
Genotype Combinations Drive Variability in the Microbiome Configuration of the Rhizosphere of Maize/Bean Intercropping System
 , , , , ,
, , , , , 
Abstract
:1. Introduction
2. Results
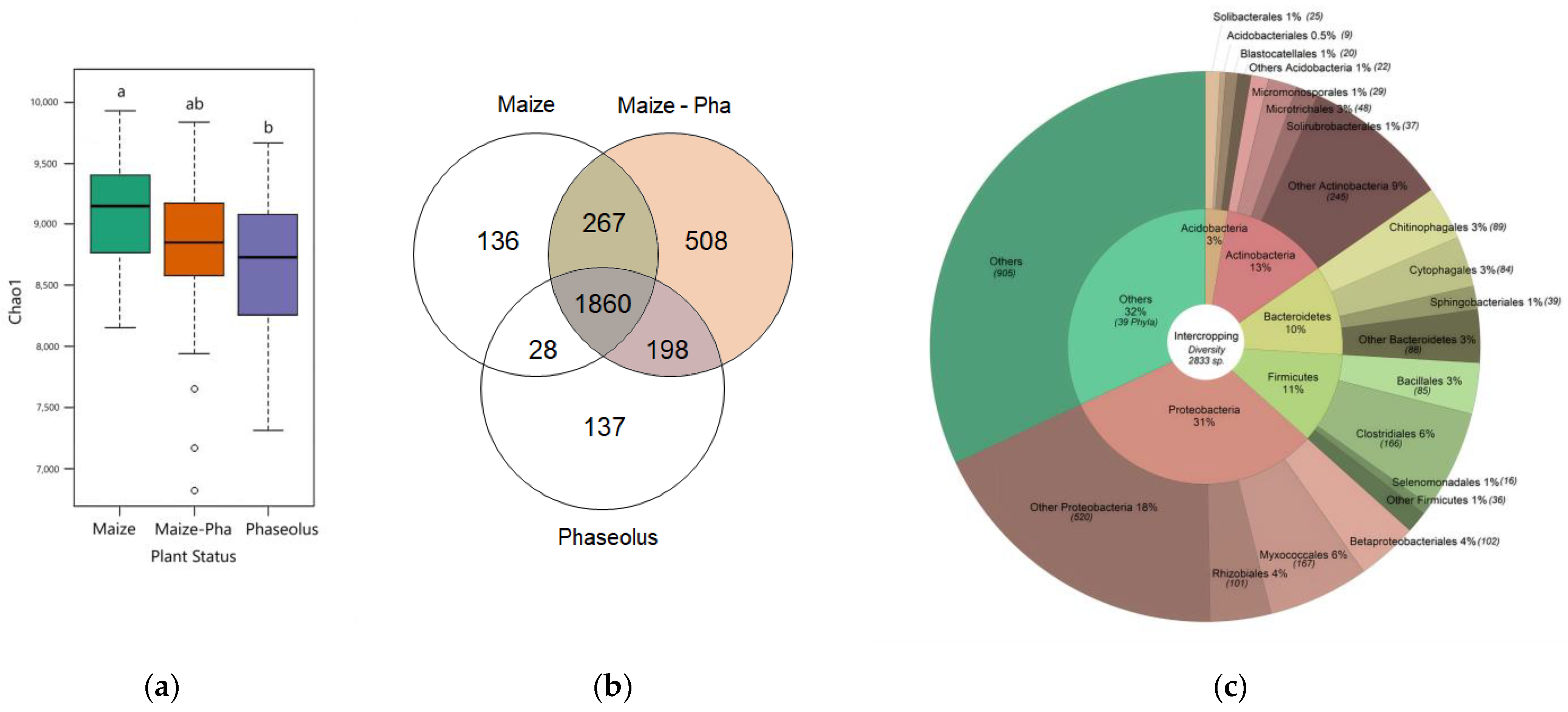
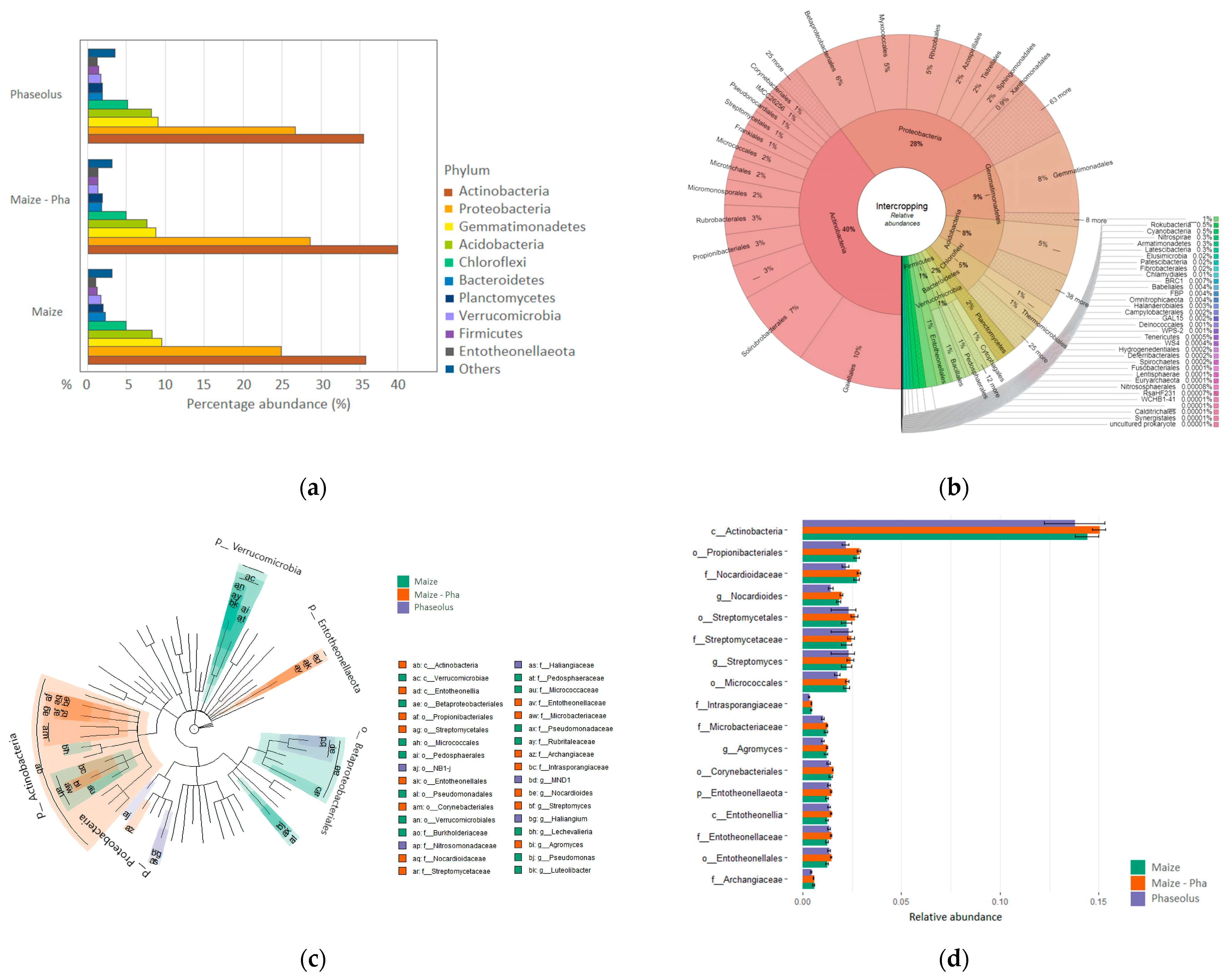
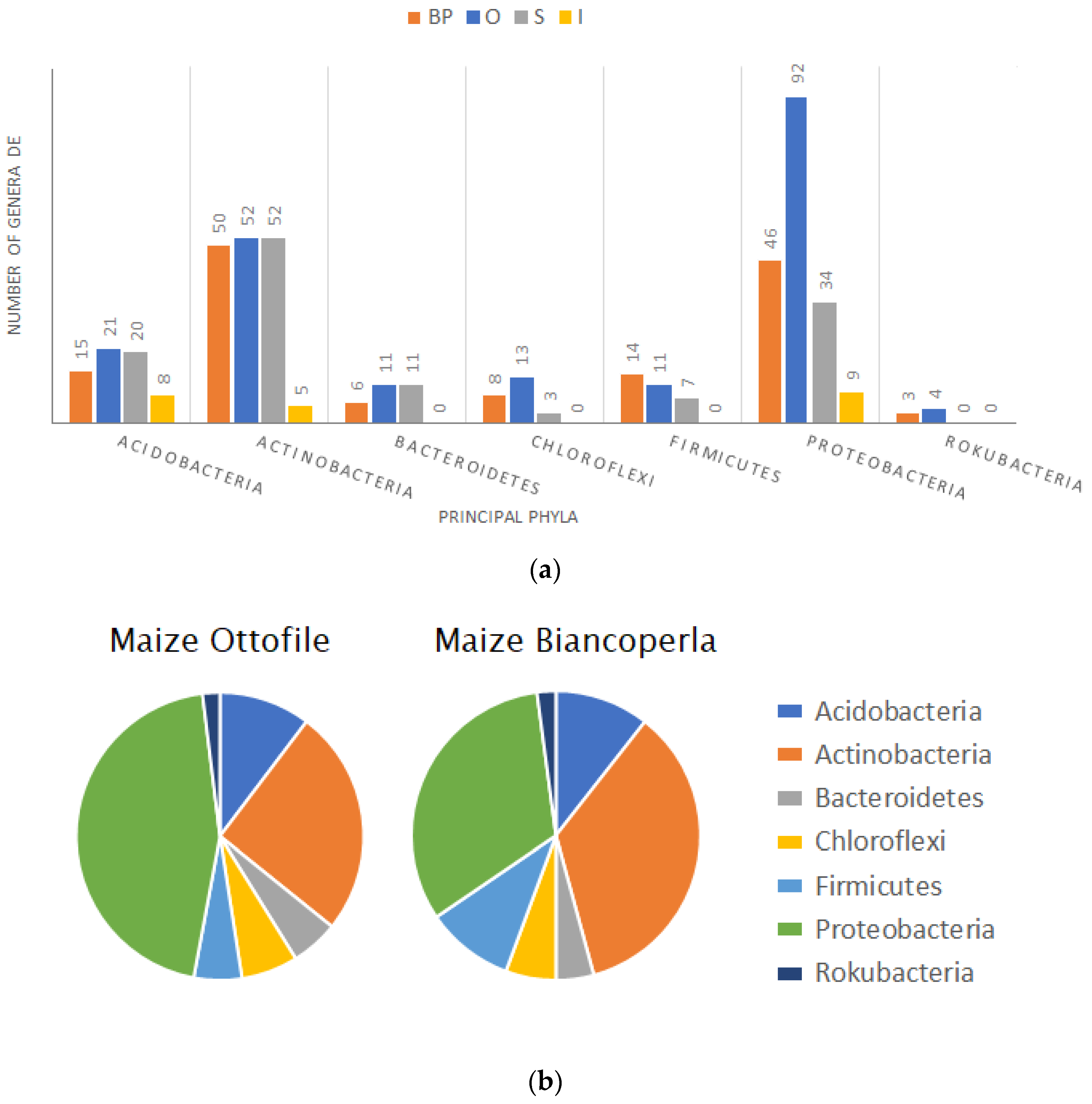
2.1. Composition of the Bacterial Community in the Rhizosphere of Maize/Bean Intercropping System
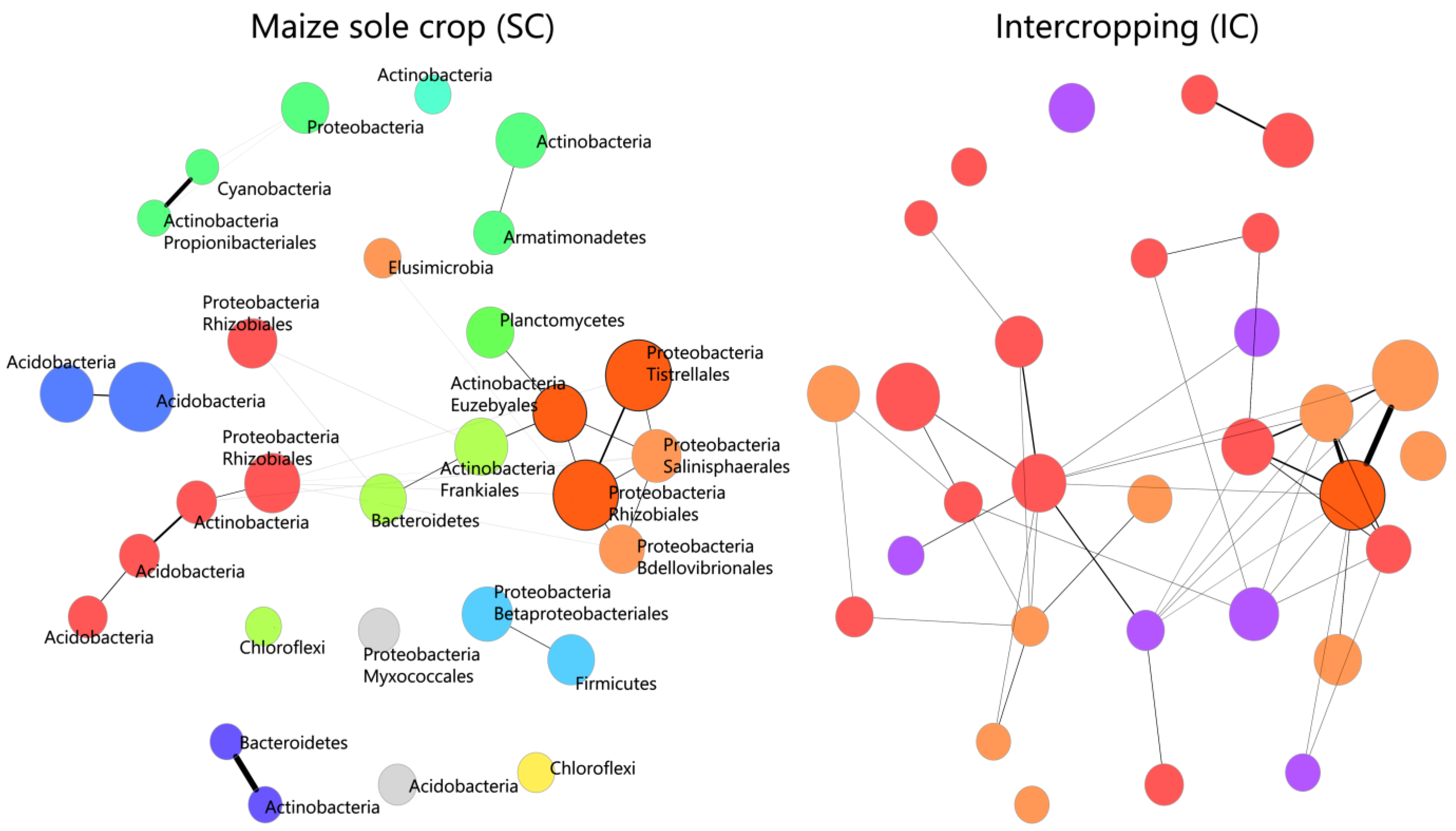
2.2. Differentiation of the Bacterial Community Structure in the Rhizosphere of the Maize/Bean Intercropping System Compared to Sole Crop
2.3. Composition of Rhizosphere Bacterial Community in Different Maize/Bean Intercropping Combinations
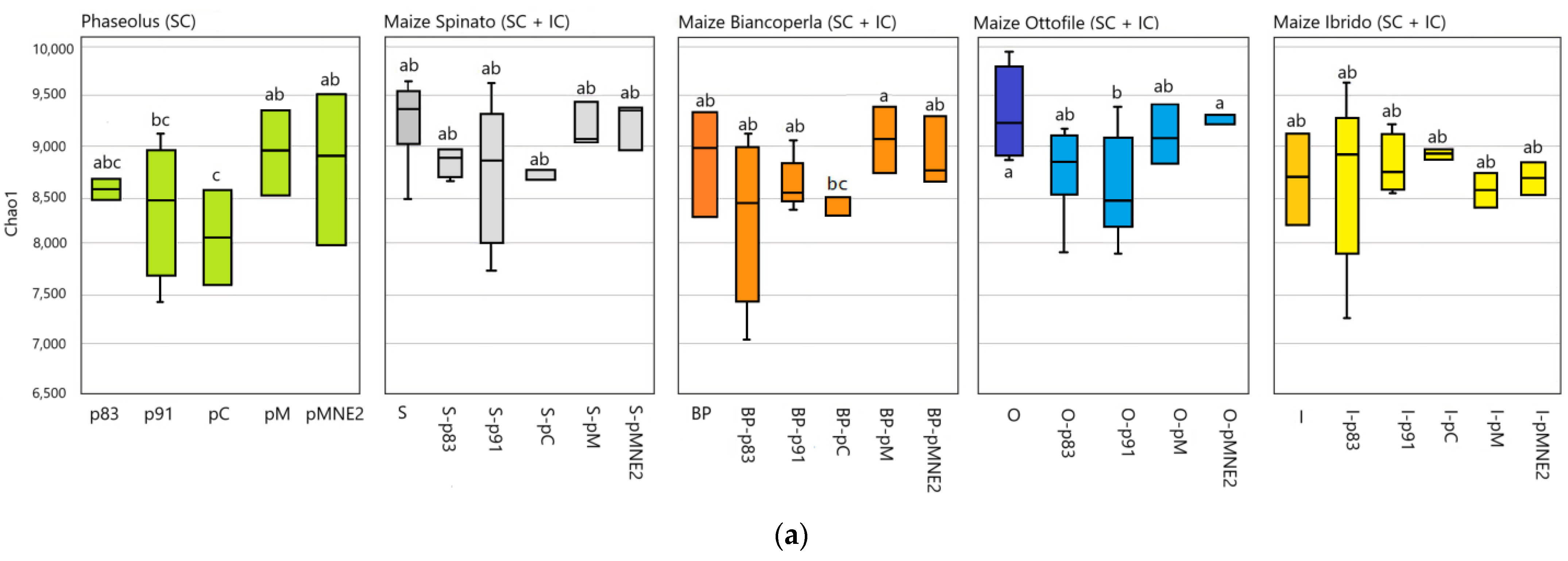
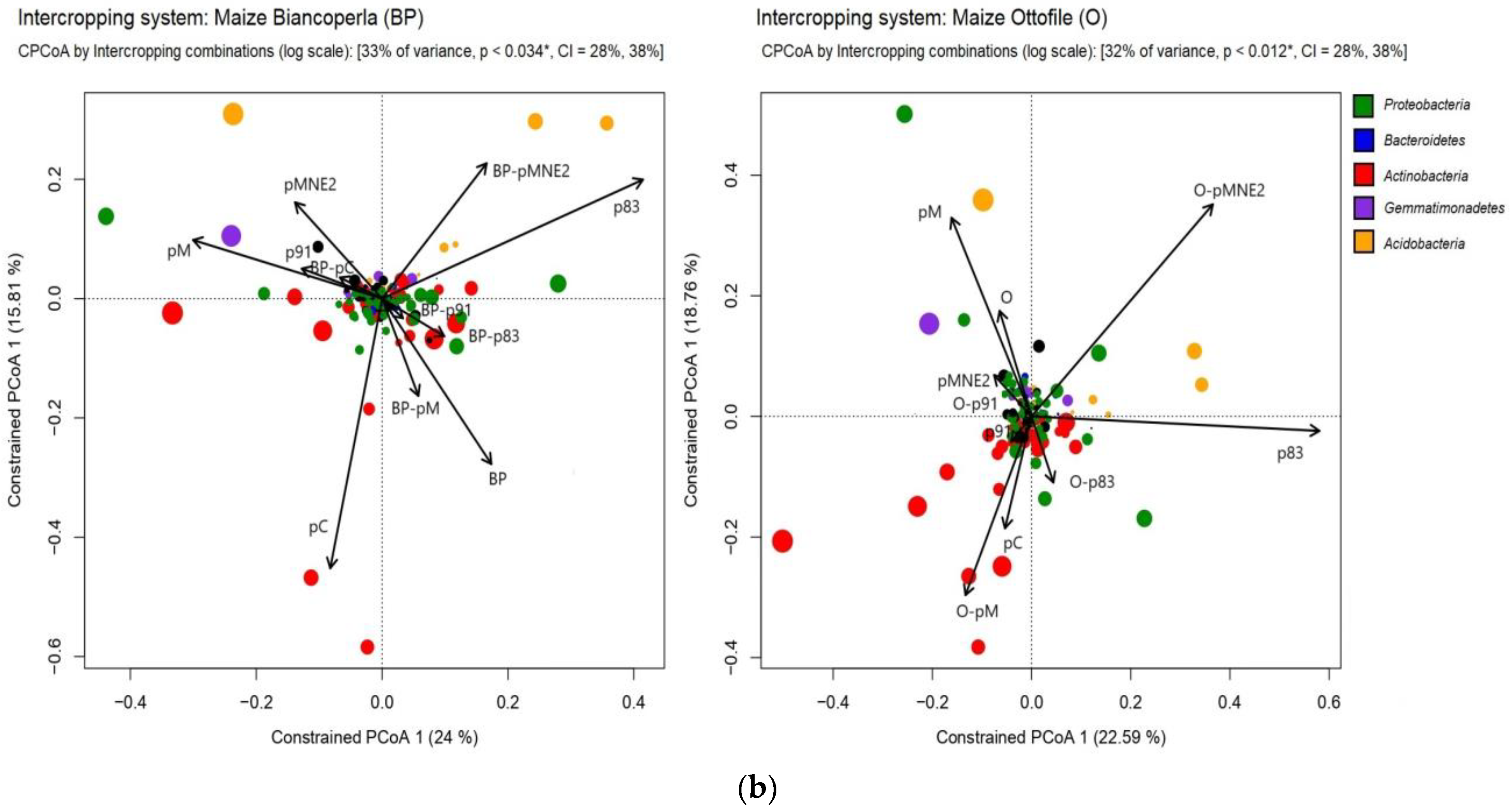
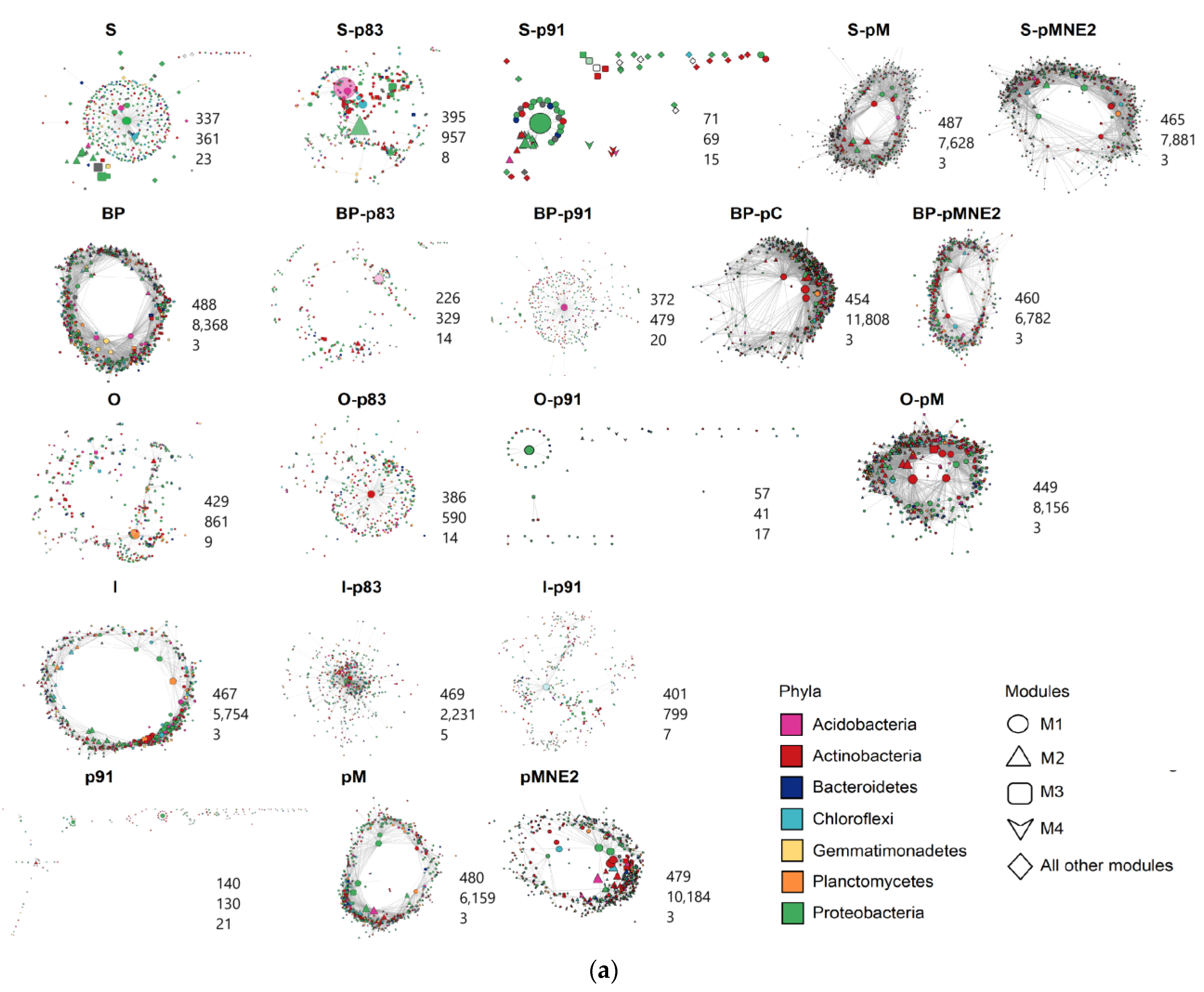
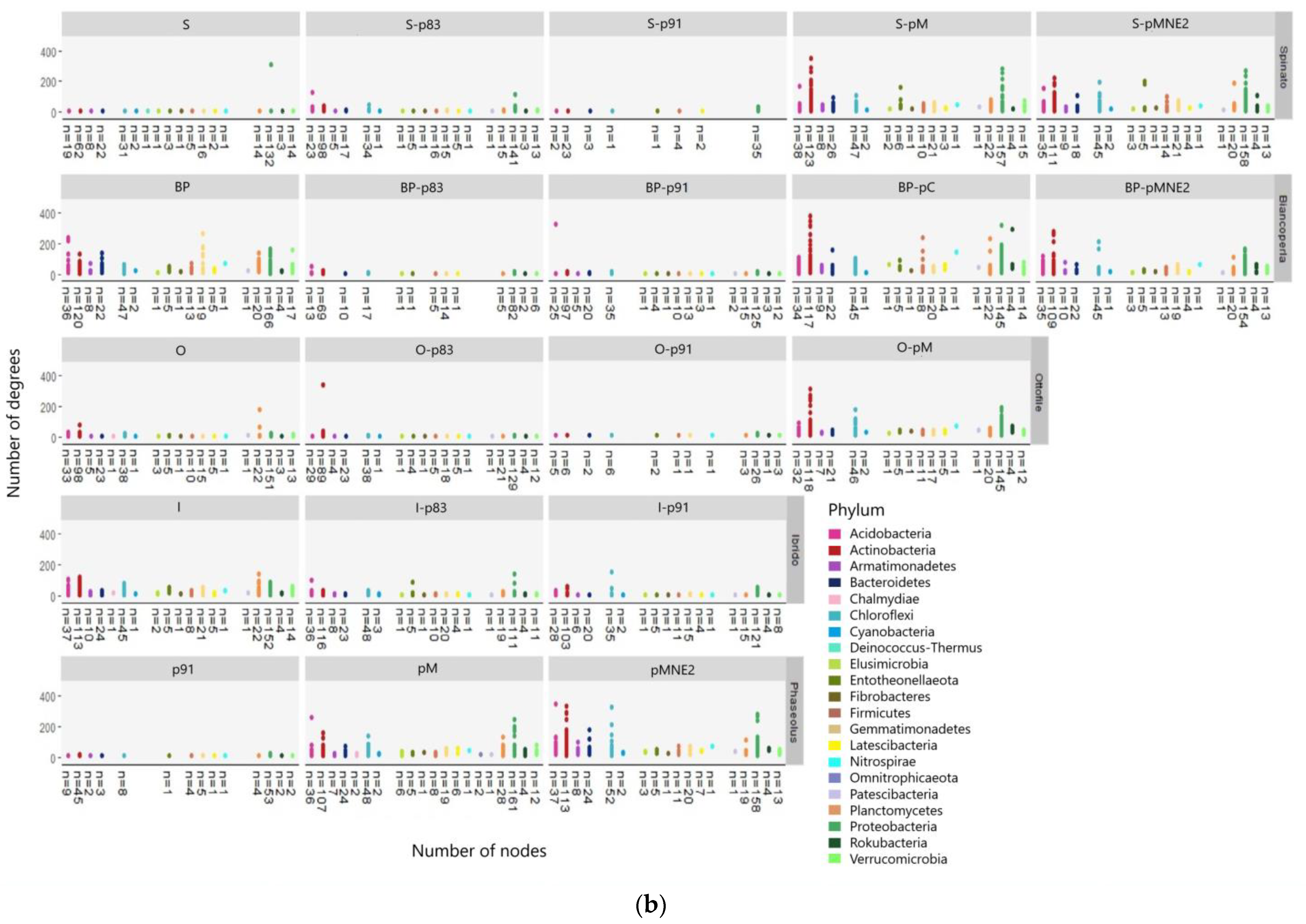
2.4. Bacterial Community Structure in the Rhizosphere of Different Maize/Bean Intercropping Combinations
3. Discussion
3.1. Members of Proteobacteria and Actinobacteria Phyla Are at the Basis of the Reconfiguration of the Bacterial Community in the Rhizosphere of Maize/Bean Intercropping System
3.2. Combination of Different Maize/Bean Lines in Intercropping Significantly Alters the Composition and Structure of the Rhizosphere Microbiome
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Metabarcoding Analysis
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duchene, O.; Vian, J.-F.; Celette, F. Intercropping with Legume for Agroecological Cropping Systems: Complementarity and Facilitation Processes and the Importance of Soil Microorganisms. A Review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Jensen, E.; Bedoussac, L.; Carlsson, G.; Journet, E.; Justes, E.; Hauggaard-Nielsen, H. Enhancing Yields in Organic Crop Production by Eco-Functional Intensification. Sustain. Agric. Res. 2015, 4, 42–50. [Google Scholar] [CrossRef]
- Willey, R. Evaluation and Presentation of Intercropping Advantages. Exp. Agric. 1985, 21, 119–133. [Google Scholar] [CrossRef]
- Altieri, M.A. Applying Agroecology to Enhance the Productivity of Peasant Farming Systems in Latin America. Environ. Dev. Sustain. 1999, 1, 197–217. [Google Scholar] [CrossRef]
- Knörzer, H.; Graeff-Hönninger, S.; Guo, B.; Wang, P.; Claupein, W. The Rediscovery of Intercropping in China: A Traditional Cropping System for Future Chinese Agriculture—A Review. In Climate Change, Intercropping, Pest Control and Beneficial Microorganisms; Springer: Dordrecht, The Netherlands, 2009; pp. 13–44. [Google Scholar] [CrossRef]
- Maitra, S.; Hossain, A.; Brestic, M.; Skalicky, M.; Ondrisik, P.; Gitari, H.; Brahmachari, K.; Shankar, T.; Bhadra, P.; Palai, J.B. Intercropping—A Low Input Agricultural Strategy for Food and Environmental Security. Agronomy 2021, 11, 343. [Google Scholar] [CrossRef]
- Loreau, M.; Barbier, M.; Filotas, E.; Gravel, D.; Isbell, F.; Miller, S.J.; Montoya, J.M.; Wang, S.; Aussenac, R.; Germain, R. Biodiversity as Insurance: From Concept to Measurement and Application. Biol. Rev. 2021, 96, 2333–2354. [Google Scholar] [CrossRef]
- Jensen, E.S.; Chongtham, I.R.; Dhamala, N.R.; Rodriguez, C.; Carton, N.; Carlsson, G. Diversifying European Agricultural Systems by Intercropping Grain Legumes and Cereals. Cienc. Investig. Agrar. Rev. Latinoam. Cienc. Agric. 2020, 47, 174–186. [Google Scholar]
- De Deyn, G.B.; Cornelissen, J.H.; Bardgett, R.D. Plant Functional Traits and Soil Carbon Sequestration in Contrasting Biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Shiel, R.S.; Ostle, N.J.; McNamara, N.P.; Oakley, S.; Young, I.; Freeman, C.; Fenner, N.; Quirk, H.; Bardgett, R.D. Additional Carbon Sequestration Benefits of Grassland Diversity Restoration. J. Appl. Ecol. 2011, 48, 600–608. [Google Scholar] [CrossRef]
- Raji, S.G.; Dörsch, P. Effect of Legume Intercropping on N2O Emissions and CH 4 Uptake during Maize Production in the Great Rift Valley, Ethiopia. Biogeosciences 2020, 17, 345–359. [Google Scholar] [CrossRef]
- Fréville, H.; Montazeaud, G.; Forst, E.; David, J.; Papa, R.; Tenaillon, M.I. Shift in Beneficial Interactions during Crop Evolution. Evol. Appl. 2022, 15, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Bokern, D.; Peeters, A.; Westhoek, H. The Role of Legumes in Bringing Protein to the Table. In Legumes in Cropping Systems; CABI: Wallingford, UK, 2017; pp. 18–36. [Google Scholar] [CrossRef]
- Bellucci, E.; Mario Aguilar, O.; Alseekh, S.; Bett, K.; Brezeanu, C.; Cook, D.; De la Rosa, L.; Delledonne, M.; Dostatny, D.F.; Ferreira, J.J. The INCREASE Project: Intelligent Collections of Food-legume Genetic Resources for European Agrofood Systems. Plant J. 2021, 108, 646–660. [Google Scholar] [CrossRef] [PubMed]
- Mamine, F.; Farès, M. Barriers and Levers to Developing Wheat–Pea Intercropping in Europe: A Review. Sustainability 2020, 12, 6962. [Google Scholar] [CrossRef]
- Alcon, F.; Marín-Miñano, C.; Zabala, J.A.; de-Miguel, M.-D.; Martínez-Paz, J.M. Valuing diversification benefits through intercropping in Mediterranean agroecosystems: A choice experiment approach. Ecol. Econ. 2020, 171, 106593. [Google Scholar] [CrossRef]
- Lakshmanan, V.; Selvaraj, G.; Bais, H.P. Functional Soil Microbiome: Belowground Solutions to an Aboveground Problem. Plant Physiol. 2014, 166, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Layek, J.; Das, A.; Mitran, T.; Nath, C.; Meena, R.S.; Yadav, G.S.; Shivakumar, B.; Kumar, S.; Lal, R. Cereal+ Legume Intercropping: An Option for Improving Productivity and Sustaining Soil Health. Legumes for Soil Health and Sustainable Management; Springer: Berlin/Heidelberg, Germany, 2018; pp. 347–386. [Google Scholar] [CrossRef]
- Bedoussac, L.; Journet, E.-P.; Hauggaard-Nielsen, H.; Naudin, C.; Corre-Hellou, G.; Jensen, E.S.; Prieur, L.; Justes, E. Ecological Principles Underlying the Increase of Productivity Achieved by Cereal-Grain Legume Intercrops in Organic Farming. A Review. Agron. Sustain. Dev. 2015, 35, 911–935. [Google Scholar] [CrossRef]
- Bedoussac, L.; Justes, E. The Efficiency of a Durum Wheat-Winter Pea Intercrop to Improve Yield and Wheat Grain Protein Concentration Depends on N Availability during Early Growth. Plant Soil 2010, 330, 19–35. [Google Scholar] [CrossRef]
- Bedoussac, L.; Justes, E. Dynamic Analysis of Competition and Complementarity for Light and N Use to Understand the Yield and the Protein Content of a Durum Wheat–Winter Pea Intercrop. Plant Soil 2010, 330, 37–54. [Google Scholar] [CrossRef]
- Hinsinger, P.; Betencourt, E.; Bernard, L.; Brauman, A.; Plassard, C.; Shen, J.; Tang, X.; Zhang, F. P for Two, Sharing a Scarce Resource: Soil Phosphorus Acquisition in the Rhizosphere of Intercropped Species. Plant Physiol. 2011, 156, 1078–1086. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant Species and Soil Type Cooperatively Shape the Structure and Function of Microbial Communities in the Rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar] [CrossRef]
- Peiffer, J.A.; Spor, A.; Koren, O.; Jin, Z.; Tringe, S.G.; Dangl, J.L.; Buckler, E.S.; Ley, R.E. Diversity and Heritability of the Maize Rhizosphere Microbiome under Field Conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6548–6553. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yang, Y.; Liu, P.; Xu, R.; Rensing, C.; Fu, X.; Liao, H. Genotype and Rhizobium Inoculation Modulate the Assembly of Soybean Rhizobacterial Communities. Plant Cell Environ. 2019, 42, 2028–2044. [Google Scholar] [CrossRef] [PubMed]
- Pivato, B.; Mazurier, S.; Lemanceau, P.; Siblot, S.; Berta, G.; Mougel, C.; Van Tuinen, D. Medicago Species Affect the Community Composition of Arbuscular Mycorrhizal Fungi Associated with Roots. New Phytol. 2007, 176, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Pivato, B.; Bru, D.; Busset, H.; Deau, F.; Matejicek, A.; Philippot, L.; Moreau, D. Positive Effects of Plant Association on Rhizosphere Microbial Communities Depend on Plant Species Involved and Soil Nitrogen Level. Soil Biol. Biochem. 2017, 114, 1–4. [Google Scholar] [CrossRef]
- Tkacz, A.; Bestion, E.; Bo, Z.; Hortala, M.; Poole, P.S. Influence of Plant Fraction, Soil, and Plant Species on Microbiota: A Multikingdom Comparison. MBio 2020, 11, 10–1128. [Google Scholar] [CrossRef]
- Li, S.; Wu, F. Diversity and Co-Occurrence Patterns of Soil Bacterial and Fungal Communities in Seven Intercropping Systems. Front. Microbiol. 2018, 9, 1521. [Google Scholar] [CrossRef]
- Chamkhi, I.; Cheto, S.; Geistlinger, J.; Zeroual, Y.; Kouisni, L.; Bargaz, A.; Ghoulam, C. Legume-Based Intercropping Systems Promote Beneficial Rhizobacterial Community and Crop Yield under Stressing Conditions. Ind. Crops Prod. 2022, 183, 114958. [Google Scholar] [CrossRef]
- Lai, H.; Gao, F.; Su, H.; Zheng, P.; Li, Y.; Yao, H. Nitrogen Distribution and Soil Microbial Community Characteristics in a Legume–Cereal Intercropping System: A Review. Agronomy 2022, 12, 1900. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Liu, Q.; Zhao, X.; Xie, S.; Wang, Z.; Wen, R.; Zhang, M.; Chen, B. Effect of Two Different Sugarcane Cultivars on Rhizosphere Bacterial Communities of Sugarcane and Soybean upon Intercropping. Front. Microbiol. 2021, 11, 596472. [Google Scholar] [CrossRef]
- Pivato, B.; Semblat, A.; Guégan, T.; Jacquiod, S.; Martin, J.; Deau, F.; Moutier, N.; Lecomte, C.; Burstin, J.; Lemanceau, P. Rhizosphere Bacterial Networks, but Not Diversity, Are Impacted by Pea-Wheat Intercropping. Front. Microbiol. 2021, 12, 674556. [Google Scholar] [CrossRef]
- Salse, J.; Barnard, R.L.; Veneault-Fourrey, C.; Rouached, H. Strategies for Breeding Crops for Future Environments. Trends Plant Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Homulle, Z.; George, T.S.; Karley, A.J. Root Traits with Team Benefits: Understanding Belowground Interactions in Intercropping Systems. Plant Soil 2021, 471, 1–26. [Google Scholar] [CrossRef]
- Lin, W.; Li, N.; Chen, L.; Wu, Z.; Lin, W.; Shen, L. Effects of Maize and Sobean Interspecific Interactions on Rhizospheric Bacteria Community Structure and Diversity. Chin. J. Eco Agric. 2022, 30, 26–37. [Google Scholar] [CrossRef]
- Fu, Z.; Li, Z.; Ping, C.; Qing, D.; Ting, P.; Chun, S.; Wang, X.; Liu, W.; Yang, W.; Yong, T. Effects of Maize-Soybean Relay Intercropping on Crop Nutrient Uptake and Soil Bacterial Community. J. Integr. Agric. 2019, 18, 2006–2018. [Google Scholar] [CrossRef]
- Zhao, R.; Li, X.; Bei, S.; Li, D.; Li, H.; Christie, P.; Bender, S.F.; Zhang, J. Enrichment of nosZ-type Denitrifiers by Arbuscular Mycorrhizal Fungi Mitigates N2O Emissions from Soybean Stubbles. Environ. Microbiol. 2021, 23, 6587–6602. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; He, W.; Shen, Y.; Zhu, L.; Yao, X.; Sun, R.; Dai, C.; Sun, B.; Chen, Y. Interspecific Neighbor Stimulates Peanut Growth Through Modulating Root Endophytic Microbial Community Construction. Front. Plant Sci. 2022, 13, 830666. [Google Scholar] [CrossRef] [PubMed]
- Vora, S.M.; Joshi, P.; Belwalkar, M.; Archana, G. Root Exudates Influence Chemotaxis and Colonization of Diverse Plant Growth Promoting Rhizobacteria in the Pigeon Pea–Maize Intercropping System. Rhizosphere 2021, 18, 100331. [Google Scholar] [CrossRef]
- Li, Q.; Chen, J.; Wu, L.; Luo, X.; Li, N.; Arafat, Y.; Lin, S.; Lin, W. Belowground Interactions Impact the Soil Bacterial Community, Soil Fertility, and Crop Yield in Maize/Peanut Intercropping Systems. Int. J. Mol. Sci. 2018, 19, 622. [Google Scholar] [CrossRef]
- Chen, J.; Arafat, Y.; Wu, L.; Xiao, Z.; Li, Q.; Khan, M.A.; Khan, M.U.; Lin, S.; Lin, W. Shifts in Soil Microbial Community, Soil Enzymes and Crop Yield under Peanut/Maize Intercropping with Reduced Nitrogen Levels. Appl. Soil Ecol. 2018, 124, 327–334. [Google Scholar] [CrossRef]
- Li, L.; Tang, C.; Rengel, Z.; Zhang, F. Chickpea Facilitates Phosphorus Uptake by Intercropped Wheat from an Organic Phosphorus Source. Plant Soil 2003, 248, 297–303. [Google Scholar] [CrossRef]
- Liao, H.; Li, Y.; Yao, H. Biochar Amendment Stimulates Utilization of Plant-Derived Carbon by Soil Bacteria in an Intercropping System. Front. Microbiol. 2019, 10, 1361. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.N.; Sun, Y.M.; Wang, E.T.; Yang, J.S.; Yuan, H.L.; Scow, K.M. Effects of Intercropping and Rhizobial Inoculation on the Ammonia-Oxidizing Microorganisms in Rhizospheres of Maize and Faba Bean Plants. Appl. Soil Ecol. 2015, 85, 76–85. [Google Scholar] [CrossRef]
- Zhang, N.N.; Sun, Y.M.; Li, L.; Wang, E.T.; Chen, W.X.; Yuan, H.L. Effects of Intercropping and Rhizobium Inoculation on Yield and Rhizosphere Bacterial Community of Faba Bean (Vicia faba L.). Biol. Fertil. Soils 2010, 46, 625–639. [Google Scholar] [CrossRef]
- Song, Y.; Marschner, P.; Li, L.; Bao, X.; Sun, J.; Zhang, F. Community Composition of Ammonia-Oxidizing Bacteria in the Rhizosphere of Intercropped Wheat (Triticum aestivum L.), Maize (Zea mays L.), and Faba Bean (Vicia faba L.). Biol. Fertil. Soils 2007, 44, 307–314. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, F.; Marschner, P.; Fan, F.; Gao, H.; Bao, X.; Sun, J.; Li, L. Effect of Intercropping on Crop Yield and Chemical and Microbiological Properties in Rhizosphere of Wheat (Triticum aestivum L.), Maize (Zea mays L.), and Faba Bean (Vicia faba L.). Biol. Fertil. Soils 2007, 43, 565–574. [Google Scholar] [CrossRef]
- Mwakilili, A.; Mwaikono, K.S.; Herrera, S.L.; Midega, C.; Magingo, F.; Alsanius, B.; Dekker, T.; Lyantagaye, S.L. Long-Term Maize-Legume Intercropping Shifts Structure and Composition of Soil Microbiome with Potential Impacts on Ecosystem Services and Food Safety; Research Square: Durham, NC, USA, 2021. [Google Scholar] [CrossRef]
- Yu, R.-P.; Yang, H.; Xing, Y.; Zhang, W.-P.; Lambers, H.; Li, L. Belowground Processes and Sustainability in Agroecosystems with Intercropping. Plant Soil 2022, 476, 263–288. [Google Scholar] [CrossRef]
- Bilyera, N.; Zhang, X.; Duddek, P.; Fan, L.; Banfield, C.C.; Schlüter, S.; Carminati, A.; Kaestner, A.; Ahmed, M.A.; Kuzyakov, Y. Maize Genotype-Specific Exudation Strategies: An Adaptive Mechanism to Increase Microbial Activity in the Rhizosphere. Soil Biol. Biochem. 2021, 162, 108426. [Google Scholar] [CrossRef]
- Mehta, S.; Singh, B.; Patra, A.; Tripathi, A.; Easwaran, M.; Choudhary, J.R.; Choudhary, M.; Aggarwal, S. Maize Microbiome: Current Insights for the Sustainable Agriculture. In Microbiomes and Plant Health; Elsevier: Amsterdam, The Netherlands, 2021; pp. 267–297. [Google Scholar] [CrossRef]
- Andrews, M.; Andrews, M.E. Specificity in Legume-Rhizobia Symbioses. Int. J. Mol. Sci. 2017, 18, 705. [Google Scholar] [CrossRef]
- Becraft, E.D.; Woyke, T.; Jarett, J.; Ivanova, N.; Godoy-Vitorino, F.; Poulton, N.; Brown, J.M.; Brown, J.; Lau, M.C.; Onstott, T. Rokubacteria: Genomic Giants among the Uncultured Bacterial Phyla. Front. Microbiol. 2017, 8, 2264. [Google Scholar] [CrossRef]
- Jingjing, Y.; Huiqin, G.; Fry, E.L.; Jonathan, R.; Shiming, T.; Ting, Y.; Weibo, R. Plant Roots Send Metabolic Signals to Microbes in Response to Long-Term Overgrazing. Sci. Total Environ. 2022, 842, 156241. [Google Scholar] [CrossRef]
- Orata, F.D.; Meier-Kolthoff, J.P.; Sauvageau, D.; Stein, L.Y. Phylogenomic Analysis of the Gammaproteobacterial Methanotrophs (Order Methylococcales) Calls for the Reclassification of Members at the Genus and Species Levels. Front. Microbiol. 2018, 9, 3162. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Du, W.; Gu, K.; Xu, M.; Yin, Y.; Sun, Y.; Wu, J.; Zhu, J.; Guo, H. Elevated CO2 Exacerbates Effects of TiO2 Nanoparticles on Rice (Oryza sativa L.) Leaf Transcriptome and Soil Bacteria. Sci. Total Environ. 2023, 857, 159689. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, Y.; He, T.; Liao, R.; Liu, R.; Yi, M.; Huang, L.; Yang, Z.; Fu, T.; Li, X. Soil Nitrogen Leaching Decreases as Biogas Slurry DOC/N Ratio Increases. Appl. Soil Ecol. 2017, 111, 105–113. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Beletsky, A.V.; Ivanova, A.A.; Danilova, O.V.; Begmatov, S.; Kulichevskaya, I.S.; Mardanov, A.V.; Ravin, N.V. Peat-Inhabiting Verrucomicrobia of the Order Methylacidiphilales Do Not Possess Methanotrophic Capabilities. Microorganisms 2021, 9, 2566. [Google Scholar] [CrossRef] [PubMed]
- Emmett, B.D.; Lévesque-Tremblay, V.; Harrison, M.J. Conserved and Reproducible Bacterial Communities Associate with Extraradical Hyphae of Arbuscular Mycorrhizal Fungi. ISME J. 2021, 15, 2276–2288. [Google Scholar] [CrossRef] [PubMed]
- Horner, A.; Browett, S.S.; Antwis, R.E. Mixed-Cropping between Field Pea Varieties Alters Root Bacterial and Fungal Communities. Sci. Rep. 2019, 9, 16953. [Google Scholar] [CrossRef] [PubMed]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.-W. Effects of Actinobacteria on Plant Disease Suppression and Growth Promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant Beneficial Endophytic Bacteria: Mechanisms, Diversity, Host Range and Genetic Determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Hamedi, J.; Mohammadipanah, F. Biotechnological Application and Taxonomical Distribution of Plant Growth Promoting Actinobacteria. J. Ind. Microbiol. Biotechnol. 2015, 42, 157–171. [Google Scholar] [CrossRef]
- Ghodhbane-Gtari, F.; Nouioui, I.; Hezbri, K.; Lundstedt, E.; D’Angelo, T.; McNutt, Z.; Laplaze, L.; Gherbi, H.; Vaissayre, V.; Svistoonoff, S. The Plant-Growth-Promoting Actinobacteria of the Genus Nocardia Induces Root Nodule Formation in Casuarina Glauca. Antonie Van Leeuwenhoek 2019, 112, 75–90. [Google Scholar] [CrossRef]
- Sousa, J.A.d.J.; Olivares, F.L. Plant Growth Promotion by Streptomycetes: Ecophysiology, Mechanisms and Applications. Chem. Biol. Technol. Agric. 2016, 3, 24. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria Associated with a Commercial Mycorrhizal Inoculum: Community Composition and Multifunctional Activity as Assessed by Illumina Sequencing and Culture-Dependent Tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef] [PubMed]
- McBride, M. The Family Flavobacteriaceae. In The Prokaryotes: Other Major Lineages of Bacteria and the Archaea; Rosenberg, E., DeLong, E., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 643–676. [Google Scholar] [CrossRef]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.L.; Bouffaud, M.-L.; Buée, M.; Dimmers, W. Soil Networks Become More Connected and Take up More Carbon as Nature Restoration Progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heijden, M.G.; Hartmann, M. Networking in the Plant Microbiome. PLoS Biol. 2016, 14, e1002378. [Google Scholar] [CrossRef] [PubMed]
- Stopnisek, N.; Shade, A. Persistent Microbiome Members in the Common Bean Rhizosphere: An Integrated Analysis of Space, Time, and Plant Genotype. ISME J. 2021, 15, 2708–2722. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, M.; Huang, S.; Li, L.; Gao, Q.; Wang, Y.; Zhang, S.; Huang, S.; Yuan, L.; Wen, Y. A Highly Conserved Core Bacterial Microbiota with Nitrogen-Fixation Capacity Inhabits the Xylem Sap in Maize Plants. Nat. Commun. 2022, 13, 3361. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and Significance of Burkholderia Species Occupying Diverse Ecological Niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef]
- Li, Y.; Pan, F.; Yao, H. Response of Symbiotic and Asymbiotic Nitrogen-Fixing Microorganisms to Nitrogen Fertilizer Application. J. Soils Sediments 2019, 19, 1948–1958. [Google Scholar] [CrossRef]
- Kumar, U.; Nayak, A.K.; Shahid, M.; Gupta, V.V.; Panneerselvam, P.; Mohanty, S.; Kaviraj, M.; Kumar, A.; Chatterjee, D.; Lal, B. Continuous Application of Inorganic and Organic Fertilizers over 47 Years in Paddy Soil Alters the Bacterial Community Structure and Its Influenceon Rice Production. Agric. Ecosyst. Environ. 2018, 262, 65–75. [Google Scholar] [CrossRef]
- Anzuay, M.S.; Viso, N.P.; Ludueña, L.M.; Morla, F.D.; Angelini, J.G.; Taurian, T. Plant Beneficial Rhizobacteria Community Structure Changes through Developmental Stages of Peanut and Maize. Rhizosphere 2021, 19, 100407. [Google Scholar] [CrossRef]
- Sun, B.; Gu, L.; Bao, L.; Zhang, S.; Wei, Y.; Bai, Z.; Zhuang, G.; Zhuang, X. Application of Biofertilizer Containing Bacillus subtilis Reduced the Nitrogen Loss in Agricultural Soil. Soil Biol. Biochem. 2020, 148, 107911. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H.; Cheng, Q. Nitrogen Fixation and Diazotrophs—A Review. Rom. Biotechnol. Lett 2021, 26, 2834–2845. [Google Scholar] [CrossRef]
- Nouioui, I.; Cortés-Albayay, C.; Carro, L.; Castro, J.F.; Gtari, M.; Ghodhbane-Gtari, F.; Klenk, H.-P.; Tisa, L.S.; Sangal, V.; Goodfellow, M. Genomic Insights into Plant-Growth-Promoting Potentialities of the Genus Frankia. Front. Microbiol. 2019, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using Network Analysis to Explore Co-Occurrence Patterns in Soil Microbial Communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Widder, S. Deciphering Microbial Interactions and Detecting Keystone Species with Co-Occurrence Networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef]
- de Vries, F.T.; Griffiths, R.I.; Bailey, M.; Craig, H.; Girlanda, M.; Gweon, H.S.; Hallin, S.; Kaisermann, A.; Keith, A.M.; Kretzschmar, M. Soil Bacterial Networks Are Less Stable under Drought than Fungal Networks. Nat. Commun. 2018, 9, 3033. [Google Scholar] [CrossRef]
- Jacquiod, S.; Puga-Freitas, R.; Spor, A.; Mounier, A.; Monard, C.; Mougel, C.; Philippot, L.; Blouin, M. A Core Microbiota of the Plant-Earthworm Interaction Conserved across Soils. Soil Biol. Biochem. 2020, 144, 107754. [Google Scholar] [CrossRef]
- Bitocchi, E.; Nanni, L.; Rossi, M.; Bellucci, E.; Papa, R. Exploring Molecular Variation in Maize Landraces from Central Italy. In Proceedings of the 2nd International Symposium on Genomics of Plant Genetic Resources, Bologna, Italy, 24–27 April 2010. [Google Scholar]
- Giupponi, L.; Leoni, V.; Colombo, F.; Cassani, E.; Hejna, M.; Rossi, L.; Pilu, R. Characterization of “Mais delle Fiorine” (Zea mays L.) and nutritional, morphometric and genetic comparison with other maize landraces of Lombardy region (Northern Italy). Genet. Resour. Crop. Evol. 2021, 68, 2075–2091. [Google Scholar] [CrossRef]
- Bressan, M. Cereali del Veneto: Le Varietá di Frumento Tenero e Mais della Tradizione Veneta; Regione Veneto: Provincia di Vicenza, Italy, 2003. [Google Scholar]
- Mais Ottofile di Roccacontrada. Available online: https://portalecustodibiodiversita.it/prodotto.php?id=80 (accessed on 1 January 2018).
- Cortinovis, G.; Oppermann, M.; Neumann, K.; Graner, A.; Gioia, T.; Marsella, M.; Alseekh, S.; Fernie, A.R.; Papa, R.; Bellucci, E. Towards the Development, Maintenance, and Standardized Phenotypic Characterization of Single-seed-descent Genetic Resources for Common Bean. Curr. Protoc. 2021, 1, e133. [Google Scholar] [CrossRef]
- Bellucci, E.; Benazzo, A.; Xu, C.; Bitocchi, E.; Rodriguez, M.; Alseekh, S.; Di Vittori, V.; Gioia, T.; Neumann, K.; Cortinovis, G. Selection and Adaptive Introgression Guided the Complex Evolutionary History of the European Common Bean. Nat. Commun. 2023, 14, 1908. [Google Scholar] [CrossRef] [PubMed]
- Team, R.D.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2010. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Anderson, M.J. Distance-based Redundancy Analysis: Testing Multispecies Responses in Multifactorial Ecological Experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Garrido-Oter, R.; Münch, P.C.; Weiman, A.; Dröge, J.; Pan, Y.; McHardy, A.C.; Schulze-Lefert, P. Structure and Function of the Bacterial Root Microbiota in Wild and Domesticated Barley. Cell Host Microbe 2015, 17, 392–403. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N.; Stine, O.C.; Bravo, H.C.; Pop, M. Differential Abundance Analysis for Microbial Marker-Gene Surveys. Nat. Methods 2013, 10, 1200–1202. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; Carrión, V.J.; Bosse, M.; Ferrão, L.F.; De Hollander, M.; Garcia, A.A.; Ramírez, C.A.; Mendes, R.; Raaijmakers, J.M. Linking Rhizosphere Microbiome Composition of Wild and Domesticated Phaseolus Vulgaris to Genotypic and Root Phenotypic Traits. ISME J. 2017, 11, 2244–2257. [Google Scholar] [CrossRef]
- Friedman, J.; Alm, E.J. Inferring Correlation Networks from Genomic Survey Data. PLoS Comput. Biol. 2012, 8, e1002687. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phyla | Bacterial Genera | M_SC | IC |
|---|---|---|---|
| Acidobacteria | Holophagae_Subgroup7_uncultured bacterium gp7 | 0 | 9 |
| Subgroup5_uncultured Acidobacterium sp. | 4 | 18 | |
| Subgroup6_unculture microorganism | 2 | 30 | |
| Thermoanaerobaculia_Thermoanaerobaculiales_Thermoanaerobaculaceae_Sub10 | 2 | 20 | |
| Acidimicrobiaa_Micotrichales_Ilumatobacteraceae_uncultured bacterium | 8 | 44 | |
| Actinobacteria | Actinobacteria_Elev-16S-976_uncultured bacterium | 1 | 2 |
| Actinobacteria_Frankiales_uncultured | 9 | 59 | |
| Actinobacteria_Micromonosporales_Micromonosporaceae_Actinorabdospora | 2 | 17 | |
| Actinobacteria_Propionibacteriales_Propionibacteraceae_Haloactinopolyspora | 1 | 6 | |
| Actinobacteria_Streptosporangiales_Thermomonosporaceae_Actinocoralia | 2 | 11 | |
| Nitriliruptoria_Euzebyales_Euzebyaceae_uncultured | 12 | 80 | |
| Thermoleophilia_Solitubrobacterales_67-14_uncultiured Rubrobacteraceae | 3 | 17 | |
| Armatimonadetes | Uncultured bacterium_#0319-6E2 | 1 | 28 |
| Bacteroidetes | Bacteroidia_Chitinophagales_Chitinophagaceae_Vibrionimonas | 1 | 22 |
| Ignavibacteria_OPB56_uncultured bacterium_#0319-6E22 | 5 | 8 | |
| Anaerolinae_R8G-13-54-9_uncultured bacterium | 2 | 30 | |
| Chloroflexi | TK10_bacterium Ellin6543_bacterium Ellis6543 | 2 | 67 |
| Cyanobacteria | Oxyphotobacteria_Chloroplast_Trifolium pratense | 1 | 19 |
| Elusimicrobia | Lineage 1b | 0 | 11 |
| Firmicutes | Bacill_Bacillales_Bacillaceae | 3 | 14 |
| Planctomycetes | VadinHA49_uncultured bacterium | 2 | 12 |
| Proteobacteria | Alphaproteobacteria_Rhizobiales_Beijerinckiaceae | 20 | 97 |
| Alphaproteobacteria_Rhizobiales_KF-JG30-B3 | 20 | 81 | |
| Alphaproteobacteria_Rhizobiales_Rhizobiales Incertae Sedis_Bauldia | 4 | 27 | |
| Alphaproteobacteria_Tistrellales_Germinicoccaceae_Candidatus Alysiosphaera | 17 | 77 | |
| Deltaproteobacteria_Bdellovibrionales_Bdellovibionaceae_Bdellovibrio | 5 | 48 | |
| Delaproteobacteria_Myxococcales_Bfdi19 | 1 | 1 | |
| Deltaproteobacteria_Myxococcales_uncultered | 0 | 40 | |
| Gammaproteobacteria_Betaproteobacteriales_TRA3-20_uncultured Alcaligenaceae | 3 | 31 | |
| Gammaproteobacteria_Salinisphaerales_Solimonadaceae | 8 | 11 |
| Source | Df | Sum of Sqs | Mean Sqs | F Model | R2 | Pr (>F) |
|---|---|---|---|---|---|---|
| Maize Spinato (S) | ||||||
| Plant status | 2 | 0.058 | 0.029 | 1.316 | 0.066 | 0.144 |
| Accession | 8 | 0.250 | 0.031 | 1.410 | 0.283 | 0.05 * |
| Residuals | 26 | 0.577 | 0.022 | 0.651 | ||
| Total | 36 | 0.886 | 1.00000 | |||
| Maize Ibrido (I) | ||||||
| Plant status | 2 | 0.072 | 0.036 | 1.516 | 0.088 | 0.084 |
| Accession | 8 | 0.249 | 0.031 | 1.305 | 0.303 | 0.116 |
| Residuals | 21 | 0.500 | 0.024 | 0.609 | ||
| Total | 31 | 0.821 | 1 | |||
| Maize Ottofile (O) | ||||||
| Plant status | 2 | 0.048 | 0.024 | 1.242 | 0.061 | 0.216 |
| Accession | 7 | 0.255 | 0.036 | 1.864 | 0.322 | 0.004 ** |
| Residuals | 25 | 0.488 | 0.020 | 0.617 | ||
| Total | 34 | 0.791 | 1 | |||
| Maize Biancoperla (BP) | ||||||
| Plant status | 2 | 0.045 | 0.023 | 1.061 | 0.054 | 0.357 |
| Accession | 9 | 0.290 | 0.032 | 1.503 | 0.341 | 0.02 * |
| Residuals | 24 | 0.515 | 0.021 | 0.605 | ||
| Total | 35 | 0.85 | 1 |
| Acronyms | Species | Description | Origin | Common Name | Genetic Material |
|---|---|---|---|---|---|
| p83 | P. vulgaris | Landrace | Italy | - | Single Seed Descent (SSD) |
| p91 | P. vulgaris | Landrace | Greece | - | Single Seed Descent (SSD) |
| pC | P. coccineus | Landrace | Italy | Fagiolo di Clusven | Single Seed Descent (SSD) |
| pM | P. vulgaris | Landrace | Italy | Monachello | Single Seed Descent (SSD) |
| pMNE2 | P. vulgaris | Landrace | Montenegro | - | Single Seed Descent (SSD) |
| S | Zea mays | Landrace | Italy | Spinato | Multiplied by custodian farmers |
| BP | Zea mays | Landrace | Italy | Biancoperla | Multiplied by custodian farmers |
| O | Zea mays | Landrace | Italy | Ottofile | Multiplied by custodian farmers |
| I | Zea mays | Commercial hybrid | - | Ibrido (tipo Marano) | Multiplied by custodian farmers |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lanzavecchia, G.; Frascarelli, G.; Rocchetti, L.; Bellucci, E.; Bitocchi, E.; Di Vittori, V.; Sillo, F.; Ferraris, I.; Carta, G.; Delledonne, M.; et al. Genotype Combinations Drive Variability in the Microbiome Configuration of the Rhizosphere of Maize/Bean Intercropping System. Int. J. Mol. Sci. 2024, 25, 1288. https://doi.org/10.3390/ijms25021288
Lanzavecchia G, Frascarelli G, Rocchetti L, Bellucci E, Bitocchi E, Di Vittori V, Sillo F, Ferraris I, Carta G, Delledonne M, et al. Genotype Combinations Drive Variability in the Microbiome Configuration of the Rhizosphere of Maize/Bean Intercropping System. International Journal of Molecular Sciences. 2024; 25(2):1288. https://doi.org/10.3390/ijms25021288
Chicago/Turabian StyleLanzavecchia, Giovanna, Giulia Frascarelli, Lorenzo Rocchetti, Elisa Bellucci, Elena Bitocchi, Valerio Di Vittori, Fabiano Sillo, Irene Ferraris, Giada Carta, Massimo Delledonne, and et al. 2024. "Genotype Combinations Drive Variability in the Microbiome Configuration of the Rhizosphere of Maize/Bean Intercropping System" International Journal of Molecular Sciences 25, no. 2: 1288. https://doi.org/10.3390/ijms25021288