
Non-Canonical Functions of Myeloperoxidase in Immune Regulation, Tissue Inflammation and Cancer

Department of Pathology, Feinberg School of Medicine, Northwestern University, 300 East Superior St., Chicago, IL 60611, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(20), 12250; https://doi.org/10.3390/ijms232012250
Submission received: 26 August 2022
/
Revised: 5 October 2022
/
Accepted: 11 October 2022
/
Published: 14 October 2022
(This article belongs to the Special Issue Myeloperoxidase)
Abstract
:Myeloperoxidase (MPO) is one of the most abundantly expressed proteins in neutrophils. It serves as a critical component of the antimicrobial defense system, facilitating microbial killing via generation of reactive oxygen species (ROS). Interestingly, emerging evidence indicates that in addition to the well-recognized canonical antimicrobial function of MPO, it can directly or indirectly impact immune cells and tissue responses in homeostatic and disease states. Here, we highlight the emerging non-canonical functions of MPO, including its impact on neutrophil longevity, activation and trafficking in inflammation, its interactions with other immune cells, and how these interactions shape disease outcomes. We further discuss MPO interactions with barrier forming endothelial and epithelial cells, specialized cells of the central nervous system (CNS) and its involvement in cancer progression. Such diverse function and the MPO association with numerous inflammatory disorders make it an attractive target for therapies aimed at resolving inflammation and limiting inflammation-associated tissue damage. However, while considering MPO inhibition as a potential therapy, one must account for the diverse impact of MPO activity on various cellular compartments both in health and disease.
1. Introduction
Myeloperoxidase (MPO) is a heme peroxidase enzyme predominantly expressed by polymorphonuclear neutrophils and to a much lesser degree monocytes and macrophage subsets. MPO is a major component of the anti-microbial defense system, facilitating the killing of ingested pathogens within the neutrophil phagosome [1]. MPO constitutes up to 5% of the dry cellular mass in activated neutrophils [2,3], consistent with its critical role as a neutrophil effector of host defense. This high molecular mass (~150 kDa) protein is produced in a sequential process and stored within the primary azurophilic granules of neutrophils until activation [4]. Upon neutrophil stimulation (for example, in response to phagocytosis of pathogenic bacteria), the majority of MPO is sequestered to the phagosome, however some of it may escape into the local inflamed environment, impacting the function of neutrophils and neighboring cells [5,6,7,8].
Given its inherent cytotoxicity, MPO escape into the surrounding tissue can lead to substantial cellular damage and exacerbate inflammation. There are several reports that illustrate the ability of MPO to cause extensive tissue damage after it is released from activated neutrophils (recently reviewed by [9,10]). The generation of MPO is tightly linked to neutrophil maturation in the bone marrow. As promyelocytes transition to become mature neutrophils, many proteolytic enzymes, including MPO are segregated in a tightly regulated sequential process into distinct granular compartments, to be released upon specific cues [11]. MPO is strictly localized to the azurophilic or primary granules, which are formed first in developing neutrophils at the promyelocyte stage [12]. In addition to MPO, azurophilic granules contain several other proteolytic enzymes, such as lysozyme, cathepsins, defensins and neutrophil elastase (NE), all of which play important roles in neutrophil effector function [13].
In maturing neutrophils, MPO synthesis begins with the generation of an inactive precursor known as apoproMPO [14]. ApoproMPO interacts with several chaperone molecules in the ER that allow for the addition of an iron protoporphyrin IX (heme) to generate proMPO [15]. Further processing produces the enzymatically active MPO molecule consisting of a homodimer with monomers composed of heavy and light chains, and with each heavy chain covalently linked to a heme group [14,16].
Upon neutrophil activation, an NADPH oxidase (NOX) complex, specifically NOX2 associates with the membrane of the phagosome to catalyze the transfer of electrons from cytoplasmic NADPH to molecular oxygen, giving rise to superoxide radical anions [17]. To limit extensive damage within the cell, superoxide is rapidly converted to hydrogen peroxide and oxygen, either spontaneously or via the action of superoxide dismutase [18]. MPO interacts with hydrogen peroxide to generate some of the strongest antimicrobial compounds, such as hypochlorous acid (HOCl), reactive halide species HOBr and HOSCN, and singlet oxygen [1,19]. Reactive oxidants generated by MPO within the phagosome exert their antimicrobial effects to provide host immunity [20,21,22,23].
Given the important function of MPO in host defense, its mode of action and the anti-bacterial activity has been extensively studied. Interestingly, in additional to its canonical function within the phagosome, MPO has been shown to directly bind select pathogens, and this has been proposed as a mechanism to locally target the activity of reactive oxidants to limit collateral tissue damage [24]. This was demonstrated to be due to the rapid consumption of H2O2 via the catalytic activity of MPO and others provide evidence for a similar protective effect of MPO on H2O2-induced cytotoxicity in murine macrophages [24,25]. Conversely, MPO can be self-degraded through the action of its catalytic products to provide a source of free iron, which can cause tissue damage via the formation of additional ROS and increase the risk of proliferation of pathogenic microbes [26]. Free iron can generate significant levels of toxic reactive oxidants through interactions with hydrogen peroxide, a process commonly referred to as the Fenton reaction. The formation of chloramines due to HOCl and other MPO oxidative products significantly impacts protein activity and structure (recently reviewed by [27]). Since proteins are a major target for HOCl in biological systems, 3-chlorotyrosine is often used as an effective marker of MPO activity [27,28,29,30,31]. MPO products have also been shown to impact the activity of other proteins. For example, HOCl can interact with zinc-histidine/cysteine clusters, impacting protein structure and disrupting enzymatic activity of yeast alcohol dehydrogenase [32].
Neutrophil extracellular trap (NET) formation is perhaps another intriguing non-canonical function of MPO [33]. NETs have been described as a mechanism of trapping and killing of invading pathogens, as well as promoting recruitment of other immune cells to combat microorganisms. NET formation follows nuclear DNA decondensation and requires the presence of MPO, neutrophil elastase (NE), and peptidylarginine deiminase type IV (PAD-4) [34,35]. How MPO activity mediates NETs release is not completely understood, but its presence on NETs themselves likely contributes to local ROS generation and killing of trapped pathogens [36]. Interestingly, recent work demonstrates the direct inhibition of NET-associated MPO by muscle-derived mesenchymal stem cells (MSCs) [37]. MSCs are known to elicit significant immunomodulatory effects, including the ability to limit excessive inflammation (reviewed by [38,39,40]). Franck and colleagues demonstrated that MSCs inhibit NET-MPO even after the release of NETs from activated neutrophils, indicating that MSCs may have therapeutic potential to treat autoimmune diseases or inflammatory conditions that are driven by the catalytic activity of MPO [37,41].
MPO is a well-established neutrophil marker and consistent with many pathological roles of neutrophils in inflammatory diseases and with the fact that although well-contained, it can leak into surrounding tissue, elevated MPO serum levels are found in several inflammatory conditions [42,43,44,45]. Despite frequent detection of partial or complete MPO deficiency in the population (~1:4000), in many cases patients experience no complications and can clear most bacterial, fungal, or viral infections [20,46]. This suggests redundancy (as one would expect for such critical function) or an induction of compensatory antimicrobial mechanisms in mammalian hosts. Clinical data also suggest that in some cases MPO-deficient individuals take longer to clear certain pathogens such as S. aureus and often experience recurring fungal infections with organisms such as C. albicans [47,48]. The antimicrobial activity of MPO is therefore an important aspect of healthy immune function, but evidence emerges for additional, regulatory roles for MPO.
Neutrophil hematopoiesis requires a substantial energy investiture, as ~2 × 1011 neutrophils are produced by the bone marrow every day. This roughly amounts 60% of the total energy devoted to hematopoiesis [11]. Synthesis of MPO protein, being ~5% of the dry mass of neutrophils, thus similarly represents a significant metabolic investment [3]. This highlights the likelihood of important regulatory functions of MPO, both canonical and non-canonical in mammalian hosts. In the following sections we will discuss the emerging non-canonical functions of MPO, including regulating neutrophil trafficking, signaling, interactions with other immune cells and its role in inflammatory diseases and cancer.
2. MPO and Inflammatory Disorders
As we have discussed above, neutrophils are a major source of MPO, and although most of it is contained within neutrophil granules or the lysosome, MPO release and accumulation in tissue, for example, endothelial cells has been observed [49,50,51]. Several mechanisms aiding in MPO dissemination to various tissue compartments have been described. For example, MPO released by circulating neutrophils can reach distal organs via binding to erythrocytes [52,53] or by being transported by plasma proteins such as albumin and various lipoproteins [49,54]. The interplay between MPO and proteins are likely mediated via electrostatic interactions due to the polycationic nature of the molecule. Ceruloplasmin can bind MPO and inhibit its catalytic activity, potentially contributing to MPO transport in the circulation [55]. MPO can also be shuttled by extracellular vesicles (EVs) or microparticles (MPs) to mediate potent effects on surrounding cells [5,56]. MPO localization to EVs is intriguing, especially given that fact that it requires a specific stimulus leading to mobilization and release of primary granules. These interactions allowing for the dissemination of MPO are thus likely potentiating the pathologic impact of MPO in inflammation.
MPO is a common inflammatory marker, and elevated MPO levels are observed in many pathological conditions, including inflammatory bowel disease (IBD) [45,57,58,59], cardiovascular disease [60,61,62,63], obesity [43,64,65], liver disease [66,67], arthritis [68], multiple sclerosis [69,70], Alzheimer’s disease [71], stroke [72] and cancer [73,74] (Table 1). MPO levels are elevated in IBD, and MPO inhibition in animal models of colitis is protective [58,75,76]. Similarly, MPO inhibition improves colonic wound healing, indicating an important detrimental role of MPO in colon inflammation [5]. In cardiovascular disease, MPO oxidizes low density lipoproteins, promoting the formation of atherosclerotic lesions and potentially contributing to plaque rupture [77,78]. MPO inhibition improves plaque stability in models of atherosclerosis and has been suggested as a target to prevent plaque rupture [79]. In the lungs, MPO is used as a marker for systemic inflammation in smokers, and elevated MPO levels are observed in patients with chronic obstructive pulmonary disease [80,81]. MPO inhibition ameliorates oxidative stress and decreases morbidity in a murine model of cystic fibrosis-related infection [82]. These few examples help illuminate the broad-spectrum of MPO activity in heath and disease. MPO also plays important roles in neuroinflammation and cancer, which we will elaborate upon in the following sections.
Interestingly in the context of various inflammatory diseases, MPO function is mainly considered as detrimental since the relative abundance of MPO positively correlates with disease severity [83]. This is consistent with MPO being a neutrophil marker and the evidence of neutrophil pathologic functions in disease. However, some reports implicate MPO activity as protective in specific disease settings [24,84,85,86]. Thus, while MPO plays a critical role in host defense via its antimicrobial activity, its emerging non-canonical roles both ROS-generation dependent and independent may play important roles in disease pathogenesis and progression. For example, MPO is protective in murine models of LPS-induced inflammation [87]. Mortality, hypothermia and proinflammatory cytokine release after LPS administration are increased in MPO KO mice compared with WTs, suggesting that MPO is beneficial in cases of sepsis [87]. Additionally, MPO may protect surrounding cells from excessive damage by effectively targeting reactive oxidant release specifically to the area of colonizing intracellular pathogens such as Salmonella [24]. Authors of this study demonstrated that MPO KO mice released high amounts of H2O2, resulting in significant tissue damage that is perpetuated by other immune effectors such as macrophages [24]. On the other hand, inhibition of MPO with a pharmacological agent (AZM198) attenuated degranulation, inflammation, and neutrophil-mediated endothelial damage in patients with antineutrophil cytoplasmic antibody (ANCA)-associated vasculitis [88]. These contrasting studies from different inflammatory disorders highlight the fact that MPO can be beneficial or detrimental in the development and progression of inflammation. The various impacts and the non-canonical functions associated with the MPO activity in inflammatory disorders are summarized in Table 1.

{kind=link}
Table 1.
Pathological roles of MPO in tissue inflammation and disease.
| Disease | MPO Activity | References |
|---|---|---|
| Cardiovascular Disease | Elevated MPO in atherosclerosis associated with disease severity MPO oxidizes low density lipoproteins, promoting the formation of atherosclerotic lesions MPO binds the ECM of smooth muscle cells and alters ECM composition MPO alters the rigidity of platelets, promoting aggregation and vascular inflammation | [60,77,89,90,91,92] |
| Neurodegenerative Disorders | MPO modulates neuronal, microglial cells, and astrocyte activity MPO-derived ROS cause vascular inflammation in the brain MPO accumulation is observed Alzheimer’s disease and contributes to severity of symptoms (demonstrated via improved health in KO mice) MPO levels positively correlate with MS severity | [69,70,71,93,94,95] |
| Cancer | MPO induces DNA damage and promotes mutational burden MPO enhances breast cancer progression in murine models MPO converts procarcinogens into carcinogens | [96,97,98] |
| Inflammatory Bowel Disease | Elevated MPO levels associated with increased morbidity MPO levels are higher in the stool of patients with active IBD MPO impairs resolution of mucosal injury via suppression of IEC migration and proliferation MPO inhibition limits severity of colitis MPO released on NETs exacerbates colitis, promotes tissue damage, and is positively correlated with severity of Crohn’s disease | [42,57,75,76,99] |
| Obesity | Obese individuals have significantly increased MPO in serum MPO participates in the regulation of obesity, however the specific function remains to be determined MPO inhibition attenuates liver damage associated with obesity | [65,100,101] |
| Pulmonary Inflammation | MPO degrades the glycocalyx, which is important in neutrophil adhesion and lung injury MPO deficiency promotes or suppresses inflammation in the lung in different models of pulmonary infection MPO inhibition shows limited efficacy as a potential therapeutic for tuberculosis | [82,102,103,104] |
| Arthritis | Experimentally induced arthritis is attenuated in MPO deficient individuals Elevated MPO levels promote oxidative stress in patients with arthritis | [68,105,106] |
| Ischemic Stroke | MPO levels positively correlate with the risk for recurrent acute stroke MPO inhibition is protective against inflammation-associated cell death following stroke | [107,108] |
3. Exogenous MPO Regulates Neutrophil Activation and Trafficking
Critical effector functions of neutrophils take place following activation and crossing of the vascular barrier as they navigate their way to sites of inflammation in the tissue. Regulatory mechanisms of neutrophil activation, including their interactions with barrier forming endothelial and epithelial cells have been extensively studied both in the bloodstream and tissues [109,110,111,112]. However, new molecular mechanisms for these vital processes are still emerging. This includes intriguing evidence of the roles of exogenous MPO in these processes.
Exogenous MPO has been suggested to mimic the action of proinflammatory cytokines by promoting activation of NF-κβ and MAPK signaling, leading to mobilization of intracellular calcium stores and neutrophil degranulation [113,114,115]. MPO can directly bind CD11b on human neutrophils, leading to nuclear translocation of NF-κβ, increased surface expression of CD11b and the release of chemokines and cytokines that promote inflammation [113]. In contrast, neutrophils from MPO KO mice showed an increased ability to engulf zymosan particles in vitro [116]. The authors found that zymosan-stimulated neutrophils from MPO KO animals had elevated surface expression of CD11b compared with WT animals, and that this increased expression was due to enhanced FAK and ERK pathway activity in MPO-deficient animals, leading to increased phagocytic ability [116]. Additional studies on zymosan-stimulated neutrophils demonstrate that MPO-deficiency promotes production of proinflammatory cytokines such as MIP-2, MIP-1α, MIP-1β, IL-1α, IL-1β and TNF-α in vitro and in vivo [85,117]. Thus, it appears that under certain conditions MPO may serve suppressive roles in neutrophil activation. Conversely, others demonstrate that MPO-deficiency attenuates LPS-induced acute lung inflammation and negatively regulates proinflammatory cytokine production [118]. This contrasting evidence suggests a functional dualism for MPO in both suppressing or activating neutrophil effector functions and that this likely depends on the experimental model or timing. Therefore, further research is warranted to uncover the specific role of exogenous MPO in neutrophil activation.
Interestingly, similar controversial evidence suggests that MPO can either delay neutrophil apoptosis (independently of its catalytic activity) [119] or promote apoptosis by enhancing surface phosphatidylserine (PS) expression [120]. In apoptotic neutrophils, MPO has been shown to co-localize with PS at the cell surface [121], suggesting a potential contribution to programmed cell death, but the role of MPO in the regulation of apoptosis remains undefined. Furthermore, MPO release correlates with the number of apoptotic cells in human patients undergoing dialysis, providing additional evidence that MPO is an important regulator of apoptosis [122]. The association of MPO with PS at the cell surface may also have implications for macrophage recruitment and subsequent clearance of apoptotic cells, therefore additional studies that investigate neutrophil-MPO-macrophage interactions would be an interesting avenue to explore.
After sensing inflammatory cues, neutrophils activate their migratory machinery to marginate towards the vessel wall and cross the endothelial barrier. One regulatory component of neutrophil transendothelial migration is the glycocalyx, which lines the apical endothelial surface. The glycocalyx serves to regulate solute exchange by preventing the movement of large or negatively charged particles (reviewed by [123,124]), however, the depth of the glycocalyx has also been shown to impact PMN adhesion in inflammatory models of pulmonary sepsis [103]. In this model, degradation of the glycocalyx by TNF-α and LPS was shown to promote neutrophil adhesion and enhance tissue damage [103]. Interestingly, circulating MPO has also been shown to reduce the thickness of the endothelial glycocalyx, thus likely indirectly assisting in neutrophil recruitment and transendothelial migration [104]. Given the cationic nature of MPO, it has been shown to bind to the endothelium and promote neutrophil binding by acting as linker between neutrophils and endothelial cells [125]. The ability of MPO to bind biological structures allows for targeted oxidative activity and is likely important in the induction of site-specific effects which may encourage neutrophil recruitment [9,90,125]. In contrast, recent work from our laboratory as well as another independent study demonstrated a negative regulatory role of MPO in neutrophil transendothelial migration [6,126]. Using in vivo intravital imaging and ex vivo adhesion/chemotaxis approaches we found that following neutrophil activation, MPO localization to the neutrophil surface was inhibitory towards neutrophil adhesion to endothelial cells and transendothelial migration [6]. In MPO KO mice or following antibody-mediated inhibition of MPO, the number of neutrophils migrating into inflamed tissue in several models of inflammation was significantly elevated, suggesting a protective or beneficial role for MPO. The above inconsistencies in the underlying MPO function in neutrophil trafficking suggest the potential for context-dependent roles and highlights the need for further investigations into MPO-directed migration.
4. MPO Can Serve as a Secondary Messenger to Inform Neighboring Cell Function
MPO can impact the function of many other cells residing in or recruited to inflamed sites. MPO can directly bind platelets [92], fibroblasts [127], endothelial cells [51], and macrophages [105]; however, detailed mechanistic evidence of its impact is still limited and includes both protective and detrimental functions. For example, MPO binding to platelets promotes neutrophil-platelet interactions, stimulating intracellular calcium release and cytoskeletal reorganization, which alters platelet rigidity [91,92]. This has been proposed to promote inflammation and tissue damage. On the other hand, MPO stimulates fibroblast cell migration and promotes the release of collagens I and VI, enhancing tissue regeneration in vivo [127].
Several studies report MPO binding to endothelial cells, and it has even been proposed that endothelial cells may express low levels of MPO or accumulate exogenous MPO [51,56]. Perplexingly, elevated MPO levels are observed in cardiovascular diseases in humans, yet MPO KO animals are more prone to developing atherosclerosis [128]. Further evidence of a beneficial role of MPO in vascular repair has been reported, via MPO-induced activation of ERK, FAK and Akt signaling, and subsequent tube formation [51]. Conversely, MPO inhibition prevents endothelial cell dysfunction in several murine models of vascular inflammation [129,130]. Likewise, neutrophil-derived microparticles containing active MPO exert a cytotoxic effect and damage endothelial cells, suggesting a proinflammatory role for MPO [56]. Finally, similar to endothelial cell studies, we have recently shown that neutrophil-derived microparticles shuttling active MPO destabilize the integrity of epithelial monolayers, resulting in disrupted intestinal epithelial cell proliferation, migration, and overall wound healing processes [5]. Given the evidence presented above, further work is required to dissect the circumstances of seemingly conflicting outcomes of released MPO activity in tissue.
5. MPO and Inflammation of the Central Nervous System
Neutrophils have recently emerged as important modulators of neuro-inflammation, specifically impacting the function of vasculature of the brain and immune effector cells of the central nervous system (CNS) (reviewed by [131]). For instance, neutrophil products such as proteinase-3 can degrade endothelial junctions within the cerebral vasculature [132]. Since neutrophils are involved in the maintenance of vascular homeostasis, it is not surprising that MPO has also been implicated in inflammatory conditions of the CNS. For example, neural MPO concentrations are elevated in models of sepsis and several neurodegenerative disorders [133,134]. MPO influences the activity of astrocytes, microglial cells, and neurons by promoting cell death and attenuating neuronal cell responses to stimuli [94]. Additionally, MPO promotes cognitive impairment in murine models of Alzheimer’s disease, as MPO KO animals showed improved performance in behavioral, learning and memory assessments [95].
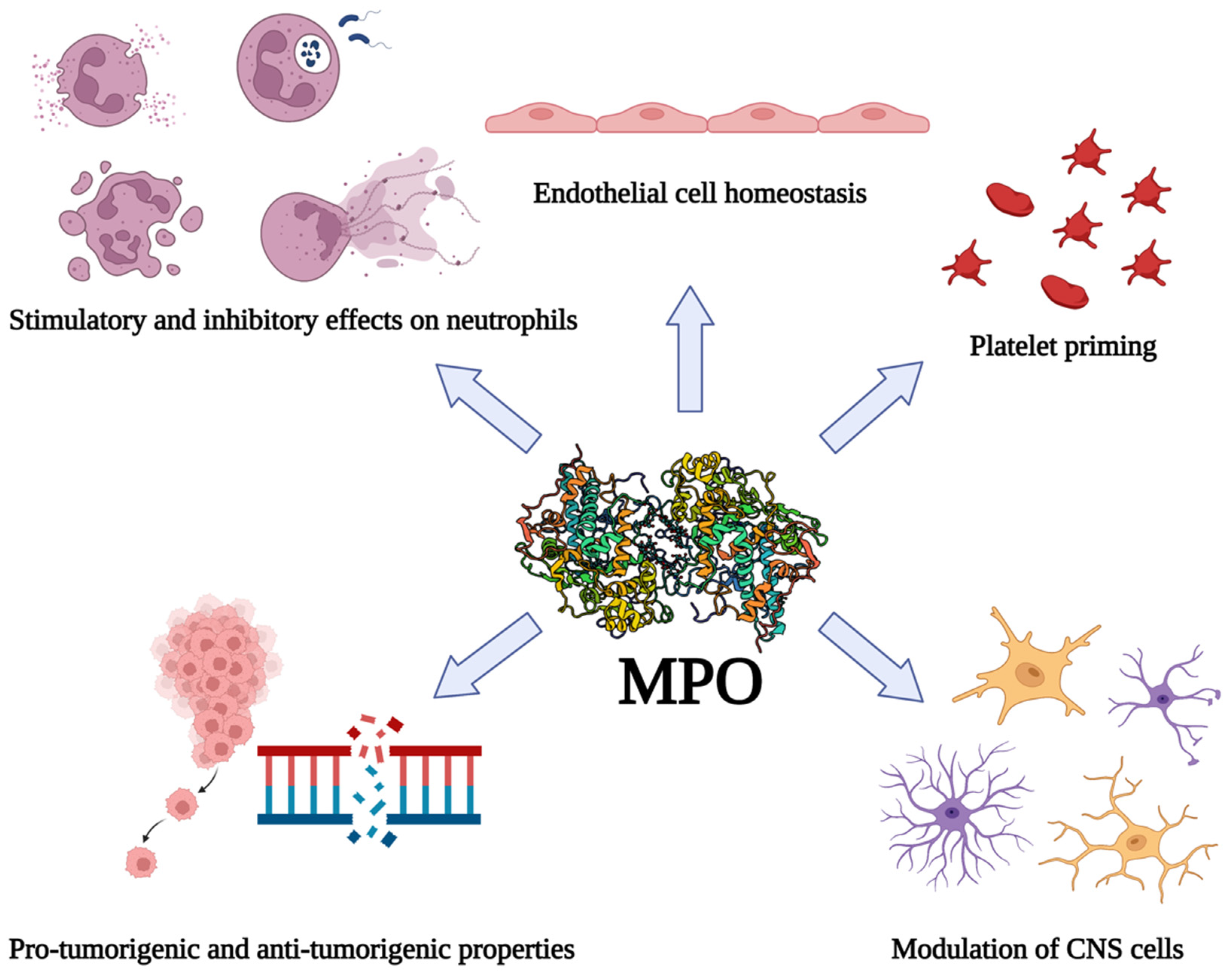
The catalytic activity of MPO results in the generation of substantial amounts of ROS and other reactive products such as HOCl, and these reactive compounds can impact homeostasis in the vasculature of the brain. For instance, hypothiocyanous acid disrupts the barrier function of brain endothelial cells, and other MPO-derived oxidants cause blood–brain barrier dysfunction [135,136,137]. For these reasons, MPO has been considered as a therapeutic target for several conditions related to inflamed brain tissue (reviewed by [138]). MPO KO mice are resistant to the neuronal cell dysfunction typically observed in murine models of ischemic stroke, and inhibition of MPO with 4-aminobenzoic acid hydrazide (ABAH) protects from tissue damage, suggesting that MPO is a realistic target for therapeutics to mitigate the inflammation associated with brain trauma [108]. Furthermore, MPO inhibition promotes cell proliferation/neurogenesis in these models, and therefore targeting MPO days after the initial injury may provide additional remedial value [139]. Other studies have investigated the efficacy of N-acetyl lysyltyrosylcysteine amide as a therapeutic for treating excess inflammation in the brain [140]. This tripeptide inhibitor of MPO has been shown to limit neuronal damage and inflammation in murine models of stroke and protects bovine aortic endothelial cells from MPO-mediated damage [140,141,142]. It is evident that MPO participates as a regulator of vascular and neuronal cell function in pathologies of the CNS, however further study is warranted to elucidate the specific mechanisms by which MPO directs inflammation in the brain. As described in the sections above, free MPO may possess certain anti-inflammatory benefits, such as inhibition of neutrophil adhesion and transendothelial migration, and so could act to limit tissue injury associated with neutrophil-mediated inflammation, however this has not yet been established in the CNS. The influences of MPO on various biological processes in tissue inflammation are summarized in Figure 1.
6. MPO and Cancer
As with tissue inflammation, neutrophils have emerged as important players in the development of many solid tumors. Tumor Associated Neutrophils (TANs) have been shown to be highly plastic, exerting both anti- and pro-tumorigenic activity. Intriguingly, such functional diversity has been suggested to be driven by the cytokine composition of the tumor niche, with TGF-β being one of the major drivers of pro-tumorigenic phenotype acquisition [143]. Alternatively, our group has recently identified temporal differences in the TAN function, where in early/low grade colon tumors TANs exhibited cytotoxic anti-tumorigenic activity, however in advanced CRC, TANs promoted tumor growth. The pro-tumorigenic function of TANs is commonly associated with oxidative burst and damaging ROS generation, to which MPO is a major contributor. Interestingly, MPO interactions with DNA were initially described as protective from oxidative damage [144]. However, it has now been well-established that in fact, MPO-generated ROS and other products such as HOCl potently oxidize and damage DNA, leading to elevated mutational burden and tumor development [145]. HOCl derived from MPO can damage nucleic acids via reactions that form chloramines and nitrogen-centered radicals [146,147,148,149,150]. MPO can also enter the nucleus and directly bind to DNA, as has been shown to occur during the initiation of NETosis to locally promote DNA oxidation [151]. Additionally, MPO can act on various procarcinogens to convert them to their subsequent carcinogenic forms [98]. This is particularly important in cancers related to cigarette smoke and patients with lung cancer, which have elevated MPO levels in lung tissue and serum. MPO-generated HOCl can cause structural changes, strand breaks and impact DNA binding properties, thereby leading to DNA irregularities, which ultimately may promote cancer progression [96]. As mentioned above, other MPO products such as HOBr and HOCl can oxidize components of DNA, resulting in the generation of 5-bromouracil, 5-chlorocytosine and other DNA-centered radicals and this has been proposed for consideration as a potential therapeutic target to prevent MPO-associated carcinogen formation [152,153]. Collectively, these studies demonstrate a major mechanism, where reactive oxidants generated by MPO can promote cancer progression. It is worth noting that a recent work from our laboratory has identified an additional MPO and ROS independent mechanism of PMN-driven DNA damage and genomic instability [154]. We found that extracellular vesicles/microparticles released by activated tissue PMNs shuttled proinflammatory miRNAs to neighboring epithelial cells to downregulate Lamin B1 and RAD-51, respectively destabilizing the nuclear envelope to promote double stranded break (DSB) generation and suppress DSB repair via homologous recombination [154]. Long term activity of these miRNAs potentiated the progression of colorectal cancer [155].
As has been discussed in previous sections, MPO is required for NET release and NETs have been linked to cancer progression in several preclinical models [156]. As with MPO function, NETs have been suggested to both promote and inhibit tumor progression. As such, NET release inhibited metastasis and exhibited cytotoxicity towards melanoma cells in vitro [157]. In contrast, NETs promoted metastasis in models of breast cancer [158], indicating that perhaps the outcome of NET activity is model-dependent and perhaps that of canonical vs. non-canonical MPO function.
Further highlighting the important role of MPO in tumorigenesis, both neutrophils, and specifically MPO have been corelated to tumor size in murine models of lung cancer [159,160]. While neutrophils promoted the development of butylated hydroxytoluene-driven lung carcinogenesis [159], developing tumors in MPO KO mice were significantly smaller compared to tumors in WT animals. In this model, inhibition of MPO with N-acetyl lysyltyrosylcysteine amide also reduced tumor size [160]. Similarly, the addition of exogenous MPO in a murine mammary tumor model stimulated significant increases in tumor size compared to control animals [97]. Authors in that work concluded that the observed impact of MPO on tumor growth was due to its interaction with surrounding fibroblasts to promote collagen release and remodeling of the ECM. In the same report, authors also observed augmented pulmonary metastasis in animals treated with exogenous MPO, providing further evidence for pro-tumorigenic properties of MPO. In contrast, MPO-derived HOCl has been suggested to protect against tumorigenesis by inhibition of NF-κβ and as such, MPO inhibition or in MPO KO mice melanoma growth was augmented [161]. Thus, the impact of MPO on tumor progression is likely multi-faceted, involving numerous signal transduction pathways and more work is warranted to gain a better understanding of the specific role of MPO in cancer.
7. Therapeutic Potential of Targeting MPO Activity
MPO targeting as a therapeutic for inflammatory disorders is an emerging area of research (reviewed by [162]). MPO has been advocated as a potential therapeutic target in essentially every inflammatory condition discussed throughout this review. The inhibition of MPO has been proposed as a strategy for resolution of inflammation in the context of IBD [57], atherosclerosis [130,163], neurodegenerative disorders [138,140], cancer [164], and infections [82,165]. While many of these studies show beneficial anti-inflammatory outcomes of MPO inhibition, it is important to remember that MPO can suppress inflammation in certain situations. Additionally, the evidence put forth here regarding the ability of MPO to act as an expansive regulator of inflammation should be carefully considered when attempting pan-inhibition of MPO as a therapeutic strategy. The likelihood for unanticipated off-target consequences is high when one considers the many cell types, processes, and pathways that are impacted by MPO. Furthermore, while designing strategies of MPO inhibition, one cannot disregard the fact that MPO serves a critical role in host defense.
Inhibitors of MPO are broadly classified as irreversible or reversible. Irreversible inhibitors such as the benzoyl hydrazides (eg. ABAH) and 2-thioxanthines act to disrupt the catalytic activity of MPO and have been shown to improve inflammation in several animal models [108,166,167]. For example, the synthetic 2-thioxanthine derivative AZD3241 limits inflammation and reduces the severity of DSS-induced experimental colitis in murine models [76]. Other irreversible 2-thioxanthine inhibitors of MPO, such as AZM198 and AZD5904 show promise as potential therapeutics in a variety of inflammatory models [82,88,101,168]. Reversible inhibitors of MPO include the small peptide N-acetyl lysyltyrosylcysteine amide, which has anti-tumorigenic effects and reduces inflammation in models of stroke and endothelial inflammation [140,142,160]. Many other compounds have been demonstrated to reversibly target MPO, for example hydroxamate compounds [169]. A comprehensive review of clinically relevant MPO inhibitors was recently published [170]. Several MPO inhibitors have gone to clinical trials, however; despite some progress and several assigned patents, there is no MPO inhibitor currently in use in the clinic today. This is likely due to the complex nature of the interactions of MPO with a diverse group of host cells, resulting in distinct physiological effects under different conditions.
Investigations into naturally occurring compounds in plants have revealed several potent inhibitors of MPO, and recently an automated assay to detect MPO inhibition was developed [171]. Alkaloids from the organism Peganum harmala can directly bind to the active site of MPO and inhibit its catalytic activity [172]. The plant has long been known to possess anti-inflammatory properties and this study demonstrates that MPO is the target of certain natural remedies [172,173]. Trigonelline, another plant alkaloid, reduces inflammation in rat models of ischemic stroke [174].
Similarly, various microorganisms have developed defensive mechanisms against MPO cytotoxicity. For example, Staphylococcus aureus produces a compound known as staphylococcal peroxidase inhibitor (SPIN), which is upregulated following phagocytosis and binds directly to MPO to protect the bacterium from MPO-mediated killing [175,176]. The presence of homologous MPO-inhibiting SPIN proteins was later confirmed in other species of Staphylococcus, suggesting that these immune evasion compounds are important factors for bacterial survival [177]. Another example is Propofol, which is a common general anesthetic drug with anti-inflammatory benefits, and which has recently been shown to at least in part exert its anti-inflammatory effect by inhibiting MPO [178,179]. These are just a few examples of natural compounds that impact MPO activity, however, given the exciting therapeutic potential of targeting MPO, many other reversible and irreversible MPO inhibitors have been developed or are currently in development through ongoing work.
In conclusion, the evidence put forth in this review suggest that MPO is an important regulator of host immune system function in the context of tissue inflammation. Ongoing research employing human tissues and MPO-deficient animals has already provided novel insights into how MPO may directly or indirectly impact the function of barrier forming epithelial and endothelial cells as well as various cells of the immune system. MPO inhibition strategies have emerged as interesting and relevant therapeutic approaches towards many inflammatory disorders, however, when designing such therapies, one must consider the multifaceted actions of MPO both within the phagosome (canonical MPO function) and interactions with other cells (the non-canonical MPO activity). Future studies that delve into the unknown mechanisms of MPO function will likely allow for the development of targeted therapeutic strategies to limit excessive inflammation.
Funding
This work was supported by grants from the American Cancer Society Research Scholar Award, National Institutes of Health NIDDK R01DK124199 and NIAID R01AI153568.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Winterbourn, C.C.; Hampton, M.B.; Livesey, J.H.; Kettle, A.J. Modeling the Reactions of Superoxide and Myeloperoxidase in the Neutrophil Phagosome: Implications for Microbial Killing. J. Biol. Chem. 2006, 281, 39860–39869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klebanoff, S.J. Myeloperoxidase: Friend and Foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, J.; Kaminker, K. Myeloperoxidase of the Leucocyte of Normal Human Blood. I. Content and Localization. Arch. Biochem. Biophys. 1962, 96, 465–467. [Google Scholar] [CrossRef]
- Hansson, M.; Olsson, I.; Nauseef, W.M. Biosynthesis, Processing, and Sorting of Human Myeloperoxidase. Arch. Biochem. Biophys. 2006, 445, 214–224. [Google Scholar] [CrossRef]
- Slater, T.W.; Finkielsztein, A.; Mascarenhas, L.A.; Mehl, L.C.; Butin-Israeli, V.; Sumagin, R. Neutrophil Microparticles Deliver Active Myeloperoxidase to Injured Mucosa To Inhibit Epithelial Wound Healing. J. Immunol. 2017, 198, 2886–2897. [Google Scholar] [CrossRef] [Green Version]
- Rehring, J.F.; Bui, T.M.; Galán-Enríquez, C.S.; Urbanczyk, J.M.; Ren, X.; Wiesolek, H.L.; Sullivan, D.P.; Sumagin, R. Released Myeloperoxidase Attenuates Neutrophil Migration and Accumulation in Inflamed Tissue. Front. Immunol. 2021, 12, 654259. [Google Scholar] [CrossRef]
- Ferrante, G.; Nakano, M.; Prati, F.; Niccoli, G.; Mallus, M.T.; Ramazzotti, V.; Montone, R.A.; Kolodgie, F.D.; Virmani, R.; Crea, F. High Levels of Systemic Myeloperoxidase Are Associated with Coronary Plaque Erosion in Patients with Acute Coronary Syndromes: A Clinicopathological Study. Circulation 2010, 122, 2505–2513. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Alsahli, M.; Rahmani, A. Myeloperoxidase as an Active Disease Biomarker: Recent Biochemical and Pathological Perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.L.; Davies, M.J. Role of Myeloperoxidase and Oxidant Formation in the Extracellular Environment in Inflammation-Induced Tissue Damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef]
- Borregaard, N. Neutrophils, from Marrow to Microbes. Immunity 2010, 33, 657–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedouhène, S.; Dang, P.M.C.; Hurtado-Nedelec, M.; El-Benna, J. Neutrophil Degranulation of Azurophil and Specific Granules. Methods Mol. Biol. 2020, 2087, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Cowland, J.B.; Borregaard, N. Granulopoiesis and Granules of Human Neutrophils. Immunol. Rev. 2016, 273, 11–28. [Google Scholar] [CrossRef]
- Grishkovskaya, X.I.; Paumann-Page, M.; Tscheliessnig, R.; Stampler, J.; Hofbauer, X.S.; Soudi, M.; Sevcnikar, B.; Oostenbrink, X.C.; Furtmüller, X.P.G.; Djinović-Carugo, X.K.; et al. Structure of Human Promyeloperoxidase (ProMPO) and the Role of the Propeptide in Processing and Maturation. J. Biol. Chem. 2017, 292, 8244–8261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauseef, W.M.; McCormick, S.J.; Goedken, M. Coordinated Participation of Calreticulin and Calnexin in the Biosynthesis of Myeloperoxidase. J. Biol. Chem. 1998, 273, 7107–7111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtmüller, P.G.; Zederbauer, M.; Jantschko, W.; Helm, J.; Bogner, M.; Jakopitsch, C.; Obinger, C. Active Site Structure and Catalytic Mechanisms of Human Peroxidases. Arch. Biochem. Biophys. 2006, 445, 199–213. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.C.; Gougerot-Pocidalo, M.A.; Dang, P.M.C. Priming of the Neutrophil Respiratory Burst: Role in Host Defense and Inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Karlsson, A.; Dahlgren, C. Assembly and Activation of the Neutrophil NADPH Oxidase in Granule Membranes. Antioxid. Redox Signal. 2002, 4, 49–60. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [Green Version]
- Klebanoff, S.J.; Kettle, A.J.; Rosen, H.; Winterbourn, C.C.; Nauseef, W.M. Myeloperoxidase: A Front-Line Defender against Phagocytosed Microorganisms. J. Leukoc. Biol. 2013, 93, 185–198. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J. Redox Reactions and Microbial Killing in the Neutrophil Phagosome. Antioxid. Redox Signal. 2013, 18, 642–660. [Google Scholar] [CrossRef] [PubMed]
- Paiva, C.N.; Bozza, M.T. Are Reactive Oxygen Species Always Detrimental to Pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivaloganathan, D.M.; Brynildsen, M.P. Phagosome-Bacteria Interactions from the Bottom Up. Annu. Rev. Chem. Biomol. Eng. 2021, 12, 309–331. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, N.; Forrer, P.; Casse, O.; Li, J.; Felmy, B.; Burgener, A.V.; Ehrenfeuchter, N.; Hardt, W.D.; Recher, M.; Hess, C.; et al. Myeloperoxidase Targets Oxidative Host Attacks to Salmonella and Prevents Collateral Tissue Damage. Nat. Microbiol. 2017, 2, 16268. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Sileikaite, I.; Davies, M.J.; Hawkins, C.L. Myeloperoxidase Modulates Hydrogen Peroxide Mediated Cellular Damage in Murine Macrophages. Antioxidants 2020, 9, 1255. [Google Scholar] [CrossRef]
- Maitra, D.; Shaeib, F.; Abdulhamid, I.; Abdulridha, R.M.; Saed, G.M.; Diamond, M.P.; Pennathur, S.; Abu-Soud, H.M. Myeloperoxidase Acts as a Source of Free Iron during Steady-State Catalysis by a Feedback Inhibitory Pathway. Free Radic. Biol. Med. 2013, 63, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.L. Hypochlorous Acid-Mediated Modification of Proteins and Its Consequences. Essays Biochem. 2020, 64, 75–86. [Google Scholar] [CrossRef]
- Hendrikje Buss, I.; Senthilmohan, R.; Darlow, B.A.; Mogridge, N.; Kettle, A.J.; Winterbourn, C.C. 3-Chlorotyrosine as a Marker of Protein Damage by Myeloperoxidase in Tracheal Aspirates from Preterm Infants: Association with Adverse Respiratory Outcome. Pediatr. Res. 2003, 53, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Nybo, T.; Dieterich, S.; Gamon, L.F.; Chuang, C.Y.; Hammer, A.; Hoefler, G.; Malle, E.; Rogowska-Wrzesinska, A.; Davies, M.J. Chlorination and Oxidation of the Extracellular Matrix Protein Laminin and Basement Membrane Extracts by Hypochlorous Acid and Myeloperoxidase. Redox Biol. 2019, 20, 496–513. [Google Scholar] [CrossRef]
- Nybo, T.; Cai, H.; Chuang, C.Y.; Gamon, L.F.; Rogowska-Wrzesinska, A.; Davies, M.J. Chlorination and Oxidation of Human Plasma Fibronectin by Myeloperoxidase-Derived Oxidants, and Its Consequences for Smooth Muscle Cell Function. Redox Biol. 2018, 19, 388–400. [Google Scholar] [CrossRef]
- Hazen, S.L.; Heinecke, J.W. 3-Chlorotyrosine, a Specific Marker of Myeloperoxidase-Catalyzed Oxidation, Is Markedly Elevated in Low Density Lipoprotein Isolated from Human Atherosclerotic Intima. J. Clin. Investig. 1997, 99, 2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, N.L.; Pattison, D.I.; Davies, M.J. Myeloperoxidase-Derived Oxidants Rapidly Oxidize and Disrupt Zinc–Cysteine/Histidine Clusters in Proteins. Free Radic. Biol. Med. 2012, 53, 2072–2080. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Rohrbach, A.S.; Slade, D.J.; Thompson, P.R.; Mowen, K.A. Activation of PAD4 in NET Formation. Front. Immunol. 2012, 3, 360. [Google Scholar] [CrossRef] [Green Version]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil Elastase and Myeloperoxidase Regulate the Formation of Neutrophil Extracellular Traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Franck, T.; Ceusters, J.; Graide, H.; Mouithys-Mickalad, A.; Serteyn, D. Muscle Derived Mesenchymal Stem Cells Inhibit the Activity of the Free and the Neutrophil Extracellular Trap (NET)-Bond Myeloperoxidase. Cells 2021, 10, 3486. [Google Scholar] [CrossRef]
- Gao, F.; Chiu, S.M.; Motan, D.A.L.; Zhang, Z.; Chen, L.; Ji, H.L.; Tse, H.F.; Fu, Q.L.; Lian, Q. Mesenchymal Stem Cells and Immunomodulation: Current Status and Future Prospects. Cell Death Dis. 2016, 7, e2062. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Xu, J. Immune Modulation by Mesenchymal Stem Cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef] [Green Version]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by Mesenchymal Stem Cells (MSCs): Mechanisms of Action of Living, Apoptotic, and Dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef]
- Odobasic, D.; Holdsworth, S.R. Emerging Cellular Therapies for Anti-Myeloperoxidase Vasculitis and Other Autoimmune Diseases. Front. Immunol. 2021, 12, 2964. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.G.B.; Sangfelt, P.; Wagner, M.; Hansson, T.; Lettesjö, H.; Carlson, M. Fecal Levels of Leukocyte Markers Reflect Disease Activity in Patients with Ulcerative Colitis. Scand. J. Clin. Lab. Investig. 2007, 67, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Olza, J.; Aguilera, C.M.; Gil-Campos, M.; Leis, R.; Bueno, G.; Martínez-Jiménez, M.D.; Valle, M.; Canẽte, R.; Tojo, R.; Moreno, L.A.; et al. Myeloperoxidase Is an Early Biomarker of Inflammation and Cardiovascular Risk in Prepubertal Obese Children. Diabetes Care 2012, 35, 2373–2376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldus, S.; Heeschen, C.; Meinertz, T.; Zeiher, A.M.; Eiserich, J.P.; Münzel, T.; Simoons, M.L.; Hamm, C.W. Myeloperoxidase Serum Levels Predict Risk in Patients with Acute Coronary Syndromes. Circulation 2003, 108, 1440–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansberry, D.R.; Shah, K.; Agarwal, P.; Agarwal, N. Fecal Myeloperoxidase as a Biomarker for Inflammatory Bowel Disease. Cureus 2017, 9, e1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parry, M.F.; Root, R.K.; Metcalf, J.A.; Delaney, K.K.; Kaplow, L.S.; Richar, W.J. Myeloperoxidase Deficiency. Prevalence and Clinical Significance. Ann. Intern. Med. 1981, 95, 293–301. [Google Scholar] [CrossRef]
- Hampton, M.B.; Kettle, A.J.; Winterbourn, C.C. Involvement of Superoxide and Myeloperoxidase in Oxygen-Dependent Killing of Staphylococcus Aureus by Neutrophils. Infect. Immun. 1996, 64, 3512–3517. [Google Scholar] [CrossRef] [Green Version]
- Aratani, Y.; Koyama, H.; Nyui, S.I.; Suzuki, K.; Kura, F.; Maeda, N. Severe Impairment in Early Host Defense against Candida Albicans in Mice Deficient in Myeloperoxidase. Infect. Immun. 1999, 67, 1828–1836. [Google Scholar] [CrossRef]
- Tiruppathi, C.; Naqvi, T.; Wu, Y.; Vogel, S.M.; Minshall, R.D.; Malik, A.B. Albumin Mediates the Transcytosis of Myeloperoxidase by Means of Caveolae in Endothelial Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 7699–7704. [Google Scholar] [CrossRef] [Green Version]
- Hartman, C.L.; Ford, D.A. MPO (Myeloperoxidase) Caused Endothelial Dysfunction. Arter. Thromb. Vasc. Biol. 2018, 38, 1676–1677. [Google Scholar] [CrossRef]
- Khalil, A.; Medfai, H.; Poelvoorde, P.; Kazan, M.F.; Delporte, C.; van Antwerpen, P.; EL-Makhour, Y.; Biston, P.; Delrée, P.; Badran, B.; et al. Myeloperoxidase Promotes Tube Formation, Triggers ERK1/2 and Akt Pathways and Is Expressed Endogenously in Endothelial Cells. Arch. Biochem. Biophys. 2018, 654, 55–69. [Google Scholar] [CrossRef]
- Adam, M.; Gajdova, S.; Kolarova, H.; Kubala, L.; Lau, D.; Geisler, A.; Ravekes, T.; Rudolph, V.; Tsao, P.S.; Blankenberg, S.; et al. Red Blood Cells Serve as Intravascular Carriers of Myeloperoxidase. J. Mol. Cell. Cardiol. 2014, 74, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Gorudko, I.V.; Sokolov, A.V.; Shamova, E.V.; Grigorieva, D.V.; Mironova, E.V.; Kudryavtsev, I.V.; Gusev, S.A.; Gusev, A.A.; Chekanov, A.V.; Vasilyev, V.B.; et al. Binding of Human Myeloperoxidase to Red Blood Cells: Molecular Targets and Biophysical Consequences at the Plasma Membrane Level. Arch. Biochem. Biophys. 2016, 591, 87–97. [Google Scholar] [CrossRef]
- Carr, A.C.; Myzak, M.C.; Stocker, R.; McCall, M.R.; Frei, B. Myeloperoxidase Binds to Low-Density Lipoprotein: Potential Implications for Atherosclerosis. FEBS Lett. 2000, 487, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Bakhautdin, B.; Goksoy Bakhautdin, E.; Fox, P.L. Ceruloplasmin Has Two Nearly Identical Sites That Bind Myeloperoxidase. Biochem. Biophys. Res. Commun. 2014, 453, 722–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitanga, T.N.; de Aragão França, L.; Rocha, V.C.J.; Meirelles, T.; Matos Borges, V.; Gonçalves, M.S.; Pontes-de-Carvalho, L.C.; Noronha-Dutra, A.A.; Dos-Santos, W.L.C. Neutrophil-Derived Microparticles Induce Myeloperoxidase-Mediated Damage of Vascular Endothelial Cells. BMC Cell Biol. 2014, 15, 21. [Google Scholar] [CrossRef] [Green Version]
- Chami, B.; Martin, N.J.J.; Dennis, J.M.; Witting, P.K. Myeloperoxidase in the Inflamed Colon: A Novel Target for Treating Inflammatory Bowel Disease. Arch. Biochem. Biophys. 2018, 645, 61–71. [Google Scholar] [CrossRef]
- Garrity-Park, M.; Loftus, E.V.; Sandborn, W.J.; Smyrk, T.C. Myeloperoxidase Immunohistochemistry as a Measure of Disease Activity in Ulcerative Colitis: Association with Ulcerative Colitis-Colorectal Cancer, Tumor Necrosis Factor Polymorphism and RUNX3 Methylation. Inflamm. Bowel Dis. 2012, 18, 275–283. [Google Scholar] [CrossRef]
- Lettesjö, H.; Hansson, T.; Peterson, C.; Ung, K.A.; Ringström, G.; Abrahamsson, H.; Simrén, M. Detection of Inflammatory Markers in Stools from Patients with Irritable Bowel Syndrome and Collagenous Colitis. Scand. J. Gastroenterol. 2009, 41, 54–59. [Google Scholar] [CrossRef]
- Kimak, E.; Ziȩba, B.; Duma, D.; Solski, J. Myeloperoxidase Level and Inflammatory Markers and Lipid and Lipoprotein Parameters in Stable Coronary Artery Disease. Lipids Health Dis. 2018, 17, 71. [Google Scholar] [CrossRef]
- Loria, V.; Dato, I.; Graziani, F.; Biasucci, L.M. Myeloperoxidase: A New Biomarker of Inflammation in Ischemic Heart Disease and Acute Coronary Syndromes. Mediat. Inflamm. 2008, 2008, 135625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkel, S.J.; Wolters, F.J.; Ikram, M.A.; de Maat, M.P.M. Circulating Myeloperoxidase (MPO)-DNA Complexes as Marker for Neutrophil Extracellular Traps (NETs) Levels and the Association with Cardiovascular Risk Factors in the General Population. PLoS ONE 2021, 16, e0253698. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Kremer Hovinga, J.A.; Schatzberg, D.; Wagner, D.D.; Lämmle, B. Circulating DNA and Myeloperoxidase Indicate Disease Activity in Patients with Thrombotic Microangiopathies. Blood 2012, 120, 1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, A.G.; Rodríguez, M.R.; Alonso, C.G.; Ochoa, D.Y.R.; Aguilar, C.A. Myeloperoxidase Is Associated with Insulin Resistance and Inflammation in Overweight Subjects with First-Degree Relatives with Type 2 Diabetes Mellitus. Diabetes Metab. J. 2015, 39, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Qaddoumi, M.G.; Alanbaei, M.; Hammad, M.M.; al Khairi, I.; Cherian, P.; Channanath, A.; Thanaraj, T.A.; Al-Mulla, F.; Abu-Farha, M.; Abubaker, J. Investigating the Role of Myeloperoxidase and Angiopoietin-like Protein 6 in Obesity and Diabetes. Sci. Rep. 2020, 10, 6170. [Google Scholar] [CrossRef] [Green Version]
- Rensen, S.S.; Slaats, Y.; Nijhuis, J.; Jans, A.; Bieghs, V.; Driessen, A.; Malle, E.; Greve, J.W.; Buurman, W.A. Increased Hepatic Myeloperoxidase Activity in Obese Subjects with Nonalcoholic Steatohepatitis. Am. J. Pathol. 2009, 175, 1473–1482. [Google Scholar] [CrossRef] [Green Version]
- Boussif, A.; Rolas, L.; Weiss, E.; Bouriche, H.; Moreau, R.; Périanin, A. Impaired Intracellular Signaling, Myeloperoxidase Release and Bactericidal Activity of Neutrophils from Patients with Alcoholic Cirrhosis. J. Hepatol. 2016, 64, 1041–1048. [Google Scholar] [CrossRef]
- Odobasic, D.; Yang, Y.; Muljadi, R.C.M.; O’Sullivan, K.M.; Kao, W.; Smith, M.; Morand, E.F.; Holdsworth, S.R. Endogenous Myeloperoxidase Is a Mediator of Joint Inflammation and Damage in Experimental Arthritis. Arthritis Rheumatol. 2014, 66, 907–917. [Google Scholar] [CrossRef]
- Nagra, R.M.; Becher, B.; Tourtellotte, W.W.; Antel, J.P.; Gold, D.; Paladino, T.; Smith, R.A.; Nelson, J.R.; Reynolds, W.F. Immunohistochemical and Genetic Evidence of Myeloperoxidase Involvement in Multiple Sclerosis. J. Neuroimmunol. 1997, 78, 97–107. [Google Scholar] [CrossRef]
- Minohara, M.; Matsuoka, T.; Li, W.; Osoegawa, M.; Ishizu, T.; Ohyagi, Y.; Kira, J. ichi Upregulation of Myeloperoxidase in Patients with Opticospinal Multiple Sclerosis: Positive Correlation with Disease Severity. J. Neuroimmunol. 2006, 178, 156–160. [Google Scholar] [CrossRef]
- Smyth, L.C.D.; Murray, H.C.; Hill, M.; van Leeuwen, E.; Highet, B.; Magon, N.J.; Osanlouy, M.; Mathiesen, S.N.; Mockett, B.; Singh-Bains, M.K.; et al. Neutrophil-Vascular Interactions Drive Myeloperoxidase Accumulation in the Brain in Alzheimer’s Disease. Acta Neuropathol. Commun. 2022, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Forghani, R.; Kim, H.J.; Wojtkiewicz, G.R.; Bure, L.; Wu, Y.; Hayase, M.; Wei, Y.; Zheng, Y.; Moskowitz, M.A.; Chen, J.W. Myeloperoxidase Propagates Damage and Is a Potential Therapeutic Target for Subacute Stroke. J. Cereb. Blood Flow Metab. 2015, 35, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Yue, Y.; Wu, D.; Zhou, C.; Guo, M.; Sun, C.; Liao, Q.; Sun, M.; Zhou, D.; Miao, C. Increased MPO in Colorectal Cancer Is Associated With High Peripheral Neutrophil Counts and a Poor Prognosis: A TCGA With Propensity Score-Matched Analysis. Front. Oncol. 2022, 12, 940706. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Sanjuán, J.; Calvo-Nieves, M.D.; Aguirre-Gervás, B.; Herreros-Rodríguez, J.; Velayos-Jiménez, B.; Castro-Alija, M.J.; Muñoz-Moreno, M.F.; Sánchez, D.; Zamora-González, N.; Bajo-Grañeras, R.; et al. Early Detection of High Oxidative Activity in Patients With Adenomatous Intestinal Polyps and Colorectal Adenocarcinoma: Myeloperoxidase and Oxidized Low-Density Lipoprotein in Serum as New Markers of Oxidative Stress in Colorectal Cancer. Lab. Med. 2015, 46, 123–135. [Google Scholar] [CrossRef]
- Saiki, T. Myeloperoxidase Concentrations in the Stool as a New Parameter of Inflammatory Bowel Disease. Kurume Med. J. 1998, 45, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, G.; Chami, B.; Liu, Y.; Schroder, A.L.; San Gabriel, P.T.; Gao, A.; Fong, G.; Wang, X.S.; Witting, P.K. The Synthetic Myeloperoxidase Inhibitor AZD3241 Ameliorates Dextran Sodium Sulfate Stimulated Experimental Colitis. Front. Pharmacol. 2020, 11, 556020. [Google Scholar] [CrossRef]
- Daugherty, A.; Dunn, J.L.; Rateri, D.L.; Heinecke, J.W. Myeloperoxidase, a Catalyst for Lipoprotein Oxidation, Is Expressed in Human Atherosclerotic Lesions. J. Clin. Investig. 1994, 94, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Teng, N.; Maghzal, G.J.; Talib, J.; Rashid, I.; Lau, A.K.; Stocker, R. The Roles of Myeloperoxidase in Coronary Artery Disease and Its Potential Implication in Plaque Rupture. Redox Rep. 2017, 22, 51–73. [Google Scholar] [CrossRef] [Green Version]
- Roth Flach, R.J.; Su, C.; Bollinger, E.; Cortes, C.; Robertson, A.W.; Opsahl, A.C.; Coskran, T.M.; Maresca, K.P.; Keliher, E.J.; Yates, P.D.; et al. Myeloperoxidase Inhibition in Mice Alters Atherosclerotic Lesion Composition. PLoS ONE 2019, 14, e0214150. [Google Scholar] [CrossRef] [Green Version]
- Andelid, K.; Bake, B.; Rak, S.; Lindén, A.; Rosengren, A.; Ekberg-Jansson, A. Myeloperoxidase as a Marker of Increasing Systemic Inflammation in Smokers without Severe Airway Symptoms. Respir. Med. 2007, 101, 888–895. [Google Scholar] [CrossRef]
- Zhu, A.; Ge, D.; Zhang, J.; Teng, Y.; Yuan, C.; Huang, M.; Adcock, I.M.; Barnes, P.J.; Yao, X. Sputum Myeloperoxidase in Chronic Obstructive Pulmonary Disease. Eur. J. Med. Res. 2014, 19, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickerhof, N.; Huang, J.; Min, E.; Michaëlsson, E.; Lindstedt, E.L.; Pearson, J.F.; Kettle, A.J.; Day, B.J. Myeloperoxidase Inhibition Decreases Morbidity and Oxidative Stress in Mice with Cystic Fibrosis-like Lung Inflammation. Free Radic. Biol. Med. 2020, 152, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Faith, M.; Sukumaran, A.; Pulimood, A.B.; Jacob, M. How Reliable an Indicator of Inflammation Is Myeloperoxidase Activity? Clin. Chim. Acta 2008, 396, 23–25. [Google Scholar] [CrossRef] [PubMed]
- Kremserova, S.; Perecko, T.; Soucek, K.; Klinke, A.; Baldus, S.; Eiserich, J.P.; Kubala, L. Lung Neutrophilia in Myeloperoxidase Deficient Mice during the Course of Acute Pulmonary Inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 5219056. [Google Scholar] [CrossRef] [Green Version]
- Endo, D.; Saito, T.; Umeki, Y.; Suzuki, K.; Aratani, Y. Myeloperoxidase Negatively Regulates the Expression of Proinflammatory Cytokines and Chemokines by Zymosan-Induced Mouse Neutrophils. Inflamm. Res. 2016, 65, 151–159. [Google Scholar] [CrossRef]
- Takeuchi, K.; Umeki, Y.; Matsumoto, N.; Yamamoto, K.; Yoshida, M.; Suzuki, K.; Aratani, Y. Severe Neutrophil-Mediated Lung Inflammation in Myeloperoxidase-Deficient Mice Exposed to Zymosan. Inflamm. Res. 2012, 61, 197–205. [Google Scholar] [CrossRef]
- Reber, L.L.; Gillis, C.M.; Starkl, P.; Jönsson, F.; Sibilano, R.; Marichal, T.; Gaudenzio, N.; Bérard, M.; Rogalla, S.; Contag, C.H.; et al. Neutrophil Myeloperoxidase Diminishes the Toxic Effects and Mortality Induced by Lipopolysaccharide. J. Exp. Med. 2017, 214, 1249–1258. [Google Scholar] [CrossRef] [Green Version]
- Antonelou, M.; Michaëlsson, E.; Evans, R.D.R.; Wang, C.J.; Henderson, S.R.; Walker, L.S.K.; Unwin, R.J.; Salama, A.D. Therapeutic Myeloperoxidase Inhibition Attenuates Neutrophil Activation, ANCA-Mediated Endothelial Damage, and Crescentic GN. J. Am. Soc. Nephrol. 2020, 31, 350–364. [Google Scholar] [CrossRef]
- McMillen, T.S.; Heinecke, J.W.; LeBoeuf, R.C. Expression of Human Myeloperoxidase by Macrophages Promotes Atherosclerosis in Mice. Circulation 2005, 111, 2798–2804. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Chuang, C.Y.; Hawkins, C.L.; Davies, M.J. Binding of Myeloperoxidase to the Extracellular Matrix of Smooth Muscle Cells and Subsequent Matrix Modification. Sci. Rep. 2020, 10, 666. [Google Scholar] [CrossRef]
- Gorudko, I.V.; Sokolov, A.; Shamova, E.; Grudinina, N.A.; Drozd, E.S.; Shishlo, L.M.; Grigorieva, D.; Bushuk, S.; Bushuk, B.A.; Chizhik, S.A.; et al. Myeloperoxidase Modulates Human Platelet Aggregation via Actin Cytoskeleton Reorganization and Store-Operated Calcium Entry. Biol. Open 2013, 2, 916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolarova, H.; Klinke, A.; Kremserova, S.; Adam, M.; Pekarova, M.; Baldus, S.; Eiserich, J.P.; Kubala, L. Myeloperoxidase Induces the Priming of Platelets. Free Radic. Biol. Med. 2013, 61, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Kantarci, O.H.; Atkinson, E.J.; Hebrink, D.D.; McMurray, C.T.; Weinshenker, B.G. Association of a Myeloperoxidase Promoter Polymorphism with Multiple Sclerosis. J. Neuroimmunol. 2000, 105, 189–194. [Google Scholar] [CrossRef]
- Coulibaly, A.P.; Pezuk, P.; Varghese, P.; Gartman, W.; Triebwasser, D.; Kulas, J.A.; Liu, L.; Syed, M.; Tvrdik, P.; Ferris, H.; et al. Neutrophil Enzyme Myeloperoxidase Modulates Neuronal Response in a Model of Subarachnoid Hemorrhage by Venous Injury. Stroke 2021, 52, 3374–3384. [Google Scholar] [CrossRef]
- Volkman, R.; Ben-Zur, T.; Kahana, A.; Garty, B.Z.; Offen, D. Myeloperoxidase Deficiency Inhibits Cognitive Decline in the 5XFAD Mouse Model of Alzheimer’s Disease. Front. Neurosci. 2019, 13, 990. [Google Scholar] [CrossRef]
- Tantry, I.Q.; Waris, S.; Habib, S.; Khan, R.H.; Mahmood, R.; Ali, A. Hypochlorous Acid Induced Structural and Conformational Modifications in Human DNA: A Multi-Spectroscopic Study. Int. J. Biol. Macromol. 2018, 106, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulos, V.; Leach, D.A.; Zinonos, I.; Ponomarev, V.; Licari, G.; Liapis, V.; Ingman, W.V.; Anderson, P.; DeNichilo, M.O.; Evdokiou, A. Inflammatory Peroxidases Promote Breast Cancer Progression in Mice via Regulation of the Tumour Microenvironment. Int. J. Oncol. 2017, 50, 1191–1200. [Google Scholar] [CrossRef] [Green Version]
- Petruska, J.M.; Mosebrook, D.R.; Jakab, G.J.; Trush, M.A. Myeloperoxidase-Enhanced Formation of (+-)-Trans-7,8-Dihydroxy-7,8-Dihydrobenzo[a]Pyrene-DNA Adducts in Lung Tissue in Vitro: A Role of Pulmonary Inflammation in the Bioactivation of a Procarcinogen. Carcinogenesis 1992, 13, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Schroder, A.L.; Chami, B.; Liu, Y.; Doyle, C.M.; el Kazzi, M.; Ahlenstiel, G.; Ahmad, G.; Pathma-Nathan, N.; Collins, G.; Toh, J.; et al. Neutrophil Extracellular Trap Density Increases With Increasing Histopathological Severity of Crohn’s Disease. Inflamm. Bowel Dis. 2022, 28, 586. [Google Scholar] [CrossRef]
- Borato, D.C.K.; Parabocz, G.C.; Ribas, J.T.; Netto, H.P.; Erdmann, F.C.; Wiecheteck, L.D.; Manente, F.A.; de Almeida Mello, L.R.; Belló, C.; André Dos Santos, F.; et al. Biomarkers in Obesity: Serum Myeloperoxidase and Traditional Cardiac Risk Parameters. Exp. Clin. Endocrinol. Diabetes 2016, 124, 49–54. [Google Scholar] [CrossRef]
- Piek, A.; Koonen, D.P.Y.; Schouten, E.M.; Lindtstedt, E.L.; Michaëlsson, E.; de Boer, R.A.; Silljé, H.H.W. Pharmacological Myeloperoxidase (MPO) Inhibition in an Obese/Hypertensive Mouse Model Attenuates Obesity and Liver Damage, but Not Cardiac Remodeling. Sci. Rep. 2019, 9, 18765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnemann, L.C.; Schaible, U.E.; Dallenga, T.K. Evaluation of Myeloperoxidase as Target for Host-Directed Therapy in Tuberculosis In Vivo. Int. J. Mol. Sci. 2022, 23, 2554. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, E.P.; Yang, Y.; Janssen, W.J.; Gandjeva, A.; Perez, M.J.; Barthel, L.; Zemans, R.L.; Bowman, J.C.; Koyanagi, D.E.; Yunt, Z.X.; et al. The Pulmonary Endothelial Glycocalyx Regulates Neutrophil Adhesion and Lung Injury during Experimental Sepsis. Nat. Med. 2012, 18, 1217–1223. [Google Scholar] [CrossRef] [Green Version]
- Manchanda, K.; Kolarova, H.; Kerkenpaß, C.; Mollenhauer, M.; Vitecek, J.; Rudolph, V.; Kubala, L.; Baldus, S.; Adam, M.; Klinke, A. MPO (Myeloperoxidase) Reduces Endothelial Glycocalyx Thickness Dependent on Its Cationic Charge. Arter. Thromb. Vasc. Biol. 2018, 38, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, D.L.; Gelderman, M.P.; Fuhrmann, S.R.; Graham, S.; Starnes, J.D.; Lefkowitz, S.S.; Bollen, A.; Moguilevsky, N. Neutrophilic Myeloperoxidase-Macrophage Interactions Perpetuate Chronic Inflammation Associated with Experimental Arthritis. Clin. Immunol. 1999, 91, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; Khalilova, I.; Tarr, J.M.; Senthilmohan, R.; Turner, R.; Haigh, R.C.; Winyard, P.G.; Kettle, A.J. Myeloperoxidase and Oxidative Stress in Rheumatoid Arthritis. Rheumatology 2012, 51, 1796–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganz, P.; Amarenco, P.; Goldstein, L.B.; Sillesen, H.; Bao, W.; Preston, G.M.; Welch, K.M.A. Association of Osteopontin, Neopterin, and Myeloperoxidase With Stroke Risk in Patients With Prior Stroke or Transient Ischemic Attacks: Results of an Analysis of 13 Biomarkers From the Stroke Prevention by Aggressive Reduction in Cholesterol Levels Trial. Stroke 2017, 48, 3223–3231. [Google Scholar] [CrossRef]
- Kim, H.J.; Wei, Y.; Wojtkiewicz, G.R.; Lee, J.Y.; Moskowitz, M.A.; Chen, J.W. Reducing Myeloperoxidase Activity Decreases Inflammation and Increases Cellular Protection in Ischemic Stroke. J. Cereb. Blood Flow Metab. 2019, 39, 1864–1877. [Google Scholar] [CrossRef]
- Ley, K.; Laudanna, C.; Cybulsky, M.I.; Nourshargh, S. Getting to the Site of Inflammation: The Leukocyte Adhesion Cascade Updated. Nat. Rev. Immunol. 2007, 7, 678–689. [Google Scholar] [CrossRef]
- Zemans, R.L.; Colgan, S.P.; Downey, G.P. Transepithelial Migration of Neutrophils: Mechanisms and Implications for Acute Lung Injury. Am. J. Respir. Cell Mol. Biol. 2009, 40, 519–535. [Google Scholar] [CrossRef]
- Parkos, C.A. Neutrophil-Epithelial Interactions: A Double-Edged Sword. Am. J. Pathol. 2016, 186, 1404–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumagin, R.; Parkos, C.A. Epithelial Adhesion Molecules and the Regulation of Intestinal Homeostasis during Neutrophil Transepithelial Migration. Tissue Barriers 2015, 3, e969100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, D.; Mollnau, H.; Eiserich, J.P.; Freeman, B.A.; Daiber, A.; Gehling, U.M.; Brümmer, J.; Rudolph, V.; Münzel, T.; Heitzer, T.; et al. Myeloperoxidase Mediates Neutrophil Activation by Association with CD11b/CD18 Integrins. Proc. Natl. Acad. Sci. USA 2005, 102, 431–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grigorieva, D.; Gorudko, I.V.; Sokolov, A.; Kostevich, V.A.; Vasilyev, V.B.; Cherenkevich, S.N.; Panasenko, O.M. Myeloperoxidase Stimulates Neutrophil Degranulation. Bull. Exp. Biol. Med. 2016, 161, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Gorudko, I.V.; Grigorieva, D.V.; Sokolov, A.V.; Shamova, E.V.; Kostevich, V.A.; Kudryavtsev, I.V.; Syromiatnikova, E.D.; Vasilyev, V.B.; Cherenkevich, S.N.; Panasenko, O.M. Neutrophil Activation in Response to Monomeric Myeloperoxidase. Biochem. Cell Biol. 2018, 96, 592–601. [Google Scholar] [CrossRef]
- Fujimoto, K.; Motowaki, T.; Tamura, N.; Aratani, Y. Myeloperoxidase Deficiency Enhances Zymosan Phagocytosis Associated with Up-Regulation of Surface Expression of CD11b in Mouse Neutrophils. Free Radic. Res. 2016, 50, 1340–1349. [Google Scholar] [CrossRef]
- Tateno, N.; Matsumoto, N.; Motowaki, T.; Suzuki, K.; Aratani, Y. Myeloperoxidase Deficiency Induces MIP-2 Production via ERK Activation in Zymosan-Stimulated Mouse Neutrophils. Free Radic. Res. 2013, 47, 376–385. [Google Scholar] [CrossRef]
- Haegens, A.; Heeringa, P.; van Suylen, R.J.; Steele, C.; Aratani, Y.; O’Donoghue, R.J.J.; Mutsaers, S.E.; Mossman, B.T.; Wouters, E.F.M.; Vernooy, J.H.J. Myeloperoxidase Deficiency Attenuates Lipopolysaccharide-Induced Acute Lung Inflammation and Subsequent Cytokine and Chemokine Production. J. Immunol. 2009, 182, 7990–7996. [Google Scholar] [CrossRef] [Green Version]
- el Kebir, D.; József, L.; Pan, W.; Filep, J.G. Myeloperoxidase Delays Neutrophil Apoptosis through CD11b/CD18 Integrins and Prolongs Inflammation. Circ. Res. 2008, 103, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Kremserová, S.; Kocurková, A.; Chorvátová, M.; Klinke, A.; Kubala, L. Myeloperoxidase Deficiency Alters the Process of the Regulated Cell Death of Polymorphonuclear Neutrophils. Front. Immunol. 2022, 13, 707085. [Google Scholar] [CrossRef]
- Flemmig, J.; Leßig, J.; Reibetanz, U.; Dautel, P.; Arnhold, J. Non-Vital Polymorphonuclear Leukocytes Express Myeloperoxidase on Their Surface. Cell. Physiol. Biochem. 2008, 21, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Fukushi, T.; Yamamoto, T.; Yoshida, M.; Fujikura, E.; Miyazaki, M.; Nakayama, M. Enhanced Neutrophil Apoptosis Accompanying Myeloperoxidase Release during Hemodialysis. Sci. Rep. 2020, 10, 21747. [Google Scholar] [CrossRef] [PubMed]
- Weinbaum, S.; Tarbell, J.M.; Damiano, E.R. The Structure and Function of the Endothelial Glycocalyx Layer. Annu. Rev. Biomed. Eng. 2007, 9, 121–167. [Google Scholar] [CrossRef]
- Uchimido, R.; Schmidt, E.P.; Shapiro, N.I. The Glycocalyx: A Novel Diagnostic and Therapeutic Target in Sepsis. Crit. Care 2019, 23, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinke, A.; Nussbaum, C.; Kubala, L.; Friedrichs, K.; Rudolph, T.K.; Rudolph, V.; Paust, H.J.; Schröder, C.; Benten, D.; Lau, D.; et al. Myeloperoxidase Attracts Neutrophils by Physical Forces. Blood 2011, 117, 1350–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, A.; Kim, K.; Li, J.; Cho, J. Myeloperoxidase Negatively Regulates Neutrophil-Endothelial Cell Interactions by Impairing AMβ2 Integrin Function in Sterile Inflammation. Front. Med. 2018, 5, 134. [Google Scholar] [CrossRef] [Green Version]
- DeNichilo, M.O.; Panagopoulos, V.; Rayner, T.E.; Borowicz, R.A.; Greenwood, J.E.; Evdokiou, A. Peroxidase Enzymes Regulate Collagen Extracellular Matrix Biosynthesis. Am. J. Pathol. 2015, 185, 1372–1384. [Google Scholar] [CrossRef]
- Brennan, M.L.; Anderson, M.M.; Shih, D.M.; Qu, X.D.; Wang, X.; Mehta, A.C.; Lim, L.L.; Shi, W.; Hazen, S.L.; Jacob, J.S.; et al. Increased Atherosclerosis in Myeloperoxidase-Deficient Mice. J. Clin. Investig. 2001, 107, 419. [Google Scholar] [CrossRef] [Green Version]
- Eiserich, J.P.; Baldus, S.; Brennan, M.L.; Ma, W.; Zhang, C.; Tousson, A.; Castro, L.; Lusis, A.J.; Nauseef, W.M.; White, C.R.; et al. Myeloperoxidase, a Leukocyte-Derived Vascular NO Oxidase. Science 2002, 296, 2391–2394. [Google Scholar] [CrossRef]
- Cheng, D.; Talib, J.; Stanley, C.P.; Rashid, I.; Michaëlsson, E.; Lindstedt, E.L.; Croft, K.D.; Kettle, A.J.; Maghzal, G.J.; Stocker, R. Inhibition of MPO (Myeloperoxidase) Attenuates Endothelial Dysfunction in Mouse Models of Vascular Inflammation and Atherosclerosis. Arter. Thromb. Vasc. Biol. 2019, 39, 1448–1457. [Google Scholar] [CrossRef]
- Kanashiro, A.; Hiroki, C.H.; da Fonseca, D.M.; Birbrair, A.; Ferreira, R.G.; Bassi, G.S.; Fonseca, M.D.; Kusuda, R.; Cebinelli, G.C.M.; da Silva, K.P.; et al. The Role of Neutrophils in Neuro-Immune Modulation. Pharmacol. Res. 2020, 151, 104580. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.M.H.; Patterson, E.K.; Clarson, C.; Cepinskas, G.; Bani-Yaghoub, M.; Stanimirovic, D.B.; Fraser, D.D. Elevated Leukocyte Azurophilic Enzymes in Human Diabetic Ketoacidosis Plasma Degrade Cerebrovascular Endothelial Junctional Proteins. Crit. Care Med. 2016, 44, e846–e853. [Google Scholar] [CrossRef] [PubMed]
- Comim, C.M.; Vilela, M.C.; Constantino, L.S.; Petronilho, F.; Vuolo, F.; Lacerda-Queiroz, N.; Rodrigues, D.H.; da Rocha, J.L.; Teixeira, A.L.; Quevedo, J.; et al. Traffic of Leukocytes and Cytokine Up-Regulation in the Central Nervous System in Sepsis. Intensive Care Med. 2011, 37, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Gellhaar, S.; Sunnemark, D.; Eriksson, H.; Olson, L.; Galter, D. Myeloperoxidase-Immunoreactive Cells Are Significantly Increased in Brain Areas Affected by Neurodegeneration in Parkinson’s and Alzheimer’s Disease. Cell Tissue Res. 2017, 369, 445. [Google Scholar] [CrossRef] [Green Version]
- van Leeuwen, E.; Hampton, M.B.; Smyth, L.C.D. Hypothiocyanous Acid Disrupts the Barrier Function of Brain Endothelial Cells. Antioxidants 2022, 11, 608. [Google Scholar] [CrossRef]
- Üllen, A.; Singewald, E.; Konya, V.; Fauler, G.; Reicher, H.; Nusshold, C.; Hammer, A.; Kratky, D.; Heinemann, A.; Holzer, P.; et al. Myeloperoxidase-Derived Oxidants Induce Blood-Brain Barrier Dysfunction in Vitro and in Vivo. PLoS ONE 2013, 8, e64034. [Google Scholar] [CrossRef]
- Goeritzer, M.; Bernhart, E.; Plastira, I.; Reicher, H.; Leopold, C.; Eichmann, T.O.; Rechberger, G.; Madreiter-Sokolowski, C.T.; Prasch, J.; Eller, P.; et al. Myeloperoxidase and Septic Conditions Disrupt Sphingolipid Homeostasis in Murine Brain Capillaries In Vivo and Immortalized Human Brain Endothelial Cells In Vitro. Int. J. Mol. Sci. 2020, 21, 1143. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Chen, H.; Du, Q.; Shen, J. Targeting Myeloperoxidase (MPO) Mediated Oxidative Stress and Inflammation for Reducing Brain Ischemia Injury: Potential Application of Natural Compounds. Front. Physiol. 2020, 11, 433. [Google Scholar] [CrossRef]
- Kim, H.J.; Wei, Y.; Lee, J.Y.; Wu, Y.; Zheng, Y.; Moskowitz, M.A.; Chen, J.W. Myeloperoxidase Inhibition Increases Neurogenesis after Ischemic Stroke. J. Pharmacol. Exp. Ther. 2016, 359, 262–272. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Liang, Y.; Zheng, S.; Zhang, H. Inhibition of Myeloperoxidase by N-Acetyl Lysyltyrosylcysteine Amide Reduces Oxidative Stress-Mediated Inflammation, Neuronal Damage, and Neural Stem Cell Injury in a Murine Model of Stroke. J. Pharmacol. Exp. Ther. 2018, 364, 311–322. [Google Scholar] [CrossRef]
- Yu, G.; Liang, Y.; Huang, Z.; Jones, D.W.; Pritchard, K.A.; Zhang, H. Inhibition of Myeloperoxidase Oxidant Production by N-Acetyl Lysyltyrosylcysteine Amide Reduces Brain Damage in a Murine Model of Stroke. J. Neuroinflamm. 2016, 13, 119. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Jing, X.; Shi, Y.; Xu, H.; Du, J.; Guan, T.; Weihrauch, D.; Jones, D.W.; Wang, W.; Gourlay, D.; et al. N-Acetyl Lysyltyrosylcysteine Amide Inhibits Myeloperoxidase, a Novel Tripeptide Inhibitor. J. Lipid Res. 2013, 54, 3016–3029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of Tumor-Associated Neutrophil Phenotype by TGF-Beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murao, S.I.; Stevens, F.J.; Ito, A.; Huberman, E. Myeloperoxidase: A Myeloid Cell Nuclear Antigen with DNA-Binding Properties. Proc. Natl. Acad. Sci. USA 1988, 85, 1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güngör, N.; Knaapen, A.M.; Munnia, A.; Peluso, M.; Haenen, G.R.; Chiu, R.K.; Godschalk, R.W.L.; van Schooten, F.J. Genotoxic Effects of Neutrophils and Hypochlorous Acid. Mutagenesis 2010, 25, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prütz, W.A. Hypochlorous Acid Interactions with Thiols, Nucleotides, DNA, and Other Biological Substrates. Arch. Biochem. Biophys. 1996, 332, 110–120. [Google Scholar] [CrossRef]
- Masuda, M.; Suzuki, T.; Friesen, M.D.; Ravanat, J.L.; Cadet, J.; Pignatelli, B.; Nishino, H.; Ohshima, H. Chlorination of Guanosine and Other Nucleosides by Hypochlorous Acid and Myeloperoxidase of Activated Human Neutrophils. Catalysis by Nicotine and Trimethylamine. J. Biol. Chem. 2001, 276, 40486–40496. [Google Scholar] [CrossRef] [Green Version]
- Whiteman, M.; Spencer, J.P.E.; Jenner, A.; Halliwell, B. Hypochlorous Acid-Induced DNA Base Modification: Potentiation by Nitrite: Biomarkers of DNA Damage by Reactive Oxygen Species. Biochem. Biophys. Res. Commun. 1999, 257, 572–576. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-Induced Damage to DNA, RNA, and Polynucleotides: Formation of Chloramines and Nitrogen-Centered Radicals. Chem. Res. Toxicol. 2002, 15, 83–92. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Pattison, D.I.; Davies, M.J. Reaction of Protein Chloramines with DNA and Nucleosides: Evidence for the Formation of Radicals, Protein-DNA Cross-Links and DNA Fragmentation. Biochem. J. 2002, 365, 605–615. [Google Scholar] [CrossRef]
- Metzler, K.D.; Fuchs, T.A.; Nauseef, W.M.; Reumaux, D.; Roesler, J.; Schulze, I.; Wahn, V.; Papayannopoulos, V.; Zychlinsky, A. Myeloperoxidase Is Required for Neutrophil Extracellular Trap Formation: Implications for Innate Immunity. Blood 2011, 117, 953–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, J.P.; Byun, J.; Takeshita, J.; Heinecke, J.W. Phagocytes Produce 5-Chlorouracil and 5-Bromouracil, Two Mutagenic Products of Myeloperoxidase, in Human Inflammatory Tissue. J. Biol. Chem. 2003, 278, 23522–23528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Mejiba, S.E.; Zhai, Z.; Gimenez, M.S.; Ashby, M.T.; Chilakapati, J.; Kitchin, K.; Mason, R.P.; Ramirez, D.C. Myeloperoxidase-Induced Genomic DNA-Centered Radicals. J. Biol. Chem. 2010, 285, 20062–20071. [Google Scholar] [CrossRef] [Green Version]
- Butin-Israeli, V.; Bui, T.M.; Wiesolek, H.L.; Mascarenhas, L.; Lee, J.J.; Mehl, L.C.; Knutson, K.R.; Adam, S.A.; Goldman, R.D.; Beyder, A.; et al. Neutrophil-Induced Genomic Instability Impedes Resolution of Inflammation and Wound Healing. J. Clin. Investig. 2019, 129, 712–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bui, T.M.; Butin-Israeli, V.; Wiesolek, H.L.; Zhou, M.; Rehring, J.F.; Wiesmüller, L.; Wu, J.D.; Yang, G.Y.; Hanauer, S.B.; Sebag, J.A.; et al. Neutrophils Alter DNA Repair Landscape to Impact Survival and Shape Distinct Therapeutic Phenotypes of Colorectal Cancer. Gastroenterology 2021, 161, 225–238.e15. [Google Scholar] [CrossRef]
- Masucci, M.T.; Minopoli, M.; del Vecchio, S.; Carriero, M.V. The Emerging Role of Neutrophil Extracellular Traps (NETs) in Tumor Progression and Metastasis. Front. Immunol. 2020, 11, 1749. [Google Scholar] [CrossRef]
- Schedel, F.; Mayer-Hain, S.; Pappelbaum, K.I.; Metze, D.; Stock, M.; Goerge, T.; Loser, K.; Sunderkötter, C.; Luger, T.A.; Weishaupt, C. Evidence and Impact of Neutrophil Extracellular Traps in Malignant Melanoma. Pigment Cell Melanoma Res. 2020, 33, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Zhang, X.; Sun, S.; Fu, Y.; Xie, L.; Ai, P. NF-ΚB and Neutrophil Extracellular Traps Cooperate to Promote Breast Cancer Progression and Metastasis. Exp. Cell Res. 2021, 405, 112707. [Google Scholar] [CrossRef]
- Vikis, H.G.; Gelman, A.E.; Franklin, A.; Stein, L.; Rymaszewski, A.; Zhu, J.; Liu, P.; Tichelaar, J.W.; Krupnick, A.S.; You, M. Neutrophils Are Required for 3-Methylcholanthrene-Initiated, Butylated Hydroxytoluene-Promoted Lung Carcinogenesis. Mol. Carcinog. 2012, 51, 993. [Google Scholar] [CrossRef] [Green Version]
- Rymaszewski, A.L.; Tate, E.; Yimbesalu, J.P.; Gelman, A.E.; Jarzembowski, J.A.; Zhang, H.; Pritchard, K.A.; Vikis, H.G. The Role of Neutrophil Myeloperoxidase in Models of Lung Tumor Development. Cancers 2014, 6, 1111–1127. [Google Scholar] [CrossRef]
- Liu, T.W.; Gammon, S.T.; Yang, P.; Fuentes, D.; Worms, D.P. Myeloid Cell-Derived HOCl Is a Paracrine Effector That Trans-Inhibits IKK/NF-ΚB in Melanoma Cells and Limits Early Tumor Progression. Sci. Signal. 2021, 14, eaax5971. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Myeloperoxidase: Mechanisms, Reactions and Inhibition as a Therapeutic Strategy in Inflammatory Diseases. Pharmacol. Ther. 2021, 218, 107685. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, P.; Narasimhulu, C.A.; Maiseyeu, A.; Durairaj, R.; Rao, S.; Rajagopalan, S.; Parthasarathy, S.; Desikan, R. Methoxyphenol Derivatives as Reversible Inhibitors of Myeloperoxidase as Potential Antiatherosclerotic Agents. Futur. Med. Chem. 2019, 12, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Valadez-Cosmes, P.; Raftopoulou, S.; Mihalic, Z.N.; Marsche, G.; Kargl, J. Myeloperoxidase: Growing Importance in Cancer Pathogenesis and Potential Drug Target. Pharmacol. Ther. 2022, 236, 108052. [Google Scholar] [CrossRef]
- Lazarevic-Pasti, T.; Leskovac, A.; Vasic, V. Myeloperoxidase Inhibitors as Potential Drugs. Curr. Drug Metab. 2015, 16, 168–190. [Google Scholar] [CrossRef]
- Kettle, A.J.; Gedye, C.A.; Winterbourn, C.C. Mechanism of Inactivation of Myeloperoxidase by 4-Aminobenzoic Acid Hydrazide. Biochem. J. 1997, 321, 503. [Google Scholar] [CrossRef] [Green Version]
- Tiden, A.K.; Sjögren, T.; Svensson, M.; Bernlind, A.; Senthilmohan, R.; Auchère, F.; Norman, H.; Markgren, P.O.; Gustavsson, S.; Schmidt, S.; et al. 2-Thioxanthines Are Mechanism-Based Inactivators of Myeloperoxidase That Block Oxidative Stress during Inflammation. J. Biol. Chem. 2011, 286, 37578–37589. [Google Scholar] [CrossRef] [Green Version]
- Ramachandra, C.J.A.; Kp, M.M.J.; Chua, J.; Hernandez-Resendiz, S.; Liehn, E.A.; Knöll, R.; Gan, L.M.; Michaëlsson, E.; Jonsson, M.K.B.; Ryden-Markinhuhta, K.; et al. Inhibiting Cardiac Myeloperoxidase Alleviates the Relaxation Defect in Hypertrophic Cardiomyocytes. Cardiovasc. Res. 2021, 118, 517–530. [Google Scholar] [CrossRef]
- Forbes, L.V.; Sjögren, T.; Auchère, F.; Jenkins, D.W.; Thong, B.; Laughton, D.; Hemsley, P.; Pairaudeau, G.; Turner, R.; Eriksson, H.; et al. Potent Reversible Inhibition of Myeloperoxidase by Aromatic Hydroxamates. J. Biol. Chem. 2013, 288, 36636–36647. [Google Scholar] [CrossRef] [Green Version]
- Soubhye, J.; van Antwerpen, P.; Dufrasne, F. A Patent Review of Myeloperoxidase Inhibitors for Treating Chronic Inflammatory Syndromes (Focus on Cardiovascular Diseases, 2013–2019). Expert Opin. Ther. Pat. 2020, 30, 595–608. [Google Scholar] [CrossRef]
- Mota, F.A.R.; Pereira, S.A.P.; Araújo, A.R.T.S.; Gullón, B.; Passos, M.L.C.; Saraiva, M.L.M.F.S. Automatic Identification of Myeloperoxidase Natural Inhibitors in Plant Extracts. Molecules 2022, 27, 1825. [Google Scholar] [CrossRef]
- Bensalem, S.; Soubhye, J.; Aldib, I.; Bournine, L.; Nguyen, A.T.; Vanhaeverbeek, M.; Rousseau, A.; Boudjeltia, K.Z.; Sarakbi, A.; Kauffmann, J.M.; et al. Inhibition of Myeloperoxidase Activity by the Alkaloids of Peganum harmala L. (Zygophyllaceae). J. Ethnopharmacol. 2014, 154, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Bremner, P.; Rivera, D.; Calzado, M.A.; Obón, C.; Inocencio, C.; Beckwith, C.; Fiebich, B.L.; Muñoz, E.; Heinrich, M. Assessing Medicinal Plants from South-Eastern Spain for Potential Anti-Inflammatory Effects Targeting Nuclear Factor-Kappa B and Other pro-Inflammatory Mediators. J. Ethnopharmacol. 2009, 124, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Pravalika, K.; Sarmah, D.; Kaur, H.; Vats, K.; Saraf, J.; Wanve, M.; Kalia, K.; Borah, A.; Yavagal, D.R.; Dave, K.R.; et al. Trigonelline Therapy Confers Neuroprotection by Reduced Glutathione Mediated Myeloperoxidase Expression in Animal Model of Ischemic Stroke. Life Sci. 2019, 216, 49–58. [Google Scholar] [CrossRef] [PubMed]
- de Jong, N.W.M.; Ramyar, K.X.; Guerra, F.E.; Nijland, R.; Fevre, C.; Voyich, J.M.; McCarthy, A.J.; Garcia, B.L.; van Kessel, K.P.M.; van Strijp, J.A.G.; et al. Immune Evasion by a Staphylococcal Inhibitor of Myeloperoxidase. Proc. Natl. Acad. Sci. USA 2017, 114, 9439–9444. [Google Scholar] [CrossRef] [Green Version]
- de Jong, N.W.M.; Ploscariu, N.T.; Ramyar, K.X.; Garcia, B.L.; Herrera, A.I.; Prakash, O.; Katz, B.B.; Leidal, K.G.; Nauseef, W.M.; van Kessel, K.P.M.; et al. A Structurally Dynamic N-Terminal Region Drives Function of the Staphylococcal Peroxidase Inhibitor (SPIN). J. Biol. Chem. 2018, 293, 2260–2271. [Google Scholar] [CrossRef] [Green Version]
- PLoScariu, N.T.; de Jong, N.W.M.; van Kessel, K.P.M.; van Strijp, J.A.G.; Geisbrecht, B.V. Identification and Structural Characterization of a Novel Myeloperoxidase Inhibitor from Staphylococcus Delphini. Arch. Biochem. Biophys. 2018, 645, 1–11. [Google Scholar] [CrossRef]
- Nyssen, P.; Franck, T.; Serteyn, D.; Mouithys-Mickalad, A.; Hoebeke, M. Propofol Metabolites and Derivatives Inhibit the Oxidant Activities of Neutrophils and Myeloperoxidase. Free Radic. Biol. Med. 2022, 191, 164–175. [Google Scholar] [CrossRef]
- Chen, M.S.; Lin, W.C.; Yeh, H.t.; Hu, C.L.; Sheu, S.M. Propofol Specifically Reduces PMA-Induced Neutrophil Extracellular Trap Formation through Inhibition of p-ERK and HOCl. Life Sci. 2019, 221, 178–186. [Google Scholar] [CrossRef]
Figure 1.
Non-canonical functions of MPO in tissue inflammation. In addition to its canonical antimicrobial activity, MPO can impact many other calls and biological processes in inflammation. Figure generated with Biorender (Publication License #BL24B6KVKU).
Figure 1.
Non-canonical functions of MPO in tissue inflammation. In addition to its canonical antimicrobial activity, MPO can impact many other calls and biological processes in inflammation. Figure generated with Biorender (Publication License #BL24B6KVKU).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lockhart, J.S.; Sumagin, R. Non-Canonical Functions of Myeloperoxidase in Immune Regulation, Tissue Inflammation and Cancer. Int. J. Mol. Sci. 2022, 23, 12250. https://doi.org/10.3390/ijms232012250
AMA Style
Lockhart JS, Sumagin R. Non-Canonical Functions of Myeloperoxidase in Immune Regulation, Tissue Inflammation and Cancer. International Journal of Molecular Sciences. 2022; 23(20):12250. https://doi.org/10.3390/ijms232012250
Chicago/Turabian StyleLockhart, Joey S., and Ronen Sumagin. 2022. "Non-Canonical Functions of Myeloperoxidase in Immune Regulation, Tissue Inflammation and Cancer" International Journal of Molecular Sciences 23, no. 20: 12250. https://doi.org/10.3390/ijms232012250
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.