
The First Deep-Sea Stylasterid (Hydrozoa, Stylasteridae) of the Red Sea

 , , ,
, , ,  ,
, 
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
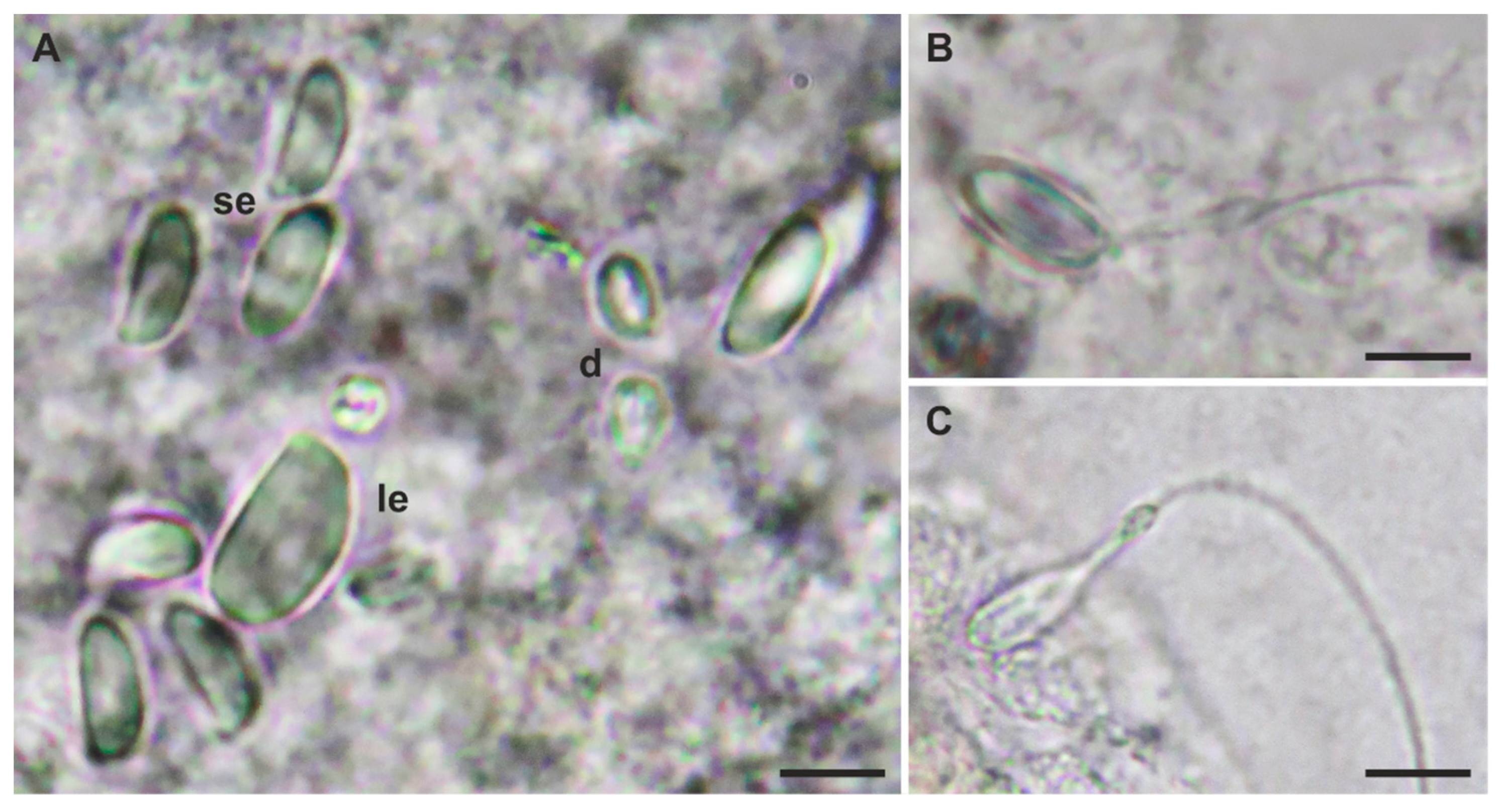{kind=link}
1. Introduction
2. Materials and Methods
3. Results
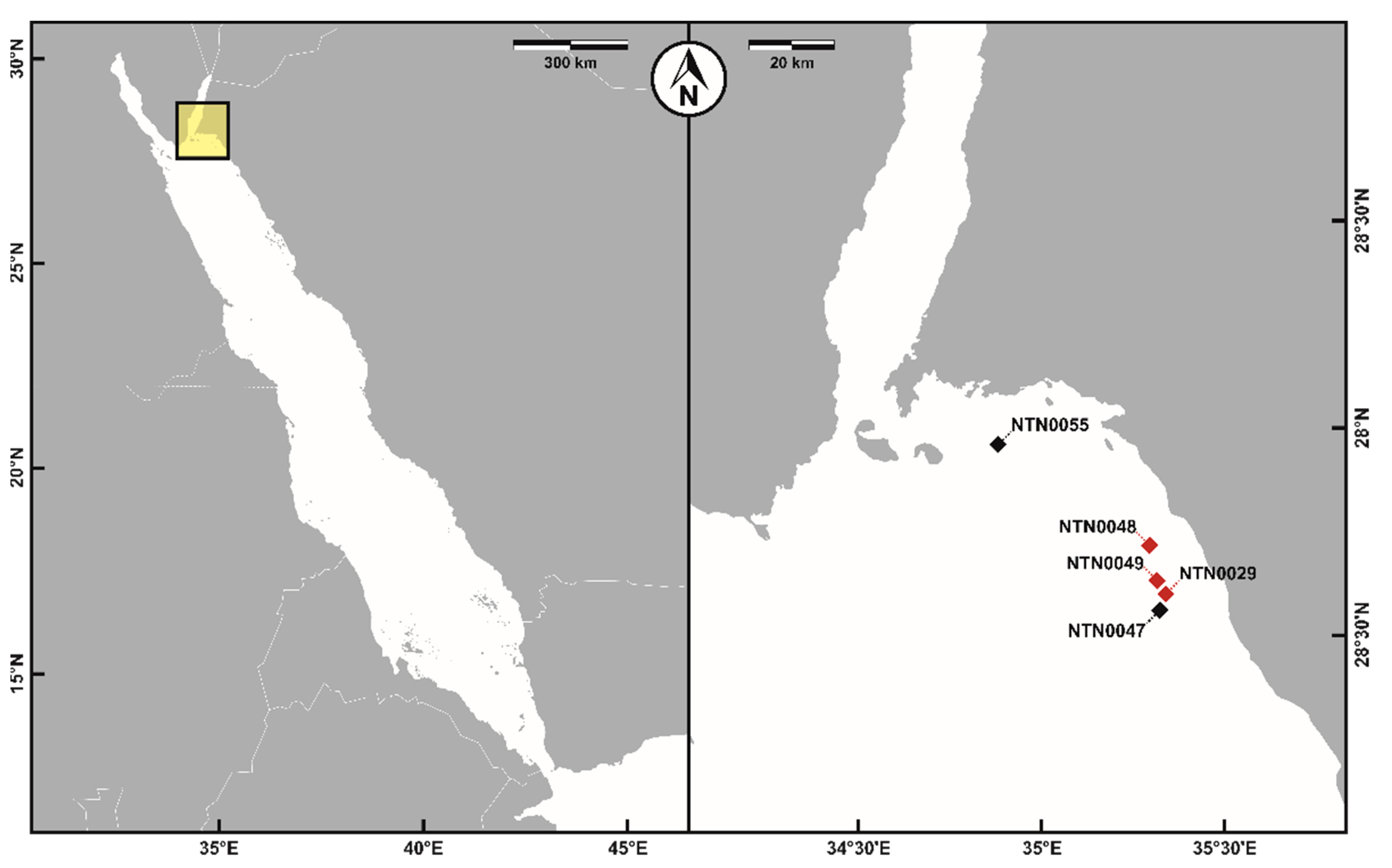
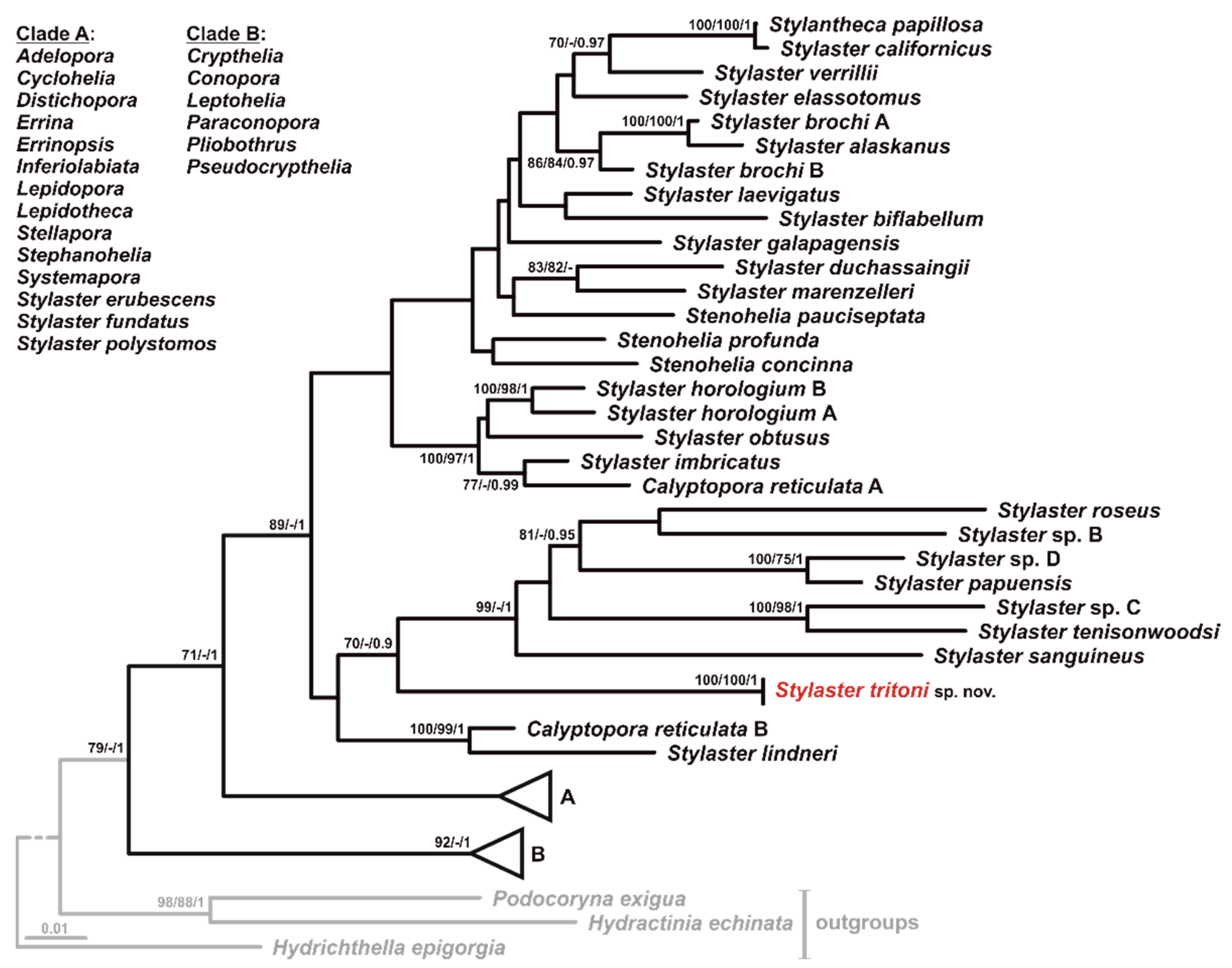
3.1. Sampling and Phylogenetic Analyses
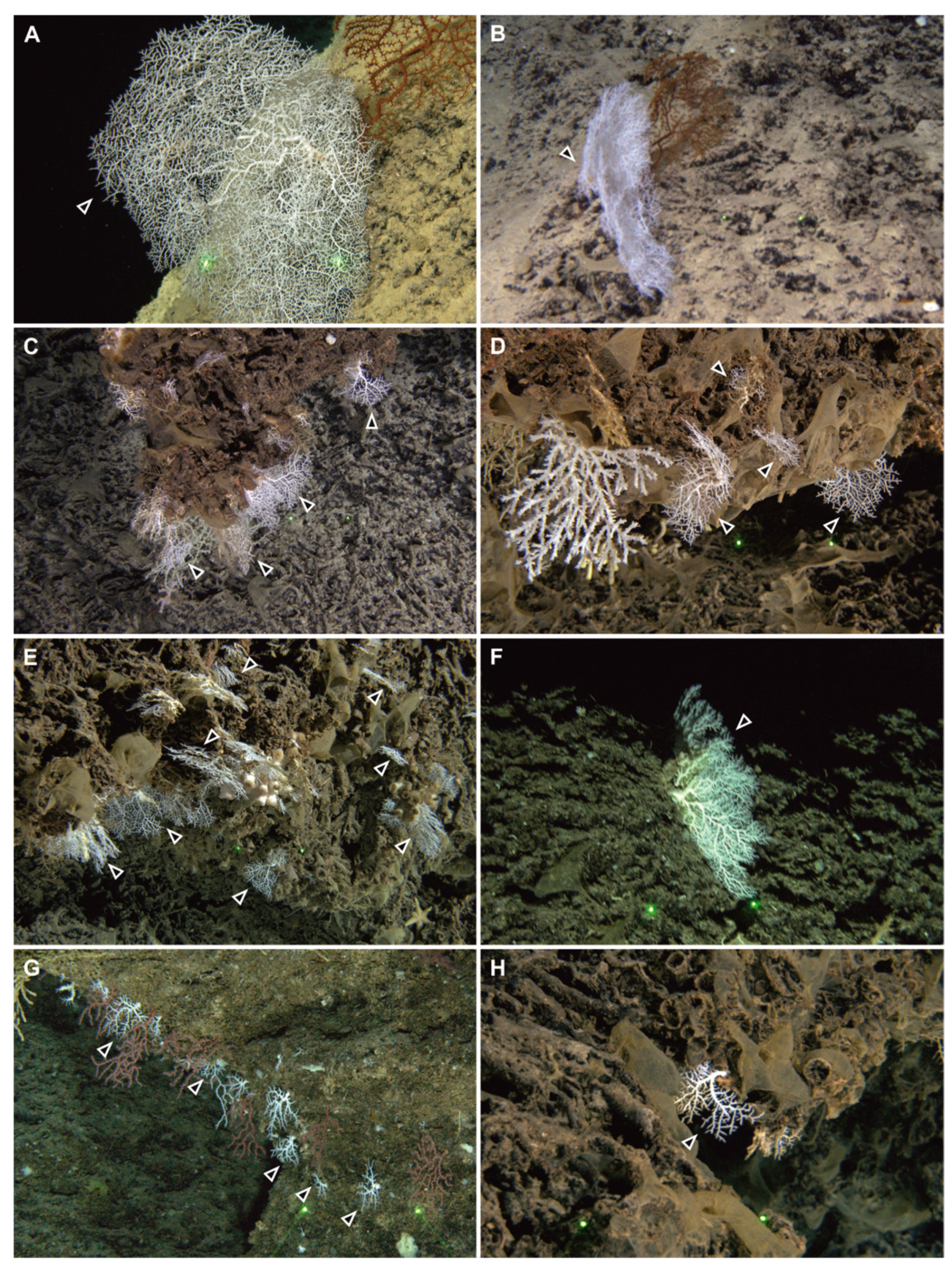
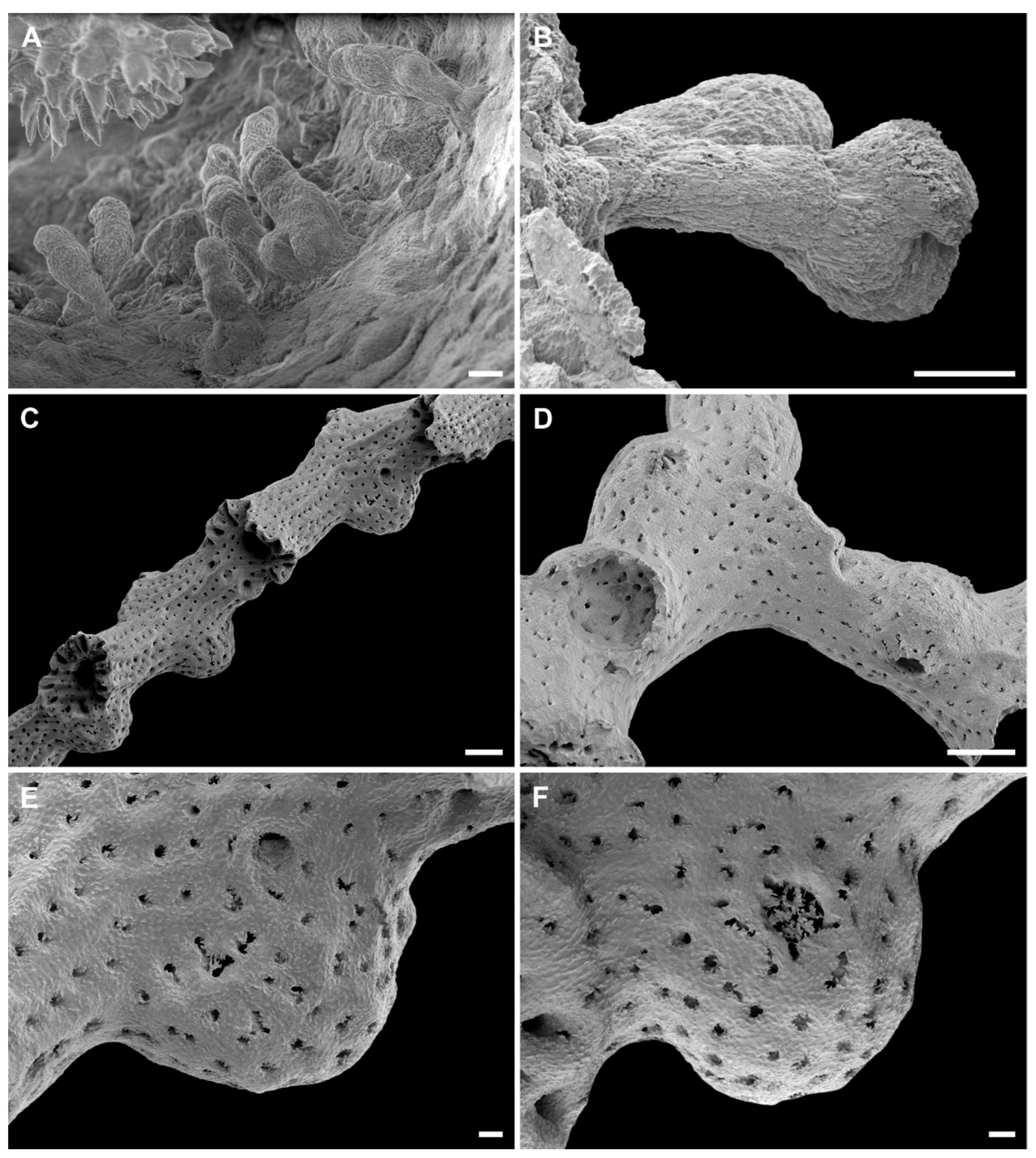
3.2. Taxonomic Account
- Class: Hydrozoa (Owen, 1843)
- Order: Anthoathecata (Cornelius, 1992)
- Family: Stylasteridae (Gray, 1847)
- Genus: Stylaster (Gray, 1831)
- Stylaster tritoni sp. nov. Maggioni, Cairns, Pica & Benzoni
- urn:lsid:zoobank.org:act:194FD2C1-8FEC-4611-AA4E-07B5A9C20B31.
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, J.M.; Wheeler, A.J.; Freiwald, A.; Cairns, S.D. Cold-Water Corals: The Biology and Geology of Deep-Sea Coral Habitats; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-051-158-158-8. [Google Scholar]
- Rex, M.A.; Etter, R.J. Deep-Sea Biodiversity: Pattern and Scale; Harvard University Press: Cambridge, MA, USA, 2010; ISBN 978-067-403-607-9. [Google Scholar]
- Danovaro, R.; Snelgrove, P.V.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Angeletti, L.; Taviani, M.; Canese, S.; Foglini, F.; Mastrototaro, F.; Argnani, A.; Trincarrdi, F.; Bakran-Petricioli, T.; Ceregato, G.; Chimienti, G.; et al. New deep-water cnidarian sites in the southern Adriatic Sea. Mediterr. Mar. Sci. 2014, 15, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Chimienti, G.; Aguilar, R.; Gebruk, A.V.; Mastrototaro, F. Distribution and swimming ability of the deep-sea holothuroid Penilpidia ludwigi (Holothuroidea: Elasipodida: Elpidiidae). Mar. Biodivers. 2019, 49, 2369–2380. [Google Scholar] [CrossRef]
- Chimienti, G.; Angeletti, L.; Furfaro, C.; Taviani, M. Habitat, morphology and trophism of Tritonia callogorgiae sp. nov., a large nudibranch inhabiting Callogorgia verticillata forests in the Mediterranean Sea. Deep Sea Res. I Oceanogr. Res. Pap. 2020, 165, 103364. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Giddens, J.; Ballesteros, E.; Blum, S.; Brown, E.K.; Caselle, J.E.; Henning, B.; Jost, C.; Salinas-de-Léon, P.; Sala, E. Marine biodiversity from zero to a thousand meters at Clipperton Atoll (Île de La Passion), Tropical Eastern Pacific. PeerJ 2019, 7, e7279. [Google Scholar] [CrossRef] [Green Version]
- Augustin, N.; van der Zwan, F.M.; Devey, C.W.; Brandsdóttir, B. 13 million years of seafloor spreading throughout the Red Sea Basin. Nat. Commun. 2021, 12, 2427. [Google Scholar] [CrossRef]
- Berumen, M.L.; Voolstra, C.R.; Daffonchio, D.; Agusti, S.; Aranda, M.; Irigoien, X.; Jones, B.H.; Morán, X.A.G.; Duarte, C.M. The Red Sea: Environmental gradients shape a natural laboratory in a nascent ocean. In Coral Reefs of the Red Sea; Voolstra, C.R., Berumen, M.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 1–10. ISBN 978-303-005-802-9. [Google Scholar]
- Sheppard, C.; Price, A.; Roberts, C. Marine Ecology of the Arabian Region: Patterns and Processes in Extreme Tropical Environments; Academic Press: London, UK, 1992; ISBN 978-012-639-490-0. [Google Scholar]
- Roder, C.; Berumen, M.L.; Bouwmeester, J.; Papathanassiou, E.; Al-Suwailem, A.; Voolstra, C.R. First biological measurements of deep-sea corals from the Red Sea. Sci. Rep. 2013, 3, 2802. [Google Scholar] [CrossRef] [Green Version]
- Qurban, M.A.; Krishnakumar, P.K.; Joydas, T.V.; Manikandan, K.P.; Ashraf, T.T.M.; Quadri, S.I.; Wafar, M.; Qasem, A.; Cairns, S.D. In-situ observation of deep water corals in the northern Red Sea waters of Saudi Arabia. Deep-Sea Res. I Oceanogr. Res. Pap. 2014, 89, 35–43. [Google Scholar] [CrossRef]
- Riegl, B.M.; Bruckner, A.W.; Rowlands, G.P.; Purkis, S.J.; Renaud, P. Red Sea coral reef trajectories over 2 decades suggest increasing community homogenization and decline in coral size. PLoS ONE 2012, 7, e38396. [Google Scholar] [CrossRef]
- Berumen, M.L.; Hoey, A.S.; Bass, W.H.; Bouwmeester, J.; Catania, D.; Cochran, J.E.M.; Khalil, M.T.; Miyake, S.; Mughal, M.R.; Spaet, J.L.Y.; et al. The status of coral reef ecology research in the Red Sea. Coral Reefs 2013, 32, 737–748. [Google Scholar] [CrossRef]
- Loya, Y.; Genin, A.; El-Zibdeh, M.; Naumann, M.S.; Wild, C. Reviewing the status of coral reef ecology of the Red Sea: Key topics and relevant research. Coral Reefs 2014, 33, 1179–1180. [Google Scholar] [CrossRef]
- Rowlands, G.; Purkis, S.; Bruckner, A. Tight coupling between coral reef morphology and mapped resilience in the Red Sea. Mar. Pollut. Bull. 2016, 105, 575–585. [Google Scholar] [CrossRef] [PubMed]
- DiBattista, J.D.; Roberts, M.B.; Bouwmeester, J.; Bowen, B.W.; Coker, D.J.; Lozano-Cortés, D.F.; Choat, J.H.; Gaither, M.R.; Hobbs, J.-P.A.; Khalil, M.T.; et al. A review of contemporary patterns of endemism for shallow water reef fauna in the Red Sea. J. Biogeogr. 2016, 43, 423–439. [Google Scholar] [CrossRef] [Green Version]
- Berumen, M.L.; Arrigoni, R.; Bouwmeester, J.; Terraneo, T.I.; Benzoni, F. Corals of the Red Sea. In Coral Reefs of the Red Sea; Voolstra, C.R., Berumen, M.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 123–155. ISBN 978-303-005-802-9. [Google Scholar]
- DiBattista, J.D.; Howard Choat, J.; Gaither, M.R.; Hobbs, J.-P.A.; Lozano-Cortés, D.F.; Myers, R.F.; Paulay, G.; Rocha, L.A.; Toonen, R.J.; Westneat, M.W.; et al. On the origin of endemic species in the Red Sea. J. Biogeogr. 2016, 43, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Joydas, T.V.; Manokaran, S.; Borja, A.; Rabaoui, L.; Manikandan, K.P.; Ashraf, T.T.M.; Aarif, K.M.; Hussain, S.A.; Baig, M.H.; Shoeb, M.; et al. A baseline quantitative assessment of deep-sea benthic fauna of the Gulf of Aqaba (Northern Saudi Arabia, Red Sea). Mar. Pollut. Bull. 2021, 164, 112028. [Google Scholar] [CrossRef]
- Gravili, C.; Di Camillo, C.G.; Piraino, S.; Boero, F. Hydrozoan species richness in the Mediterranean Sea: Past and present. Mar. Ecol. 2013, 34, 41–62. [Google Scholar] [CrossRef]
- Pica, D.; Bastari, A.; Vaga, C.F.; Di Camillo, C.G.; Montano, S.; Puce, S. Hydroid diversity of Eilat Bay with the description of a new Zanclea species. Mar. Biol. Res. 2017, 13, 469–479. [Google Scholar] [CrossRef]
- Maggioni, D.; Montano, S.; Arrigoni, R.; Galli, P.; Puce, S.; Pica, D.; Berumen, M.L. Genetic diversity of the Acropora-associated hydrozoans: New insight from the Red Sea. Mar. Biodivers. 2017, 47, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Maggioni, D.; Schiavo, A.; Ostrovsky, A.N.; Seveso, D.; Galli, P.; Arrigoni, R.; Francesca, B.; Montano, S. Cryptic species and host specificity in the bryozoan-associated hydrozoan Zanclea divergens (Hydrozoa, Zancleidae). Mol. Phylogenet. Evol. 2020, 151, 106893. [Google Scholar] [CrossRef]
- Maggioni, D.; Arrigoni, R.; Seveso, D.; Galli, P.; Berumen, M.L.; Denis, V.; Hoeksema, B.W.; Huang, D.; Manca, F.; Pica, D.; et al. Evolution and biogeography of the Zanclea-Scleractinia symbiosis. Coral Reefs 2020, 1–17. [Google Scholar] [CrossRef]
- Maggioni, D.; Schuchert, P.; Arrigoni, R.; Hoeksema, B.W.; Huang, D.; Strona, G.; Seveso, D.; Berumen, M.L.; Montalbetti, E.; Collins, R.; et al. Integrative systematics illuminates the relationships in two sponge-associated hydrozoan families (Capitata: Sphaerocorynidae and Zancleopsidae). Contr. Zool. 2021, 90, 487–525. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–417. [Google Scholar] [CrossRef]
- Randall, J.E.; DiBattista, J.D. A new species of damselfish (Pomacentridae) from the Indian Ocean. AQUA 2013, 19, 1–16. [Google Scholar]
- Voigt, O.; Erpenbeck, D.; González-Pech, R.A.; Al-Aidaroos, A.M.; Berumen, M.L.; Wörheide, G. Calcinea of the Red Sea: Providing a DNA barcode inventory with description of four new species. Mar. Biodivers. 2017, 47, 1009–1034. [Google Scholar] [CrossRef]
- Perry, O.; Bronstein, O.; Simon-Blecher, N.; Atkins, A.; Kupriyanova, E.; ten Hove, H.; Levy, O.; Fine, M. On the genus Spirobranchus (Annelida, Serpulidae) from the northern Red Sea, and a description of a new species. Invertebr. Syst. 2018, 32, 605–626. [Google Scholar] [CrossRef]
- Loya, Y. Community structure and species diversity of hermatypic corals at Eilat, Red Sea. Mar. Biol. 1972, 13, 100–123. [Google Scholar] [CrossRef]
- Perkol-Finkel, S.; Benayahu, Y. Community structure of stony and soft corals on vertical unplanned artificial reefs in Eilat (Red Sea): Comparison to natural reefs. Coral Reefs 2004, 23, 195–205. [Google Scholar] [CrossRef]
- Arrigoni, R.; Maggioni, D.; Montano, S.; Hoeksema, B.W.; Seveso, D.; Shlesinger, T.; Terraneo, T.I.; Tietbohl, M.D.; Berumen, M.L. An integrated morpho-molecular approach to delineate species boundaries of Millepora from the Red Sea. Coral Reefs 2018, 37, 967–984. [Google Scholar] [CrossRef] [Green Version]
- Boschma, H. Revision of the Indo-Pacific species of the genus Distichopora. Bijdr. Dierkd. 1959, 29, 121–171. [Google Scholar] [CrossRef] [Green Version]
- Cairns, S.D. Global diversity of the Stylasteridae (Cnidaria: Hydrozoa: Athecatae). PLoS ONE 2011, 6, e21670. [Google Scholar] [CrossRef] [Green Version]
- Lindner, A.; Cairns, S.D.; Cunningham, C.W. From offshore to onshore: Multiple origins of shallow-water corals from deep-sea ancestors. PLoS ONE 2008, 3, e2429. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.D. Deep-water corals: An overview with special reference to diversity and distribution of deep-water scleractinian corals. Bull. Mar. Sci. 2007, 81, 311–322. [Google Scholar]
- Hoarau, L.; Rouzé, H.; Boissin, É.; Gravier-Bonnet, N.; Plantard, P.; Loisil, C.; Bigot, L.; Chabanet, P.; Labarrère, P.; Penin, L.; et al. Unexplored Refugia with High Cover of Scleractinian Leptoseris spp. and Hydrocorals Stylaster flabelliformis at Lower Mesophotic Depths (75–100 m) on Lava Flows at Reunion Island (Southwestern Indian Ocean). Diversity 2021, 13, 141. [Google Scholar] [CrossRef]
- Cairns, S.D. Worldwide distribution of the Stylasteridae (Cnidaria: Hydrozoa). Sci. Mar. 1992, 56, 125–130. [Google Scholar]
- Cairns, S.D. Stylasteridae (Cnidaria: Hydrozoa: Anthoathecata) of the New Caledonian Region. Mém. Mus. Natl. Hist. Nat. 2015, 207, 1–361. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Cunningham, C.W.; Buss, L.W. Molecular evidence for multiple episodes of paedomorphosis in the family Hydractiniidae. Biochem. Syst. Ecol. 1993, 21, 57–69. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Lindner, A.; Cairns, S.D.; Zibrowius, H. Leptohelia flexibilis gen. nov. et sp. nov., a remarkable deep-sea stylasterid (Cnidaria; Hydrozoa: Stylasteridae) from the southwest Pacific. Zootaxa 2014, 3900, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Cairns, S.D.; Lindner, A. A revision of the Stylasteridae (Cnidaria, Hydrozoa, Filifera) from Alaska and adjacent waters. ZooKeys 2011, 158, 1–88. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Available online: http://www.mesquiteproject.org (accessed on 1 October 2021).
- Lanfear, R.; Calcott, B.; Ho, S.Y.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony (and Other Methods), Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggioni, D.; Garese, A.; Huang, D.; Hoeksema, B.W.; Arrigoni, R.; Seveso, D.; Galli, P.; Berumen, M.L.; Montalbetti, E.; Pica, D.; et al. Diversity, host specificity and biogeography in the Cladocorynidae (Hydrozoa, Capitata), with description of a new genus. Cladistics 2021, 38, 13–37. [Google Scholar] [CrossRef]
- Cairns, S.D. A generic revision of the Stylasterina (Coelenterata: Hydrozoa). Part 1. Description of the genera. Bull. Mar. Sci. 1983, 33, 427–508. [Google Scholar]
- Broch, H. Stylasteridae (Hydrocorals) of the John Murray Expedition to the Indian Ocean. Sci. Rep. John Murray Exped. 1947, 26, 33–46. [Google Scholar]
- Boschma, H. Stylasterina in the collection of the Paris Museum: III. Stylaster flabelliformis (Lamarck). Zool. Meded. 1957, 35, 261–282. [Google Scholar]
- Schuchert, P. World Hydrozoa Database. Available online: https://www.marinespecies.org/hydrozoa (accessed on 20 February 2022).
- Cairns, S.D.; Samimi-Namin, K. A new species of Stylaster (Cnidaria: Hydrozoa: Stylasteridae) from the Arabian Sea, off Oman. Proc. Biol. Soc. Wash. 2015, 128, 209–215. [Google Scholar] [CrossRef]
- Cairns, S.D. Revision of the Hawaiian Stylasteridae (Cnidaria: Hydrozoa: Athecata). Pac. Sci. 2005, 59, 439–451. [Google Scholar] [CrossRef] [Green Version]
- Brooke, S.; Stone, R. Reproduction of deep-water hydrocorals (family Stylasteridae) from the Aleutian Islands, Alaska. Bull. Mar. Sci. 2007, 81, 519–532. [Google Scholar]
- Miller, K.J.; Mundy, C.N.; Chadderton, W.L. Ecological and genetic evidence of the vulnerability of shallow-water populations of the stylasterid hydrocoral Errina novaezelandiae in New Zealand’s fiords. Aquat. Conserv. 2004, 14, 75–94. [Google Scholar] [CrossRef]
- Purkis, S.J.; Harris, P.M.; Ellis, J. Patterns of sedimentation in the contemporary Red Sea as an analog for ancient carbonates in rift settings. J. Sediment. Res. 2012, 82, 859–870. [Google Scholar] [CrossRef]
- Giles, E.C.; Saenz-Agudelo, P.; Hussey, N.E.; Ravasi, T.; Berumen, M.L. Exploring seascape genetics and kinship in the reef sponge Stylissa carteri in the Red Sea. Ecol. Evol. 2015, 5, 2487–2502. [Google Scholar] [CrossRef]
- Saenz-Agudelo, P.; Dibattista, J.D.; Piatek, M.J.; Gaither, M.R.; Harrison, H.B.; Nanninga, G.B.; Berumen, M.L. Seascape genetics along environmental gradients in the Arabian Peninsula: Insights from ddRAD sequencing of anemonefishes. Mol. Ecol. 2015, 24, 6241–6255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terraneo, T.I.; Fusi, M.; Hume, B.C.; Arrigoni, R.; Voolstra, C.R.; Benzoni, F.; Forsman, Z.H.; Berumen, M.L. Environmental latitudinal gradients and host-specificity shape Symbiodiniaceae distribution in Red Sea Porites corals. J. Biogeogr. 2019, 46, 2323–2335. [Google Scholar] [CrossRef]
- Terraneo, T.I.; Berumen, M.L.; Arrigoni, R.; Waheed, Z.; Bouwmeester, J.; Caragnano, A.; Stefani, F.; Benzoni, F. Pachyseris inattesa sp. n. (Cnidaria, Anthozoa, Scleractinia): A new reef coral species from the Red Sea and its phylogenetic relationships. ZooKeys 2014, 433, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, J.; Benzoni, F.; Baird, A.H.; Berumen, M.L. Cyphastrea kausti sp. n. (Cnidaria, Anthozoa, Scleractinia), a new species of reef coral from the Red Sea. ZooKeys 2015, 496, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Terraneo, T.I.; Benzoni, F.; Baird, A.H.; Arrigoni, R.; Berumen, M.L. Morphology and molecules reveal two new species of Porites (Scleractinia, Poritidae) from the Red Sea and the Gulf of Aden. Syst. Biodivers. 2019, 17, 491–508. [Google Scholar] [CrossRef]
- Puce, S.; Pica, D.; Schiaparelli, S.; Negrisolo, E. Integration of morphological data into molecular phylogenetic analysis: Toward the identikit of the stylasterid Ancestor. PLoS ONE 2016, 11, e0161423. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maggioni, D.; Terraneo, T.I.; Chimienti, G.; Marchese, F.; Pica, D.; Cairns, S.D.; Eweida, A.A.; Rodrigue, M.; Purkis, S.J.; Benzoni, F. The First Deep-Sea Stylasterid (Hydrozoa, Stylasteridae) of the Red Sea. Diversity 2022, 14, 241. https://doi.org/10.3390/d14040241
Maggioni D, Terraneo TI, Chimienti G, Marchese F, Pica D, Cairns SD, Eweida AA, Rodrigue M, Purkis SJ, Benzoni F. The First Deep-Sea Stylasterid (Hydrozoa, Stylasteridae) of the Red Sea. Diversity. 2022; 14(4):241. https://doi.org/10.3390/d14040241
Chicago/Turabian StyleMaggioni, Davide, Tullia I. Terraneo, Giovanni Chimienti, Fabio Marchese, Daniela Pica, Stephen D. Cairns, Ameer A. Eweida, Mattie Rodrigue, Sam J. Purkis, and Francesca Benzoni. 2022. "The First Deep-Sea Stylasterid (Hydrozoa, Stylasteridae) of the Red Sea" Diversity 14, no. 4: 241. https://doi.org/10.3390/d14040241