
A Phylogenetic Re-Evaluation of the Stenakrine Opecoelids (Trematoda, Digenea: Opecoeloidea) with Some Taxonomic Novelties †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Morphological Study
2.3. DNA Amplification and Sequencing
2.4. Phylogenetic Analysis
3. Results
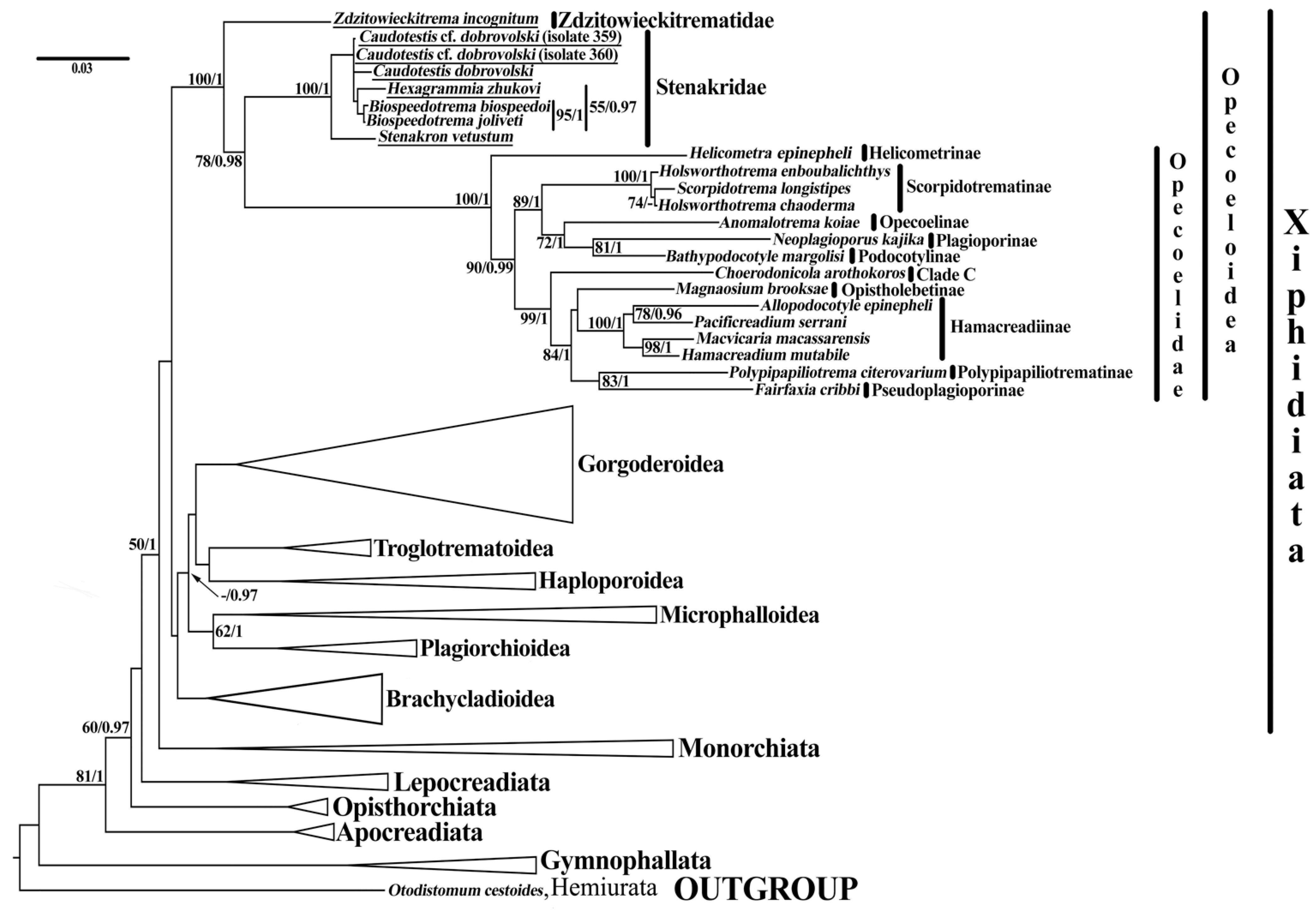
3.1. Phylogeny
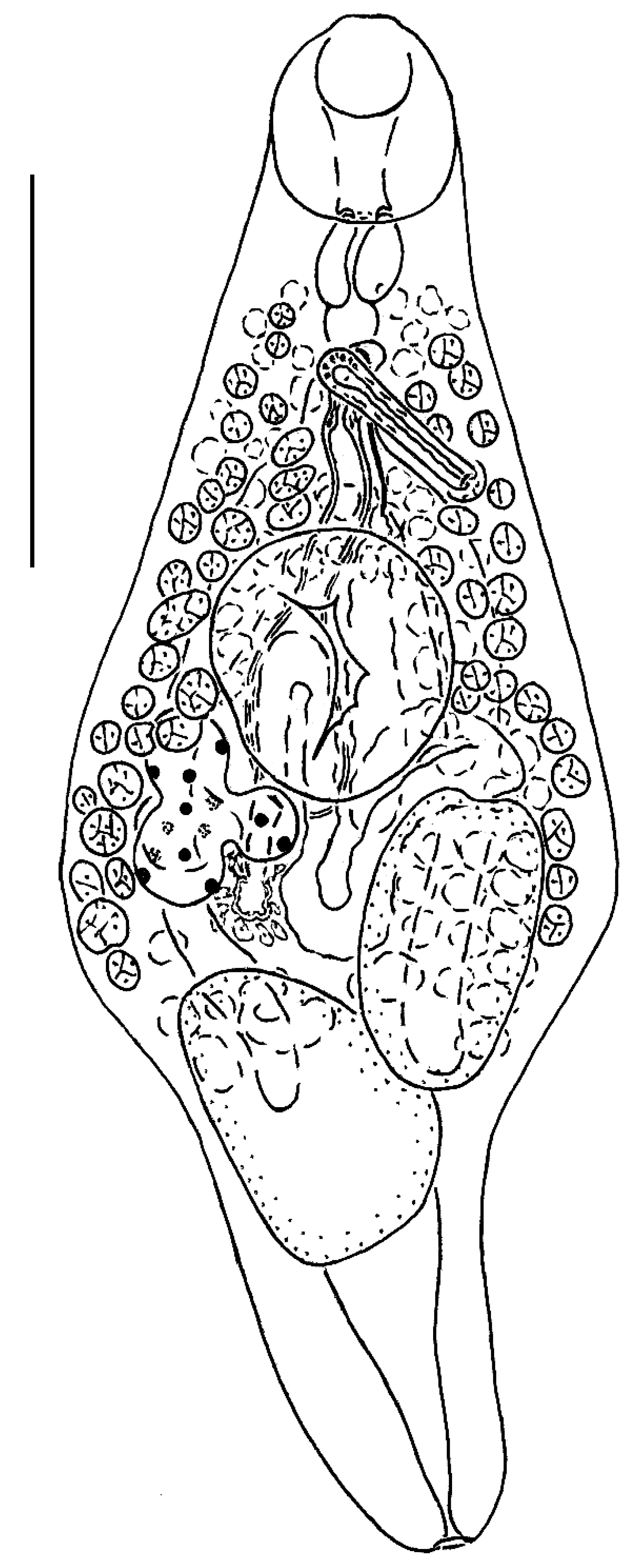
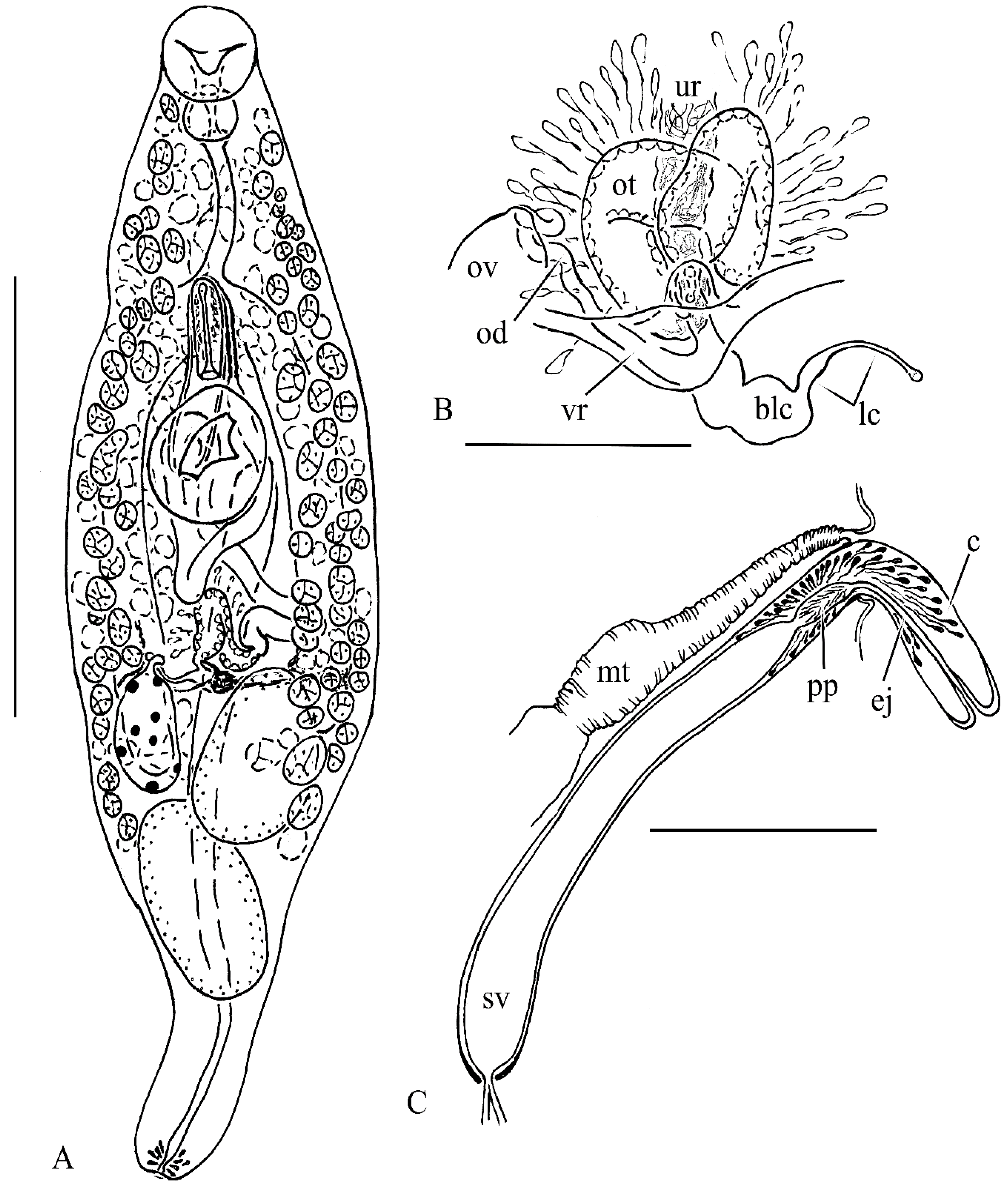
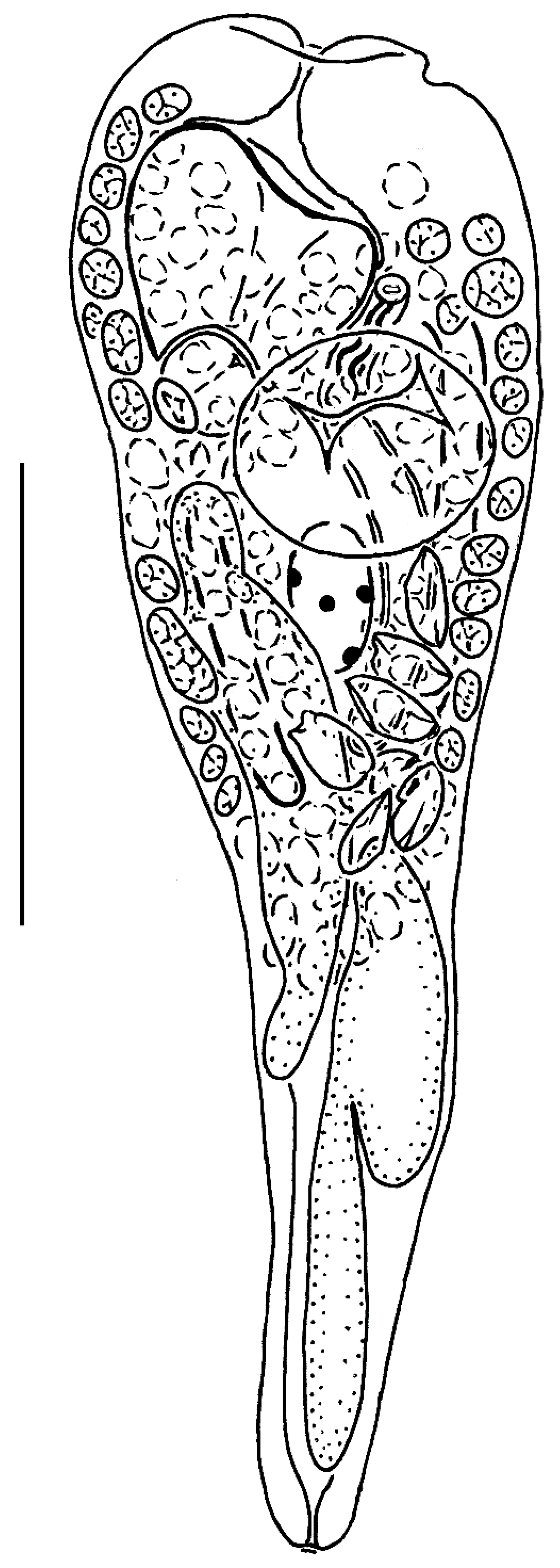
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Littlewood, D.T.J.; Bray, R.A.; Waeschenbach, A. Phylogenetic patterns of diversity in cestodes and trematodes. In Parasite Diversity and Diversification: Evolutionary Ecology Meets Phylogenetics; Morand, S., Krasnov, B., Littlewood, D.T.J., Eds.; Cambridge University Press: Cambridge, UK, 2015; pp. 304–319. [Google Scholar] [CrossRef]
- Pérez-Ponce de León, G.; Hernández-Mena, D.I. Testing the higher-level phylogenetic classification of Digenea (Platyhelminthes, Trematoda) based on nuclear rDNA sequences before entering the age of the ‘next-generation’ Tree of Life. J. Helminthol. 2019, 93, 260–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.B.; Cutmore, S.C.; Cribb, T.H. The Pseudoplagioporinae, a new subfamily in the Opecoelidae Ozaki, 1925 (Trematoda) for a small clade parasitizing mainly lethrinid fishes, with three new species. J. Zool. Syst. Evol. Res. 2020, 58, 79–113. [Google Scholar] [CrossRef]
- Martin, S.B.; Downie, A.J.; Cribb, T.H. A new subfamily for a clade of opecoelids (Trematoda: Digenea) exploiting marine fishes as second-intermediate hosts, with the first report of opecoelid metacercariae from an elasmobranch. Zool. J. Linn. Soc. 2020, 188, 455–472. [Google Scholar] [CrossRef]
- Karar, Y.F.M.; Blend, C.K.; Mohamadain, H.S.; Khalifa, R.M.A.; Dronen, N.O. Unusual opecoelids from Red Sea triggerfishes with special reference to characteristic concepts of the Opistholebetinae Fukui, 1929 (Digenea: Opecoelidae). Zootaxa 2020, 4834, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, S.G.; Shchenkov, S.V.; Gordeev, I.I. Phylogenetic position of deep-sea opecoelid digenean Tellervotrema beringi (Mamaev, 1965) (Trematoda: Opecoelidae) based on novel genetic data. Syst. Parasitol. 2020, 97, 379–387. [Google Scholar] [CrossRef]
- Cribb, T.H. Family Opecoelidae Ozaki, 1925. In Keys to the Trematoda; Jones, A., Bray, R.A., Gibson, D.I., Eds.; CABI Publishing and The Natural History Museum: Wallingford, UK, 2005; Volume 2, pp. 443–531. [Google Scholar] [CrossRef]
- Bray, R.A.; Waeschenbach, A.; Dyal, P.; Littlewood, D.T.J.; Morand, S. New digeneans (Opecoelidae) from hydrothermal vent fishes in the south eastern Pacific Ocean, including one new genus and five new species. Zootaxa 2014, 3768, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.B.; Huston, D.C.; Cutmore, S.C.; Cribb, T.H. A new classification for deep-sea opecoelid trematodes based on the phylogenetic position of some unusual taxa from shallow-water, herbivorous fishes off south-west Australia. Zool. J. Linn. Soc. 2019, 186, 385–413. [Google Scholar] [CrossRef]
- Yamaguti, S. Digenetic Trematodes of Hawaiian Fishes; Keigaku Publishing Company: Tokyo, Japan, 1970. [Google Scholar]
- Bray, R.A.; Cribb, T.H.; Littlewood, D.T.J.; Waeschenbach, A. The molecular phylogeny of the digenean family Opecoelidae Ozaki, 1925 and the value of morphological characters, with the erection of a new subfamily. Folia Parasitol. 2016, 63, 013. [Google Scholar] [CrossRef]
- Bray, R.A. Digenea in marine fishes from the eastern seaboard of Canada. J. Nat. Hist. 1979, 13, 399–431. [Google Scholar] [CrossRef]
- Baeva, O.M. Helminth fauna of Far Eastern mackerel. In Parasitic Worms of Domestic and Wild Animals; Papers on Helminthology Presented to Prof. A.A. Sobolev on the 40th Anniversary of His Scientific and Teaching Activity; Leonov, V.A., Mamaev, Y.L., Oshmarin, P.G., Eds.; Dalnevostochnii Gosudarstvennii Universitet: Vladivostok, Russia, 1965; pp. 23–32. (In Russian) [Google Scholar]
- Issaitschikov, I.M. Contributions to parasitic worms of some groups of vertebrates from Russian Arctic. Tr. Morskogo Nauchnogo Inst. 1928, 3, 1–79. (In Russian) [Google Scholar]
- Manter, H.W. Some digenetic trematodes from deep-water fish of Tortugas, Florida. Pap. Tortugas Lab. 1934, 28, 257–345. [Google Scholar]
- Srivastava, H.D. New allocreadiids (Trematoda) from Indian marine food fishes. Part IV. The morphology and systematic position of a new genus, Horatrema, of digenetic trematodes. Parasitology 1942, 34, 128–132. [Google Scholar] [CrossRef]
- Yamaguti, S. Systema helminthum, Volume I.; Interscience Publishers: New York, NY, USA; London, UK, 1958. [Google Scholar]
- Sokolov, S.G.; Lebedeva, D.I.; Shchenkov, S.V.; Gordeev, I.I. Caudotestis dobrovolski n. sp. (Trematoda, Xiphidiata) in North Pacific scorpaeniform fish: A crisis of concept of the opecoelid subfamily Stenakrinae Yamaguti, 1970. J. Zool. Syst. Evol. Res. 2020, 58, 1111–1122. [Google Scholar] [CrossRef]
- Sokolov, S.G.; Lebedeva, D.I.; Gordeev, I.I.; Khasanov, F.K. Zdzitowieckitrema incognitum gen. et sp. nov. (Trematoda: Xiphidiata) from the Antarctic fish Muraenolepis marmorata Günther, 1880 (Gadiformes: Muraenolepidae): Ordinary morphology but unclear family affiliation. Mar. Biodivers. 2019, 49, 451–462. [Google Scholar] [CrossRef]
- Martin, S.B.; Crouch, K.; Cutmore, S.C.; Cribb, T.H. Expansion of the concept of the Opistholebetinae Fukui, 1929 (Digenea: Opecoelidae Ozaki, 1925), with Magnaosimum brooksae n. g., n. sp. from Tripodichthys angustifrons (Hollard) (Tetraodontiformes: Triacanthidae) in Moreton Bay, Australia. Syst. Parasitol. 2018, 95, 121–132. [Google Scholar] [CrossRef]
- Martin, S.B.; Cutmore, S.C.; Cribb, T.H. Revision of Neolebouria Gibson, 1976 (Digenea: Opecoelidae), with Trilobovarium n. g., for species infecting tropical and subtropical shallow-water fishes. Syst. Parasitol. 2017, 94, 307–338. [Google Scholar] [CrossRef]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Bartosová, P.; Fiala, I.; Hypsa, V. Concatenated SSU and LSU rDNA data confirm the main evolutionary trends within myxosporeans (Myxozoa: Myxosporea) and provide an effective tool for their molecular phylogenetics. Mol. Phylogenet. Evol. 2009, 53, 81–93. [Google Scholar] [CrossRef]
- Petrov, N.B.; Vladychenskaya, N.S. Phylogeny of molting Protostomes (Ecdysozoa) as inferred from 18S and 28S rRNA gene sequences. Mol. Biol. 2005, 39, 503–513. [Google Scholar] [CrossRef]
- Jamy, M.; Foster, R.; Barbera, P.; Czech, L.; Kozlov, A.; Stamatakis, A.; Bending, G.; Hilton, S.; Bass, D.; Burki, F. Long-read metabarcoding of the eukaryotic rDNA operon to phylogenetically and taxonomically resolve environmental diversity. Mol. Ecol. Resour. 2020, 20, 429–443. [Google Scholar] [CrossRef]
- Gadagkar, S.R.; Rosenberg, M.S.; Kumar, S. Inferring species phylogenies from multiple genes: Concatenated sequence tree versus consensus gene tree. J. Exp. Zool. B Mol. Dev. Evol. 2005, 304, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Littlewood, D.T.J.; Olson, P. Small subunit rDNA and the Platyhelminthes: Signal, noise, conflict and compromise. In Interrelationships of the Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor and Francis: London, UK, 2001; pp. 262–278. [Google Scholar]
- Tkach, V.; Pawlowski, J.; Mariaux, J. Phylogenetic analysis of the suborder Plagiorchiata (Platyhelminthes, Digenea) based on partial lsrDNA sequences. Int. J. Parasitol. 2000, 30, 83–93. [Google Scholar] [CrossRef]
- Morgan, J.A.T.; Blair, D. Relative merits of nuclear ribosomal internal transcribed spacers and mitochondrial CO1 and ND1 genes for distinguishing among Echinostoma species (Trematoda). Parasitology 1998, 116, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Blasco-Costa, I.; Balbuena, J.A.; Kostadinova, A.; Olson, P.D. Interrelationships of the Haploporinae (Digenea: Haploporidae): A molecular test of the taxonomic framework based on morphology. Parasitol. Int. 2009, 58, 265–269. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Gibson, D.I.; Balbuena, J.A.; Raga, J.A.; Kostadinova, A. A revision of the Haploporinae Nicoll, 1914 (Digenea: Haploporidae) from mullets (Mugilidae): Haploporus Looss, 1902 and Lecithobotrys Looss, 1902. Syst. Parasitol. 2009, 73, 107–133. [Google Scholar] [CrossRef]
- Bray, R.A.; Cribb, T.H.; Waeschenbach, A.; Littlewood, D.T.J. A new species of Stephanostomum Looss, 1899 (Digenea, Acanthocolpidae) with a bizarre oral sucker: S. adlardi sp. nov. from the common coral trout Plectropomus leopardus (Lacepède, 1802) (Perciformes, Serranidae) from Lizard Island, Great Barrier Reef. Acta Parasitol. 2007, 52, 206–212. [Google Scholar] [CrossRef]
- Bray, R.A.; Foster, G.N.; Waeschenbach, A.; Littlewood, D.T.J. The discovery of progenetic Allocreadium neotenicum Peters, 1957 (Digenea: Allocreadiidae) in water beetles (Coleoptera: Dytiscidae) in Great Britain. Zootaxa 2012, 3577, 58–70. [Google Scholar] [CrossRef]
- Bray, R.A.; Webster, B.; Bartoli, P.; Littlewood, D.T.J. Relationships within the Acanthocolpidae Lühe, 1906 and their place among the Digena. Acta Parasiol. 2005, 50, 281–291. [Google Scholar]
- Briscoe, A.G.; Bray, R.A.; Brabec, J.; Littlewood, D.T.J. The mitochondrial genome and ribosomal operon of Brachycladium goliath (Digenea: Brachycladiidae) recovered from a stranded minke whale. Parasitol. Int. 2016, 65, 271–275. [Google Scholar] [CrossRef]
- Devi, K.R.; Narain, K.; Agatsuma, T.; Blair, D.; Nagataki, M.; Wickramasinghe, S.; Yatawara, L.; Mahanta, J. Morphological and molecular characterization of Paragonimus westermani in northeastern India. Acta Trop. 2010, 116, 311–318. [Google Scholar] [CrossRef]
- Hill-Spanik, K.M.; Sams, C.; Connors, V.A.; Bricker, T.; de Buron, I. Molecular data reshape our understanding of the life cycles of three digeneans (Monorchiidae and Gymnophallidae) infecting the bivalve, Donax variabilis: It’s just a facultative host! Parasite 2021, 28, 34. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.B.; Cribb, T.H.; Cutmore, S.C.; Huston, D.C. The phylogenetic position of Choerodonicola Cribb, 2005 (Digenea: Opecoelidae) with a partial life-cycle for a new species from the blue-barred parrotfish Scarus ghobban Forsskål (Scaridae) in Moreton Bay, Australia. Syst. Parasitol. 2018, 95, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.B.; Sasal, P.; Cutmore, S.C.; Ward, S.; Aeby, G.S.; Cribb, T.H. Intermediate host switches drive diversification among the largest trematode family: Evidence from the Polypipapiliotrematinae n. subf. (Opecoelidae), parasites transmitted to butterflyfishes via predation of coral polyps. Int. J. Parasitol. 2018, 48, 1107–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shchenkov, S.V.; Sokolov, S.G.; Tsushko, K.M.; Denisova, S.A. Is Gymnophallus Odhner, 1900 (Trematoda: Gymnophallidae) polyphyletic? A new hypothesis based on phylogenetic position of Gymnophallus deliciosus (Olsson, 1893). Parasitol. Res. 2022, 121, 2157–2160. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, S.G.; Shchenkov, S.V.; Gordeev, I.I. Records of opecoeline species Pseudopecoelus cf. vulgaris and Anomalotrema koiae Gibson & Bray, 1984 (Trematoda, Opecoelidae, Opecoelinae) from fish of the North Pacific, with notes on the phylogeny of the family Opecoelidae. J. Helminthol. 2019, 93, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Purschke, G. The sister group relationship of Aeolosomatidae and Potamodrilidae (Annelida: “Polychaeta”)—A molecular phylogenetic approach based on 18S rDNA and cytochrome oxidase I. Zool. Anz. 2005, 243, 281–293. [Google Scholar] [CrossRef]
- Thaenkham, U.; Nawa, Y.; Blair, D.; Pakdee, W. Confirmation of the paraphyletic relationship between families Opisthorchiidae and Heterophyidae using small and large subunit ribosomal DNA sequences. Parasitol. Int. 2011, 60, 521–523. [Google Scholar] [CrossRef]
- Voronova, A.N.; Chelomina, G.N.; Besprozvannykh, V.V.; Tkach, V.V. Genetic divergence of human pathogens Nanophyetus spp. (Trematoda: Troglotrematidae) on the opposite sides of the Pacific Rim. Parasitology 2017, 144, 601–612. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: http://www.r-project.org/index.html (accessed on 5 July 2022).
- Paradis, E.; Schliep, K. Ape 5.0: An Environment for Modern Phylogenetics and Evolutionary Analyses in R. Bioinformatics 2019, 35, 526–528. [Google Scholar] [CrossRef]
- Nylander, J.A.A. catfasta2phyml. Available online: https://github.com/nylander/catfasta2phyml (accessed on 5 July 2022).
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Sweden, Uppsala, 2004. [Google Scholar]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop, New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Register of Marine Species. Stenakrinae Yamaguti. 1970. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=391437 (accessed on 5 June 2022).
- Layman, E.M. Parasitic worms from the fishes of Peter The Great Gulf. Izv. Tikhookeanskoi Nauchno-Promysl. Stantsii 1930, 3, 1–120. [Google Scholar]
- Mamaev, Y.L.; Parukhin, A.M.; Baeva, O.M. Parasitic Worms of Flatfish from the Far Eastern Seas In: Parasitic Worms of Animals of the Primorye Region and the Pacific Ocean; Oshmarin, P.G., Ed.; Akademia Nauk SSSR: Moscow, Russia, 1963; pp. 82–113. (In Russian) [Google Scholar]
- Miller, M.J. A critical study of Stafford’s report on “Trematodes of Canadian fishes” based on his trematode collection. Can. J. Res. 1941, 19, 28–52. [Google Scholar] [CrossRef]
- Gaevskaya, A.V.; Kovaleva, A.A. New species of trematodes from fishes of the Falkland-Patagonian region (South-Western Atlantic). Parazitologiya 1977, 11, 417–423. [Google Scholar]
- Gibson, D.I. Part IV. Trematoda. In Guide to the Parasites of Canadian Fishes; NRC Research Press: Ottawa, ON, Canada, 1996. [Google Scholar]
- Zhukov, E.V. Endoparasitic worms of the fishes in the Sea of Japan and South-Kuril shallow-waters. Tr. Zool. Inst. Akad. Nauk. SSSR. 1960, 28, 3–146. (In Russian) [Google Scholar]
- Kuramochi, T. Digenean trematodes of fishes from deep-sea areas off the Pacific coast of northern Honshu, Japan. In Deep-Sea Fauna and Pollutants off Pacific Coast of Northern Japan; Fujita, T., Ed.; National Museum of Nature and Sciences: Tokyo, Japan, 2009; pp. 25–37. [Google Scholar]
- Strelkov, Y.A. Endoparasitic worms of marine fishes of East Kamchatka. Tr. Zool. Inst. Akad. Nauk SSSR. 1960, 28, 147–196. (In Russian) [Google Scholar]
- Yamaguti, S. Studies on the helminth fauna of Japan. Part 2.Trematodes of fishes. I. Jpn. J. Zool. 1934, 5, 249–541. [Google Scholar]
- Yamaguti, S. Studies on the helminth fauna of Japan. Part 44.Trematodes of fishes, IX. Arb. Aus Der Med. Univ. Zu Okayama 1951, 7, 247–282. [Google Scholar] [CrossRef]
- Yamaguti, S. Synopsis of Digenetic Trematodes of Vertebrates, Volume 1; Keigaku Publishing Company: Tokyo, Japan, 1971. [Google Scholar]
- Gibson, D.I.; Bray, R.A. A study and reorganization of Plagioporus Stafford, 1904 (Digenea: Opecoelidae) and related genera, with special reference to forms from European Atlantic waters. J. Nat. Hist. 1982, 16, 529–559. [Google Scholar] [CrossRef]
- Price, E.W. New digenetic trematodes from marine fishes. Smithson. Misc. Collect. 1934, 91, 1–8. [Google Scholar]
- Manter, H.W. Some digenetic trematodes from fishes of New Zealand. Trans. Proc. R. Soc. N. Z. 1954, 82, 475–568. [Google Scholar]
- Paggi, L.; Orecchia, P. Su di una nuova specie Plagioporus (Caudotestis) tyrrhenicus sp. n. e su due nuovi reperti parassitologici in Blennius pavo Risso, 1810. Parassitologia 1976, 18, 21–32. (In Italian) [Google Scholar] [PubMed]
- Wang, P.Q. Some digenetic trematodes of marine fishes from Fujian Province, China. Oceanol. Limnol. Sin. 1982, 3, 179–194. (In Chinese) [Google Scholar]
- Toman, G. Digenetic trematodes of marine teleost fishes from the Seychelles, Indian Ocean. III. Acta Parasitol. 1992, 37, 119–126. [Google Scholar]
- Yamaguti, S. Studies on the Helminth Fauna of Japan; Part 21. Trematodes of Fishes, IV.; Yamaguti, S., Ed.; Publisher: Kyöto, Japan, 1938. [Google Scholar]
- Yamaguti, S. Studies on the helminth fauna of Japan. Part 54. Trematodes of fishes, XII. Publ. Seto Mar. Biol. Lab. 1959, 7, 241–262. [Google Scholar] [CrossRef] [Green Version]
- Pogoril’tseva, T.P. New species of digenetic trematodes from fishes in the Black Sea. Nauk. Zap. Kiev. Derjavnogo Pedagog. Inst. 1954, 25, 133–137. (In Ukrainian) [Google Scholar]
- Machida, M. Four new species of digenean trematodes from wrasses of southern Japan and neighboring waters. Bull. Natn. Sci. Mus. Tokyo Ser. A 2004, 30, 105–111. [Google Scholar]
- Brinkmann, A. Trematodes from Greenland. Medd. Om Grnland Udgivne Af Komm. Vidensk. Undersgelser I Grnland 1975, 205, 1–88. [Google Scholar]
- Poljansky, Y.I. Parasites of the fish of the Barents Sea. Tr. Zool. Inst. Akad. Nauk SSSR. 1955, 19, 5–170. (In Russian) [Google Scholar]
- Cantatore, D.M.P.; Lancia, J.P.; Lanfranchi, A.L.; Timi, J.T. A new species of Caudotestis (Digenea: Opecoelidae) parasitizing the horsefish, Congiopodus peruvianus (Scorpaeniformes: Congiopodidae) from the Patagonian Shelf, Argentina. J. Parasitol. 2012, 98, 1144–1147. [Google Scholar] [CrossRef]
- Zdzitowiecki, K. Digenea of fishes of the Weddell Sea. IV. Three opecoelid species of the genera Neolebouria, Helicometra and Stenakron. Acta Parasitol. 1997, 42, 138–143. [Google Scholar]
- Prudhoe, S.; Bray, R.A. Digenetic trematodes from fishes. B.A.N.Z. Antarct. Res. Exped. Rep. Ser. B 1973, 8, 195–225. [Google Scholar]
- Schell, S.C. Three new species of digenetic trematodes from Puget Sound fishes. Proc. Helminthol. Soc. Wash. 1973, 40, 227–230. [Google Scholar]
- Avise, J.C.; Johns, G.C. Proposal for a standardized temporal scheme of biological classification for extant species. Proc. Natl. Acad. Sci. USA 1999, 96, 7358–7363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galaktionov, K.V.; Dobrovolskij, A.A. The Biology and Evolution of Trematodes: An Essay on the Biology, Morphology, Life Cycles, Transmissions, and Evolution of Digenetic Trematodes; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003. [Google Scholar]
- Chubrik, G.K. Life cycle of Rhodotrema quadrilobata Basicalova, 1932, intestinal parasite of flounder. Dokl. Akad. Nauk SSSR 1952, 83, 981–983. [Google Scholar] [PubMed]
- Chubrik, G.K. Fauna and ecology of larval trematodes in molluscs of Barents and White Seas. In Life Cycles of Parasitic Worms of North Seas; Poljansky, Y.I., Ed.; Nauka: Leningrad, Russia; Moscow, Russia, 1966; pp. 78–158. [Google Scholar]
- Meenakshi, M.; Madhavi, R.; Swarnakumari, V.G.M. The life-cycle of Helicometra gibsoni n. sp. (Digenea: Opecoelidae). Syst. Parasitol. 1993, 25, 63–72. [Google Scholar] [CrossRef]
- Dobrovolny, C.G. Life history of Plagioporus sinitsini Mueller and embryology of new cotylocercous cercariae (Trematoda). Trans. Am. Microsc. Soc. 1939, 58, 121–155. [Google Scholar] [CrossRef]
- Kakui, K. Digenean metacercariae parasitic in a staurozoan cnidarian. Zoolog. Sci. 2022, 39, 215–218. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokolov, S.G.; Shchenkov, S.V.; Frolov, E.V.; Gordeev, I.I. A Phylogenetic Re-Evaluation of the Stenakrine Opecoelids (Trematoda, Digenea: Opecoeloidea) with Some Taxonomic Novelties. Diversity 2022, 14, 949. https://doi.org/10.3390/d14110949
Sokolov SG, Shchenkov SV, Frolov EV, Gordeev II. A Phylogenetic Re-Evaluation of the Stenakrine Opecoelids (Trematoda, Digenea: Opecoeloidea) with Some Taxonomic Novelties. Diversity. 2022; 14(11):949. https://doi.org/10.3390/d14110949
Chicago/Turabian StyleSokolov, Sergey G., Sergei V. Shchenkov, Eugeniy V. Frolov, and Ilya I. Gordeev. 2022. "A Phylogenetic Re-Evaluation of the Stenakrine Opecoelids (Trematoda, Digenea: Opecoeloidea) with Some Taxonomic Novelties" Diversity 14, no. 11: 949. https://doi.org/10.3390/d14110949