
A Case Study of the Morphological and Molecular Variation within a Ciliate Genus: Taxonomic Descriptions of Three Dysteria Species (Ciliophora, Cyrtophoria), with the Establishment of a New Species

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Dysteria paracrassipes n. sp.
2.1.1. ZooBank Registration
2.1.2. Diagnosis
2.1.3. Type Locality
2.1.4. Etymology
2.1.5. Type Materials
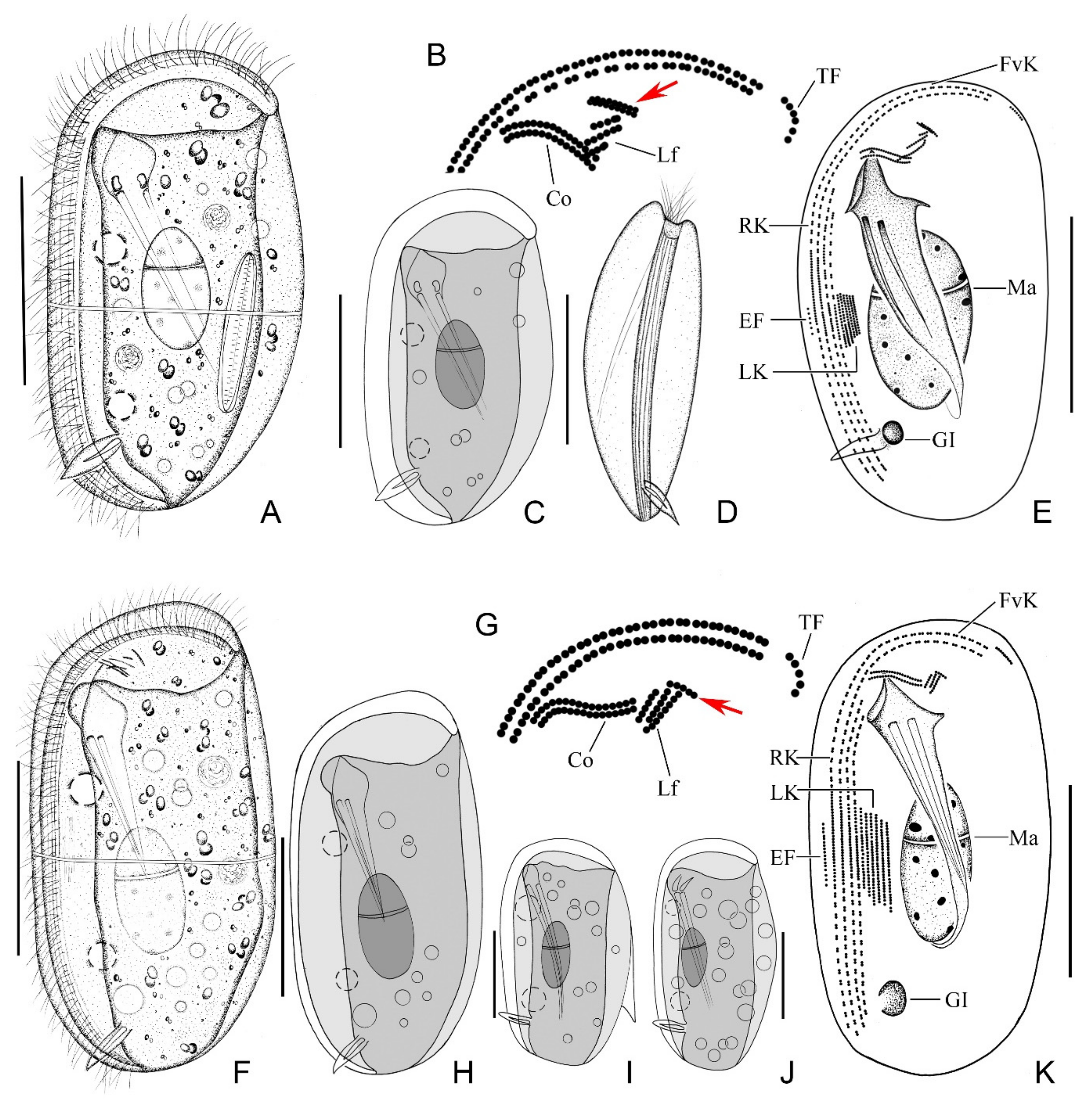
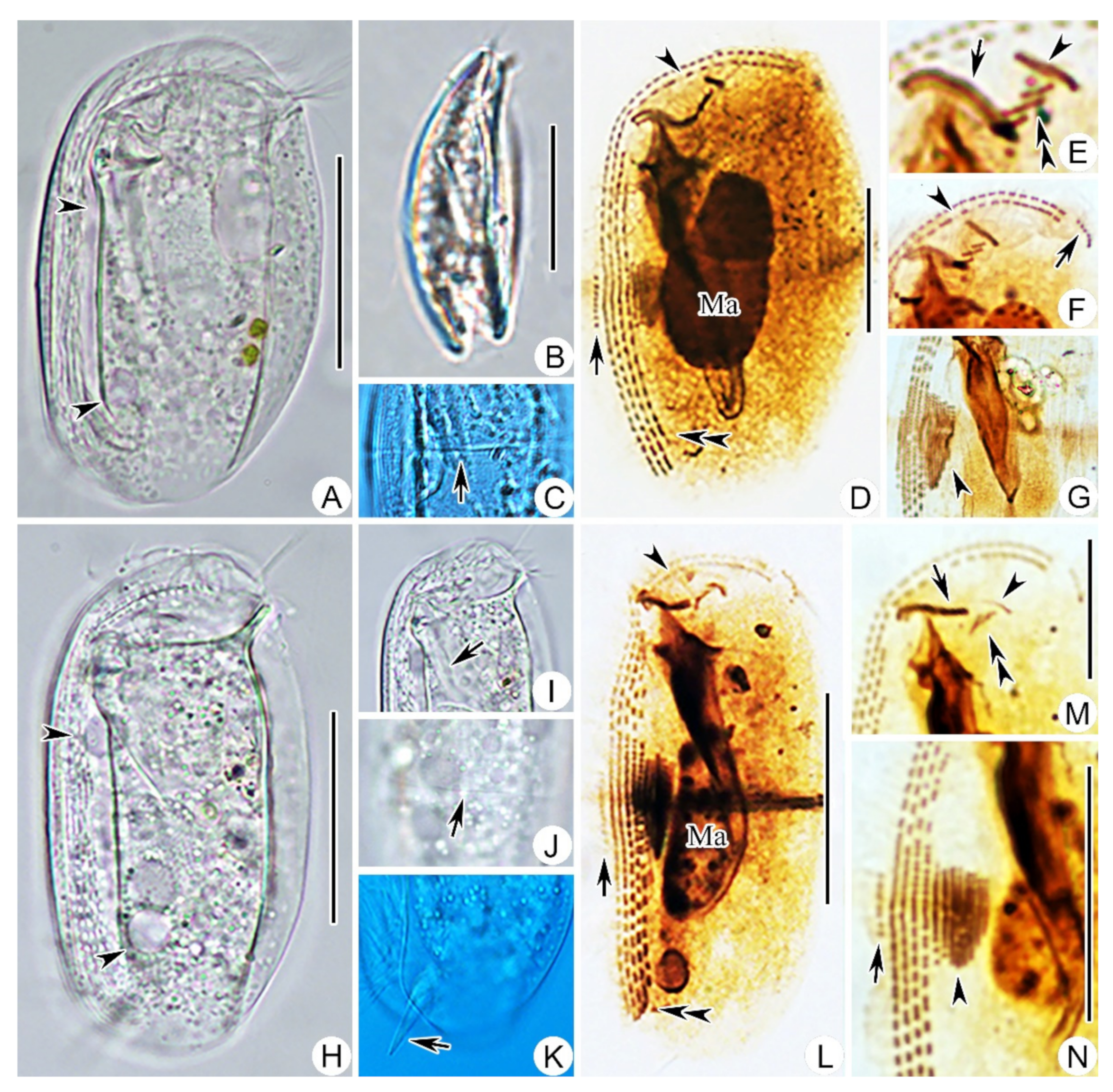
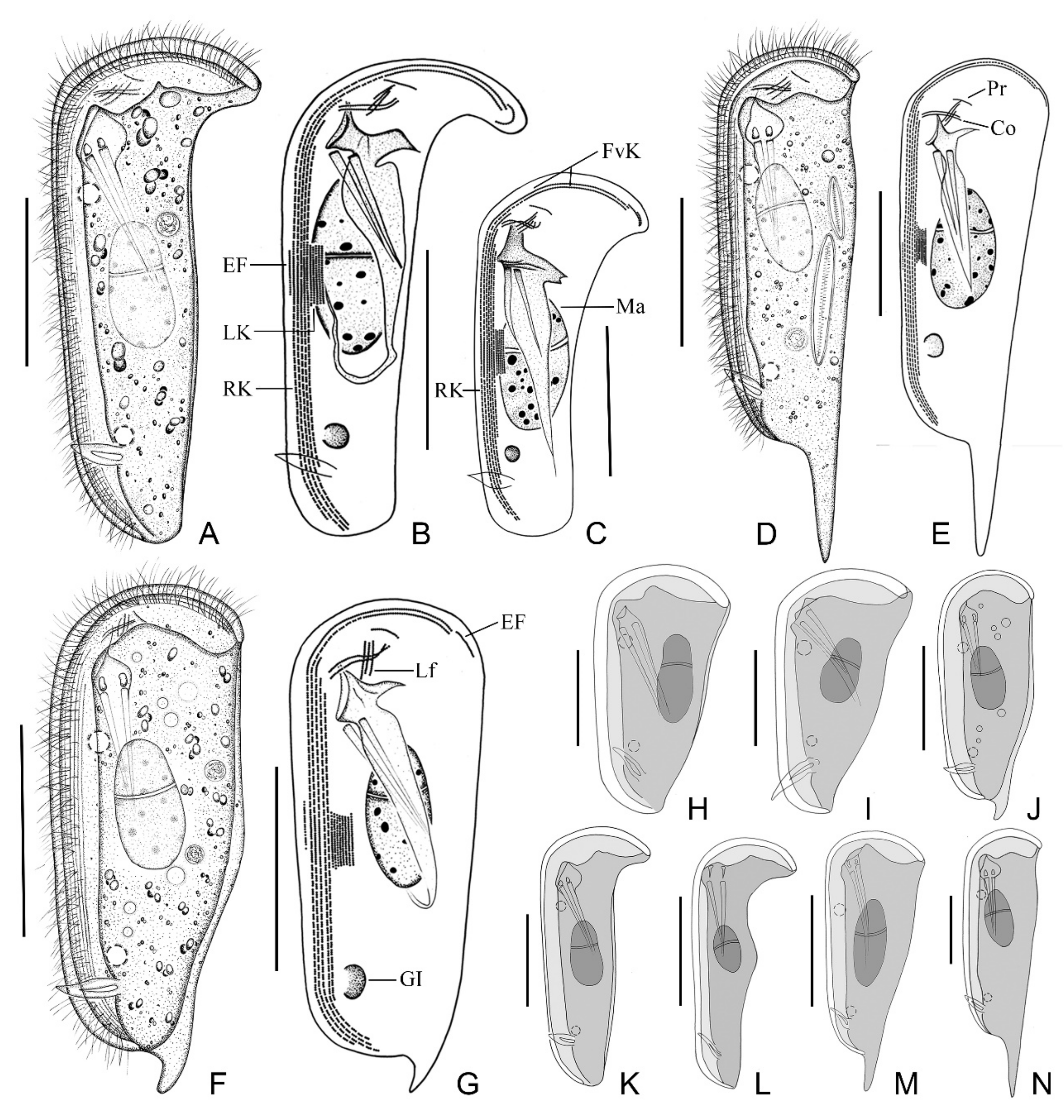
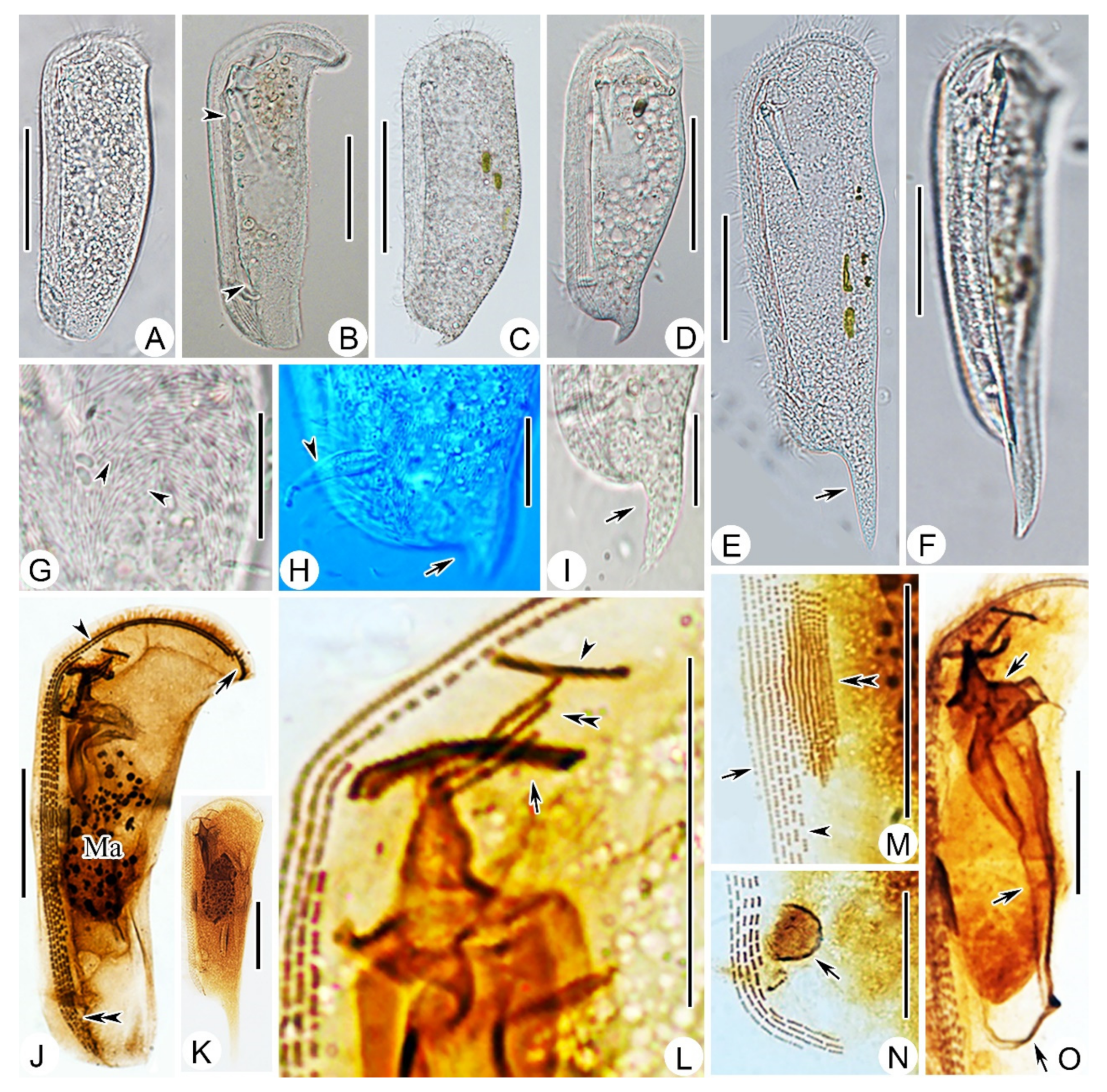
2.1.6. Morphological Description
2.2. Dysteria Crassipes Claparède & Lachmann, 1859
2.2.1. Voucher Slides
2.2.2. Morphological Description of Ningbo Population
2.3. Dysteria Brasiliensis Faria et al., 1922
2.3.1. Voucher Slides
2.3.2. Morphological Description of Ningbo Population-I
2.3.3. Morphological Description of Ningbo Population-II
2.3.4. Morphological Description of Ningbo Population-III
2.3.5. Morphological Description of Ningbo Population-IV
2.3.6. Morphological Description of Ningbo Population-V
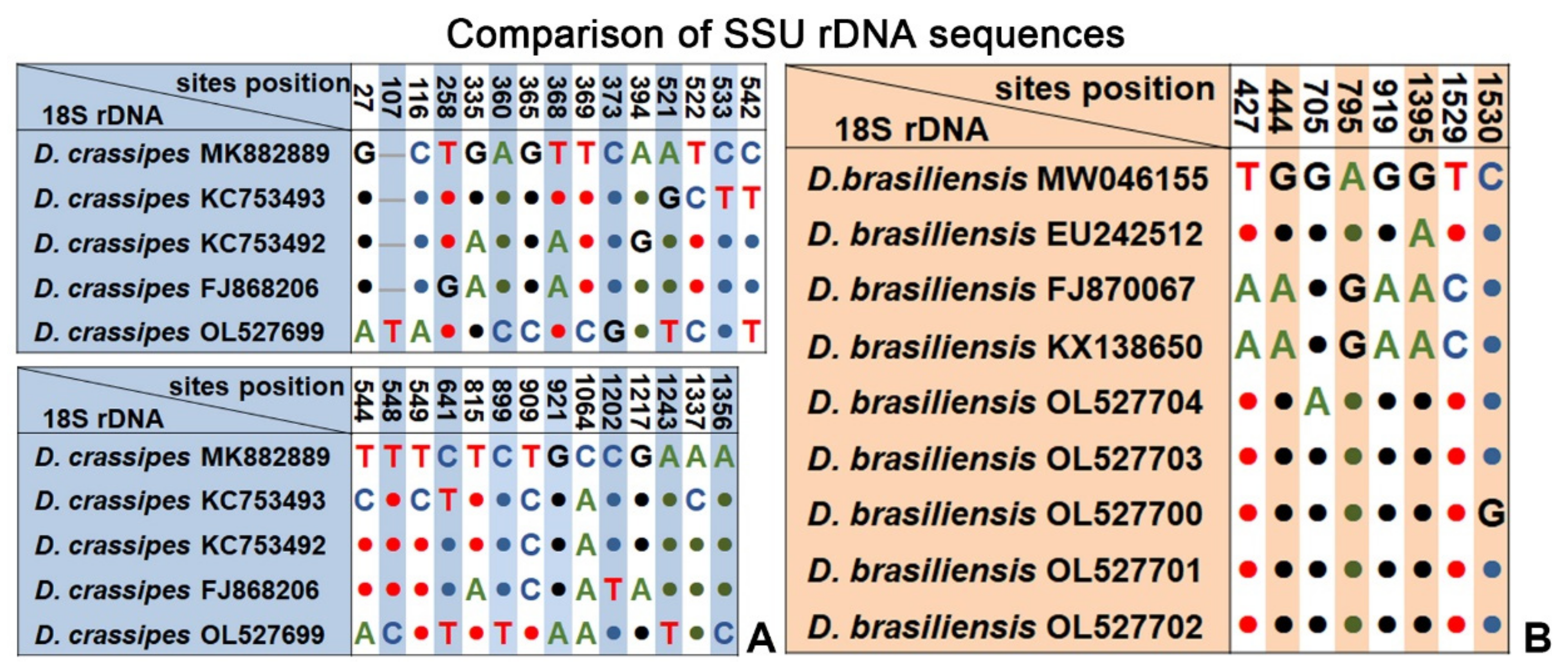
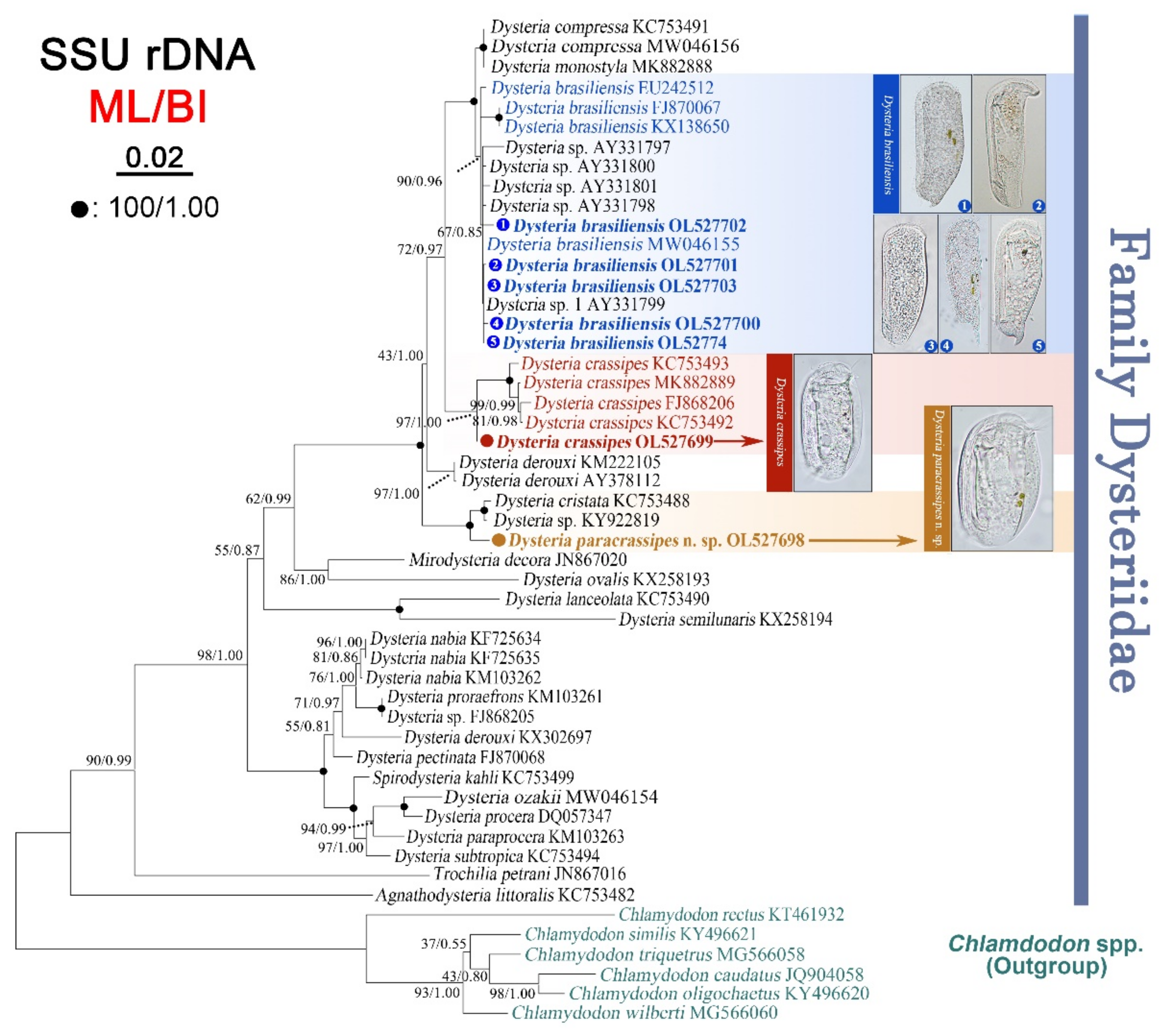
2.4. SSU rRNA Gene Sequences and Phylogenetic Analyses
3. Discussion
3.1. Comments on Dysteria paracrassipes n. sp.
3.2. Comments on Dysteria crassipes Claparède & Lachmann, 1859
3.3. Comments on Dysteria Brasiliensis Faria et al., 1922
3.4. Phylogenetic Analyses Based on SSU rDNA Sequences
4. Materials and Methods
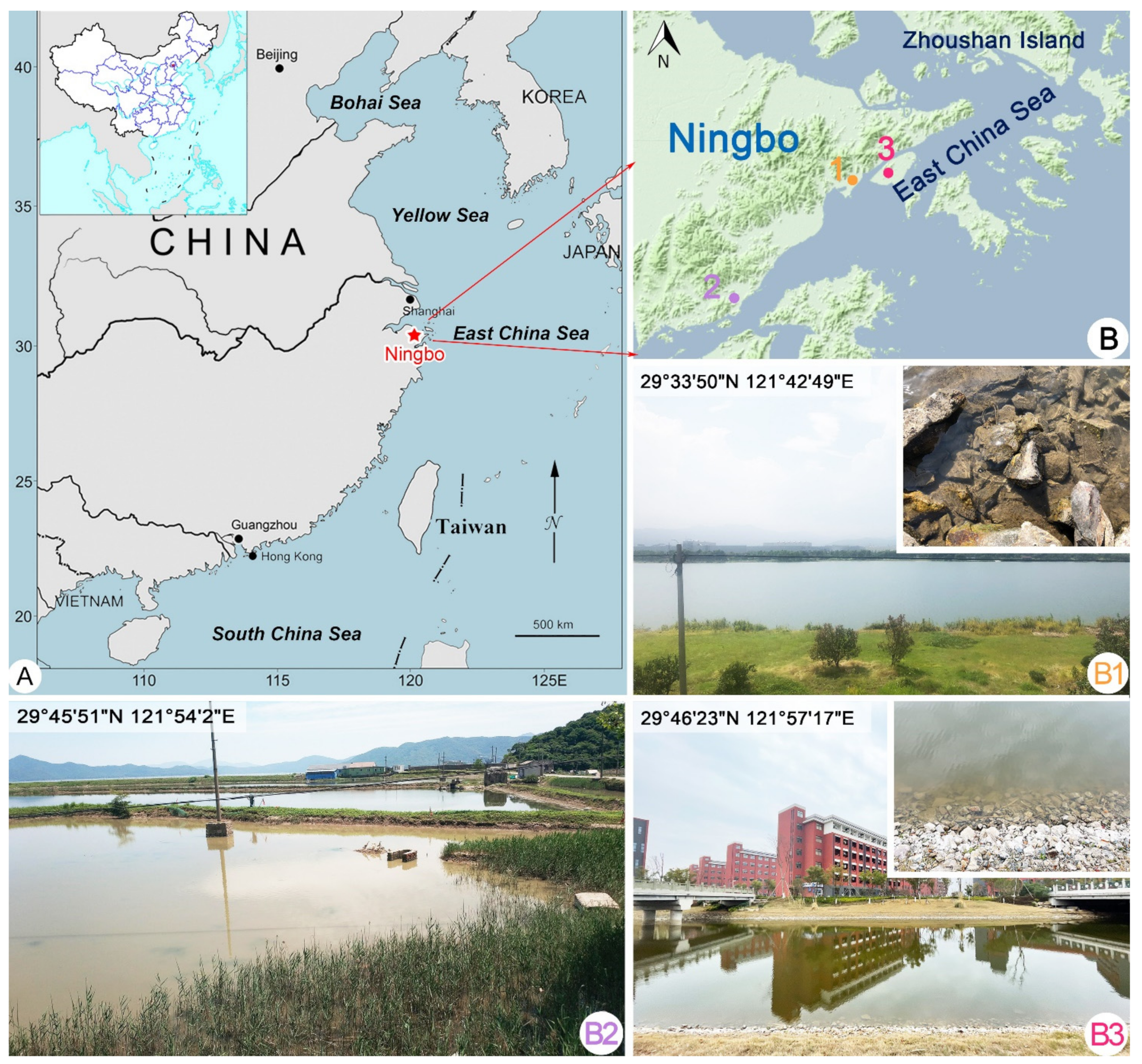
4.1. Sample Collection, Observation, and Identification
4.2. DNA Extraction, PCR Amplification, and Gene Sequencing
4.3. Phylogenetic Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Hu, X.; Gong, J.; AL-Rasheid, K.A.S.; AL-Farraj, S.A. Morphology and infraciliature of two new marine ciliates, Paracyrtophoron tropicum nov. gen., nov. spec. and Aegyria rostellum nov. spec. (Ciliophora, Cyrtophorida), isolated from tropical waters in southern China. Eur. J. Protistol. 2012, 48, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Deroux, G. Origine des cinéties antérieures, gauches et buccales dans le genre Dysteria Huxley. Cr. Acad. Bulg. Sci. 1965, 260, 6689–6691. [Google Scholar]
- Song, W.; Warren, A.; Hu, X. Free-Living Ciliates in the Bohai and Yellow Seas, China; Science Press: Beijing, China, 2009. [Google Scholar]
- Song, W.; Wilbert, N. Faunistic studies on marine ciliates from the Antarctic benthic area, including descriptions of one epizoic form, 6 new species and, 2 new genera (Protozoa: Ciliophora). Acta Protozool. 2002, 41, 23–62. [Google Scholar]
- Wang, C.; Qu, Z.; Hu, X. Morphology and SSU rDNA sequences of four cyrtophorian ciliates from China, with description of a new species (Protista, Ciliophora, Phyllopharyngea). Zootaxa 2019, 4664, 206–220. [Google Scholar] [CrossRef]
- Lynn, D.H. The Ciliated Protozoa: Characterization, Classification, and Guide to the Literature, 3rd ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1–605. [Google Scholar]
- Warren, A.; Patterson, D.J.; Dunthorn, M.; Clamp, J.C.; Achilles-Day, U.E.M.; Aescht, E.; Carr, M.; Day, J.G. Beyond the “Code”: A guide to the description and documentation of biodiversity in ciliated protists (Alveolata, Ciliophora). J. Eukaryot. Microbiol. 2017, 64, 539–554. [Google Scholar] [CrossRef] [Green Version]
- Dunthorn, M.; Stoeck, T.; Clamp, J.; Warren, A.; Mahe, F. Ciliates and the rare biosphere: A review. J. Eukaryot. Microbiol. 2014, 61, 404–409. [Google Scholar] [CrossRef]
- Gong, J.; Song, W.; Warren, A.; Lin, X.; Roberts, D.M. Microscopical observations on four marine Dysteria species (Ciliophora, Cyrtophorida). Eur. J. Protistol. 2007, 43, 147–161. [Google Scholar] [CrossRef]
- Qu, Z.; Wang, C.; Gao, F.; Li, J.; Al-Rasheid, K.A.S.; Hu, X. Taxonomic studies on seven species of Dysteria (Ciliophora, Cyrtophoria), including a description of Dysteria paraprocera sp. n. Eur. J. Protistol. 2015, 51, 241–258. [Google Scholar] [CrossRef]
- Wilbert, N.; Song, W. New contributions to the marine benthic ciliates from the Antarctic area, including description of seven new species (Protozoa, Ciliophora). J. Nat. Hist. 2005, 39, 935–973. [Google Scholar] [CrossRef]
- Wang, C.; Jiang, L.; Qu, Z.; Al-Farraj, S.A.; Hu, X. Taxonomy and phylogeny of three species of Dysteria (Ciliophora, Phyllopharyngea) including the description of a new species. Protist 2021, 172, 125831. [Google Scholar] [CrossRef]
- Zhao, L.; Pan, H.; Ma, H.; Xu, H. Morphological redescription of five marine cyrtophorid ciliates (Protozoa, Ciliophora). Period. Ocean Univ. China 2014, 44, 52–56. [Google Scholar] [CrossRef]
- Wu, T.; Li, Y.; Lu, B.; Shen, Z.; Song, W.; Warren, A. Morphology, taxonomy and molecular phylogeny of three marine peritrich ciliates, including two new species: Zoothamnium apoarbuscula n. sp. and Z. apohentscheli n. sp. (Protozoa, Ciliophora, Peritrichia). Mar. Life Sci. Technol. 2020, 2, 334–348. [Google Scholar] [CrossRef]
- Liu, W.; Shin, M.K.; Yi, Z.; Tan, Y. Progress in studies on the diversity and distribution of planktonic ciliates (Protista, Ciliophora) in the South China Sea. Mar. Life Sci. Technol. 2021, 3, 28–43. [Google Scholar] [CrossRef]
- Jankowski, A.V. Phylum Ciliophora Doflein, 1901. Review of Taxa. In Protista: Handbook on Zoology, 2nd ed.; Nauka: St. Petersburg, Russia, 2007; pp. 415–993. [Google Scholar]
- Chen, X.; Al-Farraj, A.; Warren, A. New considerations on the phylogeny of cyrtophorian ciliates (Protozoa, Ciliophora): Expanded sampling to understand their evolutionary relationships. Zool. Scr. 2016, 45, 334–348. [Google Scholar] [CrossRef]
- Gao, S.; Huang, J.; Li, J.; Song, W. Molecular phylogeny of the cyrtophorid ciliates (Protozoa, Ciliophora, Phyllopharyngea). PLoS ONE 2012, 7, e33198. [Google Scholar] [CrossRef]
- Hu, X.; Lin, X.; Song, W. Ciliate Atlas: Species Found in the South China Sea, China; Science Press: Beijing, China, 2019. [Google Scholar]
- Hu, X.; Suzuki, T. Light microscopical observations on two marine dysteriid ciliates from Japan, including a description of Dysteria yagiui nov. spec. (Ciliophora, Cyrtophorida). Eur. J. Protistol. 2005, 41, 29–36. [Google Scholar] [CrossRef]
- Kahl, A. Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 2. Holotricha außer den im 1. Teil behandelten Prostomata. Tierwelt Dtl. 1931, 21, 181–398. [Google Scholar]
- Jin, D.; Qu, Z.; Wei, B.; Montagnes, D.; Fan, X.; Chen, X. Two parasitic ciliates (Protozoa: Ciliophora: Phyllopharyngea) isolated from respiratory-mucus of an unhealthy beluga whale: Characterization, phylogeny and an assessment of morphological adaptations. Zool. J. Linn. Soc. 2020, 4, 1–20. [Google Scholar] [CrossRef]
- Gong, J.; Song, W. Morphology and infraciliature of two marine benthic ciliates, Dysteria procera Kahl, 1931 and Dysteria magna nov. spec. (Protozoa, Ciliophora, Cyrtophorida), from China. Eur. J. Protistol. 2003, 39, 301–310. [Google Scholar] [CrossRef]
- Park, M.H.; Min, G.S. A new marine cyrtophorid ciliate, Dysteria nabia nov. spec. (Ciliophora: Phyllopharyngea: Cyrtophorida: Dysteriidae), from South Korea. Acta Protozool. 2014, 53, 257–268. [Google Scholar] [CrossRef]
- Pan, H.; Hu, X.; Gong, J.; Lin, X.; Al-Rasheid, K.A.S.; Al-Farraj, S.A.; Warren, A. Morphological redescriptions of four marine ciliates (Ciliophora: Cyrtophorida: Dysteriidae) from Qingdao, China. Eur. J. Protistol. 2011, 47, 197–207. [Google Scholar] [CrossRef]
- Gismervik, I. Podite carrying ciliates dominate the benthic ciliate community in the kelp forest. Aquat. Microb. Ecol. 2004, 36, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Song, W.; Warren, A. Redescriptions of two marine cyrtophorid ciliates, Dysteria cristata (Gourret & Roeser, 1888) Kahl, 1931 and Dysteria monostyla (Ehrenberg, 1838) Kahl, 1931 (Protozoa, Ciliophora, Cyrtophorida), from China. Eur. J. Protistol. 2002, 38, 213–222. [Google Scholar] [CrossRef]
- Gong, J.; Song, W. Description of a new marine cyrtophorid ciliate, Dysteria derouxi nov. spec., with an updated key to 12 well-investigated Dysteria species (Ciliophora, Cyrtophorida). Eur. J. Protistol. 2004, 40, 13–19. [Google Scholar] [CrossRef]
- Liu, W.; Gong, J.; Lin, X.; Shen, Z.; Li, J.; Zhu, M.; Wang, M.; Song, W. Redescriptions of two cyrtophorid ciliates, Dysteria procera Kahl, 1931 and Pseudochilodonopsis marina Song, 1991 (Protozoa, Ciliophora). Acta Hydrobiol. Sin. 2008, 32, 84–89. [Google Scholar] [CrossRef]
- Pan, H.; Jiang, J.; Fan, X.; Al-Farraj, S.A.; Gao, S. Phylogeny and taxonomy of five poorly known species of cyrtophorian ciliates (Protozoa: Ciliophora: Phyllopharyngea) from China Seas. Zool. J. Linn. Soc. 2017, 180, 475–492. [Google Scholar] [CrossRef]
- Fauré-Fremiet, E. Morphologie des Dysteriidae (Ciliata Cyrtophorina). Comptes Rendus Hebd. Des Seances De L Acad. Des Sci. 1965, 260, 6679–6684. [Google Scholar]
- Claparède, E.; Lachmann, J. Études sur les infusoires et les rhizopodes. Mém. Inst. Natn. Genèv. 1859, 5, 1–260. [Google Scholar]
- Faria, J.G.; da Cunha, A.M.; Pinto, C. Estudos sobre protozoarios do mar. Mem. Inst. Oswaldo Cruz. 1922, 15, 186–208. [Google Scholar] [CrossRef]
- Song, W.; Packroff, G. Taxonomische Untersuchungen an marinen Ciliaten aus China mit Beschreibungen von zwei neuen Arten, Strombidium globosaneum nov. spec. und S. platum nov. spec. (Protozoa, Ciliophora). Arch. Protistenkd. 1997, 147, 331–360. [Google Scholar] [CrossRef]
- Wilbert, N. Eine verbesserte Technik der Protargolimpräg-nation für Ciliaten. Mikrokosmos 1975, 64, 171–179. [Google Scholar]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Osnat, P.; Eyal, P.; Haim, A.; Giddy, L.; Dan, G.; Tal, P. Guidance: A web server for assessing alignment confidence scores. Nucleic Acids Res. 2010, 38, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J. MrModeltest V2. Program Distributed by the Author. Bioinformatics 2004, 24, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Page, R.D.M. Tree view: An application to display phylogenetic trees on personal computers. Comput. Appl. Bio. 1996, 12, 357–358. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Warren, A.; Zhang, Q.; Gong, J.; Miao, M.; Sun, P.; Xu, D.; Huang, J.; Yi, Z.; Song, W. The all-data-based evolutionary hypothesis of ciliated protists with a revised classification of the phylum Ciliophora (Eukaryota, Alveolata). Sci. Rep. 2016, 6, 24874. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Species | Max | Min | Mean | M | SD | CV% | n |
|---|---|---|---|---|---|---|---|---|
| Body length (μm) | bra1 | 133 | 87 | 110.9 | 111 | 2.52 | 2.27 | 25 |
| bra2 | 144 | 104 | 121.2 | 118 | 9.82 | 8.10 | 25 | |
| bra3 | 145 | 101 | 118.2 | 119 | 10.86 | 9.19 | 25 | |
| bra4 | 240 | 177 | 204.1 | 206 | 16.39 | 8.03 | 16 | |
| bra5 | 157 | 96 | 126.5 | 126 | 13.49 | 10.66 | 25 | |
| cra | 68 | 45 | 57.8 | 57 | 1.00 | 1.73 | 25 | |
| par | 56 | 42 | 50.6 | 51 | 3.91 | 7.73 | 25 | |
| Body width (μm) | bra1 | 49 | 30 | 39.3 | 40 | 0.94 | 2.39 | 25 |
| bra2 | 58 | 44 | 50.3 | 50 | 3.20 | 6.36 | 25 | |
| bra3 | 58 | 42 | 49.8 | 49 | 3.75 | 7.53 | 25 | |
| bra4 | 64 | 43 | 52.4 | 51.5 | 6.12 | 11.68 | 16 | |
| bra5 | 63 | 42 | 51.4 | 52 | 5.51 | 10.72 | 25 | |
| cra | 68 | 27 | 33.7 | 32 | 1.49 | 4.41 | 25 | |
| par | 36 | 24 | 31.2 | 32 | 2.79 | 8.94 | 25 | |
| Number of right kineties | bra1 | 5 | 5 | 5.0 | 5 | 0.00 | 0.00 | 25 |
| bra2 | 5 | 5 | 5.0 | 5 | 0.00 | 0.00 | 25 | |
| bra3 | 6 | 5 | 5.2 | 5 | 0.41 | 7.88 | 25 | |
| bra4 | 5 | 5 | 5.0 | 5 | 0.00 | 0.00 | 16 | |
| bra5 | 5 | 5 | 5.0 | 5 | 0.00 | 0.00 | 25 | |
| cra | 4 | 4 | 4.0 | 4 | 0.00 | 0.00 | 25 | |
| par | 4 | 4 | 4.0 | 4 | 0.00 | 0.00 | 25 | |
| Number of frontoventral kineties | bra1 | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 |
| bra2 | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 | |
| bra3 | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 | |
| bra4 | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 16 | |
| bra5 | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 | |
| cra | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 | |
| par | 2 | 2 | 2.0 | 2 | 0.00 | 0.00 | 25 | |
| Number of left kineties | bra1 | 9 | 5 | 6.9 | 7 | 0.23 | 5.04 | 25 |
| bra2 | 9 | 4 | 6.7 | 7 | 1.46 | 21.79 | 25 | |
| bra3 | 8 | 3 | 6.2 | 6 | 1.36 | 21.93 | 25 | |
| bra4 | 8 | 6 | 7.1 | 7 | 0.81 | 11.41 | 16 | |
| bra5 | 10 | 4 | 6.6 | 7 | 1.50 | 22.73 | 25 | |
| cra | 9 | 5 | 6.9 | 7 | 0.23 | 3.28 | 25 | |
| par | 8 | 4 | 5.8 | 6 | 0.97 | 16.72 | 25 | |
| Number of basal bodies in a frontoventral kinety | bra1 | 283 | 182 | 225.8 | 223 | 5.54 | 2.45 | 25 |
| bra2 | 293 | 206 | 258 | 253 | 21.40 | 8.30 | 25 | |
| bra3 | 271 | 186 | 217.9 | 217 | 26.17 | 12.01 | 25 | |
| bra4 | 353 | 188 | 272.2 | 288.5 | 50.88 | 18.69 | 25 | |
| bra5 | 261 | 189 | 218.3 | 215 | 22.79 | 10.44 | 25 | |
| cra | 131 | 93 | 109.0 | 107 | 1.92 | 1.76 | 25 | |
| par | 110 | 84 | 95.3 | 95 | 7.13 | 7.48 | 25 | |
| Number of basal bodies in terminal fragment | bra1 | 17 | 11 | 14.0 | 14 | 0.29 | 2.06 | 25 |
| bra2 | 17 | 9 | 13.2 | 13 | 2.07 | 15.68 | 25 | |
| bra3 | 16 | 8 | 12.1 | 12 | 2.30 | 19.01 | 25 | |
| bra4 | 17 | 10 | 14.1 | 14.5 | 2.11 | 14.96 | 16 | |
| bra5 | 15 | 9 | 11.6 | 12 | 1.66 | 14.31 | 25 | |
| cra | 7 | 3 | 5.5 | 6 | 0.17 | 3.18 | 25 | |
| par | 7 | 4 | 5.4 | 5 | 0.76 | 14.07 | 25 | |
| Number of basal bodies in equatorial fragment | bra1 | 33 | 8 | 17.4 | 14 | 1.52 | 8.75 | 25 |
| bra2 | 26 | 7 | 15.6 | 14 | 5.69 | 36.47 | 25 | |
| bra3 | 22 | 7 | 13 | 13 | 4.07 | 31.31 | 25 | |
| bra4 | 30 | 9 | 20.6 | 21.5 | 6.27 | 30.44 | 16 | |
| bra5 | 25 | 7 | 13.5 | 12 | 4.66 | 34.57 | 25 | |
| cra | 20 | 5 | 11.6 | 11 | 1.01 | 8.72 | 25 | |
| par | 18 | 4 | 11.2 | 12 | 4.37 | 39.02 | 25 | |
| Length of macronucleus (μm) | bra1 | 40 | 24 | 33.6 | 33 | 0.70 | 2.10 | 25 |
| bra2 | 59 | 31 | 42.96 | 42 | 6.60 | 15.36 | 25 | |
| bra3 | 60 | 23 | 42.2 | 43 | 9.23 | 21.87 | 25 | |
| bra4 | 67 | 44 | 53.1 | 53 | 6.55 | 12.34 | 16 | |
| bra5 | 75 | 40 | 55.6 | 53 | 9.64 | 17.34 | 25 | |
| cra | 27 | 14 | 19.9 | 20 | 0.54 | 2.70 | 25 | |
| par | 27 | 16 | 23.4 | 23 | 2.27 | 9.70 | 25 | |
| Width of macronucleus (μm) | bra1 | 19 | 12 | 14.8 | 15 | 0.32 | 2.15 | 25 |
| bra2 | 27 | 17 | 22.3 | 22 | 2.56 | 11.48 | 25 | |
| bra3 | 38 | 11 | 23.0 | 21 | 7.33 | 31.87 | 25 | |
| bra4 | 32 | 18 | 24.4 | 23 | 4.32 | 17.70 | 16 | |
| bra5 | 41 | 19 | 27.6 | 27 | 6.01 | 21.78 | 25 | |
| cra | 11 | 6 | 8.8 | 9 | 0.27 | 3.08 | 25 | |
| par | 19 | 12 | 14.9 | 15 | 2.27 | 15.23 | 25 | |
| Diameter of glandule | bra1 | 10 | 7 | 8.4 | 8 | 0.13 | 1.53 | 25 |
| bra2 | 8 | 4 | 7.1 | 7 | 0.91 | 12.81 | 25 | |
| bra3 | 12 | 5 | 7.9 | 8 | 1.82 | 23.04 | 25 | |
| bra4 | 11 | 7 | 8.6 | 9 | 1.15 | 13.37 | 16 | |
| bra5 | 11 | 7 | 8.4 | 8 | 1.35 | 16.07 | 25 | |
| cra | 6 | 3 | 4.6 | 5 | 0.15 | 3.32 | 25 | |
| par | 6 | 3 | 3.6 | 3 | 0.81 | 22.50 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhang, H.; Zhang, Q.; Qu, Z.; Warren, A.; Wu, D.; Chen, X. A Case Study of the Morphological and Molecular Variation within a Ciliate Genus: Taxonomic Descriptions of Three Dysteria Species (Ciliophora, Cyrtophoria), with the Establishment of a New Species. Int. J. Mol. Sci. 2022, 23, 1764. https://doi.org/10.3390/ijms23031764
Zhao X, Zhang H, Zhang Q, Qu Z, Warren A, Wu D, Chen X. A Case Study of the Morphological and Molecular Variation within a Ciliate Genus: Taxonomic Descriptions of Three Dysteria Species (Ciliophora, Cyrtophoria), with the Establishment of a New Species. International Journal of Molecular Sciences. 2022; 23(3):1764. https://doi.org/10.3390/ijms23031764
Chicago/Turabian StyleZhao, Xuetong, Hui Zhang, Qianqian Zhang, Zhishuai Qu, Alan Warren, Di Wu, and Xiangrui Chen. 2022. "A Case Study of the Morphological and Molecular Variation within a Ciliate Genus: Taxonomic Descriptions of Three Dysteria Species (Ciliophora, Cyrtophoria), with the Establishment of a New Species" International Journal of Molecular Sciences 23, no. 3: 1764. https://doi.org/10.3390/ijms23031764