
Molecular Mechanisms of Shigella Pathogenesis; Recent Advances


1
Werner Siemens Chair of Synthetic Biotechnology, Department of Chemistry, Technical University of Munich (TUM), Lichtenberg Str. 4, 85748 Garching bei München, Germany
2
Institute for Life Technologies, University of Applied Sciences Western Switzerland Valais-Wallis, 1950 Sion, Switzerland
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(3), 2448; https://doi.org/10.3390/ijms24032448
Submission received: 30 December 2022
/
Revised: 21 January 2023
/
Accepted: 23 January 2023
/
Published: 26 January 2023
(This article belongs to the Section Molecular Microbiology)
Abstract
:Shigella species are the main cause of bacillary diarrhoea or shigellosis in humans. These organisms are the inhabitants of the human intestinal tract; however, they are one of the main concerns in public health in both developed and developing countries. In this study, we reviewed and summarised the previous studies and recent advances in molecular mechanisms of pathogenesis of Shigella Dysenteriae and non-Dysenteriae species. Regarding the molecular mechanisms of pathogenesis and the presence of virulence factor encoding genes in Shigella strains, species of this bacteria are categorised into Dysenteriae and non-Dysenteriae clinical groups. Shigella species uses attachment, invasion, intracellular motility, toxin secretion and host cell interruption mechanisms, causing mild diarrhoea, haemorrhagic colitis and haemolytic uremic syndrome diseases in humans through the expression of effector delivery systems, protein effectors, toxins, host cell immune system evasion and iron uptake genes. The investigation of these genes and molecular mechanisms can help us to develop and design new methods to detect and differentiate these organisms in food and clinical samples and determine appropriate strategies to prevent and treat the intestinal and extraintestinal infections caused by these enteric pathogens.
1. Introduction
Shigella are facultative anaerobe, non-spore-forming, rod-shaped, non-motile and Gram-negative bacteria belonging to the Enterobacteriaceae family [1]. Genotypically and phenotypically, these bacteria are closely related to Escherichia coli strains; however, Shigella strains are less active in carbohydrate utilisation, by which they can be differentiated and identified biochemically from the Escherichia coli strains. Shigella strains are among the oldest human (Homo sapiens) specific pathogens, which evolved 170,000 years ago [2]. In 1888, Chantemesse and Widel isolated a bacterium from faecal samples collected from human patients with acute dysentery. Ten years later, Kiyoshi Shiga isolated and identified Shigella strains as humans’ main causative agents of bacillary dysentery [3]. Then, twenty years later, different serological groups of Shigella strains were distinguished and differentiated [1]. Using novel genetic methods, more than 46 serotypes of this bacteria have been identified and classified. Four species subgroups of Shigella have been identified, including S. dysenteriae (subgroup A; 15 serotypes), S. flexneri (subgroup B; 6 serotypes), S. boydii (subgroup C; 20 serotypes) and S. sonnei (subgroup D; 1 serotype) [4]. These four species are differentiated based on differences in biochemical reactions and their O antigen lipopolysaccharide. Four species of Shigella are widely transmitted among humans and foods via the faecal–oral route [5].
Shigella strains have been classified as waterborne and foodborne pathogens [6]. The natural reservoirs and hosts for these bacteria are humans and primates. Shigella strains are found in the faeces of infected people (or primate animals) and it can be transmitted via a vehicle such as contaminated food and water to cause any infection and diseases in humans [7]. Several outbreaks have occurred via the consumption of contaminated food and water sources. Shigella foodborne outbreaks are usually common when consuming foods subjected to processing by hand, exposed to a limited thermal treatment or raw foods [8]. Shigella strains have commonly been isolated from contaminated ground beef, oysters, potato salads, bean dip, raw vegetables and fish [9]. Person-to-person transmission of Shigella strains also can occur through the faecal–oral route in foodborne and waterborne outbreaks [10]. Clinical symptoms of gastrointestinal infections caused by Shigella species vary from mild diarrhoea for a few days by S. sonnei to dysentery (bloody stools with small and painful mucoid), vomiting and nausea by S. dysenteriae. The infective dose of Shigella species ranges from 10 cells of S. dysenteriae to 500 cells of S. sonnei in humans [11,12].
Shigellosis, a global health problem in developed and developing countries, is a contagious infectious disease caused by different species of Shigella via the consumption of contaminated food and water [13]. However, the incidence of shigellosis is also significantly high among homosexual males via person-to-person transmission [14]. It is caused by invasion of the colon, ileum and rectum epithelial cells. Shigella infections occur in people of all ages; however, it has been reported more in sensitive populations. More than one million deaths annually are caused by diarrhoeal diseases worldwide [15]. However, approximately 164,000 deaths are attributed to shigellosis annually, particularly in children under 5 years old. It is also estimated that more than 125 million diarrhoeal episodes are annually caused by Shigella species [11,13]. In addition to public health concerns generated by this disease, severe economic losses are caused by shigellosis worldwide. Investigation of the pathogenicity mechanisms of Shigella species helps us to determine more successful, preventive, specific and effective strategies against shigellosis [16,17]. This article will review and provide new insights into the novel aspects of molecular mechanisms of Shigella pathogenesis for intestinal and extraintestinal diseases in humans. We summarised these mechanisms and listed all virulence factor encoding genes associated with different pathogenesis mechanisms of Dysenteriae and non-Dysenteriae species of Shigella.
2. Bacterial Pathogenesis
A wide spectrum of microbial strains, including several bacteria, fungi, parasites and viruses, have been identified, classified and characterised as pathogens causing different types of chronic and acute infectious diseases in humans. Bacterial strains are the main causative agents of mortality and morbidity among the infected persons and hospitalised patients worldwide [18,19]. According to recent documented reports, more than 90% of infections in hospitalised patients have been caused by bacterial agents; consequently, it is estimated that a considerable number of bacterial infections occur in the general population [20]. Most microbial pathogens in humans have food product origin and are considered foodborne pathogens. These organisms contribute to foodborne diseases in humans [21]. According to the World Health Organization, foodborne illnesses are toxic or infectious diseases caused by consuming contaminated food or water [22]. Foodborne bacterial pathogens cause human intestinal and extraintestinal diseases through different pathogenicity mechanisms [23].
2.1. Bacterial Pathogenicity Definition
Bacterial pathogenicity is defined as the ability of bacterial strains to induce infection, release toxins, produce virulence and cause diseases and mortality in the host (humans). All bacterial pathogens have pathogenicity activity [24]. Specific mechanisms mediate this ability between bacterial pathogens and human host cells. Pathogenesis of a bacterial strain is defined as the mechanisms and series of events by which the bacterial cells induce and develop a morbid state or disease in the host (humans) [25]. The degree of virulence of a bacterial strain is the measure of the pathogenicity of a bacterium. Susceptibility to a bacterial infection depends on the bacteria’s virulence degree and the host’s physiologic conditions (humans) [26]. There is a complex binary interaction between the bacterial pathogen and the host throughout the development of the disease [27]. Consequently, pathogenicity mechanisms refer to the mechanisms by which the disease is developed in the host and to the pathogenesis mechanisms mediated by the bacterial strains interacting with the host. Considering the host-pathogen interactions, various virulence factors in pathogenic bacterial cells mediate the pathogenesis mechanisms in interaction with the host cells [28,29].
2.2. Virulence Factors in Bacterial Pathogenesis
Understanding the virulence factors is important in studying bacterial pathogenesis in establishing disease in the host [30]. As previously discussed, diseases caused by Enterobacteriaceae family members are mediated via intestinal and extraintestinal pathogenesis mechanisms. These mechanisms are regulated and developed via several mechanism-specific virulence factors of the pathogenic bacteria [31,32]. Virulence factors in bacterial pathogens are divided into 6 main classes including (1) membrane proteins, (2) capsule, (3) secretory proteins, (4) outer membrane proteins, (5) biofilm formation and (6) iron acquisition. The membrane proteins class includes adhesion, invasion, colonisation, and surface virulence factors. Secretory proteins are subclassified into immune response inhibitors, toxins, and transport groups [33,34,35].
Regarding the pathogenesis mechanism of each bacterial pathogen, the expression and release of these virulence factors has been regulated and induced by the bacterial strain [20]. Adhesion factors lead to binding to the host epithelial cells and mucosal layers. Different types of adhesin proteins are expressed and released on bacterial cell surfaces. Adhesion can be mediated by either fimbrial or afimbrial structures [36,37,38]. Protein secretion pathways also play a key role in the pathogenesis of Gram-negative bacterial pathogens, and there are seven distinct types of secretion systems in these bacteria. Host cell invasion, intracellular multiplication and cell-to-cell spread are prominent virulence factors in the pathogenicity mechanisms of invasive bacterial pathogens such as Shigella species [16,17]. Several pathogenic bacteria release some specific polypeptides with toxic effects on host cell processes and structures. These toxins allow the pathogens to cause damage and grow within the host cells [39]. Toxins are secreted via lysis of the bacterial cell (endotoxin) or through a specific secretion system (exotoxin). Bacterial pathogens can also evade the host immune system by employing specific mechanisms [40,41]. Regarding the mechanism of pathogenesis, the type of virulence factors and the symptoms of the disease, Shigella species are categorised into two distinct pathogenic groups: Dysenteriae and non-Dysenteriae species [16,42]. In the following sections of this paper, we will investigate the pathogenic molecular mechanisms and virulence factors of these two pathogenic groups separately.
3. Pathogenic Molecular Mechanisms of Non-Dysenteriae Shigella Species Pathogenesis
3.1. Non-Dysenteriae Shigella Species
The epidemiological studies of shigellosis showed that the species of S. sonnei and S. flexneri are the main causes of this disease [43]. S. dysenteriae type 1 causes more severe and fatal disease in human as an extraintestinal pathogen, which has been investigated in the next section. Regarding the epidemiological studies, S. sonnei and S. flexneri are more prevalent for endemic shigellosis in high-income and low-income (developed and developing) countries, respectively [11,12]. All species of Shigella except S. dysenteriae type 1 have been considered as non-Dysenteriae species of Shigella, causing intestinal shigellosis [16]. Regarding public health concerns, three major characteristics have been identified for shigellosis: first, as a paediatric disease with more than 60% of the cases between the ages of 1 to 5 years old; second, as a general diarrhoeal disease with more than 150 million cases each year; and third, the deadly disease, mostly in young children and infants [11,12,13]. Non-Dysenteriae Shigella species cause the first and second groups. Non-Dysenteriae species of Shigella are invasive bacteria and cause mild diarrhoea in humans via intracellular multiplication and gastrointestinal epithelial cell manipulation mechanisms [6,16,17].
3.2. Molecular Mechanisms of Non-Dysenteriae Shigella Species Pathogenesis
S. flexneri and S. sonnei (non-Shiga-toxin producing strains) are mainly responsible for non-Dysenteriae disease (mild diarrhoea) caused by Shigella strains [5,8,16]. The pathogenesis of Shigella possesses unique and multiple well-established pathogenic mechanisms mediated by seven distinct steps, including attachment to the epithelial host cell, entry to the cell, host cell autophagy evasion, vacuole formation, vacuole rapture, intracellular life and immune response [44,45]. Shigella employs these seven steps, and releases different effector proteins to damage, invade and suppress the immune system of the host gastrointestinal epithelial cells, contributing, eventually, to mild diarrhoeal symptoms in the patient [3,46]. Infection by Shigella initiates in the colon by reaching the underlying submucosa through transcytosis and passing through microfold cells. As the first barrier, Mucin glycol-protein is remodelled and glycosylated by Shigella during the attachment process [43,47,48]. Gel-forming mucin is induced by proinflammatory cytokines such as tumour necrosis factor-α (TNFα) and interferon-gamma (IFN-γ). Mu5AC factor, secreted by Shigella strains, stimulates specific gel-forming mucin and induces the accumulation of gel-like structures on the host cell surface, leading to bacterial invasion. This mechanism is dependent on the presence of the type III secretion system (T3SS) in the bacterial cell [3,12,46].
The virulence and pathogenesis of Shigella strains require T3SS, the most important pathogenicity mechanism of Shigella species. T3SS is a needle-like molecular structure in the bacterial cell wall encoded on the Mxi-spa locus, located on a 220 kb virulence plasmid (pWR100) in Shigella strains [49]. All virulence factor encoded genes providing different mechanisms of Dysenteriae and non-Dysenteriae Shigella species are described in Table 1. It provides a direct channel between the host cytoplasm and the bacterial cell to inject several bacterial protein effectors through this syringe-like structure. Mxi-spa locus encodes Mxi (A-N) and spa genes, expressing the components of T3SS structure [50,51]. Shigella strains use T3SS, mainly, to induce its uptake into the host epithelial cells, escape from the vacuole, mediate cell-to-cell spreading and manipulate the host cell processes. These mechanisms are mediated through different protein effectors [46,49]. After the initial attachment by accumulating a gel-like structure on the surface of the host cells, complementary attachment is mediated by a specific outer membrane protein (IcsA) encoded by VirG gene. The binding site of the IcsA protein on the host cell is Neural Wiskott-Aldrich syndrome protein (N-WASP), a WASP family and CDC-42 dependent compound mediating the actin nucleation in the host epithelial cell cytoplasm through the Arp2/3 complex [25,44,52]. IpaB and IpaC are involved in pore formation in host cell. IpaD, as a hydrophilic protein, binds with T3SS tips and blocks the needle pore. Following the sensing of cholesterol and sphingomyelin on the host cell membrane, IpaD stimulates IpaB in the T3SS tip to mediate the invasion of the host cell [44,51]. Key protein effectors such as VirA, IPgD, IpaA, IpaB and IpaC secreted through T3SS into the host cell mediate epithelial cell signalling, cellular uptake, cytoskeletal rearrangements and lysis of the endocytic vacuole [45,53,54]. VirA, IPgD and IpaA are involved in microtubule and actin destabilisation to develop the invasion into the phagosome and establish the endocytic vacuole [44,55]. IpaH, IpaB, IpaC and IpaD are the protein effectors responsible for phagosome escape in the host cell cytoplasm [44,52,56].
In addition to T3SS, type VI secretion system (T6SS) is also employed by S. sonnei strains and tss (A-K) genes encode this mechanism. The type II secretion system (T2SS), encoded by gsp (A-O) genes, is also used by S. boydii Sb227 and S. dysenteriae Sd197 strains [44,57,58].
The intracellular dissemination of Shigella species through host epithelial cells is mediated via the manipulation of cell membrane and cytoskeleton. Some T3SS effector proteins, such as IpgB1 and IpgB2, are responsible for these mechanisms through the regulation of Rho family GTPases in host cells [44,45]. These effectors are guanine nucleotide exchange factors and are defined and grouped together by the WxxxE motif (tryptophan-xxx-glutamic acid). These effectors mimic the low molecular weight GTPases and activate the signalling cascades in the host cells. They are mainly encoded in T3SS gene domains [50,52]. However, WxxxE sequences have also been detected in TLR domains of both Shigella species and human host cells. WxxxE family proteins such as IpgB2 interreact with human RhoA via inducing conformational changes. The activation of the Rho protein family induces host cell contraction via cell membrane ruffling through the activation of GTP-binding proteins such as Rac1 and Cdc42 [44,46,51]. It has also been shown that WxxxE effectors can bind to the host engulfment and modulate the host immune system. IpgB1 is also involved in the subversion of actin cytoskeleton dynamics. IpgB2 activates the nuclear factors through the activation of Rho-associated kinases. IpgB1 and IpgB2 also have a key role in the tight junctions by the pathways of unidentified signalling because of their Rho and Rac mimicking abilities [44,45,50]. After the cell entry, pyroptosis is mediated through interleukins and activated by nod- and Toll-like receptors contributing to the activation of inflammatory caspases. Pyroptosis causes membrane rupture in the host cells, contributing to inflammatory response and ion venting. However, Shigella bacterial cells can prevent host cell death and mediate intracellular life by IpaH family effectors such as IpaH9.8, IpaH7.8, IpaH1.4 and IpaH4.5 [44,46]. It should be noted that different proteins in host cells have key roles in Shigella pathogenesis, including N-WASP, ARP2/3, NLR, TLR, Integrin, Talin, Actin, Rho family proteins, TNF-α, IFN-γ, Mu5AC, NF-κB and interleukins [44].
After entering into the host epithelial cells, Shigella strains promote their survival by using different protein effectors and subverting the host cell processes. IcsA protein (VirG) triggers actin nucleation and induces the actin microfilament growth at the one pole of the organism, mediating the movement of the bacterial cell through the host cell cytoplasm [59,60]. The VirA effector also destabilises the microtubules and enables the bacteria to control the actin polymerization leading to efficient spread of the bacterial cells throughout the host cell. This mechanism provides the intracellular motility and dissemination characteristics of Shigella bacterial cells [45,47]. Shigella strains employ three main mechanisms to prevent host cell death and provide the replication niche for bacterial infection. IpaB effector has been shown to induce host cell cycle arrest via targeting Mitotic Arrest Deficient 2 Like 2 (MAD2L2, an anaphase inhibitor) protein to prevent the host cell turnover. Preventing the host cell detachment, OspE protein effector interacts with the integrin-linked kinase. IpgD also prevents apoptosis in the host epithelial cell via the activation of Akt proteins and stimulation of phosphoinositide-3-kinase [44,45,47]. Shigella bacterial cells must evade the innate immune responses to persist inside the host cells, and Osp genes mediate this mechanism. OspF induces the dephosphorylation of the mitogen-activated protein kinases required for the regulation of nuclear factor kappa B (NF-κB) gene expression in the host cell. OspG and OspI effectors also bind the E2 protein and inhibit the activation of NF-κB. OspF and OspB effectors reduce the interleukin-8 (IL-8) secretion level in the host cells. OspD2 inhibits necrosis, and OspC3 mediates the inhibition of pyroptosis in the infected host cells [44,61,62]. As a non-Osp effector, ipaH can interact with the splicing factors and inhibits the expression of inflammatory cytokines. The IpaH effectors family also are involved in other host cellular processes, including protein degradation, cell cycle and endocytosis [63,64]. Shigella lipopolysaccharide (LPS), composed of core polysaccharides, O-antigen and lipid A and encoded by gtr genes, also mediates intracellular spread and resistance to host cell defence [65,66]. The expression of iron uptake genes is critical for intracellular growth, and this protective mechanism is mediated by aerobactin proteins encoded by iuc (A-D) genes [44,52,67].
Autophagy is a protective and defence mechanism of the host cells which enables them to survive during infections via the degradation of macromolecules to recycle the damaged organelles and the energy in the cell cytoplasm [68]. This defence mechanism is mediated by autophagy related proteins (ARP). The IcsB effector can be recognised as ARP, masks the region that is unmasked by ARP and inhibits the autophagy mechanism of the host cell [69,70]. Non-Dysenteriae Shigella species inside the host cells secrete different toxins and toxic compounds. Two main types of exotoxins, including Shigella enterotoxin 1 (ShET1) and Shigella enterotoxin 2 (ShET2), are secreted by non-Dysenteriae Shigella species. ShET1 is chromosomally encoded by set1A and set1B genes. This exotoxin is an iron-dependent toxin (55 KDa), mainly released by S. flexneri strains, and predominantly accounts for the early phase of diarrhoea in shigellosis. ShET2 is a plasmid-borne toxin (inv plasmid) encoded by the senB gene and also responsible for the early phase of diarrhoea [12,71,72]. Other toxins have also been described for the Shigella species, including protein involved in intestinal colonisation and Shigella IgA-like protease homology toxins encoded by pic and sigA genes, respectively. Pic is a protease non-cytotoxic toxin, secreted via type V secretion system (T5SS), mainly released by S. flexneri strains and belonging to the serine protease autotransporters of the Enterobacteriaceae family (SPATEs). SigA is a protease cytotoxin, also secreted through T5SS and belonging to the SPATEs. This toxin is a serine protease involved in the accumulation of intestinal fluid during shigellosis [43,44,73].
Since Shigella species are intracellular pathogens, there are several host proteins and engulfment that affect the pathogenesis mechanisms of these bacterial pathogens. The invasion of host epithelial cells by Shigella species requires a comprehensive adaptation of the bacterial pathogen to the new environment through accessing different metabolic cofactors and nutrients mediating intracellular growth. Shigella also employs some specific structures in host cells, such as N-WASP, Arp2/3 complex and actin molecules during their pathogenesis mechanisms [58,59]. Several in vivo studies revealed that human host cells elaborate specific mechanisms, such as immune response signalling and autophagy against Shigella invasion, intracellular growth and dissemination through the host cells. In the initial steps of the epithelial host cell infection by Shigella, host cells release IL-8 and pro-inflammatory cytokines, including IL-1β and IL-18 by macrophages, induce inflammation and activate the immune cells, including neutrophils and natural killer cells, against Shigella invasion [50,53,54]. Host cells mediate the autophagy mechanism against pathogenic bacterial intracellular dissemination. Autophagy in human epithelial cells is mediated through recruiting TNF receptor-associated factor 6, ubiquitin, LC3, NDP52, ATG5, P62, calpains, caspase-8 and Beclin-1 factors during Shigella infection and intoxication (by Shiga-toxin) [44,46,49,50,54]. Shigella species elaborate specific mechanisms to suppress these antimicrobial strategies throughout intracellular growth, dissemination and activities which are also comprehensively described in this review paper.
4. Pathogenic Molecular Mechanisms of Dysenteriae Shigella Species Pathogenesis
4.1. Dysenteriae Shigella Species Diseases
S. dysenteriae is one of the four serological groups of Shigella bacteria. Type 1 of S. dysenteriae is the only bacterial strain of Shigella species considered as the clinical group of Dysenteriae Shigella species and differentiated from other species due to the production of Shiga toxins (Stxs); however, some studies recently reported the production of Stxs by S. sonnei isolated from clinical samples [8,74]. S. dysenteriae type 1 is also considered one of the leading causes of bacillary dysentery or shigellosis and is transmitted to humans through the ingestion of contaminated water or food. It is also transmitted person-to-person via the faecal–oral route [44,75]. S. dysenteriae infection and intoxication are primarily more prevalent in lower-middle- and low-income countries, especially among children, contributing to long-term diarrhoea and malnutrition due to intestinal protein loss [76,77]. S. dysenteriae serotype 1 is also known for producing explosive pandemics with high fatality rates, including pandemics which have occurred in the past fifty years in central America, south Asia, and central and east areas of Africa [12,13,78]. Another public health concern regarding S. dysenteriae type 1 is that this organism is resistant to all the currently used antibiotics [5]. The incubation period of shigellosis caused by Shigella non-dysenteriae species ranges from 1 to 4 days, and typically for S. dysenteriae type 1 it is up to 8 days. In the first phase, S. dysenteriae type 1 causes watery diarrhoea, mediated by the secretion of an enterotoxin, for 1 or 2 days. Then, the organism invades the large intestine epithelial cells and produces cramps, fever, tenesmus and bloody diarrhoea. At this phase, S. dysenteriae type 1 is isolated from the stool culture of more than 50% of the cases [79,80,81]. As a Shiga-toxin-producing strain, S. dysenteriae type 1 mainly causes haemolytic uraemic syndrome (HUS) in patients. Shiga toxin secreted by S. dysenteriae type 1 accounts for more than 2,800,000 acute diseases, leading to approximately 3900 HUS cases annually. Around 2–7% of S. dysenteriae infections cause HUS in humans [82].
4.2. Molecular Mechanisms of Dysenteriae Shigella Species Pathogenesis
S. dysenteriae is a normal microflora of the human gastrointestinal system; however, it is considered one of the most important public health concerns, and a major cause of bloody diarrhoea and HUS (S. dysenteriae type 1) in humans [74]. The serotype 1 of this organism is closely related to Escherichia coli serotype O157: H7, regarding the fact that both of these organisms can secret Stxs [83,84]. Foodborne outbreaks caused by S. dysenteriae type 1 are highly associated with the consumption of undercooked processed foods and vegetable salads [74,85]. HUS is the only disease caused by S. dysenteriae type 1 infection, and the typical symptoms of this disease are renal failure, thrombocytopenia and microangiopathic haemolytic anaemia [58]. The fatality rate of this disease is 35%. S. dysenteriae type 1 adheres to the human colonic mucin and attaches to the epithelial cells by using the same mechanisms and virulence factor genes employed by other species [12,13]. After attachment and adherence, Stxs are secreted by this organism [74]. In addition to the presence of Stxs encoding genes, there are some minor differences between the virulence factors of S. dysenteriae type 1 and other species of Shigella bacteria. S. dysenteriae type 1 uses type 2 secretion system (T2SS) as an effector delivery system to inject protein effectors into the host cell. This needle-like structure is encoded and expressed by gsp (A-O) genes in S. dysenteriae type 1 [86,87]. S. dysenteriae type 1 also uses a specific heme uptake system as a protective virulence factor encoded by Shu genes. This mechanism mediates the direct binding of heme-containing proteins and the secretion of hemophores to provide iron for the organism during the pathogenesis and survival processes. This mechanism also protects the DNA of S. dysenteriae type 1 from heme-mediated oxidative damage [44,67].
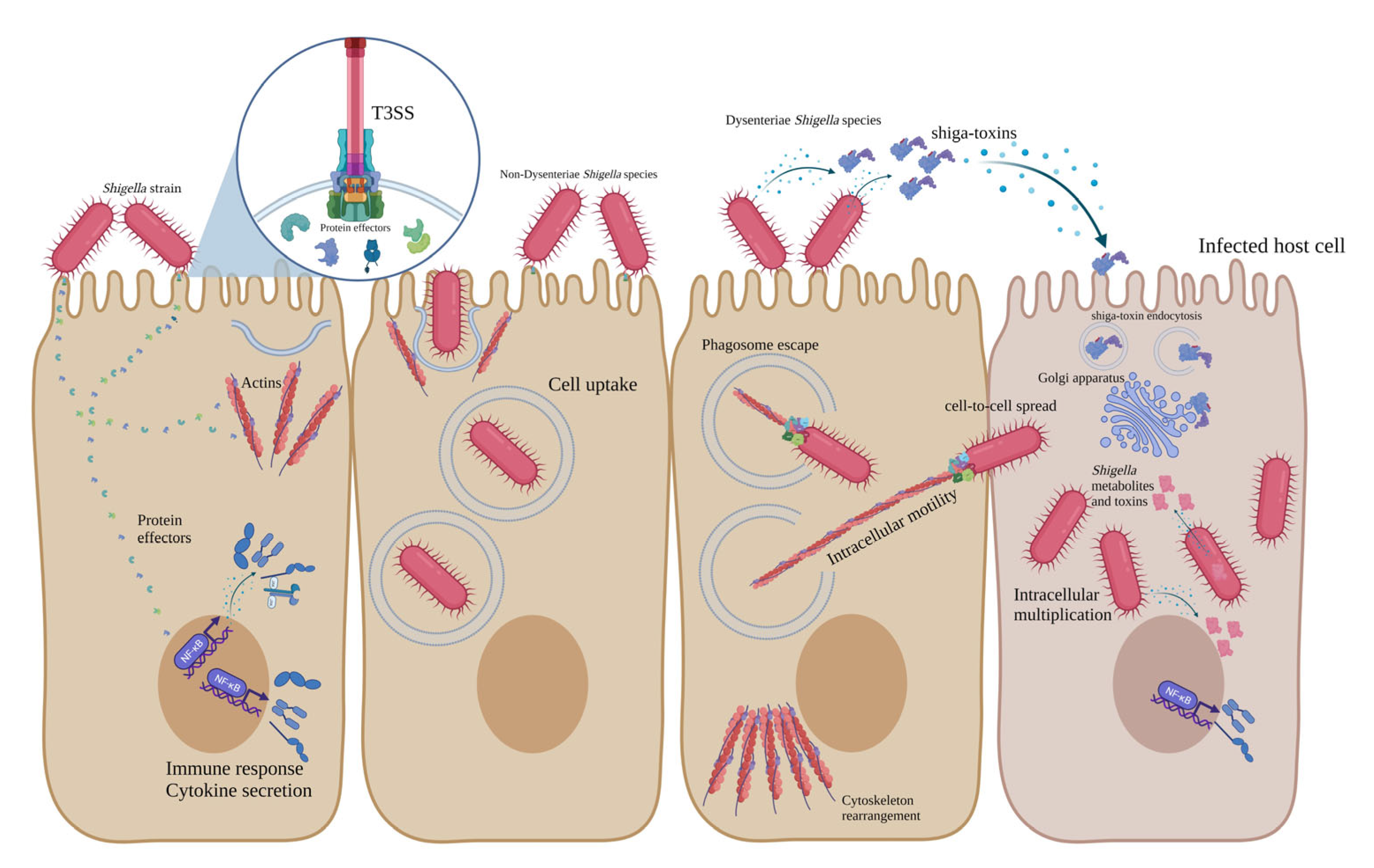
Stxs, also known as Shiga-like toxins, verocytotoxins and verotoxins, are lethal toxins first isolated from S. dysenteriae in 1975 [88,89]. In addition to S. dysenteriae type 1, some other bacterial pathogens secret these toxins, such as some E. coli serogroups (and more than 200 serotypes), including O157, O26, O91, O103, O104, O145 and S. sonnei, which has recently been reported [90,91]. Two types of Stxs have been identified, including Stx1 and Stx2. Additionally, different variants of these toxins have been characterised, such as Stx1 (c-d) and Stx2 (c-k). Stx2 is 56% identical to Stx1, considering the amino acid sequence. Stx members are AB5 proteins composed of the A subunit (32 KDa), which is noncovalently bound to five B subunits (each one is 7.7 KDa) [90,92,93]. A and B subunits of Stxs are encoded by chromosomal stxA and stxB genes. The pentameric structure of Stx (B5) binds to globotriaosylceramide (GB3), its receptor on the surface of the host Intestinal and kidney epithelial cells. After binding, Stx enters the host cell through the endocytosis mechanism and is trafficked into the Golgi apparatus, where the A subunit is activated [94,95,96,97]. A subunit is the active component of the Stx and is composed of two disulfide-bonded fragments (A1-A2). Subunit A presents proteolytic activity, deactivates 28 s rRNA and inhibits peptide synthesis in host cells, leading to apoptosis, necrosis and cell death [11,12,89,98,99]. Stxs are responsible for HUS, haemorrhagic colitis and bloody diarrhoea in the human host, since they target both intestinal and kidney epithelial cells [13,44,74]. All pathogenesis mechanisms of Dysenteriae and non-Dysenteriae Shigella species are summarised in Figure 1.
5. Limitations and Perspectives
Shigella is one of the most important bacterial infectious challenges in food safety and public health, causing diarrhoea and extra-intestinal disorders including hemolytic colitis and haemorrhagic uremic syndrome in humans. So far, several mechanisms of pathogenesis and virulence factors of Shigella species have been defined and specified. However, various types of new protein effectors and the main roles of different effectors have not been known and identified clearly [44,51,58]. Bacterial Quorum sensing systems (QS) have recently been known as novel virulence factors specified for Enterobacteriaceae family pathogens such as Salmonella, Escherichia and Shigella species. QS is a bacterial cell-to-cell communication mechanism contributing to the adaptation of the organism to the new environment. Through this pathogenesis mechanism, bacterial strains release specific QS molecules as signals which are received and detected by other strains and allow them to respond to challenging environmental conditions [100]. These mechanisms have also been characterised for Shigella species, especially S. flexneri; however, more studies need to be implemented to deeply understand the key role of QS molecules and mechanisms in Shigella pathogenesis [101]. On the other hand, using QS inhibitors has widely been developed as a practical, affordable and efficient anti-bacterial strategy in food industries and clinical treatments [102].
6. Conclusions
Shigella species are one of the main concerns in public health, and they cause mild diarrhoea, hemolytic colitis and haemorrhagic uremic syndrome in humans. Regarding the severity and type of the caused diseases and the presence of different virulence factor encoding genes, species of Shigella are categorised into two groups: Dysenteriae and non-Dysenteriae species. The pathogenesis of non-Dysenteriae Shigella species includes attachment, invasion, vacuole escape, intracellular motility, toxin secretion and host cell interruption. The pathogenesis mechanisms of non-Dysenteriae and Dysenteriae species of Shigella are mediated by the expression of effector delivery systems (T3SS and T6SS), different protein effectors, toxins, iron uptake genes and the secretion of the Shiga-toxin (only for S. dysenteriae type 1). S. dysenteriae type 1 uses T2SS as the effector delivery system. Shiga-toxins contribute to HUS and bloody diarrhoea in humans, with a high rate of fatality. Regarding the specific genes in each group, invasion and Shiga-toxin encoding genes can be considered as the target genes to detect and differentiate Dysenteriae and non-Dysenteriae species of Shigella in food and clinical samples. Additionally, considering the different pathogenicity molecular mechanisms and virulence factor encoding genes in each clinical group, distinct and relative therapeutic strategies can be used to prevent and treat the intestinal and extraintestinal infections caused by these bacterial enteric pathogens.
Author Contributions
Conceptualisation, B.P.; W.M.B. and T.B.B.; investigation, B.P.; writing—original draft preparation, B.P.; writing—review and editing, W.M.B. and T.B.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
We confirm that all data supporting the findings of this study are available within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lampel, K.A.; Formal, S.B.; Maurelli, A.T. A brief history of Shigella. EcoSal Plus 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yang, J.; Jin, Q. The molecular evolutionary history of Shigella spp. and enteroinvasive Escherichia coli. Infect. Genet. Evol. 2009, 9, 147–152. [Google Scholar] [CrossRef]
- Anderson, M.; Sansonetti, P.J.; Marteyn, B.S. Shigella diversity and changing landscape: Insights for the twenty-first century. Front. Cell. Infect. Microbiol. 2016, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthuirulandi Sethuvel, D.; Devanga Ragupathi, N.; Anandan, S.; Veeraraghavan, B. Update on: Shigella new serogroups/serotypes and their antimicrobial resistance. Lett. Appl. Microbiol. 2017, 64, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Puzari, M.; Sharma, M.; Chetia, P. Emergence of antibiotic resistant Shigella species: A matter of concern. J. Infect. Public Health 2018, 11, 451–454. [Google Scholar] [CrossRef]
- Pakbin, B.; Amani, Z.; Allahyari, S.; Mousavi, S.; Mahmoudi, R.; Brück, W.M.; Peymani, A. Genetic diversity and antibiotic resistance of Shigella spp. isolates from food products. Food Sci. Nutr. 2021, 9, 6362–6371. [Google Scholar] [CrossRef]
- Moxley, R.A. Enterobacteriaceae: Shigella. In Veterinary Microbiology; John Wiley and Sons: Hoboken, NJ, USA, 2022; pp. 100–107. [Google Scholar]
- Pakbin, B.; Didban, A.; Brück, W.M.; Alizadeh, M. Phylogenetic analysis and antibiotic resistance of Shigella sonnei isolates. FEMS Microbiol. Lett. 2022, 369, fnac042. [Google Scholar] [CrossRef]
- Warren, B.; Parish, M.; Schneider, K. Shigella as a foodborne pathogen and current methods for detection in food. Crit. Rev. Food Sci. Nutr. 2006, 46, 551–567. [Google Scholar] [CrossRef]
- Pakbin, B.; Zolghadr, L.; Rafiei, S.; Brück, W.M.; Brück, T.B. FTIR differentiation based on genomic DNA for species identification of Shigella isolates from stool samples. Sci. Rep. 2022, 12, 2780. [Google Scholar] [CrossRef]
- Kotloff, K.L.; Riddle, M.S.; Platts-Mills, J.A.; Pavlinac, P.; Zaidi, A.K. Shigellosis. Lancet 2018, 391, 801–812. [Google Scholar] [CrossRef]
- Bennish, M.L.; Ahmed, S. Shigellosis. In Hunter’s Tropical Medicine and Emerging Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2020; pp. 492–499. [Google Scholar]
- Keusch, G.T. Shigellosis. In Bacterial Infections of Humans; Springer: Berlin/Heidelberg, Germany, 2009; pp. 699–724. [Google Scholar]
- Chen, T.; Leung, R.K.-K.; Zhou, Z.; Liu, R.; Zhang, X.; Zhang, L. Investigation of key interventions for shigellosis outbreak control in China. PLoS ONE 2014, 9, e95006. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.A.; Troeger, C.; Blacker, B.F.; Rao, P.C.; Brown, A.; Atherly, D.E.; Brewer, T.G.; Engmann, C.M.; Houpt, E.R.; Kang, G. Morbidity and mortality due to Shigella and enterotoxigenic Escherichia coli diarrhoea: The Global Burden of Disease Study 1990–2016. Lancet Infect. Dis. 2018, 18, 1229–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakbin, B.; Basti, A.A.; Khanjari, A.; Azimi, L.; Brück, W.M.; Karimi, A. RAPD and ERIC-PCR coupled with HRM for species identification of non-dysenteriae Shigella species; as a potential alternative method. BMC Res. Notes 2021, 14, 345. [Google Scholar] [CrossRef] [PubMed]
- Pakbin, B.; Didban, A.; Monfared, Y.K.; Mahmoudi, R.; Peymani, A.; Modabber, M.R. Antibiotic susceptibility and genetic relatedness of Shigella species isolated from food and human stool samples in Qazvin, Iran. BMC Res. Notes 2021, 14, 144. [Google Scholar] [CrossRef]
- Ashrafi Tamai, I.; Mohammadzadeh, A.; Zahraei Salehi, T.; Mahmoodi, P.; Pakbin, B. Investigation of antimicrobial susceptibility and virulence factor genes in Trueperella pyogenes isolated from clinical mastitis cases of dairy cows. Food Sci. Nutr. 2021, 9, 4529–4538. [Google Scholar] [CrossRef]
- Tamai, I.A.; Mohammadzadeh, A.; Mahmoodi, P.; Pakbin, B.; Salehi, T.Z. Antimicrobial susceptibility, virulence genes and genomic characterization of Trueperella pyogenes isolated from abscesses in dairy cattle. Res. Vet. Sci. 2023, 154, 29–36. [Google Scholar] [CrossRef]
- Tamai, I.A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P.; Pakbin, B. Expression of virulence factor genes in co-infections with Trueperella pyogenes isolates and other bacterial pathogens; an in vivo study. Microb. Pathog. 2022, 164, 105435. [Google Scholar] [CrossRef]
- Pakbin, B.; Mahmoudi, R.; Mousavi, S.; Allahyari, S.; Amani, Z.; Peymani, A.; Qajarbeygi, P.; Hoseinabadi, Z. Genotypic and antimicrobial resistance characterizations of Cronobacter sakazakii isolated from powdered milk infant formula: A comparison between domestic and imported products. Food Sci. Nutr. 2020, 8, 6708–6717. [Google Scholar] [CrossRef]
- Pakbin, B.; Allahyari, S.; Amani, Z.; Brück, W.M.; Mahmoudi, R.; Peymani, A. Prevalence, Phylogroups and Antimicrobial Susceptibility of Escherichia coli Isolates from Food Products. Antibiotics 2021, 10, 1291. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W.M.; Rossen, J.W. Virulence factors of enteric pathogenic Escherichia coli: A review. Int. J. Mol. Sci. 2021, 22, 9922. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W.M.; Brück, T.B.; Allahyari, S.; Ashrafi Tamai, I. A quantitative prevalence of Escherichia coli O157 in different food samples using real-time qPCR method. Food Sci. Nutr. 2022, 11, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.A.; Winkler, M.; Ho, B.T. Bacterial Pathogenesis: A Molecular Approach; John Wiley and Sons: Hoboken, NJ, USA, 2020. [Google Scholar]
- Abebe, E.; Gugsa, G.; Ahmed, M. Review on major foodborne zoonotic bacterial pathogens. J. Trop. Med. 2020, 2020, 4674235. [Google Scholar] [CrossRef] [PubMed]
- Gyles, C.; Prescott, J. Themes in bacterial pathogenic mechanisms. In Pathogenesis of Bacterial Infections in Animals; John Wiley and Sons: Hoboken, NJ, USA, 2010; Volume 3. [Google Scholar]
- Bliven, K.A.; Maurelli, A.T. Evolution of bacterial pathogens within the human host. Microbiol. Spectr. 2016, 4, 4.1.05. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, J.L. Bacterial pathogens: From natural ecosystems to human hosts. Environ. Microbiol. 2013, 15, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Yang, Y.; Zhang, J.-R. Molecular basis of host specificity in human pathogenic bacteria. Emerg. Microbes Infect. 2014, 3, e23. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [Green Version]
- Donnenberg, M.S. Pathogenic strategies of enteric bacteria. Nature 2000, 406, 768–774. [Google Scholar] [CrossRef]
- Wu, H.-J.; Wang, A.H.; Jennings, M.P. Discovery of virulence factors of pathogenic bacteria. Curr. Opin. Chem. Biol. 2008, 12, 93–101. [Google Scholar] [CrossRef]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.I. Bacterial virulence factors. In Bacterial Pathogens and Their Virulence Factors; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–38. [Google Scholar]
- Van Belkum, A.; Almeida, C.; Bardiaux, B.; Barrass, S.V.; Butcher, S.J.; Çaykara, T.; Chowdhury, S.; Datar, R.; Eastwood, I.; Goldman, A. Host-pathogen adhesion as the basis of innovative diagnostics for emerging pathogens. Diagnostics 2021, 11, 1259. [Google Scholar] [CrossRef]
- Milles, L.F.; Schulten, K.; Gaub, H.E.; Bernardi, R.C. Molecular mechanism of extreme mechanostability in a pathogen adhesin. Science 2018, 359, 1527–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solanki, V.; Tiwari, M.; Tiwari, V. Host-bacteria interaction and adhesin study for development of therapeutics. Int. J. Biol. Macromol. 2018, 112, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Ghazaei, C. Advances in the Study of Bacterial Toxins, Their Roles and Mechanisms in Pathogenesis. Malays. J. Med. Sci. 2022, 29, 4. [Google Scholar] [CrossRef]
- Martinović, T.; Andjelković, U.; Gajdošik, M.Š.; Rešetar, D.; Josić, D. Foodborne pathogens and their toxins. J. Proteom. 2016, 147, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Banerji, R.; Karkee, A.; Kanojiya, P.; Saroj, S.D. Pore-forming toxins of foodborne pathogens. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2265–2285. [Google Scholar] [CrossRef] [PubMed]
- Pakbin, B.; Basti, A.A.; Khanjari, A.; Brück, W.M.; Azimi, L.; Karimi, A. Development of high-resolution melting (HRM) assay to differentiate the species of Shigella isolates from stool and food samples. Sci. Rep. 2022, 12, 473. [Google Scholar] [CrossRef]
- Torres, A.G. Current aspects of Shigella pathogenesis. Rev. Latinoam. Microbiol. 2004, 46, 89–97. [Google Scholar]
- Nasser, A.; Mosadegh, M.; Azimi, T.; Shariati, A. Molecular mechanisms of Shigella effector proteins: A common pathogen among diarrheic pediatric population. Mol. Cell. Pediatr. 2022, 9, 12. [Google Scholar] [CrossRef]
- Ashida, H.; Suzuki, T.; Sasakawa, C. Shigella infection and host cell death: A double-edged sword for the host and pathogen survival. Curr. Opin. Microbiol. 2021, 59, 1–7. [Google Scholar] [CrossRef]
- Agaisse, H. Molecular and cellular mechanisms of Shigella flexneri dissemination. Front. Cell. Infect. Microbiol. 2016, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Schnupf, P.; Sansonetti, P.J. Shigella pathogenesis: New insights through advanced methodologies. Microbiol. Spectr. 2019, 7, 7.2.28. [Google Scholar] [CrossRef]
- Killackey, S.A.; Sorbara, M.T.; Girardin, S.E. Cellular aspects of Shigella pathogenesis: Focus on the manipulation of host cell processes. Front. Cell. Infect. Microbiol. 2016, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Argudo, I.; Blocker, A.J. The Shigella T3SS needle transmits a signal for MxiC release, which controls secretion of effectors. Mol. Microbiol. 2010, 78, 1365–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthuramalingam, M.; Whittier, S.K.; Picking, W.L.; Picking, W.D. The Shigella Type III Secretion System: An Overview from Top to Bottom. Microorganisms 2021, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Bajunaid, W.; Haidar-Ahmad, N.; Kottarampatel, A.H.; Ourida Manigat, F.; Silué, N.; Tchagang, C.F.; Tomaro, K.; Campbell-Valois, F.-X. The T3SS of Shigella: Expression, structure, function, and role in vacuole escape. Microorganisms 2020, 8, 1933. [Google Scholar] [CrossRef]
- Belotserkovsky, I.; Sansonetti, P.J. Shigella and enteroinvasive Escherichia coli. In Escherichia coli, a Versatile Pathogen; Springer: Cham, Switzerland, 2018; pp. 1–26. [Google Scholar]
- De Jong, M.F.; Alto, N.M. Cooperative immune suppression by Escherichia coli and Shigella effector proteins. Infect. Immun. 2018, 86, e00560-17. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-C.; Hung, C.-F.; Aljuffali, I.A.; Fang, J.-Y. The roles of the virulence factor IpaB in Shigella spp. in the escape from immune cells and invasion of epithelial cells. Microbiol. Res. 2015, 181, 43–51. [Google Scholar] [CrossRef]
- Mou, X.; Souter, S.; Du, J.; Reeves, A.Z.; Lesser, C.F. Synthetic bottom-up approach reveals the complex interplay of Shigella effectors in regulation of epithelial cell death. Proc. Natl. Acad. Sci. USA 2018, 115, 6452–6457. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Wang, W.; Cao, M.; Zhu, Q.; Ma, T.; Zhang, Y.; Liu, G.; Zhou, X.; Li, B.; Shi, Y. Virulence factors and molecular characteristics of Shigella flexneri isolated from calves with diarrhea. BMC Microbiol. 2021, 21, 214. [Google Scholar] [CrossRef]
- Parsot, C. Shigella spp. and enteroinvasive Escherichia coli pathogenicity factors. FEMS Microbiol. Lett. 2005, 252, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Cruz, C.B.N.d.; Souza, M.C.S.d.; Serra, P.T.; Santos, I.; Balieiro, A.; Pieri, F.A.; Nogueira, P.A.; Orlandi, P.P. Virulence factors associated with pediatric shigellosis in Brazilian Amazon. BioMed Res. Int. 2014, 2014, 539697. [Google Scholar] [CrossRef]
- Kühn, S.; Bergqvist, J.; Gil, M.; Valenzuela, C.; Barrio, L.; Lebreton, S.; Zurzolo, C.; Enninga, J. Actin assembly around the Shigella-containing vacuole promotes successful infection. Cell Rep. 2020, 31, 107638. [Google Scholar] [CrossRef] [PubMed]
- Alwan, H.R.; Maaroof, M.N. Determination of some virulence genes of Shigella sonnei isolated from children stool suffering from diarrhea and investigation about isolates producing Shiga toxin. AIP Conf. Proc. 2022, 2394, 020048. [Google Scholar]
- Wright, S.S.; Vanaja, S.K. Shigella “Osp” pression of innate immunity. Cell 2022, 185, 2205–2207. [Google Scholar] [CrossRef]
- Faherty, C.S.; Lampel, K.A. Shigella. In Food Microbiology Fundamentals and Frontiers; John Wiley and Sons: Hoboken, NJ, USA, 2019; pp. 317–345. [Google Scholar]
- Ashida, H.; Sasakawa, C. Shigella IpaH family effectors as a versatile model for studying pathogenic bacteria. Front. Cell. Infect. Microbiol. 2016, 5, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kueltzo, L.A.; Osiecki, J.; Barker, J.; Picking, W.L.; Ersoy, B.; Picking, W.D.; Middaugh, C.R. Structure-function analysis of invasion plasmid antigen C (IpaC) from Shigella flexneri. J. Biol. Chem. 2003, 278, 2792–2798. [Google Scholar] [CrossRef] [Green Version]
- Cohen, D.; Meron-Sudai, S.; Bialik, A.; Asato, V.; Goren, S.; Ariel-Cohen, O.; Reizis, A.; Hochberg, A.; Ashkenazi, S. Serum IgG antibodies to Shigella lipopolysaccharide antigens—A correlate of protection against shigellosis. Hum. Vaccines Immunother. 2019, 15, 1401–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nezhad, A.Y.; Heidary, S.; Far, A.M. Investigation of Shigella lipopolysaccharides Effects on Immunity Stimulation of Host Cells. Int. Trans. J. Eng. Manag. Appl. Sci. Technol. 2019, 10, 465–476. [Google Scholar]
- Mey, A.R.; Gómez-Garzón, C.; Payne, S.M. Iron Transport and Metabolism in Escherichia, Shigella, and Salmonella. EcoSal Plus 2021, 9, eESP-0034. [Google Scholar] [CrossRef]
- Hu, W.; Chan, H.; Lu, L.; Wong, K.T.; Wong, S.H.; Li, M.X.; Xiao, Z.G.; Cho, C.H.; Gin, T.; Chan, M.T. Autophagy in intracellular bacterial infection. Semin. Cell Dev. Biol. 2020, 101, 41–50. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, Y.; Peng, T.; Zhou, P.; Ding, X.; Li, Z.; Zhong, H.; Xu, Y.; Chen, S.; Hang, H.C. Nε-fatty acylation of multiple membrane-associated proteins by Shigella IcsB effector to modulate host function. Nat. Microbiol. 2018, 3, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Yoshimori, T.; Suzuki, T.; Sagara, H.; Mizushima, N.; Sasakawa, C. Escape of intracellular Shigella from autophagy. Science 2005, 307, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Farfán, M.J.; Toro, C.S.; Barry, E.M.; Nataro, J.P. Shigella enterotoxin-2 is a type III effector that participates in Shigella-induced interleukin 8 secretion by epithelial cells. FEMS Immunol. Med. Microbiol. 2011, 61, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Moosavian, M.; Seyed-Mohammadi, S.; Sheikh, A.F.; Khoshnood, S.; Dezfuli, A.A.; Saki, M.; Ghaderian, G.; Shahi, F.; Abdi, M.; Abbasi, F. Prevalence of enterotoxin-encoding genes among diverse Shigella strains isolated from patients with diarrhea, southwest Iran. Acta Microbiol. Immunol. Hung. 2019, 66, 91–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingersoll, M.; Groisman, E.A.; Zychlinsky, A. Pathogenicity islands of Shigella. In Pathogenicity Islands and the Evolution of Pathogenic Microbes; Springer: Berlin/Heidelberg, Germany, 2002; pp. 49–65. [Google Scholar]
- Pakbin, B.; Akhondzadeh Basti, A.; Khanjari, A.; Azimi, L.; Karimi, A. Differentiation of stx1A gene for detection of Escherichia coli serotype O157: H7 and Shigella dysenteriae type 1 in food samples using high resolution melting curve analysis. Food Sci. Nutr. 2020, 8, 3665–3672. [Google Scholar] [CrossRef]
- Strockbine, N.A.; Maurelli, A.T. Shigella. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley and Sons: Hoboken, NJ, USA, 2015; pp. 1–26. [Google Scholar]
- Mark Taylor, C. Enterohaemorrhagic Escherichia coli and Shigella dysenteriae type 1-induced haemolytic uraemic syndrome. Pediatr. Nephrol. 2008, 23, 1425–1431. [Google Scholar] [CrossRef] [Green Version]
- Nabatanzi, M.; Kwesiga, B.; Bulage, L.; Lubwama, B.; Ario, A.R.; Harris, J. Epidemiology of Dysentery, Uganda, 2014–2018. J. Interv. Epidemiol. Public Health 2022, 5, 22. [Google Scholar]
- Kotloff, K.L. Shigella infection in children and adults: A formidable foe. Lancet Glob. Health 2017, 5, e1166–e1167. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Wang, W.; Zhang, L.; Liu, G.; Zhou, X.; Li, B.; Shi, Y.; Zhu, Z.; Zhang, J. Epidemic and molecular characterization of fluoroquinolone-resistant Shigella dysenteriae 1 isolates from calves with diarrhea. BMC Microbiol. 2021, 21, 6. [Google Scholar] [CrossRef]
- Mattock, E.; Blocker, A.J. How do the virulence factors of Shigella work together to cause disease? Front. Cell. Infect. Microbiol. 2017, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Al-Dahmoshi, H.O.; Al-Khafaji, N.S.; Al-Allak, M.H.; Salman, W.K.; Alabbasi, A.H. A review on shigellosis: Pathogenesis and antibiotic resistance. Drug Invent. Today 2020, 14, 793–798. [Google Scholar]
- Baker, S.; The, H.C. Recent insights into Shigella: A major contributor to the global diarrhoeal disease burden. Curr. Opin. Infect. Dis. 2018, 31, 449. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular mechanisms of Escherichia coli pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T. Molecular characterization of multidrug-resistant Shigella spp. of food origin. Int. J. Food Microbiol. 2015, 194, 78–82. [Google Scholar] [CrossRef]
- Yang, F.; Yang, J.; Zhang, X.; Chen, L.; Jiang, Y.; Yan, Y.; Tang, X.; Wang, J.; Xiong, Z.; Dong, J. Genome dynamics and diversity of Shigella species, the etiologic agents of bacillary dysentery. Nucleic Acids Res. 2005, 33, 6445–6458. [Google Scholar] [CrossRef]
- Basak, C.; Chakraborty, R. A novel strain of Shigella isolated from the gut of Lepidocephalichthys guntea has in its genome a complete gene package for Typell secretion system, and elaborate repertoire of genes responsible for multiple antibiotic-resistance and metal resistance via specific efflux channels. Lett. Appl. Microbiol. 2022, 76, ovac049. [Google Scholar]
- Gyles, C. Shiga toxin-producing Escherichia coli: An overview. J. Anim. Sci. 2007, 85, E45–E62. [Google Scholar] [CrossRef]
- Bergan, J.; Lingelem, A.B.D.; Simm, R.; Skotland, T.; Sandvig, K. Shiga toxins. Toxicon 2012, 60, 1085–1107. [Google Scholar] [CrossRef]
- Melton-Celsa, A.R. Shiga toxin (Stx) classification, structure, and function. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.L.; Fratamico, P.M.; Gunther IV, N.W. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar] [PubMed]
- Johannes, L.; Römer, W. Shiga toxins—From cell biology to biomedical applications. Nat. Rev. Microbiol. 2010, 8, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.C.; Zhang, Y.; Bai, X.; Xiong, Y.; Wang, Y.; Yang, X.; Xu, Q.; He, X. Structural and functional characterization of Stx2k, a new subtype of Shiga toxin 2. Microorganisms 2019, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Joseph, A.; Cointe, A.; Mariani Kurkdjian, P.; Rafat, C.; Hertig, A. Shiga toxin-associated hemolytic uremic syndrome: A narrative review. Toxins 2020, 12, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.S.; Ng, T.B. Shiga toxins: From structure and mechanism to applications. Appl. Microbiol. Biotechnol. 2016, 100, 1597–1610. [Google Scholar] [CrossRef]
- Pezeshkian, W.; Gao, H.; Arumugam, S.; Becken, U.; Bassereau, P.; Florent, J.-C.; Ipsen, J.H.; Johannes, L.; Shillcock, J.C. Mechanism of Shiga toxin clustering on membranes. ACS Nano 2017, 11, 314–324. [Google Scholar] [CrossRef]
- Sandvig, K.; Kavaliauskiene, S.; Skotland, T. The Protein Toxins Ricin and Shiga Toxin as Tools to Explore Cellular Mechanisms of Internalization and Intracellular Transport. Toxins 2021, 13, 377. [Google Scholar] [CrossRef]
- Álvarez, R.S.; Gómez, F.D.; Zotta, E.; Paton, A.W.; Paton, J.C.; Ibarra, C.; Sacerdoti, F.; Amaral, M.M. Combined action of shiga toxin type 2 and subtilase cytotoxin in the pathogenesis of hemolytic uremic syndrome. Toxins 2021, 13, 536. [Google Scholar] [CrossRef]
- Legros, N.; Pohlentz, G.; Steil, D.; Müthing, J. Shiga toxin-glycosphingolipid interaction: Status quo of research with focus on primary human brain and kidney endothelial cells. Int. J. Med. Microbiol. 2018, 308, 1073–1084. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, B.; Ding, X.; Bin, P.; Yang, Y.; Zhu, G. Regulatory Mechanisms between Quorum Sensing and Virulence in Salmonella. Microorganisms 2022, 10, 2211. [Google Scholar] [CrossRef]
- Xu, P.; Yang, J.; Lu, L.; Feng, E.; Wang, H.; Lu, Y.; Zhu, L. The effect of quorum sensing system for growth competitiveness on Shigella flexneri. Yi Chuan Hered. 2015, 37, 487–493. [Google Scholar]
- Kalia, V.C.; Patel, S.K.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2019, 37, 68–90. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic representation of pathogenesis mechanisms of Dysenteriae and non-Dysenteriae species of Shigella.
Figure 1.
Schematic representation of pathogenesis mechanisms of Dysenteriae and non-Dysenteriae species of Shigella.


{kind=link}
Table 1.
Virulence factor encoding genes in Dysenteriae and non-Dysenteriae Shigella species.
| Class | Virulence Factor Gene | Function | Type (D/ND) 1 |
|---|---|---|---|
| Effector delivery | GspC, GspD, GspE, GspF, GspG, GspH, GspI, GspJ, GspK, GspL, GspM, GspO | T2SS structure | D |
| MxiA, MxiC, MxiD, spa9, spa13, spa15, spa24, spa29, spa33, spa40, spa47 | T3SS structure | D/ND | |
| TssA, TssB, TssC, TssD, TssE, TssF, TssG, TssH, TssI, TssJ, TssK, TssL, TssM | T6SS structure | ND | |
| Effectors | IcsA | intracellular motility | D/ND |
| IcsB | autophagy evasion and actin destabilisation | D/ND | |
| IpaA | actin depolymerisation | D/ND | |
| IpaB | endocytosis, phagosome escape, cell apoptosis inhibition | D/ND | |
| IpaC | adhesion to host cell and phagosome escape | D/ND | |
| IpaH | inflammatory response suppression, phagosome escape, protein degradation, cell cycle intervention and pyroptosis induction | D/ND | |
| WxxxE effectors (IpgB1 and IpgB2) | cell entry, invasion, host cell process manipulation, inflammation regulation, actin organisation, vacuole formation, apoptosis and tight junction disruption | D/ND | |
| IpgD | actin destabilisation, endocytic vacuole lysis and apoptosis prevention | D/ND | |
| ospB | immune response suppression | D/ND | |
| ospC | cell pyroptosis inhibition | D/ND | |
| ospD | cell pyroptosis and necrosis inhibition | D/ND | |
| ospE | bacterial dissemination | D/ND | |
| ospF | immune response suppression | D/ND | |
| ospG | immune response suppression | D/ND | |
| ospI | immune response suppression | D/ND | |
| virA | actin destabilisation and endocytic vacuole lysis | D/ND | |
| Toxins | Set1A | encoding Shigella enterotoxin 1, early diarrhoea | ND |
| Set1B | encoding Shigella enterotoxin 1, early diarrhoea | ND | |
| senB | encoding Shigella enterotoxin 2, early diarrhoea | ND | |
| gtrA | encoding lipopolysaccharide, intracellular spread, immune modulation, host cell defence inhibition | D/ND | |
| gtrB | encoding lipopolysaccharide, intracellular spread, immune modulation, host cell defence inhibition | D/ND | |
| pic | encoding protein involved in intestinal colonisation toxin, SPATEs, haemagglutinin activity | ND | |
| sigA | encoding Shigella IgA-like protease homology, SPATEs, intestinal fluid accumulation | ND | |
| stxA | encoding Shiga-toxin subunit A, proteolytic activity, peptide synthesis inhibition, haemorrhagic colitis and the haemolytic uremic syndrome | D | |
| stxB | encoding Shiga-toxin B subunits, binding to the cellular receptors, haemorrhagic colitis and the haemolytic uremic syndrome | D | |
| Metabolic | iucA, iucB, iucC, iucD, iutA | encoding aerobactin, iron uptake | D/ND |
| ShuA, ShuS, ShuT, ShuU, ShuV, ShuY, ShuX | encoding hemophores, heme uptake system, DNA protection against oxidative damages | D |
1 D, Dysenteriae Shigella species; ND, non-Dysenteriae Shigella species.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pakbin, B.; Brück, W.M.; Brück, T.B. Molecular Mechanisms of Shigella Pathogenesis; Recent Advances. Int. J. Mol. Sci. 2023, 24, 2448. https://doi.org/10.3390/ijms24032448
AMA Style
Pakbin B, Brück WM, Brück TB. Molecular Mechanisms of Shigella Pathogenesis; Recent Advances. International Journal of Molecular Sciences. 2023; 24(3):2448. https://doi.org/10.3390/ijms24032448
Chicago/Turabian StylePakbin, Babak, Wolfram Manuel Brück, and Thomas B. Brück. 2023. "Molecular Mechanisms of Shigella Pathogenesis; Recent Advances" International Journal of Molecular Sciences 24, no. 3: 2448. https://doi.org/10.3390/ijms24032448
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.