
New Species and New Records of Sponge-Inhabiting Barnacles (Cirripedia, Balanidae, Acastinae) from Australia


1
Collections and Research, Western Australian Museum, 49 Kew St, Welshpool, WA 6106, Australia
2
Curtin Medical School, Curtin University, Bentley, WA 6102, Australia
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(7), 290; https://doi.org/10.3390/d13070290
Submission received: 11 May 2021
/
Revised: 17 June 2021
/
Accepted: 17 June 2021
/
Published: 27 June 2021
(This article belongs to the Special Issue Australian Marine Invertebrate Biodiversity)
Abstract
:The subfamily Acastinae contains a diverse group of barnacles that are obligate symbionts of sponges and alcyonacean and antipatharian corals. Integrating morphological and genetic (COI) data to compare against known species, this paper reports on nine species of sponge-inhabiting barnacles of the subfamily Acastinae, including three undescribed species (Acasta caveata sp. nov., Euacasta acutaflava sp. nov., and E. excoriatrix sp. nov.) and three species previously not recorded in Australian waters (A. sandwichi, Pectinoacasta cancellorum, and P. sculpturata). The new species are distinguished from similar species by a suite of morphological characters as well as genetic distances. A lectotype for Pectinoacasta cancellorum is designated. Sponge hosts were identified for all specimens where possible and are represented by 19 species from eight families and five orders.
1. Introduction
Barnacles can be found as obligate commensals with a variety of taxa, but those in symbiosis with sponges are proving to be particularly diverse [1,2,3,4]. The majority of sponge symbionts are classified under the subfamily Acastinae [5], with approximately 90 described species, and this number is expected to at least double [1,4]. Historically, this group had been arranged within informal groups based on characters of the fourth cirrus by Broch [6] and opercular plates by Hiro [7] under the genus Acasta sensu Leach, 1817 [8]. Neither author took this further, largely due to the close affinity with Conopea Say, 1812 [9], and Membranobalanus Hoek, 1913 [10], whose members are obligate commensals with cnidarians and sponges, respectively, but also Armatobalanus [10], which contained a mixture of free-living and commensal species. This caution has proven to be warranted, as a number of species have been transferred between these genera by subsequent workers [11]. Further to this, phylogenetic studies have also shown that the subfamily and family level arrangement is also unsupported within the Balanoidea [12,13]; as a result, the family Archaeobalanidae was recently synonymised with the Balanidae [14]. Kolbasov [5], in his revision of Acasta, was the first to evaluate host usage in the Acasta and how that was reflected in his newly proposed phylogeny and generic arrangement, which revealed some loose trends, but was hampered by a distinct lack of previously published host data.
Since then, more studies on sponge-inhabiting barnacles have been placing emphasis on host identification [1,2,3,15,16] to build a better understanding of host usage. Not only have the evolutionary relationships of the hosts begun to be looked at, but also the morphology of the host has been examined to test larval barnacle biology [17]. Integrating modern techniques such as genetic sequencing and computed tomography [18,19] has also provided greater precision when evaluating inter- and intra-specific variation by shedding light on character evolution.
The waters surrounding the Australian coast are inhabited by a highly diverse sponge fauna [20,21,22], leading to an expectation of a correspondingly diverse community of sponge-inhabiting barnacles. This paper reports on a collection of sponge-inhabiting barnacles from across Australian territorial waters as part of a broader study on the biodiversity of commensal barnacles.
2. Materials and Methods
2.1. Sampling and Morphological Examination
Specimens of both the barnacles and the sponges examined in this study are deposited in the following institutions: Western Australian Museum, Perth (WAM), Queensland Museum, Brisbane (QM), University of Copenhagen Zoological Museum, Denmark (ZMUC), and Seto Marine Biological Laboratory, Japan (SMBL). For direct morphological examination of barnacle shell plates and arthropodal characters, the body and associated soft tissues were removed from the shell. The remnants of the barnacle tissue and host sponge on the surfaces of the parietes, scutum and tergum, were removed using a brush and forceps. The shell was then immersed in 2% bleach for <2 h to completely digest the organic tissue and was subsequently rinsed in water. Any remaining debris or contaminants were then removed by cleaning in an ultrasonic cleaner for < 5 s. The specimens were examined under a Leica M205 C (Leica, Germany) stereomicroscope and digital photographs produced with a Leica DMC4500. All images and figures were processed using GIMP 2.10 (www.gimp.org accessed on 20 June 2021) or Inkscape 0.92 (www.inkscape.org accessed on 20 June 2021). Barnacle size is measured in millimetres along the rostro-carinal (RC) axis at the basal rim. Setal terminology follows that of Chan et al. [23].
Sponge identifications were partly facilitated by examining gross morphology, surface characteristics, and microscopic skeletal characteristics of specimens. Subsamples of sponge tissue were sectioned at right angles to the surface of the sponge and processed for microscopic examination using a Shandon Elliott tissue processor. This included ethanol dehydration, histolene to clear the tissue, and subsequent paraffin impregnation. Thick sections (~90 μm) were cut with a Leitz slide microtome from wax blocks. Sections were de-waxed in histolene and mounted using Shandon EZ-Mountant. Skeletal slides were examined with an Olympus BX50.
2.2. Molecular Analysis
Adductor or depressor muscle tissues of barnacles were subsampled from specimens, and genomic DNA was extracted using either a Bioline Isolate II or Qiagen DNeasy extraction kit following the manufacturer’s instructions. Partial fragments of the cytochrome c oxidase I (COI) gene were amplified using the primers dgLCO1490 5’-GTCAACAAATCATAAAGAYATYGG-3’ and dgHCO2198 5’- TAAACTTCAGGGTGACCAAARAAYCA-3’ [24] in a 25 µL reaction volume and consisting of 2 µL DNA extract, 1 unit MyTaq DNA polymerase, MyTaq PCR buffer, and 0.1 µM of each primer. The following polymerase chain reaction conditions were used: 2 min at 95 °C for initial denaturing, then 35 cycles of 30 s at 95 °C, 30 s at 46 °C, 45 s at 72 °C, and a final extension for 7 min at 72 °C. The resulting amplicons were sequenced at the Australian Genome Research Facility, Perth, using the same primers via Sanger (cycle) sequencing.
The sequences were assembled and trimmed using Geneious Prime and submitted to GenBank (Table 1, https://www.ncbi.nlm.nih.gov/genbank/ accessed on 20 June 2021). Quality control of sequences involved trimming low-quality bases, inspecting for stop codons, verifying the reading frame, checking for matches on BLAST, and re-examining original specimens.
Representative COI sequences identified as belonging to the genera studied herein were downloaded from GenBank and BoLD (www.boldsystems.org) (Table 1). Membranobalanus porphyrophilus Hosie and Jones, 2019 [18], was selected to serve as an outgroup in the phylogenetic analyses. The combined dataset was aligned in Geneious Prime 2020.2.5 (https://www.geneious.com) and checked manually. The Assemble Species by Automatic Partitioning (ASAP; [25]) software was used to compare the genetic data with the morphological identifications by delimiting putative species under three substitution models: Jukes-Cantor (JC69) [26], Kimura 2-parameter model (K2P) [27], and uncorrected p-distance. Pairwise genetic distances were calculated using the software MEGA v10 [28]. Phylogenetic reconstructions using a maximum likelihood analysis were conducted using the IQ-TREE webserver [29]. Model selection was automatically assessed by the webserver, with the COI data partitioned by codon, and branch support was calculated by conducting 1000 ultra-fast bootstraps [30]. Bayesian inference analyses were conducted in MrBayes 3.2.6 [31] utilising the CIPRES webserver [32] with four heated chains, a General Time Reversible model with proportion of invariable sites and rate heterogeneity across sites (GTR+I+G), an MCMC of 11 million generations, and a burn-in of 10%.
3. Results
3.1. Systematics
Balanomorpha Pilsbry, 1916 [34]
Balanidae Leach, 1817 [8]
Acastinae Kolbasov, 1993 [5]
3.1.1. Acasta Leach, 1817 [8]
Acasta aspera Yu et al., 2017 [3]
Acasta aspera Yu et al., 2017: 155, Figures 1–10 [3]
Material examined: WAM C58326, WAM C58835–8, all from NE of Ah Chong Island, Montebello Islands, WA, 20°29′57″ S 115°35′23″ E, 14.5 m, NCB Muirons and Montebellos April 2015 Expedition, stn. 6/MM15, coll. A.M. Hosie & A. Hara, host: WAM Z84165 Hamigera PB1. WAM C58332, WAM C58812–15, WAM C71837–8, all from NE of Ah Chong Island, Montebello Islands, WA, 20°29′57″ S 115°35′23″ E, 14.5 m, NCB Muirons and Montebellos Expedition, stn. 6/MM15, 16 Apr 2015, coll. A.M. Hosie & A. Hara, host: WAM Z84171 Lissodendoryx (Acanthodoryx) KMB1.
Diagnosis: Shell pinkish or yellow-brown, orifice edge toothed. Basis cup-shaped with crenate edge. Radii with creased edges. Externally, parietes rugged with several irregularly spaced calcareous projections; internally, sheath extending over half of parietes with vesicular structure, inner surface below sheath with longitudinal ribs. Scutum with feeble growth ridges and longitudinal striations, articular furrow shallow. Tergum beaked, apex tinged with red, spur truncated, width less than 0.5 of basal margin. Curved teeth on anterior ramus of cirrus IV feeble. Cirrus V protopod with vertical row of denticles on posterior edge.
Distribution: Australia: Perth—Montebello Islands, WA. Taiwan [3]. 0–16 m.
Hosts: Lissodendoryx (Acanthodoryx) KMB1 (Coelosphaeridae), and Hamigera PB1 (Hymedesmiidae). Previous records: Crella (Yvesia) spinulata (Hentschel, 1911) [35] (Crellidae), and Iotroata sp. [3] (Iotrochotidae).
Remarks: This species was originally identified inhabiting Iotroata sp. and Crella (Yvesia) spinulata from the central western coast of Australia and Taiwan, respectively. Here, we report two additional host species from the families Coelosphaeridae and Hymedesmiidae, demonstrating that A. aspera is a relative generalist, capable of colonising sponge species from several families within the order Poecilosclerida.
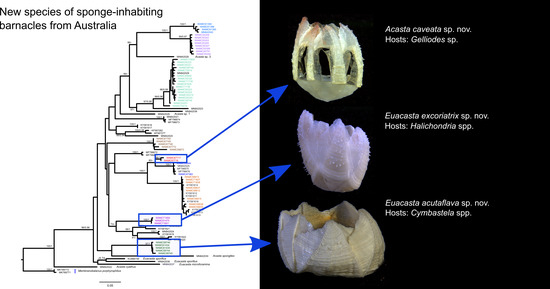
Acasta caveata sp. nov. (Figure 1, Figure 2, Figure 3 and Figure 4)
Material examined: Holotype: WAM C67716, 5.5 mm RC, 3 slides, S of Sponge Garden, NW side of Exmouth Gulf, WA, 21°49′15″ S 114°12′34″ E, 22 m, NCB Exmouth/Muiron Expedition, stn. 14/EM16, 6 Jun 2016, coll. A.M. Hosie & A. Hara, Host: WAM Z97140 Gelliodes KMB1.
Paratype: WAM C67717, 5.2 mm RC, same data as holotype.
Other material: WAM C66772, specimens still with host, same data as holotype.
Diagnosis: Shell fenestrate; very large, subrectangular windows present between each parietal junction, occupying over half of parietal height, as wide as or wider than adjacent parietes at basal rim. Parietes with blunt, calcareous projections, spaced irregularly in longitudinal rows. Basis saucer-shaped, rim undulating, crenate where it interlocks with parietes, otherwise smooth. Cirrus IV protopod with single horizontal tooth, anterior and posterior rami with hooked teeth arranged in triangular shape. Cirrus V anterior ramus with hooked teeth. Penis basidorsal point prominent.
Description: Shell (Figure 1A–M) erect, slightly elongate vertically, plates semitransparent, white or tinged red or burgundy near apices. Externally, parietes with fine, horizontal growth lines and short, irregularly spaced spines; apices of parietes curved toward orifice. Radii and alae summits oblique, not reaching basal margin of corresponding parietes, leaving elongate, subrectangular membranous windows between parietes. With ontogeny, windows secondarily filled with calcareous shell, maintaining lower margin approximately level with basal margin of sheath. Internally, sheath adpressed with vesicular structure, faint horizontal growth lines, lower margin separated from inner lamina except on carina; inner lamina with weak longitudinal ribs extending to basal rim, lateral margins inflected or thickened.
Basis (Figure 1H,M) saucer-shaped, making up approximately a quarter of total shell height, with six wide and shallow longitudinal furrows corresponding to the bases of parietes; rim undulating, crenate where interlocking with parietes, inter-parietal rim concave, smooth.
Scutum (Figure 1N–P) semitransparent, triangular, approximately as high as wide, growth lines prominent with fine crenulations and fringed with short, inconspicuous setae; basitergal angle broadly rounded; occludent margin toothed. Internally smooth, adductor muscle pit weakly defined, lateral depressor muscle pit well defined; articular ridge low, parallel with tergal margin, with shallow articular furrow, extending approximately 0.75 length of tergal margin.
Tergum (Figure 1Q,R) semitransparent, apex acute, tinged with red; external growth lines fringed with short inconspicuous setae; articular margin concave in apical half, carinal margin evenly arcuate, basal margin slightly sinuous on carinal side of spur; scutal side straight, but obtuse to, articular margin; spur length approximately twice width, occupying approximately half of basal margin, positioned approximately half own width from scutal margin; spur and furrow margins coincident, defined by abrupt change in growth lines and corresponding groove on articular side and gentle change on carinal side; distal margin convex, obliquely truncate. Internally smooth, crests for depressor muscles faint or absent.
Labrum (Figure 2A,B) bilobed, lobes separated by deep, medial, v-shaped notch, each rounded lobe with two or three marginal teeth and numerous fine setae.
Mandibular palp (Figure 2A,C) rhomboid, apex obliquely truncate, anterior margin slightly concave, almost straight, posterior margin straight; setae heavily serrulate, becoming longer and denser distally; distal angle with long, simple setae.
Mandible (Figure 2D–F) with five teeth, second to third teeth bifid, fourth and fifth teeth serrate of moloriform, inferior angle with denticles. Dense, short setae regularly spaced on inner and outer faces, longer, fine setae on inferior and superior margin.
Maxillule (Figure 2G,H) cutting margin straight, with 11 cuspidate setae, upper and lower pairs longer than remaining setae, medial setae serrulate on inferior margin. Dense, short setae regularly spaced on inner and outer faces, longer, fine setae on inferior and superior margin.
Maxilla (Figure 2I) bilobed, lobes ovate, distal lobe elongate, curved towards mouth, serrulate setae arranged on anterior margins, becoming more dense at apex, longer on distal lobe than those on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 3A) protopod with pappose setae basally on posterior margin. Rami unequal, anterior ramus approximately 2.3 times longer than posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus II (Figure 3B) protopod anterior margin lined with long, plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami unequal, anterior ramus approximately 1.25 times longer than posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus III (Figure 3C–E) protopod anterior margin lined with long, plumose setae, posterior margin with pappose setae basally and plumose setae distally. Rami unequal, anterior ramus approximately 1.25 times longer than posterior ramus, setation lasiopod, segments with serrulate and simple setae on mesial face, less densely arranged than cirrus I and II. Anterior ramus with row of sharp, erect spines on anterodistal portion of all segments, except terminal segment, most prominent on medial segments. Posterior ramus similar but spines smaller.
Cirrus IV (Figure 4A–E) protopod elongate, length 1.8 times width, with simple setae on anterior and posterior margins, tuft of simple setae at posterodistal angles of both segments; basis with single tooth at anterodistal angel. Rami subequal, setation ctenopod, segments with 3 pairs of serrulate setae on anterior margin. Anterior ramus with up to 5 hooked teeth arranged in a rough triangular formation on anterior margin of first 15 segments, anterodistal angle with group of erect spines. Posterior ramus with 1 or 2 hooked teeth on segments 10–21, anterodistal angle with group of erect spines.
Cirrus V (Figure 4F–H) protopod elongate, length 1.38 times width, with simple setae on anterior and posterior margins, anterodistal angle unarmed, tuft of simple cuspidate setae at posterodistal angles of both segments. Rami subequal, setation ctenopod, anterior ramus with 3 and posterior ramus with 4 pairs of serrulate setae on anterior margin, posterior ramus with four and both rami of cirrus VI with 4 pairs of serrulate setae; posterodistal angles with tuft of simple setae. Anterior ramus with 1 or 2 hooked teeth on segments 13–21.
Cirrus VI (Figure 4I) protopod elongate, length 1.4 times width simple setae on anterior and posterior margins, lacking denticles or erect spines on anterodistal angles, tuft of simple cuspidate setae at posterodistal angles of both segments. Rami subequal, setation ctenopod, with 4 pairs of serrulate setae; posterodistal angles with tuft of simple setae; anterior margins without teeth or erect spines.
Penis (Figure 4I,J) longer than cirrus VI, annulated, sparsely setose; basidorsal point triangular, prominent, apex rounded.
Hosts: Gelliodes KMB1 (Niphatidae).
Distribution: Western Australia: Exmouth Gulf. 23 m.
Etymology: The specific epithet is derived from the Latin noun cavea (cage, hollow, cavity), in reference to the large membrane-covered windows that give the shell a cage-like appearance.
Remarks: This species, with its very conspicuous membranous windows, is externally similar to a number of species of Acasta. The most similar are A. sandwichi Yu et al. 2017 [2] and A. pertusa Kolbasov 1990 [36], which exhibit close morphological similarities in the form of the windows and opercular plates. The external shell and opercular plates are nearly identical, but with some potential differences in the relative widths of the windows and adjacent parietes. The windows of A. caveata sp. nov. occupy more than half the shell height (excluding basis) and are level with, or just above, the lower margin of the sheath, thus larger than in Acasta sandwichi and A. pertusa where the margin is distinctly lower than the sheath. In A. caveata, the free, lower margins of the parietes are parallel with the basis, giving the windows a more rectangular shape, whereas in A. pertusa and A. sandwichi, they are curved, giving them a more arched look. The key arthropodal character to distinguish these species is the armature of cirri IV and V. For A. caveata, both the anterior and posterior rami of cirrus IV and the anterior ramus of cirrus V bear strong hooked teeth, whereas only a single, straight “denticle” is found on each segment of the anterior ramus of cirrus IV in A. sandwichi, while such armature is completely absent in A. pertusa.
Of the other species with conspicuous windows (A. armata Gravier, 1921 [37], A. crucibasis Yu et al., 2017 [2], A. fenestrata Darwin, 1854 [38], A. foraminifera Broch, 1931 [6], and A. tzetlini Kolbasov, 1992 [39]), only A. tzetlini bears teeth, but only on cirrus IV anterior ramus, and it has much smaller windows with the parietes articulated with the basis over most of its circumference. In addition to lacking teeth on cirrus IV, A. armata and A. fenestrata lack longitudinal ribs on the inner lamina of the parietes; A. armata also has very distinct opercular plates, and the windows of A. fenestrata are significantly smaller. The carinolatus of A. foraminifera and A. crucibasis do not reach the basis at all but are instead suspended between the latus and the carina.
Figure 1.
Acasta caveata sp. nov. shell plates (all WAM C 67716, holotype). (A–C) right, rostral, and left external views, respectively, of whole specimen; (D–H) external views of rostrum, right lateral, right carinolateral, carina, and basis; (I–L) internal views of rostrum, right lateral, right carinolateral, and carina; (M) close up of carinal portion of basis rim; (N–O) external and internal view of right scutum; (P) close up of growth lines on scutum showing setae; (Q–R) internal and external views of right tergum. Scale bars: A–L = 2 mm; M, P = 0.5 mm; N–O, Q–R = 1 mm.
Figure 1.
Acasta caveata sp. nov. shell plates (all WAM C 67716, holotype). (A–C) right, rostral, and left external views, respectively, of whole specimen; (D–H) external views of rostrum, right lateral, right carinolateral, carina, and basis; (I–L) internal views of rostrum, right lateral, right carinolateral, and carina; (M) close up of carinal portion of basis rim; (N–O) external and internal view of right scutum; (P) close up of growth lines on scutum showing setae; (Q–R) internal and external views of right tergum. Scale bars: A–L = 2 mm; M, P = 0.5 mm; N–O, Q–R = 1 mm.

Figure 2.
Acasta caveata sp. nov. mouthparts (all WAM C67716, holotype). (A) labrum and right mandibular palp; (B) crest of labrum; (C) left mandibular palp; (D) left mandible; (E) close up of inferior angle of mandible showing fourth and fifth serrated teeth; (F) right mandible; (G) left maxillule; (H) right maxillule; (I) left and right maxilla, lateral view. Scale bars: A, C–D, F–I = 200 µm; B, E = 50 µm.
Figure 2.
Acasta caveata sp. nov. mouthparts (all WAM C67716, holotype). (A) labrum and right mandibular palp; (B) crest of labrum; (C) left mandibular palp; (D) left mandible; (E) close up of inferior angle of mandible showing fourth and fifth serrated teeth; (F) right mandible; (G) left maxillule; (H) right maxillule; (I) left and right maxilla, lateral view. Scale bars: A, C–D, F–I = 200 µm; B, E = 50 µm.

Figure 3.
Acasta caveata sp. nov. cirri I–III (all WAM C67716, holotype). (A) left cirrus I; (B) left cirrus II; (C) left cirrus III; (D) anterior margin of medial segments of left cirrus III anterior ramus; (E) anterior margin of medial segments of right cirrus III anterior ramus. Scale bars: A–C = 200 µm, D–E = 50 µm.
Figure 3.
Acasta caveata sp. nov. cirri I–III (all WAM C67716, holotype). (A) left cirrus I; (B) left cirrus II; (C) left cirrus III; (D) anterior margin of medial segments of left cirrus III anterior ramus; (E) anterior margin of medial segments of right cirrus III anterior ramus. Scale bars: A–C = 200 µm, D–E = 50 µm.

Figure 4.
Acasta caveata sp. nov. cirri IV–VI (all WAM C67716, holotype). (A) left cirrus IV; (B) hooked teeth on medial segments of left cirrus IV anterior ramus; (C) close up of tooth on basis of right cirrus IV protopod; (D) medial segments of cirrus IV; (E) detail of erect spines on medial segments of cirrus IV anterior ramus; (F) cirrus V; (G) setae on medial segments of cirrus V; (H) hooked teeth on medial segments of cirrus V anterior ramus; (I) cirrus VI and penis; (J) basidorsal point of penis. Scale bars: A, F, I = 300 µm; B, D = 100 µm; C, E = 50 µm; J = 200 µm.
Figure 4.
Acasta caveata sp. nov. cirri IV–VI (all WAM C67716, holotype). (A) left cirrus IV; (B) hooked teeth on medial segments of left cirrus IV anterior ramus; (C) close up of tooth on basis of right cirrus IV protopod; (D) medial segments of cirrus IV; (E) detail of erect spines on medial segments of cirrus IV anterior ramus; (F) cirrus V; (G) setae on medial segments of cirrus V; (H) hooked teeth on medial segments of cirrus V anterior ramus; (I) cirrus VI and penis; (J) basidorsal point of penis. Scale bars: A, F, I = 300 µm; B, D = 100 µm; C, E = 50 µm; J = 200 µm.

Acasta fenestrata Darwin, 1854 [38] (Figure 5, Figure 6, Figure 7 and Figure 8)
Acasta fenestrata Darwin, 1854: p. 316, Tab. 9, Figure 7 [38]. Weltner 1897: p. 259 [40]. 1922, p. 104 [41]. Nilsson-Cantell 1938: p. 57 [42]. Hiro, 1939: p. 243 [43].Newman & Ross 1976: p. 53 [44]. Ren 1984 [45]. Rosell 1989: p. 33, Pl. 9b [46]. Liu & Ren 2007: p. 417, Figure 187 [47]. Wibowo et al. 2011: p. 227, Figures 10–13 [48]. Sulistiono et al. 2014: p. 182, Figure 4 [49]. Jones & Hosie 2016: p. 275 [50].
Acasta sp.4 Yu et al., 2020 [17]
? Acasta fenestrata: Utinomi 1958: p. 57, Figures 6–7 [51]. Rosell 1972: p. 194, Pl. XXI, XXII 1–3, XXIII 3, 4 [52]. Foster & Buckeridge 1994: p. 374, Figure 15A–I [53].
Material examined: WAM C20856, between Powerful and Survey Islands, Buccaneer Archipelago, WA, 16°05′ S 123°27′ E, intertidal, WAM Kimberley Islands & Reefs Survey, stn. 41, 26 Aug 1991, coll. D.S. Jones, host: Neopetrosia chaliniformis. WAM C43897, Montgomery Reef, Camden Sound, WA, 15°53.815′ S 124°19.531′ E, intertidal, Woodside Kimberley Survey, stn. 14/K09-A, 19 Oct 2009, coll. A.M. Hosie, host: Neopetrosia chaliniformis. WAM C45274, Montgomery Reef, Camden Sound, WA, 15°52.588′ S 124°19.793′ E, intertidal, Woodside Kimberley Survey, stn. 18/K09-T2, 20 Oct 2009, coll. A.M. Hosie, host: Neopetrosia chaliniformis. WAM C45275, WAM C55291, 4.9 mm RC, Montgomery Reef, Camden Sound, WA, 16°00.865′ S 124°10.389′ E, intertidal, Woodside Kimberley Survey, stn. 24/K09-T1, 23 Oct 2009, coll. A.M. Hosie; host: WAM Z29255 Neopetrosia chaliniformis. WAM C45278, Montgomery Reef, Camden Sound, WA, 15°51.323′ S 124°18.875′ E, intertidal, Woodside Kimberley Survey, stn. 17/K09-A, 20 Oct 2009, coll. A.M. Hosie, host: Neopetrosia chaliniformis. WAM C45280, Montgomery Reef, Camden Sound, WA, 15°55’08.6″ S 124°17’43.9″ E, intertidal, Woodside Kimberley Survey, stn. 16/K09, 19 Oct 2009, coll. A.M. Hosie; host: Neopetrosia chaliniformis. WAM C46096, Pt Cloates, Ningaloo Reef, WA, 22°46.35′ S 113°39.996′ E, 30 m, CReefs Ningaloo, stn. NR10-085, 31 May 2010, coll. L.L. Betterridge, host: Neopetrosia chaliniformis. WAM C46640, WAM C55273–4, all from Long Reef, Admiralty Gulf, WA, 13°48.593′ S 125°49.448′ E, intertidal, Woodside Kimberley Survey, stn. 52/K10-T1, 22 Oct 2010, coll. A.M. Hosie, host: WAM Z29256 Neopetrosia chaliniformis. WAM C46669, 5.7 mm RC, Long Reef, WA, 13°49.925′ S 125°48.712′ E, 12 m, Woodside Kimberley Biodiversity Survey 2010, stn. 54/K10-Q1, 23 Oct 2010, coll. A.M. Hosie, Host: WAM Z29257 Neopetrosia chaliniformis. WAM C48490, WAM C55409, WAM C58748,WAM C58749, 3 slides, all from E of Numanbu Island, Champagny Islands, Bonaparte Archipelago, 15°19.844′ S 124°13.015′ E, intertidal, Woodside Kimberley Survey, stn. 63/K11-T1, 14 Oct 2011, coll. A.M. Hosie, host: WAM Z54173 Neopetrosia chaliniformis. WAM C48495 from E of Numanbu Island, Champagny Islands, Bonaparte Archipelago, 15°19.844′ S 124°13.015′ E, intertidal, Woodside Kimberley Survey, stn. 63/K11-T2, 14 Oct 2011, coll. A.M. Hosie, host: WAM Z54220 Neopetrosia chaliniformis. WAM C54382, SE corner of Hibernia Reef, WA, 11°58’33.8″ S 123°23’22.8″ E, 13 m, Woodside Kimberley Survey 2013, stn. 145/K13, 5 Oct 2013, coll. A.M. Hosie, Host: WAM Z69888 Neopetrosia chaliniformis. WAM C55251–4, all from Ashmore Reef, 12°16.638′ S 123°08.166′ E, intertidal, Woodside Kimberley Survey, stn. 129/K13-T1, 28 Sept 2013, coll. A.M. Hosie, host: WAM Z29257 Neopetrosia chaliniformis. WAM C56252, WAM C55319–22, all from Mermaid Reef, Rowley Shoals, WA, 17°04.344′ S 119°37.626′ E, 12–14 m, Woodside Kimberley Survey, stn. 150/K14-T1, 2 Oct 2014, coll. A.M. Hosie; host: WAM Z84014 Neopetrosia chaliniformis. WAM C56867, WAM C55323–5 all from Clerke Reef, Rowley Shoals, WA, 17°18.858′ S 119°22.5′ E, 6–8 m, Woodside Kimberley Survey, stn. 171/K14-T2, 10 Oct 2014, coll. A.M. Hosie; host: WAM Z83364 Neopetrosia chaliniformis. WAM C66924, WAM C74354, WAM C74419–20, QM W29554 all from Ian Pt, Hook Island, Whitsunday Islands, QLD, 20°06′ 15.84″ S 148°54′ 8.99″ E, 2 m, Bush Blitz Coral Sea Expedition, Stn. BBCS16/2, 14 Jun 2016, coll. A.M. Hosie, Host Neopetrosia chaliniformis. WAM C67829, 3 slides, WAM C71738–9, WAM C71884, all from West Lewis Island, Dampier Archipelago, WA, 20°35′25" S 116°36′20″ E, 9.7 m, NCB Dampier Archipelago Expedition, 23/D17, 29 Mar 2017, coll. A.M. Hosie & A. Hara, host: WAM Z86889 Neopetrosia chaliniformis. WAM C72909–10, WAM C72914 all from Norwegian Bay, Ningaloo Reef, WA, 8–10 m, 22°39’45.2″ S 113°38’22.6″ E, 4 Jan 2017, coll. Abdul Wahab, M.A., host: Neopetrosia chaliniformis.
Diagnosis: Shell typically globuloconic, parietes with irregularly spaced spines, radii and alae oblique, membranous windows present between each shell plate, windows may reach half height of shell plates, maximum width approximately same as carinolatus at basal margin. Tergal spur truncate, basal margin concave with disto-articular angle projecting. Cirri III and IV anterior rami with row of sharp erect spines on anterodistal portion segments. Both rami of cirri IV–VI with specialised multicuspidate setae at posterodistal angles of basal-most segments.
Description: Shell (Figure 5A–G) plates white or tinged red or burgundy, typically globuloconic, but may elongate to match growth of host. Externally, parietes with fine, horizontal growth lines and short, irregularly spaced spines; apices of parietes curved toward orifice. Radii and alae oblique, not reaching basal margin of corresponding parietes, leaving elongate membranous windows between parietes. With ontogeny, windows secondarily filled with calcareous shell to maintain apical margin with internal basal margin of sheath. Internally, smooth below sheath, without longitudinal ribs, lateral margins not inflected or thickened. Sheath adpressed.
Basis (Figure 5G) cup-shaped, depth varying from nearly flat to making up more than twice total shell height (Figure 5A); rim smooth.
Scutum (Figure 5H–I) triangular, apex coloured dark red or purple, slightly higher than wide, growth lines prominent with fine crenulations and fringed with short, inconspicuous setae; basitergal angle broadly rounded; occludent margin strongly toothed. Internally smooth with slight depressions for adductor and lateral depressor muscles; articular ridge prominent, with deep furrow, extending halfway along tergal margin.
Tergum (Figure 5J,K) apex acute, tinged dark red or purple, growth lines fringed with short, inconspicuous setae; articular margin concave in apical half, carinal margin slightly convex almost straight, basal margin straight; spur length 0.5 times width, occupying approximately half of basal margin, positioned less than half own width from scutal margin; spur and furrow margins coincident, defined by abrupt change in growth lines and corresponding groove on articular side and gentle change on carinal side; spur truncate distal margin slightly concave with, disto-articular angle projecting (Figure 5J–K). Internally smooth, crests for depressor muscles faint.
Labrum (Figure 6A,B) bilobed, lobes separated by deep medial v-shaped notch, each rounded lobe with two or three marginal teeth and numerous fine setae.
Mandibular palp (Figure 6A) rhomboid, apex obliquely truncate, anterior margin concave, posterior margin straight; setae heavily serrulate, becoming longer and denser distally; distal angle broadly rounded with long, simple setae.
Mandible (Figure 6C,D) with five teeth, second to fourth teeth bifid, inferior angle with denticles. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxillule (Figure 6E–G) cutting margin straight, with 9–11 cuspidate setae, upper and lower pairs longer than remaining setae, 3 medial setae serrulate on inferior margin. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxilla (Figure 6H) bilobed, lobes ovate, serrulate setae arranged on anterior margins, becoming more dense at apex, longer on distal lobe than those on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 7A–B) protopod with pappose setae basally on protopod posterior margin. Rami unequal, anterior ramus twice length of posterior ramus, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus II (Figure 7C) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami unequal, anterior ramus 1.25 times longer that posterior ramus, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus III (Figure 7D,E) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally and plumose setae distally. Rami subequal, anterior ramus 1.15 length of posterior, segments with serrulate and simple setae on mesial face, less densely arranged than cirrus I and II. Anterior ramus with row of sharp, erect spines on anterodistal portion of all segments, except terminal segment, most prominent on medial segments.
Cirrus IV (Figure 8A–D) protopod elongate, length 1.83–1.95 times width, with simple setae on anterior and posterior margins, lacking denticles or erect spines on anterodistal angles, tuft of simple, cuspidate setae at posterodistal angles of both segments. Rami subequal, setation ctenopod, segments with 3 pairs of serrulate setae on anterior margin; posterodistal angles of basal-most segments with specialised, multicuspidate setae with pair of denticles situated midway along setae. Anterior ramus with row of sharp, erect spines on anterodistal portion of all segments except terminal segment, most prominent on medial segments.
Cirrus V (Figure 8E–G) and VI similar, protopod elongate length 1.32–1.38 and 1.21–1.31 times width, respectively, with simple setae on anterior and posterior margins, lacking denticles or erect spines on anterodistal angles, tuft of simple cuspidate setae at posterodistal angles of both segments. Rami subequal, setation ctenopod, cirrus V anterior ramus with 3 pairs of serrulate setae margin, posterior ramus and both rami of cirrus VI with 4 pairs of serrulate setae; posterodistal angles of basal-most segments with specialised multicuspidate setae with 1 pair of denticles situated midway along setae.
Penis longer than cirrus VI, annulated, sparsely setose, basidorsal point may be low and rounded or triangular and pointed.
Distribution: Australia: northern Western Australia and Queensland. Japan, Thailand, Indonesia [50]. Intertidal–195 m.
Hosts: Neopetrosia chaliniformis (Thiele, 1899) [54] (Petrosiidae). Previous records: Discodermia japonica Döderlein, 1884 [40,55] (Theonellidae); N. similis (Ridley & Dendy, 1886) [42,56]; “Geodia-like hard tetraxon sponge” and “soft siliceous sponge” [51]; Haliclona sp. [5] (Chalinidae); and N. chaliniformis (as N. exigua) and Neopetrosia sp. [48,49].
Remarks: In the literature, descriptions attributed to this species show significant variations that lead us to conclude that more than one species has been confounded under the name A. fenestrata. Darwin [38] only described the shell and opercular plates, stating that the body was poorly preserved, but that as far as he could tell, the cirri and mouthparts were similar to Neoacasta glans (Lamarck, 1818) [57]. The first description of the mouthparts and cirri to be attributed to A. fenestrata was provided by Utinomi [51], who stated that cirrus IV had hooked teeth on the anterior ramus, a feature not found in N. glans. Another departure from Darwin’s description is that the tergum, as figured, is much narrower in Utinomi’s specimens with the spur rounded, rather than truncated, and situated confluent with the basiscutal angle. Rosell [52] noted several differences between his accounts and those by Darwin and Utinomi, but maintained the identification based on the similarity with Utinomi’s description of the mouthparts and cirri. Key differences that Rosell highlighted were the presence of pliable, chitinous spines seated on calcareous “elevations” on the parietes, as opposed to the simple, short, blunt, calcareous spines described in other accounts, internal basal ribs on the parietes, and a crenated rim on the basis. The specimen figured by Foster and Buckeridge [53] has a generally globose appearance, with relatively broader radii, smaller windows, and hooked teeth on cirrus IV. Their records also extended the known depth range from 65 to 195 m.
These records contrast with the present specimens and the descriptions and illustrations given by Ren [45], Liu and Ren [47], Sulistiono et al. [49], and Wibowo et al. [48], where the examined specimens only have erect spines on the anterior margins of cirrus IV. Our material also presents a novel character not mentioned by previous authors: the multicuspidate setae on the posterodistal angles of cirri IV–VI (Figure 8D,G). A re-examination of type specimens, and possibly type host, along with additional specimens from across the reported geographic and host range would provide clarity to this species concept.
Figure 5.
Acasta fenestrata Darwin, 1854 shell plates (A, WAM C66924; B–D WAM C46669; E–J, WAM C55291). (A) whole, enormously elongate specimen still embedded in host; (B,C) left and right lateral views of whole specimen; (D) close up of fenestrae on right side; (E,F) ventral and lateral view of specimen (basis removed); (G) internal view of basis; (H,I) external and internal view of right scutum; (J,K) internal and external view of right tergum. Scale bars: A = 5 mm; B, C, E–G = 2 mm; H–K = 1 mm.
Figure 5.
Acasta fenestrata Darwin, 1854 shell plates (A, WAM C66924; B–D WAM C46669; E–J, WAM C55291). (A) whole, enormously elongate specimen still embedded in host; (B,C) left and right lateral views of whole specimen; (D) close up of fenestrae on right side; (E,F) ventral and lateral view of specimen (basis removed); (G) internal view of basis; (H,I) external and internal view of right scutum; (J,K) internal and external view of right tergum. Scale bars: A = 5 mm; B, C, E–G = 2 mm; H–K = 1 mm.

Figure 6.
Acasta fenestrata Darwin, 1854 mouth parts (all WAM C58749 except G, WAM C67829). (A) labrum and right mandibular palp; (B) close up of labrum crest; (C,D) left and right mandible; (E,F) left and right maxillule; (G) close up of setae on left maxillule; (H) left and right maxillae. Scale bars: A–B = 100 µm; C–F, H = 200 µm; G = 50 µm.
Figure 6.
Acasta fenestrata Darwin, 1854 mouth parts (all WAM C58749 except G, WAM C67829). (A) labrum and right mandibular palp; (B) close up of labrum crest; (C,D) left and right mandible; (E,F) left and right maxillule; (G) close up of setae on left maxillule; (H) left and right maxillae. Scale bars: A–B = 100 µm; C–F, H = 200 µm; G = 50 µm.

Figure 7.
Acasta fenestrata Darwin, 1854 cirri I–III (all WAM C58749). (A) left cirrus I; (B) serrulate setae on left cirrus I distal segments of posterior ramus; (C) left cirrus II; (D), left cirrus III; (E) erect spines on left cirrus III anterior ramus. Scale bars: A, B–C = 200 µm; D, E = 50 µm.
Figure 7.
Acasta fenestrata Darwin, 1854 cirri I–III (all WAM C58749). (A) left cirrus I; (B) serrulate setae on left cirrus I distal segments of posterior ramus; (C) left cirrus II; (D), left cirrus III; (E) erect spines on left cirrus III anterior ramus. Scale bars: A, B–C = 200 µm; D, E = 50 µm.

Figure 8.
Acasta fenestrata Darwin, 1854 cirri IV–VI (all WAM C58749). (A) left cirrus IV; (B) basal segments of left cirrus IV; (C) erect spines on medial segments of cirrus IV anterior ramus; (D) multicuspidate seta on posterodistal angle of cirrus IV posterior ramus; (E) left cirrus V; (F) setation on medial segments of cirrus V anterior ramus; (G) multicuspidate setae on basal segments of cirrus V anterior ramus; (H) left cirrus VI; (I) setation on intermediate segments of cirrus VI posterior and anterior rami. Scale Bars: A, E, H = 200 µm; B =100 µm; C, F, I = 50 µm; D, G 20 µm.
Figure 8.
Acasta fenestrata Darwin, 1854 cirri IV–VI (all WAM C58749). (A) left cirrus IV; (B) basal segments of left cirrus IV; (C) erect spines on medial segments of cirrus IV anterior ramus; (D) multicuspidate seta on posterodistal angle of cirrus IV posterior ramus; (E) left cirrus V; (F) setation on medial segments of cirrus V anterior ramus; (G) multicuspidate setae on basal segments of cirrus V anterior ramus; (H) left cirrus VI; (I) setation on intermediate segments of cirrus VI posterior and anterior rami. Scale Bars: A, E, H = 200 µm; B =100 µm; C, F, I = 50 µm; D, G 20 µm.

Acasta sandwichi Yu, Chan, Achituv & Kolbasov, 2017 [2]
Acasta sandwichi Yu et al., 2017: p. 606, Figures 22–28 [2].
Material examined: WAM C47383, Long Reef, WA, 13°49.121′ S 125°46.7796′ E, 12 m, Woodside Kimberley Biodiversity Survey 2010, stn. 53/K10, 23 Oct 2010, coll. A.M. Hosie, host: WAM Z29261 Xestospongia testudinaria. WAM C53344, S of Lesueur Island, WA, 28.3–29.5 m, 13°49′52″ S 127°15′52″ E to 13°49′56″ S 127°15′49″ E, CSIRO King George River Expedition 2013, stn. S04, 6 Jun 2013, coll. A.M. Hosie, C.S. Whisson & A. Hara, host: Xestospongia testudinaria.
Diagnosis: Shell fenestrate, windows present between each parietal junction, very large, occupying over half of shell height, as wide as or wider than adjacent parietes at basal rim. Parietes with blunt, calcareous projections, spaced irregularly in longitudinal rows. Basis shallow saucer-shaped, rim undulating, crenate where it interlocks with parietes, otherwise smooth. Cirrus IV anterior ramus with single denticle on anterior margin of segments.
Distribution: Western Australia: Kimberley. Taiwan [2]. 18–30 m.
Hosts: Xestospongia testudinaria (Lamarck, 1815) [58] (Petrosiidae).
Remarks: This species was only recently described from Taiwanese waters [2], and thus the specimens examined herein represent the first records from Australian waters.
This species has a morphologically and genetically close relationship to A. caveata sp. nov., and the morphological characters separating the two are dealt with under the treatment for that species. Distinguishing A. sandwichi from other Acasta with large windows was detailed by Yu et al. [2], and of these, the only A. fenestrata currently known is from Australia. This species can be distinguished by the relatively smaller windows, the lack of longitudinal ribs on the internal surface of the parietes, and the broader and more truncate tergal spur; the cirrus IV anterior ramus has erect spines on the anterodistal angles of segments and the stout multicuspidate setae are present on the posterodistal angles of cirri IV–VI. Further to this, there is no indication that either species inhabit the same hosts.
3.1.2. Euacasta Kolbasov, 1993 [5]
Euacasta acutaflava sp. nov. (Figure 9, Figure 10, Figure 11 and Figure 12)
Material examined: Holotype: WAM C58744, 4.6 mm RC, 3 slides, Montgomery Reef, Camden Sound, WA, 15°53.895′S 124°10.901′E, intertidal, Woodside Kimberley Biodiversity Survey 2009, stn. 20/K09-T2, 21 Oct 2009, coll. A.M. Hosie; host: WAM Z29216 Cymbastela stipitata.
Paratypes: WAM C58745–7, WAM C76401, 3 slides, same data as for holotype. WAM C61434, WAM C61435, 5.6 mm RC, E of Trimouille Island, Montebello Islands, WA, 20°22′18″ S 115°33′50″ E, 11 m, NCB Montebello/Muiron Islands Expedition, 1/MM15, 14 Apr 2015, coll. A.M. Hosie & A. Hara, Host: WAM Z84101 Cymbastela stipitata. WAM C 76451, 3.0 mm RC, 3 slides, Montgomery Reef, Camden Sound, WA, 15°54′ S 124°08′ E, intertidal, 15 Jul 1990, coll. L.M. Marsh, host: WAM Z59175 Cymbastela stipitata.
Other material: WAM C43667, 4.0 mm RC, more specimens still with host, same data as for holotype. WAM C54629, WAM C55289, both from Montgomery Reef, Camden Sound, WA, 15°54′ S 124°08′ E, intertidal, 15 Jul 1990, coll. L.M. Marsh, host: WAM Z59175 Cymbastela stipitata. WAM C54631, Enderby Island, Dampier Archipelago, WA, 20°35.15′ S 116°35.63′E, 17 m, Woodside Dampier Expedition, stn. DA3/99/55, 2 Sep 1999, coll. J. Fromont, host: WAM Z5468 Cymbastela vespertina. WAM C71476, Camden Sound, WA, 15°25′42.72″ S 124°16′23.39″ E, 16.9–17.2 m, WAMSI Survey 1B Camden Sound, stn. SOL160/WA025, 21 May 2015, coll. O.A. Gomez & J.A. Ritchie, Host: WAM Z87011 Cymbastela cf. stipitata.
Diagnosis: Shell squat, cuboid; parietes thick, strongly articulated, growth lines pronounced, inflected upwards at lateral margins, inner lamina with broad, prominent longitudinal ribs. Basis flat, square in outline with angular corners, diagonal grooves running from each corner. Opercular plates with yellow-orange, chitinous cuticle in apical section, particularly prominent on tergum; scutum with growth lines scalloped by longitudinal ridges. Cirrus IV with 2–4 hooked teeth on basis of protopod; anterior ramus with up to 3 teeth on basal segments.
Description: The shell (Figure 9A–Q) is white and squat with maximum diameter greater than shell height and square in outline. The parietes are thick and strongly articulated; externally having variably pronounced and irregularly spaced longitudinal ribs, with horizontal growth lines inflected upward on raised lateral margins; the parietes are slightly curved toward the orifice, the latter only slightly smaller than the diameter of the shell. The opercular plates are exposed, with the apices extending beyond the apices of the parietes. The radii and alae reach the basal margin of the corresponding parietes, and the summits are oblique and broad with horizontal striations. Rostrum, laterals, and carina are approximately equal in width; carinolateral is exceptionally narrow, the parietal area is represented by a beaded strip, and radii and alae are similar to other plates. Internally, the sheath has conspicuous horizontal growth lines, and the lower margin is separated from the inner lamina; below the sheath, broad, longitudinal, parietal ribs are very prominent.
The basis (Figure 9H,I) is flat and quadrangular with distinct angular corners; the concentric growth lines are conspicuous; the basal rim is strongly crenate, interlocking with internal ribs on the parietes.
The scutum (Figure 9R,S) is triangular, with height 1.35 times its width, and growth lines scalloped by longitudinal ridges; the ridges are increasing in number basally; the basitergal angle is rounded, and the tergal section is narrow; the occludent margin is strongly toothed. Internally smooth, the apex has a yellow chitinous cuticle, and adductor and lateral depressor muscle pits are present; the articular groove is deep, bounded by a distinct articular ridge, the latter extending halfway along the tergal margin.
The tergum (Figure 9T,U) apex is acute and beaked, with calcification giving way to a yellow–orange chitinous point, and the apex projecting beyond scutum when articulated; growth lines are prominent, fringed with short, inconspicuous setae; the yellow chitinous cuticle extends onto the carinal half; and the articular margin is concave in the apical half. The carinal margin is evenly concave; the basal margin is straight; the spur is truncate, the length is 0.5 times the width, occupying approximately half the basal margin, positioned less than half its own width from the scutal margin; the spur and furrow margins are coincident, the spur furrow is defined by an abrupt change in growth lines and a corresponding groove on the articular side, and a gentle change on the carinal side; internally smooth, the chitinous cuticle covers the upper half, and the crests for depressor muscles are faint. The articular furrow is broad, the articular ridge is low and rounded, occupying 0.25 of the scutal margin.
The labrum (Figure 10A,B) is bilobed, divided by a deep, medial, v-shaped notch, and each rounded lobe has two or three marginal teeth and numerous fine setae.
The mandibular palp (Figure 10A,C) is rhomboid, distal, and obliquely truncate, the anterior margin is concave, and the posterior margin is straight; the setae are heavily serrulate, becoming longer and denser distally; the distal angle has long, simple setae.
The mandible (Figure 10D–G) cutting margin has four distinct teeth, the second to fourth teeth are bifid, and the fifth tooth is congruent with molariform or spinose inferior angle; the superior and inferior margins have a row of long, fine, simple setae, with dense short setae covering the inner and outer faces.
The maxillule (Figure 10H) cutting margin is straight with 9–10 cuspidate setae, the upper and lower pairs are larger and more robust than the remaining setae, and the inferior angle has a tuft of shorter cuspidate setae. Dense, short setae are regularly spaced on the inner and outer faces, with longer fine setae on the inferior and superior margin.
The maxilla (Figure 10I) is bilobed, the lobes are ovate, and serrulate setae are arranged on the anterior margins, becoming denser at the apex and longer on the distal lobe than those on the basal lobe.
Cirral segment counts are given in Table 2.
The cirrus I (Figure 11A) protopod has pappose setae basally on the posterior margin. The rami are unequal, with anterior ramus 3.9 times the length of the posterior ramus and lasiopod setation, and segments have densely arranged serrulate and simple setae on the mesial face.
The cirrus II (Figure 11B,C) protopod anterior margin is lined with long plumose setae, posterior margin with pappose setae basally and plumose setae distally. The rami are unequal, the anterior ramus is 1.25 times longer than the posterior ramus, and segments have densely arranged serrulate and simple setae on the mesial faces.
The cirrus III (Figure 11D–F) protopod anterior margin is lined with long plumose setae, the posterior margin with pappose setae basally and plumose setae distally. The rami are subequal with anterior ramus 1.1 times the length of the posterior ramus, and segments have serrulate and simple setae on the mesial face, less densely arranged than cirrus I and II. The anterior ramus has a row of ctenoid scales on the anterodistal portion of all segments, most prominent on medial segments.
The cirrus IV (Figure 12A–F) protopod has simple setae on the anterior and posterior margins, and a tuft of simple, short setae at the posterodistal angles of both segments; the basis is elongate, with length 3.1–3.4 times the width, and the distal quarter of the anterior margin has a row of 2–4 hooked teeth increasing in size distally. The rami are subequal with ctenopod setation. The anterior ramus has 1–3 hooked teeth on the first nine segments, basal segments have one pair of serrulate setae at the anterodistal angle, and intermediate segments have two pairs. The posterior ramus with three pairs of setae on intermediate segments.
The cirrus V (Figure 12G–I) and VI (Figure 12J,K) protopods both have simple setae on the anterior and posterior margins and a tuft of simple, short setae at the posterodistal angles of both segments; the basis is less elongate than cirrus IV, with length 1.7–2 and 1.5–1.7 times width, respectively, without hooked teeth; the rami are subequal with intermediate segments having three or four pairs of setae. The anterior rami lack hooked teeth or erect spines.
The penis (Figure 12L) is longer than cirrus VI, annulated, and sparsely setose. The basidorsal point is low and rounded.
Distribution: Western Australia: Onslow to northern Kimberley, 0–15 m.
Hosts: Cymbastela stipitata (Bergquist and Tizard, 1967) [59], C. cf stipitata, and C. vespertina (Hooper and Bergquist, 1992) [60] (Axinellidae).
Etymology: The species epithet is derived from the Latin acuta (point, sharp) and flava (yellow) in reference to the yellow–orange chitinous tips of the terga.
Remarks: This species presents interesting variability in the details of the parietes. The longitudinal ribbing may be very faint, as it is in the holotype, to very pronounced (compare Figure 9A–E with Figure 9F,G). In some specimens, the basal margins are markedly convex, making the basal perimeter scalloped. The junctions between the rostrum, carina, and latera in these specimens cause the basal plate to form acute angles that are drawn up from the basal plane. This in turn causes grooves to run diagonally across the basis. In small specimens (~2 mm diameter), the horizontal growth lines appear denticulate and fringed with chitinous setae. The growth lines are especially pronounced at the lateral margin, where they inflect upwards and seem almost spine-like.
These projecting growth lines, however, are not comparable with the sharp, acicular, and fragile parietal spines present on the majority of the Euacasta: E. abnormis (Kolbasov, 1991) [61], E. aculeata (Nilsson-Cantell 1921) [62], E. ctenodentia (Rosell, 1972) [52], E. dofleini (Krüger, 1911) [63], E. microforaminia (Rosell, 1970) [64], E. porata (Nilsson-Cantell, 1921) [62], and E. tabachniki (Kolbasov, 1990) [65]. In E. zuiho (Hiro, 1936) [66] and E. excoriatrix sp. nov., the parietes have blunt calcareous projections, but are otherwise smooth, lacking any trace of longitudinal ridges. In E. sporillus (Darwin, 1854) [38], the parietes are perforated in a regular, grid-like arrangement. The only species with smooth parietes is E. antipathidis (Broch, 1916) [67], described based solely on dead, empty shells embedded in the skeleton of a moribund alcyonacean [11]. Despite the lack of opercular plates or cirri, it can be separated easily by the overall globose shape of the shell as compared to the box-like shape of E. acutaflava sp. nov.
The chitinous apex to the opercular plates is also a character unique to the E. acutaflava sp. nov. among the Euacasta. Further distinguishing features include the hooked teeth of cirrus IV protopod, which are restricted to the distal-most portion, whereas E. excoriatrix sp. nov., E. ctenodentia, E. dofleini, E. porata, and E. zuiho all have numerous teeth lining at least half of the basis (but see Yu et al. [68]). Both E. abnormis and E. tabachniki completely lack hooked teeth on cirrus IV, while E. sporillus, as figured by Kolbasov [5] and Yu [68], has three well-spaced teeth.
Figure 9.
Euacasta acutaflava sp. nov. (A–E) top, rostral, left, right and basal views, respectively, of holotype (WAM C58744), opercular plates removed; (F,G) left lateral and basal views, respectively, of paratype (WAM C61435), opercular plates removed; (H) internal view of basis; (I) enlarged portion of basis rim showing crenate margin; (J–M) external and (N–Q) internal views of rostrum, right lateral, right carinolateral and carina, respectively of paratype (WAM C76451); (R,S) external and internal view of scutum; (T,U) external and internal view of tergum of holotype (WAM C58744). Scale bars: A–G = 2 mm; H, J–U = 1 mm; I = 0.5 mm.
Figure 9.
Euacasta acutaflava sp. nov. (A–E) top, rostral, left, right and basal views, respectively, of holotype (WAM C58744), opercular plates removed; (F,G) left lateral and basal views, respectively, of paratype (WAM C61435), opercular plates removed; (H) internal view of basis; (I) enlarged portion of basis rim showing crenate margin; (J–M) external and (N–Q) internal views of rostrum, right lateral, right carinolateral and carina, respectively of paratype (WAM C76451); (R,S) external and internal view of scutum; (T,U) external and internal view of tergum of holotype (WAM C58744). Scale bars: A–G = 2 mm; H, J–U = 1 mm; I = 0.5 mm.

Figure 10.
Euacasta acutaflava sp. nov. mouthparts of holotype (WAM C58744) (A) labrum and right mandible palp; (B) close up of crest of labrum; (C) right mandible palp; (D) right mandible; (E) close up of inferior angle of right mandible; (F) left mandible; (G) close up of inferior angle of left mandible; (H) left maxillule; (I) left and right maxillae. Scale bars: A, I = 200 µm; B, E, G = 50 µm; C–D, F, H = 100 µm.
Figure 10.
Euacasta acutaflava sp. nov. mouthparts of holotype (WAM C58744) (A) labrum and right mandible palp; (B) close up of crest of labrum; (C) right mandible palp; (D) right mandible; (E) close up of inferior angle of right mandible; (F) left mandible; (G) close up of inferior angle of left mandible; (H) left maxillule; (I) left and right maxillae. Scale bars: A, I = 200 µm; B, E, G = 50 µm; C–D, F, H = 100 µm.

Figure 11.
Euacasta acutaflava sp. nov. cirri I–III of holotype (WAM C58744) (A) left cirrus I; (B) left cirrus II; (C) terminal segment of left cirrus II posterior ramus; (D) left cirrus III; (E) serrulate setae on terminal segment of right cirrus III posterior ramus; (F) medial segments of left cirrus III anterior ramus, arrows indicate ctenoid scales. Scale bars: A–B, D = 200 µm; C, E–F = 50 µm.
Figure 11.
Euacasta acutaflava sp. nov. cirri I–III of holotype (WAM C58744) (A) left cirrus I; (B) left cirrus II; (C) terminal segment of left cirrus II posterior ramus; (D) left cirrus III; (E) serrulate setae on terminal segment of right cirrus III posterior ramus; (F) medial segments of left cirrus III anterior ramus, arrows indicate ctenoid scales. Scale bars: A–B, D = 200 µm; C, E–F = 50 µm.

Figure 12.
Euacasta acutaflava sp. nov. cirri IV–VI of holotype (WAM C58744) (A) left cirrus IV; (B) basal segments of rami and basis of left cirrus IV protopod; (C,D) medial segments of left cirrus IV anterior ramus; (E) teeth on basis of right cirrus IV protopod; (F), medial segments of left cirrus IV posterior ramus; (G) left cirrus V; (H) basis of left cirrus V protopod; (I) medial segments of right cirrus V; (J) left cirrus VI; (K) medial segments of left cirrus VI; (L) basidorsal point of penis. Scale bars: A, G, J =200 µm; B, H–I, K–L 100 µm; C–E = 50 µm.
Figure 12.
Euacasta acutaflava sp. nov. cirri IV–VI of holotype (WAM C58744) (A) left cirrus IV; (B) basal segments of rami and basis of left cirrus IV protopod; (C,D) medial segments of left cirrus IV anterior ramus; (E) teeth on basis of right cirrus IV protopod; (F), medial segments of left cirrus IV posterior ramus; (G) left cirrus V; (H) basis of left cirrus V protopod; (I) medial segments of right cirrus V; (J) left cirrus VI; (K) medial segments of left cirrus VI; (L) basidorsal point of penis. Scale bars: A, G, J =200 µm; B, H–I, K–L 100 µm; C–E = 50 µm.

Euacasta excoriatrix sp. nov. (Figure 13, Figure 14, Figure 15 and Figure 16)
Material examined: Holotype: WAM C71800, 5.0 mm RC, 3 slides, Flying Foam Passage, Dampier Archipelago, WA, 20°28′3″ S 116°49′40″ E, 9 m, NCB Dampier Archipelago Expedition, stn. 4/D17, 23 Mar 2017, coll. A.M. Hosie & A. Hara, host: WAM Z86895 Halichondria BAR1
Paratypes: WAM C61475, 3 slides, S of Champagny Island, Buccaneer Archipelago, WA, 15°19′56.71″ S 124°14′09.49″ E, intertidal, Woodside Kimberley Survey, stn. 62/K11-T3, 15 Oct 2011, coll. O.A. Gomez, Host: WAM Z54014 Halichondria NW1; WAM C71801, same data as for holotype.
Other material: WAM C58573, same data as for holotype.
Diagnosis: Shell white, parietes studded with irregularly spaced, short calcareous spines. Basis concave, quadrangular, drawn up into angular corners at junctions of parietes, diagonal grooves crossing at the centre. Scutum elongate, prominent growth ridges crossed with longitudinal ridges, giving a beaded appearance. Tergum narrow, falcate apex beaked, spur broad, occupying over half of basal margin. Mandibular palp with distal corner produced into long narrow projection. Cirrus I posterior ramus ~5 times length of anterior ramus. Cirrus III anterior ramus with small, erect spines on anterodistal angle. Cirrus IV protopod with row of hooked teeth on 0.75 of anterior margin and group of erect spines on anterodistal angle of basis; anterior ramus with 2-3 hooked teeth on anterior margin and group of erect spines on anterodistal angle. Cirrus V with or without hooked teeth on protopod, anterior ramus with single tooth on medial segments.
Description: Shell (Figure 13A–O) white, slightly globose, parietes divergent, only slightly curved toward orifice, diameter of orifice equal to or greater than basis. Parietes externally with irregularly spaced, short, blunt calcareous projections, horizontal growth lines faint, basal margins convex. Radii and alae reaching basal margin of corresponding parietes, summits oblique, broad with fine horizontal and oblique striations. Rostrum and carina broadest plates, approximately equal in width; carinolateral exceptionally narrow, parietal area represented by thin strip, radii and alae similar to those on other plates. Internally, sheath occupying one third of inner lamina, with conspicuously raised, horizontal, growth ridges, lower margin separated from inner lamina, below sheath, narrow, longitudinal, parietal ribs extending to basal margin.
Basis (Figure 13J,O) shallow, subrectangular in outline, with rounded corners and convex sides; corners drawn up into junctions between parietes; conspicuous grooves running diagonally from each corner in “x”-shape; concentric growth lines conspicuous; basal rim finely crenate, interlocking with internal ribs on parietes.
Scutum (Figure 13P,Q) triangular, height 1.5 times width, concave on apicobasal axis, growth lines scalloped by longitudinal ridges; basitergal angle rounded, tergal section narrow; occludent margin strongly toothed. Internally smooth, adductor and lateral depressor muscle pits faint, not well defined; articular groove narrow, bounded by distinct articular ridge, latter extending halfway along tergal margin, not projecting beyond tergal margin.
Tergum (Figure 13R,S) narrow, apex tinged with red, acute, strongly beaked, projecting beyond scutum when articulated; growth lines fine, well-spaced, fringed with short, inconspicuous setae; carinal margin evenly concave; basal margin straight; spur distally truncate, broad, length 0.3 times width, occupying approximately 0.6 of basal margin, positioned approximately 0.15 own width from scutal margin; spur and furrow margins coincident, spur furrow defined by abrupt change in growth lines on both sides; internally smooth, crests for depressor muscles absent; articular furrow narrow, articular ridge low, rounded occupying approximately 0.3 scutal margin.
Labrum (Figure 14A,B) bilobed, divided by deep, medial, v-shaped notch, each rounded lobe with 3 marginal teeth and numerous fine setae.
Mandibular palp (Figure 14A,C) rhomboid, distal angle produced ventrally into long, narrow projection, anterior margin concave, posterior margin straight; setae heavily serrulate, becoming longer and denser distally; distal angle with long, simple setae.
Mandible (Figure 14D,E) cutting margin with five distinct teeth, second and third teeth bifid, fourth tooth becoming molariform, fifth tooth congruent with molariform or spinose inferior angle; superior and inferior margins with row of long, fine simple setae; dense, short setae covering inner and outer faces.
Maxillule (Figure 14F,G) cutting margin straight with 10–12 cuspidate setae, upper and lower pairs larger and more robust than remaining setae; inferior angle with tuft of shorter cuspidate setae. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxilla (Figure 14H) bilobed, lobes ovate, serrulate setae arranged on anterior margins, becoming more dense at apex, longer on distal lobe than those on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 15A) protopod with pappose setae basally on posterior margin. Rami unequal, anterior ramus approximately 5 times length of posterior ramus, setation lasiopod; segments with densely arranged serrulate and simple setae on mesial face.
Cirrus II (Figure 15B,C) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami unequal, anterior ramus 1.33 times longer that posterior ramus, segments with densely arranged serrulate and simple setae on mesial face, transverse row of ctenoid scales on distal margin of each segment.
Cirrus III (Figure 15D,E) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami subequal, anterior ramus 1.1 times longer that posterior ramus segments with serrulate and simple setae on mesial face, less densely arranged than on cirrus I and II. Anterior ramus with row of small, erect spines and ctenoid scales on anterodistal portion of all segments, except terminal segment, most prominent on medial segments.
Cirrus IV (Figure 16A–G) protopod with simple setae on anterior and posterior margins, tuft of simple, short setae at posterodistal angles of both segments; basis elongate, length 3.1–3.4 times width, anterior margin with row of 9–12 hooked teeth extending 0.75 length, evenly sized or increasing in size distally, anterodistal angle with additional upward turned teeth and erect spines. Rami subequal, setation ctenopod. Anterior ramus with 1–3 hooked teeth on first 10–11 segments, anterodistal angle with erect spines and ctenoid scales; basal segments with 1 pair of serrulate setae at anterodistal angle, intermediate segments with 2 pairs. Posterior ramus without hooked teeth, anterodistal angle with small erect spines, ctenoid scales on distal margin, 3 pairs of setae on intermediate segments.
Cirrus V (Figure 16H–J) protopod with simple setae on anterior and posterior margins, tuft of simple, short setae at posterodistal angles of both segments; basis less elongate than cirrus IV, length 2.2–2.5 times width, up to 3 teeth distally on anterior margin or teeth absent. Rami subequal, intermediate segments with 3 or 4 pairs of setae. Anterior ramus with 1 hooked tooth on 11th–18th segments; posterior ramus lacking teeth.
Cirrus VI (Figure 16L,M) similar, protopod with simple setae on anterior and posterior margins, tuft of simple, short setae at posterodistal angles of both segments; basis less elongate than cirrus IV, length 1.6–2.1 times width, respectively, without hooked teeth. Rami subequal, intermediate segments with 3 or 4 pairs of setae. Anterior rami lacking hooked teeth or short erect spinules along distal margins.
Penis (Figure 16L) shorter than cirrus VI, annulated, sparsely setose. Basidorsal point (Figure 16N), triangular, apex rounded, acute.
Hosts: Halichondria BAR1, H. NW1 (Halichondriidae).
Distribution: Western Australia: Dampier Archipelago to the Kimberley. Intertidal–9 m.
Etymology: The species epithet translates as “the flayer” and is derived from the Latin excorior (to flay, to skin), this is in reference to the long whip-like cirri and the strong hooked teeth on cirrus IV and V that are used to scrape away encroaching host tissues to prevent being overgrown.
Remarks: This species is characterised by several features that separate it from the remainder of the genus. In particular, the beaked tergum, the relative lengths and armature of the cirri, and mandible palp clearly define the morphological boundaries of this species. The upwardly directed teeth on the anterodistal angle of the protopod of cirrus IV also appear to be unique within the genus, as the other species have only been documented with teeth that are horizontal or hooked downward. Both E. abnormis and E. tabachniki have a tergum with a similarly beaked apex, but these species lack teeth on cirrus IV and V; the mandibular palps are not elongated; in E. abnormis, there are no internal parietal ribs; and E. tabachniki has long spines on the parietes [61,65]. In external appearance, this species is most similar to E. zuiho (sensu Hiro 1936 [45]), but this species has a typically truncated mandibular palp, shorter cirri (segment counts can be up to half of those in the present specimens), and the anterior ramus of cirrus I is approximately 2.5 times longer than the posterior, compared with 5 in E. excoriatrix [66]. However, the figures of E. zuiho given by Ren ([45] Figure 11 and Plate III 18–22) and Yu et al. ([68] Figure 14, Supplementary Figures S10 and S12) show a prominently beaked tergum, and, in the case of Ren, a mandible with a slightly produced distal angle. Their accounts still differ from the present species, having relatively short cirri and no erect spines on the protopod basis of cirrus IV. Further to this, the ribs on the inner lamina of the parietes are restricted to the basal margin, and the basidorsal point of the penis is absent in Ren’s specimens. Lastly, in Yu et al., the tergal spur is distinctly separated from the basi-scutal angle. These records show some differences with those of the original description [66] and potentially also represent different species.
The armature of cirrus IV is rather constant, with only small changes in the number of teeth on the protopod and segments, but the variation seen in the protopod of cirrus V is peculiar and not symmetrical. One of the paratypes (WAM C61475) bears two small, upturned teeth on the left and three hooked teeth on the right, while the holotype bears no trace of teeth.
Figure 13.
Euacasta excoriatrix sp. nov. shell and shell plates (all WAM C71800). (A–E) left, rostral, right, carinal and basal views, respectively, of whole shell; (F–J) external view of rostrum, right latus, right carinolatus, carina, and basis, respectively; (K–N) internal view of rostrum, right latus, right carinolatus and carina, respectively; (O) close up of basal rim; (P,Q), external and internal view of right scutum; (R,S), internal and external view of right tergum. Scale bars: A–N, P–S = 2 mm; O = 1 mm.
Figure 13.
Euacasta excoriatrix sp. nov. shell and shell plates (all WAM C71800). (A–E) left, rostral, right, carinal and basal views, respectively, of whole shell; (F–J) external view of rostrum, right latus, right carinolatus, carina, and basis, respectively; (K–N) internal view of rostrum, right latus, right carinolatus and carina, respectively; (O) close up of basal rim; (P,Q), external and internal view of right scutum; (R,S), internal and external view of right tergum. Scale bars: A–N, P–S = 2 mm; O = 1 mm.

Figure 14.
Euacasta excoriatrix sp. nov. mouthparts (A,B, WAM C71800, holotype; C–H, WAM C61475 paratype). (A) labrum with right mandible palp; (B) crest of labrum; C, right mandible palp; (D,E) left and right mandible; (F,G) left and right maxillule; (H) left and right maxillae. Scale bars: A, C–H = 200 µm; B = 100 µm.
Figure 14.
Euacasta excoriatrix sp. nov. mouthparts (A,B, WAM C71800, holotype; C–H, WAM C61475 paratype). (A) labrum with right mandible palp; (B) crest of labrum; C, right mandible palp; (D,E) left and right mandible; (F,G) left and right maxillule; (H) left and right maxillae. Scale bars: A, C–H = 200 µm; B = 100 µm.

Figure 15.
Euacasta excoriatrix sp. nov. Cirri I–III (WAM C71800, holotype). (A) left cirrus I; (B) left cirrus II; (C) ctenoid scales on cirrus II anterior ramus segment; (D) left cirrus III; (E) small erect spines on the anterodistal angle of cirrus III anterior ramus. Scale bars: A–B, D = 200 µm; C, E = 50 µm.
Figure 15.
Euacasta excoriatrix sp. nov. Cirri I–III (WAM C71800, holotype). (A) left cirrus I; (B) left cirrus II; (C) ctenoid scales on cirrus II anterior ramus segment; (D) left cirrus III; (E) small erect spines on the anterodistal angle of cirrus III anterior ramus. Scale bars: A–B, D = 200 µm; C, E = 50 µm.

Figure 16.
Euacasta excoriatrix sp. nov. cirri IV–VI and penis (A–C, E–M WAM C71800, holotype; D, WAM C61475, paratype). (A) left cirrus IV; (B) basis of cirrus IV protopod; (C) close up of anterodistal angle of same; (D) basis of cirrus IV protopod; (E) medial segments of cirrus IV anterior and posterior rami; (F,G) detail of teeth and erect spines on segments of cirrus IV anterior ramus; (H) left cirrus V; (I,J) teeth on distal portion of left and right cirrus V protopod; (K) tooth on medial segment of right cirrus V; (L) left cirrus VI and penis; (M) medial segments of cirrus VI anterior and posterior rami; (N) basidorsal point of penis. Scale bars: A, H, L = 400 µm; B, D–E, I–J, M = 100 µm; C, F, G, K, N = 50 µm.
Figure 16.
Euacasta excoriatrix sp. nov. cirri IV–VI and penis (A–C, E–M WAM C71800, holotype; D, WAM C61475, paratype). (A) left cirrus IV; (B) basis of cirrus IV protopod; (C) close up of anterodistal angle of same; (D) basis of cirrus IV protopod; (E) medial segments of cirrus IV anterior and posterior rami; (F,G) detail of teeth and erect spines on segments of cirrus IV anterior ramus; (H) left cirrus V; (I,J) teeth on distal portion of left and right cirrus V protopod; (K) tooth on medial segment of right cirrus V; (L) left cirrus VI and penis; (M) medial segments of cirrus VI anterior and posterior rami; (N) basidorsal point of penis. Scale bars: A, H, L = 400 µm; B, D–E, I–J, M = 100 µm; C, F, G, K, N = 50 µm.

3.1.3. Pectinoacasta Kolbasov, 1993 [5]
Pectinoacasta cancellorum (Hiro, 1931) [69] (Figure 17, Figure 18, Figure 19 and Figure 20)
Acasta cancellorum Hiro, 1931: p. 151, Figures 8A–C, Pl. XIII 1–1d [69]. Hiro 1937: p. 459, Figures 36, 37 [7].
Pectinoacasta cancellorum Kolbasov 1993: p. 411 [5].
Material examined: Lectotype: SMBL #149, Seto, Kii Province, Japan
Australia: WAM C50791, WAM C55262–4, all from Echuca Shoal, WA, 13°54′02.48″ S 123°53′36.42″ E, 20 m, Woodside Kimberley Survey 2012, stn. 108/K12, 18 Oct 2012, coll. A.M. Hosie, host: WAM Z54694 Agelas KMB1. WAM C57105, WAM C55302–7, all from Mermaid Reef, Rowley Shoals, WA, 17°01’35.7″ S 119°37’08.0″ E, 25 m, Woodside Kimberley Survey 2014, stn. 181/K14, 14 Oct 2014, coll. A.M. Hosie, host: Agelas KMB1. WAM C55340, 10.1 mm RC, Wheatstone, off Onslow, WA, 21°35′46″ S 115°00′43″ E, 12.3 m, WAMSI Onslow Survey I March 2013, stn. RVS5737/D11-T1, 27 Mar 2013, coll. C.L. Schonberg, host: WAM Z65287 Agelas cf. mauritiana. WAM C56830, WAM C55308–9, all from Clerke Reef, WA, 17°19’01.1″ S 119°23’01.6″ E, 18 m, Woodside Kimberley Survey 2014, stn. 170/K14, 10 Oct 2014, coll. A.M. Hosie, Host: Agelas KMB1.
Diagnosis: Shell globose, parietes permeated with narrow longitudinal incisions extending from basal rim and small membranous windows at parietal junctions; externally surfaces studded with short sharp calcareous spines; internally longitudinal ribs extend below sheath and interlock with crenate basal rim. Scutum with longitudinal ridges crossing growth ridges. Tergum with setose, yellow chitinous cuticle covering external surface of carinal side of spur furrow, growth ridges crossed with longitudinal ridges giving a latticed appearance. Cirrus IV protopod with row of 6–7 teeth on anterior margin, anterior ramus basal segments with 1–2 recurved teeth on anterior margins.
Description: Shell (Figure 17A–D,J,K), white or grey; orifice small relative to shell diameter, rim bluntly toothed. Parietes with longitudinal incisions extending from basal rim to over half parietal height, creating narrow, membrane-covered slits, incisions secondarily calcified in upper portions of parietes; external surfaces with numerous short, sharp, calcareous projections arranged irregularly, or in longitudinal rows where confined by incisions; parietes strongly curved toward orifice. Radii and alae not reaching basal rim, leaving small membranous windows between parietes; broad with striations perpendicular to oblique summits. Rostrum widest shell plate; internally, sheath with conspicuous horizontal growth lines; lower margin separated from internal parietal wall, extending to just above apical extent of parietal incisions.
Basis circular, shallow to deeply cupped, not more than half total shell height; concentric growth lines conspicuous; basal rim irregularly crenate, interlocking with internal ribs of parietes.
Scutum (Figure 17L,M) triangular, as wide as or slightly wider than high, growth ridges strongly crenate with raised points and short setae, longitudinal ridges variably pronounced; basitergal angle rounded, tergal section indistinctly separated; occludent margin strongly toothed. Internally smooth, apex with yellow chitinous cuticle, adductor and lateral depressor muscle pits present; articular ridge short, occupying 0.3 length of tergal margin, prominently protruding beyond tergal margin, with deep articular furrow, extending entire length of tergal margin.
Tergum (Figure 17N,O) apex blunt, external surface covered by yellow, setose, chitinous cuticle on carinal side of spur furrow; growth lines prominent, crossed by strong longitudinal ridges giving a lattice-like or cancellated appearance, fringed with short inconspicuous setae; articular margin concave in apical half. Carinal margin almost straight, only curved near apex; basal margin straight; spur length approximately 1.25 times width, occupying approximately 0.25 basal margin, positioned less than half own width from scutal margin; spur furrow defined by abrupt change in growth lines, margins infolded, furrow open in basal half; distal margin rounded or truncate. Internally roughened with raised tubercles in apical section, chitinous cuticle on carinal and basal margins, crests for depressor muscles prominent, particularly in larger specimens. Articular furrow broad, articular ridge low, rounded occupying 0.25 scutal margin.
Labrum (Figure 18A,B) bilobed, divided by deep, medial, v-shaped notch, each rounded lobe with 3 prominent, marginal teeth and numerous fine setae.
Mandibular palp (Figure 18A,C) rhomboid, distal end obliquely truncate, anterior margin concave, posterior margin straight; setae heavily serrulate, becoming longer and more dense distally; distal angle with long, simple setae
Mandible (Figure 18D–F) cutting margin with 4 distinct teeth, second to fourth tooth bifid, fifth tooth congruent with molariform or spinose inferior angle; superior and inferior margins with row of long, fine, simple setae, dense short setae covering inner and outer faces.
Maxillule (Figure 18G,H) cutting margin straight, with small notch between second and third setae; 10–11 cuspidate setae, upper and lower pair larger and more robust than remaining setae, 3–4 medial setae serrate on inferior margin. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxilla (Figure 18I) bilobed, lobes ovate, distal lobe elongate, length more than twice width, serrulate setae arranged on anterior margins, becoming more dense at apex, setae longer on distal lobe than on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 19A,B) protopod with pappose setae on basal segment posterior margin, basis with tuft of setae at posterodistal angle. Rami unequal, anterior ramus 3 times length of posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face, tuft of simple setae on posterodistal angle.
Cirrus II (Figure 19C) protopod lined with long serrulate setae on anterior margin and pappose or plumose setae on posterior margin on basal segment; basis with long serrulate setae on anterior margin. Rami unequal, anterior ramus 1.25 times longer than posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus III (Figure 19D–F) protopod lined with long serrulate setae on anterior margin and pappose or plumose setae on posterior margin on basal segment. Rami unequal, anterior ramus 1.25 times length of posterior ramus; setation lasiopod, segments with serrulate and simple setae on mesial face, less densely arranged than on cirrus I and II. Anterior ramus with transverse row of distally directed spines or ctenoid scales at anterodistal angle on all but distal-most segments, spines most prominent on medial segments; posterodistal angle with transverse row of ctenoid scales. Posterior ramus with similar spination but much reduced in size.
Cirrus IV (Figure 20A–F) protopod basal segment posterior margin with few plumose setae, tuft of simple, short setae at posterodistal angle; anterior margin with widely spaced plumose setae; basis elongate, length 2.09–2.15 width, with tuft of simple, long setae at posterodistal angle; anterior margin lined with up to 7 well-spaced, hooked teeth, lower third lacking teeth. Rami subequal in length, setation ctenopod. Anterior ramus with 1-2 hooked teeth on anterior margins of first 14 segments, anterodistal angles with transverse row of erect spines on all but distal-most segments; 3 pairs of serrulate setae on medial segments. Posterior ramus with similar setation as anterior ramus, with erect spines on anterodistal angles, with or without hooked teeth.
Cirrus V (Figure 20G) protopod with well-spaced, long setae and dense row of short spinules on anterior margins; posterior margin lacking setae; basis elongate, length 1.7–1.75 width, without hooked teeth, row of long simple setae on anterior margin and tuft of setae at posterodistal angle. Rami subequal, setation ctenopod, 3 pairs of serrulate setae on anterior margins; without teeth or erect spines.
Cirrus VI (Figure 20H–K) protopod and rami setation as in cirrus V; protopod basis elongate, length 1.78–1.97 width, without hooked teeth, longitudinal row of small spinules on anterior and posterior margins of basal segment. Anterior ramus without erect spines or teeth on anterior margins; both rami with 3 pairs of serrulate setae on medial segments.
Penis (Figure 20H) shorter than cirrus VI, annulated, sparsely setose; basidorsal point rounded, inconspicuous.
Distribution: Australia: oceanic coral reefs and shoals off northern WA. Southern Japan [69].
Remarks: The presence of longitudinal slits in the parietes immediately distinguishes P. cancellorum from all other members of the genus except P. sculpturata [6], with which it also shares the chitinous cuticle on the opercular plates. The two species share an obvious morphological affinity, as recognised by Hiro [7], although what he considered to be “remarkable differences between… the sculpture of the opercular valves” of the two species was presumably in reference to the presence of the yellow cuticle on the tergum, which was omitted from Broch’s description of P. sculpturata [6]. This leaves a finer morphological distinction between the two species than at first conceived. Key differences include the presence of hooked teeth on the protopod of cirrus IV of P. cancellorum (absent in P. sculpturata), the basis of which is more elongate (height/width ratio >2.0 in P. cancellorum, <2.0 in P. sculpturata), and the basidorsal point, which is rounded and inconspicuous (triangular and prominent in P. sculpturata). Furthermore, the calcareous spines on the parietes appear smaller and more numerous in P. cancellorum compared to the stout, blunt spines on P. sculpturata. This morphological affinity also corresponds with closely related host usage and a close genetic relationship (see Section 3.2).
There are no slide-mounted specimens among the extant cotypes with which to compare to our material. Only the specimen selected to serve as lectotype still has the prosoma present, which has at some point dried out. This has distorted the prosoma and makes dissection exceptionally difficult without risking further damage. However, some key characters are still observable, e.g., the teeth on cirrus IV are arranged in a similar fashion. There is some variability in the armature of cirrus IV within this species. Hiro describes the posterior ramus of cirrus IV as typically, but not always, bearing a single hooked tooth on the segments; in the present material, only one specimen was found to have this character. The number and development of the teeth on the protopod are also variable, with some specimens having few teeth restricted to the upper portion of the protopod and others having irregular spacing, and the left and right cirri do not necessarily have the same arrangement.
The type series also lacks any trace of the host and the accounts by Hiro [7,69], only mention that it was found in association with A. sulcata [57]. This latter species has now been recorded from three orders of sponge: Haplosclerida, Poecilosclerida, and Dictyoceratida [3,5,19]. The present specimens were all collected in species of Agelas (order Agelasida), the same host genus as P. sculpturata, albeit morphologically different species.
Figure 17.
Pectinoacasta cancellorum (Hiro, 1931) shell and shell plates (A–I, Lectotype SMBL #149; J,K, WAM C55340; L–O, WAM C55305): (A,B) external and internal view of rostrum; (C,D) external and internal view of articulated right lateral and carinolateral (radius broken) plates; (E,F) internal and external view of left scutum; (G,H) external and internal view of left tergum; (I) close up of depressor muscle crests on right tergum; (J,K) left and right views, respectively, of whole specimen; (L,M) external and internal views of right scutum; (N,O) internal and external views of right tergum. Scale bars: A–D, J,K = 2 mm, E–H, L–O = 1 mm; I = 0.5 mm.
Figure 17.
Pectinoacasta cancellorum (Hiro, 1931) shell and shell plates (A–I, Lectotype SMBL #149; J,K, WAM C55340; L–O, WAM C55305): (A,B) external and internal view of rostrum; (C,D) external and internal view of articulated right lateral and carinolateral (radius broken) plates; (E,F) internal and external view of left scutum; (G,H) external and internal view of left tergum; (I) close up of depressor muscle crests on right tergum; (J,K) left and right views, respectively, of whole specimen; (L,M) external and internal views of right scutum; (N,O) internal and external views of right tergum. Scale bars: A–D, J,K = 2 mm, E–H, L–O = 1 mm; I = 0.5 mm.

Figure 18.
Pectinoacasta cancellorum (Hiro, 1931) mouthparts (WAM C55305): (A) labrum and right mandible palp; (B) close up of labrum crest; (C) left mandible palp; (D) left mandible; (E) closeup of inferior angle; (F) right mandible; (G) right maxillule; (H) close up of serrated setae of maxillule; (I) left and right maxilla. Scale bars: A, D, F, G, I = 200 µm; B = 50 µm; C = 100 µm.
Figure 18.
Pectinoacasta cancellorum (Hiro, 1931) mouthparts (WAM C55305): (A) labrum and right mandible palp; (B) close up of labrum crest; (C) left mandible palp; (D) left mandible; (E) closeup of inferior angle; (F) right mandible; (G) right maxillule; (H) close up of serrated setae of maxillule; (I) left and right maxilla. Scale bars: A, D, F, G, I = 200 µm; B = 50 µm; C = 100 µm.

Figure 19.
Pectinoacasta cancellorum (Hiro, 1931) cirri I–III (WAM C55340). (A) left cirrus I; (B) serrulate setae on cirrus I anterior ramus terminal segment; (C) cirrus II; (D) Cirrus III; E, erect spines on cirrus III anterior ramus; (E) ctenoid scales on cirrus III posterior ramus. Scale Bars: A, C–D = 200 µm; B, E–F = 50 µm.
Figure 19.
Pectinoacasta cancellorum (Hiro, 1931) cirri I–III (WAM C55340). (A) left cirrus I; (B) serrulate setae on cirrus I anterior ramus terminal segment; (C) cirrus II; (D) Cirrus III; E, erect spines on cirrus III anterior ramus; (E) ctenoid scales on cirrus III posterior ramus. Scale Bars: A, C–D = 200 µm; B, E–F = 50 µm.

Figure 20.
Pectinoacasta cancellorum (Hiro, 1931) cirri IV–VI (WAM C55340): (A) left cirrus IV; (B) basis of left cirrus IV protopod; (C) same of right cirrus IV; (D) medial segments of cirrus IV; (E) small spinules on medial segments of cirrus IV posterior ramus; (F) erect spines on medial segments of right cirrus IV anterior ramus; (G) right cirrus V; (H) left cirrus VI with penis; (I) medial segments of left cirrus VI; (J,K) close up of posterior and anterior margin of cirrus VI protopod basal segment. Scale Bars: A, G, H = 500 µm; B, C, I = 100 µm; D = 200 µm; E, F, J, K = 50 µm.
Figure 20.
Pectinoacasta cancellorum (Hiro, 1931) cirri IV–VI (WAM C55340): (A) left cirrus IV; (B) basis of left cirrus IV protopod; (C) same of right cirrus IV; (D) medial segments of cirrus IV; (E) small spinules on medial segments of cirrus IV posterior ramus; (F) erect spines on medial segments of right cirrus IV anterior ramus; (G) right cirrus V; (H) left cirrus VI with penis; (I) medial segments of left cirrus VI; (J,K) close up of posterior and anterior margin of cirrus VI protopod basal segment. Scale Bars: A, G, H = 500 µm; B, C, I = 100 µm; D = 200 µm; E, F, J, K = 50 µm.

Pectinoacasta pectinipes (Pilsbry, 1912) [71] (Figure 21, Figure 22, Figure 23 and Figure 24)
Acasta pectinipes Pilsbry, 1912: p. 294 [71]. Broch 1922: p. 330 [72]. Barnard 1924: p. 86 [73]. Broch 1931: p. 105 [6]. Hiro 1937: p. 463, Figure 38 [7]. Nilsson-Cantell 1938: p. 57, Figure 19 [42]. Utinomi 1962: p. 221 [74]. Zullo 1968: p. 227, Figure 6 [75]. Newman & Ross 1976: p. 54 [44]. Foster 1980: p. 219, Figure 4C. Ren 1984 [45]: p. 189, Figure 4, Pl. 2 10–18. Jones 1990: p. 222 [76] —Jones et al. 1990: p. 15 [77].
Acasta nitida Hoek, 1913: 237, Pl. 24 Figures 17–19, Pl. 25 Figures 1–3 [10].
Acasta komaii Hiro, 1931: 149, Pl. 12 Figures 1–1d [69].
Pectinoacasta pectinipes. Kolbasov, 1993: 411, Figure 14 [5]. Jones et al. 2000: p. 273 [78]. Liu & Ren 2007: [47]: Jones 2010: p. 217 [79]. Van Syoc et al. 2015: Tab. 1–2 [1]. Jones and Hosie 2016: p. 277 [50].
Material examined: WAM C48061, Cockburn Sound, WA, 32°12′S 115°44′ E, stn. RW3 Q3 (rep 3), Aug 2008, host: Trachytedania L2. WAM C58141, WAM C58870, 3.4 mm RC, WAM C58871, 3 slides, WAMC 58872–3, all from South Beach, Fremantle, WA, 32°01’25.3″ S 115°45’00.1″ E, washed up on beach, 10 Jun 2015, coll. G. Raymond, host: WAM Trachytedania L1. WAM C66680, WAM C67748, WAM C67749, 3 slides, WAM C67750, all from west side of North Muiron Island, WA, 21°37′40″ S 114°22′25″ E, 17 m, NCB Exmouth & Muirons Expedition, stn. 7/EM16, 3 Jun 2016, coll. A. Hara, Host Z97093 Trachytedania MM1. WAM C66678, WAM C67772, 7.4mm RC, all from west side of North Muiron Island, WA, 21°37′40″ S 114°22′25″ E, 17 m, NCB Exmouth & Muirons 2016 Expedition, stn. 7/EM16, 3 Jun 2016, coll. A. Hara, Host Z97092 Trachytedania MM1. WAM C67123, South Beach, Fremantle, WA, washed up on beach, 32°04′ S 115°45′ E, 22 May 2016, coll. J. Raymond, Host: Trachytedania L1. WAM C70035, W of Adele Island, Lynher Bank, 63.2–64.3 m, 15°32′41.520″ S 122°12′25.680″ E to 15°32′38.760″ S 122°12′27.240″ E WAMSI Survey 4 Lynher Bank, stn. SOL23/WA112, 30 Oct 2016, coll. J. Fromont & J.A. Ritchie, host: WAM Z90726 Tedania sp.1. QM W27677, near Viscount Shoals, Hay Pt, Queensland, 21°25’54.9″ S 149°31’02.9″ E, 27 May 1997, host unknown.
Diagnosis: Shell globose, basis shallow to deeply cupped, small membranous slits between parietal junctions. Externally, parietes with variously pronounced longitudinal ribs lined with small, sharp points; internally with prominent longitudinal ribs interlocking with crenate rim of basis. Scutum transversely elongate, with longitudinal ridges crossing growth ridges. Tergum growth ridges crossed with strong longitudinal ridges giving a latticed appearance. Cirrus IV protopod with 14–15 curved teeth, anterior ramus basal segments with 1–4 recurved teeth on anterior margins. Cirrus V with or without 1–3 teeth on medial segments of anterior ramus.
Description: Shell (Figure 21A–H) globose, ranging from white to orange or pink; orifice small relative to shell diameter, rim toothed. Parietes externally with variously pronounced longitudinal ribs studded with small, sharp points; parietes strongly curved over opercular plates, creating narrow orifice. Radii and alae not reaching basal rim, leaving small membranous slits between parietes; summits oblique, broad with horizontal striations. Rostrum widest shell plate. Internally, sheath with conspicuous horizontal growth lines, vesicular structure where sheath adjoins parietes; lower margin separated from internal parietal wall, below sheath longitudinal ribs prominent.
Basis (Figure 21H) circular, shallow to deeply cupped, making up to half total shell height; concentric growth lines conspicuous; basal rim strongly crenate, interlocking with internal ribs on parietes.
Scutum (Figure 21I,J) triangular, transversely elongate, height 0.6 times width, growth lines marked by longitudinal ridges, ridges increasing in number basally; basitergal angle angular, tergal section narrow; occludent margin strongly toothed. Internally smooth, apex with yellow chitinous cuticle, adductor and lateral depressor muscle pits present; articular ridge short, occupying 0.3 length of tergal margin, prominently protruding beyond tergal margin with deep articular furrow, extending entire length of tergal margin.
Tergum (Figure 21K,L) apex blunt, growth lines prominent, crossed by strong longitudinal ridges giving a lattice-like or cancellated appearance, fringed with short, inconspicuous setae; articular margin concave in apical half. Carinal margin straight, only curved near apex; basal margin straight; spur length 0.3 times width, occupying approximately 0.4 of basal margin, positioned less than half own width from scutal margin; spur furrow defined by abrupt change in growth lines, margins infolded, furrow open in basal half; distal margin concave with disto-articular angle projecting. Internally smooth, crests for depressor muscles faint. Articular furrow broad, articular ridge low, rounded occupying 0.25 scutal margin.
Labrum (Figure 22A,B) bilobed, divided by deep, medial, v-shaped notch, each rounded lobe with 2 marginal teeth and numerous fine setae.
Mandibular palp (Figure 22A,C) rhomboid, distal end obliquely truncate, anterior margin concave, posterior margin straight; setae heavily serrulate, becoming longer and more dense distally; distal angle with long, simple setae.
Mandible (Figure 22D–F) cutting margin straight with 4 distinct teeth, second to fourth tooth bifid, fifth tooth congruent with molariform or spinose inferior angle; superior and inferior margins with row of long, fine simple setae, dense short setae covering inner and outer faces.
Maxillule (Figure 22G,H) cutting margin with 11 cuspidate setae, upper and lower pair larger, more robust than remaining setae. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxilla (Figure 22I) bilobed, lobes ovate, serrulate setae arranged on anterior margins, becoming more dense at apex, longer on distal lobe than those on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 23A,B) protopod with pappose setae basally on protopod posterior margin. Rami unequal, anterior ramus 2.5 times length of posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus II (Figure 23C,D) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami unequal, anterior ramus 1.25 times longer that posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus III (Figure 23E,F) protopod anterior margin lined with long plumose setae, posterior margin with pappose setae basally, plumose setae distally. Rami subequal, setation lasiopod, segments with serrulate and simple setae on mesial face, less densely arranged than on cirrus I and II. Anterior ramus with row of ctenoid scales on anterodistal portion of all segments, except terminal segment, most prominent on medial segments.
Cirrus IV (Figure 24A–E) posterior margin of protopod basal segment margin lined with plumose setae, tuft of simple, short setae at posterodistal angle; basis elongate, length 2.09–2.15 times width, with tuft of simple, short setae at posterodistal angle; anterior margin lined with 14–15 closely-spaced, hooked teeth. Rami subequal, setation ctenopod. Anterior ramus with 1-4 hooked teeth on first 8 segments, distal segments with up to 3 erect spines on anterior margins; long serrulate setae arranged in row at anterodistal angle. Posterior ramus lacking hooked teeth or erect spines on anterior margins; 3–6 pairs of long, serrulate setae arranged in row at anterodistal angle.
Cirrus V (Figure 24F–H) protopod with sparse setae on anterior margins, tufts of short, simple setae at posterodistal angles; basis elongate, length 1.7–1.75 times width, without hooked teeth. Rami subequal, setation ctenopod; anterior ramus with 1-3 teeth on segments 4–13, basal 3 segments without teeth; 2–3 pairs of serrulate setae on distal portion of anterior margins of each segment; posterior ramus setation as in anterior ramus, segments without hooked teeth or erect spines.
Cirrus VI (Figure 24I,J) protopod with sparse setae on anterior margins, tufts of short, simple setae at posterodistal angles; basis elongate, length 1.78–1.97 width, without hooked teeth. Rami equal, setation ctenopod, 2–3 pairs of long, serrulate setae on distal portion of anterior margins of each segment.
Penis long, annulated, with sparse, short setae; basidorsal point low, rounded.
Distribution: Australia, from Perth north to the Kimberley region, south to New South Wales and Victoria. South Africa, Andaman Islands, Java Sea, Sulu Archipelago, Malay Archipelago, South China Sea, Hong Kong, China, Philippines, Japan. Depth: 0–170 m.
Hosts: Trachytedania L1, Trachytedania L2, Trachytedania MM1, and Tedania sp.1 (all Tedaniidae). Previous records: Halichondria (Halichondria) okadai (Kadota, 1922) [80], H. (H.) japonica (Kadota, 1922) [7,69,80]; Tedania sp. [81]; and Halichondriidae [5].
Remarks: This distinctive species has been reported across the Indo-Pacific, but host identifications have been rare. Morphological variation has been reported in the armature of cirri IV and V with regards to the number of teeth present on the protopodite and anterior ramus, as well as the degree of the sculpturing on the shell plates, including the scutum [7,69]. Given the vast geographic range reported for this species, this raises the potential for more than one species to be present. These records provide the first COI sequences for this species. Coupling these with the description and host identifications will greatly aid further workers in identifying this species and distinguishing any potential cryptic species.
The remaining species currently assigned to the genus Pectinoacasta can be readily separated from P. pectinipes. Firstly, P. cancellorum and P. sculpturata may be considered fenestrate, but in addition to the small membranous windows between parietes are longitudinal slits within parietal sections of the wall. These two species also have a yellow chitinous cuticle on the external surface of the tergum, and their respective tergal spurs are also distinctly more elongate. Pectinoacasta angusticalcar (Broch, 1931) [6] does not have fenestra between the parietes, the tergal spur is elongate, and the furrow is completely closed along its length. Lastly, in P. zevinae (Kolbasov, 1991) [61], the orifice is large, the parietes do not curve over the opercular plates, exposing the opercular plates from above, the tergum lacks a distinct spur or spur furrow, and the armature of the posterior cirri is reduced to small erect spines, lacking the hooked teeth of P. pectinipes.
Figure 21.
Pectinoacasta pectinipes (Pilsbry, 1916) shell and shell plates (A–E, I–L: WAM C67772; F–H: WAM C58870), (A–D) left, right, rostral and carinal views, respectively, of whole shell; (E) membranous slit between junction of rostrum and right lateral; (F) internal view of rostrum, (G) internal view of right lateral and carinolateral; (H) internal view of basis; (I,J) external and internal views of scutum; (K,L) internal and external views of tergum. Scale bars: A–D = 5 mm; E = 0.5 mm; F–H = 1 mm; I–L = 2 mm.
Figure 21.
Pectinoacasta pectinipes (Pilsbry, 1916) shell and shell plates (A–E, I–L: WAM C67772; F–H: WAM C58870), (A–D) left, right, rostral and carinal views, respectively, of whole shell; (E) membranous slit between junction of rostrum and right lateral; (F) internal view of rostrum, (G) internal view of right lateral and carinolateral; (H) internal view of basis; (I,J) external and internal views of scutum; (K,L) internal and external views of tergum. Scale bars: A–D = 5 mm; E = 0.5 mm; F–H = 1 mm; I–L = 2 mm.

Figure 22.
Pectinoacasta pectinipes (Pilsbry, 1916) mouthparts (WAM C58871) (A) labrum and right mandible palp; (B) crest of labrum; (C) left mandible palp; (D) right mandible; (E) close up of inferior angle of right mandible; (F) left mandible; (G,H) right and left maxillules; (I) right and left maxillae. Scale bars: A, C,D, F–I = 100 µm; B, E = 50 µm.
Figure 22.
Pectinoacasta pectinipes (Pilsbry, 1916) mouthparts (WAM C58871) (A) labrum and right mandible palp; (B) crest of labrum; (C) left mandible palp; (D) right mandible; (E) close up of inferior angle of right mandible; (F) left mandible; (G,H) right and left maxillules; (I) right and left maxillae. Scale bars: A, C,D, F–I = 100 µm; B, E = 50 µm.

Figure 23.
Pectinoacasta pectinipes (Pilsbry, 1916) cirri I–III (WAM C58871). (A) Left cirrus I; (B) terminal segment of cirrus I anterior ramus; (C) left cirrus II; (D) terminal segment of left cirrus II posterior ramus; (E) left cirrus III; (F) medial segments of cirrus III anterior ramus. Scale bars: A, C, E = 200 µm; B, D, F = 50 µm.
Figure 23.
Pectinoacasta pectinipes (Pilsbry, 1916) cirri I–III (WAM C58871). (A) Left cirrus I; (B) terminal segment of cirrus I anterior ramus; (C) left cirrus II; (D) terminal segment of left cirrus II posterior ramus; (E) left cirrus III; (F) medial segments of cirrus III anterior ramus. Scale bars: A, C, E = 200 µm; B, D, F = 50 µm.

Figure 24.
Pectinoacasta pectinipes (Pilsbry, 1916) cirri IV–VI (WAM C58871). (A) left cirrus IV; (B,C) left and right cirrus IV protopod basis; (D) hooked teeth on medial segments of left cirrus IV anterior ramus; (E) setation on distal segments of left cirrus IV anterior ramus; (F) left cirrus V; (G) hooked teeth on medial segments of right cirrus V anterior ramus; (H) protopod of left cirrus V; (I) left cirrus VI; (J) medial segments of left cirrus VI. Scale bars: A, F, I = 200 µm; B, D,E = 50 µm; C, G,H, J = 100 µm.
Figure 24.
Pectinoacasta pectinipes (Pilsbry, 1916) cirri IV–VI (WAM C58871). (A) left cirrus IV; (B,C) left and right cirrus IV protopod basis; (D) hooked teeth on medial segments of left cirrus IV anterior ramus; (E) setation on distal segments of left cirrus IV anterior ramus; (F) left cirrus V; (G) hooked teeth on medial segments of right cirrus V anterior ramus; (H) protopod of left cirrus V; (I) left cirrus VI; (J) medial segments of left cirrus VI. Scale bars: A, F, I = 200 µm; B, D,E = 50 µm; C, G,H, J = 100 µm.

Pectinoacasta sculpturata (Broch, 1931) [6] (Figure 25, Figure 26, Figure 27 and Figure 28)
Acasta sculptura: Newman and Ross 1976: p. 54 [44].
Material examined: Holotype: ZMUC CRU9003, 8 mm RC, Indonesia, Java Sea 5°47’ S 106°7’ E, 49 m, Danish Expedition to the Kei Islands, stn. 107, 5 Aug 1922, Host: Agelas sp.
Paratypes: ZMUC CRU9003, 3.5 mm RC, more specimens still with host, same data as holotype.
Other material: WAM C58282, WAM C61394, WAM C61395, 3 slides, WAM C61396, all from Pansy Island, Montebello Islands, WA, 20°21′19″ S 115°33′02″ E, 26 m, NCB Montebello and Muiron Islands Expedition, stn. 3/MM15, 15 Apr 2015, coll. A.M. Hosie & A. Hara, host: WAM Amphinomia sulphurea. WAM C74480, Y Island, Exmouth Gulf, WA, 9–12.8 m, 21°57′19″ S 114°24′29″ E, Bush Blitz Cape Range Expedition, stn. CR19/07, 19 Jun 2019, coll. A.M. Hosie & A. Hara, host: WAM Z88930 Amphinomia sulphurea. WAM C75199, Ningaloo Reef, Ningaloo Marine Park, 22°40′52.61″ S 113°36′05.76″ E, CReefs Ningaloo Expedition, stn. NR09-025, 18 May 2009, coll. J. Fromont, host: WAM Z45362 Amphinomia sulphurea. QM W29555, southern Great Barrier Reef, QLD, 21°46’38.3″ S 151°50’12.9″ E, 57.6 m, CSIRO SBD Project, 26 Nov 2005, coll. S.D. Cook, host: Agelas nemoechinata.
Diagnosis: Shell globose, parietes permeated with narrow, longitudinal incisions extending from basal rim and small windows at parietal junctions; externally surfaces studded with short, blunt, calcareous projections; internally longitudinal ribs extending below sheath and interlock with crenate basal rim. Scutum with longitudinal ridges crossing growth ridges. Tergum with setose, yellow, chitinous cuticle, growth ridges crossed with longitudinal ridges, giving a latticed appearance. Cirrus IV protopod lacking curved teeth, anterior margin with longitudinal row of small spinules, anterior ramus basal segments with 1–2 recurved teeth on anterior margins.
Description: Shell (Figure 25A–L) globose, white or grey; orifice small relative to shell diameter, rim toothed. Parietes with longitudinal incisions extending from basal rim to over half parietal height, creating narrow, membranous slits, incisions secondarily calcified in upper portions of parietes; external surfaces with short, blunt calcareous projections arranged irregularly, or in longitudinal rows where confined by incisions; yellow, chitinous cuticle on basal sections; parietes strongly curved toward orifice. Radii and alae not reaching basal rim, leaving small membranous slits between parietes; broad with striations perpendicular to oblique summits. Rostrum widest shell plate. Internally, sheath with conspicuous horizontal growth lines; lower margin adpressed to inner lamina, extending to just above apical extent of parietal incisions; longitudinal ribs below sheath formed either side of incisions.
Basis circular, shallow to deeply cupped; concentric growth lines conspicuous; basal rim irregularly crenate, interlocking with internal ribs of parietes.
Scutum (Figure 25M,N) triangular, approximately as high as wide, growth lines marked by longitudinal ridges, ridges increasing in number basally; basitergal angle rounded, tergal section narrow; occludent margin strongly toothed. Internally smooth, apex with yellow chitinous cuticle, adductor and lateral depressor muscle pits present; articular ridge short, occupying 0.5 length of tergal margin, prominently protruding beyond tergal margin with deep articular furrow extending entire length of tergal margin.
Tergum (Figure 25O,P) apex blunt, external surface covered by yellow, setose, chitinous cuticle; growth lines prominent, crossed by strong longitudinal ridges, giving a latticed or cancellated appearance, fringed with short, inconspicuous setae; articular margin concave in apical half. Carinal margin straight, only curved near apex; basal margin straight; spur length 2 times width, occupying approximately 0.3 basal margin, positioned less than half own width from scutal margin; spur furrow defined by abrupt change in growth lines, margins infolded, furrow open in basal half; distal margin rounded. Internally roughened, chitinous cuticle extending beyond carinal margin of calcareous part, crests for depressor muscles distinct. Articular furrow narrow, articular ridge low, rounded occupying 0.25 scutal margin.
Labrum (Figure 26A) bilobed, divided by deep, medial, v-shaped notch, each rounded lobe with 2 prominent, marginal teeth and numerous fine setae.
Mandibular palp (Figure 26B) rhomboid, distal end obliquely truncate, anterior margin concave, posterior margin straight; setae heavily serrulate, becoming longer and more dense distally; distal angle with long, simple setae.
Mandible (Figure 26C,D) with five teeth, second to fourth teeth bifid, fifth tooth confluent with molariform inferior angle; inferior angle with denticles. Regularly spaced, dense, short setae on inner and outer faces, longer fine setae on inferior and superior margin.
Maxillule (Figure 26E–G) cutting margin straight, with 10 cuspidate setae, upper and lower pairs longer than remaining setae; three medial, shorter setae serrulate on inferior margin; tuft of short setae at basal angle. Dense, short setae regularly spaced on inner and outer faces, longer fine setae on inferior and superior margin.
Maxilla (Figure 26H) bilobed, lobes ovate, serrulate setae arranged on anterior margins, becoming more dense at apex, longer on distal lobe than those on basal lobe.
Cirral segment counts are given in Table 2.
Cirrus I (Figure 27A) protopod with pappose setae on basal segment posterior margin, basis with tuft of setae at posterodistal angle. Rami unequal, anterior ramus 3.5 times length of posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face, tuft of simple setae on posterodistal angle.
Cirrus II (Figure 27B) protopod lined with long serrulate setae on anterior margin, with pappose or plumose setae on posterior margin on basal segment; basis with long serrulate setae on anterior margin. Rami unequal, anterior ramus 1.25 times longer that posterior ramus, setation lasiopod, segments with densely arranged serrulate and simple setae on mesial face.
Cirrus III (Figure 27C–E) protopod lined with long, serrulate setae on anterior margin, pappose or plumose setae on posterior margin of basal segment. Rami subequal, anterior ramus 1.1 times length of posterior ramus, setation lasiopod, segments with serrulate and simple setae on mesial face, less densely arranged than on cirrus I and II. Anterior ramus with transverse row of distally directed spines at anterodistal angle on all but distal-most segments, spines most prominent on medial segments; posterodistal angle with transverse row of ctenoid scales. Posterior ramus with similar spination but much reduced in size.
Cirrus IV (Figure 28A–F) protopod basal segment anterior and posterior margins with well-spaced row of short, simple setae; tuft of simple, short setae at posterodistal angle; dense row of short, sharp spinules on anterior margin; basis elongate, length 1.80–1.91 times width, with tuft of simple, long setae at posterodistal angle; anterior margin lined with short, simple and short, sharp spinules. Rami subequal in length, setation ctenopod. Anterior ramus with 1-2 hooked teeth on anterior margins of first 10 segments, anterodistal angles with transverse row of erect spines on all but distal-most segments; 3 pairs of serrulate setae on medial segments. Posterior ramus with similar setation to anterior ramus, with erect spines on anterodistal angles, but lacking hooked teeth.
Cirrus V (Figure 28G) and VI (Figure 28H,J) similar, protopod with well-spaced, long setae and dense row of short spinules on anterior margins; posterior margin lacking setae; basis elongate, cirrus V length 1.76–1.82 width, cirrus VI length 1.71–1.75 width, hooked teeth absent, row of long simple setae on anterior margin, tuft of setae at posterodistal angle. Rami subequal, setation ctenopod; 3 pairs of serrulate setae on distal portion of anterior margins of each segment; rami without teeth or erect spines.
Penis longer than cirrus VI, annulated sparsely setose. basidorsal point (Figure 28I) prominent, triangular, as wide as high.
Distribution: Australia: Ningaloo to Montebello Islands, WA; southern Great Barrier Reef, QLD. Java Sea Philippines, Taiwan. 12–97 m.
Hosts: Agelas sp. (type host), A. nemoechinata Hoshino, 1985 [83], and Amphinomia sulphurea Hooper, 1991 [84] (all Agelasidae). Previous records: “Alcyonarian” [6]; Agelas nemoechinata Hoshino, 1985 [17].
Remarks: Re-examination of the type series of this species was key in determining the identity of the Australian material, as the original description failed to mention the chitinous cuticle, a rather prominent feature of the species. The type series of this species includes mounted slide material, disarticulated shell plates, extracted whole specimens, and specimens still embedded in the host. Unfortunately, it is not possible to connect the slides to the dissected shell plates. It is also not clear which, if any, of these specimens is the one reportedly extracted from an alcyonacean. The specimen designated as “type” in Broch’s figure 35a is clearly still extant and whole. It is this specimen we treat as being the holotype.
Distinguishing P. sculpturata from P. cancellorum and P. pectinipes has been summarised under the treatment of these latter species. The remaining species of Pectinoacasta, P. angusticalcar and P. zevinae, both lack the incisions in the parietes seen in P. sculpturata. Further to this, P. zevinae lacks a distinct tergal spur or teeth on the protopod or rami of cirrus IV. While in P. angusticalcar, the spur furrow is fully closed, the scutum is distinctly wider than high, and the protopod of cirrus IV has a row of teeth. Given Broch’s omission of the chitinous cuticle in, it is not clear if it was similarly omitted from his description of P. angusticalcar in the same paper [6].
Examination of the type-host reveals it to be a species of Agelas, albeit distinct from the species inhabited by the Australian specimens, as well as the Agelas inhabited by P. cancellorum. The addition of A. sulphurea as a host broadens the generic range but affirms this species’ affinity for sponges of the Agelasidae.
3.2. Molecular Results
The top-ranked partitions produced by the ASAP analyses suggested between 26 or 28 putative species. The three substitution models provided similar results and the first two highest-ranked partitions received close scores. Under the JC69 model and p-distance partitions of 26 species and 28 species were received an ASAP score of 2.0 and 2.50, respectively, at a distance threshold of 7% or 4%. The difference between these partitions was the lumping or splitting of A. huangi and A. sp.2, and A. sandwichi and A. caveata sp. nov. The K2P model ranked partitioning the dataset into 26 or 27 species with scores of 1.0 and 3.0, respectively, at a distance threshold of 2.9% or 3.5%. The difference between these two partitions is the lumping or splitting of A. sandwichi and A. caveata sp. nov.
The intraspecific and the interspecific K2P genetic distances ranged from 0–0.019 and 0.060−0.261, respectively across the dataset (Figure 29, Table S1). The intraspecific K2P distances of the three new species were: A. caveata sp. nov. 0.006, E. acutaflava sp. nov. 0.005, E. excoriatrix sp. nov. 0.002. The greatest intraspecific distance was within P. pectinipes (0.19), while Acasta sandwichi and A. caveata sp. nov. exhibited the closest interspecific genetic similarity (0.060).
IQTree selected the general time-reversible model with a proportion of invariant sites and gamma-distributed rate heterogeneity (GTR+I+G) as this was ranked highest under the Bayesian information criterion [85] and this was utilised in the BI analysis as well. The phylogenetic tree of sponge barnacles, with ML and BI support values, exhibited very similar topologies and statistical support (Figure 29). All new species concepts presented herein formed reciprocally monophyletic clades under all analyses; however, none of the genera were recovered as monophyletic. Although there was little support for the branching patterns observed except at nodes separating closely related species such as the nodes between P. cancellorum and P. sculpturata and the clade containing A. undulaterga, A. radenta, A. huangi, and A. sp. 2 MCY-2020.
Figure 25.
Pectinoacasta sculpturata (Broch, 1931) shell and shell plates (A–C, holotype ZMUC CRU9003; D, ZMUC CRU9003; E–L, ZMUC CRU9003; M–P, WAM C61395). (A–D) left, right and top views, respectively, of whole specimens; (E–H) external views of disarticulated rostrum, right latus, right carinolatus and carina; (I–L) internal views of same; (M,N) external and internal views of right scutum; (O,P) internal and external views of right tergum. Scale Bars: A–P = 2 mm.
Figure 25.
Pectinoacasta sculpturata (Broch, 1931) shell and shell plates (A–C, holotype ZMUC CRU9003; D, ZMUC CRU9003; E–L, ZMUC CRU9003; M–P, WAM C61395). (A–D) left, right and top views, respectively, of whole specimens; (E–H) external views of disarticulated rostrum, right latus, right carinolatus and carina; (I–L) internal views of same; (M,N) external and internal views of right scutum; (O,P) internal and external views of right tergum. Scale Bars: A–P = 2 mm.

Figure 26.
Pectinoacasta sculpturata (Broch, 1931) mouthparts (WAM C61395). (A) crest of labrum; (B) right mandibular palp; (C,D) left and right mandibles; (E) left maxillule; (F) serrated setae on left maxillule; (G) right maxillule; (H) left and right maxillae. Scale bars: A = 100 µm; B–F, H = 200 µm; G = 50 µm.
Figure 26.
Pectinoacasta sculpturata (Broch, 1931) mouthparts (WAM C61395). (A) crest of labrum; (B) right mandibular palp; (C,D) left and right mandibles; (E) left maxillule; (F) serrated setae on left maxillule; (G) right maxillule; (H) left and right maxillae. Scale bars: A = 100 µm; B–F, H = 200 µm; G = 50 µm.

Figure 27.
Pectinoacasta sculpturata (Broch, 1931) cirri I–III (WAM C61395). (A) left cirrus I; (B) left cirrus II; (C) right cirrus III; (D,E) detail of spines on medial segments of anterior ramus of left and right cirrus III, respectively. Scale bars: A–C = 200 µm; D–E = 50 µm.
Figure 27.
Pectinoacasta sculpturata (Broch, 1931) cirri I–III (WAM C61395). (A) left cirrus I; (B) left cirrus II; (C) right cirrus III; (D,E) detail of spines on medial segments of anterior ramus of left and right cirrus III, respectively. Scale bars: A–C = 200 µm; D–E = 50 µm.

Figure 28.
Pectinoacasta sculpturata (Broch, 1931) cirri IV–VI (WAM C61395). (A) left cirrus IV; (B) spinules on the anterior margin of cirrus IV protopod; (C–E) teeth and erect spines on basal segments of cirrus IV anterior ramus; (F) erect spines on medial segment of cirrus IV posterior ramus; (G) right cirrus V; (H) right cirrus VI; (I) basidorsal point of penis; (J) setation on medial segments of cirrus VI. Scale bars: A, G–H = 500 µm; B, E–F, J = 50 µm; C–D, I = 100 µm.
Figure 28.
Pectinoacasta sculpturata (Broch, 1931) cirri IV–VI (WAM C61395). (A) left cirrus IV; (B) spinules on the anterior margin of cirrus IV protopod; (C–E) teeth and erect spines on basal segments of cirrus IV anterior ramus; (F) erect spines on medial segment of cirrus IV posterior ramus; (G) right cirrus V; (H) right cirrus VI; (I) basidorsal point of penis; (J) setation on medial segments of cirrus VI. Scale bars: A, G–H = 500 µm; B, E–F, J = 50 µm; C–D, I = 100 µm.

Figure 29.
Maximum likelihood (ML) tree based on COI dataset. Tips are labelled with institutional accession numbers for newly published sequences (coloured by species) or GenBank accession numbers for previously published data (black). Node labels are supporting bootstrap (bs) and posterior probability (pp) values, respectively, with only values ≥ 70% bs or 0.7 pp shown (- indicates where only one method did not meet these thresholds). Scale bar indicates number of expected substitutions per site.
Figure 29.
Maximum likelihood (ML) tree based on COI dataset. Tips are labelled with institutional accession numbers for newly published sequences (coloured by species) or GenBank accession numbers for previously published data (black). Node labels are supporting bootstrap (bs) and posterior probability (pp) values, respectively, with only values ≥ 70% bs or 0.7 pp shown (- indicates where only one method did not meet these thresholds). Scale bar indicates number of expected substitutions per site.

4. Discussion
The present study documents an additional six species of the Australian Acastinae; however, there are still many more species to be formally described, and the total number of species is likely to be well over 60 [4]. Hosts for most of the previously reported barnacle species are still unknown (e.g. [86,87]), and given how integral this is to understanding barnacle biodiversity and patterns of speciation [1,4,17], it is the other key gap in our knowledge.
Most of the Australian specimens documented in this study were collected from a restricted host range, typically inhabiting a single or few sponge genera. Only A. aspera appears to have a broad host range, with hosts from multiple families, but all from the same sponge order (Poecilosclerida). Species such as P. pectinipes and A. fenestrata have been documented from multiple host orders in other parts of their geographic range. However, once more material of A. fenestrata is sequenced and type specimens examined, our expectation is for a more restricted range of hosts.
In the case of P. sculpturata, molecular data proved that specimens reported from Taiwan [17] are conspecific with the Australian specimens, while identifying the type-host shows that this species appears to specialise in species within the genera of the Agelasidae: Agelas and Amphinomia (this latter genus is monotypic). However, the report of P. sculpturata inhabiting an alcyonacean coral [6] requires further investigation, as it seems unlikely for a species to specialise in one family of sponges but also embed in hosts from a different phylum. Pectinoacasta cancellorum was recovered as sister to P. sculpturata and is not only morphologically similar, but also inhabits species of Agelas, albeit different species. Both the genetic results and host used show these two species to be rather distantly related to P. pectinipes, suggesting that the morphological similarities are superficial.
The proposed new species, A. caveata sp. nov., E. acutaflava sp. nov., and E. excoriatrix sp. nov., were each identified from single host genera, but it is too soon to tell if they are host specialists, as they have so far been identified from only a few host individuals. The factors involved in determining the range of hosts used by individual barnacle species are unknown and would be a valuable avenue of study.
It was already known that A. aspera was distributed in Australia and Taiwan with no discernible genetic differentiation [3]. In an additional three species, specimens from Taiwanese waters were nested within clades of Western Australian specimens: A. sandwichi, A. fenestrata and P. sculpturata. Interestingly, the single sequence of A. fenestrata from Queensland was sister to the clade formed by the WA and Taiwanese specimens. This result suggests that there is potentially greater connectivity between WA and Taiwan than with the east coast of Australia, for this species at least, but further specimens and sequences will be needed to test this hypothesis.
The types of armature found on the fourth cirrus have long been seen as critical characters upon which to base taxonomic decisions at the species level or above [5,6]. The key morphological distinction between A. sandwichi and A. caveata sp. nov. is that the former bears a single, straight, and sharp “denticle” on each segment [2] compared to the multiple, robust, and hooked teeth of the latter. The close, but distinct, genetic relationship of these apparent sister species indicates that the evolution, or loss, of these character states may occur independently in different lineages. The ASAP analysis did not provide unequivocal support for separating these species. However, this analysis is looking for a partitioning threshold that can give predictive power to molecular identification as a starting point for assessing biological or adaptive significance [25]. The interspecific genetic distance between these two species (0.60) is still much higher than the greatest intraspecific distance (0.19) observed in this dataset and supports the hypothesis that the specimens represent separate species. Therefore, while there is more work needed on the intraspecific variation of the fourth cirrus in the Acastinae, it is considered a reliable character for delimiting species and a key support for erecting this new species and is believed to be a functional adaptation to the host [5,16]. Further evidence that supports this decision includes the separate host families (Niphatidae and Petrosiidae) suggesting ecological separation and that the Australian specimens identified as A. sandwichi are indistinguishable morphologically and genetically from those from Taiwan but are easily separated from those of A. caveata sp. nov.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d13070290/s1, Table S1: genetic distances.
Author Contributions
Conceptualization, A.M.H., J.F., K.M., and D.S.J.; methodology, A.M.H.; formal analysis, A.M.H.; investigation, A.M.H. and J.F.; resources, A.M.H., J.F., and K.M.; data curation, A.M.H. and J.F.; writing—original draft preparation, A.M.H.; writing—review and editing, A.M.H., J.F., K.M., and D.S.J.; visualization, A.M.H.; supervision, K.M., J.F., and D.S.J.; project administration, A.M.H., J.F., and K.M.; funding acquisition, A.M.H., J.F., and D.S.J. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Australian Biological Resources Study Grant #RF213-25; Gorgon Barrow Island Net Conservation Benefits Fund; WAM/Woodside Dampier & Kimberley Biodiversity Projects; Bush Blitz Species Discovery Program; Total Corporate Foundation, Paris; Western Australian Marine Science Institution Kimberley Marine Research Program (Project 1.1.1 Benthic Biodiversity and Dredging Science Node (Project 6.3 Field Surveys: Sponges), Census of Marine Life CReefs program and the Australian Government Research Training Program.
Institutional Review Board Statement
Ethical review and approval were waived for this study, as the animals (marine sponges and barnacles) used in this study are invertebrates that are not subject to animal ethics regulations in Australia.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in article or Supplementary Materials.
Acknowledgments
The authors would like to thank the following people and groups for their assistance during this study. Merrick Ekins (QM) identified sponge specimens from the Queensland Museum and made them available for study. Danny Eibye-Jacobsen (ZMUC) and Shigeyuki Yamato (SMBL) provided loans of the essential type specimens examined herein. Ana Hara, Oliver Gomez, Linette Umbrello, Kara Layton, Nerida Wilson (all WAM), and Jarunee Siengsanan-Lamont (Curtin University) assisted with specimen handling, processing, and sequencing in the lab and field. The comments of two reviewers greatly improved the manuscript. We are grateful to the Traditional Owners of the sea country where the studied specimens were collected.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Van Syoc, R.J.; Van Soest, R.W.; Xavier, J.R.; Hooper, J.N. A phylogenetic overview of sponge-inhabiting barnacles and their host specificity (Crustacea, Cirripedia). Proc. Calif. Acad. Sci. Ser. 2015, 4, 331–357. [Google Scholar]
- Yu, M.-C.; Chan, B.K.K.; Achituv, Y.; Kolbasov, G.A. Four new sponge-inhabiting barnacles of the genus Acasta (Thoracica: Archaeobalanidae: Acastinae) from the Indo-Pacific. Raffles Bull. Zool. 2017, 65, 585–615. [Google Scholar]
- Yu, M.-C.; Kolbasov, G.A.; Hosie, A.M.; Lee, T.-M.; Chan, B.K.K. Descriptions of four new sponge-inhabiting barnacles (Thoracica: Archaeobalanidae: Acastinae). Zootaxa 2017, 4277, 151–198. [Google Scholar] [CrossRef] [PubMed]
- Hosie, A.M.; Fromont, J.; Munyard, K.; Wilson, N.G.; Jones, D.S. Surveying keratose sponges (Porifera, demospongiae, Dictyoceratida) reveals hidden diversity of host specialist barnacles (Crustacea, Cirripedia, Balanidae). Mol. Phylogenetics Evol. 2021, 161, 107179. [Google Scholar] [CrossRef]
- Kolbasov, G.A. Revision of the genus Acasta Leach (Cirripedia: Balanoidea). Zool. J. Linn. Soc. 1993, 109, 395–427. [Google Scholar] [CrossRef]
- Broch, H. Papers from Dr. Th. Mortensen’s Pacific expédition 1914–1916. LVI. Indomalayan Cirripedia. Vidensk. Medd. Fra Dan. Nat. Foren. 1931, 91, 1–142. [Google Scholar]
- Hiro, F. Studies on cirripedian fauna of Japan II: Cirripeds found in the vicinity of the Seto marine biological laboratory. Mem. Coll. Sci. Kyoto Imp. University. Ser. B 1937, 12, 385–478. [Google Scholar]
- Leach, W.E. The Zoological Miscellany: Being Descriptions of New Or Interesting Animals; E. Nodder & Sons: London, UK, 1817; Volume 3. [Google Scholar]
- Say, T. An account of some marine shells of the United States. J. Acad. Nat. Sci. Phila. 1822, 2, 221–248; 302–325. [Google Scholar]
- Hoek, P.P.C. The Cirripedia of the Siboga-expedition. B. In Siboga-Expeditie Mongographes; E.J. Brill: Leyden, The Netherlands, 1913; Volume XXXIb, pp. 129–275. [Google Scholar]
- Kolbasov, G.A.; Chan, B.K.K.; Molodstova, T.N.; Achituv, Y. Revision of the coral-inhabiting genus Conopea (Cirripedia: Archaeobalanidae) with description of two new species of the genera Conopea and Acasta. Zootaxa 2016, 4178, 182–208. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Harp, M.; Høeg, J.T.; Achituv, Y.; Jones, D.; Watanabe, H.; Crandall, K.A. The tempo and mode of barnacle evolution. Mol. Phylogenetics Evol. 2008, 46, 328–346. [Google Scholar] [CrossRef]
- Tsang, L.M.; Chu, K.H.; Nozawa, Y.; Chan, B.K.K. Morphological and host specificity evolution in coral symbiont barnacles (Balanomorpha: Pyrgomatidae) inferred from a multi-locus phylogeny. Mol. Phylogenetics Evol. 2014, 77, 11–22. [Google Scholar] [CrossRef]
- Chan, B.K.K.; Dreyer, N.; Gale, A.S.; Glenner, H.; Ewers-Saucedo, C.; Pérez-Losada, M.; Kolbasov, G.A.; Crandall, K.A.; Høeg, J.T. The evolutionary diversity of barnacles, with an updated classification of fossil and living forms. Zool. J. Linn. Soc. 2021, zlaa160. [Google Scholar] [CrossRef]
- Yu, M.-C.; Kolbasov, G.; Chan, B.K.K. A new species of sponge inhabiting barnacle Bryozobia (Archaeobalanidae, Bryozobiinae) in the West Pacific. ZooKeys 2016, 571, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Van Syoc, R.J.; Winther, R. Sponge-inhabiting barnacles of the Americas: A new species of Acasta (Cirripedia, Archaeobalanidae), first record from the eastern Pacific, including discussion of the evolution of cirral morphology. Crustaceana 1999, 72, 467–486. [Google Scholar]
- Yu, M.-C.; Dreyer, N.; Kolbasov, G.A.; Høeg, J.T.; Chan, B.K.K. Sponge symbiosis is facilitated by adaptive evolution of larval sensory and attachment structures in barnacles. Proc. R. Soc. B 2020, 287, 20200300. [Google Scholar] [CrossRef]
- Hosie, A.M.; Fromont, J.; Munyard, K.; Jones, D.S. Description of a new species of Membranobalanus (Crustacea, Cirripedia) from southern Australia. ZooKeys 2019, 873, 25–42. [Google Scholar] [CrossRef]
- Pitriana, P.; Wessel, A.; Aschenbach, T.; Von Rintelen, K. Exploring sponge-inhabiting barnacles of eastern Indonesia using micro-CT scanning. Treubia 2020, 47, 77–98. [Google Scholar] [CrossRef]
- Fromont, J.; Abdul Wahab, M.A.; Gomez, O.; Ekins, M.; Grol, M.; Hooper, J.N.A. Patterns of sponge biodiversity in the Pilbara, Northwestern Australia. Diversity 2016, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Heyward, A.; Fromont, J.; Schönberg, C.H.L.; Colquhoun, J.; Radford, B.; Gomez, O. The sponge gardens of Ningaloo reef, Western Australia. Open Mar. Biol. J. 2010, 4, 3–11. [Google Scholar]
- Hooper, J.N.; Kennedy, J.A.; Quinn, R.J. Biodiversity ‘hotspots’, patterns of richness and endemism, and taxonomic affinities of tropical Australian sponges (Porifera). Biodivers. Conserv. 2002, 11, 851–885. [Google Scholar] [CrossRef]
- Chan, B.K.K.; Garm, A.; Høeg, J. Setal morphology and cirral setation of thoracican barnacle cirri: Adaptations and implications for thoracican evolution. J. Zool. 2008, 275, 294–306. [Google Scholar] [CrossRef]
- Meyer, C.P. Molecular systematics of cowries (Gastropoda: Cypraeidae) and diversification patterns in the tropics. Biol. J. Linn. Soc. 2003, 79, 401–459. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2020, 1–12. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. Mamm. Protein Metab. 1969, 3, 21–132. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Schwartz, T.; Hoover, P.; Yoshimoto, K.; Sivagnanam, S.; Majumdar, A. The CIPRES workbench: A flexible framework for creating science gateways. In Proceedings of the 2015 XSEDE Conference: Scientific Advancements Enabled by Enhanced Cyberinfrastructure, St. Louis, MO, USA, 26–30 July 2015; pp. 1–8. [Google Scholar]
- Van Syoc, R.; Carrison-Stone, D.; Madrona, L.; Williams, G. Barnacle symbionts of gorgonian sea fans, description of seven new species (Archaeobalanidae: Cirripedia) from the Philippines, Including a Key to the Western Pacific species of Conopea. In The Coral Triangle, The 2011 Hearst Philippine Biodiversity Expedition; Williams, G., Gosliner, T., Eds.; California Academy of Sciences: San Francisco, CA, USA, 2014; pp. 171–211. [Google Scholar]
- Pilsbry, H.A. The sessile barnacles (Cirripedia) contained in the US National Museum; including a monograph of the American species. Bull. United States Natl. Mus. 1916, 93, 1–366. [Google Scholar]
- Hentschel, E. Tetraxonida. 2. Teil. In Die Fauna Südwest-Australiens. Ergebnisse der Hamburger südwest-australischen Forschungsreise 1905; Michaelsen, W., Hartmeyer, R., Eds.; Gustav Fischer: Jena, Germany, 1911; Volume 3, pp. 279–393. [Google Scholar]
- Kolbasov, G.A. Acasta pertusa sp n (Cirripedia, Thoracica) from the Red Sea. Zool. Zhurnal 1990, 69, 142–145. [Google Scholar]
- Gravier, C. Sur deux espèces de Cirripèdes du genre Acasta Leach vivant a la côte française des Somalis. Bull. Muséum D’histoire Nat. Paris 1921, 353–357. [Google Scholar]
- Darwin, C. A Monograph on the Sub-Class Cirripedia; The Ray Society: London, UK, 1854. [Google Scholar]
- Kolbasov, G.A. Two new species of Acasta (Cirripedia, Thoracica) from southwest part of Indian Ocean. Zool. Zhurnal 1992, 71, 140–145. [Google Scholar]
- Weltner, W. Verzeichnis der bisher beschriebenen recenten Cirripedienarten. Arch. Naturgeschichte. 1897, 63, 227–280. [Google Scholar]
- Weltner, W. Cirripedia der deutschen Tiefsee-expedition. In Wissenschaftliche Ergebnisse der deutschen Tiefsee-Expedition auf dem Dampfer Valdivia 1898–1899; Gustav Fischer: Jena, Germany, 1922; Volume 23, pp. 59–112. [Google Scholar]
- Nilsson-Cantell, C.A. Cirripeds from the Indian Ocean in the collection of the Indian Museum, Calcutta. Mem. Indian Mus. 1938, 13, 1–81. [Google Scholar]
- Hiro, F. Studies on the cirripedian fauna of Japan. III. Supplementary notes on the cirripeds found in the vicinity of Seto. Mem. Coll. Sci. Kyoto Imp. Univ. Ser. B 1939, 15, 237–244. [Google Scholar]
- Newman, W.A.; Ross, A. Revision of the balanomorph barnacles, including a catalog of the species. Mem. San Diego Soc. Nat. Hist. 1976, 9, 1–108. [Google Scholar]
- Ren, X. Studies on Chinese Cirripedia (Crustacea) V. Genus Acasta. Studia Mar. Sin. 1984, 23, 183–214. [Google Scholar]
- Rosell, N.C. Thoracic Cirripeds from the MUSORSTOM 2 expedition. Mémoires du Muséum Natl. d’Histoire naturelle. Ser. A Zool. 1989, 144, 9–35. [Google Scholar]
- Liu, R.; Ren, X. Crustacea Cirripedia Thoracica; Science Press: Beijing, China, 2007; Volume 42. [Google Scholar]
- Wibowo, R.A.; Prabowo, R.E.; Nuryanto, A. Biodiversitas teritip yang hidup pada spons di Perairan Pantai Kepulauan Karimunjawa. Semin. Nas. I Mataki 2011, 20, 219–235. [Google Scholar]
- Sulistiono, S.; Kawaroe, M.; Madduppa, H.; Prabowo, R. Karakteristik morfologi teritip spons Indonesia. DEPIK J. Ilmu-Ilmu Perair. Pesisir Dan Perikan. 2014, 3, 178–186. [Google Scholar]
- Jones, D.S.; Hosie, A.M. A checklist of the barnacles (Cirripedia: Thoracica) of Singapore and neighbouring waters. Raffles Bull. Zool. 2016, 34, 241–311. [Google Scholar]
- Utinomi, H. Studies on the Cirripedian fauna of Japan VII. Cirripeds from Sagami Bay. Publ. Seto Mar. Lab. 1958, 6, 281–311. [Google Scholar] [CrossRef] [Green Version]
- Rosell, N. Some barnacles (Cirripedia Thoracica) of Puerto Galera found in the vicinity of the UP Marine Biological Laboratory. Nat. Appl. Sci. Bull. Univ. Philipp. 1972, 24, 143–285. [Google Scholar]
- Foster, B.A.; Buckeridge, J.S. Barnacles (Cirripedia, Thoracica) of seas off Reunion Island and the east indies. Bull. Muséum Natl. D’histoire Nat. Sect. A Zool. Biol. Écologie Anim. 1994, 16, 345–382. [Google Scholar]
- Thiele, J. Studien über pazifische Spongien. II. Ueber einige Spongien von Celebes. Zoologica. Orig. Abh. Aus Dem Gesamtgeb. Zoologie. Stuttg. 1899, 24, 1–33. [Google Scholar]
- Döderlein, L. Studien an japanischen Lithistiden. Z. Fürwissenschaftliche Zool. 1884, 40, 62–104. [Google Scholar]
- Ridley, S.O.; Dendy, A. Preliminary report on the Monaxonida collected by H.M.S. Challenger. Part I. Ann. Mag. Nat. Hist. 1886, 18, 470–493. [Google Scholar] [CrossRef]
- Lamarck, J.-B.P.A. Histoire Naturelle des Animaux sans Vertèbres; Deterville: Paris, France, 1818; Volume 5, p. 612. [Google Scholar]
- Lamarck, J.-B.P.A. Histoire Naturelle des Animaux sans Vertèbres Précédée d’une Introduction Offrant la Détermination des Caractères Essentiels de L’animal, sa Distinction du Végétal et des Autres Corps Naturels, enfin, L’exposition des Principes Fondamentaux de la Zoologie; Verdière: Paris, France, 1815; Volume 7. [Google Scholar]
- Bergquist, P.R.; Tizard, C.A. Australian intertidal sponges from the Darwin area. Micronesica 1967, 3, 175–202. [Google Scholar]
- Hooper, J.N.A.; Bergquist, P.R. Cymbastela, a new genus of lamellate coral reef sponges. Mem. Qld. Mus. 1992, 32, 99–137. [Google Scholar]
- Kolbasov, G.A. New species of the genus Acasta (Cirripedia, Thoracica) from western Pacific. Zool. Žurnal 1991, 70, 32–38. [Google Scholar]
- Nilsson-Cantell, C.A. Cirripeden-studien zur kenhtnis der biologie, anatomie und systematik dieser gruppe. Zool. Bidr. Från Upps. 1921, 7. [Google Scholar]
- Krüger, P. Beitrage zur Cirripedienfauna Ostasiens. Konglige Bayer. Akad. Der Wiss. Munich Math. Phys. Klasse. Abh. Suppl. 1911, 2, 1–72. [Google Scholar]
- Rosell, N.C. A gorgonacean inhabiting barnacle Genus Acasta from Cebu. Nat. Appl. Sci. Bull. 1970, 22, 103–111. [Google Scholar]
- Kolbasov, G.A. Acasta tabachniki sp. nov. (Cirripedia, Thoracica) from South China Sea. Zool. Zhurnal 1990, 69, 135–137. [Google Scholar]
- Hiro, F. Report on the Cirripedia collected in the Malayan waters by the ship “Zuihomaru”. Jpn. J. Zool. 1936, 6, 621–636. [Google Scholar]
- Broch, H. Cirripedien. Results of Dr. E. Mjoberg’s Swedish scientific expedition to Australia, 1910–1913, No. 8. K. Sven. Vetensk. Handl. 1916, 52, 1–16. [Google Scholar]
- Yu, M.-C.; Chan, B.K.K.; Kolbasov, G.A.; Ganmanee, M. Biodiversity and host specificity of sponge-associated barnacles (Cirripedia: Thoracica) in Thailand. J. Crustacean Biol. 2020, 40, 839–865. [Google Scholar] [CrossRef]
- Hiro, F. Notes on some new Cirripedia from Japan. Mem. Coll. Sci. Kyoto Imp. Univ. 1931, 7, 143–158. [Google Scholar]
- Carter, H.J. Contributions to our knowledge of the Spongida. Ann. Mag. Nat. Hist. 1883, 12, 308–329. [Google Scholar] [CrossRef]
- Pilsbry, H.A. Diagnoses of new barnacles from the Philippine Archipelago and China Sea. Proc. United States Natl. Mus. 1912, 42, 291–294. [Google Scholar] [CrossRef] [Green Version]
- Broch, H. Papers from Dr. Th. Mortensen’s Pacific expedition 1914-1916. X. Studies on Pacific cirripedes. Vidensk. Meddr. Dan. Naturh. Foren. Kbh. 1922, 73, 215–358. [Google Scholar]
- Barnard, K.H. Contributions to the Crustacean fauna of South Africa. N7. Cirripedia. Ann. S. Afr. Mus. 1924, 20, 1–103. [Google Scholar]
- Utinomi, H. Studies on the cirripedian fauna of Japan. VIII. Thoracic cirripeds from western Kyusyu. Publ. Seto Mar. Lab. 1962, 10, 211–239. [Google Scholar] [CrossRef] [Green Version]
- Zullo, V.A. Catalog of the Cirripedia named by Henry A. Pilsbry. Proc. Acad. Nat. Sci. Phila. 1968, 120, 209–235. [Google Scholar]
- Jones, D. A guide to the shallow-water barnacles (Cirripedia: Lepadomorpha, Balanomorpha) of the Shark Bay area, Western Australia. In Research in Shark Bay: Report of the France-Australe Bicentenary Expedition Committee; IUCN: Perth, Australia, 1990; pp. 209–229. [Google Scholar]
- Jones, D.S.; Anderson, J.T.; Anderson, D.T. Checklist of the Australian Cirripedia. Tech. Rep. Aust. Mus. 1990, 3, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.S.; Hewitt, M.A.; Sampey, A. A checklist of the Cirripedia of the South China Sea. Raffles Bull. Zool. 2000, 48, 233–308. [Google Scholar]
- Jones, D. The littoral and shallow-water barnacles (Crustacea: Cirripedia) of south-eastern Queensland. In Proceedings of the Thirteenth International Marine Biological Workshop, The Marine Fauna and Flora of Moreton Bay; Davie, P.J.F., Phillips, J.A., Eds.; Queensland Museum: Brisbane, Australia, 2010; pp. 199–233. [Google Scholar]
- Kadota, J. Observations of two new species of the genus Reniera of monaxonid sponges. Zool. Mag. 1922, 34, 700–711. [Google Scholar]
- Foster, B. Shallow water barnacles from Hong Kong. Proc. First Int. Mar. Biol. Work. Mar. Flora Fauna Hong Kong S. China 1980, 1, 207–232. [Google Scholar]
- Rosell, N.C. Crustacea Cirripedia Thoracica: MUSORSTOM 3 Philippine collection. Mémoires Muséum Natl. D’histoire Nat. Ser. A Zool. 1991, 152, 9–61. [Google Scholar]
- Hoshino, T. Description of two new species in the genus Agelas (Demospongia) from Zamami Island, the Ryukyus, Japan. Proc. Jpn. Soc. Syst. Zool. 1985, 30, 1–10. [Google Scholar]
- Hooper, J.N.A. Revision of the family Raspailiidae (Porifera: Demospongiae), with description of Australian species. Invertebr. Taxon. 1991, 5, 1179–1418. [Google Scholar] [CrossRef]
- Watanabe, S. A widely applicable Bayesian information criterion. J. Mach. Learn. Res. 2013, 14, 867–897. [Google Scholar]
- Jones, D.S. The shallow-water barnacles (Cirripedia: Lepadomorpha, Balanomorpha) of southern Western Australia. In Proceedings of the Third International Marine Biological Workshop: The Marine Flora and Fauna of Albany, Western Australia; Western Australian Museum: Perth, WA, Australia, 1990; pp. 332–437. [Google Scholar]
- Krüger, P. Cirripedia. In Die Fauna Südwest-Australiens. Ergebnisse der Hamburger Südwest-Australischen Forschungsreise 1905 Herausgegeben von Prof. Dr. W. Michaelsen und Dr. R. Hartmeyer.; Michaelsen, W., Ed.; Gustav Fischer: Jena, Germany, 1914; Volume 4, pp. 427–441. [Google Scholar]

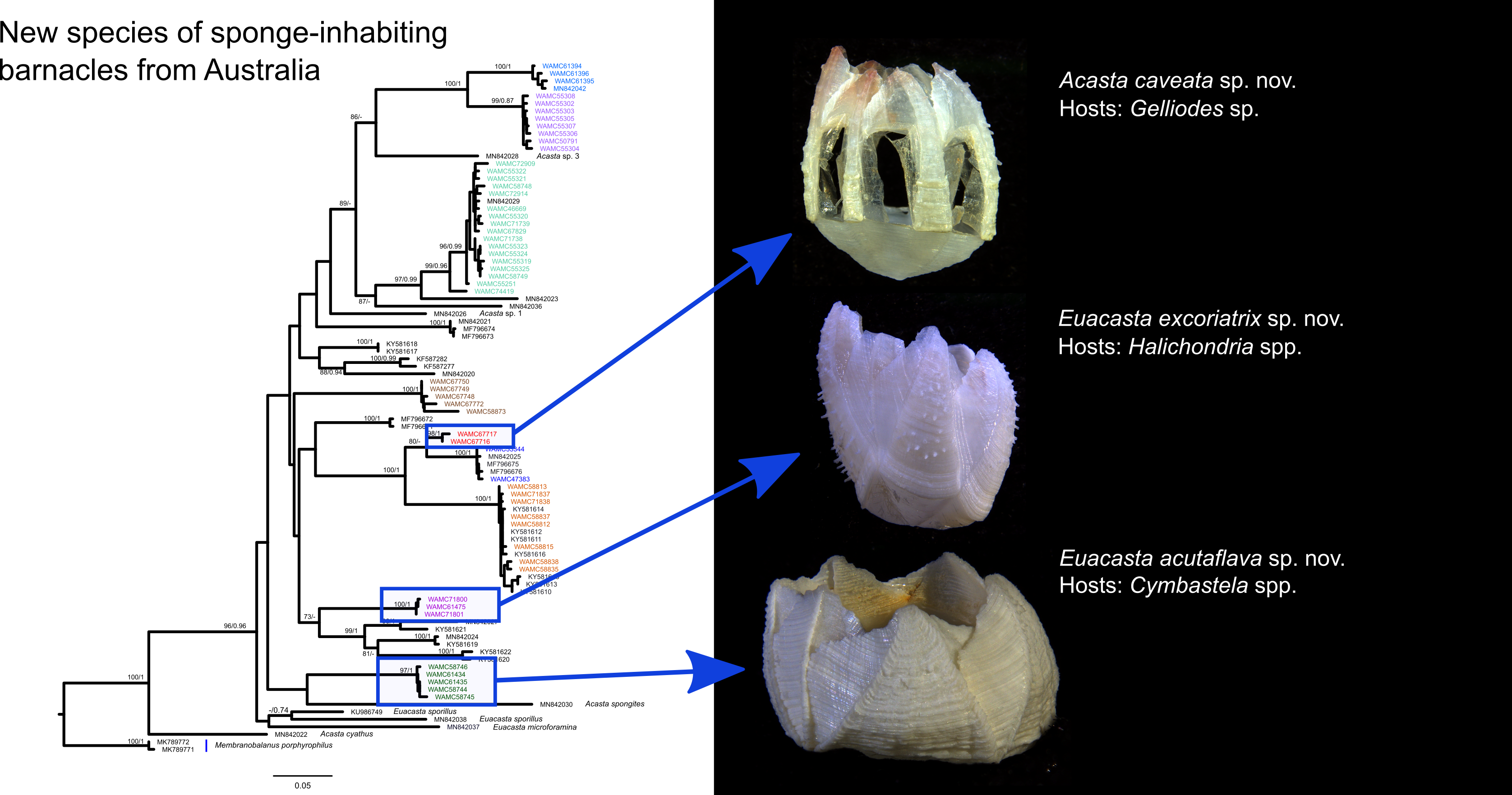{kind=link}
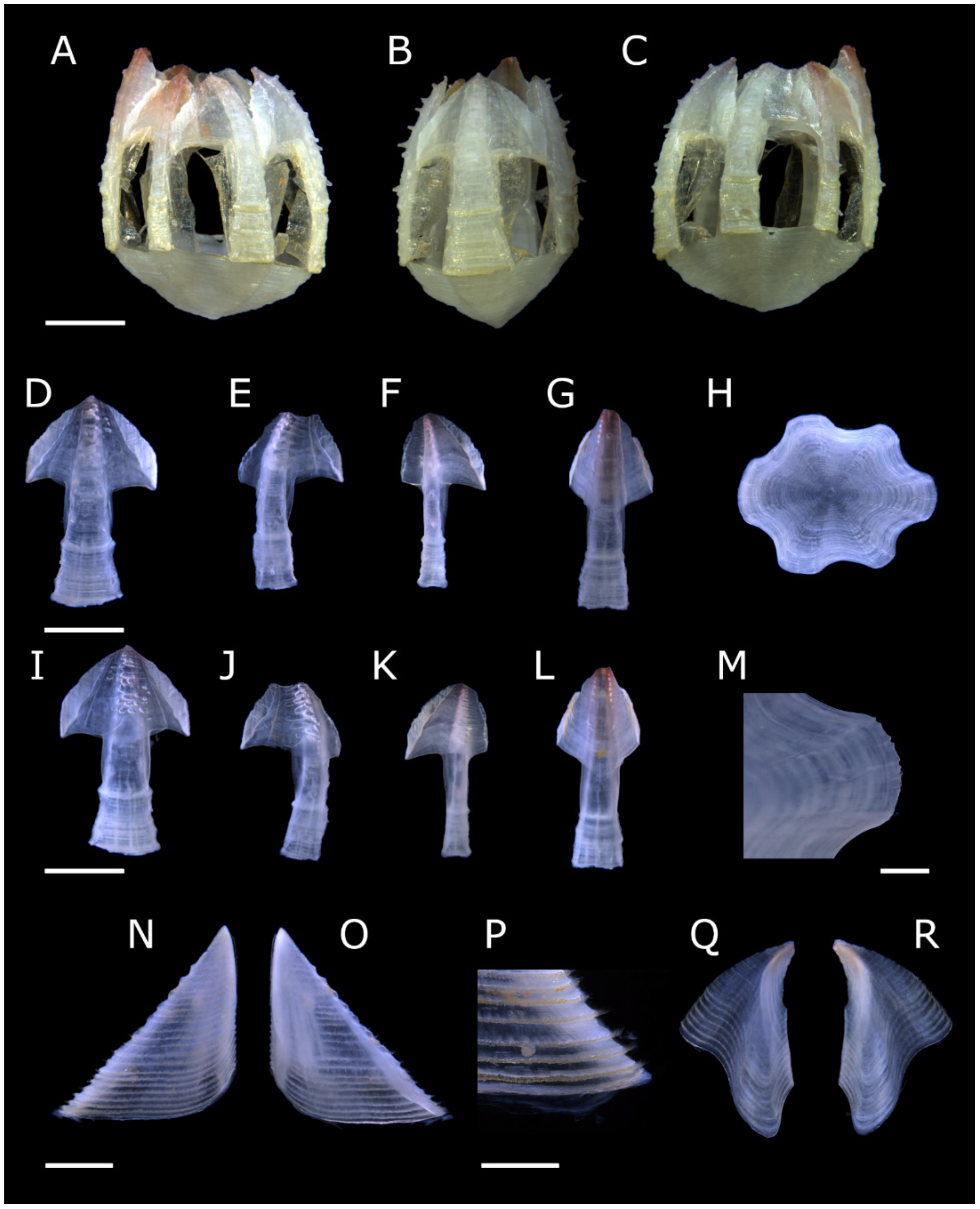{kind=link}
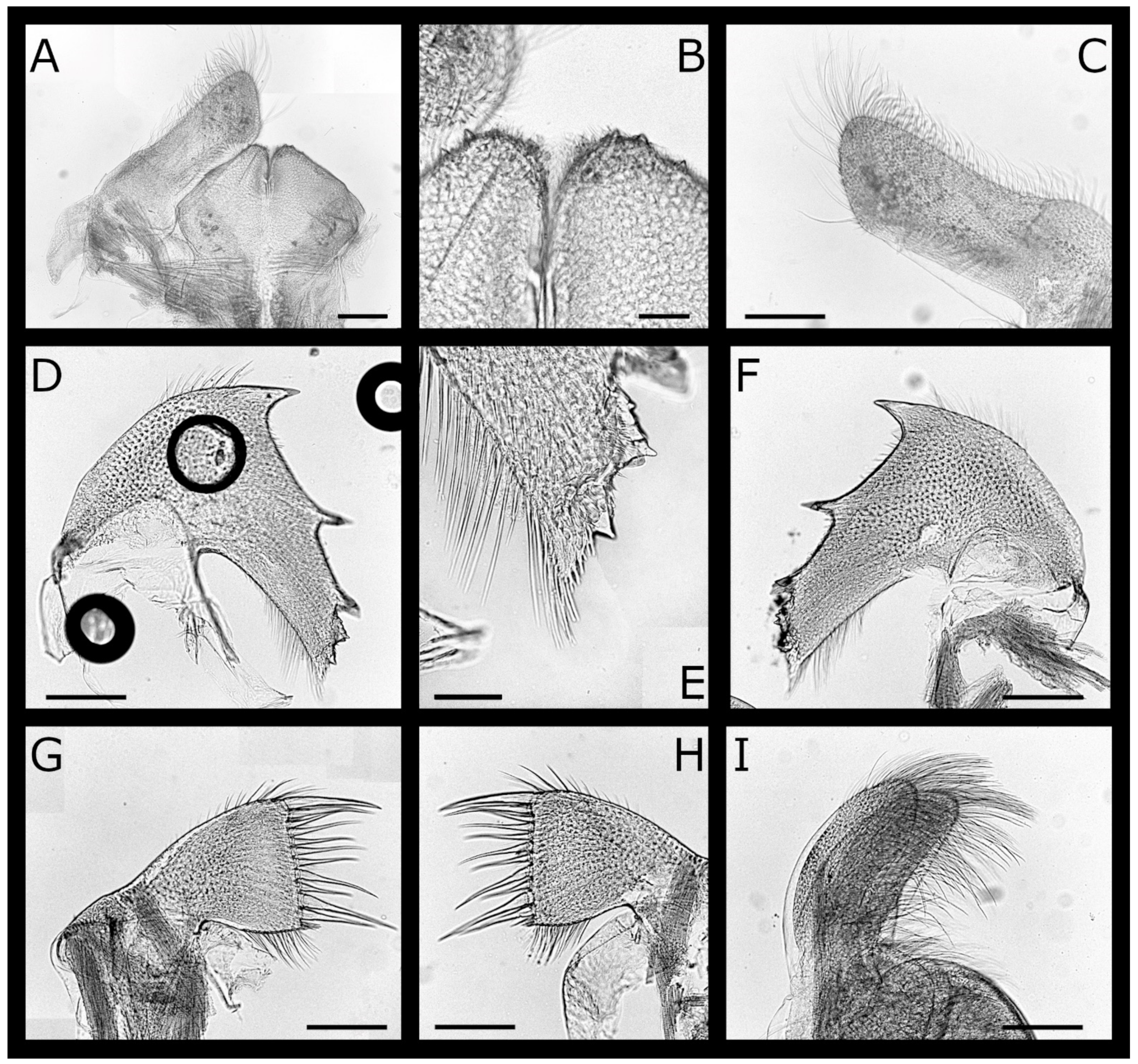{kind=link}
{kind=link}
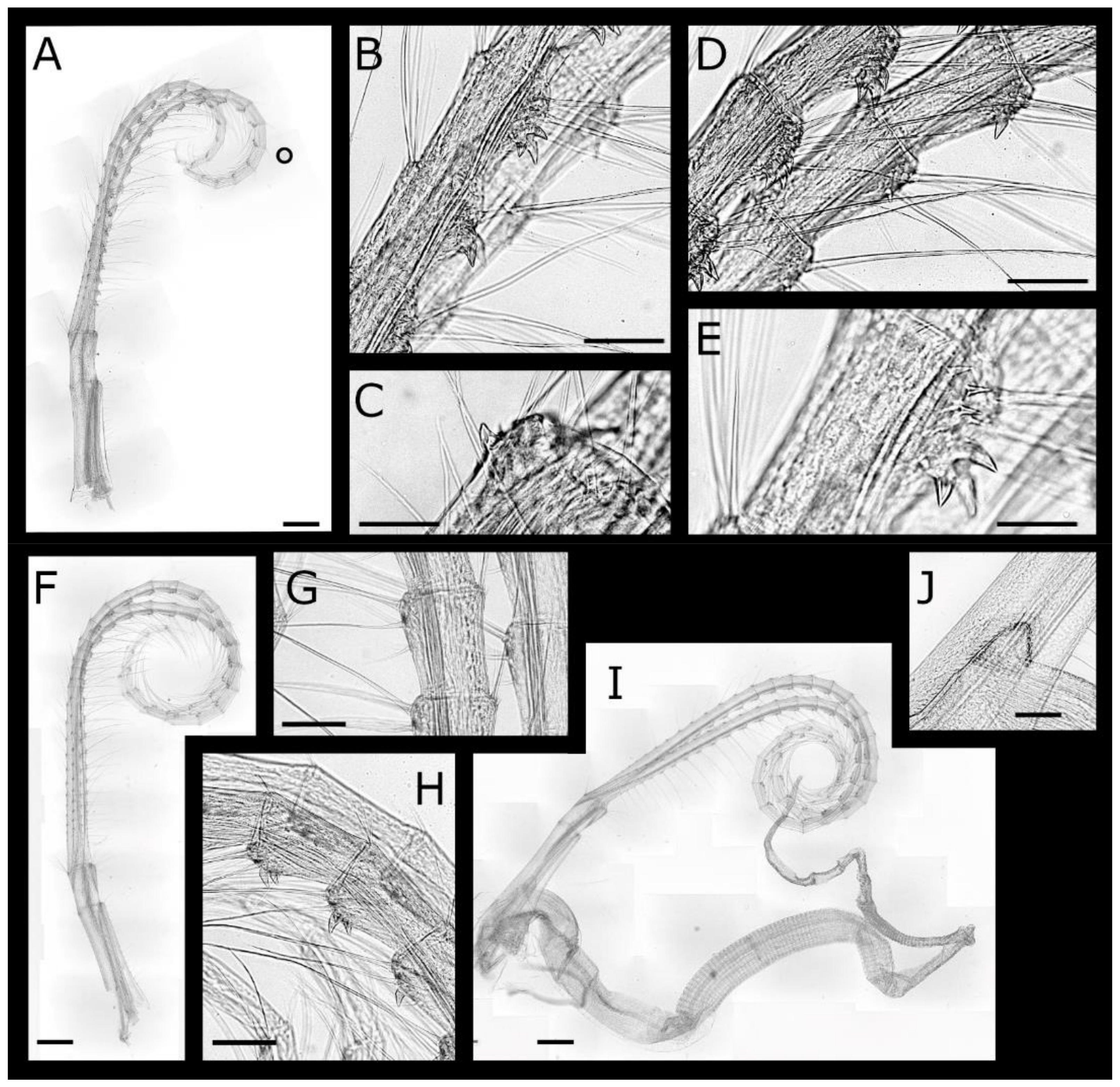{kind=link}
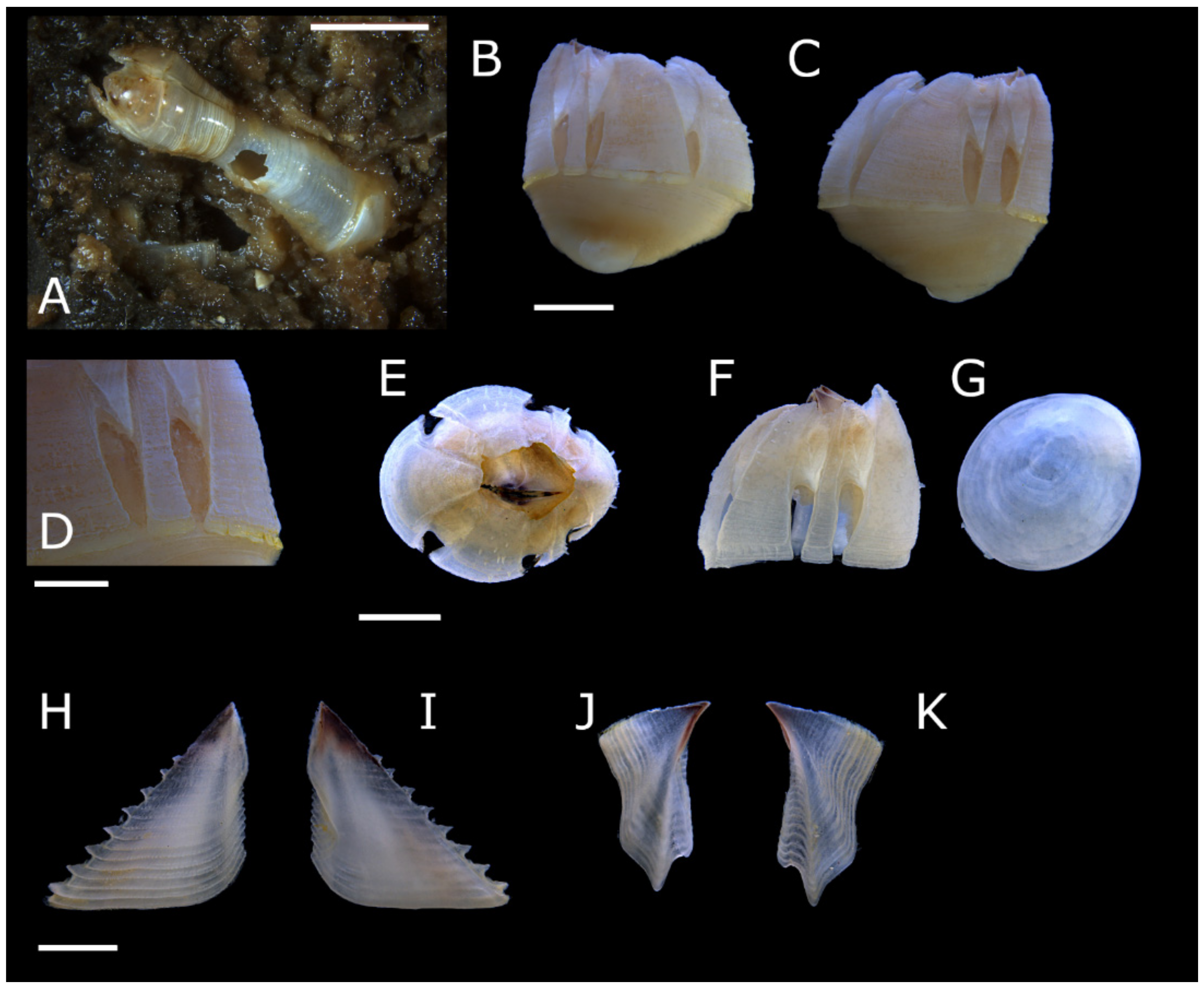{kind=link}
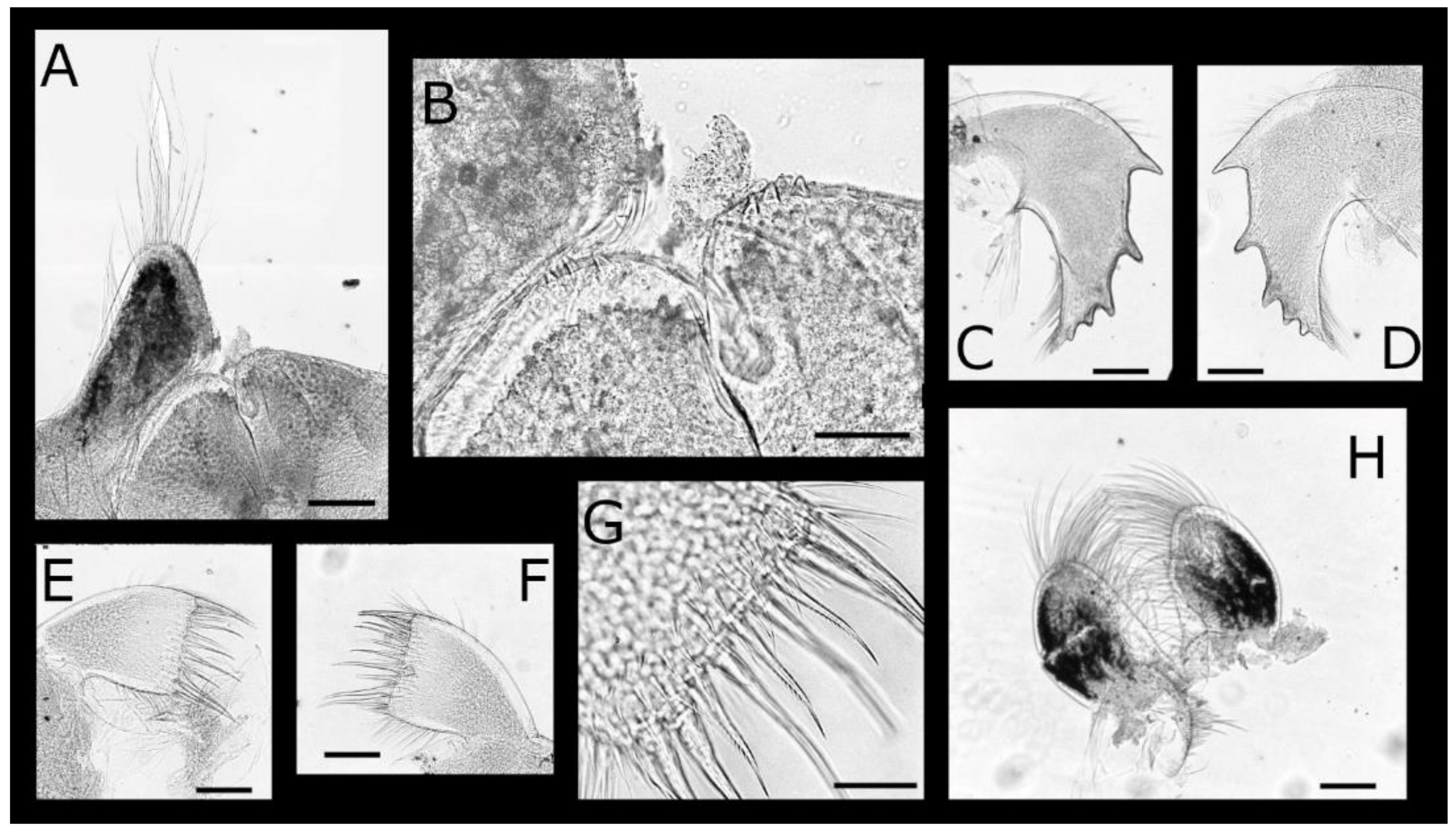{kind=link}
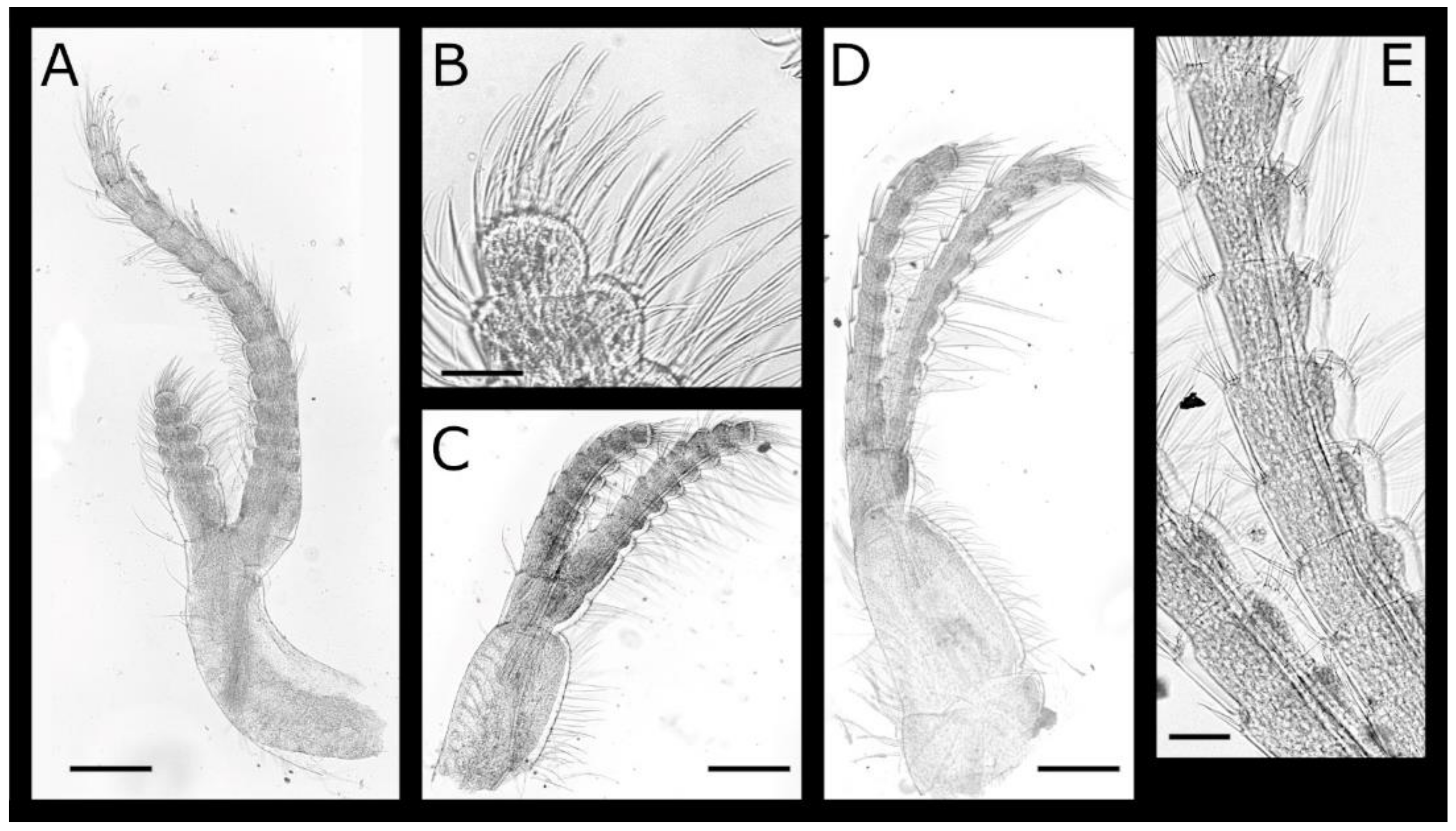{kind=link}
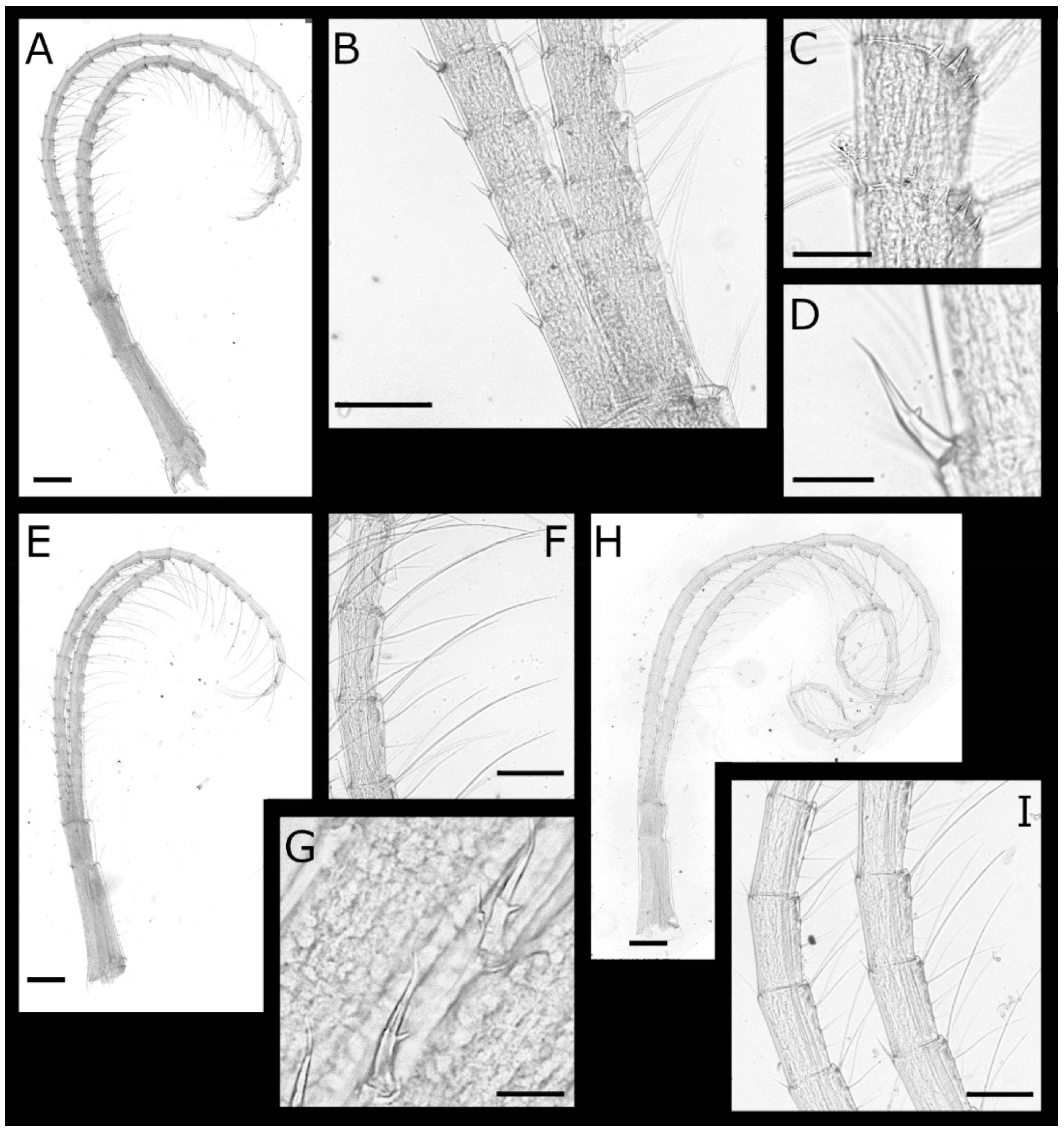{kind=link}
{kind=link}
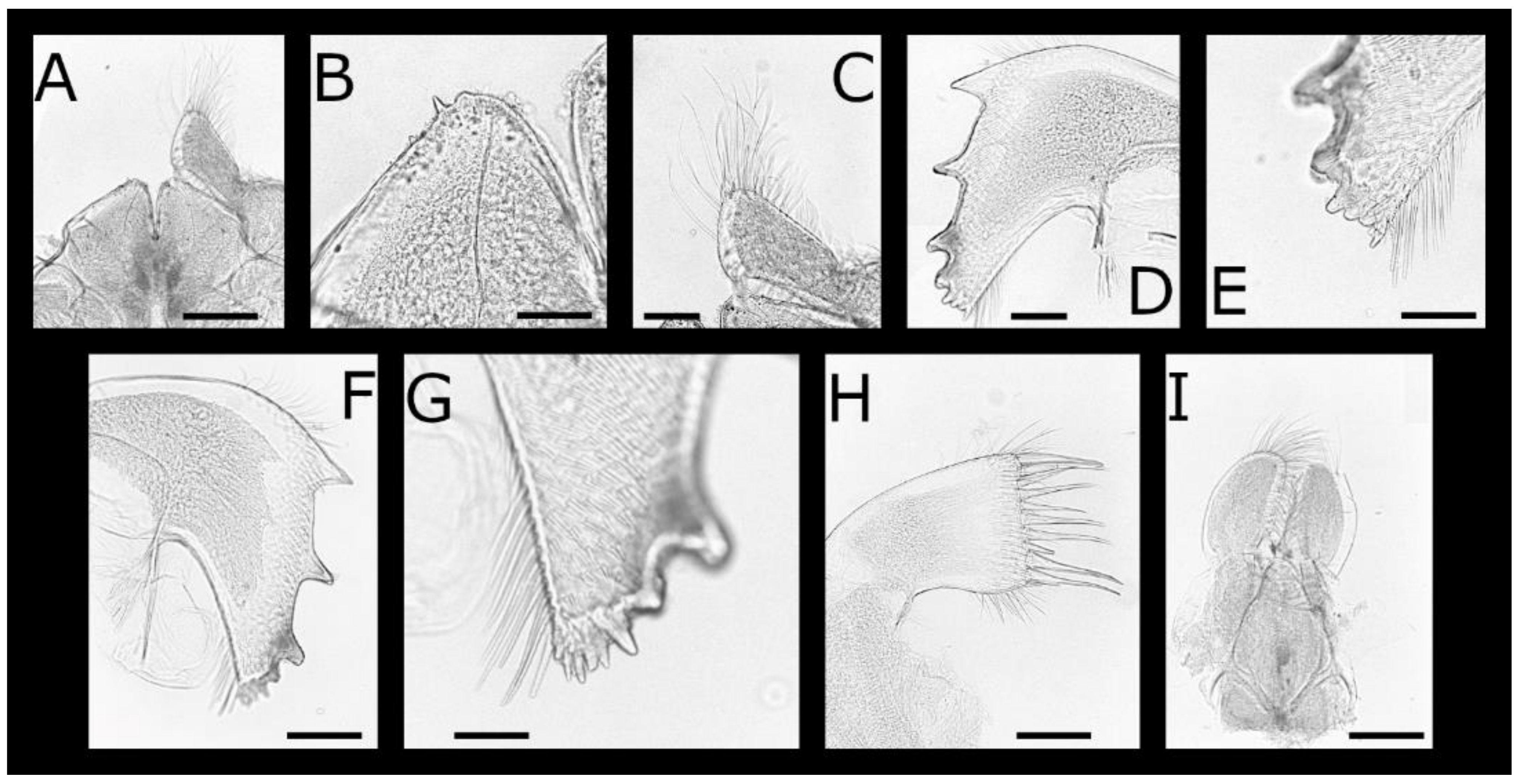{kind=link}
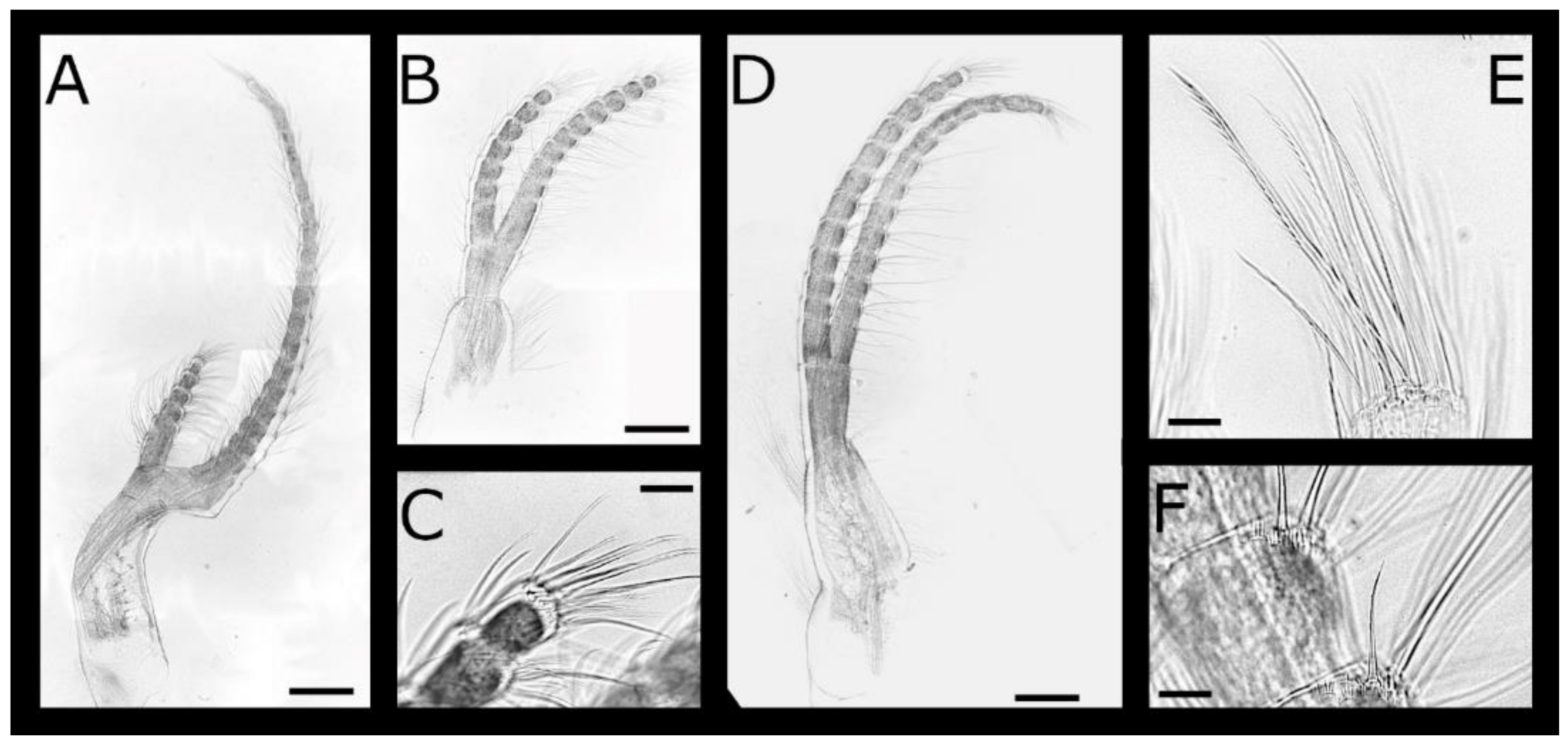{kind=link}
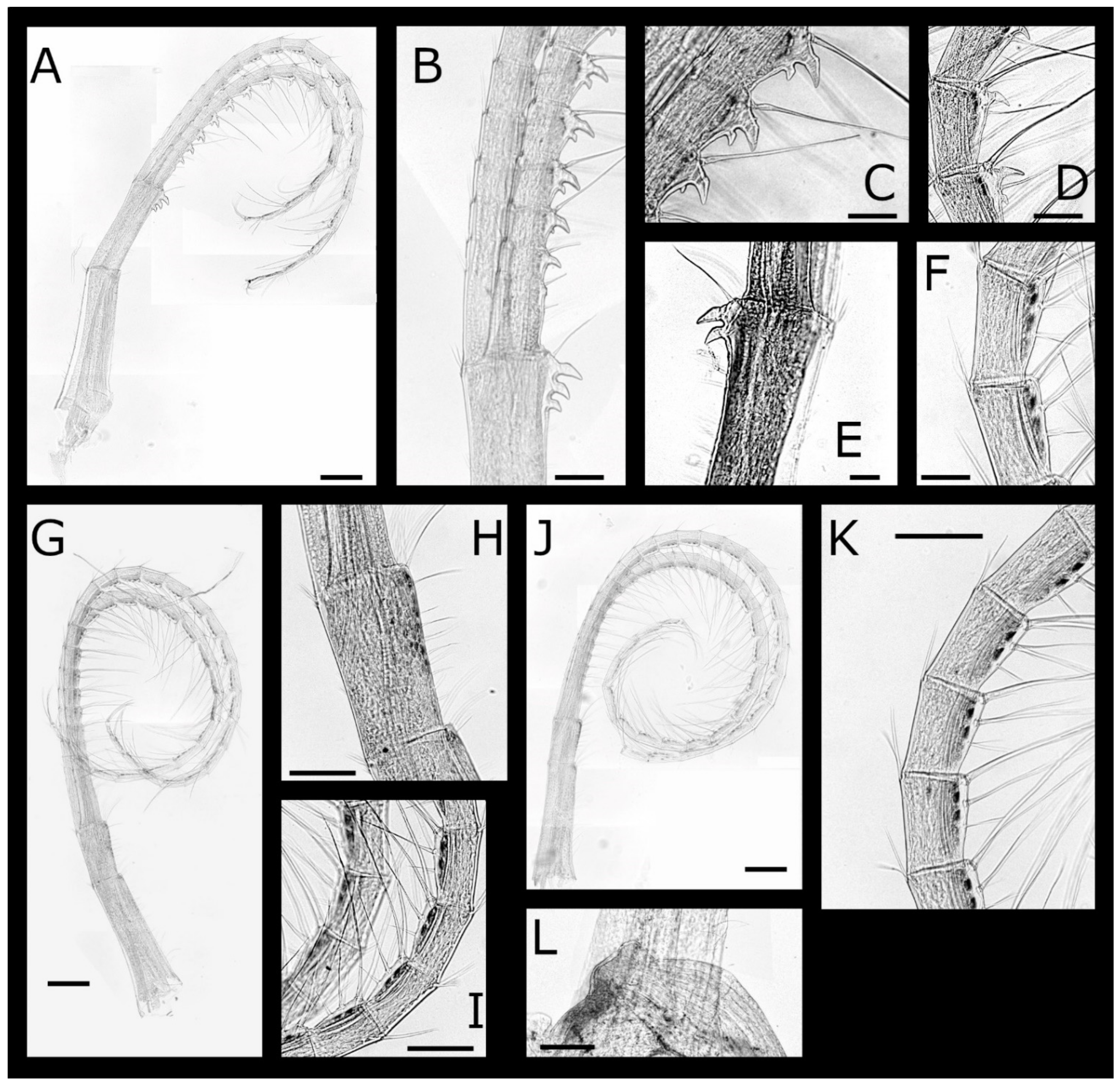{kind=link}
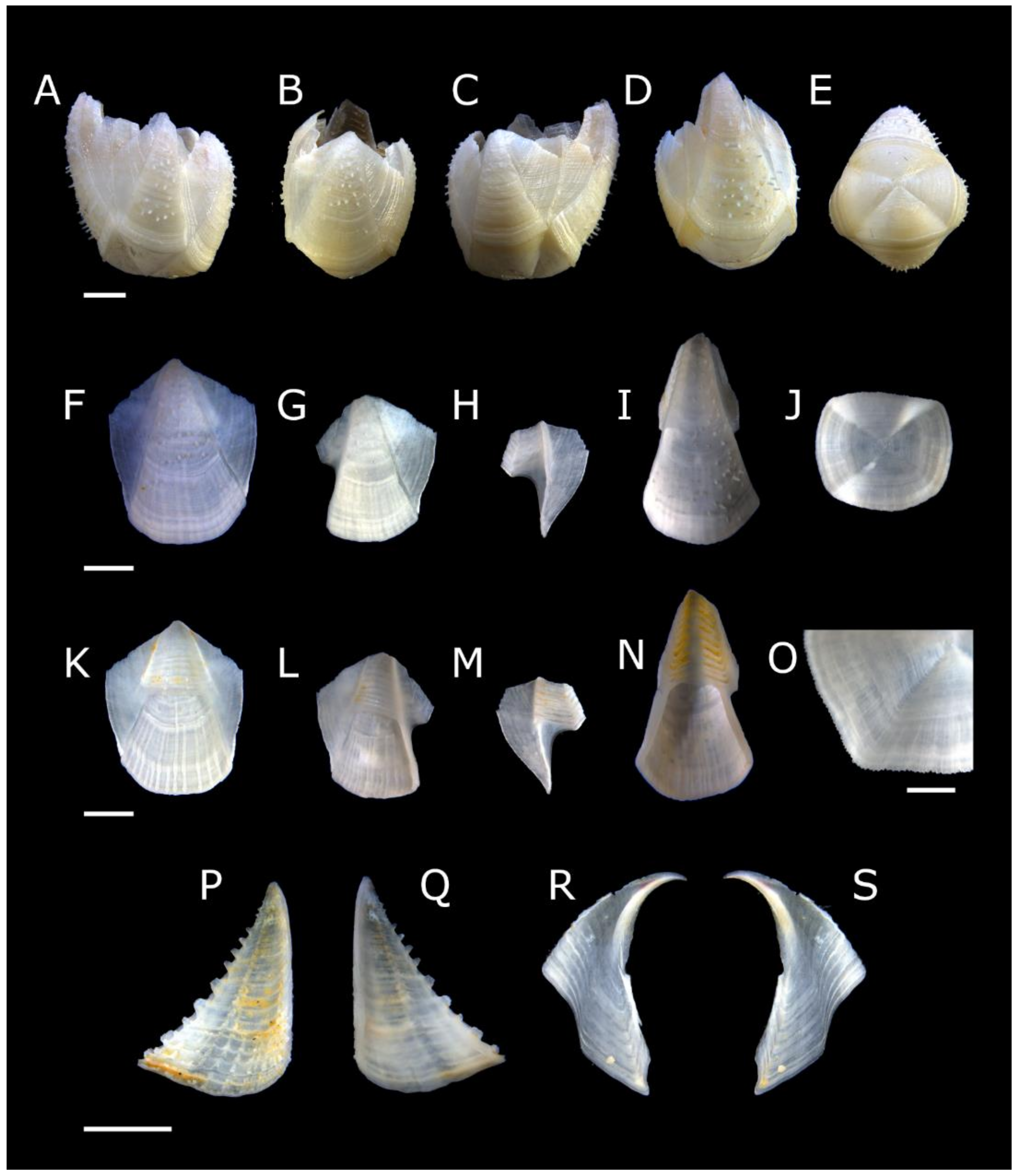{kind=link}
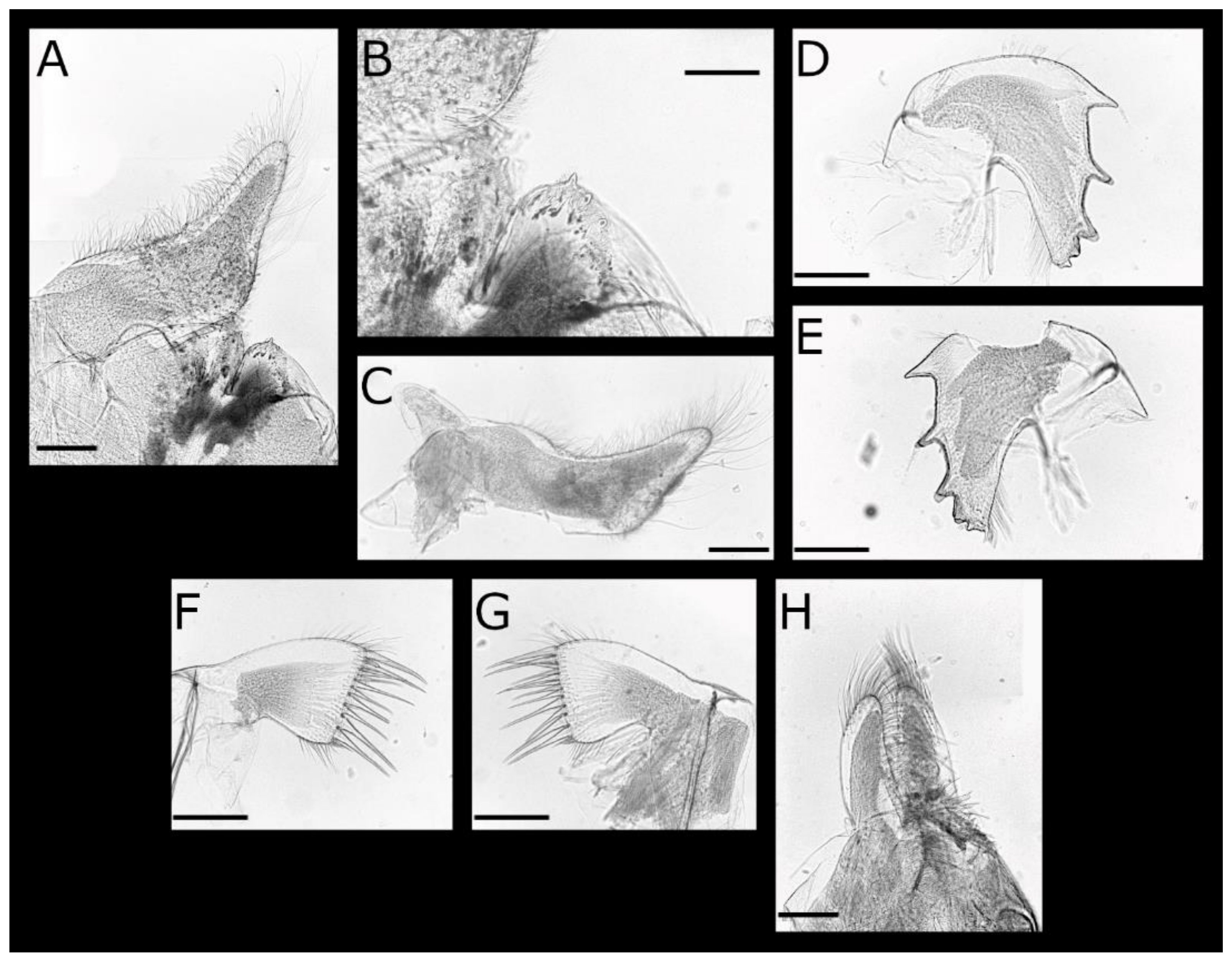{kind=link}
{kind=link}
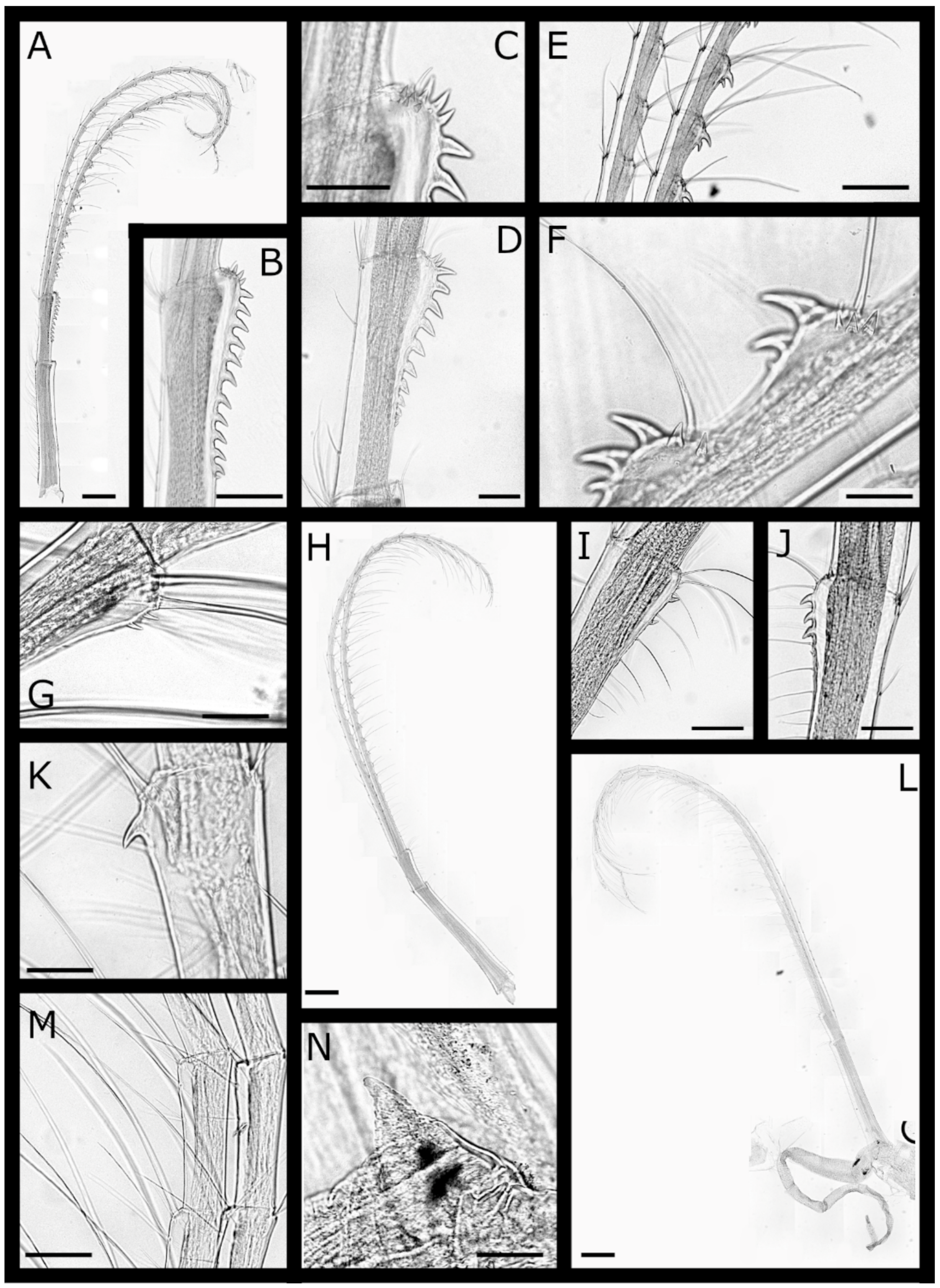{kind=link}
{kind=link}
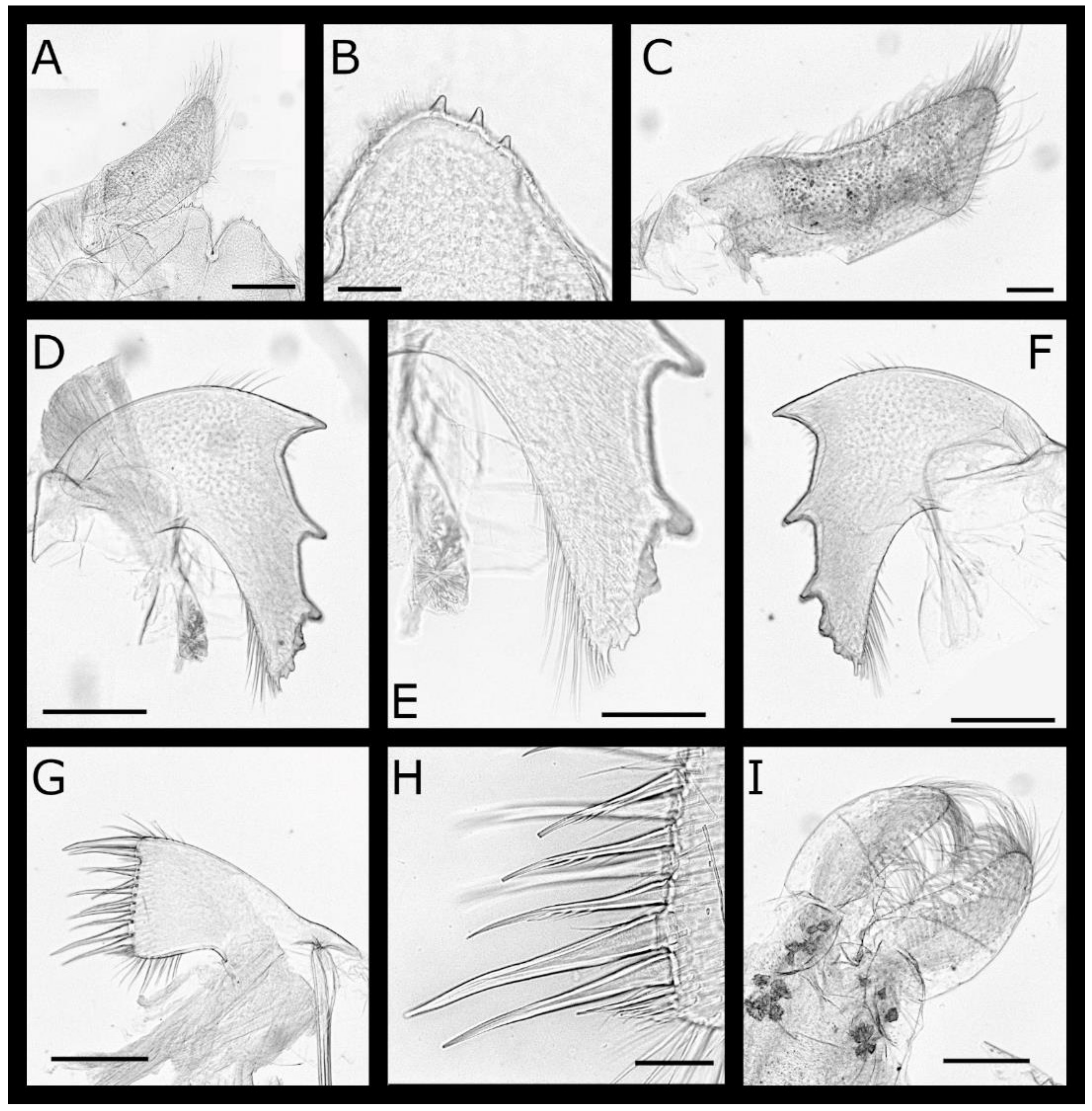{kind=link}
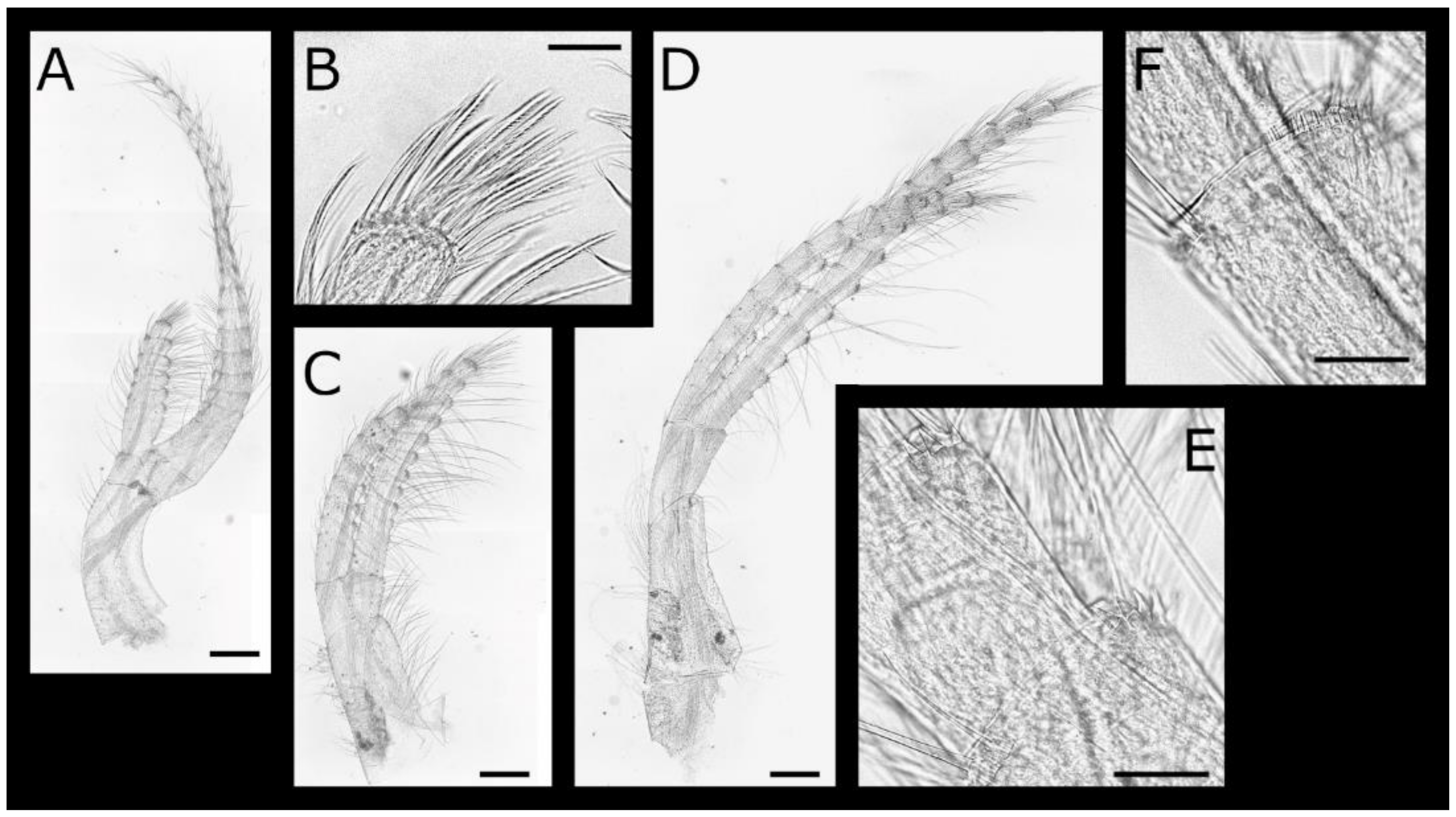{kind=link}
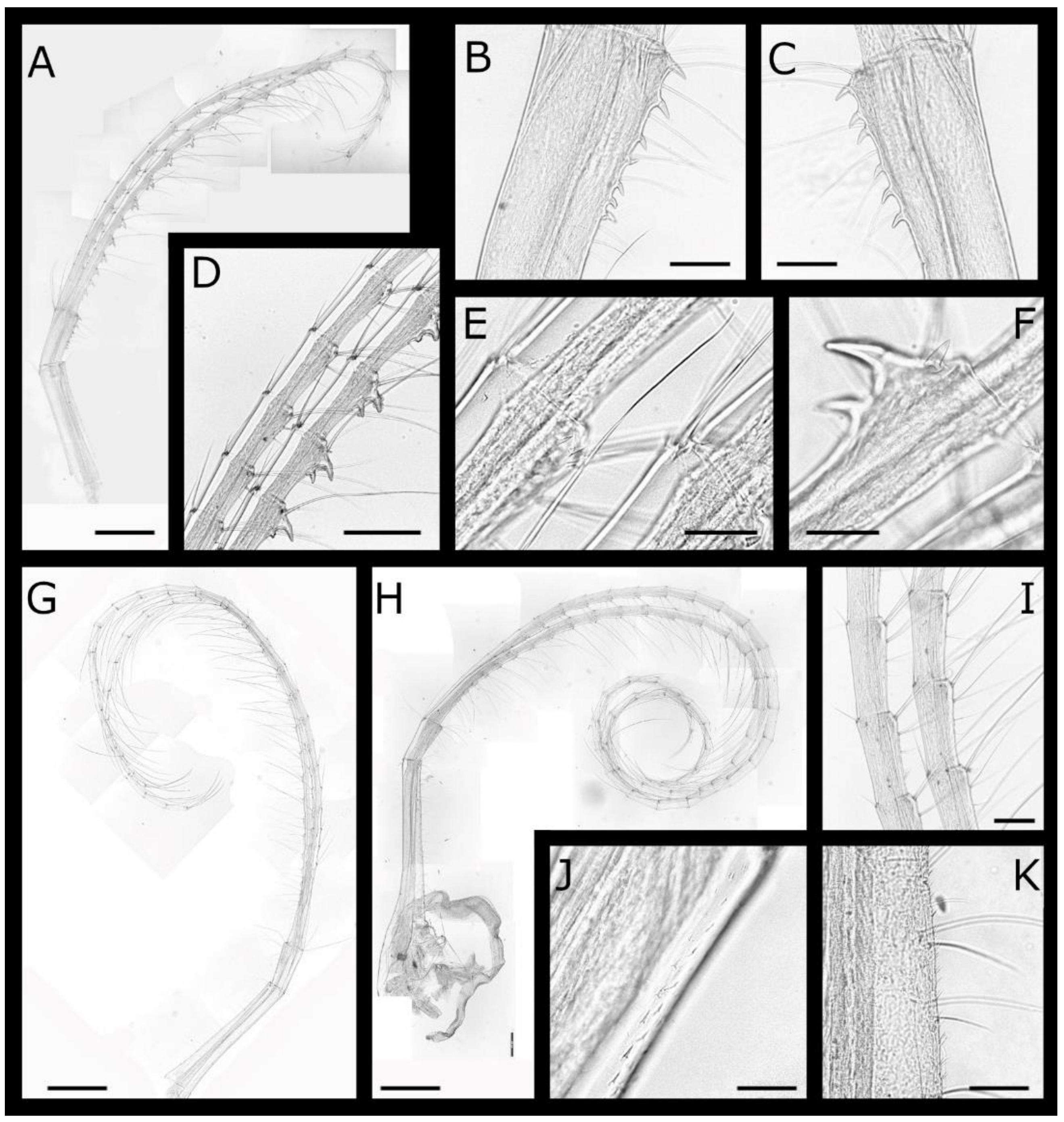{kind=link}
{kind=link}
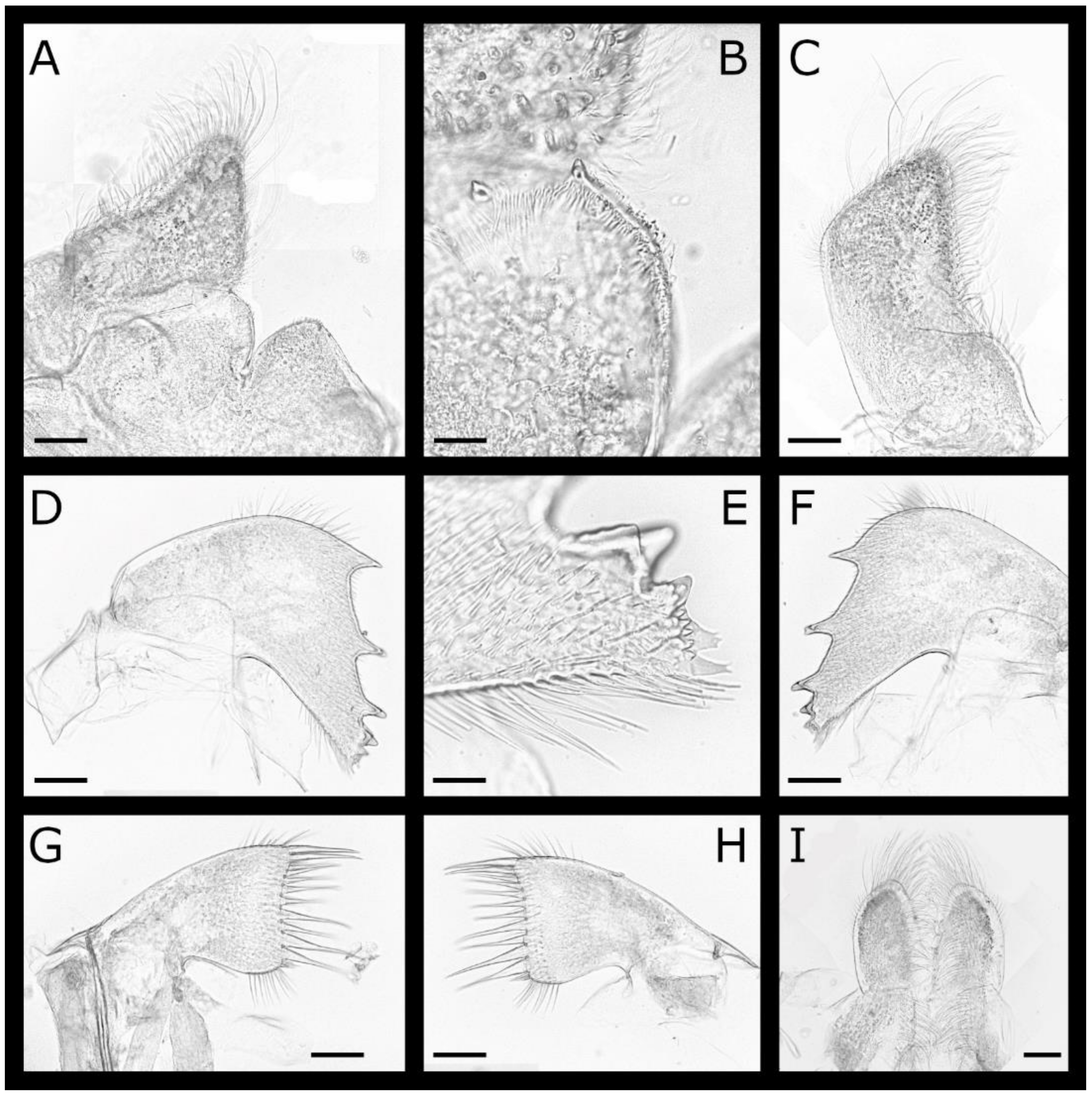{kind=link}
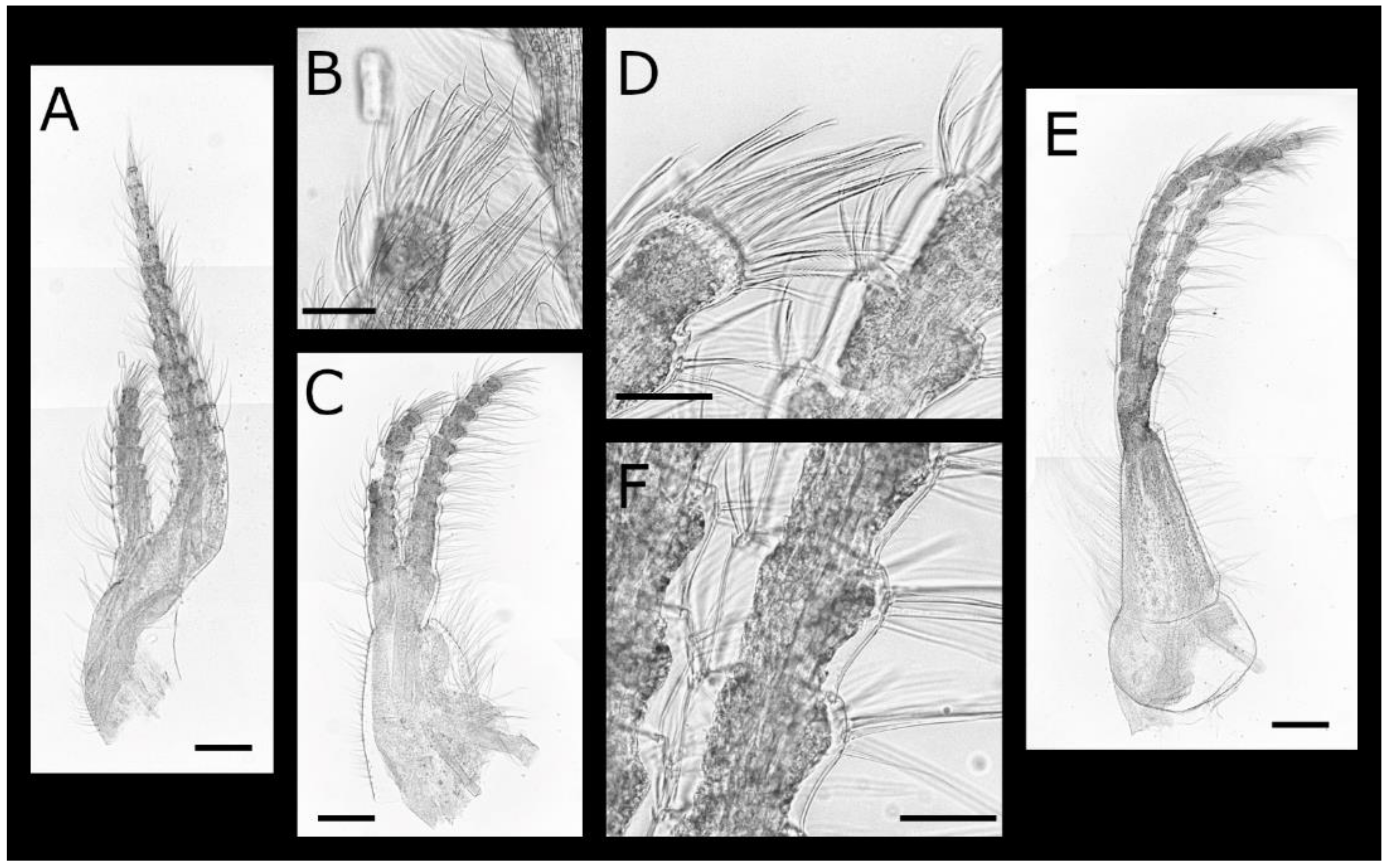{kind=link}
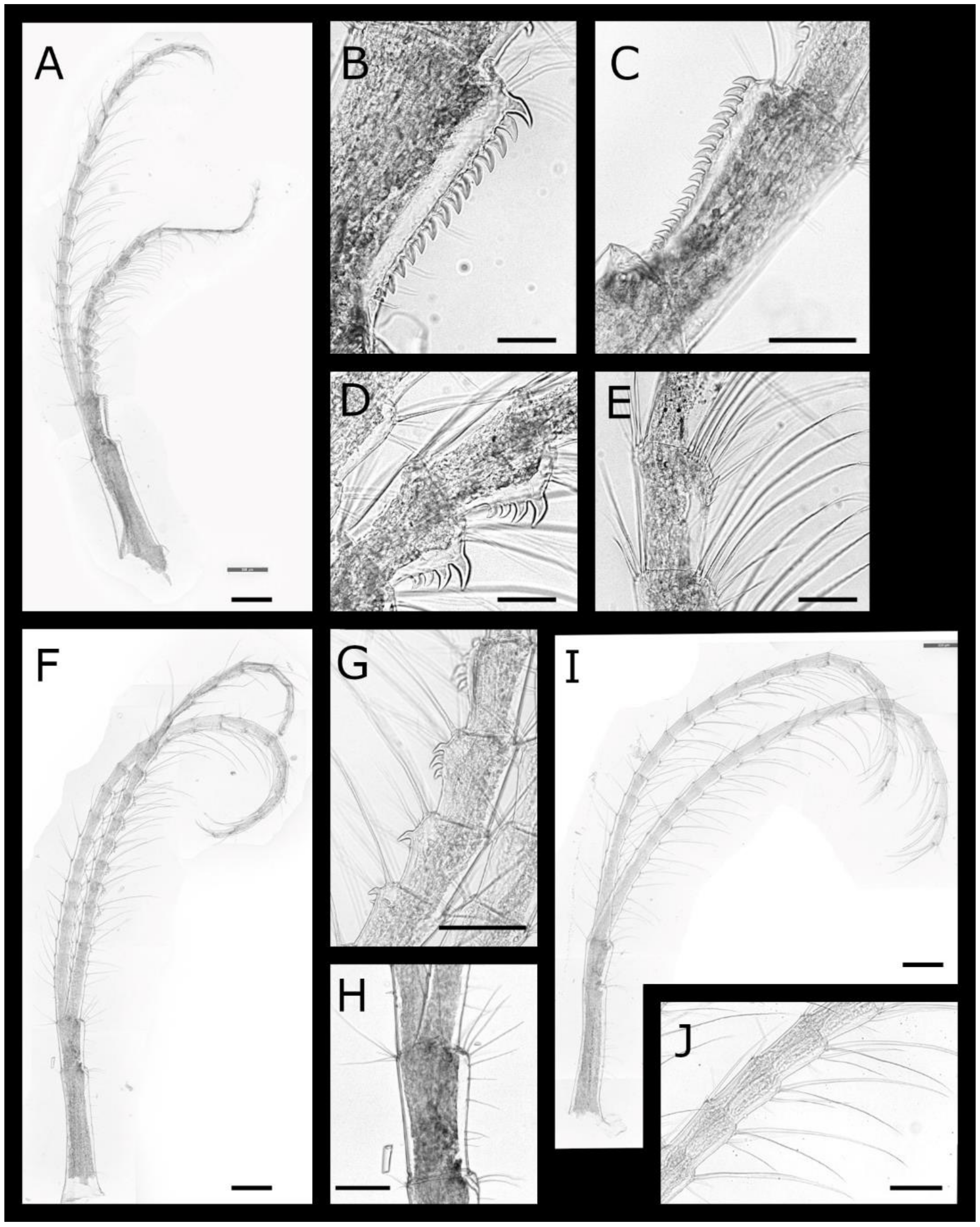{kind=link}
{kind=link}
{kind=link}
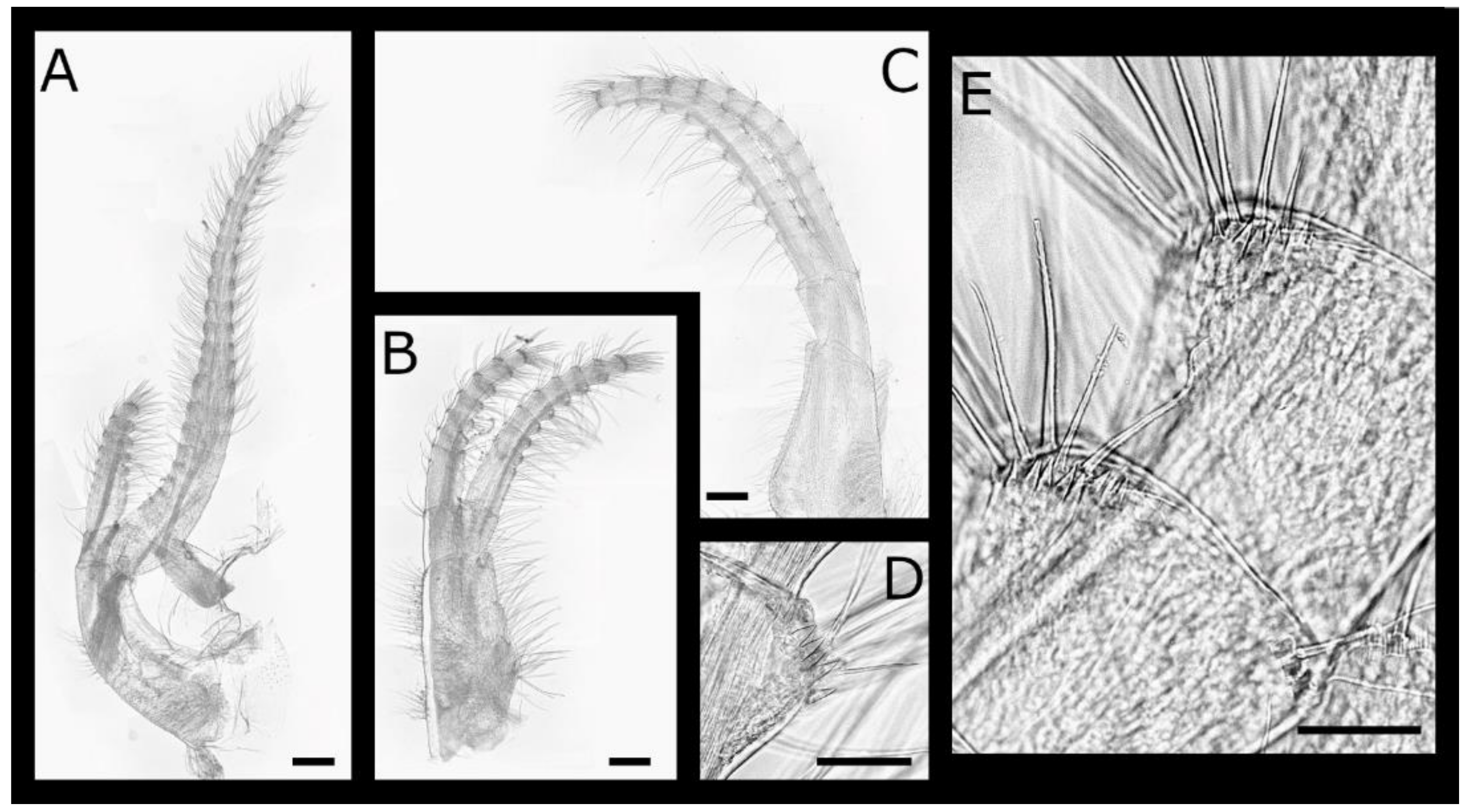{kind=link}
{kind=link}
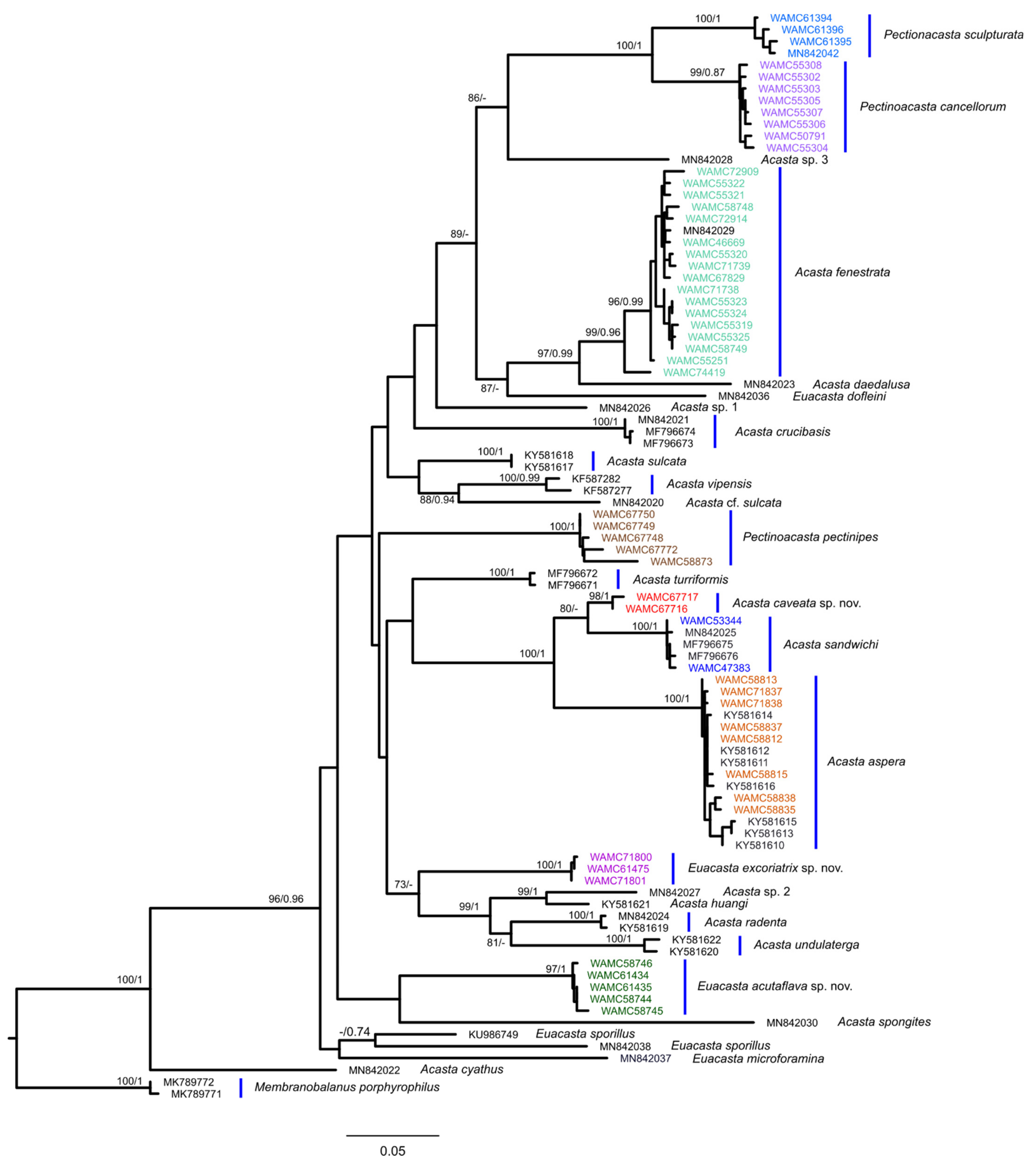{kind=link}
Table 1.
Specimen details of sequences used in the molecular analysis. Records in bold indicate specimens newly sequenced for this study.
Table 1.
Specimen details of sequences used in the molecular analysis. Records in bold indicate specimens newly sequenced for this study.
| Species | Accession | Host Species | GenBank |
|---|---|---|---|
| Acasta aspera | SGI20-1 | Iotroata sp. | KY581615 [3] |
| Acasta aspera | SGI20-2 | Iotroata sp. | KY581616 [3] |
| Acasta aspera | SGI2-1 | Iotroata sp. | KY581614 [3] |
| Acasta aspera | WAM C55057 | Crella (Yvesia) spinulata | KY581613 [3] |
| Acasta aspera | WAM C58741 | Crella (Yvesia) spinulata | KY581610 [3] |
| Acasta aspera | WAM C58742 | Crella (Yvesia) spinulata | KY581611 [3] |
| Acasta aspera | WAM C58743 | Crella (Yvesia) spinulata | KY581612 [3] |
| Acasta aspera | WAM C58812 | Lissodendoryx (Acanthodoryx) KMB1 | MZ086820 |
| Acasta aspera | WAM C58813 | Lissodendoryx (Acanthodoryx) KMB1 | MZ086821 |
| Acasta aspera | WAM C58815 | Lissodendoryx (Acanthodoryx) KMB1 | MZ086822 |
| Acasta aspera | WAM C58835 | Hamigera PB1 | MZ086823 |
| Acasta aspera | WAM C58837 | Hamigera | MZ086824 |
| Acasta aspera | WAM C58838 | Hamigera | MZ086825 |
| Acasta aspera | WAM C71837 | Lissodendoryx (Acanthodoryx) KMB1 | MZ086844 |
| Acasta aspera | WAM C71838 | Lissodendoryx (Acanthodoryx) KMB1 | MZ086845 |
| Acasta caveata sp. nov. | WAM C67716 | Gelliodes KMB1 | MZ086833 |
| Acasta caveata sp. nov. | WAM C67717 | Gelliodes KMB1 | MZ086834 |
| Acasta crucibasis | MF796674 | Unknown | MF796674 |
| Acasta crucibasis | S32-1 | Xestospongia vansoesti | MN842021 [17] |
| Acasta cyathus | SGI265-1 | Xestospongia testudinaria | MN842022 [17] |
| Acasta daedalusa | SGI28-2 | Petrosia (Petrosia) sp. | MN842023 [17] |
| Acasta fenestrata | WAM C46669 | Neopetrosia chaliniformis | MZ086796 |
| Acasta fenestrata | WAM C55251 | Neopetrosia chaliniformis | MZ086800 |
| Acasta fenestrata | WAM C55319 | Neopetrosia chaliniformis | MZ086808 |
| Acasta fenestrata | WAM C55320 | Neopetrosia chaliniformis | MZ086809 |
| Acasta fenestrata | WAM C55321 | Neopetrosia chaliniformis | MZ086810 |
| Acasta fenestrata | WAM C55322 | Neopetrosia chaliniformis | MZ086811 |
| Acasta fenestrata | WAM C55323 | Neopetrosia chaliniformis | MZ086812 |
| Acasta fenestrata | WAM C55324 | Neopetrosia chaliniformis | MZ086813 |
| Acasta fenestrata | WAM C55325 | Neopetrosia chaliniformis | MZ086814 |
| Acasta fenestrata | WAM C58748 | Neopetrosia chaliniformis | MZ086818 |
| Acasta fenestrata | WAM C58749 | Neopetrosia chaliniformis | MZ086819 |
| Acasta fenestrata | WAM C67829 | Neopetrosia chaliniformis | MZ086839 |
| Acasta fenestrata | WAM C71738 | Neopetrosia chaliniformis | MZ086840 |
| Acasta fenestrata | WAM C71739 | Neopetrosia chaliniformis | MZ086841 |
| Acasta fenestrata | WAM C72909 | Neopetrosia chaliniformis | MZ086846 |
| Acasta fenestrata | WAM C72914 | Neopetrosia chaliniformis | MZ086847 |
| Acasta fenestrata | WAM C74419 | Neopetrosia chaliniformis | MZ086848 |
| Acasta huangi | SNE47-4 | Jaspis splendens | KY581621 [3] |
| Acasta radenta | SLQ32-1 | Jaspis splendens | MN842024 [17] |
| Acasta radenta | SNE47-1 | Jaspis splendens | KY581619 [3] |
| Acasta sandwichi | SGI208-1 | Xestospongia testudinaria | MN842025 [17] |
| Acasta sandwichi | SN77-1 | Unknown | MF796675 |
| Acasta sandwichi | SN77-2 | Unknown | MF796676 |
| Acasta sandwichi | WAM C47383 | Xestospongia testudinaria | MZ086797 |
| Acasta sandwichi | WAM C53344 | Xestospongia testudinaria | MZ086799 |
| Acasta sp. 1 MCY-2020 | SJP55-N1 | Mycale sp. | MN842026 [17] |
| Acasta sp. 2 MCY-2020 | SLQ20-2 | Haliclona sp. | MN842027 [17] |
| Acasta sp. 3 MCY-2020 | SN81-3 | Petrosia (Petrosia) sp. | MN842028 [17] |
| Acasta sp. 4 MCY-2020 | SDS4-2 | Unknown | MN842029 [17] |
| Acasta spongites | STI18-1 | Clathria sp. | MN842030 [17] |
| Acasta sulcata | SN48-1 | Callyspongia sp. | KY581617 [3] |
| Acasta sulcata | SN48-3 | Callyspongia sp. | KY581618 [3] |
| Acasta turriformis | SGI178-1 | Unknown | MF796671 |
| Acasta turriformis | SGI178-2 | Unknown | MF796672 |
| Acasta undulaterga | SNE47-3 | Jaspis splendens | KY581620 [3] |
| Acasta undulaterga | SNE47-5 | Jaspis splendens | KY581622 [3] |
| Acasta vipensis | CAS:IZ:187687A | Menella sp. | KF587277 [33] |
| Acasta vipensis | CAS:IZ:187764A | Menella sp. | KF587282 [33] |
| Euacasta acutaflava sp. nov. | WAM C58744 | Cymbastelastipitata | MZ086815 |
| Euacasta acutaflava sp. nov. | WAM C58745 | Cymbastelastipitata | MZ086816 |
| Euacasta acutaflava sp. nov. | WAM C58746 | Cymbastelastipitata | MZ086817 |
| Euacasta acutaflava sp. nov. | WAM C61434 | Cymbastela cf. stipitata | MZ086830 |
| Euacasta acutaflava sp. nov. | WAM C61435 | Cymbastela cf. stipitata | MZ086831 |
| Euacasta excoriatrix sp. nov. | WAM C61475 | Halichondria NW1 | MZ086832 |
| Euacasta excoriatrix sp. nov. | WAM C71800 | Halichondria BAR1 | MZ086842 |
| Euacasta excoriatrix sp. nov. | WAM C71801 | Halichondria BAR1 | MZ086843 |
| Euacasta dofleini | SGI168-2 | Haliclona sp. | MN842036 [17] |
| Euacasta microforamina | SJJ2-1 | Haliclona sp. | MN842037 [17] |
| Euacasta sporillus | RMNH:CRUS:C.10235 | Axinyssa sp. | KU986749 |
| Euacasta sporillus | S14-1 | Aaptos suberitoides | MN842038 [17] |
| Membranobalanus porphyrophilus | WAM C71853 | Spheciospongia purpurea | MK789771 [18] |
| Membranobalanus porphyrophilus | WAM C71881 | Spheciospongia purpurea | MK789772 [18] |
| Pecitnoacasta sculpturata | WAM C61394 | Amphinomiasulphurea | MZ086827 |
| Pecitnoacasta sculpturata | WAM C61395 | Amphinomiasulphurea | MZ086828 |
| Pecitnoacasta sculpturata | WAM C61396 | Amphinomiasulphurea | MZ086829 |
| Pectinoacasta cancellorum | WAM C50791 | Agelas KMB1 | MZ086798 |
| Pectinoacasta cancellorum | WAM C55302 | Agelas KMB1 | MZ086801 |
| Pectinoacasta cancellorum | WAM C55303 | Agelas KMB1 | MZ086802 |
| Pectinoacasta cancellorum | WAM C55304 | Agelas KMB1 | MZ086803 |
| Pectinoacasta cancellorum | WAM C55305 | Agelas KMB1 | MZ086804 |
| Pectinoacasta cancellorum | WAM C55306 | Agelas KMB1 | MZ086805 |
| Pectinoacasta cancellorum | WAM C55307 | Agelas KMB1 | MZ086806 |
| Pectinoacasta cancellorum | WAM C55308 | Agelas KMB1 | MZ086807 |
| Pectinoacasta pectinipes | WAM C58873 | Tedania (Trachytedania) L1 | MZ086826 |
| Pectinoacasta pectinipes | WAM C67748 | Tedania (Trachytedania) MM1 | MZ086835 |
| Pectinoacasta pectinipes | WAM C67749 | Tedania (Trachytedania) MM1 | MZ086836 |
| Pectinoacasta pectinipes | WAM C67750 | Tedania (Trachytedania) MM1 | MZ086837 |
| Pectinoacasta pectinipes | WAM C67772 | Tedania (Trachytedania) MM1 | MZ086838 |
| Pectinoacasta sculpturata | SGI131-1 | Agelas nemoechinata | MN842042 [17] |
Table 2.
Number of segments on each left (L) and right (R) cirrus (CI–CVI; anterior, posterior ramus) of species described herein, * denotes damaged cirrus.
Table 2.
Number of segments on each left (L) and right (R) cirrus (CI–CVI; anterior, posterior ramus) of species described herein, * denotes damaged cirrus.
| Specimen | CI | C II | C III | C IV | C V | C VI | |
|---|---|---|---|---|---|---|---|
| Acasta caveata sp. nov. | |||||||
| WAM C67716 | L | 17, 8 | 11, 8 | 15, 13 | 23, 26 | 32, 34 | 34, 33 |
| R | 17, 9 | 11, 10 | 15, 12 | 24, 30 | 33, 38 | 34, 36 | |
| Acasta fenestrata | |||||||
| WAM C58749 | L | 16, 6 | 9, 8 | 12, 10 | 26, 27 | 17*, 26 | 32, 33 |
| R | 16, 6 | 8, 7 | 11, 9 | 27, 26 | 23, 23 | 34, 31 | |
| Euacasta acutaflava sp. nov. | |||||||
| WAM C58744 | L | 20, 6 | 10, 7 | 14, 12 | 17, 22 | 29, 28 | 28*, 31 |
| R | 20, 6 | *,* | 11, 11 | 17*, 21 | 27, 28 | 29, 29 | |
| WAM C74601 | L | 17, 6 | 9, 7 | 12, 11 | 18, 24 | 28, 29 | 11*, 26* |
| R | 17, 6 | 11, 6 | 12, 12 | 15*, 22 | 29, 31 | 31, 22* | |
| WAM C76451 | L | 17, 5 | 8, 6 | 10, 9 | 19, 18* | 29, 30 | 33, 34 |
| R | 18, 5 | 8, 6 | 10, 9 | 19, 26 | 31, 32 | 33, 12* | |
| Euacasta excoriatrix sp. nov. | |||||||
| WAM C71800 | L | 30, 7 | 13, 9 | 17, 14 | 27, 29 | 32,29* | 33, 34 |
| R | 30, 7 | 12, 8 | 16, 13 | 22, 30 | 25*,31 | 37, 26* | |
| WAM C61475 | L | 29, 7 | 12, 8 | 16, 14 | 20, 21* | 26*,28* | 17*, 23* |
| R | 28, 6 | 13, 9 | 14, 15 | 14*, 22* | 30,22* | 20*, 22* | |
| Pectinoacasta cancellorum | |||||||
| WAM C55305 | L | 20, 6 | 10, 8 | 13, 11 | 21*, 16* | 15*, 17* | 35, 33 |
| R | 20, 6 | 10, 7 | 13, 11 | 22, 21 | 28, 29 | 36, 32 | |
| Pectinoacasta pectinipes | |||||||
| WAM C58871 | L | 15, 7 | 9, 8 | 11, 11 | 17, 19 | 21*, 23 | 22, 23 |
| R | 12, 7* | 9, 7 | 9, 9 | 3*, 18 | 23, 22 | 23, 23 | |
| Pectinoacasta sculpturata | |||||||
| WAM C61395 | L | 22, 6 | 9, 7 | 12, 5* | 14*, 23 | 24, 2* | 24, 24 |
| R | *,* | 9, 7 | 11, 10 | 22, 17* | 24, 25 | 26, 24 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hosie, A.M.; Fromont, J.; Munyard, K.; Jones, D.S. New Species and New Records of Sponge-Inhabiting Barnacles (Cirripedia, Balanidae, Acastinae) from Australia. Diversity 2021, 13, 290. https://doi.org/10.3390/d13070290
AMA Style
Hosie AM, Fromont J, Munyard K, Jones DS. New Species and New Records of Sponge-Inhabiting Barnacles (Cirripedia, Balanidae, Acastinae) from Australia. Diversity. 2021; 13(7):290. https://doi.org/10.3390/d13070290
Chicago/Turabian StyleHosie, Andrew M., Jane Fromont, Kylie Munyard, and Diana S. Jones. 2021. "New Species and New Records of Sponge-Inhabiting Barnacles (Cirripedia, Balanidae, Acastinae) from Australia" Diversity 13, no. 7: 290. https://doi.org/10.3390/d13070290
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.