

Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Strain Isolation
2.2. Morphological Observation
2.3. DNA Extraction and PCR Amplification
2.4. Detection for Microystins Synthesis Genes
2.5. Phylogenetic Analyses
2.6. Analyses of 16S–23S Internal Transcribed Spacer (ITS)
3. Results
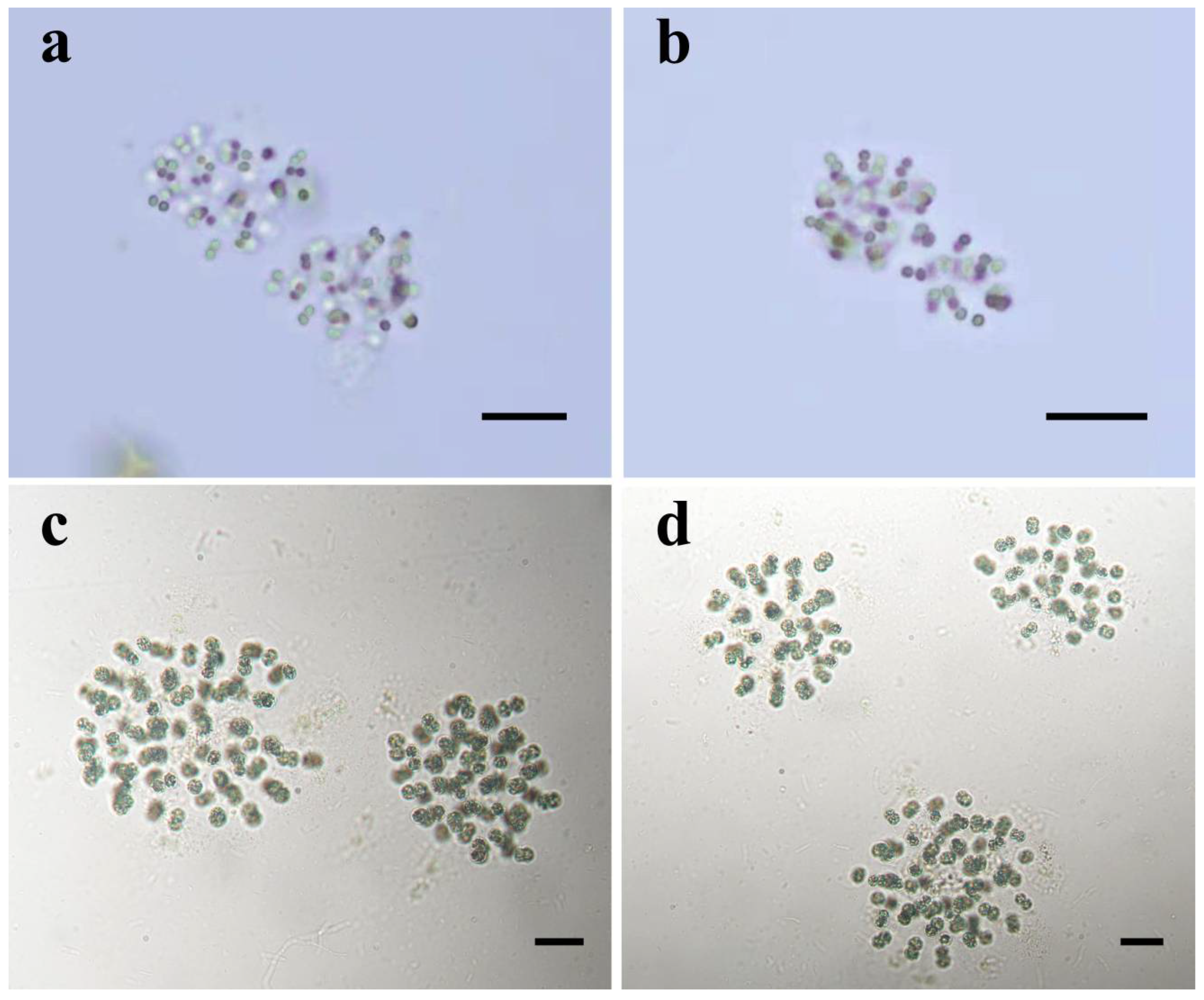
3.1. Morphological Description
3.2. Molecular and Phylogenetic Analyses
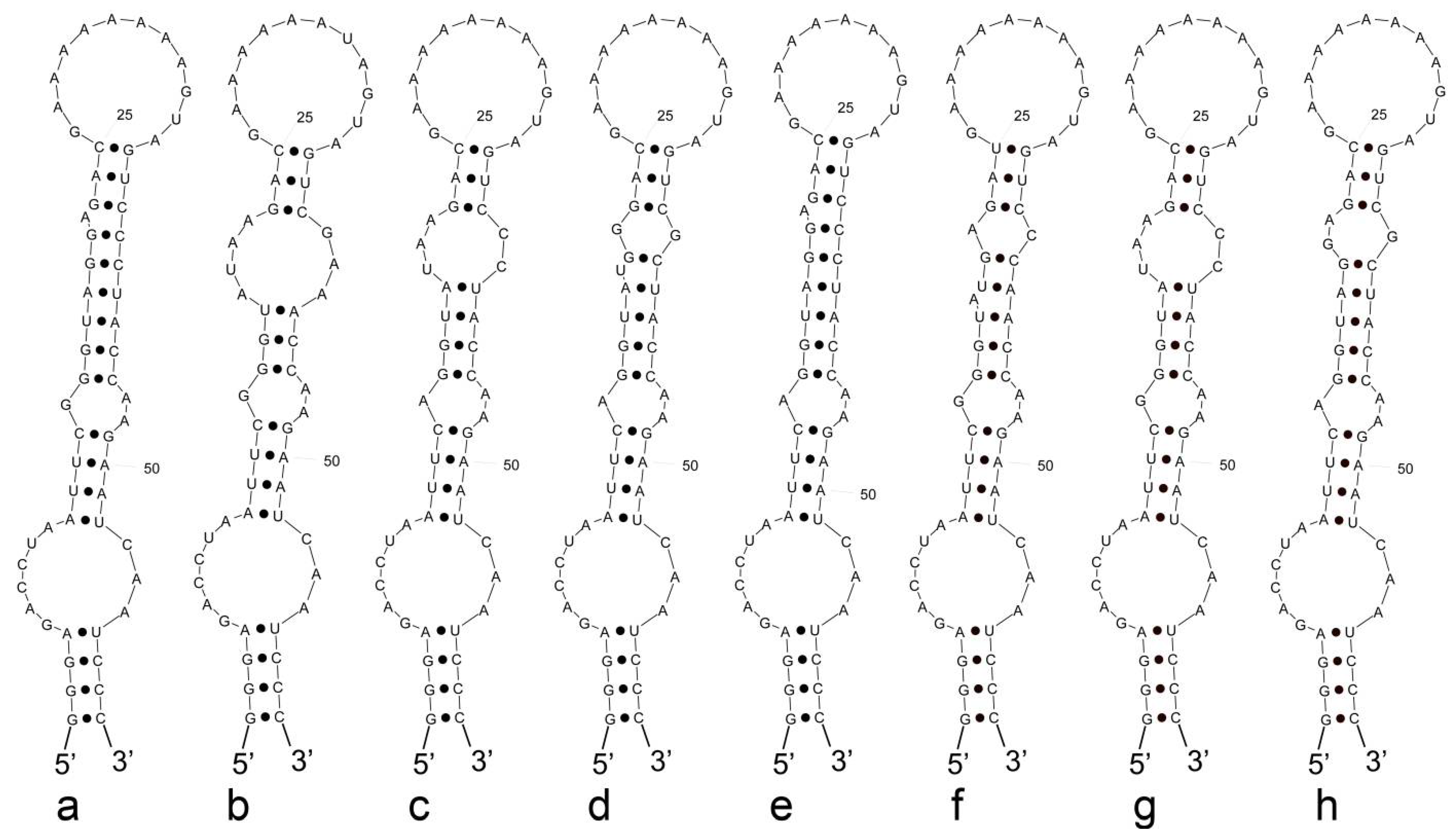
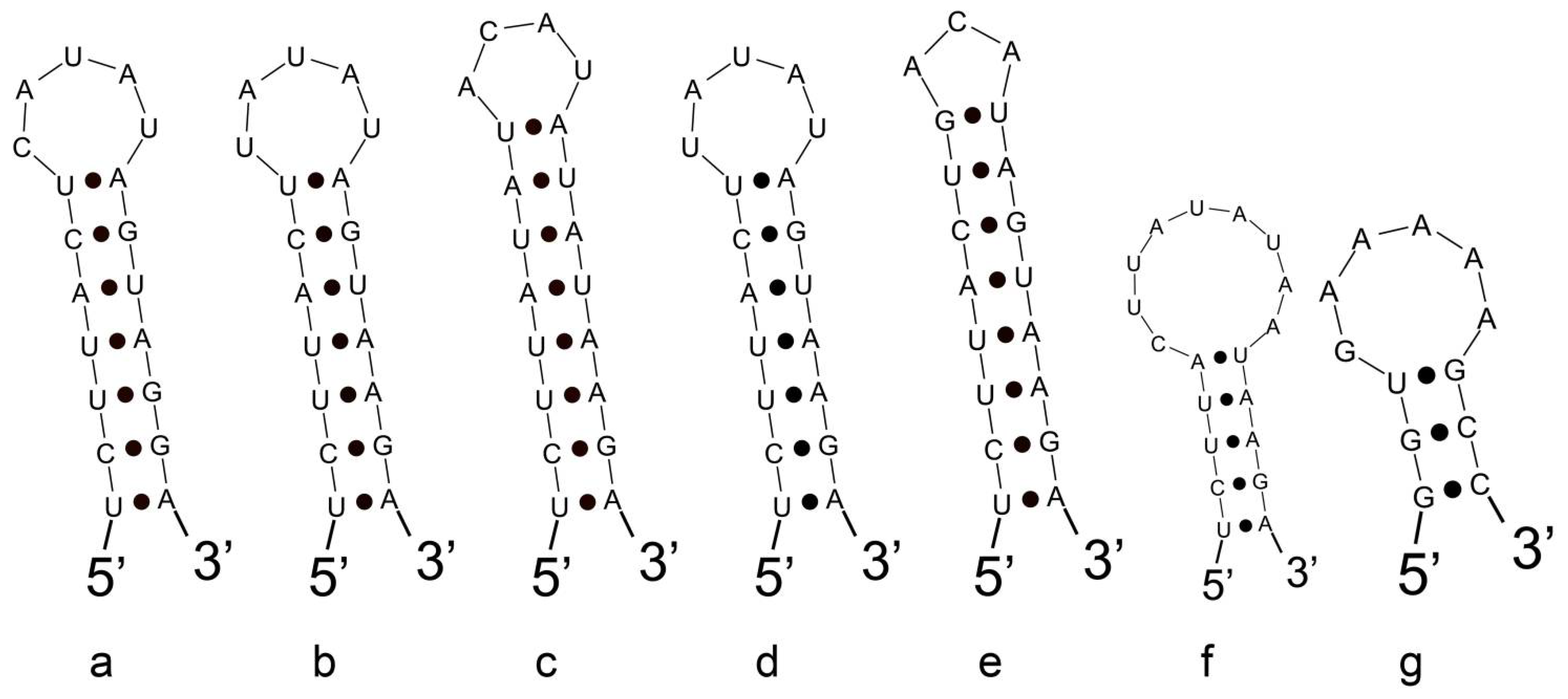
3.3. Analyses of ITS between 16S and 23S rRNA Gene and Secondary Structures
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Heisler, J.; Glibert, P.M.; Burkholder, J.M.; Anderson, D.M.; Cochlan, W.; Dennison, W.C.; Dortch, Q.; Gobler, C.J.; Heil, C.A.; Humphries, E.; et al. Eutrophication and harmful algal blooms: A scientific consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Climate—Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, J.M.; Davis, T.W.; Burford, M.A.; Gobler, C.J. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae 2012, 14, 313–334. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef]
- Huo, D.; Gan, N.; Geng, R.; Cao, Q.; Song, L.; Yu, G.; Li, R. Cyanobacterial blooms in China: Diversity, distribution, and cyanotoxins. Harmful Algae 2021, 109, 102106. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Komáková, J. Review of the European Microcystis-morphospecies (Cyanoprokaryotes) from nature. Czech Phycol. Olomouc 2002, 2, 1–24. [Google Scholar]
- Komárek, J.; Katovsk, J.; Mare, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Skuja, H. Taxonomie des Phytoplanktons einiger Seen in Uppland, Schweden. Symb. Bot. Ups. 1948, 9, 1–399. [Google Scholar]
- Hinoák, F. Two new species of the genus Radiocystis (Chroococcales, Cyanophyta/Cyanobacteria). Eur. J. Pediatr. Surg. 1996, 19, 2, 113–116. [Google Scholar] [CrossRef]
- Komárek, J.; Komárková-Legnerová, J. Radiocystis fernandoii, a new planktic cyanoprokariotic from tropical freshwater reservoirs. Preslia 1993, 65, 355–357. [Google Scholar]
- Rios, J.F.; Retz, D.C.L.; Leal, C.R.; Sant’Anna, C.L. The influence of temperature on Radiocystis fernandoi strain (cyanobacteria) growth and microcystin production. Braz. J. Bot. 2018, 41, 675–680. [Google Scholar]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen–Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef] [PubMed]
- Neilan, B.A.; Jacobs, D.; Goodman, A.E. Genetic diversity and phylogeny of toxic cyanobacteria determined by DNA polymorphisms within the phycocyanin locus. Appl. Environ. Microbiol. 1995, 61, 3875–3883. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blocker, H.; Emde, M.; Bottger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Rajaniemi, P.; Vardaka, E.; Moustaka-gouni, M.; Lanaras, T.; Sivonen, K. Limnothrix redekei (Van Goor) Meffert (Cyanobacteria) strains from Lake Kastoria, Greece form a separate phylogenetic group. Microb. Ecol. 2005, 49, 176–182. [Google Scholar] [CrossRef]
- Briand, E.; Escoffier, N.; Straub, C.; Sabart, M.; Quiblier, C.; Humbert, J.F. Spatiotemporal changes in the genetic diversity of a bloom-forming Microcystis aeruginosa (cyanobacteria) population. ISME J. 2009, 3, 4, 419–429. [Google Scholar] [CrossRef]
- Rinta-Kanto, J.M.; Ouellette, A.J.A.; Boyer, G.L.; Twiss, M.R.; Bridgeman, T.B.; Wilhelm, S.W. Quantification of toxic Microcystis spp. during the 2003 and 2004 blooms in western Lake Erie using quantitative real-time PCR. Environ. Sci. Technol. 2005, 39, 4198–4205. [Google Scholar] [CrossRef]
- Zuo, J.; Chen, L.; Shan, K.; Hu, L.; Song, L.; Gan, N. Assessment of different mcy genes for detecting the toxic to non-toxic Microcystis ratio in the field by multiplex qPCR. J. Oceanol. Limnol. 2018, 36, 1132–1144. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nuclc-Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovli, I.; Zou, H.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 7, 3022–3027. [Google Scholar] [CrossRef]
- Page, R.D.M. TreeView: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids-Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Starr, M.P. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic note: A placefor DNA–DNA reassociation and 16S rRNA sequence analysisin the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Ebers, J. Taxonomic parameters revisited: Tarnished gold standards. Microbiol. Today 2006, 33, 152–155. [Google Scholar]
- Xu, Y.; Wu, Z.; Yu, B.; Peng, X.; Yu, G.; Wei, Z.; Wang, G.; Li, R. Non-microcystin producing Microcystis wesenbergii (Komárek) Komárek (Cyanobacteria) representing a main water bloom-forming species in Chinese waters. Environ. Pollut. 2008, 156, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J. A review of water-bloom forming Microcystis species, with regard to population from Japan. Arch. Hydrobiol. (Suppl.)/Algol. Stud. 1991, 73, 115–127. [Google Scholar]
- Yu, G.L.; Song, L.R.; Li, R.H. Taxonomic notes on water bloom forming Microcystis species (Cyanophyta) from China—An example from samples of the Dianchi Lake. Acta Phytotaxon. Sin. 2007, 45, 727–741. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, B.; Wu, Z.; Xu, T.; Lu, Z. Microcystis panniformis—A newly recorded species of Microcystis in China. Sci. Limnol. Sin. 2012, 24, 647–650. [Google Scholar]
- Komárek, J.; Johansen, J. Freshwater Algae of North America; Wehr, J.D., Sheath, R., Kociolek, J.P., Eds.; Academic Press: Cambridge, MA, USA, 2015; p. 1050. [Google Scholar]
- Nägeli, C. Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschr. Der Allg. Schweiz. Ges. Für Die Gesammten Nat. 1849, 10, 1–139. [Google Scholar]
- Vieira, J.; Azevedo, M.; Azevedo, S.; Honda, R.Y.; Corrêa, B. Microcystin production by Radiocystis fernandoi (Chroococcales, cyanobacteria) isolated from a drinking water reservoir in the city of belém, pa, brazilian amazonia region. Toxicon 2003, 42, 709–713. [Google Scholar] [CrossRef]
- Vieira, J.M.D.S.; Azevedo, M.T.D.P.; Azevedo, S.M.F.D.O.; Honda, R.Y.; Corrêa, B. Toxic cyanobacteria and microcystin concentrations in a public water supply reservoir in the Brazilian Amazonia region. Toxicon 2005, 45, 7, 901–909. [Google Scholar] [CrossRef]
- Sant’Anna, C.L.A.; Werner, V.R.T.P.; Dogo, C.R.; Rios, F.R.D.C.; Azevedo, M.T.D.P.; De Carvalho, L.R. Review of toxic species of Cyanobacteria in Brazil. Algol. Stud. 2008, 126, 251–265. [Google Scholar] [CrossRef]
- Borges, P.A.F.; Train, S.; Dias, J.D.; Bonecker, C.C. Effects of fish farming on plankton structure in a Brazilian tropical reservoir. Hydrobiologia 2010, 649, 279–291. [Google Scholar] [CrossRef]
- Ferrari, G.; Pérez, M.D.C.; Dabezies, M.; Míguez, D.; Saizar, C. Planktic Cyanobacteria in the Lower Uruguay River, South America. J. Czech Phycol. Soc. 2011, 11, 225–234. [Google Scholar] [CrossRef]
- Fonseca, I.A.; Maniglia, T.C.; Rodrigues, L.; Prioli, A.J.; Prioli, S. Identification of the mcyA gene in natural blooms of Radiocystis fernandoi from a tributary of the Rosana reservoir, Brazil. Acta Sci. Biol. Sci. 2011, 33, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, M.; Pinto, F.C.R.; Vieira, J.M.S.; Honda, R.Y.; Pimenta, A.M.; Bemquerer, M.P.; Carvalho, L.; Kiyota, S. Isolation and structural characterization of microcystin-LR and three minor oligopeptides simultaneously produced by Radiocystis fernandoi (Chroococcales, Cyanobacteriae): A Brazilian toxic cyanobacterium. Toxicon Off. J. Int. Soc. Toxinol. 2006, 47, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.A.F.; Train, S.; Rodrigues, L.C. Estrutura do fitoplâncton, em curto período de tempo, em um braço do reservatório de Rosana (ribeirão do Corvo, Paraná, Brasil). Acta Sci. Anim. Sci. 2008, 30, 57–65. [Google Scholar]
- Azevedo, M.T.D.P.; Sant’Anna, C.L. Sphaerocavum, a new genus of planktic cyanobacteria from continental water bodies in Brazil. Algol. Stud. 2003, 109, 79–92. [Google Scholar]
- Wood, S.; Crowe, A.L.M.; Ruck, J.G.; Wear, R.G. New records of planktonic cyanobacteria in New Zealand freshwaters. N. Zeal. J. Bot. 2005, 43, 479–492. [Google Scholar] [CrossRef]
- Rigonato, J.; Sant’Anna, C.L.; Giani, A.; Azevedo, M.T.P.; Gama, W.A.; Viana, V.F.L.; Fiore, M.F.; Werner, V.R. Sphaerocavum: A coccoid morphogenus identical to Microcystis in terms of 16S-rDNA and ITS sequence phylogenies. Hydrobiologia 2018, 811, 35–48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. Microcystis fernandoi WZU1501 | |||||||||||||||
| 2. Microcystis aeruginosa BKP SB69 | 99.85 | ||||||||||||||
| 3. Microcystis aeruginosa FACHB-905 | 99.54 | 99.70 | |||||||||||||
| 4. Microcystis wesenbergii TAC52 | 99.70 | 99.85 | 99.85 | ||||||||||||
| 5. Microcystis wesenbergii NRERC-212 | 99.62 | 99.77 | 99.77 | 99.92 | |||||||||||
| 6. Microcystis viridis NIES-102 | 99.70 | 99.85 | 99.70 | 99.70 | 99.62 | ||||||||||
| 7. Microcystis viridis TAC17 | 99.70 | 99.85 | 99.70 | 99.70 | 99.62 | 100.00 | |||||||||
| 8. Microcystis ichthyoblabe VN325 | 99.70 | 99.85 | 98.70 | 99.85 | 99.77 | 99.70 | 99.70 | ||||||||
| 9. Microcystis ichthyoblabe NRERC-217 | 99.77 | 99.92 | 99.77 | 99.92 | 99.85 | 99.77 | 99.77 | 99.92 | |||||||
| 10. Microcystis flos-aquae NRERC 218 | 99.62 | 99.77 | 99.62 | 99.77 | 99.70 | 99.62 | 99.62 | 99.92 | 99.85 | ||||||
| 11. Microcystis flos-aquae NRERC-211 | 99.62 | 99.77 | 99.62 | 99.77 | 99.70 | 99.62 | 99.62 | 99.92 | 99.85 | 99.85 | |||||
| 12. Microcystis novacekii CHAB1444 | 99.54 | 99.70 | 99.70 | 99.85 | 99.77 | 99.54 | 99.54 | 99.70 | 99.77 | 99.62 | 99.62 | ||||
| 13. Microcystis novacekii TAC65 | 99.47 | 99.62 | 99.47 | 99.62 | 99.54 | 99.62 | 99.62 | 99.62 | 99.70 | 99.54 | 99.54 | 99.62 | |||
| 14. Microcystis bengalensis VN486 | 99.62 | 99.77 | 99.62 | 99.62 | 99.54 | 99.77 | 99.77 | 99.62 | 99.70 | 99.54 | 99.54 | 99.62 | 99.70 | ||
| 15. Radiocystis sp. JJ30-3 | 91.05 | 91.22 | 91.31 | 91.31 | 91.22 | 91.22 | 91.22 | 91.31 | 91.22 | 91.22 | 91.22 | 91.14 | 90.88 | 90.96 | |
| 16. Radiocystis geminata TAU-MAC 1214 | 90.71 | 90.88 | 90.97 | 90.97 | 90.88 | 90.88 | 90.88 | 90.97 | 90.88 | 90.88 | 90.88 | 90.80 | 90.53 | 90.64 | 99.45 |
| Strain | Genbank | Complete ITS (nt) | D1-D1′ Helix (nt) | tRNAIle | tRNAAla | Box-B Helix (nt) | Box-A Helix (nt) | D4 | V3 Helix (nt) | D5 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1. Microcystis fernandoi WZU1501 | OP279644 | 357 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 2. Microcystis wesenbergii NIES111 | AB015388.1 | 361 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 3. Microcystis viridis NIES 103 | AB254444.1 | 358 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 4. Microcystis novacekii VN517 | AB665999.1 | 359 | 58 | + | - | 19 | 14 | 7 | 12 | 17 |
| 5. Microcystis ichthyoblabe VN512 | AB666031.1 | 362 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 6. Microcystis panniformis VN516 | AB666041.1 | 359 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 7. Microcystis pseudofilamentosa VN511 | AB666052.1 | 358 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 8. Microcystis aeruginosa PCC 7941 | MH620004.1 | 359 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| 9. Microcystis aeruginosa IPPAS B-1527 | MW396712.1 | 364 | 59 | + | - | 19 | 14 | 7 | 12 | 17 |
| Strain | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| 1. Microcystis fernandoi WZU1501 | ||||||||
| 2. Microcystis wesenbergii NIES111 | 95.44 | |||||||
| 3. Microcystis viridis NIES 103 | 95.73 | 98.86 | ||||||
| 4. Microcystis novacekii VN517 | 96.30 | 96.30 | 96.58 | |||||
| 5. Microcystis ichthyoblabe VN512 | 94.87 | 96.87 | 96.87 | 98.29 | ||||
| 6. Microcystis panniformis VN516 | 95.44 | 96.30 | 96.30 | 97.72 | 97.72 | |||
| 7. Microcystis pseudofilamentosa VN511 | 97.15 | 94.87 | 94.87 | 94.59 | 94.30 | 94.87 | ||
| 8. Microcystis aeruginosa PCC 7941 | 96.87 | 95.16 | 95.16 | 94.87 | 94.59 | 94.59 | 99.72 | |
| 9. Microcystis aeruginosa IPPAS B-1527 | 97.44 | 95.16 | 95.16 | 94.87 | 95.16 | 95.73 | 97.44 | 97.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, X.; Cheng, Y.; Zhang, S.; Hu, Z.; Xiao, P.; Zhang, H.; Geng, R.; Li, R. Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China. Diversity 2022, 14, 816. https://doi.org/10.3390/d14100816
Lv X, Cheng Y, Zhang S, Hu Z, Xiao P, Zhang H, Geng R, Li R. Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China. Diversity. 2022; 14(10):816. https://doi.org/10.3390/d14100816
Chicago/Turabian StyleLv, Xingju, Yao Cheng, Silong Zhang, Zhaowen Hu, Peng Xiao, He Zhang, Ruozhen Geng, and Renhui Li. 2022. "Polyphasic Characterization and Taxonomic Evaluation of a Bloom-Forming Strain Morphologically Resembling Radiocystis fernandoi (Chroococcales, Cyanobacteria) from Lake Erhai, China" Diversity 14, no. 10: 816. https://doi.org/10.3390/d14100816