
The Evidence for Effective Inhibition of INa Produced by Mirogabalin ((1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid), a Known Blocker of CaV Channels

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Inhibitory Effect of MGB on Voltage-Gated Na+ Current (INa) Measured from Pituitary GH3 Cells
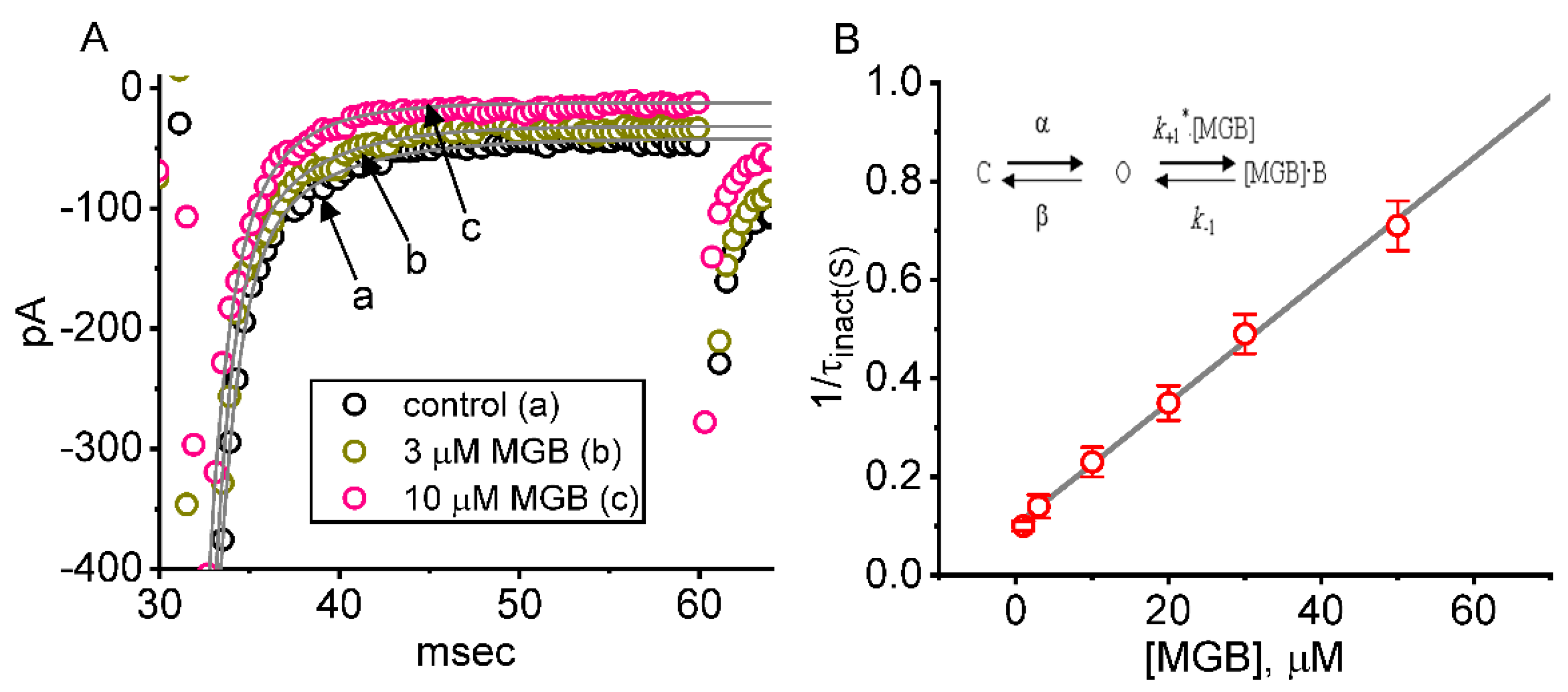
2.2. Kinetic Evaluation of Time-Dependent Block by MGB on INa(T) in GH3 Cells
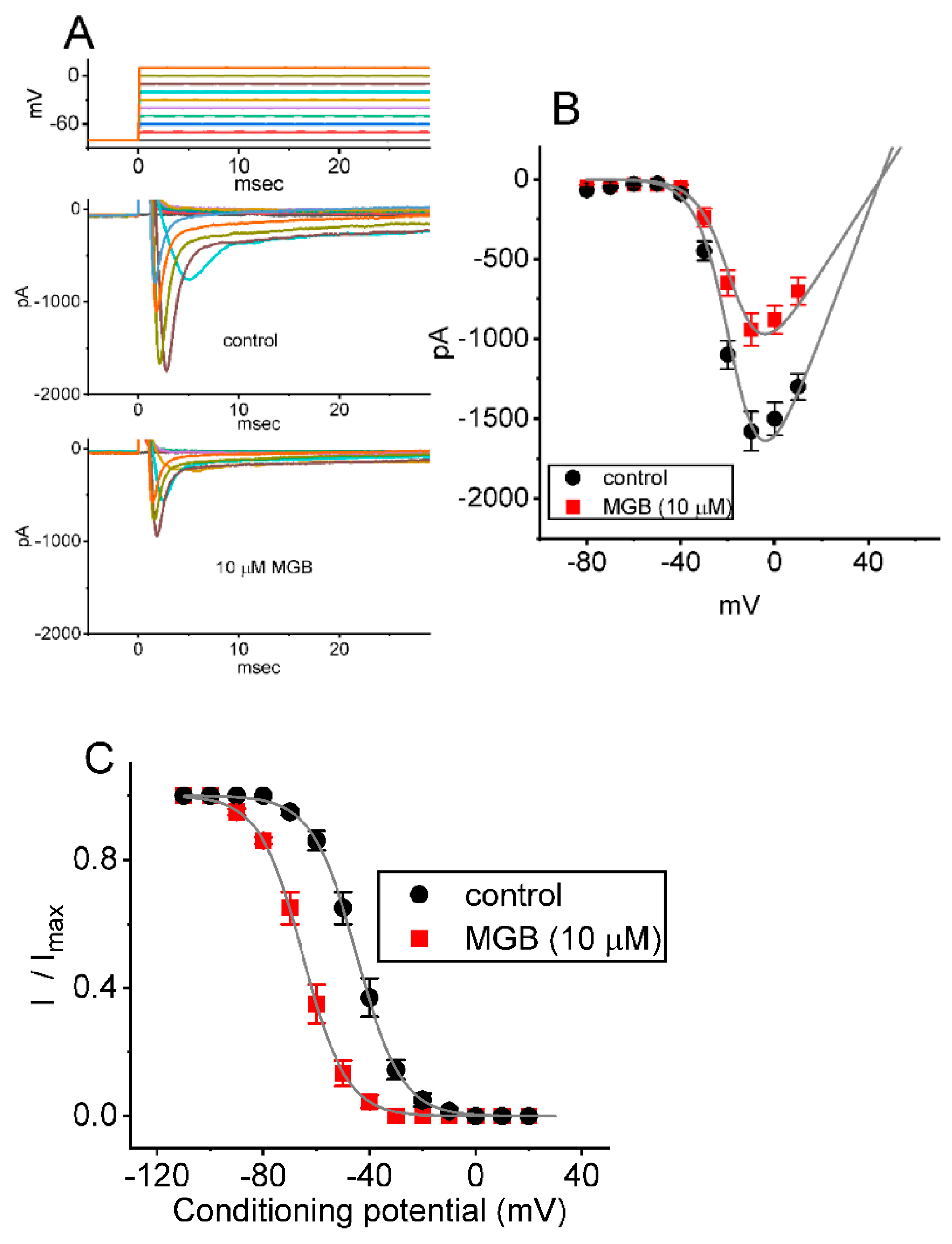
2.3. Mean Current-Voltage (I-V) Relationship of INa(T) Caused by MGB
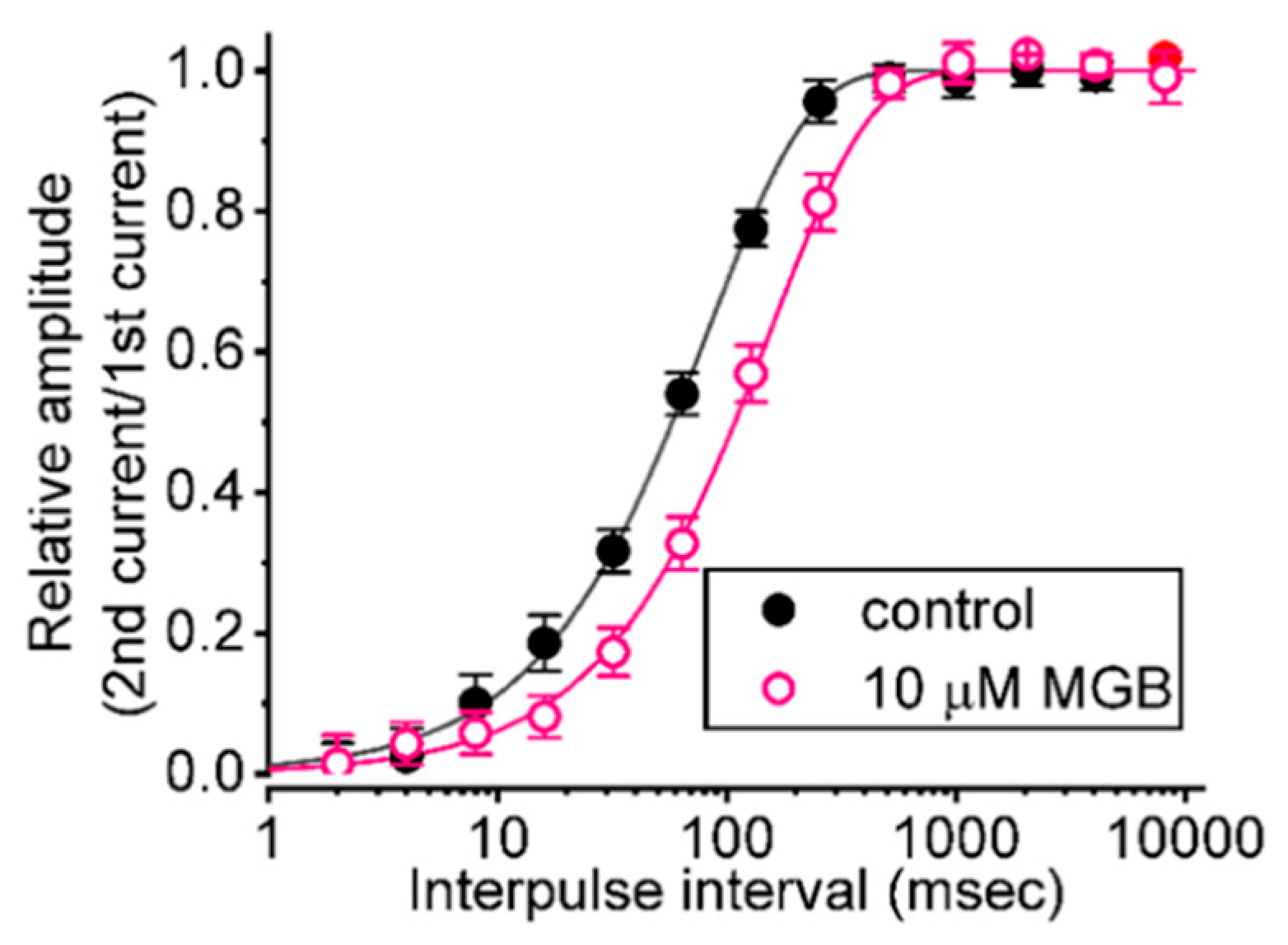
2.4. Effect of MGB on the Recovery from INa(T) Inactivation Evoked during Varying Interpulse Intervals
2.5. Effect of MGB on the Window Component of INa (INa(W)) Measured from GH3 Cells
2.6. Suppressive Effect of MGB on Resurgent INa (INa(R)) Seen in GH3 Cells
2.7. Effect of MGB on the Hysteretic Behavior of Persistent Na+ Current (INa(P)) Triggered by Isosceles Triangular Ramp Voltage (Vramp)
2.8. MGB-Induced Increase in Cumulative Inhibition of INa(T) Inactivation
3. Discussion
4. Materials and Methods
4.1. Chemicals, Drugs and Solutions Used in This Work
4.2. Cell Culture
4.3. Electrophysiological Measurements
4.4. Data Recordings and Analyses
4.5. Curve-Fitting Procedures and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CaV channel | voltage-gated Ca2+ channel |
| I-V | current versus voltage |
| IC50 | concentration required for 50% inhibition |
| INa | voltage-gated Na+ current |
| INa(L) | late Na+ current |
| INa(P) | persistent Na+ current |
| INa(R) | resurgent Na+ current |
| INa(T) | transient Na+ current |
| INa(W) | window Na+ current |
| KD | dissociation constant |
| MGB | mirogabalin (Tarlige®, 1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid) |
| NaV channel | voltage-gated Na+ channel |
| SEM | standard error of the mean |
| TEA | tetraethylammonium chloride |
| Tef | tefluthrin |
| TTX | tetrodotoxin |
| Vhys | voltage-dependent hysteresis |
| Vramp | ramp voltage |
References
- Calandre, E.P.; Rico-Villademoros, F.; Slim, M. Alpha2delta ligands, gabapentin, pregabalin and mirogabalin: A review of their clinical pharmacology and therapeutic use. Expert Rev. Neurother. 2016, 16, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Hutmacher, M.M.; Frame, B.; Miller, R.; Truitt, K.; Merante, D. Exposure-response modeling of average daily pain score, and dizziness and somnolence, for mirogabalin (DS-5565) in patients with diabetic peripheral neuropathic pain. J. Clin. Pharm. 2016, 56, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Domon, Y.; Kitano, Y.; Makino, M. Analgesic effects of the novel α2δ ligand mirogabalin in a rat model of spinal cord injury. Pharmazie 2018, 73, 659–661. [Google Scholar] [PubMed]
- Domon, Y.; Arakawa, N.; Inoue, T.; Matsuda, F.; Takahashi, M.; Yamamura, N.; Kai, K.; Kitano, Y. Binding characteristics and analgesic effects of mirogabalin, a novel ligand for the α2δ subunit of voltage-gated calcium channels. J. Pharm. Exp. 2018, 365, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Javed, S.; Alam, U.; Malik, R.A. Mirogabalin and emerging therapies for diabetic neuropathy. J. Pain Res. 2018, 11, 1559–1566. [Google Scholar] [CrossRef] [Green Version]
- Deeks, E.D. Mirogabalin: First global approval. Drugs 2019, 79, 463–468. [Google Scholar] [CrossRef]
- Kitano, Y.; Wakimoto, S.; Tamura, S.; Kubota, K.; Domon, Y.; Arakawa, N.; Saito, M.; Sava, B.; Buisson, B. Effects of mirogabalin, a novel ligand for the α₂δ subunit of voltage-gated calcium channels, on N-type calcium channel currents of rat dorsal root ganglion culture neurons. Pharmazie 2019, 74, 147–149. [Google Scholar]
- Saeki, K.; Yasuda, S.I.; Kato, M.; Kano, M.; Domon, Y.; Arakawa, N.; Kitano, Y. Analgesic effects of mirogabalin, a novel ligand for α2δ subunit of voltage-gated calcium channels, in experimental animal models of fibromyalgia. Naunyn Schmiedebergs Arch. Pharm. 2019, 392, 723–728. [Google Scholar] [CrossRef]
- Baba, M.; Matsui, N.; Kuroha, M.; Wasaki, Y.; Ohwada, S. Long-term safety and efficacy of mirogabalin in Asian patients with diabetic peripheral neuropathic pain. J. Diabetes Investig. 2020, 11, 693–698. [Google Scholar] [CrossRef]
- Burgess, J.; Javed, S.; Frank, B.; Malik, R.A.; Alam, U. Mirogabalin besylate in the treatment of neuropathic pain. Drugs Today 2020, 56, 135–149. [Google Scholar] [CrossRef]
- Kato, J.; Matsui, N.; Kakehi, Y.; Murayama, E.; Ohwada, S. Long-term safety and efficacy of mirogabalin in Asian patients with postherpetic neuralgia: Results from an open-label extension of a multicenter randomized, double-blind, placebo-controlled trial. Medicine 2020, 99, e21976. [Google Scholar] [CrossRef] [PubMed]
- Murasawa, H.; Kobayashi, H.; Saeki, K.; Kitano, Y. Anxiolytic effects of the novel α2δ ligand mirogabalin in a rat model of chronic constriction injury, an experimental model of neuropathic pain. Psychopharmacology 2020, 237, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Murasawa, H.; Kobayashi, H.; Yasuda, S.I.; Saeki, K.; Domon, Y.; Arakawa, N.; Kubota, K.; Kitano, Y. Anxiolytic-like effects of mirogabalin, a novel ligand for α2δ ligand of voltage-gated calcium channels, in rats repeatedly injected with acidic saline intramuscularly, as an experimental model of fibromyalgia. Pharm. Rep. 2020, 72, 571–579. [Google Scholar] [CrossRef]
- Tetsunaga, T.; Tetsunaga, T.; Nishida, K.; Misawa, H.; Takigawa, T.; Yamane, K.; Tsuji, H.; Takei, Y.; Ozaki, T. Short-term outcomes of mirogabalin in patients with peripheral neuropathic pain: A retrospective study. J. Orthop. Surg. Res. 2020, 15, 191. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Gray, E.; Wang, Y.F.; Wang, S.J. Cost-effectiveness of mirogabalin for the treatment of post-herpetic neuralgia in Taiwan. J. Med. Econ. 2020, 23, 529–536. [Google Scholar] [CrossRef]
- Oyama, M.; Watanabe, S.; Iwai, T.; Tanabe, M. Mirogabalin activates the descending noradrenergic system by binding to the α2δ-1 subunit of voltage-gated Ca2+ channels to generate analgesic effects. J. Pharmacol. Sci. 2021, 146, 33–39. [Google Scholar] [CrossRef]
- Murasawa, H.; Pawlak, A.; Kobayashi, H.; Saeki, K.; Yasuda, S.I.; Kitano, Y. Mirogabalin, a novel ligand for α2δ subunit of voltage-gated calcium channels, improves cognitive impairments in repeated intramuscular acidic saline injection model rats, an experimental model of fibromyalgia. Biomed. Pharm. 2021, 139, 111647. [Google Scholar] [CrossRef]
- Bauer, C.S.; Nieto-Rostro, M.; Rahman, W.; Tran-Van-Minh, A.; Ferron, L.; Douglas, L.; Kadurin, I.; Sri Ranjan, Y.; Fernandez-Alacid, L.; Millar, N.S.; et al. The increased trafficking of the calcium channel subunit alpha2delta-1 to presynaptic terminals in neuropathic pain is inhibited by the alpha2delta ligand pregabalin. J. Neurosci. 2009, 29, 4076–4088. [Google Scholar] [CrossRef] [Green Version]
- Alexander, S.P.; Catterall, W.A.; Kelly, E.; Marrion, N.; Peters, J.A.; Benson, H.E.; Faccenda, E.; Pawson, A.J.; Sharman, J.L.; Southan, C.; et al. The Concise guide to PHARMACOLOGY 2015/16: Voltage-gated ion channels. Br. J. Pharm. 2015, 172, 5904–5941. [Google Scholar] [CrossRef] [Green Version]
- Zajączkowska, R.; Mika, J.; Leppert, W.; Kocot-Kępska, M.; Malec-Milewska, M.; Wordliczek, J. Mirogabalin-A novel selective ligand for the α2δ calcium channel subunit. Pharmaceuticals 2021, 14, 112. [Google Scholar] [CrossRef]
- Simasko, S.M. A background sodium conductance is necessary for spontaneous depolarizations in rat pituitary cell line GH3. Am. J. Physiol. 1994, 266, C709–C719. [Google Scholar] [CrossRef] [PubMed]
- England, S.; Rawson, D. Isoform-selective voltage-gated Na+ channel modulators as next-generation analgesics. Future Med. Chem. 2010, 2, 775–790. [Google Scholar] [CrossRef] [PubMed]
- Jukič, M.; Kikelj, D.; Anderluh, M. Isoform selective voltage-gated sodium channel modulators and the therapy of pain. Curr. Med. Chem. 2014, 21, 164–186. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; So, E.C.; Liao, Y.K.; Huang, Y.M. Reversal by ranolazine of doxorubicin-induced prolongation in the inactivation of late sodium current in rat dorsal root ganglion neurons. Pain Med. 2015, 16, 1032–1034. [Google Scholar] [CrossRef] [Green Version]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Cummins, T.R.; Sheets, L.; Waxman, S.G. The roles of sodium channels in nociception: Implications for mechanisms of pain. Pain 2007, 131, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Lampert, A.; O’Reilly, A.O.; Reeh, P.; Leffler, A. Sodium channelopathies and pain. Pflugers Arch. 2010, 460, 249–263. [Google Scholar] [CrossRef]
- Bannister, K.; Kucharczyk, M.; Dickenson, A.H. Hopes for the future of pain control. Pain Ther. 2017, 6, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Coates, M.D.; Vrana, K.E.; Ruiz-Velasco, V. The influence of voltage-gated sodium channels on human gastrointestinal nociception. Neurogastroenterol. Motil. 2019, 31, e13460. [Google Scholar] [CrossRef]
- Rühlmann, A.H.; Körner, J.; Hausmann, R.; Bebrivenski, N.; Neuhof, C.; Detro-Dassen, S.; Hautvast, P.; Benasolo, C.A.; Meents, J.; Machtens, J.-P.; et al. Uncoupling sodium channel dimers restores the phenotype of a pain-linked Nav 1.7 channel mutation. Br. J. Pharmacol. 2020, 177, 4481–4496. [Google Scholar] [CrossRef]
- Lo, Y.C.; Tseng, Y.T.; Liu, C.M.; Wu, B.N.; Wu, S.N. Actions of KMUP-1, a xanthine and piperazine derivative, on voltage-gated Na+ and Ca2+ -activated K+ currents in GH3 pituitary tumour cells. Br. J. Pharmacol. 2015, 172, 5110–5122. [Google Scholar] [CrossRef] [PubMed]
- So, E.C.; Wu, S.N.; Lo, Y.C.; Su, K. Differential regulation of tefluthrin and telmisartan on the gating charges of INa activation and inactivation as well as on resurgent and persistent INa in a pituitary cell line (GH3). Toxicol. Lett. 2018, 285, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.; Kao, Z.H.; Lee, S.W.; Wu, S.N. Effects of sesamin, the major furofuran lignan of sesame oil, on the amplitude and gating of voltage-gated Na+ and K+ currents. Molecules 2020, 25, 3062. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Wu, S.N.; Huang, C.W. The specific effects of OD-1, a peptide activator, on voltage-gated sodium current and seizure susceptibility. Int. J. Mol. Sci. 2020, 21, 8254. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Characterization of direct perturbations on voltage-gated sodium current by esaxerenone, a nonsteroidal mineralocorticoid receptor blocker. Biomedicines 2021, 9, 549. [Google Scholar] [CrossRef]
- Chang, W.T.; Wu, S.N. Effectiveness of columbianadin, a bioactive coumarin derivative, in perturbing transient and persistent INa. Int. J. Mol. Sci. 2021, 22, 621. [Google Scholar] [CrossRef]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. The evidence for sparsentan-mediated inhibition of INa and IK(erg): Possibly unlinked to its antagonism of angiotensin II or endothelin type A receptor. Biomedicines 2021, 10, 86. [Google Scholar] [CrossRef]
- Wu, S.N.; Chen, B.S.; Hsu, T.I.; Peng, H.; Wu, Y.H.; Lo, Y.C. Analytical studies of rapidly inactivating and noninactivating sodium currents in differentiated NG108-15 neuronal cells. J. Theor. Biol. 2009, 259, 828–836. [Google Scholar] [CrossRef]
- Morris, C.E.; Boucher, A.; Joós, B. Left-shifted nav channels in injured bilayer: Primary targets for neuroprotective nav antagonists? Front. Pharmacol. 2012, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Frenz, C.T.; Hansen, A.; Dupuis, N.D.; Shultz, N.; Levinson, S.R.; Finger, T.E.; Dionne, V.E. NaV1.5 sodium channel window currents contribute to spontaneous firing in olfactory sensory neurons. J. Neurophysiol. 2014, 112, 1091–1104. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.N.; Lo, Y.C.; Shen, A.Y.; Chen, B.S. Contribution of non-inactivating Na+ current induced by oxidizing agents to the firing behavior of neuronal action potentials: Experimental and theoretical studies from NG108-15 neuronal cells. Chin. J. Physiol. 2011, 54, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, J.; Soderlund, D.M. Actions of tefluthrin on rat Na(v)1.7 voltage-gated sodium channels expressed in Xenopus Oocytes. Pestic. Biochem. Physiol. 2011, 101, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaliq, Z.M.; Gouwens, N.W.; Raman, I.M. The contribution of resurgent sodium current to high-frequency firing in Purkinje neurons: An experimental and modeling study. J. Neurosci. 2003, 23, 4899–4912. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.H.; Raman, I.M. Resurgent current of voltage-gated Na+ channels. J. Physiol. 2014, 592, 4825–4838. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, S.; Seki, S.; Terman, D.H.; Pantazis, A.; Olcese, R.; Wiedau-Pazos, M.; Chandler, S.H. Resurgent Na+ current offers noise modulation in bursting neurons. PLoS Comput. Biol. 2019, 15, e1007154. [Google Scholar] [CrossRef]
- Quattrocolo, G.; Dunville, K.; Nigro, M.J. Resurgent sodium current in neurons of the cerebral cortex. Front. Cell Neurosci. 2021, 15, 760610. [Google Scholar] [CrossRef]
- Villalba-Galea, C.A.; Chiem, A.T. Hysteretic behavior in voltage-gated channels. Front. Pharmacol. 2020, 11, 579596. [Google Scholar] [CrossRef]
- Guérineau, N.C.; Monteil, A.; Lory, P. Sodium background currents in endocrine/neuroendocrine cells: Towards unraveling channel identity and contribution in hormone secretion. Front. Neuroendocrinol. 2021, 63, 100947. [Google Scholar] [CrossRef]
- Taddese, A.; Bean, B.P. Subthreshold sodium current from rapidly inactivating sodium channels drives spontaneous firing of tuberomammillary neurons. Neuron 2002, 33, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Navarro, M.A.; Salari, A.; Lin, J.L.; Cowan, L.M.; Penington, N.J.; Milescu, M.; Milescu, L.S. Sodium channels implement a molecular leaky integrator that detects action potentials and regulates neuronal firing. Elife 2020, 9, e54940. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.V.; Espinosa, J.L.; López-Domínguez, A.M.; López-Santiago, L.F.; Navarrete, A.; Cota, G. L-type calcium channel activation up-regulates the mRNAs for two different sodium channel alpha subunits (Nav1.2 and Nav1.3) in rat pituitary GH3 cells. Brain Res. Mol. Brain Res. 2003, 116, 115–125. [Google Scholar] [CrossRef]
- Lo, Y.K.; Wu, S.N.; Lee, C.T.; Li, H.F.; Chiang, H.T. Characterization of action potential waveform-evoked L-type calcium currents in pituitary GH3 cells. Pflugers Arch. 2001, 442, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Soliman, D.; Wang, L.; Hamming, K.S.; Yang, W.; Fatehi, M.; Carter, C.C.; Clanachan, A.S.; Light, P.E. Late sodium current inhibition alone with ranolazine is sufficient to reduce ischemia- and cardiac glycoside-induced calcium overload and contractile dysfunction mediated by reverse-mode sodium/calcium exchange. J. Pharmacol. Exp. Ther. 2012, 343, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Shenoda, B. The role of Na+/Ca2+ exchanger subtypes in neuronal ischemic injury. Transl. Stroke Res. 2015, 6, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endocr. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef] [Green Version]
- Gould, H.J., 3rd; Garrett, C.; Donahue, R.R.; Paul, D.; Diamond, I.; Taylor, B.K. Ranolazine attenuates behavioral signs of neuropathic pain. Behav. Pharmacol. 2009, 20, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Gould, H.J., 3rd; Soignier, R.D.; Cho, S.R.; Hernandez, C.; Diamond, I.; Taylor, B.K.; Paul, D. Ranolazine attenuates mechanical allodynia associated with demyelination injury. Pain Med. 2014, 15, 1771–1780. [Google Scholar] [CrossRef] [Green Version]
- Gould, H.J., 3rd; Diamond, I. Ranolazine: A potential treatment for refractory neuropathic pain. J. Neurol. Sci. 2016, 369, 310–311. [Google Scholar] [CrossRef]
- Elkholy, S.E.; Elaidy, S.M.; El-Sherbeeny, N.A.; Toraih, E.A.; El-Gawly, H.W. Neuroprotective effects of ranolazine versus pioglitazone in experimental diabetic neuropathy: Targeting Nav1.7 channels and PPAR-γ. Life Sci. 2020, 250, 117557. [Google Scholar] [CrossRef]
- Casey, G.P.; Roberts, J.S.; Paul, D.; Diamond, I.; Gould, H.J., 3rd. Ranolazine attenuation of CFA-induced mechanical hyperalgesia. Pain Med. 2010, 11, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Nodera, H.; Rutkove, S.B. Changes of the peripheral nerve excitability in vivo induced by the persistent Na+ current blocker ranolazine. Neurosci. Lett. 2012, 518, 36–40. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J., 3rd. Response to letter to the editor: Reversal by ranolazine of doxorubicin-induced prolongation in the inactivation of late sodium current in rat dorsal root ganglion neurons. Pain Med 2015, 16, 1035. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; Mendell, J.; Ohwada, S.; Hsu, C.; He, L.; Warren, V.; Dishy, V.; Zahir, H. Tolerability, pharmacokinetics, and pharmacodynamics of mirogabalin in healthy subjects: Results from phase 1 studies. Pharmacol. Res. Perspect. 2018, 6, e00418. [Google Scholar] [CrossRef] [PubMed]
- Jansen, M.; Warrington, S.; Dishy, V.; Ohwada, S.; Johnson, L.; Brown, K.; Ishizuka, H. A randomized, placebo-controlled, double-blind study of the safety, tolerability, pharmacokinetics, and pharmacodynamics of single and repeated doses of mirogabalin in healthy Asian volunteers. Clin. Pharmacol. Drug Dev. 2018, 7, 661–669. [Google Scholar] [CrossRef] [Green Version]
- Amir, R.; Argoff, C.E.; Bennett, G.J.; Cummins, T.R.; Durieux, M.E.; Gerner, P.; Gold, M.S.; Porreca, F.; Strichartz, G.R. The role of sodium channels in chronic inflammatory and neuropathic pain. J. Pain 2006, 7, S1–S29. [Google Scholar] [CrossRef]
- Wu, S.N.; Peng, H.; Chen, B.S.; Wang, Y.J.; Wu, P.Y.; Lin, M.W. Potent activation of large-conductance Ca2+-activated K+ channels by the diphenylurea 1,3-bis-[2-hydroxy-5-(trifluoromethyl)phenyl]urea (NS1643) in pituitary tumor (GH3) cells. Mol. Pharmacol. 2008, 74, 1696–1704. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.T.; Liu, P.Y.; Gao, Z.H.; Lee, S.W.; Lee, W.K.; Wu, S.N. Evidence for the effectiveness of remdesivir (GS-5734), a nucleoside-analog antiviral drug in the inhibition of IK(M) or IK(DR) and in the stimulation of IMEP. Front. Pharmacol. 2020, 11, 1091. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.-L.; Chuang, C.-W.; Cho, H.-Y.; Chuang, T.-H.; Wu, S.-N. The Evidence for Effective Inhibition of INa Produced by Mirogabalin ((1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid), a Known Blocker of CaV Channels. Int. J. Mol. Sci. 2022, 23, 3845. https://doi.org/10.3390/ijms23073845
Wu C-L, Chuang C-W, Cho H-Y, Chuang T-H, Wu S-N. The Evidence for Effective Inhibition of INa Produced by Mirogabalin ((1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid), a Known Blocker of CaV Channels. International Journal of Molecular Sciences. 2022; 23(7):3845. https://doi.org/10.3390/ijms23073845
Chicago/Turabian StyleWu, Chao-Liang, Chao-Wei Chuang, Hsin-Yen Cho, Tzu-Hsien Chuang, and Sheng-Nan Wu. 2022. "The Evidence for Effective Inhibition of INa Produced by Mirogabalin ((1R,5S,6S)-6-(aminomethyl)-3-ethyl-bicyclo [3.2.0] hept-3-ene-6-acetic acid), a Known Blocker of CaV Channels" International Journal of Molecular Sciences 23, no. 7: 3845. https://doi.org/10.3390/ijms23073845