
Exploring the Antimicrobial Potential and Biofilm Inhibitory Properties of Hemocyanin from Hemifusus pugilinus (Born, 1778)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
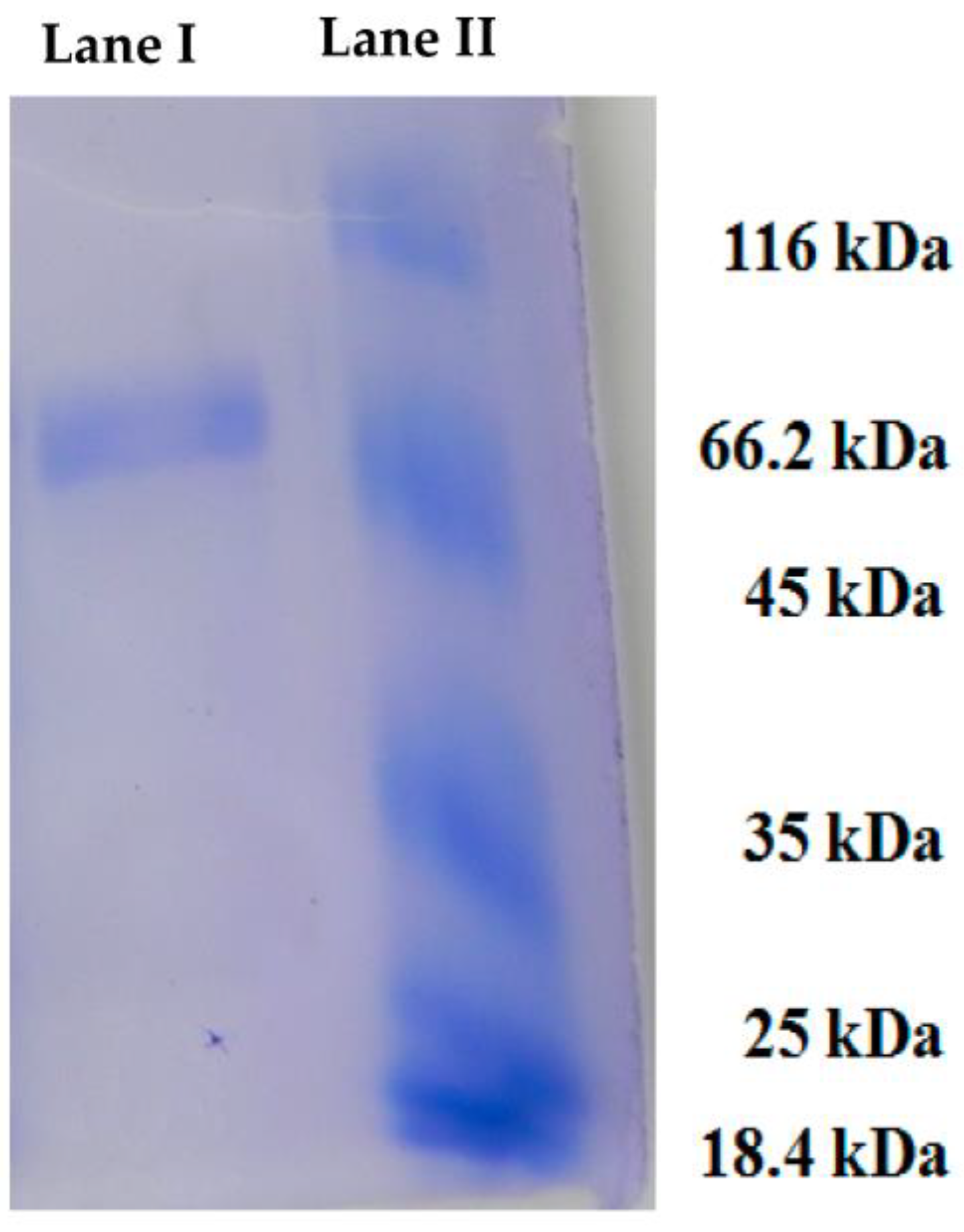
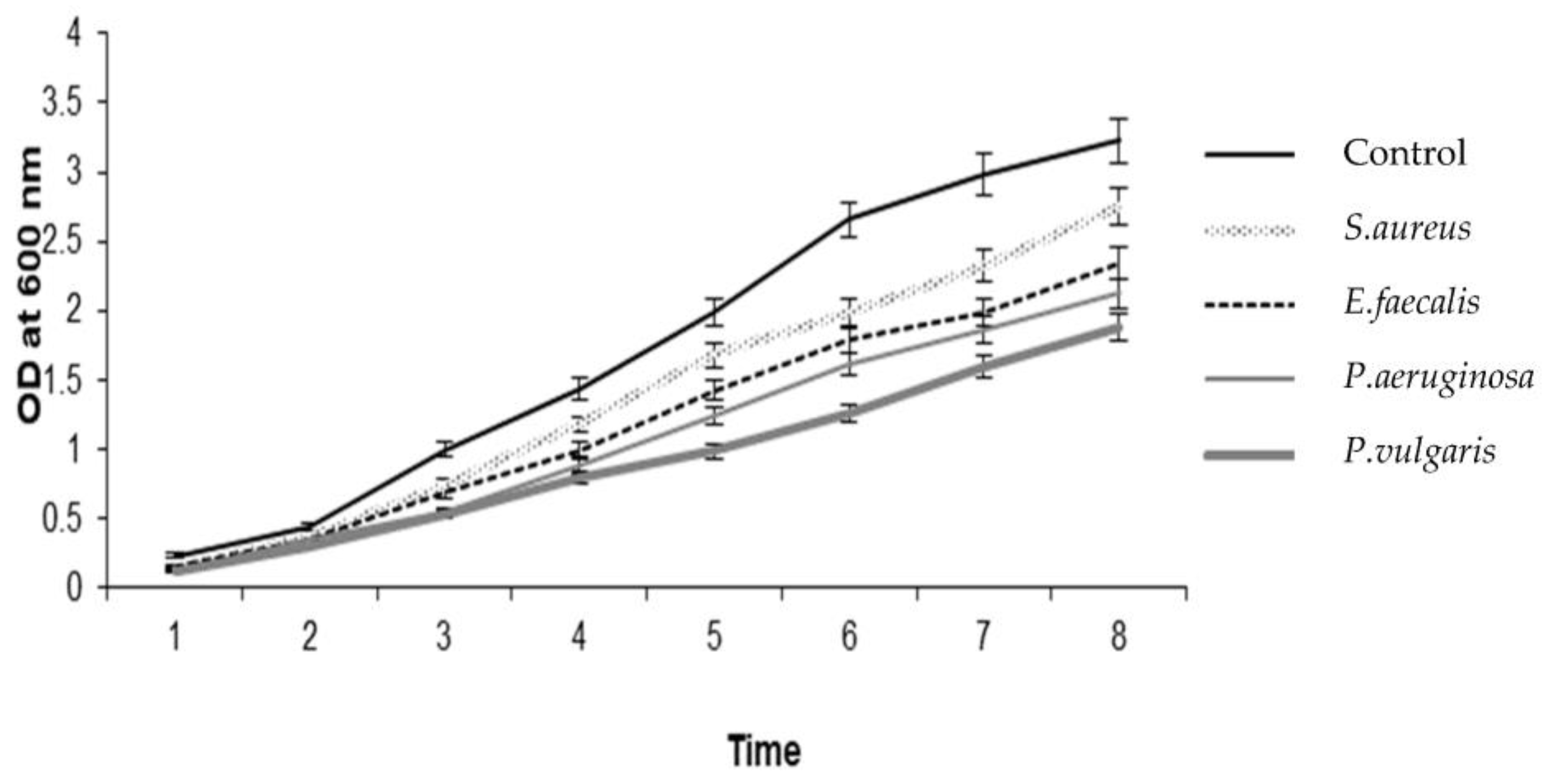
2. Results and Discussion
3. Materials and Methods
3.1. Animals
3.2. Purification of H. pugilinus Hemocyanin
3.3. Minimum Inhibitory Concentration
3.4. Antibiofilm Assay
3.5. Antifungal Activity
3.6. Statistical Analysis
3.7. Cell Cytotoxicity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Suleria, H.A.R.; Hines, B.M.; Addepalli, R.; Chen, W.; Masci, P.; Gobe, G.; Osborne, S.A. In vitro Anti-Thrombotic Activity of Extracts from Blacklip Abalone (Haliotis rubra) Processing Waste. Mar. Drugs 2016, 15, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigariga, P.; Saren, D.; Varma, R.; Sugumar, V. Assessment of trace metal bioaccumulation on the shells of edible gastropod Chicoreus ramosus and Hemifusus pugilinus. Environ. Monit. Assess. 2023, 195, 608. [Google Scholar] [CrossRef]
- Ellis, R.P.; Parry, H.; Spicer, J.I.; Hutchinson, T.H.; Pipe, R.K.; Widdicombe, S. Immunological function in marine inverte-brates: Responses to environmental perturbation. Fish Shellfish Immunol. 2011, 30, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Markl, J. Evolution of molluscan hemocyanin structures. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2013, 1834, 1840–1852. [Google Scholar] [CrossRef] [Green Version]
- Coates, C.J.; Decker, H. Immunological properties of oxygen-transport proteins: Hemoglobin, hemocyanin and hem-erythrin. Cell. Mol. Life Sci. 2017, 74, 293–317. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.K.; Go, H.J.; Kim, C.H.; Nam, B.H.; Park, N.G. Antimicrobial peptide, hdMolluscidin, purified from the gill of the abalone, Haliotis discus. Fish Shellfish Immunol. 2016, 52, 289–297. [Google Scholar] [CrossRef]
- Dolashka, P.; Dolashki, A.; Van Beeumen, J.; Floetenmeyer, M.; Velkova, L.; Stevanovic, S.; Voelter, W. Antimicrobial Activity of Molluscan Hemocyanins from Helix and Rapana Snails. Curr. Pharm. Biotechnol. 2016, 17, 263–270. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef]
- Li, C.; Zhu, J.; Wang, Y.; Chen, Y.; Song, L.; Zheng, W.; Li, J.; Yu, R. Antibacterial Activity of AI-Hemocidin 2, a Novel N-Terminal Peptide of Hemoglobin Purified from Arca inflata. Mar. Drugs 2017, 15, 205. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, F.; Hu, Z.; Zhao, X.; Min, S.; Du, Z.; Zhao, S.; Ye, X.; Li, Y. Hemocyanin from shrimp Litopenaeus vannamei shows hemolytic activity. Fish Shellfish. Immunol. 2009, 27, 330–335. [Google Scholar] [CrossRef]
- Yan, F.; Zhang, Y.; Jiang, R.; Zhong, M.; Hu, Z.; Du, H.; Lun, J.; Chen, J.; Li, Y. Identification and agglutination properties of hemocyanin from the mud crab (Scylla serrata). Fish Shellfish. Immunol. 2011, 30, 354–360. [Google Scholar] [CrossRef]
- Petit, V.W.; Rolland, J.-L.; Blond, A.; Cazevieille, C.; Djediat, C.; Peduzzi, J.; Goulard, C.; Bachère, E.; Dupont, J.; Destoumieux-Garzón, D.; et al. A hemocyanin-derived antimicrobial peptide from the penaeid shrimp adopts an alpha-helical structure that specifically permeabilizes fungal membranes. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2016, 1860, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Babu, V.S.; Wan, Q.; Muhammad, A.; Li, J.; Lan, J.; Lin, L. Antibacterial activity of hemocyanin from red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2018, 75, 391–399. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, B.L.; Söderhäll, K. Processing of an antibacterial peptide from hemocyanin of the freshwater crayfish Pacifastacus leniusculus. J. Biol. Chem. 2003, 278, 7927–7933. [Google Scholar] [CrossRef] [Green Version]
- Manubens, A.; Salazar, F.; Haussmann, D.; Figueroa, J.; Del Campo, M.; Pinto, J.M.; Huaquín, L.; Venegas, A.; Becker, M.I. Concholepas hemocyanin biosynthesis takes place in the hepatopancreas, with hemocytes being involved in its metabolism. Cell Tissue Res. 2010, 342, 423–435. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Hemocyanin-derived phenoloxidase activity: A contributing factor to hyperpigmentation in Nephrops norvegicus. Food Chem. 2013, 140, 361–369. [Google Scholar] [CrossRef]
- Peña, J.J.; Loker, E.S.; Adema, C.M. Comparative analysis of three families of hygrophilid snails shows that the egg mass fluid protein haemocyanin-like 1 (Hcl-1) is unique to planorbids. J. Molluscan Stud. 2023, 89, eyad006. [Google Scholar] [CrossRef]
- Jiang, N.; Tan, N.S.; Ho, B.; Ding, J.L. Respiratory protein–generated reactive oxygen species as an antimicrobial strategy. Nat. Immunol. 2007, 8, 1114. [Google Scholar] [CrossRef]
- Zhuang, J.; Coates, C.J.; Zhu, H.; Zhu, P.; Wu, Z.; Xie, L. Identification of candidate antimicrobial peptides derived from abalone hemocyanin. Dev. Comp. Immunol. 2015, 49, 96–102. [Google Scholar] [CrossRef]
- De Zoysa, M.; Whang, I.; Lee, Y.; Lee, S.; Lee, J.-S.; Lee, J. Defensin from disk abalone Haliotis discus discus: Molecular cloning, sequence characterization and immune response against bacterial infection. Fish Shellfish Immunol. 2010, 28, 261–266. [Google Scholar] [CrossRef]
- Bansemir, A.; Blume, M.; Schröder, S.; Lindequist, U. Screening of cultivated seaweeds for antibacterial activity against fish pathogenic bacteria. Aquaculture 2006, 252, 79–84. [Google Scholar] [CrossRef]
- Bazes, A.; Silkina, A.; Douzenel, P.; Faÿ, F.; Kervarec, N.; Morin, D.; Bourgougnon, N. Investigation of the antifouling con-stituents from the brown alga Sargassum muticum (Yendo) Fensholt. J. Appl. Phycol. 2009, 21, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Mayer, A.M.; Rodríguez, A.D.; Berlinck, R.G.; Hamann, M.T. Marine pharmacology in 2005–6: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2009, 1790, 283–308. [Google Scholar]
- Jayaraj, S.; Thiagarajan, R.; Arumugam, M.; Mullainadhan, P. Isolation, purification and characterization of β-1,3-glucan binding protein from the plasma of marine mussel Perna viridis. Fish Shellfish. Immunol. 2008, 24, 715–725. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Blas, I.; Ruizzarzuela, I.; Cunningham, D.; Vendrell, D.; Múzquiz, J.L. The role of probiotics in aquaculture. Veter-Microbiol. 2006, 114, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2011, 28, 196–268. [Google Scholar] [CrossRef]
- Maktoob, A.; Ronald, H.T. Handbook of Natural Products from Marine Invertebrates; Phyllum Mollusca Part. 1; Harwood Academic Publishers: Reading, UK, 1997. [Google Scholar]
- Kelly, M.S. Echinoderms: Their Culture and Bioactive Compounds. In Echinodermata; Springer: Berlin/Heidelberg, Germany, 2005; pp. 139–165. [Google Scholar] [CrossRef]
- Watanabe, M.; Fuda, H.; Jin, S.; Sakurai, T.; Ohkawa, F.; Hui, S.-P.; Takeda, S.; Watanabe, T.; Koike, T.; Chiba, H. Isolation and Characterization of a Phenolic Antioxidant from the Pacific Oyster (Crassostrea gigas). J. Agric. Food Chem. 2012, 60, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Rapado, L.; Nakano, E.; Ohlweiler, F.; Kato, M.; Yamaguchi, L.; Pereira, C.; Kawano, T. Molluscicidal and ovicidal activities of plant extracts of the Piperaceae on Biomphalaria glabrata (Say, 1818). J. Helminthol. 2011, 85, 66–72. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, M.; Chiang, M.; Shin, P.; Cheung, S. Characterization of subpopulations and immune-related parameters of hemocytes in the green-lipped mussel Perna viridis. Fish Shellfish Immunol. 2012, 32, 381–390. [Google Scholar] [CrossRef]
- Mitta, G.; Vandenbulcke, F.; Roch, P. Original involvement of antimicrobial peptides in mussel innate immunity. FEBS Lett. 2000, 486, 185–190. [Google Scholar] [CrossRef] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Furunaka, H.; Miyata, T.; Tokunaga, F.; Muta, T.; Iwanaga, S.; Niwa, M.; Takao, T.; Shimonishi, Y. Tachyplesin, a class of antimicrobial peptide from the hemocytes of the horseshoe crab (Tachypleus tridentatus). Isolation and chemical structure. J. Biol. Chem. 1988, 263, 16709–16713. [Google Scholar] [CrossRef] [PubMed]
- Olicard, C.; Renault, T.; Torhy, C.; Benmansour, A.; Bourgougnon, N. Putative antiviral activity in hemolymph from adult Pacific oysters, Crassostrea gigas. Antivir. Res. 2005, 66, 147–152. [Google Scholar] [CrossRef]
- Gueguen, Y.; Herpin, A.; Aumelas, A.; Garnier, J.; Fievet, J.; Escoubas, J.M.; Bachère, E. Characterization of a defensin from the oyster Crassostrea gigas Recombinant production, folding, solution structure, antimicrobial activities, and gene expression. J. Biol. Chem. 2006, 281, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Roch, P.; Yang, Y.; Toubiana, M.; Aumelas, A. NMR structure of mussel mytilin, and antiviral–antibacterial activities of derived synthetic peptides. Dev. Comp. Immunol. 2008, 32, 227–238. [Google Scholar] [CrossRef] [Green Version]
- Decker, H.; Hellmann, N.; Jaenicke, E.; Lieb, B.; Meissner, U.; Markl, J. Minireview: Recent progress in hemocyanin research. Integr. Comp. Biol. 2007, 47, 631–644. [Google Scholar] [CrossRef] [Green Version]
- García-Carrenño, F.L.; Cota, K.; Navarrete del Toro, M.A. Phenoloxidase activity of hemocyanin in whiteleg shrimp Penaeus vannamei: Conversion, characterization of catalytic properties, and role in postmortem melanosis. J. Agric. Food Chem. 2008, 56, 6454–6459. [Google Scholar] [CrossRef]
- Decker, H.; Ryan, M.; Jaenicke, E.; Terwilliger, N. SDS-induced Phenoloxidase Activity of Hemocyanins from Limulus pol-yphemus, Eurypelma californicum, and Cancer magister. J. Biol. Chem. 2001, 276, 17796–17799. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Hirata, T.; Nagai, K.; Sakaguchi, M. Hemocyanin a Most Likely Inducer of Black Spots in Kuruma Prawn Penaeus japonicus During Storage. J. Food Sci. 2001, 66, 1130–1136. [Google Scholar] [CrossRef]
- Pless, D.D.; Aguilar, M.B.; Falcón, A.; Lozano-Alvarez, E.; de la Cotera, E.P.H. Latent phenoloxidase activity and N-terminal amino acid sequence of hemocyanin from Bathynomus giganteus, a primitive crustacean. Arch. Biochem. Biophys. 2003, 409, 402–410. [Google Scholar] [CrossRef]
- Zanjani, N.T.; Sairi, F.; Marshall, G.; Saksena, M.M.; Valtchev, P.; Gomes, V.G.; Dehghani, F. Formulation of abalone he-mocyanin with high antiviral activity and stability. Eur. J. Pharm. Sci. 2014, 53, 77–85. [Google Scholar] [CrossRef]
- Dolashka-Angelova, P.; Schwarz, H.; Dolashki, A.; Stevanovic, S.; Fecker, M.; Saeed, M.; Voelter, W. Oligomeric stability of Rapana venosa hemocyanin (RvH) and its structural subunits. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2003, 1646, 77–85. [Google Scholar] [CrossRef]
- Dolashka, P.; Zal, F.; Dolashki, A.; Molin, L.; Traldi, P.; Salvato, B. ESI-MS and MALLS analysis of quaternary structure of molluscan hemocyanins. J. Mass Spectrom. 2012, 47, 940–947. [Google Scholar] [CrossRef]
- Dolashka, P.; Moshtanska, V.; Borisova, V.; Dolashki, A.; Stevanovic, S.; Dimanov, T.; Voelter, W. Antimicrobial proline-rich peptides from the hemolymph of marine snail Rapana venosa. Peptides 2011, 32, 1477–1483. [Google Scholar] [CrossRef] [PubMed]
- Dolashka, P.; Nesterova, N.; Zagorodnya, S.; Dolashki, A.; Baranova, G.; Golovan, A.; Voelter, W. Antiviral activity of he-mocyanin Rapana venosa and its isoforms against Epstein-Barr virus. Glob. J. Pharmacol. 2014, 8, 206–212. [Google Scholar]
- Dang, V.T.; Benkendorff, K.; Speck, P. In vitro antiviral activity against herpes simplex virus in the abalone Haliotis laevigata. J. Gen. Virol. 2011, 92, 627–637. [Google Scholar] [CrossRef]
- Dang, V.T.; Benkendorff, K.; Corbeil, S.; Williams, L.M.; Hoad, J.; Crane MS, J.; Speck, P. Immunological changes in response to herpesvirus infection in abalone Haliotis laevigata and Haliotis rubra hybrids. Fish Shellfish Immunol. 2013, 34, 688–691. [Google Scholar] [CrossRef]
- Dang, V.T.; Speck, P.; Doroudi, M.; Smith, B.; Benkendorff, K. Variation in the antiviral and antibacterial activity of abalone Haliotis laevigata, H. rubra and their hybrid in South Australia. Aquaculture 2011, 315, 242–249. [Google Scholar] [CrossRef]
- Dolashka, P.; Dolashki, A.; Voelter, W.; Van Beeumen, J.; Stevanovic, S. Antimicrobial activity of peptides from the hemo-lymph of Helix lucorum snails. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 1061–1071. [Google Scholar]
- Sivakamavalli, J.; Nirosha, R.; Vaseeharan, B. Purification and Characterization of a Cysteine-Rich 14-kDa Antibacterial Peptide from the Granular Hemocytes of Mangrove Crab Episesarma tetragonum and Its Antibiofilm Activity. Appl. Biochem. Biotechnol. 2015, 176, 1084–1101. [Google Scholar] [CrossRef] [PubMed]
- Sivakamavalli, J.; Park, K.; Kwak, I.S.; Vaseeharan, B. Purification and partial characterization of carbohy-drate-recognition protein C-type lectin from Hemifusus pugilinus. Carbohydr. Res. 2021, 499, 108224. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeyachandran, S.; Chellapandian, H.; Park, K.; Kwak, I.-S. Exploring the Antimicrobial Potential and Biofilm Inhibitory Properties of Hemocyanin from Hemifusus pugilinus (Born, 1778). Int. J. Mol. Sci. 2023, 24, 11494. https://doi.org/10.3390/ijms241411494
Jeyachandran S, Chellapandian H, Park K, Kwak I-S. Exploring the Antimicrobial Potential and Biofilm Inhibitory Properties of Hemocyanin from Hemifusus pugilinus (Born, 1778). International Journal of Molecular Sciences. 2023; 24(14):11494. https://doi.org/10.3390/ijms241411494
Chicago/Turabian StyleJeyachandran, Sivakamavalli, Hethesh Chellapandian, Kiyun Park, and Ihn-Sil Kwak. 2023. "Exploring the Antimicrobial Potential and Biofilm Inhibitory Properties of Hemocyanin from Hemifusus pugilinus (Born, 1778)" International Journal of Molecular Sciences 24, no. 14: 11494. https://doi.org/10.3390/ijms241411494