

Reproductive Biology of Fritillaria aurea Schott (Liliaceae), a Rare Species Endemic to Turkey



1
Institute of Science, Erzincan Binali Yıldırım University, Erzincan 24002, Turkey
2
Erzincan Horticultural Research Institute, Republic of Turkey Ministry of Agriculture and Forestry, Erzincan 24060, Turkey
3
Department of Biology, Faculty of Science and Art, Erzincan Binali Yıldırım University, Erzincan 24002, Turkey
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(12), 1052; https://doi.org/10.3390/d14121052
Submission received: 9 November 2022
/
Revised: 27 November 2022
/
Accepted: 29 November 2022
/
Published: 1 December 2022
(This article belongs to the Special Issue Diversity and Conservation of Scrublands Flora and Vegetation)
Abstract
:Fritillaria aurea, unlike other Fritillaria species, is a narrowly endemic species. It has a brown checkered speckled color on each tepal and also has regular yellow intense colors on the tepals. Its large bell-shaped appearance when the flower is open is one of its most striking ornamental features. However, F. aurea plant numbers are limited in their natural habitats, and their numbers are decreasing day by day; they are even confronted with the risk of extinction owing to the damage resulting from human activities. This detailed investigation of reproductive biology plays a vital role in determining the evolutionary success and survival of F. aurea, largely in determining the effectiveness of their reproductive performance. Our results indicate that the species exhibits a very low partial dichogamy and herkogamy structure, as well as self-incompatible pollination. The flowering period ranges from March to April. Average seed viability, average seed number, and seed germination were determined as 71%, 255, and 45%, respectively. The pollen viability, pollen grains, and stigma receptivity were found as 96%, 392.000, and 85%, respectively. The pollen/ovule ratio (P/O) and self-incompatibility index (SII) in the spontaneous cross-pollination were detected as 1537 and 0, respectively. Flowers of F. aurea were visited by several insect species, mostly honeybees (Apis mellifera); Bombus sp. and Vespa sp. also played a minor role in pollination. To sum up, these results not only lay a solid foundation for further reproductive biology investigations to more broadly reveal the mechanisms of F. aurea endangerment in the future but also provide a reliable theoretical basis for hybridization breeding of parents.
1. Introduction
Global climate change, one of the hottest topics of the last decade, has been highlighted as the most important environmental problem affecting the survival and reproduction of many plant species, including seedless plants (Cryptogamae) and seed plants (Spermatophyta) [1,2,3]. Many recent studies have indeed discussed the issue of how plant reproductive mechanisms respond to climate change, focusing on characteristics such as mating systems [4], plant–pollinator interactions [5], and flowering phenology [6,7]. Therefore, access to knowledge of the reproductive traits of plants is vital both for estimating potential adaptation pathways and for formulating reasonable protection strategies that can cope with ongoing climate change. This goal, however, cannot be effectively achieved without a comprehensive knowledge of the ecology and biology of plants, and generally, when making attempts to protect the diversity of any plant in the environment in which it grows, we face the challenge of not only inattentive picking of these plant species to generate revenue but also damage by late spring frosts due to early flowering. On the other hand, little is known about the actual biological and/or ecological traits of bulbous plants, namely tulips, inverted tulips, and orchids, and this topic still remains a mystery among researchers.
Although it seems that habitat degradation, global climate change, and human factors are the main reasons causing the extinction of the species, the most basic issue is the reproductive failure of the species [8]. Creating a tentative global list of threatened plant species is the first step toward achieving the biodiversity goal (Goal 2 of the Global Strategy for Plant Conservation [GSPC]), but efforts so far have been insufficient to achieve this goal. Therefore, extinction risk assessments of plants urgently need to be expanded to guide conservation management and government policy [9]. Having said that, the loss of bulb plants, as with many plant species, could be detrimental not only to organisms but to the health of ecosystems and humans [10,11]. This information coupled with data affecting the reproduction of species growing under harsh conditions suggests that the investigation of the actual biological and/or ecological traits of bulb plant species has a vital role.
Fritillaria L. genus, a bulbous plant species, is a group of perennial herbaceous plants found in Liliaceae, with approximately 160 species in North America, Asia, China, Japan, and Mediterranean climate regions. It is mostly distributed in temperate areas of the northern hemisphere [12,13]. Approximately 45 species have been recorded in Turkey [12]. Dried onions of some species of Fritillaria are used in traditional Turkish, Chinese, and Japanese medicine for the treatment of various diseases, especially asthma. Their bulbs are also utilized as raw materials in traditional medicine for alleviating pain, reducing phlegm, bringing down fever, relieving coughs, and detoxification [14,15]. Because Fritillaria plants are slow growing, their annual growth and medicinal yield are low, and they usually require 4–5 years from seed germination to flowering [16]. In Turkey, however, Fritillaria germplasm resources, combined with habitat degradation and animal and human intervention, have led to overexploitation and serious endangerment of wild Fritillaria resources. Collection and destruction of this species are prohibited and are updated annually in Turkey’s national key protected wild plants.
Studies on the reproductive biology and/or ecological traits of this plant group have been limited, and little is known about it though Fritillaria is both the largest genus of the Liliaceae family, and most of its species are listed as protected [17]. The reproductive systems of Fritillaria species are diverse; some have a high ratio of outcrossing [17], others favor self-fertilization [18], and some have been reported to have a specific strategy, apomixis, according to experimental data [19]. As far as we know, studies on the reproductive biology of endemic Fritillaria aurea Schott belonging to the genus Fritillaria are limited. Basically, these studies appear to have been developed on a very weak experimental foundation, since their results are mostly based on anecdotal observations, and they are often one-day or short-term studies [17,18,19]. We sought, therefore, answers to the following questions by recognizing the inadequacy of detailed long-term observational studies of Fritillaria aurea. (I) What are the reproductive traits of F. aurea? (II) What is its pollination mechanism in the cultural environment? (III) How does its reproductive system affect its breeding success? (IV) What are the key pollinators of F. aurea flowers? (V) Are there any temporal changes in pollinator activities?
2. Materials and Methods
2.1. Study Site and Material
Fritillaria aurea Schott (Liliaceae) distribution from Central and Eastern Anatolia (Malatya, Kayseri, Sivas, Kahramanmaraş, İçel, Turkey) to Turkey grows on rocky and calcareous areas and Juniperus forest at an altitude of 1650–3000 m [20] (Figure 1a,b). However, the material of the study consists of individuals belonging to F. aurea collected from its natural areas in Malatya and Kahramanmaraş and preserved in the Erzincan Horticultural Research Institute, Turkey. The morphological characters (plant width and height; vegetation time and flowering; number, size, color, and odor of tepals; and color, number, and size of the leaf size) of this plant were previously investigated by Türktaş et al. [21]. This species normally has single flowers in its natural habitat, while in its cultural habitat it has 1 to 3 flowers. Its flowers are wide bell-shaped, with a characteristic brown tessellate pattern resembling a chessboard on yellow tepals [21,22]. In the phenological stages of this species, where nectars are found, when flowers begin to secrete nectar and how long this process lasts, and how this stage corresponds to the anthers and pistil fertility stages, were considered in accordance with previous authors’ reports [17,22]. The flowering time of the plant varies from 20 March to 10 April. The flowering time of the plant is about 6–12 days, and its height is approximately 15–40 cm. The average values of leaf length, leaf width, and bulb diameter of F. aurea were detected as 64.0 ± 0.6, 14.0 ± 0.9, and 24.7 ± 0.5, respectively. The outer tepal length, outer tepal width, inner tepal length, and inner tepal width of F. aurea were determined as 34.0 ± 1.3, 15.0 ± 0.8, 38.5 ± 1.6, and 20.8 ± 1.3 mm, respectively (unpublished data). A soil containing a mixture of forest soil, peat, sand, and pumice mixture (at a ratio of 8/4/2/1, respectively) was prepared for the plants, and after this mortar was placed in pots and pans, the plants were planted in separate pots and pans. After labels were written on each pot, these plants were placed in the pans and pots, and routine cultural care was performed. Each of the plants used in the experiment had homogeneous characteristics. Bulbs collected from natural conditions were taken under protection by taking each one in a flower pot. Pans were created from the seeds (approximately 16 years) obtained from young onions and bulbs obtained from the whole plant collected from nature over the years. The area where the plants were located was surrounded by wire fences and was a sheltered area. The bulbs of these plants were collected with special permission and their natural conditions were determined. To provide this environment, the plants were grown under semi-controlled conditions. Shading was conducted between June and August to protect from high temperatures. Irrigation was carried out with drip irrigation using tap water in March, April, and October to the end of the year. The soil properties of the plant in the natural environment are as follows: pH: 6.90; lime: 0.97; organic matter: 1.01%; p = 3.2 kg/da; K: 72 kg/da.
2.2. Pollination Experiments
In both 2019 and 2020, we randomly marked 220 flowering plants at the closed bud stage for the pollination experiment of the studied population and designated them into five applications: (1) open-pollination or self-pollinated (control) (n = 50), (2) spontaneous self-pollination (bagged with a fine 1 mm2 mesh to prevent insects from visiting the flowers) (n = 50), (3) spontaneous cross-pollination (open-pollinated, flowers emasculated in buds) (n = 40), (4) apomixis: induced cross-pollination (isolated from further insect visits and supplemental pollination, flowers emasculated as above) (n = 30), and (5) induced self-pollination (the isolated flowers were hand-pollinated with self-pollen) (n = 50). We utilized either a flower from a plant that was growing at a distance of at least 3–4 m (cross-pollination) or (cross-pollen) or from one dehisced anther of the same individual flower (self-pollination), for hand pollination based on Dafni et al. [23]. The flowers were unbegged, about 15–20 days after flowering. The fruit number was counted after a further 5 weeks, when the capsules had started to ripen. The length of the fruit brought to the laboratory was determined, and the number of developed seeds in each capsule was determined. Seeds of plants were detected on the basis of size and morphology. Before this step, seeds were prepared as hand sections of 50 seeds of various sizes. Then, their endosperm and embryo development stages were examined under a light microscope. The length of the seeds obtained from the plants ranged from ~5.0 to 6.0 mm.
2.3. Flowering Dynamics
Regarding our observations, the duration of flowering of the studied population was approximately 2 weeks. We observed the flowering dynamics and flower traits of F. aurea, (from about 15 March to April each year) and classified the progress of flowering into eight stages: a, bud stage (the first stage before bud burst); b, initial flowering (stem and flower development after bud burst); c, pre-dehiscence stage, (beginning of coloration in perianths or pollen presentation and beginning of coloration and development of the anthers in the capsule); d, first dehiscence stage (i.e., about one anther dehisced, beginning of pollen presentation); e, full dehiscence, (all anthers dehisced, pollen presentation phase); f, end-dehiscence stage (anthers empty and end of pollen presentation); g, fresh perianth stage (stamens and pistils continued to develop, the perianths remained fresh); and h, perianth wilting stage (the dehydrated perianth withers, and the stigma becomes dry and brown).
2.4. Stigma Receptivity and Pollen Viability
We randomly selected three flowers of F. aurea at each of the six developmental stages (a, b, c, d, e, f, and g stage), and stigma receptivity was determined using the benzidine-hydrogen peroxide reaction solution method [23]. The benzidine-hydrogen peroxide reaction solution for stigma receptivity was prepared at a ratio of water: 3% hydrogen peroxide: 1% benzidine (23:11:4). Selected stigmas were placed on a concave glass slide, an appropriate amount of benzidine-hydrogen peroxide solution was dripped onto it, and the time of bubble formation on the stigma was recorded. Bubbling and discoloration around the stigma were observed and recorded after 1 min. Pollen of F. aurea was taken, and the presence of protoplast and the cellular condition of a pollen grain were determined in the laboratory environment using the Acetocarmine method. Stigma receptivity was determined based on bubble densities around the stigma by slight modifications to the method of Dafni and Maués [23], as follows: (1) stigma receptivity strong (if the bubbles are dense or bubble formation at approximately 9/10 of the stigma surface), (2) stigma receptivity middle (if bubbles are moderate or bubble formation at approximately 5/10 of the stigma surface), (3) stigma receptivity poor (if bubble density is very low or bubble formation at approximately 1/10 of the stigma surface).
We randomly selected ten flowers of F. aurea at the pre-dehiscence stage for pollen viability. Pollen viability was detected using the acetocarmine staining method [24]. In this study, anthers taken from flowers (at different developmental stages) were examined under the microscope as fresh pollen on a slide without wasting time (i.e., no drying in the anthers). In the study, 1% acetocarmine was dripped onto the anther sacs left on the slide, allowing the pollen to be dispersed; then, the anther sacs were removed from the slide. After spreading the pollen grains into acetocarmine, it was covered with a coverslip, and the prepared preparation was left on a heater at 50 °C for 1 min to allow the dye to be diffused by the pollen grains. Then, color changes in the pollen were observed with a 10× objective under a light microscope (Diaphot 300; NIKON Tokyo, Japan). Dark reds were considered viable pollen, light reds were considered semi-viable pollen, and undyed ones were considered as not viable pollen [24]. Data on viability pollen tube formation are presented in Supplementary Material Figure S1.
2.5. Estimation of Pollen/Ovule Ratio (P/O) and Self-Incompatibility Index (SII)
Ten different plants grown in pans were randomly marked, and an anther was taken from each of these plants during the pre-dehiscence. The anther was then blasted on a slide with a counting scale (10 × 10 mm), and pollen grains were counted by observing with a stereomicroscope. At the beginning of flowering, randomly marked ovules were taken from ten different plants grown in pans; then, they were divided with a scalpel on the slide, and young ovules were removed in groups. The young ovules obtained from the plants were examined under a stereomicroscope and counted. The number of pollen grains produced for each ovule in a flower was based on Cruden [25]. For the self-incompatibility index (SII), the seed setting success of spontaneous self-pollination and spontaneous cross-pollination plants after flowering was recorded, and considering self-pollination/cross-pollination, the SII value was determined according to the method of Zapata and Arroyo [25].
2.6. Seed Viability and Germination
We randomly marked ten fruits of F. aurea after about 30 days from the perianth wilting stage, and their seed viability was detected by the 2,3,5-triphenyl tetrazolium chloride test (TTC) [26]. Two hundred seeds from different fruits were placed in the prepared TTC solution. These seeds were then immersed in distilled water for 24 h to activate the embryo, and the soaked seeds were left in a 0.1% solution at room temperature for 24 h to expose the embryo. Embryos were observed in halved seeds after sufficient incubation time. In the seeds examined under the stereomicroscope, embryos with red color were considered as viable seeds, and those without any color change in embryos were considered as dead seeds. Additionally, seed germination experiments were carried out based on [27].
2.7. Pollinator Behavior Observation
Insects and their behavior were photographed visiting the flowers for ten consecutive days during full flowering on sunny days in 2019–2020. The wasp and honeybee were visited the flower and were photographed; thus, we could determine exactly when they entered the flower. Each observation day, insect visits (consisting of six rounds of insect capture) were recorded (each hour, from 09:00 a.m. to 15:00 p.m.; the recording was not performed if some preliminary work observations conducted in the early morning and at sunset showed no insect activity. One record round lasted about 1 h and occurred in three stages: (a) random selection of flowers based on insect visitation, (b) video recording according to insect activities (in a 30 min digital video with a Canon 5D Mark IV camera), and (c) insect capture. One or two insects of the same species arriving on the flowers for insect identification were collected using a trap. Data about pollen on the surface of the insect body is presented in Supplementary Figure S2.
2.8. Statistical Analysis
Analysis of variance was conducted using JMP statistical software (version. 7.0, SAS Institute Inc., Cary, NC, USA). Student’s t-test was used for open-pollination and spontaneous cross-pollination. However, Tukey’s HSD test (p ≤ 0.05) was used in the comparisons of pollen viability and stigma receptivity between different flowerings stages. Regression analysis was conducted to determine relationships between pollen viability and stigma receptivity in the different flowering stages of F. aurea. We generally used linear curves for the calibration equations (regression model) created from the data obtained in the different flowering stages of F. aurea.
2.9. Collection Permits of Plant Materials
Necessary collection permits were obtained within the scope of the project TUBITAK (The Scientific and Technological Research Council of Turkey).
3. Results
3.1. Pollination Experiments
This study examined five types of pollination on F. aurea, and it was detected that the rate of fruit set varied between them: open-pollination (68.0 ± 8.3) > spontaneous cross-pollination (63.0 ± 9.4) > spontaneous self-pollination (0) = apomixis (0) = hand self-pollination (0) (Table 1). The length of the seeds obtained from the plants ranged from ~5.0 to 6.0 mm. Regarding our results, the fruit setting rate of open-pollination was only slightly higher (%5) than that of spontaneous cross-pollination, but there were no significant differences between them. Unfortunately, no fruit set was observed in spontaneous self-pollination, apomixis, and hand self-pollination treatments. The fruit setting rate of F. aurea was in the natural pollination treatment higher than that in the bagging without emasculation treatment (Table 1).
3.2. Flowering Dynamics
Although F. aurea stems usually have a single flower on the top, sometimes two to three flowers may occur (Figure 2).
In this study, however, single flowers were preferred as the experimental material. Flowers of the plant opened a mean of 7 days, and pollen presentation of flowers lasted for 4.0–5.0 days. There also was no difference in flowering time for both years. The flower buds of the plant were initially green, and they were partially pendulous, in the shape of a long tube. Later, the tepal color gradually began to change as the pendulous flower opened, and it turned into a characteristic brown tessellate pattern resembling a chessboard on yellow tepals. The flowers were hermaphrodite, having an uncurled leaf-shaped bract at the apex. The six perianth pieces of flowers consisted of three outside ones and three inside tepals, with tiny brown checkered spots on each tepal and regular yellow intense color on the tepals. The flowers, which had a glabrous base, had a wide bell-shaped appearance when opened. The plant had an anther structure that released intense yellow pollen grains through longitudinal dehiscence. Each anther within a single flower had a different dehiscence time, and usually, the shorter anthers opened before the others. It had a single style of three carpellate, covered with mucus on the surface and terminated in a yellow trifid stigma. The style was undivided, with three lobes at the end, three capsules longitudinally, and numerous seed germs, arranged in two rows in each segment. A single flower had six separated stamens surrounding the pistil. They had herkogamy structures owing to the shortening of the anther length after anther dehiscence. There was a vertical distance of ~0.5 to 1 cm between the anthers and the stigma. The period from the bud to the unopened flower bud in F. aurea lasted 10–25 days. The brown tessellate pattern resembling a chessboard increased along with the perianth segments when the flower bud period continued. The flowering process of the plant lasted 12–25 days, as follows: (1) the bud stage on soil (1–2 days), (2) the initial flowering time (5–7 days), (3) pre-dehiscence perianth coloration (2–3 days), (4) full dehiscence after perianth coloration (2–3 days), (5) end-dehiscence of anthers (4–5 days), (6) the fresh perianth of flowers (5–7 days), (7) the perianth wilting time (3–4 days), and (8) the emergence of seed capsule after the dried perianth (1 day) (Figure 3).
3.3. Stigma Receptivity and Pollen Viability
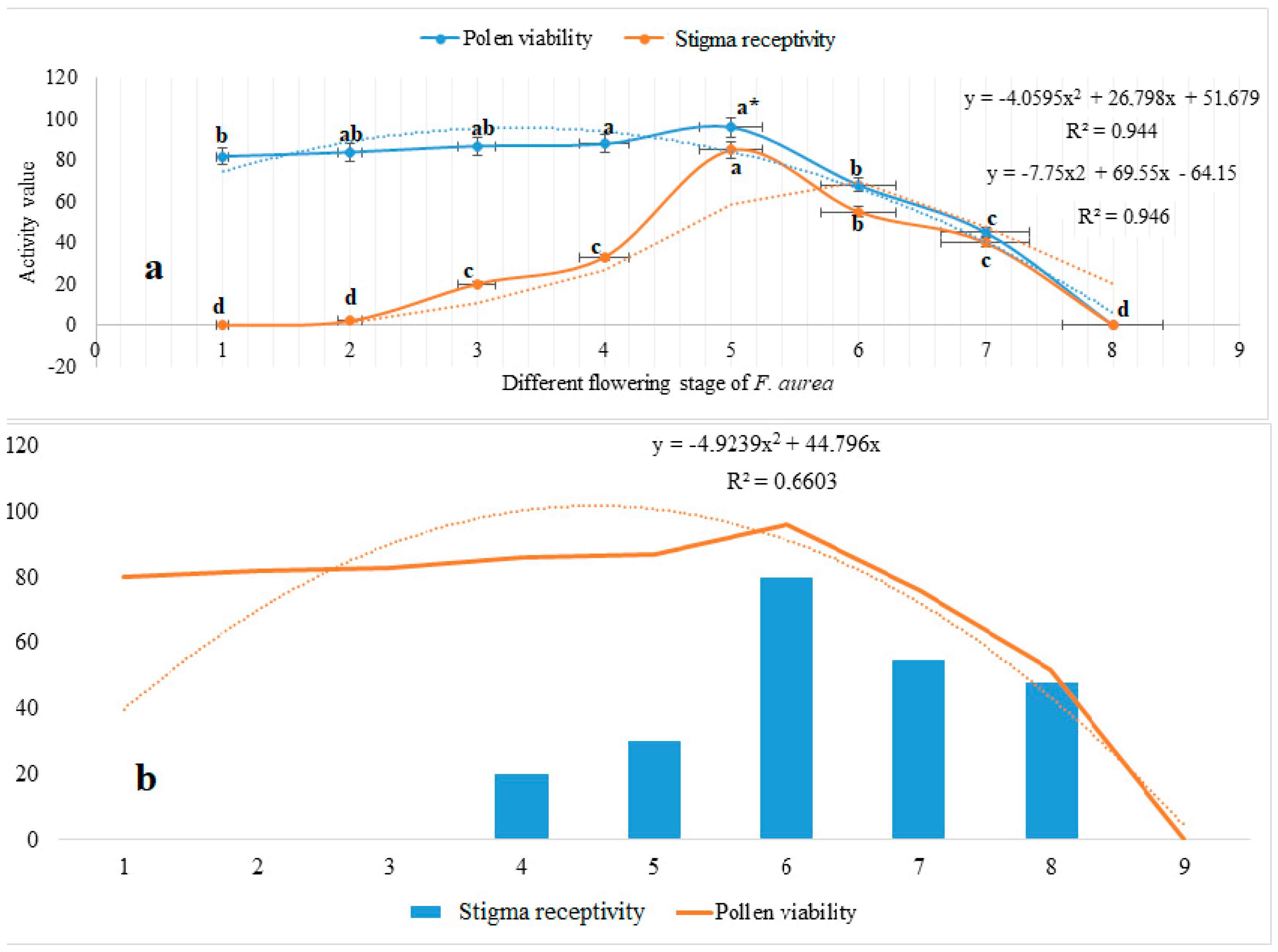
Stigma receptivity of F. aurea was lowest at the bud, initial flowering, pre-dehiscence, and perianth wilting stages (0), highest at the end-dehiscence stage (85.0%), and declined significantly during fresh perianth (55 ± 0.2) and perianth wilting stage (0). At the bud, initial flowering, pre-dehiscence, and perianth wilting stages, the stigma not showed bubbles and was not stained, indicating that it was not receptive at these stages. There also was no significant difference among these stages; however, significant differences were found among first dehiscence, full dehiscence, end-dehiscence, and the fresh perianth stages. Additionally, the results showed a positive significant correlation (p ≤ 0.05) between stigma receptivity and different flowering stages of F. aurea, with a correlation coefficient of r = 0.944*. High values of stigma receptivity of F. aurea overlapped at stages (see Figure 3d–g and Figure 4a). Pollen viability showed viability at different rates in terms of stages of flowering. Pollen viability gradually increased during flower development stages and reached a maximum level of 96% ± 3.25 at full dehiscence, decreasing to 0 at the perianth wilting stage. The pollen number and viability gradually decreased after full dehiscence. The results showed a positive significant correlation (p ≤ 0.05) between pollen viability and different flowering stages of F. aurea, determined as r = 0.946*. High values of pollen viability of F. aurea overlapped at stages (see Figure 3a–g and Figure 4a,b).
3.4. Estimation of Pollen/Ovule Ratio (P/O) and Self-Incompatibility Index (SII)
The number of pollen grains in F. aurea varied from 354.000 to 402.000 per flower, with an average value of 392.000, whereas the number of ovules varied between 240 and 270, with an average value of 255. The P/O ratio was found to be 1537 on average. On the other hand, self-pollination did not occur in the pollination treatments of this species, and thus the percentage was not established. Based on Zapata and Arroyo [25], the self-incompatibility index (SII) of F. aurea is 0.
3.5. Seed Viability and Germination
The seed viability was detected only in the spontaneous cross-pollination treatment because the fruit set was not observed in spontaneous self-pollination, apomixis, and hand self-pollination treatments.
Based on the results of the tetrazolium staining test, viable and dead ratios of seeds in the spontaneous cross-pollination treatment were 71% and 29%, respectively, whereas viable and dead ratios of seeds in open-pollination were 67% and 33%, respectively (Table 1). There were no statistical differences between the open-pollination and the spontaneous cross-pollination treatment. In our findings, there was also no significant difference between seed germination in spontaneous cross-pollination and open pollination, and these values were 40 and 45%, respectively.
3.6. Pollinator Behavior Observation
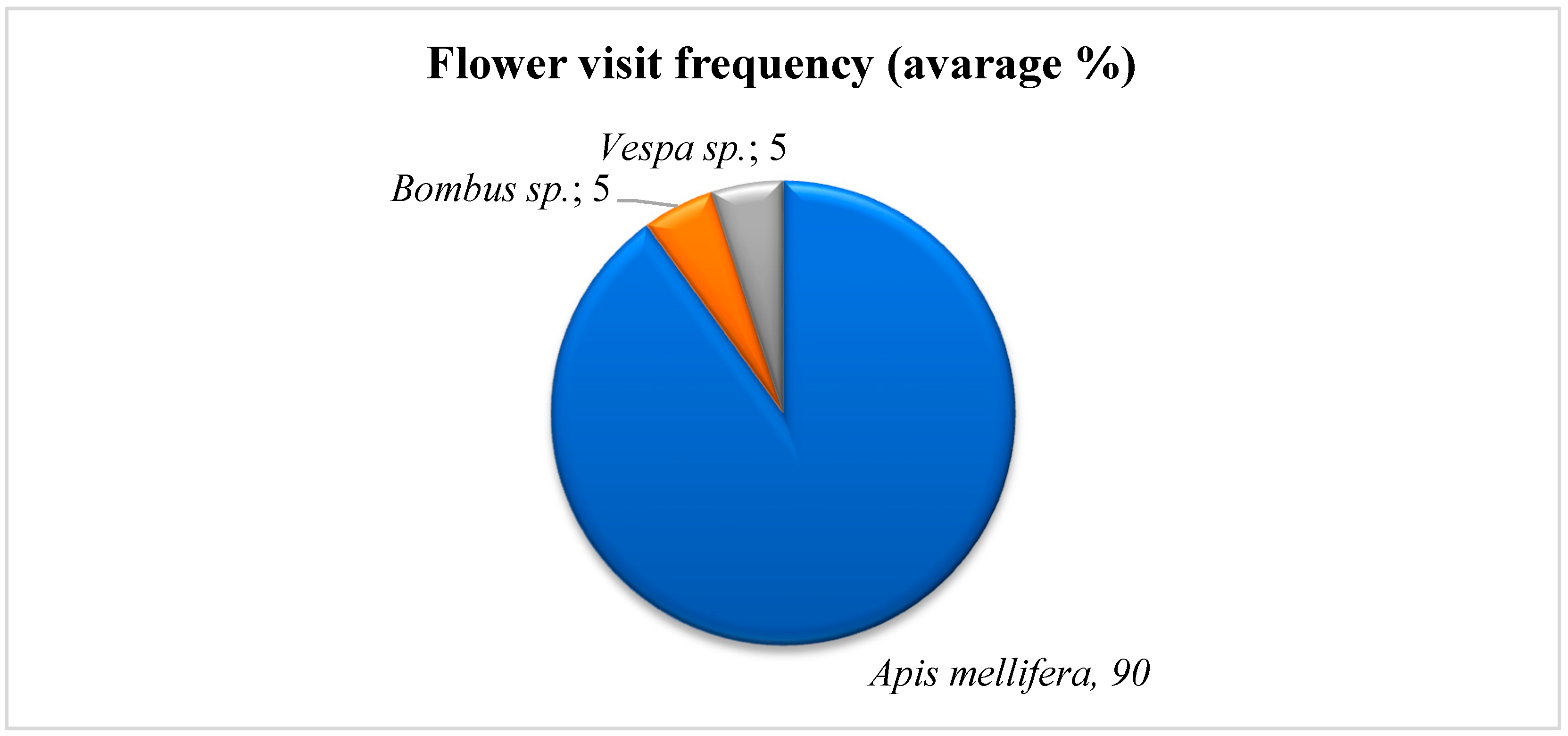
In the present study, three insect species were recorded visiting F. aurea flowers. They belonged to two taxonomic orders: Hymenoptera (Apidae: Apis mellifera L., Bombus sp., Vespa sp.) and Diptera (Musca sp.). Insect visitors were defined as pollinators who constantly came into contact with both stigmas and anthers and deposited pollen grains on both their bodies and feet [28]. A total of three Hymenoptera flower visitors were detected during the entire observation period: one honeybee (Apis mellifera) and two wasps (Bombus sp. and Vespa sp.). Honeybees (Apis mellifera), Bombus sp., and Vespa sp. were observed in 2019–2020 and, when present, performed 89–90%, 4–5%, and 4–5% of visits, respectively (Figure 5).
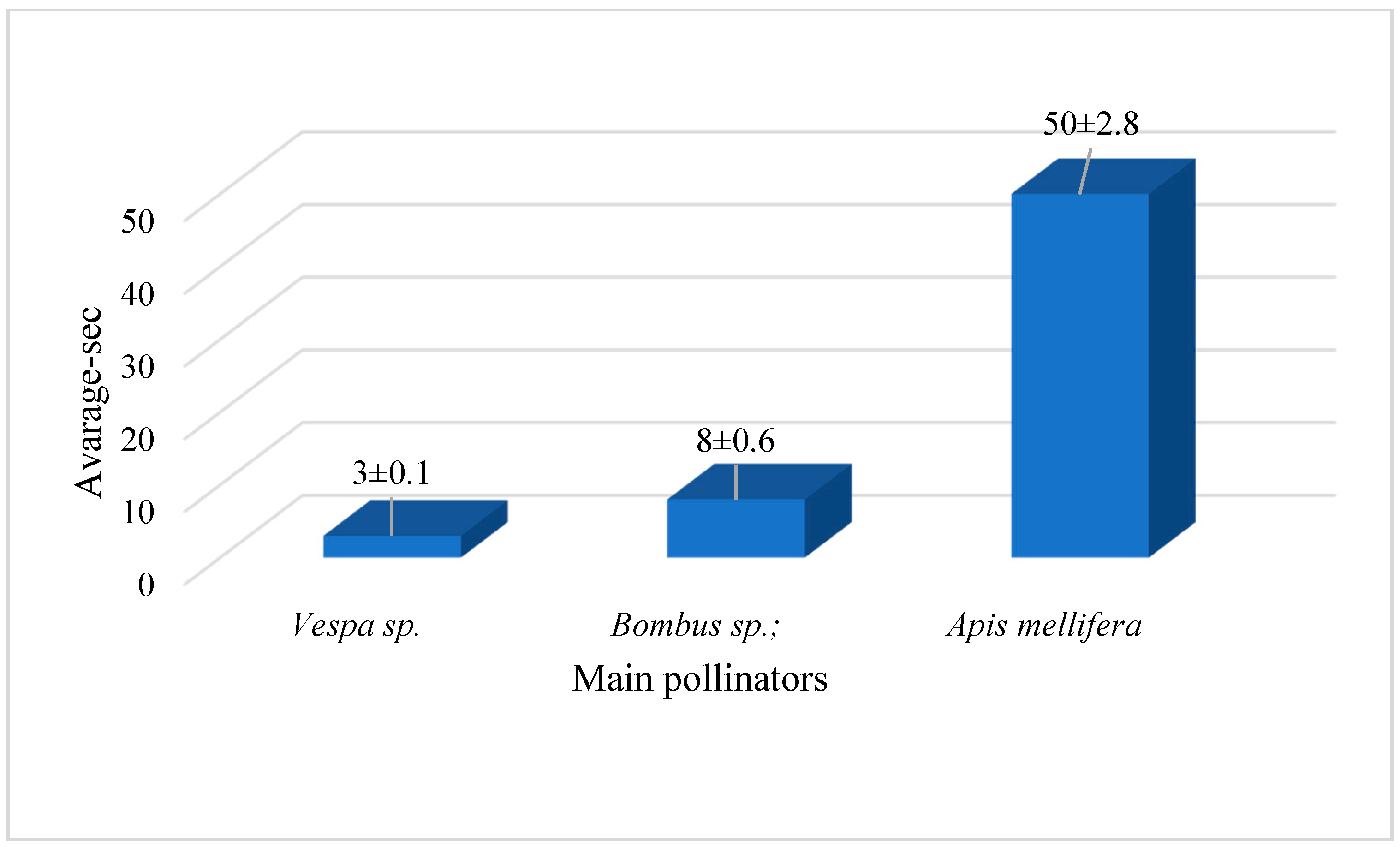
The Vespa sp. visited four or five Fritillaria flowers, with each visit lasting 5–20 s; Bombus sp. visited nine or ten Fritillaria flowers, with each visit lasting 3–4 s; Apis mellifera L. visited 40–50 fritillary flowers, with each visit lasting 10–60 s (Figure 6). The wasp and honeybee were visiting the flower when we photographed; thus, we knew exactly when they entered the flower, and it was clear which bee had the longer visit time. The Vespa sp. hovered outside the perianth when visiting flowers after climbing into the flowers of the plant along with the tepals during our field observations.
4. Discussion
4.1. Pollination Experiments
Basic information on reproductive biology is not only important for effective conservation strategies [29] but also essential for systematic and evolutionary studies [30] for threatened and endemic plant species with very few populations like F. aurea. This study reports on the reproductive biology of F. aurea, which is an endemic species in Turkey. This study performed five types of pollination on F. aurea, but fruit set was not detected in spontaneous self-pollination, apomixis, and hand self-pollination treatments; fruit setting only occurred in open-pollination and spontaneous cross-pollination treatments (Table 1). The results of these three pollination treatments without fruit setting highlighted that there was pollination limitation in F. aurea, and its reproductive strategy was only obligate outcrossing. Both herkogamy and dichogamy did not occur in the flower’s life cycle. However, F. aurea is a self-incompatible plant, with comparatively low reproductive output. The pistils and stamens of F. aurea have a degree of dichogamy, but in this process, although the pollen viability remains higher than 50%, the pollen viability decreases when the stigma receptivity is the strongest. Although both the stamen and pistil were not in a position to visibly promote herkogamy in the flower, and a transient overlap period existed between male and female fertility, seed-setting in the flower did not occur. This is likely due to the predominant self-incompatibility of F. aurea, which is directly related to selective pressure to maintain genetic variability. It has been reported that the degree of allogamy and duration of pollen viability may vary greatly within a single plant species depending on various factors, including duration of pollen viability, degree of dichogamy, and pollinator abundance [31]. It should be pointed out that insect vectors are needed to promote cross-pollination in F. aurea flowers. In our study, the seed set did not occur in self- or intra-morph crosses (self, geito, or intra), demonstrating that F. aurea is an obligately outcrossing species. Thus, these findings suggest that F. aurea could be considered as a functionally heterostylous plant species characterized by a self-incompatibility system. It is assumed to have evolved as a way of avoiding inbreeding to ensure efficient pollen exchange in inappropriate mating due to this reproductive trait, which is consistent with flower morphology [32,33]. Hand self-pollination treatments (63%) did not increase fruit set compared to open pollination (68%), and there was no significant difference between them (Table 1), suggesting that no pollen limitation occurred in F. aurea pot and pan populations, which is in contrast to the strong pollen limitation found in other Fritillaria species, such as F. meleagris L., F. maximowiczii Freyn and F. delavayi Franch [12,34,35]. On the other hand, it is interesting to investigate why F. aurea did evolve self-incompatibility systems as present in other closely related species such as F. camtschatcensis L. [36], as such systems could be advantageous in long-lived plants and this kind of perennial. The likely answer is that self-incompatibility systems appear to evolve automatically in F. aurea and, as shown in models of self-incompatibility evolution in some species, for the spread of the first self-incompatibility allele, a level of inbreeding depression >0.5 is obligatory [37]. This criterion is apparently met in F. aurea, because there was no spontaneous self-pollination fruit setting.
4.2. Floral Traits, Stigma Receptivity, and Pollen Viability
Flower color on F. aurea with brown checkered spots on each tepal and regular yellow intense color on the tepals is one of the most remarkable ornamental traits. F. imperialis L. and F. persica L. species have the showiest and most attractive flowers among Fritillaria species in Turkey, followed by F. aurea. Breeding studies were carried out by us (with about 15 years of breeding) [38,39], taking into account both the showy and attractive flower characteristics of this species, and a new variety was developed from F. aurea. As a result of this breeding, the cultivar named “Eastern Sun” (Registration number 2014/104) was registered [40] as Turkey’s first inverted tulip variety. As far as we know, however, it is difficult to compare it with studies in the literature, since no research was found on the floral features of this plant. F. aurea is an interesting plant species, and may be placed in the most fundamental position on the Fritillaria phylogenetic tree given the genomic data [13,41], perhaps implying that it has primitive morphological characteristics. On the other hand, F. aurea flowers opened for 7 days (from about 15 March to 15 April each year), and pollen presentation of flowers lasted for 4.0–5.0 days, which is in agreement with data on Fritillaria species in previous studies [10,34,35]. Our observations indicated that, contrary to the periant shapes of many Fritillaria species in the literature [10,34,42], the species has a wide bell-shaped appearance when opened. The flowering process of the plant lasted 12–25 days, and this difference in time from bud to unopened flower bud was due to temperature fluctuations in March and April. There was slight asynchrony and partial similarities among the previously studied Fritillaria species during the flowering to fruiting period. Such similarities and asynchrony among Fritillaria species may be due to microhabitat and climatic differences, as reported by the literature [10,34,35,43].
The stigma receptivity and pollen viability play a key role in the successful initiation of pollen pistil interaction [44]. The presence of certain enzymes on pistils, namely esterases, peroxidases, and acid phosphatases, indicates the receptivity of stigmas [23]. Improved stigma receptivity on the pistil plays a vital role in the pollination, fertilization, and reproductive success of plants and assures fruit set [45]. In our results, stigma receptivity of F. aurea was highest at the end-dehiscence stage and declined significantly during the fresh perianth. Our observations show a positive significant correlation (p ≤ 0.05) between stigma receptivity and different flowering stages of F. aurea, determined as r = 0.944*. High values of stigma receptivity of F. aurea overlapped at stages (Figure 3d–g and Figure 4a,b), demonstrating that this species maintains a high level of sexual fertility during these stages. Pollen viability showed viability at different rates, depending on stages of flowering. Pollen viability gradually increased during flower development stages and reached a maximum level at full dehiscence. The viability and number of pollen grains were high, and their adhesion was strong during the first six periods. These stages were vital periods for the collection of pollen. The pollen number and viability gradually decreased after full dehiscence. The results showed a positive significant correlation (p ≤ 0.05) between pollen viability and different flowering stages of F. aurea. High values of pollen viability of F. aurea overlapped at stages a–g (Figure 3a–g and Figure 4a,b), demonstrating that this species maintains a high level of sexual fertility during these stages. However, the pollen viability test and pollen load on the stigmatic surface showed clear-cut evidence that F. aurea is a self-incompatible species under pot and pan conditions. Though the number of pollen grains and viability is greater due to various factors that we do not know in detail, self-incompatibility was observed. We hypothesize that due to the inverted structure of the perianth, pollen from the burst anthers did not fully reach the stigma, and therefore fertilization did not occur. It could, however, be assumed that there may be different factors that we cannot explain in this study. Recent studies have indeed reported that various factors, including season, temperature, genetic load anther length, stigma receptivity, pollen quantity and quality, pollen grain size, shape, and anther dehiscence mode, are important in whether a flower will be pollinated or not [17,46,47,48,49,50,51].
4.3. Estimation of Pollen/Ovule Ratio (P/O) and Self-Incompatibility Index (SII)
Our estimates of the self-incompatibility index (SII), pollen/ovule ratio (P/O), and natural population pollination experiments indicated that the breeding system of F. aurea included a high level of self-incompatibility, meaning that it has a type of self-incompatible breeding system, and wind or insect vectors are required for the pollination of flowers. Therefore, its breeding system was detected to be facultative xenogamy, according to Cruden [25]. First, it was determined that the species is not a self-compatible species in any way in the pollination experiments conducted. We determined the pollination preference of the species using two different methods, which are commonly used in these experiments [25,26]. While the SII method values supported our results, the P/O method showed some difference. In other words, if zero is divided by a number in the SII method, the value becomes zero. This supports our results, but the contradiction is due to the P/O method. This method was used by Cruden [25] on a certain number of plants, and a generalization occurred. When this method was applied to the data we obtained, our results showed that the species was in the facultative xenogam range, based on the Cruden [25] method. In addition, we believe that F. aurea may not perform self-fertilization based on the assumption that the perianth grow structurally inverted and the pollen cannot reach the stigma in an upside-down flower. It has generally been reported that the whole Liliaceae family, including Fritillaria, is mostly out-crossing [52]. However, F. aurea has remarkable self-incompatibility, ensuring no sexual reproduction in the absence of pollinators. These results were generally inconsistent with previous reports of fritillary species where self-pollination is possible, such as F. koidzumiana Ohwi [53], F. meleagris [10], and F. persica [54]. In this study, however, specific studies were not conducted to examine which factors are more important in determining self-incompatibility in F. aurea. Considering the Zapata and Arroyo [25] index, the self-incompatibility index (SII) of F. aurea was 0, meaning that it has a type of self-incompatible breeding system. This is also evident from the pollination treatments, as 63% of the spontaneous cross-pollinated flowers set fruits and seeds. Some authors strongly suggest that low fruit setting in open-pollinated flowers compared to cross-pollinated flowers requires some external agents, such as insects and wind, for effective pollination in many plant species [55].
4.4. Seed Viability and Germination
A better understanding of the germination fraction and seed viability dynamics of F. aurea may allow us to better model both demographic dynamics and effective population sizes. Regarding the results of the tetrazolium staining test, seed viability in the spontaneous cross-pollination and open-pollination was 71% and 67%, respectively (Table 1). The lack of any differences or clear trends in seed viability levels for spontaneous cross-pollination and open-pollination suggests that self-incompatibility and pollen coming from any distance sufficient to reduce the chances of mating the same plant species as themselves is not equally good. On the other hand, seed germination in spontaneous cross-pollination and open pollinations was 40 and 45%, respectively, and this value was quite low compared to seed viability. Perhaps this species may also be displaying a strategy where not all seeds germinate at once, or these seeds were not as viable as the initial germinants or they did not germinate due to unknown reasons. It has been reported that seed germination is a complex process [56]. From a quiescent state of very low metabolic activity during seed germination, radicle and imbibition emergence proceed via a series of biochemical and physiological events that include processes such as enzyme and organelle activity, hydration of enzymes and other colloids, respiration (citric acid cycle, pentose phosphate pathway, glycolysis), subcellular structural changes, elongation of radical cells, and reserve mobilization and biosynthesis of macromolecules (mRNA, enzymes, hormones) [57]. The deterioration caused by the disruptions in any of these stages may have affected the germination success.
4.5. Pollinator Behavior Observation
Reproductive biology in plants has mainly focused on flowering phenology, breeding systems, pollen–pollinator interaction, floral biology, and gene flow through pollen and seeds [51]. The flowers were visited by Hymenoptera (Apidae: Apis mellifera L., Bombus sp., Vespa sp.) and Diptera (Musca sp.) in our study. Insects visited the flowers at certain times during the day after anthesis, and all these visitors played key a role in spontaneous cross-pollination. Our observations showed that honeybees (Apis mellifera) are effective pollinators for F. aurea flowers compared to other pollinators. This explained that bumblebees preferred nectar to pollen. Considering the nectar was located in the wider space inside the bell-shaped sepals and at the base of the tepals, it took less effort for both bumblebees and honeybees to get into the corollas of F. aurea. Unlike the three mentioned pollinators, we hypothesized that F. aurea might also have had some nectar robbers, namely Diptera (Musca sp.). These insects may have fed on nectar by biting from the outside of the base of the perianth where the nectar is located. Self-incompatibility for F. aurea confirms that there is a pollination deficit in this species, and higher effectiveness of the pollinators is mainly attributed to the lack of pollinators. Widely accepted findings to explain the evolution of pollination by deception in the Liliaceae suggest the fidelity of bumblebees and honeybees to Fritillaria flowers, as observed by some authors during analyses of insect pollen loads [10,58,59,60]. There also was a difference between both the frequency and duration of the insects visiting the flower; generally, insects do not have problems foraging in pendulous flowers of this fritillary. Our findings are consistent with the results that most Lasioglossum and Andrena individuals spend more time in flowers compared to bumblebees [10]. Honeybees (Apis mellifera) tend to move more chaotically within the flowers of F. aurea, sometimes leaving the stigma completely covered with pollen, and thus possibly conspecific stigmas deliver more self-pollen. These data also highlight the partial understanding of the processes underlying dependent shifts between pollination syndromes of F. aurea. However, contrary to our findings observed within the species F. aurea, it has been reported that Fritillaria flowers such as F. maximowiczii and F. meleagris were pollinated by bumblebees, spiders, and bumblebee queens [10,34,35]. Although it is difficult to explain with our current knowledge why honeybees visit more flowers of F. aurea than other bees, we hypothesize that it is because the number of other pollinators in the study area is less than the population of honeybees. Since they flower earlier than many other plants in the study area, honeybees could have preferred the flower of F. aurea for their food supply.
5. Conclusions
This study provides a detailed account of various aspects of the reproductive biology of F. aurea, such as floral biology, phenology, floral visitors, and pollen/ovule (P/O) ratio. It also reports here for the first time comprehensive research of the pollen and seed viability, pollen and seed number, and stigma receptivity of F. aurea, which is an endemic species in Turkey. Flower color of F. aurea, with brown checkered spots on each tepal and regular yellow intense color on the tepals and a wide bell-shaped appearance when opened, is one of the most remarkable ornamental traits, unlike other Fritillaria species. Our results with regard to pollen/ovule ratio (P/O), self-incompatibility index (SII), and natural population pollination treatments indicate that the breeding system of F. aurea includes a high level of self-incompatibility. All the F. aurea plants investigated exhibited high levels of pollinator dependence, and our observations showed that honeybees (Apis mellifera) are effective pollinators for F. aurea flowers compared to other pollinators, namely Bombus sp. and Vespa sp. Detailed investigation of the reproductive biology of the Fritillaria should also be considered in any restoration attempts, as they could decide the success or failure of the work by conservation biologists and may pose a great challenge, though this area receives little attention in research on restoration.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14121052/s1, Figure S1. Represents images of pollen and seed studies. Figure S2. Image of pollen of the species on the surface of honey bees (Apis mellifera) body (Photo Original; F. Yıldız).
Author Contributions
F.Y. and M.A. designed this study and performed the experiments. F.Y. and O.K. analyzed the data. O.K. wrote the manuscript. M.A., O.K., F.Y. and A.K. supervised the experiment and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported both by TUBITAK (The Scientific and Technological Research Council of Turkey), project No. 106G022 and 110G007, and by the General Directorate of Agricultural Research and Policies, Scientific Research Project-SRP (Project No. TAGEM/bbad/17/a09/p09/05) appropriated to Erzincan Horticultural Research Institute, Turkey.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data can be found in the article and Supplementary.
Acknowledgments
We express our deepest gratitude to the anonymous reviewers for their thoughtful suggestions and careful work that greatly assisted in the development of the manuscript. The TUBITAK (The Scientific and Technological Research Council of Turkey) and Erzincan Horticultural Research Institute, Republic of Turkey Ministry of Agriculture and Forestry is gratefully acknowledged for their support during our field work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giejsztowt, J.; Classen, A.T.; Deslippe, J.R. Climate change and invasion may synergistically affect native plant reproduction. Ecology 2020, 101, e02913. [Google Scholar] [CrossRef] [PubMed]
- Kaya, O.; Kose, C.; Gecim, T. An exothermic process involved in the late spring frost injury to flower buds of some apricot cultivars (Prunus armenica L.). Sci. Hortic. 2018, 241, 322–328. [Google Scholar] [CrossRef]
- Kaya, O.; Kose, C.; Donderalp, V.; Gecim, T.; Taskın, S. Last updates on cell death point, bud death time and exothermic characteristics of flower buds for deciduous fruit species by using differential thermal analysis. Sci. Hortic. 2020, 270, 109403. [Google Scholar] [CrossRef]
- Razanajatovo, M.; Fischer, L.; van Kleunen, M. Do floral traits and the selfing capacity of Mimulus guttatus plastically respond to experimental temperature changes? Oecologia 2020, 192, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Tsiftsis, S.; Djordjević, V. Modelling sexually deceptive orchid species distributions under future climates: The importance of plant pollinator interactions. Sci. Rep. 2020, 10, 10623. [Google Scholar] [CrossRef]
- Cooper, E.J.; Dullinger, S.; Semenchuk, P. Late snowmelt delays plant development and results in lower reproductive success in the High Arctic. Plant Sci. 2011, 180, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Humphreys, A.M.; Govaerts, R.; Ficinski, S.Z.; Nic Lughadha, E.; Vorontsova, M.S. Global dataset shows geography and life form predict modern plant extinction and rediscovery. Nat. Ecol. Evol. 2019, 3, 1043–1047. [Google Scholar] [CrossRef]
- Primack, R.B.; Miller-Rushing, A.J. Uncovering, collecting, and analyzing records to investigate the ecological impacts of climate change: A template from Thoreau’s Concord. BioScience 2012, 62, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Holz, H.; Segar, J.; Valdez, J.; Staude, I.R. Assessing extinction risk across the geographic ranges of plant species in Europe. Plants People Planet 2022, 4, 303–311. [Google Scholar] [CrossRef]
- Zych, M.; Stpiczyńska, M. Neither protogynous nor obligatory out crossed: Pollination biology and breeding system of the European Red List Fritillaria meleagris L. (Liliaceae). Plant Biol. 2012, 14, 285–294. [Google Scholar] [CrossRef]
- Molina-Venegas, R.; Rodríguez, M.Á.; Pardo-de-Santayana, M.; Ronquillo, C.; Mabberley, D.J. Maximum levels of global phylogenetic diversity efficiently capture plant services for humankind. Nat. Ecol. Evol. 2021, 5, 583–588. [Google Scholar] [CrossRef]
- Tekşen, M. Fritillaria L. In Resimli Türkiye Florası (Illustrated Flora of Turkey), 2nd ed.; Güner, A., Kandemir, A., Menemen, Y., Yıldırım, H., Aslan, S., Ekşi, G., Güner, I., Çimen, A.Ö., Eds.; ANG Vakfı Nezahat Gökyiğit Botanik Bahçesi Yayınları: İstanbul, Turkey, 2018; Volume 2, pp. 800–876. [Google Scholar]
- Huang, J.; Yu, Y.; Liu, Y.M.; Xie, D.F.; He, X.J.; Zhou, S.D. Comparative chloroplast genomics of Fritillaria (Liliaceae), inferences for phylogenetic relationships between Fritillaria and Lilium and plastome evolution. Plants 2020, 9, 133. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, Z.; Zhang, L.; Atanasov, A.G.; Wang, S. Characterization of the isosteroidal alkaloid chuanbeinone from bulbus of Fritillaria pallidiflora as novel antitumor agent in vitro and in vivo. Planta Med. 2016, 82, 195–204. [Google Scholar] [CrossRef]
- Wang, Y.; Aamer, M.; Aslay, M.; Sener, B.; Khan, F.; Wahab, A.; Rahman, A.; Choudhary, M. A new steroidal alkaloid from Fritillaria michailovskyi Fomin. Nat. Prod. Res. 2020, 36, 361–366. [Google Scholar] [CrossRef]
- Shi, Z.J.; Li, J.H. Research progress on key cultivation techniques and planting patterns of Fritillaria. Mod. Agric. Sci. Technol. 2017, 20, 59–61. [Google Scholar]
- Zych, M.; Stpiczyńska, M.; Roguz, K. Pollination biology and breeding system of European Fritillaria meleagris L. (Liliaceae). In Reproductive Biology of Plants; CRC Press: Boca Raton, FL, USA, 2014; pp. 147–163. [Google Scholar]
- Zhang, X.J.; Cui, D.L.; Zong, X.C.; Ren, R.Y.; Wei, J.C.; SITU, L.L.; Zhang, Y.L. Pollination biology and breeding system of Fritillaria ussuriensis Maxim. Acta Bot. Sin. 2010, 30, 1404–1408. [Google Scholar]
- Ma, S.M.; Wang, Y.F.; Ye, X.L.; Zhao, N.X.; Liang, C.Y. Progress in study of apomixis in monocotyledonous plants. Chin. Bull. Bot. 2002, 19, 530–537. [Google Scholar]
- Aslay, M.K.; Çukadar, H.M.; Ünlü, Z.; Kadıoğlu, M.; Tekşen, E.; Kaya, Ö. Determination of Some Productivity Situations of the Soils of Terslale (Fritillaria L.) Species of Eastern Anatolia Flora. V. In Proceedings of the Uluslararası Katılımlı Toprak ve Su Kaynakları Kongresi, Kırklareli, Turkey, 12–15 September 2017. [Google Scholar]
- Türktaş, M.; Aslay, M.; Kaya, E.; Ertuğrul, F. Molecular characterization of phylogeneticrelationships in Fritillaria species inferred from chloroplast trnL-trnF sequences. Turk. J. Biol. 2012, 36, 552–560. [Google Scholar] [CrossRef]
- Rix, E.M. Fritillaria L. In Flora of Turkey and the East Aegean Islands, 1st ed.; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1975; Volume 8, pp. 284–302. [Google Scholar]
- Dafni, A.; Maues, J. A rapid and simple procedure to determine stigma receptivity. Sex. Plant Reprod. 1998, 11, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Shivanna, K.R.; Tandon, R. Reproductive Ecology of Flowering Plants: A Manual; Springer: New Delhi, India, 2014; pp. 107–123. [Google Scholar]
- Cruden, R.W. Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants. Evolution 1977, 31, 32–46. [Google Scholar] [CrossRef]
- Zapata, T.R.; Arroyo, M.T.K. Plant reproductive ecology of a secondary deciduous tropical forest in Venezuela. Biotropica 1978, 10, 221–230. [Google Scholar] [CrossRef]
- Dafni, A. Pollination Ecology: A Practical Approach; University Press: New York, NY, USA, 1992. [Google Scholar]
- Aslay, M.; Çukadar, K.; Ünlü, H.M.; Kadıoğlu, Z.; Tekşen, M. Ekim Öncesi Uygulamalar ve Farklı Çimlenme Ortamlarının Tchihatchewia isatidea Boiss. (Brassicaceae) Tohumlarının Çimlenmesi Üzerine Etkisi. IV. In Proceedings of the Süs Bitkileri Kongresi, Mersin, Turkey, 20–22 October 2010. [Google Scholar]
- Bernardello, G.; Anderson, G.J.; Lopez, S.P.; Cleland, M.A.; Stuessy, T.F.; Crawford, D.J. Reproductive biology of Lactoris fernandeziana (Lactoridaceae). Am. J. Bot. 1999, 86, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S. Identifying Important Plant Areas; Plantlife International: London, UK, 2002. [Google Scholar]
- Nyman, Y. Pollination mechanisms in six Campanula species (Campanulaceae). Plant Syst. Evol. 1992, 181, 97–108. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Cruzan, M.B. Incompatibility in heterostylous plants. In Genetic Control of Self-Incompatibility and Reproductive Development in Flowering Plants; Williams, E.G., Clarke, A.E., Knox, R.B., Eds.; Kluwer: Dordrecht, The Netherland, 1994; pp. 189–219. [Google Scholar]
- Brys, R.; Jacquemyn, H.; Hermy, M. Impact of mate availability, population size, and spatial aggregation of morphs on sexual reproduction in a distylous, aquatic plant. Am. J. Bot. 2007, 94, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Lei, F.W.; Wu, Y.M.; Shen, X.L.; Xia, X.F.; Zhang, D.H.; Xian, Y.M.; Zhang, Z.X. Multiple reproductive strategies of a spring ephemeral plant, Fritillaria maximowiczii, enable its adaptation to harsh environments. Plant Species Biol. 2022, 37, 38–51. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, C.; Song, B.; Du, F. Corolla retention after pollination facilitates the development of fertilized ovules in Fritillaria delavayi (Liliaceae). Sci. Rep. 2019, 9, 729. [Google Scholar] [CrossRef] [Green Version]
- Yashima, T.; Kinoshita, E.; Shimizu, T. Flowering phenology and self-incompatibility in Fritillaria camtschatcensis (L.) Ker-Gawl. J. Phytogeogr. Taxon. 1997, 45, 129–133. [Google Scholar]
- de Jong, T.; Klinkhamer, P. Evolutionary Ecology of Plant Reproductive Strategies; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Aslay, M.; Çukadar, K.; Ünlü, H.M.; Kadıoğlu, Z.; Tekşen, M.; Kaya, E. Doğu Anadolu Fritillaria’larının Kültür Şartlarına Adaptasyonu. V. In Proceedings of the Süs Bitkileri Kongresi, Yalova, Turkey, 6–9 May 2013. [Google Scholar]
- Aslay, M.; Tekşen, M.; Alp, Ş.; Ellialtıoğlu, Ş. 13. Terslale. In Süs Bitkileri Islahı (Türler) Kitabı; Kazaz, S., Yalçın Mendi, N.Y., Eds.; Gece Kitaplığı: Ankara, Turkey, 2021. [Google Scholar]
- Aslay, M. Tescil Edilmiş Ters Lale Süs Bitkisi Çeşidi ”Doğu Güneşi”. VII. In Proceedings of the Ulusal Bahçe Bitkileri Kongresi, Çanakkale, Turkey, 25–29 August 2015. [Google Scholar]
- Karakaş Metin, Ö.; Türktaş, M.; Aslay, M.; Kaya, E. Evaluation of the Genetic Relationship Between Fritillaria Species From Turkey’s Flora Using Fluorescent-based AFLP. Turk. J. Biol. 2013, 37, 273–279. [Google Scholar]
- Yucel, G.; Mengüç, A. Production of Fritillaria imperialis L. Bulbs Using Different Methods and Monitoring the Development of Bulb Groups Under Export Size. Egypt. J. Hortic. 2021, 48, 257–266. [Google Scholar] [CrossRef]
- Newstrom, L.E.; Frankie, G.W.; Baker, H.G.; Colwell, R.K. Diversity of Long-Term Flowering Patterns. La Selva: Ecology and Natural History of A Neotropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1994; Volume 1, pp. 142–160. [Google Scholar]
- Kumari, P.; Khajuria, A.; Wani, I.A.; Khan, S.; Verma, S. Effect of floral size reduction on Pollination and reproductive efficiency of female flowers of Valeriana wallichii, a threatened medicinal plant. Nat. Acad. Sci. Lett. 2021, 44, 75–79. [Google Scholar] [CrossRef]
- Castro, S.; Silveira, P.; Navarro, L. Effect of pollination on floral longevity and costs of delaying fertilization in the out-crossing Polygala vayredae Costa (Polygalaceae). Ann. Bot. 2008, 102, 1043–1048. [Google Scholar] [CrossRef] [Green Version]
- Hermanutz, L.; Innes, D.; Denham, A.; Whelan, R. Very low fruit:flower ratios in Grevillea (Proteaceae) are independent of breeding system. Aust. J. Bot. 1998, 46, 465–478. [Google Scholar] [CrossRef] [Green Version]
- Rocha, O.J.; Aguilar, G. Reproductive biology of the dry forest tree Enterolobium cyclocarpum (Guanacaste) in Costa Rica: A comparison between trees left in pastures and in continuous forest. Am. J. Bot. 2001, 88, 1607–1614. [Google Scholar] [CrossRef]
- Byers, D.L. Pollen quantity and quality as explanations for low seed set in small populations exemplified by Eupatorium (Asteraceae). Am. J. Bot. 2004, 82, 1000–1006. [Google Scholar] [CrossRef]
- Dorken, M.E.; Eckert, C.G. Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J. Ecol. 2001, 89, 339–350. [Google Scholar] [CrossRef]
- Colling, G.; Reckinger, C.; Matthies, D. Effects of pollen quantity and quality on reproduction and offspring vigour in the rare plant Scorzonera humilis (Asteraceae). Am. J. Bot. 2004, 91, 1774–1782. [Google Scholar] [CrossRef]
- Gopalakrishnan, K.K.; Thomas, T.D. Reproductive biology of Pittosporum dasycaulon Miq., (Family Pittosporaceae) a rare medicinal tree endemic to Western Ghats. Bot. Stud. 2014, 55, 15. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.N. Liliaceae. In The Families and Genera of Vascular Plants. III. Flowering Plants. Monocotyledons, Lilianae (Except Orchidaceae); Kubitzki, K., Ed.; Springer: Berlin, Germany, 1998; pp. 343–353. [Google Scholar]
- Kawano, S.; Masuda, J.; Hayashi, K. Life history monographs of Japanese plants. 10: Fritillaria koidzumiana Ohwi (Liliaceae). Plant Species Biol. 2008, 23, 51–57. [Google Scholar] [CrossRef]
- Mancuso, E.; Peruzzi, L. Male individuals in cultivated Fritillaria persica L. (Liliaceae): Real androdioecy or gender disphasy? Turk. J. Bot. 2010, 34, 435–440. [Google Scholar] [CrossRef]
- Kulloli, S.K.; Sreekala, A.K.; Pandurangan, A.G. Floral biology of Impatiens trichocarpa Hook. f., (Balsaminaceae) an endemic Balsam of Western Ghats. Indian J. Sci. Technol. 2009, 2, 30–34. [Google Scholar] [CrossRef]
- Ramsey, M.; Vaughton, G. Effect of environment on the magnitude of inbreeding depression in seed germination in a partially self-fertile perennial herb (Blandfordia grandiflora, Liliaceae). Int. J. Plant Sci. 1998, 159, 98–104. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Mobilization of stored seed reserves. In Seeds; Springer: Boston, MA, USA, 1994; pp. 293–343. [Google Scholar]
- Goulson, D. Bumblebees: Their Behaviour and Ecology; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Nic Lughadha, E.; Bachman, S.P.; Leão, T.C.; Forest, F.; Halley, J.M.; Moat, J.; Carmen, A.; Karen, L.B.; Ryan, F.A.B.; Gildas, G.; et al. Extinction risk and threats to plants and fungi. Plants People Planet 2020, 2, 389–408. [Google Scholar] [CrossRef]
Figure 1.
A view from in-situ (a) and ex-situ (b) habitat of F. aurea (original photo by M. Aslay).

Figure 2.
A view from in-situ (a) and ex-situ (b) habitat of F. Aurea (Photo; M. Aslay original).

Figure 3.
The blossom process of Fritillaria aurea: (a) bud stage, (b) initial flowering, (c) pre-dehiscence stage, (d) first dehiscence stage, (e) full dehiscence, (f) end-dehiscence stage, (g) fresh perianth stage, and (h) perianth wilting stage (original photo by M. Aslay).
Figure 3.
The blossom process of Fritillaria aurea: (a) bud stage, (b) initial flowering, (c) pre-dehiscence stage, (d) first dehiscence stage, (e) full dehiscence, (f) end-dehiscence stage, (g) fresh perianth stage, and (h) perianth wilting stage (original photo by M. Aslay).

Figure 4.
Pollen viability and stigma receptivity of F. aurea; * Means on charts followed by the lowercase letters differ significantly between different flowering stages of F. aurea for pollen viability and stigma receptivity at 0.05 level based on adjusted p-values using Tukey HSD test (a). Correlation between pollen viability and stigma receptivity (b).
Figure 4.
Pollen viability and stigma receptivity of F. aurea; * Means on charts followed by the lowercase letters differ significantly between different flowering stages of F. aurea for pollen viability and stigma receptivity at 0.05 level based on adjusted p-values using Tukey HSD test (a). Correlation between pollen viability and stigma receptivity (b).

Figure 5.
Frequency of insect visits to flowers of F. aurea based on video recordings.

Figure 6.
Duration of insect visits to flowers of F. aurea based on video recordings (X ± SE).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mean fruit production (± SD) in F. aurea plants exposed to five experimental pollination applications.
Table 1.
Mean fruit production (± SD) in F. aurea plants exposed to five experimental pollination applications.
| Treatment | Number of Flowers | Number to Recycle * | Fruit Setting Rate % | Seed Viability (%) | Germination (%) |
|---|---|---|---|---|---|
| Open-pollination | 50 | 48 | 68.0 ± 8.3 ns | 67.0 ± 6.3 ns | 45.0 ± 5.3 ns |
| Spontaneous self-pollination | 50 | 50 | - | - | - |
| Spontaneous cross-pollination | 40 | 39 | 63.0 ± 9.4 | 71.0 ± 9.2 | 40.0 ± 4.9 |
| Apomixis | 30 | 30 | - | - | - |
| Hand self-pollination | 50 | 49 | - | - | - |
Average ± standard deviation (SD). * Number to recycle refers to the number of individuals unharmed by insects or animals in each application; ns: not significant (at 0.05 level).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yıldız, F.; Aslay, M.; Kandemir, A.; Kaya, O. Reproductive Biology of Fritillaria aurea Schott (Liliaceae), a Rare Species Endemic to Turkey. Diversity 2022, 14, 1052. https://doi.org/10.3390/d14121052
AMA Style
Yıldız F, Aslay M, Kandemir A, Kaya O. Reproductive Biology of Fritillaria aurea Schott (Liliaceae), a Rare Species Endemic to Turkey. Diversity. 2022; 14(12):1052. https://doi.org/10.3390/d14121052
Chicago/Turabian StyleYıldız, Faruk, Meral Aslay, Ali Kandemir, and Ozkan Kaya. 2022. "Reproductive Biology of Fritillaria aurea Schott (Liliaceae), a Rare Species Endemic to Turkey" Diversity 14, no. 12: 1052. https://doi.org/10.3390/d14121052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.