
Comprehensive Identification and Functional Analysis of Stress-Associated Protein (SAP) Genes in Osmotic Stress in Maize

Abstract
:1. Introduction
2. Results
2.1. The ZmSAP Members in Maize
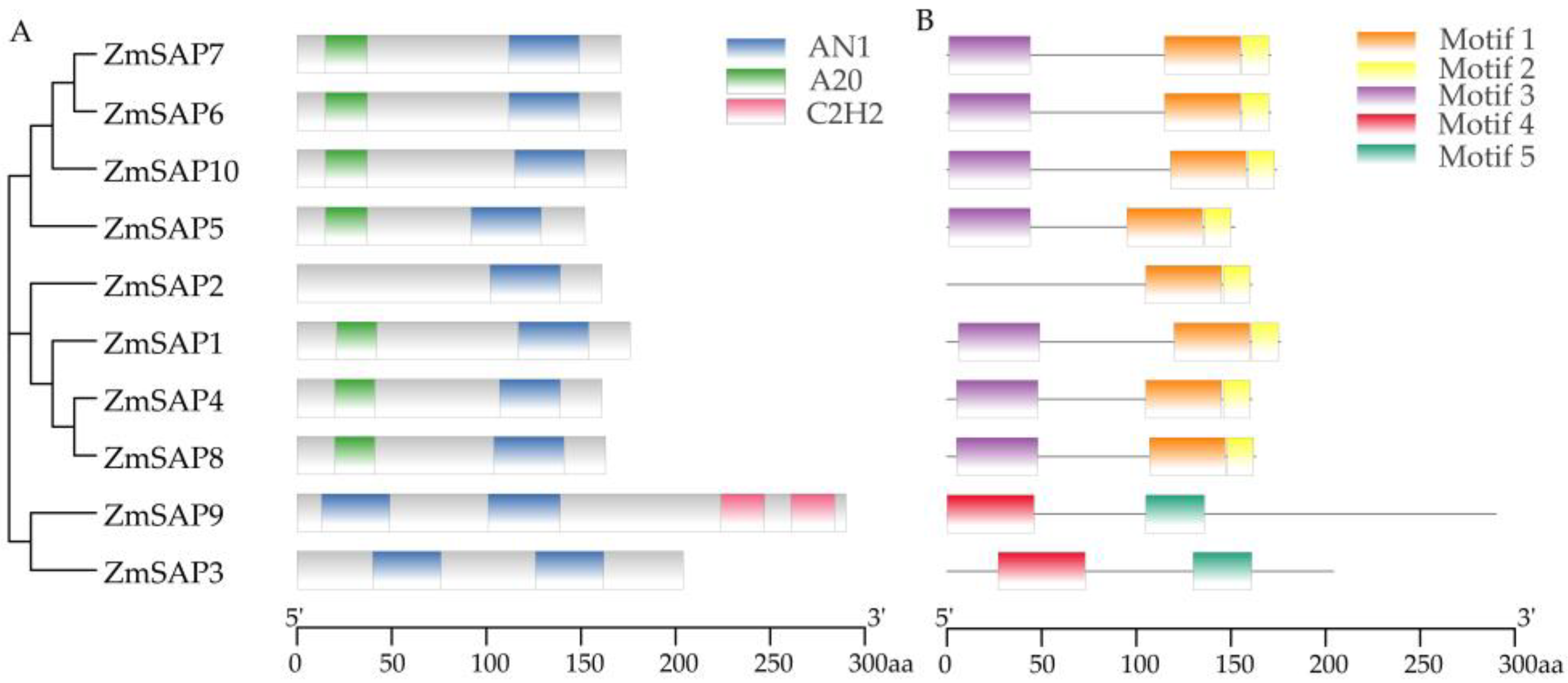
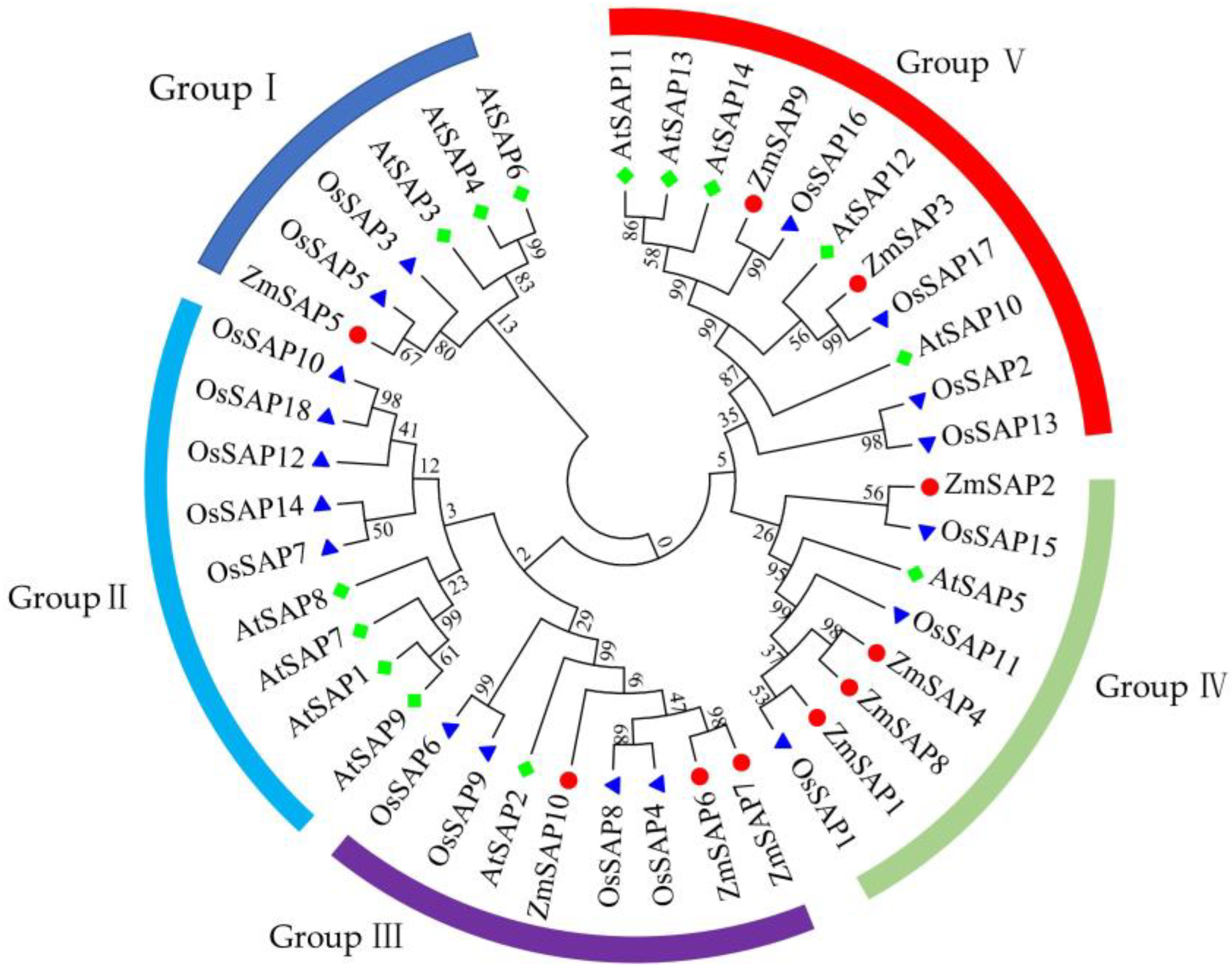
2.2. Conserved Domains and Phylogenetic Analysis of ZmSAPs
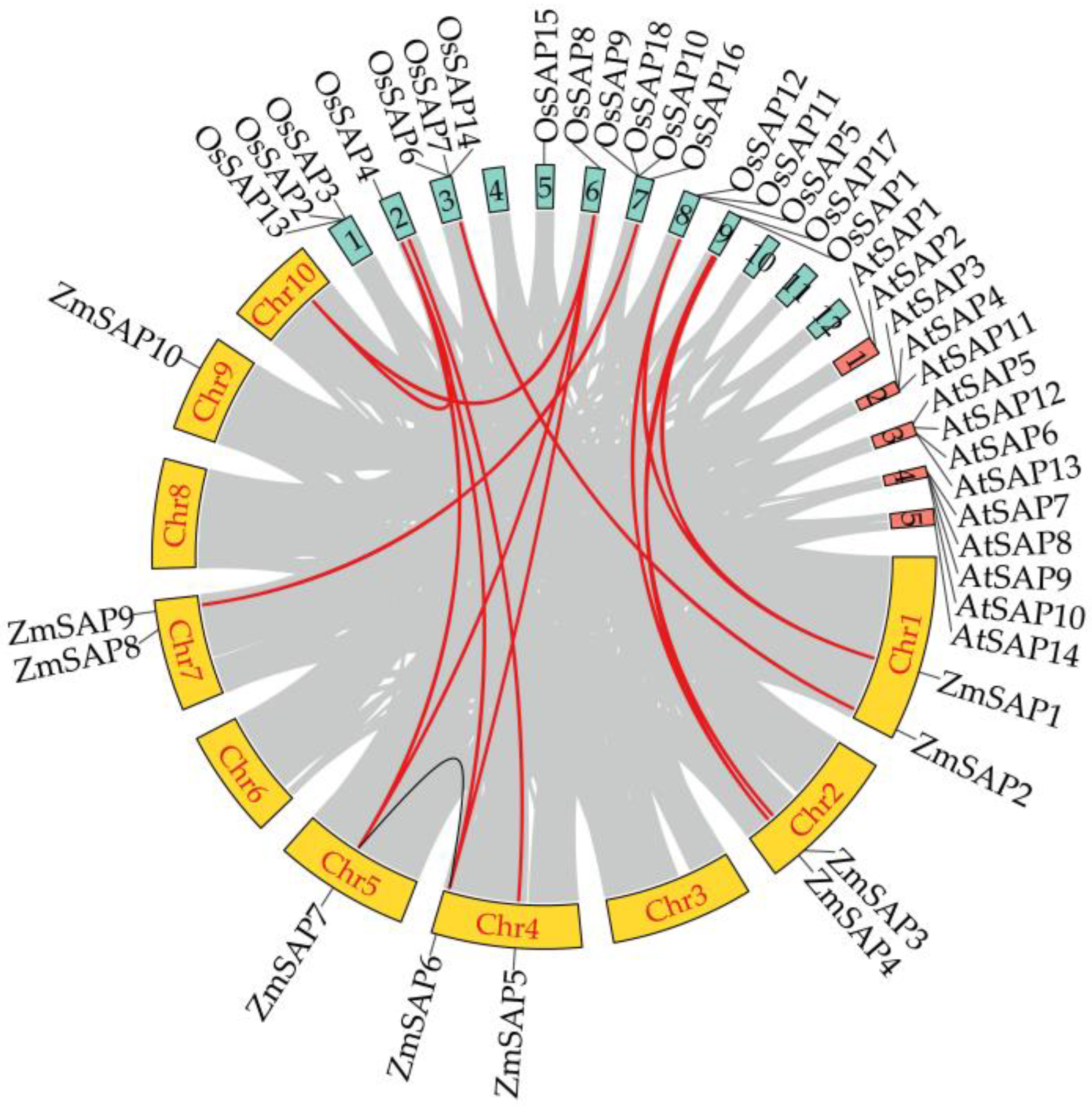
2.3. Chromosome Localization and Synteny Analysis
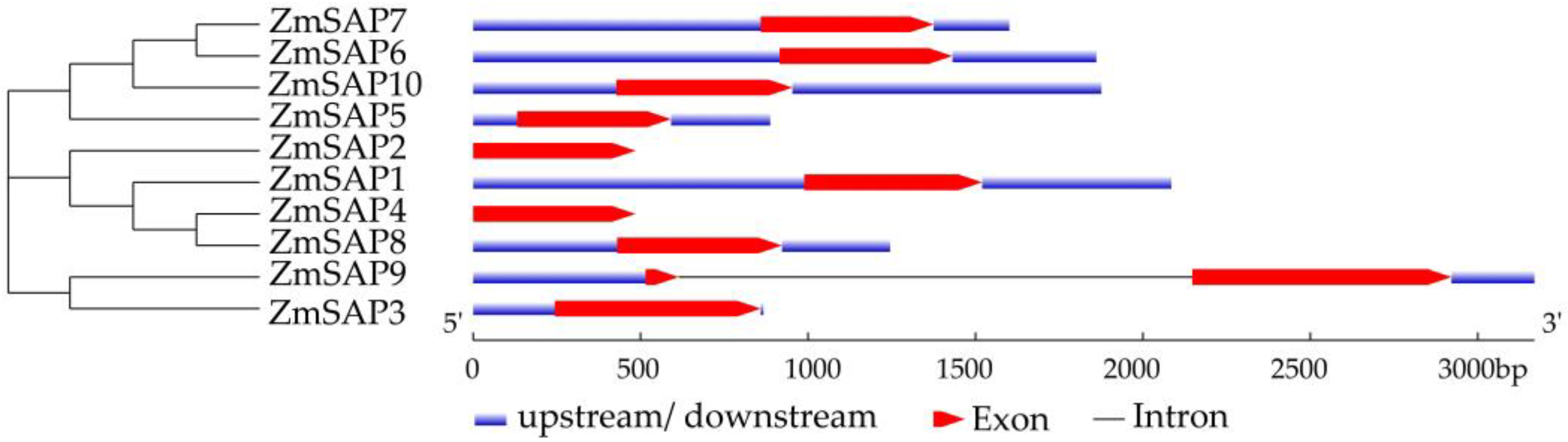
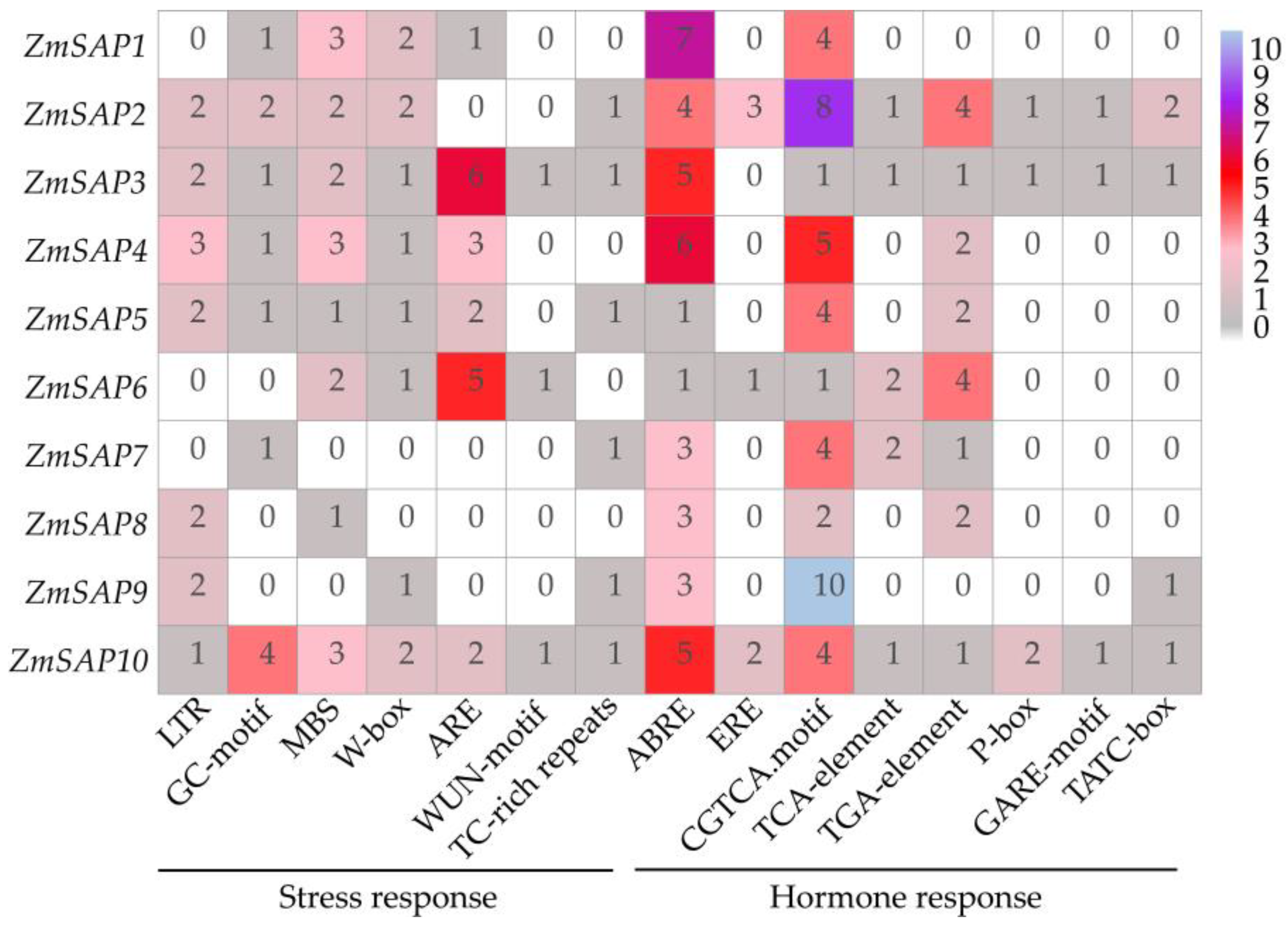
2.4. Gene Structure and Cis-Acting Elements of ZmSAPs
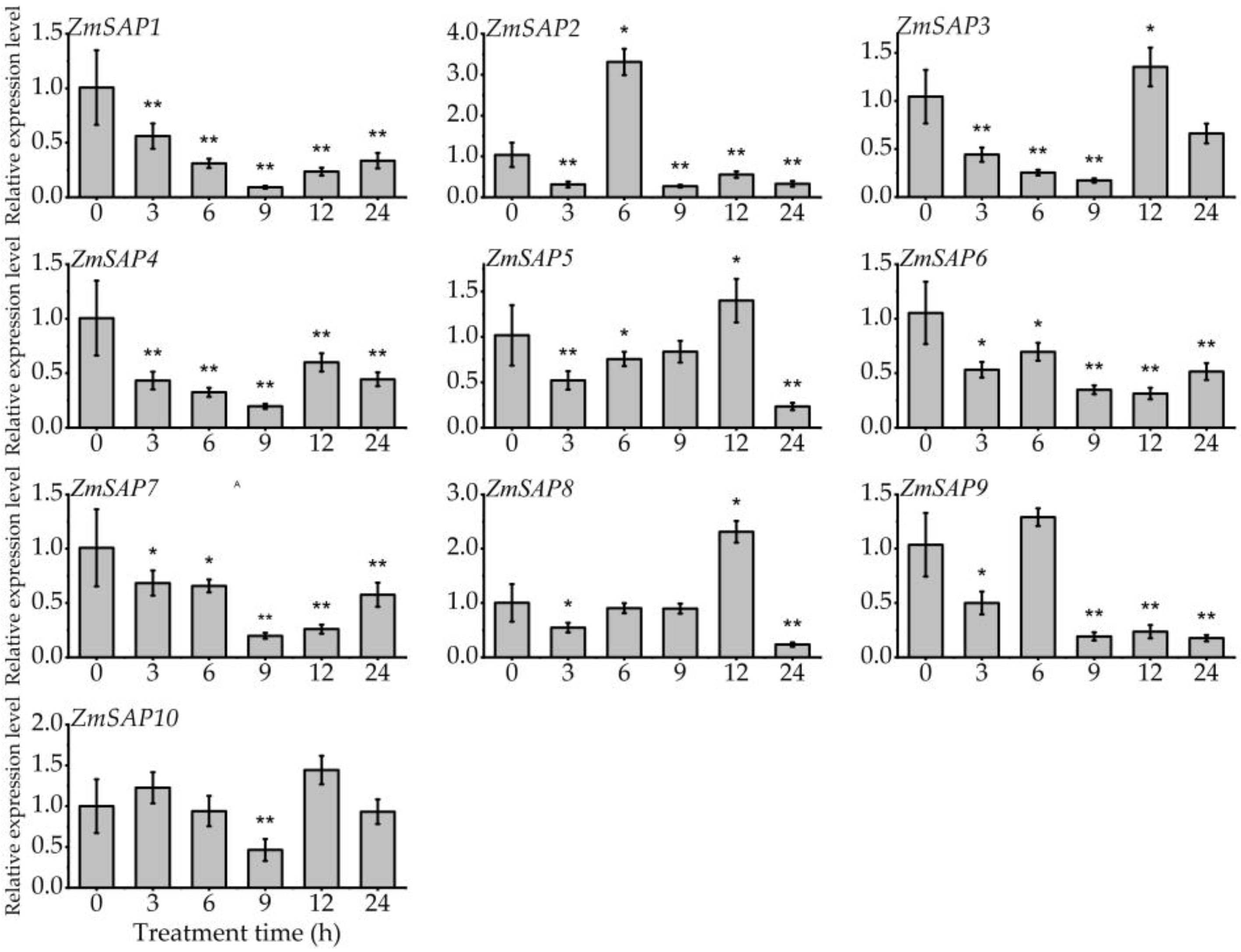
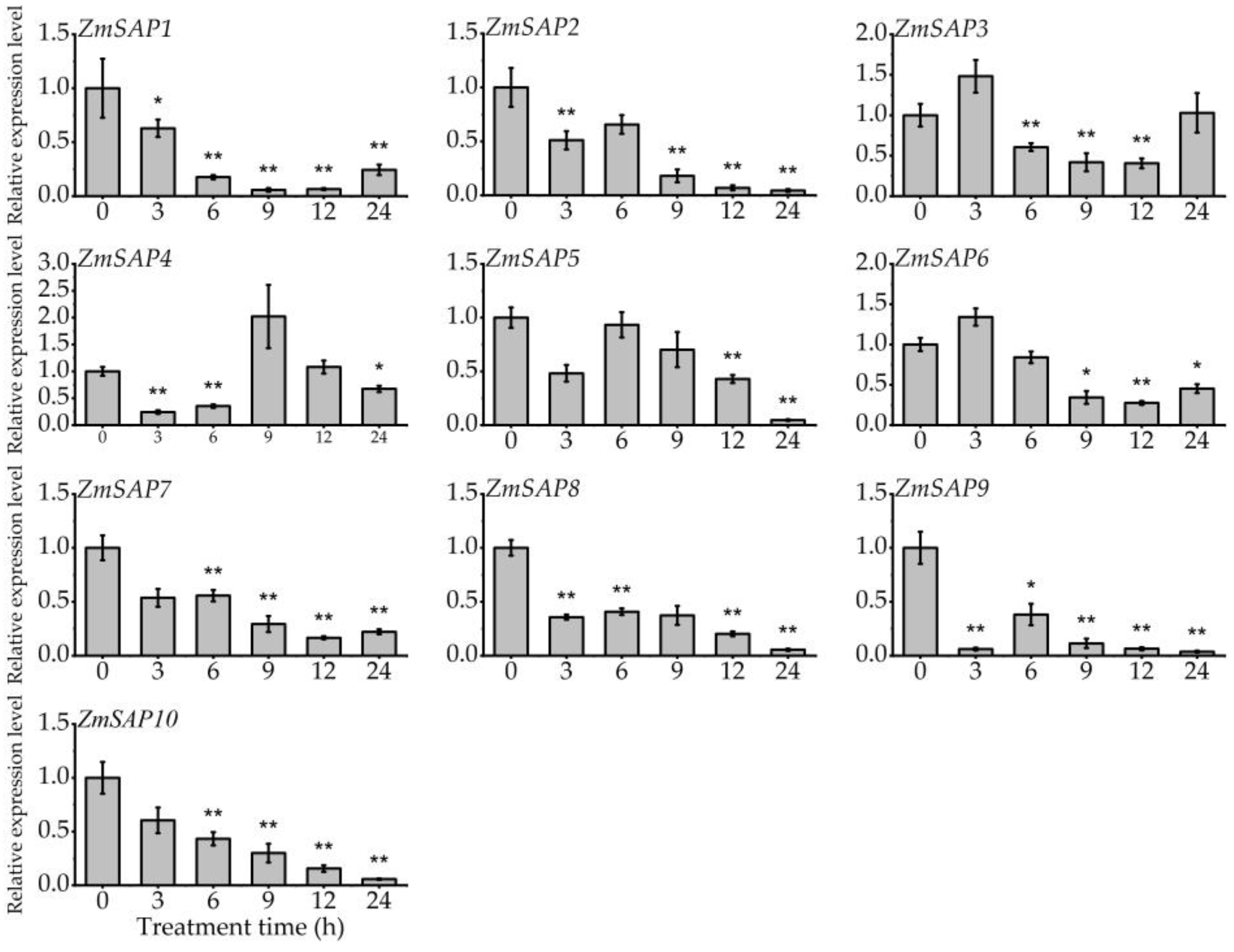
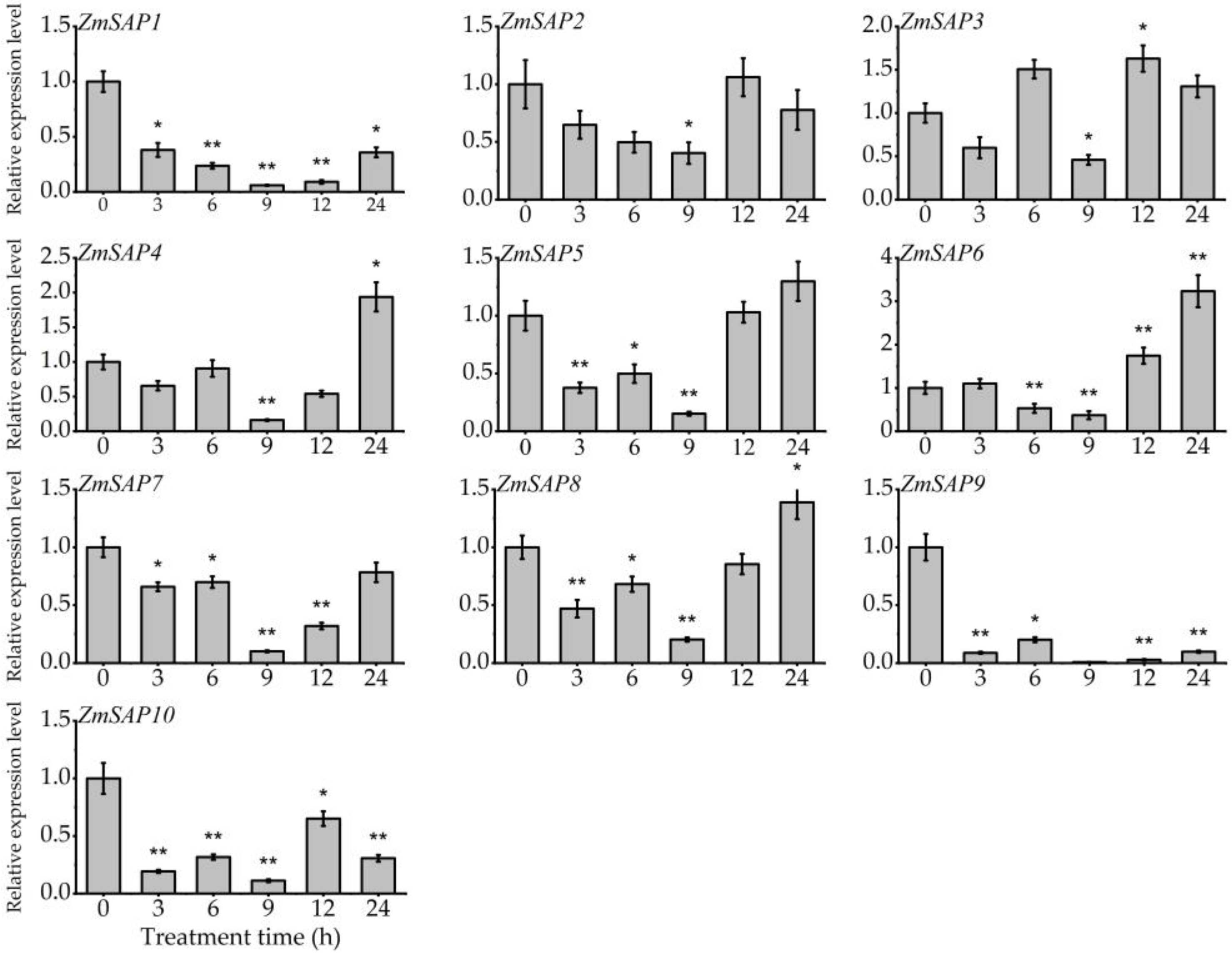
2.5. Expression Patterns of ZmSAPs
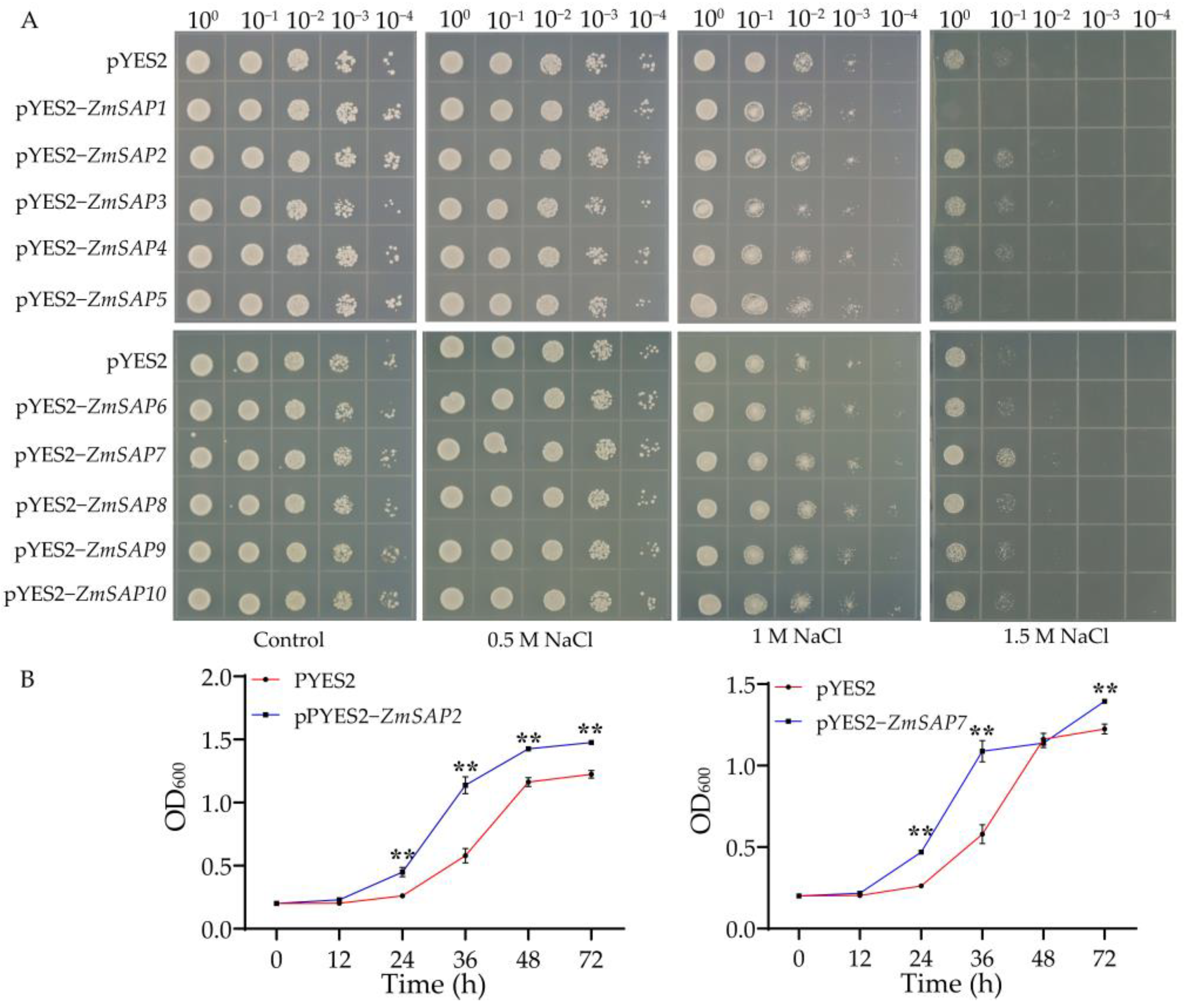
2.6. Overexpression of ZmSAP2 and ZmSAP7 Enhanced the Saline Tolerance in Yeast
3. Discussion
4. Materials and Methods
4.1. Identification of ZmSAPs in Maize
4.2. Chromosomal Location, Gene Replication and Structure analysis
4.3. Tissue Expression Analysis of ZmSAPs
4.4. Plant Materials and Stress Treatments
4.5. QRT-PCR Analysis
4.6. Stress Tolerance Test of ZmSAPs in Yeast Cells
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chávez-Arias, C.C.; Ligarreto-Moreno, G.A.; Ramírez-Godoy, A.; Restrepo-Díaz, H. Maize responses challenged by drought, elevated daytime temperature and arthropod herbivory stresses: A physiological, biochemical and molecular view. Front. Plant Sci. 2021, 12, 702841. [Google Scholar] [CrossRef] [PubMed]
- De Santis, M.A.; Soccio, M.; Laus, M.N.; Flagella, Z. Influence of drought and salt stress on durum wheat grain quality and composition: A review. Plants Basel 2021, 10, 2599. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Nykiel, M.; Gietler, M.; Fidler, J.; Prabucka, B.; Rybarczyk-Płońska, A.; Graska, J.; Boguszewska-Mańkowska, D.; Muszyńska, E.; Morkunas, I.; Labudda, M. Signal Transduction in cereal plants struggling with environmental stresses: From perception to response. Plants 2022, 11, 1009. [Google Scholar] [CrossRef]
- Lee, E.G.; Boone, D.L.; Chai, S.; Libby, S.L.; Chien, M.; Lodolce, J.P.; Ma, A. Failure to regulate TNF-induced NF-kappaB and cell death responses in A20-deficient mice. Science 2000, 289, 2350–2354. [Google Scholar] [CrossRef] [PubMed]
- Linnen, J.M.; Bailey, C.P.; Weeks, D.L. Two related localized mRNAs from Xenopus laevis encode ubiquitin-like fusion proteins. Gene 1993, 128, 181–188. [Google Scholar] [CrossRef]
- Opipari, A.W.; Boguski, M.S., Jr.; Dixit, V.M. The A20 cDNA induced by tumor necrosis factor alpha encodes a novel type of zinc finger protein. J. Biol. Chem. 1990, 265, 14705–14708. [Google Scholar] [CrossRef]
- Vij, S.; Tyagi, A.K. A20/AN1 zinc-finger domain-containing proteins in plants and animals represent common elements in stress response. Funct. Integr. Genom. 2008, 8, 301–307. [Google Scholar] [CrossRef]
- Giri, J.; Dansana, P.K.; Kothari, K.S.; Sharma, G.; Vij, S.; Tyagi, A.K. SAPs as novel regulators of abiotic stress response in plants. Bioessays 2013, 35, 639–648. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Vij, S.; Tyagi, A.K. Overexpression of a zinc-finger protein gene from rice confers tolerance to cold, dehydration, and salt stress in transgenic tobacco. Proc. Natl. Acad. Sci. USA 2004, 101, 6309–6314. [Google Scholar] [CrossRef]
- Vij, S.; Tyagi, A.K. Genome-wide analysis of the stress associated protein (SAP) gene family containing A20/AN1 zinc-finger(s) in rice and their phylogenetic relationship with Arabidopsis. Mol. Genet Genom. 2006, 276, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Giri, J.; Vij, S.; Dansana, P.K.; Tyagi, A.K. Rice A20/AN1 zinc-finger containing stress-associated proteins (SAP1/11) and a receptor-like cytoplasmic kinase (OsRLCK253) interact via A20 zinc-finger and confer abiotic stress tolerance in transgenic Arabidopsis plants. New Phytol. 2011, 191, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, M.M.; Jiang, Y.; Bao, Y.M.; Huang, X.; Sun, H.; Xu, D.Q.; Lan, H.X.; Zhang, H.S. Expression analysis of rice A20/AN1-type zinc finger genes and characterization of ZFP177 that contributes to temperature stress tolerance. Gene 2008, 420, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Fokar, M.; Abdelmageed, H.; Allen, R.D. Arabidopsis SAP5 functions as a positive regulator of stress responses and exhibits E3 ubiquitin ligase activity. Plant Mol. Biol. 2011, 75, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Ströher, E.; Wang, X.J.; Roloff, N.; Klein, P.; Husemann, A.; Dietz, K.J. Redox-dependent regulation of the stress-induced zinc-finger protein SAP12 in Arabidopsis thaliana. Mol. Plant 2009, 2, 357–367. [Google Scholar] [CrossRef]
- Kanneganti, V.; Gupta, A.K. Overexpression of OsiSAP8, a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol. Biol. 2008, 66, 445–462. [Google Scholar] [CrossRef]
- Hozain, M.; Abdelmageed, H.; Lee, J.; Kang, M.; Fokar, M.; Allen, R.D.; Holaday, A.S. Expression of AtSAP5 in cotton up-regulates putative stress-responsive genes and improves the tolerance to rapidly developing water deficit and moderate heat stress. J. Plant Physiol. 2012, 169, 1261–1270. [Google Scholar] [CrossRef]
- Dixit, A.; Tomar, P.; Vaine, E.; Abdullah, H.; Hazen, S.; Dhankher, O.P. A stress-associated protein, AtSAP13, from Arabidopsis thaliana provides tolerance to multiple abiotic stresses. Plant Cell Environ. 2018, 41, 1171–1185. [Google Scholar] [CrossRef]
- Ben Saad, R.; Fabre, D.; Mieulet, D.; Meynard, D.; Dingkuhn, M.; Al-Doss, A.; Guiderdoni, E.; Hassairi, A. Expression of the Aeluropus littoralis AlSAP gene in rice confers broad tolerance to abiotic stresses through maintenance of photosynthesis. Plant Cell Environ. 2012, 35, 626–643. [Google Scholar] [CrossRef]
- Li, J.B.; Sun, P.; Xia, Y.X.; Zheng, G.S.; Sun, J.S.; Jia, H.X. A Stress-associated protein, PtSAP13, from populus trichocarpa provides tolerance to salt stress. Int. J. Mol. Sci. 2019, 20, 5782. [Google Scholar] [CrossRef]
- Liu, J.J.; Yang, X.N.; Yang, X.Z.; Xu, M.Y.; Liu, J.; Xue, M.M.; Ma, P.D. Isolation and characterization of LcSAP, a Leymus chinensis gene which enhances the salinity tolerance of Saccharomyces cerevisiae. Mol. Biol. Rep. 2017, 44, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Sreedharan, S.; Shekhawat, U.K.; Ganapathi, T.R. MusaSAP1, a A20/AN1 zinc finger gene from banana functions as a positive regulator in different stress responses. Plant Mol. Biol. 2012, 80, 503–517. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Yin, Y.J.; Liu, X.Y.; Tong, S.M.; Xing, J.W.; Zhang, Y.; Pudake, R.N.; Izquierdo, E.M.; Peng, H.R.; Xin, M.M.; et al. The E3 Ligase TaSAP5 alters drought stress responses by promoting the degradation of drip proteins. Plant Physiol. 2017, 175, 1878–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, L.; Chang, H.H.; Chang, J.C.; Lu, H.C.; Wang, T.T.; Hsu, D.W.; Tzean, Y.; Cheng, A.P.; Chiu, Y.S.; Yeh, H.H. Plant A20/AN1 protein serves as the important hub to mediate antiviral immunity. PLoS Pathog. 2018, 14, e1007288. [Google Scholar] [CrossRef]
- Kang, M.; Lee, S.; Abdelmageed, H.; Reichert, A.; Lee, H.K.; Fokar, M.; Mysore, K.S.; Allen, R.D. Arabidopsis stress associated protein 9 mediates biotic and abiotic stress responsive ABA signaling via the proteasome pathway. Plant Cell Environ. 2017, 40, 702–716. [Google Scholar] [CrossRef]
- Tyagi, H.; Jha, S.; Sharma, M.; Giri, J.; Tyagi, A.K. Rice SAPs are responsive to multiple biotic stresses and overexpression of OsSAP1, an A20/AN1 zinc-finger protein, enhances the basal resistance against pathogen infection in tobacco. Plant Sci. 2014, 225, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Bae, E.K.; Choi, H.; Yoon, S.K.; Jang, H.A.; Choi, Y.I.; Lee, H. Knockdown of PagSAP11 confers drought resistance and promotes lateral shoot growth in hybrid poplar (Populus alba × Populus tremula var. glandulosa). Front Plant Sci. 2022, 13, 925744. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Conejero, A.; Leida, C.; Petri, C.; Gil-Muñoz, F.; Burgos, L.; Badenes, M.L.; Ríos, G. Dual regulation of water retention and cell growth by a stress-associated protein (SAP) gene in Prunus. Sci. Rep. 2017, 7, 332. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, Y.; Xiao, J.; Ma, Q.; Li, D.; Xue, Z.; Chong, K. OsDOG, a gibberellin-induced A20/AN1 zinc-finger protein, negatively regulates gibberellin-mediated cell elongation in rice. J. Plant Physiol. 2011, 168, 1098–1105. [Google Scholar] [CrossRef]
- Zhang, Y.; Lan, H.; Shao, Q.; Wang, R.; Chen, H.; Tang, H.; Zhang, H.; Huang, J. An A20/AN1-type zinc finger protein modulates gibberellins and abscisic acid contents and increases sensitivity to abiotic stress in rice (Oryza sativa). J. Exp. Bot. 2016, 67, 315–326. [Google Scholar] [CrossRef]
- Brookbank, B.P.; Patel, J.; Gazzarrini, S.; Nambara, E. Role of basal ABA in plant growth and development. Genes 2021, 12, 1936. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Jia, Z.C.; Pu, Q.; Tian, Y.; Zhu, F.Y.; Liu, Y.G. ABA mediates plant development and abiotic stress via alternative splicing. Int. J. Mol. Sci. 2022, 23, 3796. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Giri, J.; Tyagi, A.K. Rice OsiSAP7 negatively regulates ABA stress signaling and imparts sensitivity to water-deficit stress in Arabidopsis. Plant Sci. 2015, 237, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Revilla, P.; Alves, M.L.; Andelković, V.; Balconi, C.; Dinis, I.; Mendes-Moreira, P.; Redaelli, R.; de Galarreta, R.J.I.; Vaz, P.C.; Žilić, S.; et al. Traditional foods from maize (Zea mays L.) in Europe. Front. Nutr. 2022, 8, 683399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Z.; Zheng, W.J.; Cao, X.Y.; Cui, X.Y.; Zhao, S.P.; Yu, T.F.; Chen, J.; Zhou, Y.B.; Chen, M.; Chai, S.C.; et al. Genomic analysis of stress associated proteins in soybean and the role of GmSAP16 in abiotic stress responses in Arabidopsis and soybean. Front. Plant Sci. 2019, 10, 1453. [Google Scholar] [CrossRef] [Green Version]
- Solanke, A.U.; Sharma, M.K.; Tyagi, A.K.; Sharma, A.K. Characterization and phylogenetic analysis of environmental stress-responsive SAP gene family encoding A20/AN1 zinc finger proteins in tomato. Mol. Genet Genom. 2009, 282, 153–164. [Google Scholar] [CrossRef]
- Gao, W.; Long, L.; Tian, X.Q.; Jin, J.J.; Liu, H.L.; Zhang, H.; Xu, F.C. Genome-wide identification and expression analysis of stress-associated proteins (SAPs) containing A20/AN1 zinc finger in cotton. Mol. Genet. Genom. 2016, 291, 2199–2213. [Google Scholar] [CrossRef]
- Dong, Q.L.; Duan, D.Y.; Zhao, S.; Xu, B.Y.; Luo, J.W.; Wang, Q.; Huang, D.; Liu, C.H.; Li, C.; Gong, X.Q.; et al. Genome-wide analysis and cloning of the apple stress-associated protein gene family reveals MdSAP15, which confers tolerance to drought and osmotic stresses in transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2478. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Xie, S.; Xie, P.; Yao, M.; Liu, W.; Qin, L.W.; Liu, Z.S.; Zheng, M.; Liu, H.F.; Guan, M. Genome-wide identification of stress-associated proteins (SAP) with A20/AN1 zinc finger domains associated with abiotic stresses responses in Brassica napus. Environ. Exp. Bot. 2019, 165, 108–119. [Google Scholar] [CrossRef]
- Lai, W.; Zhou, Y.; Pan, R.; Liao, L.T.; He, J.C.; Liu, H.C.; Yang, Y.G.; Liu, S.Q. Identification and expression analysis of stress-associated proteins (SAPs) containing A20/AN1 zinc finger in cucumber. Plants Basel 2020, 9, 400. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Kuang, J.G.; Han, B.; Chen, S.Y.; Liu, A.Z. Genomic characterization and expression profiles of stress-associated proteins (SAPs) in castor bean (Ricinus communis). Plant Divers. 2021, 43, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Wang, M.; Fu, J.J.; Xuan, N.; Zhu, Y.; Lian, Y.; Jia, Z.W.; Zheng, J.; Wang, G.Y. Phylogenetic and expression analysis of ZnF-AN1 genes in plants. Genomics 2007, 90, 265–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Lyum, H.M.; Zhu, K.K.; Van De, P.Y.; Cheng, Z.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2021, 105, 1072–1082. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Q.; Muhammad, H.B.K.; Lu, F.Z.; Sun, F.A.; Qu, J.T.; Liu, B.L.; Li, W.C.; Fu, F.L. Isolation and identification of a vegetative organspecific promoter from maize. Physiol. Mol. Biol. Plants. 2019, 25, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Narusaka, Y.; Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Furihata, T.; Abe, H.; Narusaka, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Interaction between two cis-acting elements, ABRE and DRE, in ABA-dependent expression of Arabidopsis rd29A gene in response to dehydration and high-salinity stresses. Plant J. 2003, 34, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Kavi Kishor, P.B.; Tiozon, R.N., Jr.; Fernie, A.R.; Sreenivasulu, N. Abscisic acid and its role in the modulation of plant growth, development, and yield stability. Trends Plant Sci. 2022, 10, S1360-1385(22)00219-9. [Google Scholar] [CrossRef]
- Yu, H.Q.; Feng, W.Q.; Sun, F.A.; Zhang, Y.Y.; Qu, J.T.; Liu, B.L.; Lu, F.Z.; Yang, L.; Fu, F.L.; Li, W.C. Cloning and characterization of BES1/BZR1 transcription factor genes in maize. Plant Growth Regul. 2018, 86, 235–249. [Google Scholar] [CrossRef]
- Bae, Y.; Lim, C.W.; Lee, S.C. Differential functions of pepper stress-associated proteins in response to abiotic stresses. Front. Plant Sci. 2021, 12, 756068. [Google Scholar] [CrossRef]
- Sun, F.A.; Yu, H.Q.; Qu, J.T.; Cao, Y.; Ding, L.; Feng, W.Q.; Bin Khalid, M.H.; Li, W.C.; Fu, F.L. Maize ZmBES1/BZR1-5 decreases ABA sensitivity and confers tolerance to osmotic stress in transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Saad, R.; Zouari, N.; Ben Ramdhan, W.; Azaza, J.; Meynard, D.; Guiderdoni, E.; Hassairi, A. Improved drought and salt stress tolerance in transgenic tobacco overexpressing a novel A20/AN1 zinc-finger AlSAP gene isolated from the halophyte grass Aeluropus littoralis. Plant Mol. Biol. 2010, 72, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Ben Saad, R.; Safi, H.; Hsouna, A.B.; Brini, F.; Romdhane, W.B. Functional domain analysis of LmSAP protein reveals the crucial role of the zinc-finger A20 domain in abiotic stress tolerance. Protoplasma 2019, 256, 1333–1344. [Google Scholar] [CrossRef]
- Dansana, P.K.; Kothari, K.S.; Vij, S.; Tyagi, A.K. OsiSAP1 overexpression improves water-deficit stress tolerance in transgenic rice by affecting expression of endogenous stress-related genes. Plant Cell Rep. 2014, 33, 1425–1440. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.K.; Bae, E.K.; Lee, H.; Choi, Y.I.; Han, M.; Choi, H.; Kang, K.S.; Park, E.J. Downregulation of stress-associated protein 1 (PagSAP1) increases salt stress tolerance in poplar (Populus alba × P. glandulosa). Trees 2018, 32, 823–833. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202-8. [Google Scholar] [CrossRef]
- Wang, Y.P.; Tang, H.B.; Debarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 −ΔΔ C T method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Soni, R.; Carmichael, J.P.; Murray, J.A. Parameters affecting lithium acetate-mediated transformation of Saccharomyces cerevisiae and development of a rapid and simplified procedure. Curr. Genet. 1993, 24, 455–459. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Number of Amino Acids | Molecular Weight (KDa) | pI | CDS (bp) | GC (%) | Grand Average Hydropathy | Subcellular Locations | Instability Index |
|---|---|---|---|---|---|---|---|---|---|
| Zm00001eb034760 | ZmSAP1 | 176 | 18.75 | 9.12 | 531 | 73.07 | −0.572 | N | 68.93 |
| Zm00001eb060270 | ZmSAP2 | 161 | 16.68 | 9.53 | 486 | 65.02 | −0.515 | N | 54.33 |
| Zm00001eb099020 | ZmSAP3 | 204 | 22.15 | 9.39 | 615 | 71.22 | −0.713 | N | 48.66 |
| Zm00001eb101840 | ZmSAP4 | 161 | 16.78 | 9.19 | 486 | 73.25 | −0.276 | N | 50.93 |
| Zm00001eb181400 | ZmSAP5 | 152 | 16.00 | 9.01 | 459 | 70.81 | −0.387 | N | 51.56 |
| Zm00001eb205990 | ZmSAP6 | 171 | 18.31 | 8.28 | 516 | 54.84 | −0.323 | N | 31.15 |
| Zm00001eb236360 | ZmSAP7 | 171 | 18.29 | 8.28 | 516 | 56.59 | −0.235 | N | 30.92 |
| Zm00001eb316600 | ZmSAP8 | 163 | 17.20 | 9.45 | 492 | 73.37 | −0.458 | N | 62.8 |
| Zm00001eb324750 | ZmSAP9 | 290 | 32.04 | 8.58 | 873 | 49.37 | −0.59 | N | 38.37 |
| Zm00001eb388350 | ZmSAP10 | 174 | 18.41 | 8.48 | 525 | 59.62 | −0.198 | N | 26.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Q.; Duan, H.; Cao, Y.; Li, Y.; Lin, X.; Pang, H.; Yang, Q.; Li, W.; Fu, F.; Zhang, Y.; et al. Comprehensive Identification and Functional Analysis of Stress-Associated Protein (SAP) Genes in Osmotic Stress in Maize. Int. J. Mol. Sci. 2022, 23, 14010. https://doi.org/10.3390/ijms232214010
Fu Q, Duan H, Cao Y, Li Y, Lin X, Pang H, Yang Q, Li W, Fu F, Zhang Y, et al. Comprehensive Identification and Functional Analysis of Stress-Associated Protein (SAP) Genes in Osmotic Stress in Maize. International Journal of Molecular Sciences. 2022; 23(22):14010. https://doi.org/10.3390/ijms232214010
Chicago/Turabian StyleFu, Qiankun, Huaming Duan, Yang Cao, Yan Li, XiaoLong Lin, Haowan Pang, Qingqing Yang, Wanchen Li, Fengling Fu, Yuanyuan Zhang, and et al. 2022. "Comprehensive Identification and Functional Analysis of Stress-Associated Protein (SAP) Genes in Osmotic Stress in Maize" International Journal of Molecular Sciences 23, no. 22: 14010. https://doi.org/10.3390/ijms232214010