parasit. Vectors. 2019, 12, 317. [Google Scholar] [CrossRef] [Green Version]
- Trinchera, M.; Aronica, A.; Dall’Olio, F. Selectin ligands sialyl-Lewis a and sialyl-Lewis x in gastrointestinal cancers. Biology 2017, 6, 16. [Google Scholar] [CrossRef]
- Cagnoni, A.J.; Pérez Sáez, J.M.; Rabinovich, G.A.; Mariño, K.V. Turning-off signaling by siglecs, selectins, and galectins: Chemical inhibition of glycan-dependent interactions in cancer. Front. Oncol. 2016, 6, 109. [Google Scholar] [CrossRef] [Green Version]
- Wahrenbrock, M.G.; Varki, A. Multiple hepatic receptors cooperate to eliminate secretory mucins aberrantly entering the bloodstream: Are circulating cancer mucins the “tip of the iceberg”? Cancer Res. 2006, 66, 2433–2441. [Google Scholar] [CrossRef] [Green Version]
- Ngampasutadol, J.; Ram, S.; Gulati, S.; Agarwal, S.; Li, C.; Visintin, A.; Monks, B.; Madico, G.; Rice, P.A. Human factor H interacts selectively with Neisseria gonorrhoeae and results in species-specific complement evasion. J. Immunol. 2008, 180, 3426–3435. [Google Scholar] [CrossRef] [Green Version]
- Weiss, L. Neuraminidase, sialic acids, and cell interactions. J. Natl. Cancer Inst. 1973, 50, 3–19. [Google Scholar] [CrossRef] [Green Version]
- Adams, O.J.; Stanczak, M.A.; von Gunten, S.; Läubli, H. Targeting sialic acid-Siglec interactions to reverse immune suppression in cancer. Glycobiology 2018, 28, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Bärenwaldt, A.; Läubli, H. The sialoglycan-Siglec glyco-immune checkpoint—A target for improving innate and adaptive anti-cancer immunity. Expert Opin. Ther. Targets. 2019, 23, 839–853. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Netravali, I.A.; Cariappa, A.; Mattoo, H. Siglecs and immune regulation. Annu Rev. Immunol. 2012, 30, 357–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wuhrer, M.; Holst, S. Serum sialylation changes in cancer. Glycoconj. J. 2018, 35, 139–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loers, G.; Schachner, M. Recognition molecules and neural repair. J. Neurochem. 2007, 101, 865–882. [Google Scholar] [CrossRef]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Ruzafa, N.; Pereiro, X.; Aspichueta, P.; Araiz, J.; Vecino, E. The retina of osteopontin deficient mice in aging. Mol. Neurobiol. 2018, 55, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Wang, B. Sialic acid is an essential nutrient for brain development and cognition. Annu. Rev. Nutr. 2009, 29, 177–222. [Google Scholar] [CrossRef]
- Wang, Y.; Neumann, H. Alleviation of neurotoxicity by microglial human Siglec-11. J. Neurosci. 2010, 30, 3482–3488. [Google Scholar] [CrossRef] [Green Version]
- Wielgat, P.; Braszko, J.J. The participation of sialic acids in microglia-neuron interactions. Cell. Immunol. 2012, 273, 17–22. [Google Scholar] [CrossRef]
- Munkley, J. The role of sialyl-Tn in cancer. Int. J. Mol. Sci. 2016, 17, 275. [Google Scholar] [CrossRef] [Green Version]
- Waters, P.J.; Lewry, E.; Pennock, C.A. Measurement of sialic acid in serum and urine: Clinical applications and limitations. Ann. Clin. Biochem. 1992, 29, 625–637. [Google Scholar] [CrossRef]
- Moule, S.K.; Peak, M.; Thompson, S.; Turner, G.A. Studies of the sialylation and microheterogeneity of human serum alpha 1-acid glycoprotein in health and disease. Clin. Chim. Acta. 1987, 166, 177–185. [Google Scholar] [CrossRef]
- Katnik, I.; Goodarzi, M.T.; Turner, G.A. An improved ELISA for the determination of sialyl Lewis(x) structures on purified glycoconjugates. Glycoconj. J. 1996, 13, 1043–1047. [Google Scholar] [CrossRef]
- De Haan, N.; Reiding, K.R.; Haberger, M.; Reusch, D.; Falck, D.; Wuhrer, M. Linkage-specific sialic acid derivatization for MALDI-TOF-MS profiling of IgG glycopeptides. Anal. Chem. 2015, 87, 8284–8291. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zhu, H.; Li, L.; Guo, Y.; Gashash, E.; Ma, C.; Sun, X.; Li, J.; Zhang, L.; Wang, P.G. Sialic acid linkage-specific permethylation for improved profiling of protein glycosylation by MALDI-TOF MS. Anal. Chim. Acta 2017, 981, 53–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dědová, T.; Braicu, E.I.; Sehouli, J.; Blanchard, V. Sialic Acid Linkage Analysis Refines the Diagnosis of Ovarian Cancer. Front. Oncol. 2019, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Wielgat, P.; Walesiuk, A.; Braszko, J.J. Effects of chronic stress and corticosterone on sialidase activity in the rat hippocampus. Behav. Brain. Res. 2011, 222, 363–367. [Google Scholar] [CrossRef]
- Sandi, C.; Merino, J.J.; Cordero, M.I.; Touyarot, K.; Venero, C. Effects of chronic stress on contextual fear conditioning and the hippocampal expression of the neural cell adhesion molecule, its polysialylation, and L1. Neuroscience 2001, 102, 329–339. [Google Scholar] [CrossRef]
- Shahraz, A.; Kopatz, J.; Mathy, R.; Kappler, J.; Winter, D.; Kapoor, S.; Schütza, V.; Scheper, T.; Gieselmann, V.; Neumann, H. Anti-inflammatory activity of low molecular weight polysialic acid on human macrophages. Sci. Rep. 2015, 5, 16800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, S.S.; Matar, R.; Merheb, M.; Hodeify, R.; Vazhappilly, C.G.; Marton, J.; Shamsuddin, S.A.; Al Zouabi, H. Siglecs in Brain Function and Neurological Disorders. Cells 2019, 8, 1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielgat, P.; Holownia, A.; Braszko, J.J. Lipopolysaccharide changes sialylation pattern in the mouse central nervous system. J. Physiol. Pharmacol. 2012, 63, 555–561. [Google Scholar] [PubMed]
- Jungnickel, J.; Eckhardt, M.; Haastert-Talini, K.; Claus, P.; Bronzlik, P.; Lipokatic-Takacs, E.; Maier, H.; Gieselmann, V.; Grothe, C. Polysialyltransferase overexpression in Schwann cells mediates different effects during peripheral nerve regeneration. Glycobiology 2011, 22, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Li, Y.; Qin, K.; Du, B.; Li, T.; Yuan, H.; Han, C.; Luo, Y.; Du, B. Siglec-G Deficiency Ameliorates Hyper-Inflammation and Immune Collapse in Sepsis via Regulating Src Activation. Front. Immunol. 2019, 10, 2575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A.; Kaarniranta, K. Siglec receptors and hiding plaques in Alzheimer’s disease. J. Mol. Med. 2009, 87, 697–701. [Google Scholar] [CrossRef]
- Miles, L.A.; Hermans, S.J.; Crespi, G.A.; Gooi, J.H.; Doughty, L.; Nero, T.L.; Markulić, J.; Ebneth, A.; Wroblowski, B.; Oehlrich, D.; et al. Small Molecule Binding to Alzheimer Risk Factor CD33 Promotes Aβ Phagocytosis. iScience 2019, 19, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Estus, S.; Shaw, B.C.; Devanney, N.; Katsumata, Y.; Press, E.E.; Fardo, D.W. Evaluation of CD33 as a genetic risk factor for Alzheimer’s disease. Acta Neuropathol. 2019, 138, 187–199. [Google Scholar] [CrossRef]
- Hopperton, K.E.; Mohammad, D.; Trépanier, M.O.; Giuliano, V.; Bazinet, R.P. Markers of microglia in post-mortem brain samples from patients with Alzheimer’s disease: A systematic review. Mol. Psychiatry. 2018, 23, 177–198. [Google Scholar] [CrossRef]
- Griciuc, A.; Serrano-Pozo, A.; Parrado, A.R.; Lesinski, A.N.; Asselin, C.N.; Mullin, K.; Hooli, B.; Choi, S.H.; Hyman, B.T.; Tanzi, R.E. Alzheimer’s disease risk gene CD33 inhibits microglial uptake of amyloid beta. Neuron 2013, 78, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Mroz, R.; Holownia, A.; Wielgat, P.; Sitko, A.; Skopinski, T.; Chyczewska, E.; Braszko, J.J. Siglec-8 in Induced Sputum of COPD Patients. Atherosclerosis 2013, 788, 19–23. [Google Scholar] [CrossRef]
- Raclawska, D.S.; Ttofali, F.; Fletcher, A.A.; Harper, D.N.; Bochner, B.S.; Janssen, W.J.; Evans, C.M. Mucins and Their Sugars. Critical Mediators of Hyperreactivity and Inflammation. Ann. Am. Thorac. Soc. 2016, 13, S98–S99. [Google Scholar]
- Yu, H.; Gonzalez-Gil, A.; Wei, Y.; Fernandes, S.M.; Porell, R.N.; Vajn, K.; Paulson, J.C.; Nycholat, C.M.; Schnaar, R.L. Siglec-8 and Siglec-9 binding specificities and endogenous airway ligand distributions and properties. Glycobiology 2017, 27, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Youngblood, B.A.; Brock, E.C.; Leung, J.; Falahati, R.; Bochner, B.S.; Rasmussen, H.S.; Peterson, K.; Bebbington, C.; Tomasevic, N. Siglec-8 antibody reduces eosinophils and mast cells in a transgenic mouse model of eosinophilic gastroenteritis. JCI Insight 2019, 4, 126219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doherty, T.A.; Soroosh, P.; Broide, D.H.; Croft, M. CD4+ cells are required for chronic eosinophilic lung inflammation but not airway remodeling. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 296, L229–L235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiwamoto, T.; Kawasaki, N.; Paulson, J.C.; Bochner, B.S. Siglec-8 as a drugable target to treat eosinophil and mast cell-associated conditions. Pharmacol. Ther. 2012, 135, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Sugimoto, C.; Liu, D.X.; Midkiff, C.C.; Alvarez, X.; Lackner, A.A.; Kim, W.-K.; Didier, E.S.; Kuroda, M.J. Increased monocyte turnover is associated with interstitial macrophage accumulation and pulmonary tissue damage in SIV-infected rhesus macaques. J. Leukoc. Biol. 2015, 97, 1147–1153. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Hayakawa, T.; Yamanaka, M.; Varki, A.; Nakamura, M. Discovery of Siglec-14, a novel sialic acid receptor undergoing concerted evolution with Siglec-5 in primates. FASEB J. 2006, 20, 1964–1973. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Ishii, T.; Motegi, T.; Oka, R.; Taylor, R.E.; Soto, P.C.; Chang, Y.-C.; Secundino, I.; Gao, C.-X.; Ohtsubo, K.; et al. Loss of Siglec-14 reduces the risk of chronic obstructive pulmonary disease exacerbation. Cell. Mol. Life Sci. 2013, 70, 3199–3210. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Nizet, V. The interplay between Siglecs and sialylated pathogens. Glycobiology 2014, 24, 818–825. [Google Scholar] [CrossRef] [Green Version]
- Wielgat, P.; Trofimiuk, E.; Czarnomysy, R.; Holownia, A.; Braszko, J.J. Sialylation pattern in lung epithelial cell line and Siglecs expression in monocytic THP-1 cells as cellular indicators of cigarette smoke—Induced pathology in vitro. Exp. Lung Res. 2018, 44, 167–177. [Google Scholar] [CrossRef]
- Carlin, A.F.; Chang, Y.-C.; Areschoug, T.; Lindahl, G.; Hurtado-Ziola, N.; King, C.C.; Varki, A.; Nizet, V. Group B Streptococcus suppression of phagocyte functions by protein-mediated engagement of human Siglec-5. J. Exp. Med. 2009, 206, 1691–1699. [Google Scholar] [CrossRef]
- Ali, S.R.; Fong, J.J.; Carlin, A.F.; Busch, T.D.; Linden, R.; Angata, T.; Areschoug, T.; Parast, M.; Varki, N.; Murray, J.; et al. Siglec-5 and Siglec-14 are polymorphic paired receptors that modulate neutrophil and amnion signaling responses to group B Streptococcus. J. Exp. Med. 2014, 211, 1231–1242. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lan, C.; Ren, D.; Chen, G.Y. Induction of Siglec-1 by endotoxin tolerance suppresses the innate immune response by promoting TGF-β1 production. J. Biol. Chem. 2016, 291, 12370–12382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Pomares, L.; Gordon, S. Potential role of the mannose receptor in antigen transport. Immunol. Lett. 1999, 65, 9–13. [Google Scholar] [CrossRef]
- Nath, D.; Hartnell, A.; Happerfield, L.; Miles, D.W.; Burchell, J.M.; Taylor-Papadimitriou, J.; Crocker, P. Macrophage-tumour cell interactions: Identification of MUC1 on breast cancer cells as a potential counter-receptor for the macrophage-restricted receptor, sialoadhesin. Immunology 1999, 98, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Kumamoto, Y.; Higashi, N.; Denda-Nagai, K.; Tsuiji, M.; Sato, K.; Crocker, P.; Irimura, T. Identification of Sialoadhesin as a Dominant Lymph Node Counter-receptor for Mouse Macrophage Galactose-type C-type Lectin 1. J. Boil. Chem. 2004, 279, 49274–49280. [Google Scholar] [CrossRef] [Green Version]
- Kidder, D.; Richards, H.E.; Ziltener, H.J.; Garden, O.A.; Crocker, P.R. Sialoadhesin ligand expression identifies a subset of CD4+Foxp3- T cells with a distinct activation and glycosylation profile. J. Immunol. 2013, 190, 2593–2602. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.Y.; Brown, N.K.; Zheng, P.; Liu, Y. Siglec-G/10 in self-nonself discrimination of innate and adaptive immunity. Glycobiology 2014, 24, 800–806. [Google Scholar] [CrossRef]
- Mikulak, J.; Di Vito, C.; Zaghi, E.; Mavilio, D. Host Immune Responses in HIV-1 Infection: The Emerging Pathogenic Role of Siglecs and Their Clinical Correlates. Front. Immunol. 2017, 8, 14243. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Angata, T. Siglecs-the major subfamily of I-type lectins. Glycobiology 2006, 16, 1R–27R. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Ma, X.; Su, D.; Zhang, Y.; Yu, L.; Jiang, F.; Zhou, X.; Feng, Y.; Ma, F. The Roles of Siglec7 and Siglec9 on Natural Killer Cells in Virus Infection and Tumour Progression. J. Immunol. Res. 2020, 2020, 6243819. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, X.; Xu, Y.; Yang, H.; Gao, D.; Li, X.; Gao, L.; Ma, C.; Liang, X. Decreased Siglec-9 Expression on Natural Killer Cell Subset Associated With Persistent HBV Replication. Front. Immunol. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Pino, M.; Erkizia, I.; Benet, S.; Erikson, E.; Fernandez-Figueras, M.-T.; Guerrero, D.; Dalmau, J.; Ouchi, D.; Rausell, A.; Ciuffi, A.; et al. HIV-1 immune activation induces Siglec-1 expression and enhances viral trans-infection in blood and tissue myeloid cells. Retrovirology 2015, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Zou, Z.; Chastain, A.; Moir, S.; Ford, J.; Trandem, K.; Martinelli, E.; Cicala, C.; Crocker, P.; Arthos, J.; Sun, P.D. Siglecs Facilitate HIV-1 Infection of Macrophages through Adhesion with Viral Sialic Acids. PLoS ONE 2011, 6, e24559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.-D.; Klausing, S.; Hillier, M.; Maher, J.; Noll, T.; Crocker, P.R.; et al. The mucin MUC1 modulates the tumor immunological microenvironment through engagement of the lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraschilla, I.; Pillai, S. Viewing Siglecs through the lens of tumor immunology. Immunol. Rev. 2017, 276, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, E.; Macauley, M.S. Hypersialylation in Cancer: Modulation of Inflammation and Therapeutic Opportunities. Cancers 2018, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Wielgat, P.; Trofimiuk, E.; Czarnomysy, R.; Braszko, J.J.; Car, H. Sialic acids as cellular markers of immunomodulatory action of dexamethasone on glioma cells of different immunogenicity. Mol. Cell. Biochem. 2019, 455, 147–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielgat, P.; Czarnomysy, R.; Trofimiuk, E.; Car, H. The sialoglycan-Siglec-E checkpoint axis in dexamethasone-induced immune subversion in glioma-microglia transwell co-culture system. Immunol. Res. 2019, 67, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Engblom, C.; Pfirschke, C.; Zilionis, R.; Martins, J.D.S.; Bos, S.A.; Courties, G.; Rickelt, S.; Severe, N.; Baryawno, N.; Faget, J.; et al. Osteoblasts remotely supply lung tumors with cancer-promoting SiglecFhighneutrophils. Science 2017, 358, eaal5081. [Google Scholar] [CrossRef] [Green Version]
- Amoureux, M.C.; Coulibaly, B.; Chinot, O.; Loundou, A.; Metellus, P.; Rougon, G.; Figarella-Branger, D. Polysialic acid neural cell adhesion molecule (PSA-NCAM) is an adverse prognosis factor in glioblastoma, and regulates olig2 expression in glioma cell lines. BMC Cancer 2010, 10, 1–12. [Google Scholar] [CrossRef]
- Lin, C.H.; Yeh, Y.C.; Yang, K.D. Functions and therapeutic targets of Siglec-mediated infections, inflammations and cancers. J. Formos. Med. Assoc. 2019. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, R.; Ohtsubo, K.; Takamatsu, S.; Taniguchi, N.; Angata, T. The interaction between Siglec-15 and tumor-associated sialyl-Tn antigen enhances TGF-β secretion from monocytes/macrophages through the DAP12-Syk pathway. Glycobiology 2013, 23, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spence, S.; Greene, M.K.; Fay, F.; Hams, E.; Saunders, S.P.; Hamid, U.; Fitzgerald, M.; Beck, J.; Bains, B.K.; Smyth, P.; et al. Targeting Siglecs with a sialic acid–decorated nanoparticle abrogates inflammation. Sci. Transl. Med. 2015, 7, 303ra140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büll, C.; Boltje, T.J.; Wassink, M.; De Graaf, A.M.A.; Van Delft, F.L.; Brok, M.H.D.; Adema, G.J. Targeting Aberrant Sialylation in Cancer Cells Using a Fluorinated Sialic Acid Analog Impairs Adhesion, Migration, and In Vivo Tumor Growth. Mol. Cancer Ther. 2013, 12, 1935–1946. [Google Scholar] [CrossRef] [Green Version]
- Heise, T.; Pijnenborg, J.F.A.; Büll, C.; Van Hilten, N.; Kers-Rebel, E.D.; Balneger, N.; Elferink, H.; Adema, G.J.; Boltje, T.J. Potent Metabolic Sialylation Inhibitors Based on C-5-Modified Fluorinated Sialic Acids. J. Med. Chem. 2018, 62, 1014–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, J.; Liu, Y.; Xu, D. Sialic acid metabolism as a potential therapeutic target of atherosclerosis. Lipids Health Dis. 2019, 18, 173. [Google Scholar] [CrossRef] [Green Version]
- Gruber, S.; Hendrikx, T.; Tsiantoulas, D.; Ozsvar-Kozma, M.; Göderle, L.; Mallat, Z.; Witztum, J.L.; Shiri-Sverdlov, R.; Nitschke, L.; Binder, C.J. Sialic Acid-Binding Immunoglobulin-like Lectin G Promotes Atherosclerosis and Liver Inflammation by Suppressing the Protective Functions of B-1 Cells. Cell Rep. 2016, 14, 2348–2361. [Google Scholar] [CrossRef] [Green Version]
- Hartnell, A.; Steel, J.; Turley, H.; Jones, M.; Jackson, D.G.; Crocker, P. Characterization of human sialoadhesin, a sialic acid binding receptor expressed by resident and inflammatory macrophage populations. Blood 2001, 97, 288–296. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.-S.; Zhou, Y.-H.; Rong, G.-H.; Wu, W.-L.; Liang, Y.; Yang, Z.; Geng, H.-L.; Ren-Qian, Z. Siglec-1 on monocytes is a potential risk marker for monitoring disease severity in coronary artery disease. Clin. Biochem. 2009, 42, 1057–1063. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Li, J.; Nie, L.; Hu, Y.; Wang, F.; Liu, H.; Fernandes, S.M.; Zhong, Q.; Li, X.; et al. Immunoregulatory Siglec ligands are abundant in human and mouse aorta and are up-regulated by high glucose. Life Sci. 2019, 216, 189–199. [Google Scholar] [CrossRef]
- De Mello, C.P.P.; Drusano, G.L.; Adams, J.R.; Shudt, M.; Kulawy, R.; Brown, A. Oseltamivir-zanamivir combination therapy suppresses drug-resistant H1N1 influenza A viruses in the hollow fiber infection model (HFIM) system. Eur. J. Pharm. Sci. 2018, 111, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Enioutina, E.Y.; Daynes, R.A. The control of IL-4 gene expression in activated murine T lymphocytes: A novel role for neu-1 sialidase. J. Immunol. 1997, 158, 3070–3080. [Google Scholar] [PubMed]
- Haxho, F.; Neufeld, R.J.; Szewczuk, M.R. Neuraminidase-1: A novel therapeutic target in multistage tumorigenesis. Oncotarget 2016, 7, 40860–40881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.-Y.; Brown, N.; Wu, W.; Khedri, Z.; Yu, H.; Chen, X.; Van De Vlekkert, D.; D’Azzo, A.; Zheng, P.; Liu, Y. Broad and direct interaction between TLR and Siglec families of pattern recognition receptors and its regulation by Neu1. Life 2014, 3, e04066. [Google Scholar] [CrossRef]
- Ando, M.; Shoji, T.; Tu, W.; Higuchi, H.; Nishijima, K.-I.; Iijima, S. Lectin-dependent localization of cell surface sialic acid-binding lectin Siglec-9. Cytotechnology 2014, 67, 601–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmich, N.N.; Sivak, K.V.; Chubarev, V.N.; Porozov, Y.B.; Savateeva-Lyubimova, T.N.; Peri, F.; Porozov, Y.B. TLR4 Signaling Pathway Modulators as Potential Therapeutics in Inflammation and Sepsis. Vaccines 2017, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wei, T.-T.; Li, Y.; Li, J.; Fan, Y.; Huang, F.-Q.; Cai, Y.-Y.; Ma, G.; Liu, J.; Chen, Q.-Q.; et al. Functional Metabolomics Characterizes a Key Role forN-Acetyl-Neuraminic Acid in Coronary Artery Diseases. Circulation 2017, 137, 1374–1390. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, E.P.; Hyun, S.W.; Feng, C.; Zhang, L.; Liu, A.; Guang, W.; Nguyen, C.; Luzina, I.G.; Atamas, S.P.; Passaniti, A.; et al. NEU1 Sialidase Expressed in Human Airway Epithelia Regulates Epidermal Growth Factor Receptor (EGFR) and MUC1 Protein Signaling. J. Biol. Chem. 2012, 287, 8214–8231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, K.; Szewczuk, M.R.; Rousseau, D.; Neufeld, R.J. Oseltamivir phosphate released from injectable Pickering emulsions over an extended term disables human pancreatic cancer cell survival. Oncotarget 2018, 9, 12754–12768. [Google Scholar] [CrossRef] [Green Version]
- Muneswarao, J.; Hassali, M.A.; Ibrahim, B.; Saini, B.; Ali, I.A.H.; Verma, A.K. It is time to change the way we manage mild asthma: An update in GINA 2019. Respir. Res. 2019, 20, 183–186. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Agrawal, S.; Chakrabarti, S.; Ish, P. COPD 2020 Guidelines—What is new and why? Adv. Respir. Med. 2020, 88, 38–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumeier, A.; Keith, R. Clinical Guideline Highlights for the Hospitalist: The GOLD and NICE Guidelines for the Management of COPD. J. Hosp. Med. 2020, 15, e1–e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, A.G. Inhaled Corticosteroid Treatment in Chronic Obstructive Pulmonary Disease (COPD): Boon or Bane? J. Am. Board Fam. Med. 2020, 33, 289–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielgat, P.; Mroz, R.M.; Stasiak-Barmuta, A.; Szepiel, P.; Chyczewska, E.; Braszko, J.J.; Holownia, A. Inhaled corticosteroids increase siglec-5/14 expression in sputum cells of COPD patients. Adv. Exp. Med. Biol. 2015, 839, 1–5. [Google Scholar]
- Zeng, Z.; Li, M.; Wang, M.; Wu, X.; Li, Q.; Ning, Q.; Zhao, J.; Xu, Y.; Xie, J. Increased expression of Siglec-9 in chronic obstructive pulmonary disease. Sci. Rep. 2017, 7, 10116. [Google Scholar] [CrossRef] [Green Version]
- Steinke, J.W.; Liu, L.; Huyett, P.; Negri, J.; Payne, S.; Borish, L. Prominent role of IFN-γ in patients with aspirin-exacerbated respiratory disease. J. Allergy Clin. Immunol. 2013, 132, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, K.; Schned, A.; Fortuny, J.; Heaney, J.; Marsit, C.J.; Kelsey, K.T.; Karagas, M.R. Glucocorticoid therapy and risk of bladder cancer. Br. J. Cancer 2009, 101, 1316–1320. [Google Scholar] [CrossRef]
- Klement, R.J.; Champ, C.E. Corticosteroids compromise survival in glioblastoma in part through their elevation of blood glucose levels. Brain 2017, 140, e16. [Google Scholar] [CrossRef] [Green Version]
- Ayroldi, E.; Cannarile, L.; Adorisio, S.; Delfino, D.V.; Riccardi, C. Role of Endogenous Glucocorticoids in Cancer in the Elderly. Int. J. Mol. Sci. 2018, 19, 3774. [Google Scholar] [CrossRef] [Green Version]
- Kelly, W.J.; Giles, A.J.; Gilbert, M. T lymphocyte-targeted immune checkpoint modulation in glioma. J. Immunother. Cancer 2020, 8, e000379. [Google Scholar] [CrossRef]
- Ryken, T.C.; Kuo, J.S.; Prabhu, R.S.; Sherman, J.H.; Kalkanis, S.N.; Olson, J.J. Congress of Neurological Surgeons Systematic Review and Evidence-Based Guidelines on the Role of Steroids in the Treatment of Adults With Metastatic Brain Tumors. Neurosurgery 2019, 84, E189–E191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitter, K.L.; Tamagno, I.; Alikhanyan, K.; Hosni-Ahmed, A.; Pattwell, S.S.; Donnola, S.; Dai, C.; Ozawa, T.; Chang, M.; Chan, T.A.; et al. Corticosteroids compromise survival in glioblastoma. Brain 2016, 139, 1458–1471. [Google Scholar] [CrossRef] [Green Version]
- Dubinski, D.; Hattingen, E.; Senft, C.; Seifert, V.; Peters, K.G.; Reiss, Y.; Devraj, K.; Plate, K.H. Controversial roles for dexamethasone in glioblastoma—Opportunities for novel vascular targeting therapies. Br. J. Pharmacol. 2019, 39, 1460–1468. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.Y.; Rudra, S.; Ma, S.; Campian, J.L.; Huang, J. Impact of overall corticosteroid exposure during chemoradiotherapy on lymphopenia and survival of glioblastoma patients. J. Neuro Oncol. 2019, 143, 129–136. [Google Scholar] [CrossRef]
- Petrelli, F.; Signorelli, D.; Ghidini, M.; Ghidini, A.; Pizzutilo, E.G.; Ruggieri, L.; Cabiddu, M.; Borgonovo, K.; Dognini, G.; Brighenti, M.; et al. Association of Steroids Use with Survival in Patients Treated with Immune Checkpoint Inhibitors: A Systematic Review and Meta-Analysis. Cancers 2020, 12, 546. [Google Scholar] [CrossRef] [Green Version]
- Petrelli, F.; De Stefani, A.; Ghidini, A.; Bruschieri, L.; Riboldi, V.; Dottorini, L.; Iaculli, A.; Zaniboni, A.; Trevisan, F. Steroids use and survival in patients with glioblastoma multiforme: A pooled analysis. J. Neurol. 2020, 1–8. [Google Scholar] [CrossRef]
- Silva, Z.; Ferro, T.; Almeida, D.; Soares, H.; Ferreira, J.A.; Deschepper, F.M.; Hensbergen, P.J.; Pirro, M.; Van Vliet, S.J.; Springer, S.; et al. MHC Class I Stability is Modulated by Cell Surface Sialylation in Human Dendritic Cells. Pharmaceutics 2020, 12, 249. [Google Scholar] [CrossRef] [Green Version]
- Dusoswa, S.A.; Horrevorts, S.K.; Ambrosini, M.; Kalay, H.; Paauw, N.J.; Nieuwland, R.; Pegtel, M.D.; Würdinger, T.; Van Kooyk, Y.; García-Vallejo, J.-J. Glycan modification of glioblastoma-derived extracellular vesicles enhances receptor-mediated targeting of dendritic cells. J. Extracell. Vesicles 2019, 8, 1648995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejstříková, E.; Kalina, T.; Trka, J.; Starý, J.; Hrusak, O. Correlation of CD33 with poorer prognosis in childhood ALL implicates a potential of anti-CD33 frontline therapy. Leukemia 2005, 19, 1092–1094. [Google Scholar] [CrossRef] [PubMed]
- Erduran, E.; Tekelioglu, Y.; Karakas, T.; Gedik, Y.; Mert, F.M.; Tekelioǧlu, Y. Comparision of the Apoptotic Effects on Lymphoblasts and on Increase of Myeloid Lineage Cells of a Short-Time, High-Dose Methylprednisolone and the Conventional-Dose Prednisolone Treatments in Children with Acute Lymphoblastic Leukemia. Pediatr. Hematol. Oncol. 2006, 23, 587–598. [Google Scholar] [CrossRef]
- Chevallier, P.; Huguet, F.; Raffoux, E.; Etienne, A.; Leguay, T.; Isnard, F.; Robillard, N.; Guillaume, T.; Delaunay, J.; Charbonnier, A.; et al. Vincristine, dexamethasone and epratuzumab for older relapsed/refractory CD22+ B-acute lymphoblastic leukemia patients: A phase II study. Haematologica 2014, 100, e128–e131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Dong, Y.; Wang, H.; Culley, D.J.; Marcantonio, E.R.; Crosby, G.; Tanzi, R.E.; Zhang, Y.; Xie, Z. Peripheral Surgical Wounding and Age-Dependent Neuroinflammation in Mice. PLoS ONE 2014, 9, e96752. [Google Scholar] [CrossRef] [PubMed]
- Teter, B.; Morihara, T.; Lim, G.; Chu, T.; Jones, M.; Zuo, X.; Paul, R.; Frautschy, S.; Cole, G. Curcumin restores innate immune Alzheimer’s disease risk gene expression to ameliorate Alzheimer pathogenesis. Neurobiol. Dis. 2019, 127, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Clancy, R.M.; Halushka, M.; Rasmussen, S.E.; Lhakhang, T.; Chang, M.; Buyon, J.P. Siglec-1 Macrophages and the Contribution of IFN to the Development of Autoimmune Congenital Heart Block. J. Immunol. 2018, 202, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Lisney, A.R.; Szelinski, F.; Reiter, K.; Burmester, G.R.; Rose, T.; Dörner, T. High maternal expression of SIGLEC1 on monocytes as a surrogate marker of a type I interferon signature is a risk factor for the development of autoimmune congenital heart block. Ann. Rheum. Dis. 2017, 76, 1476–1480. [Google Scholar] [CrossRef]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Tang, J.; Zheng, P.; Liu, Y. CD24 and Siglec-10 Selectively Repress Tissue Damage-Induced Immune Responses. Science 2009, 323, 1722–1725. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Pathology | Conventional Therapy/Drugs | Human/Murine Siglec Involved | References |
|---|---|---|---|
| Infections | zanamivir, oseltamivir | Siglec-5,-9,-E | [115,116] |
| COPD | corticosteroids, LAMA, LABA | Siglec-5/14, Siglec-8 | [64,125,126] |
| Asthma | aspirin | Siglec-8 | [127] |
| Brain tumors (in vitro models) | corticosteroids | Siglec-E,-F | [98,99] |
| Leukemia | corticosteroids | CD22, CD33 | [141,142] |
| Alzheimer’s’ Disease | ibuprofen, levobupivacaine | CD33 | [143] |
| Congenital Heart Block | Hydroxychloroquine | Siglec-1 | [146] |
| Liver injury | acetaminophen | Siglec-10 | [147,148] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wielgat, P.; Rogowski, K.; Niemirowicz-Laskowska, K.; Car, H. Sialic Acid-Siglec Axis as Molecular Checkpoints Targeting of Immune System: Smart Players in Pathology and Conventional Therapy. Int. J. Mol. Sci. 2020, 21, 4361. https://doi.org/10.3390/ijms21124361
Wielgat P, Rogowski K, Niemirowicz-Laskowska K, Car H. Sialic Acid-Siglec Axis as Molecular Checkpoints Targeting of Immune System: Smart Players in Pathology and Conventional Therapy. International Journal of Molecular Sciences. 2020; 21(12):4361. https://doi.org/10.3390/ijms21124361
Chicago/Turabian StyleWielgat, Przemyslaw, Karol Rogowski, Katarzyna Niemirowicz-Laskowska, and Halina Car. 2020. "Sialic Acid-Siglec Axis as Molecular Checkpoints Targeting of Immune System: Smart Players in Pathology and Conventional Therapy" International Journal of Molecular Sciences 21, no. 12: 4361. https://doi.org/10.3390/ijms21124361