
A New Deep-Water Epilithic Green Alga, Ulvella lacustris, from an Alpine Brackish Lake in Qinghai–Tibet Plateau


1
Key Laboratory of Algal Biology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(8), 594; https://doi.org/10.3390/d14080594
Submission received: 28 June 2022
/
Revised: 20 July 2022
/
Accepted: 21 July 2022
/
Published: 26 July 2022
/
Corrected: 26 September 2022
(This article belongs to the Special Issue Diversity and Ecology of Algae in China)
Abstract
:Ulvella species are widely distributed in ocean and freshwater habitats and from high latitudes to the tropics. However, no species of this genus have been found in the Qinghai–Tibetan Plateau. In the present study, five rock specimens were collected from the bottom of Qinghai Lake at a depth of about 15–28 m, from which three unialgal strains were isolated. These strains were characterized by prostrate pseudoparenchyma, irregular or radial branch filaments, and mono- or polystromatic discs or rosettes formed by consecutive filaments, with thalli that can grow up to 180 um in length, suggesting that they may be Ulvella species. Phylogenetic analysis based on 18S rDNA and tufA cpDNA sequences revealed that the isolates formed a strong branch among other Ulvella species and were closely related to Ulvella leptochaete and Ulvella waernii. This result was also confirmed by neighbor-joining tree inference based on ITS2 secondary structure. Unlike other members of Ulvella, the isolated strain was characterized by its unique habitat, field morphology of thallus, size of vegetative cells, and number of pyrenoids. Based on these morphological differences, the phylogenetic analysis, and the comparison of ITS2 secondary structure with relative species, the strain isolated in this study was proposed to be a novel Ulvella sp. The alga was found on the bottom of brackish lakes on the Qinghai–Tibetan Plateau, which meant that our sampling range needed to be expanded further.
1. Introduction
Chlorophyta consists of several deep Prasinophyte clades and the core Chlorophytes (mainly includes three classes: Trebouxiophyceae, Chlorophyceae, and Ulvophyceae) [1,2]. Ulvophyceae are one of the most studied classes and have attracted much attention in view of their unparalleled morphological characteristics, ranging from unicellular and simple multicellular filaments to sheet-like and complex cortical thalloids [3]. The relationship between some orders of Ulvophyceae and the location of some genera have not been well-resolved [3]. Due to extensive morphological variation within genera, the generic boundaries in many Ulvophyceaen groups are often blurred [4,5]. Thus, DNA sequence data have provided more instrumental information in guiding the delimitation of those genera and correctly assigning specimens to these genera [6]. Additionally, intraspecific morphological variation and phenotypic plasticity are very common in many Ulvophycean algae [7], and therefore, DNA taxonomy seems more reliable and useful in species delimitation and identification. The family Ulvellaceae is a group of Ulvophycean algae with diverse habitats and various morphological characteristics [8]. The application of DNA taxonomy has successfully solved some taxonomic problems in Ulvellaceae [9].
The taxonomic research history of Ulvellaceae is very complex [10]. O’Kelly and Floyd (1983) emended Ulvellaceae mainly based on the characteristics of sporangia and gametangia, and only six genera were retained: Acrochaete, Endophyton, Endocladia, Ochlochaete, Pringsheimiella, and Ulvella [11]. Ulvella, proposed by Crouan et al. in 1859 [12], was originally described as a plant body 1–2 mm in diameter, formed by spherical, ovate, and angular cells. Acrochaete was established in 1862 and described as uniseriate irregularly branched filaments; the main feature of the genus is the formation of small hyaline cells from which the hair projects [13]. Endophyton was originally proposed in 1909 and described as irregularly branched filiform; the branches are usually shorter, and it has been recorded in the Pacific coast of North America, the Bering Sea, and the Antarctic Sea [14]. Ochlochaete was proposed in 1849 and recorded as a small pseudoparenchyma thallus of uniseriate branching filaments; species of this genus act as epiphytes in salt and brackish water and as part of thin algal mats covering shells, small stones, wood, etc. [15]. By observing the ultrastructure of vegetative cells, [16] confirmed the close relationship between ulvalean algae and transferred Endophyton to Entocladia. However, as a result of a phylogenetic location closer to Ulvellaceae, Ochlochaete was removed from Ulvellaceae by O’Kelly et al. [17]. Nielsen et al. [18] revised this family using the polyphasic method combined with molecular sequence and morphological characteristics, retaining only Ulvella in Ulvellaceae and transferring species originally belonging to Acrochaete, Endophyton, Ochlochaete, Entocladia, and Pringsheimela to Ulvella. Zhu et al. reported the freshwater Ulvella tongshanensis and placed Jaoa species into Ulvella. Now, Ulvella is characterized by upright or prostrate, irregular, or radial branch filaments and mono- or polystromatic discs or rosettes formed by consecutive filaments, with a uniseriate basal and parenchymatous middle part [19].
As the only remaining genus of Ulvellaceae, Ulvella comprises 66 species according to AlgaeBase and is distributed from polar regions to tropical areas [20]. Most Ulvella species are recorded in marine or brackish habitats, and only a few species are reported from freshwater habitats. The holotype species is Ulvella lens, and the vast majority of other species are often recorded as epiphytic, epizoic, endophytic, or endolithic; only rare species are found to be free-living [21,22,23,24,25]. Due to the morphological variability caused by habitat diversity and morphological plasticity, molecular methods have become more effective means of dealing with taxonomic problems of Ulvella. In the present study, three algal specimens were sampled from five different rocks collected from the bottom of the lake, of which three unialgal isolates were successfully cultivated and analyzed. According to microscopic observation, phylogenetic analysis based on 18S rDNA and tufA genes, and comparison of the ITS2 secondary structure with other Ulvella species, we proposed a novel species, U. lacustris. This is the first record that a Deep-water Epilithic Ulvella was found at the bottom of a brackish inland lake.
2. Materials and Methods
2.1. Sampling and Culture
In June 2021, algal strains were collected from the green rocks from the bottom of Qinghai Lake (37°13′ N, 100°23′ E) at a depth of about 15–28 m. The distribution of sampling points is shown in Figure 1. The algal strains were inoculated onto BG-11 solid medium containing 1.3% agar at a constant temperature of 25 °C under a 12/12 h light/dark cycle until visible colonies appeared [26]. Subsequently, the colonies were transferred to fresh medium until unialgal culture was obtained. The unialgal culture was transferred to a 96-well plate containing BG-11 liquid medium. The algal strains were stored in the freshwater algae culture bank (Freshwater Algae Culture Collection at the Institute of Hydrobiology) of the Institute of Aquatic Biology, Chinese Academy of Sciences, China (No. 7, Donghu South Road, Wuhan, Hubei), under the Accession No. FACHB-3450. To observe asexual and sexual reproduction, the algal strains were transferred to nitrogen-deficient BG-11 medium. The morphological observation of natural and cultivated algal strains was performed using a Leica DM5000B microscope, and micrographs were obtained using a Leica DFC320 digital camera. Table 1 provides the major habitat information and physical and chemical indices of the sampling points.
2.2. DNA Extraction and PCR Amplification
For DNA extraction, the pure unialgal cells were disrupted using a bead beater (3110BX, Biospec Products, Buttersville, MI, USA) with mini beads. The total DNA was extracted using the HP Plant DNA Kit (Omega Bio-Tek, Norcross, GA, USA). PCR amplification was performed using 6 μL of template DNA, 1 μL of each primer, and 42 μL of Master Mix in 50 μL reaction volume. The 18S rDNA sequences were amplified using the primers 18SR and 18SF [27], and the amplification conditions were as follows: 94 °C for 5 min, followed by 94 °C for 50 s, 55 °C for 50 s, 72 °C for 90 s, and a final extension at 72 °C for 10 min. The tufA sequence was amplified using the primers tufAF and tufAR [28], and the amplification conditions were as follows: 94 °C for 5 min, followed by 32 cycles of 94 °C for 50 s, 56.5 °C for 50 s, and 72 °C for 70 s, and a final extension at 72 °C for 10 min. The primers used to amplify the ITS sequence were NS7m and LR1850 [29], and the amplification conditions were as follows: 32 cycles of 94 °C for 5 min, 94 °C for 1 min, 55 °C for 1 min, 72 °C for 2 min, and a final extension at 72 °C or 10 min. The PCR products were sequenced by TSINGKE Biotechnologies (China), and the sequences have been deposited in GenBank (http://www.ncbi.nlm.nih.gov/, accessed on 22 November 2021) with login numbers OL604474–OL604476 and OL630150–OL630153.
2.3. Phylogenetic Analyses
A total of 64 18S rDNA and 61 tufA sequences were downloaded from GenBank based on BLAST search selection, as well as possible phylogenetic relationships and broader green algae, and preliminarily aligned using MAFFT 7.3 [29]. Manual optimization was achieved through Seaview [30]. To evaluate the mutational saturation of the three alignments’ saturation in variable positions, pairwise distances were plotted against model-corrected distances using Tamura and Nei (1993) and Kimura (1980) models estimated in MEGA (v.11.0) [19,31], and neither transversion nor transformation reached saturation. Phylogenies were estimated using maximum likelihood (ML) in PhyloSuite (v.1.2.1) [32] and Bayesian inference in MrBayes (v. 3.1.2) [33]. The best-fit evolutionary model was selected using hierarchical likelihood ratio tests and Akaike information criterion through Modeltest (v.3.06) [34]. TN93 + G + I and T92 + G + I were found to be the best-fit models for 18S rDNA and tufA, respectively. For ML analysis, tree search was realized using a heuristic search option with the random addition of sequences (10 replicates) and a tree bisection and reconnection branch-swapping algorithm. Statistical reliability was estimated by Bootstrap analysis with 1000 replicates of the dataset for ML. Four Markov chains (three heated chains, one cold chain) were run for Bayesian Markov Chain Monte Carlo analysis for 20 million generations with tree sampling performed for every 10,000 generations. When the average standard deviation of split frequencies between the two runs was lower than 0.01, stationary distribution was assumed. The first 25% of the trees were discarded, the consensus tree with the remaining samples was constructed, and a posteriori probability was inferred. The phylogenetic trees were edited using Figtree1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/, accessed on 10 July 2022).
2.4. Analyses of ITS2 Secondary Structure
The prediction of the ITS2 secondary structure was completed in the ITS2 database (http://its2.bioapps.biozentrum.uni-wuerzburg.de/). By comparing the secondary structure of ITS2, the difference between the target sequence and the sequence in the database was analyzed. Cluster W [35] multiple alignment sequence in 4SALE version 1.7 software was used to align the ITS2 sequence and the secondary structure [36]. The results obtained by comparison were entered into ProfDistS for neighbor-joining tree reconstruction [37]. The secondary structure images were edited using Varna v3.1 [38].
3. Results
3.1. Ulvella lacustris Q. YAN et H. ZHU sp. nov.
Description: Small (up to 2 mm in maximum diameter) green-to-olive spots on the rough and irregular rock surface (Figure 2a,b). Thalli often appeared as pseudoparenchyma, with a thick gelatinous envelope and thick cell wall. The plant can grow up to 180 um in length. The cell arrangement was compact, and the cell shape was severely compressed Figure 2c,e and Figure 3a). Young thalli were irregular, unicellular, and composed of a layer of cells, and the gelatinous envelope was significantly obvious (Figure 2d,f). Mature thalli consisted of several filaments or elongated parenchyma cells, and the gelatinous envelope was not obvious (Figure 2g and Figure 3b). Vegetative cells were nearly cylindrical, unbranched or with irregular filamentous branches extending outward from the base; chloroplasts and pyrenoids were clearly visible (Figure 3c–g). Suspected empty sporangium was observed (Figure 3h). The cells in the middle or marginal thalli were irregular or spherical; the distal cell width was 8–24 μm and length/width ratio was 1.0–5.0; and the middle cell width was 8–29 μm and length/width ratio was 1.0–4.0. The marginal cells were mainly cylindrical, short filamentous body, sometimes branching, and the terminal cells were slightly bluntly tapering. The vegetative chloroplast was nearly band-shape, almost filling the whole cell, with obvious one to five pyrenoids. Asexual and sexual reproduction were not observed.
Culture observations: There were significant differences in the morphology between field and culture samples, as well as in the culture morphology between solid medium and liquid medium. The field samples presented pseudoparenchyma and were nearly spherical, and the cell wall and integument were thicker (Figure 2c–d). By contrast, the culture samples had filiform branches and were creeping compact, the central part was composed of many spherical cells, and the gelatinous envelope was not obvious (Figure 3a–c). Furthermore, pyrenoids and chloroplasts were widely detected in solid medium, whereas empty sporangia were observed in liquid medium (Figure 3h). Hair was not observed on samples grown in standard medium and nitrogen-deficient medium (BBM or BG-11).
Etymology. The species epithet refers to the Holotype habitat (Qinghai Lake).
Type locality. Qinghai Lake (37°13′ N, 100°23′ E), Qinghai Province, China; on a rock surface.
Iconotype. Figure 2f.
Holotype. QH2016 (HBI), collected by Qiu–Feng Yan and Huan Zhu, June 2021; deposited in the Freshwater Algal Herbarium (HBI), Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, Hubei Province, China.
Distribution: At present, the algal isolate is only found in Qinghai Lake, China. It grows on rough and irregular rock surfaces, making the rock appear dark green in color, which is clearly visible to the naked eye.
3.2. Phylogenetic Analyses
A total of 64 Chlorophyta sequences (including 12 Ulvellaceae sequences) were used for 18S rDNA alignment and consisted of 1655 sites, of which 687 (41.5%) and 496 (30.0%) were variable and parsimony-informative sites, respectively. The tufA alignment included 58 Ulvella sequences and 855 sites, of which 313 (36.6%) and 243 (28.4%) were variable and parsimony-informative sites, respectively. Table 2 presents detailed information about the alignment and nucleotide substitution in 18S rDNA and tufA concatenated phylogenies for ML analysis.
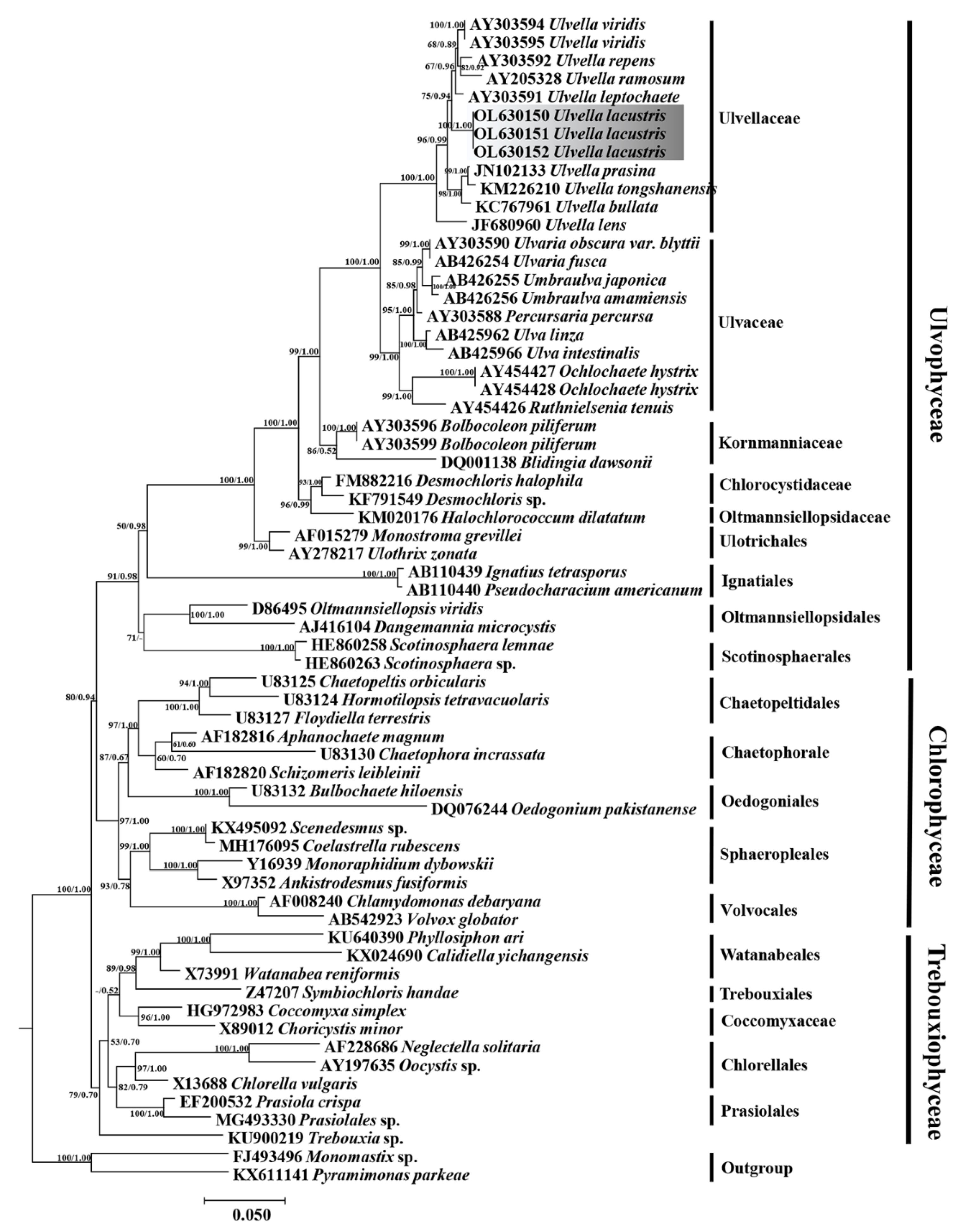
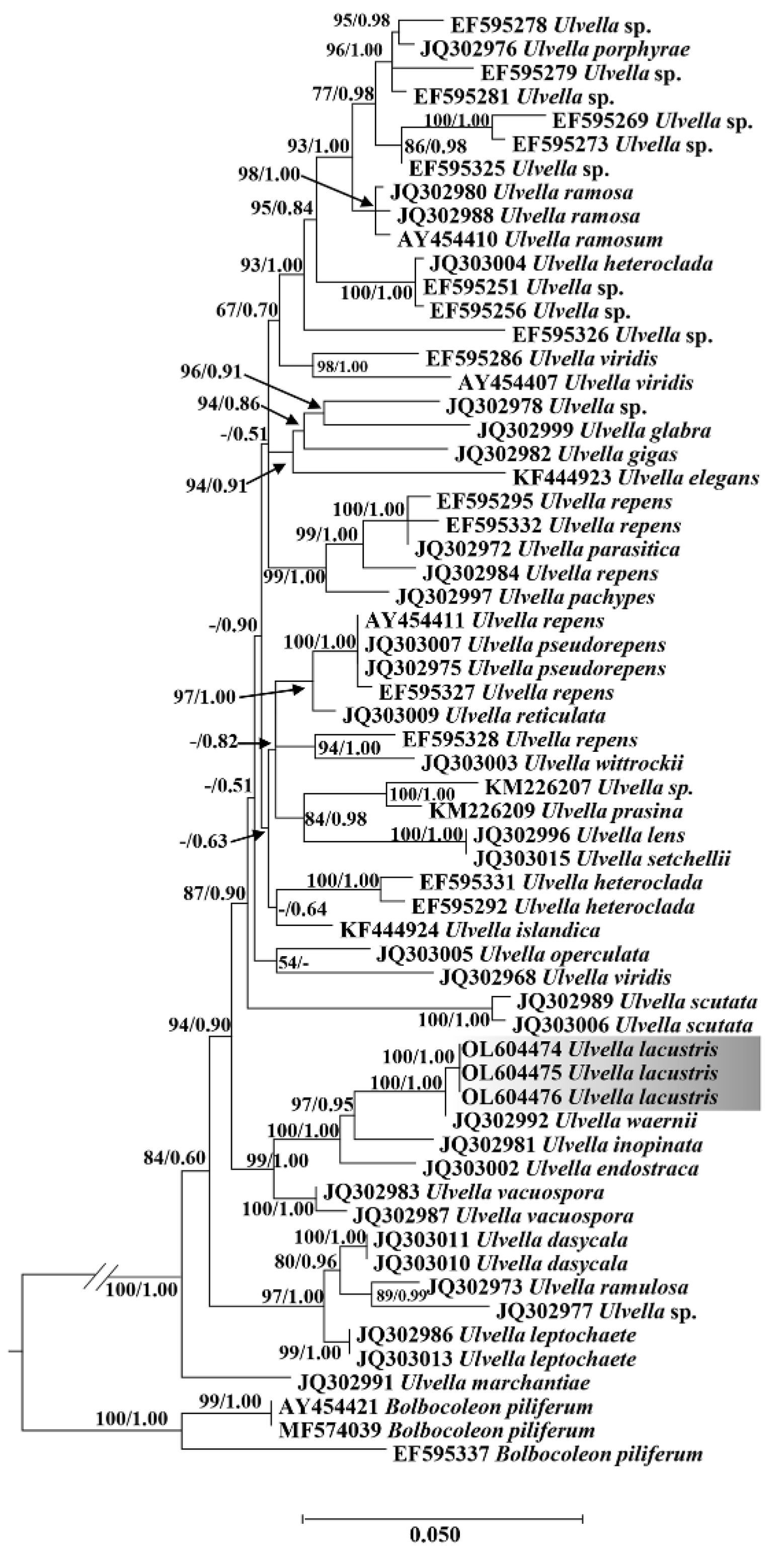
Figure 4 and Figure 5 show the ML tree based on 18S rDNA and tufA alignments, with the Bayesian posterior probabilities and indication of ML bootstrap support. The topology of our tufA and 18S rDNA phylogeny (Figure 4) is essentially consistent with that reported in previous studies [19]. The ML tree based on the tufA gene showed that U. lacustris (OL604474) formed a large clade with U. waernii (JQ302992), U. inopinata (JQ302981), U. endostraca (JQ303002), U. vacuospora (JQ302983), and U. vacuospora (JQ302987) and presented high ML support value and Bayesian posterior probability; hence, the algal isolates were classified into the genus Ulvella. The tree based on tufA (Figure 5) sequences presented the interspecific relationships within the genus Ulvella, and the algal strain isolated in this study formed a distinct separate branch and clustered with Ulvella waernii (JQ302992), which was consistent with the comparison results from the NCBI database, with Ulvella inopinata (JQ302981) forming its sister branch.
3.3. ITS2 Secondary Structure
The ITS2 secondary structure was annotated, and eight Ulvella strains were detected (Figure 6a), namely, U. lacustris (OL630153), U. heteroclada (EF595353), U. leptochaete (EF595355), U. repens (EF595349), U. viridis (EF595351), U. leptochaete (MK910764), U. leptochaete (MK910760), and U. leptochaete (MN510853). Pairwise comparison was employed to analyze the differences among these strains. A total of zero to four compensatory base changes (CBCs) and zero to four hemi-compensatory base changes (h-CBCs) were detected among these strains. On comparing our algal strain with the other seven strains, one to four CBCs were found; however, no CBCs were found in U. viridis. On comparing U. lacustris with the other algal strains, two to three h-CBCs were detected. Furthermore, on comparing the four U. leptochaete strains, zero to one CBCs and no h-CBCs were found. Moreover, comparison of U. leptochaete (EF595355) and U. viridis revealed no CBCs. Table 3 provides detailed information about the CBCs and h-CBCs in all the examined Ulvella strains.
4. Discussion
In the present study, the ML tree constructed from 18S rDNA alignments revealed that the algal isolates formed a robust branch among Ulvella and were closely related to U. leptochaete (AY303591). This finding was confirmed by the neighbor-joining tree based on ITS2 secondary structure (Figure 6b). The tufA gene is the most used marker for phylogenetic analysis in the genus Ulvella [39,40]. The ML tree based on the tufA gene showed that U. lacustris (OL604474) and U. waernii (JQ302992) were most closely related. These molecular phylogenetic data support the classification of our novel algal isolates into the genus Ulvella.
The ITS2 secondary structure analysis detected zero to four CBCs and zero to four h-CBCs among all the strains. These strains exhibited different degrees of variation. The emergence of CBCs is often used as an indicator of isolated species or genera [41,42,43]. On comparing our algal strain with the other seven strains, one to four CBCs were found, except in U. viridis (EF595351). However, two to three h-CBCs were detected between U. lacustris and the remaining algal strains, which indicated that U. lacustris (OL604474) is unique. On comparing the four U. leptochaete strains, zero to one CBCs and no h-CBCs were found. Furthermore, no CBCs were detected between U. leptochaete (EF595355) and U. viridis (EF595351). The phylogenetic tree reconstructed from the ITS2 secondary structure and the sequences of the eight algal strains suggested that U. lacustris (OL604474) formed a clear and independent branch, separated from U. viridis (EF595351). These obvious differences validate that our algal isolate is a novel Ulvella sp.
U. lacustris (OL604474) is different from other Ulvella species with respect to its unique habitat, size of vegetative cells, and number of pyrenoids. Most of the Ulvella species are found in marine or freshwater habitats, and our samples were also collected from brackish lake. This difference in geography and habitat may be one of the reasons for the unique morphology and phylogeny of U. lacustris. To better distinguish U. waernii (JQ302992) and U. lacustris (OL604474), their morphological characteristics were compared. Previous studies have proposed eight natural growth forms for the genus Ulvella [26,38]. Accordingly, the algal strain isolated in the present study could grow into calcified material [8], whereas U. waernii is heterotrichous. Furthermore, U. waernii has one to two pyrenoids, with a distal cell width of 6–8.5 μm and middle cell width of 11–16.5 μm, and contains hair and apical/semi-apical, elongate cylindrical sporangia [18]. By contrast, U. lacustris has one to five pyrenoids, with a distal cell width of 8–24 μm and middle cell width of 8–29 μm. While U. waernii is attached to the lower leaf sheath of reed, U. lacustris grows on the rough rock surface at the bottom of the lake. This habitat may be the main reason for the morphological differences between U. waernii and U. lacustris. Similarly, differences in morphology were also noted between field and culture samples, which has often been observed in the members of Ulvellaceae [18,23,44]. Many Ulvella species frequently present Acrochaete-type hair, and their location and type are used as auxiliary features to distinguish them. However, in the present study, the algal strain grown in standard medium and nitrogen-deficient medium (BBM or BG-11) did not contain hair. This characteristic is different from some of the Ulvella species. Thus, hair is not a stable morphological feature, and the presence/absence of hair probably should not be used as a feature for the classification of the genus Ulvella. Therefore, intraspecific morphological variation and phenotypic plasticity occur frequently in Ulvophycean algae [7]. Unlike most of the marine Ulvella species, the Ulvella strain isolated in the present study was derived from the bottom of brackish lake. The low light intensity and the stress of a brackish environment suggest that our isolated algal strain has tenacious vitality and that there may be many undiscovered algal species growing in extreme environments.
5. Conclusions
Based on phylogenetic analysis, comparison of the ITS2 secondary structure, and morphological characteristics, U. lacustris was proposed as a novel Ulvella speices. The unique habitat of U. lacustris, epilithic on the bottom of a brackish lake, may be the main reason for the neglect of this species, and further investigation of widely sampling similar habitats may be helpful for understanding such algal diversity and finding such novel algae.
Author Contributions
Methodology, H.Z.; investigation, Q.Y, H.Z., Q.D. and B.L.; writing—original draft preparation, Q.Y.; writing—review and editing, Q.Y. and H.Z.; project administration, G.L.; funding acquisition, H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by The Second Tibetan Plateau Scientific Expedition and Research program (Grant No. 2019QZKK0304), and by the National Key Research and Development Project of China (2019YFD0900603).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data can be found within the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Friedl, T. The evolution of the green algae. In Origins of Algae and Their Plastids; Springer: Vienna, Austria, 1997; pp. 87–101. [Google Scholar]
- Cocquyt, E.; Verbruggen, H.; Leliaert, F.; De Clerck, O. Evolution and cytological diversification of the green seaweeds (Ulvophyceae). Mol. Biol. Evol. 2010, 27, 2052–2061. [Google Scholar] [CrossRef] [PubMed]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Phylogeny and molecular evolution of the green algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef]
- Boedeker, C.; Leliaert, F.; Zuccarello, G.C. Molecular phylogeny of the Cladophoraceae (Cladophorales, Ulvophyceae), with the resurrection of Acrocladus Nägeli and Willeella Børgesen, and the description of Lurbica gen. nov. and Pseudorhizoclonium gen. nov. J. Phycol. 2016, 52, 905–928. [Google Scholar] [CrossRef] [PubMed]
- Steinhagen, S.; Barco, A.; Wichard, T.; Weinberger, F. Conspecificity of the model organism Ulva mutabilis and Ulva compressa (Ulvophyceae, Chlorophyta). J. Phycol. 2019, 55, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.A.; Hendrixson, B.E.; Brewer, M.S.; Bond, J.E. An evaluation of sampling effects on multiple DNA barcoding methods leads to an integrative approach for delimiting species: A case study of the North American tarantula genus Aphonopelma (Araneae, Mygalomorphae, Theraphosidae). Mol. Phylogenet. Evol. 2014, 71, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Schlagintweit, F.; Bover-Arnal, T. The morphological adaptation of Lithocodium aggregatum Elliott (calcareous green alga) to cryptic microhabitats (Lower Aptian, Spain): An example of phenotypic plasticity. Facies 2012, 58, 37–55. [Google Scholar] [CrossRef]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Freshwater Flora of Central Europe, Vol 13: Chlorophyta: Ulvophyceae (Süßwasserflora von Mitteleuropa, Bd. 13: Chlorophyta: Ulvophyceae) (Vol. 13); Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Wysor, B.; O’Kelly, C.J.; Bellows, W.K. 178 Molecular Systematics of the Ulvellaceae (Ulvales, Ulvophyceae) Inferred from Nuclear and Chloroplast DNA Sequences. J. Phycol. 2003, 39, 61. [Google Scholar] [CrossRef]
- Nielsen, R.; McLachlan, J. Acrochaete marchantiae comb. nov. and Trichothyra irregularis gen. et sp. nov. with notes on other species of small filamentous green algae from St. Lucia (West Indies). Nord. J. Bot. 1986, 6, 515–524. [Google Scholar] [CrossRef]
- O’kelly, C.J.; Floyd, G.L. The flagellar apparatus of Entocladia viridis motile cells, and the taxonomic position of the resurrected family Ulvellaceae (Ulvales, Chlorophyta)1. J. Phycol. 1983, 19, 153–164. [Google Scholar] [CrossRef]
- Crouan, P. Notice sur quelques especes et genres nouveaux d’algues marines de la rade de Brest. Ann. Sci. Nat. Bot. 1859, 4, 288–295. [Google Scholar]
- Pringsheim, N. Beiträge zur Morphologie der Meeres-Algen; Dümmler in Comm: Berlin, Germany, 1862. [Google Scholar]
- Gardner, N.L. New Chlorophyceae from California; The University Press: Morgantown, WV, USA, 1909. [Google Scholar]
- Harvey, W.H. Phycologia Britannica: Or a History of British Sea-Weeds, Containing Coloured Figures, Generic and Specific Characters, Synonyms, and Descriptios of All the Species of Algae Inhabiting the Shores of the British Islands; Reeve Brothers: London, UK, 1846; Volume 4. [Google Scholar]
- Leonardi, P.I.; Correa, J.A.; Cáceres, E.J. Ultrastructure and taxonomy of the genus Endophyton (Ulvales, Ulvophyceae). Eur. J. Phycol. 1997, 32, 175–183. [Google Scholar] [CrossRef]
- O’Kelly, C.J.; Wysor, B.; Bellows, W.K. Gene sequence diversity and the phylogenetic position of algae assigned to the genera Phaeophila and Ochlochaete (Ulvophyceae, Chlorophyta). J. Phycol. 2004, 40, 789–799. [Google Scholar] [CrossRef]
- Nielsen, R.; Petersen, G.; Seberg, O.; Daugbjerg, N.; Wysor, B. Revision of the genus Ulvella (Ulvellaceae, Ulvophyceae) based on morphology and tufA gene sequences of species in culture, with Acrochaete and Pringsheimiella placed in synonymy. Phycologia 2013, 52, 37–56. [Google Scholar] [CrossRef]
- Zhu, H.; Leliaert, F.; Zhao, Z.J.; Xia, S.; Hu, Z.Y.; Liu, G.X. Ulvella tongshanensis (Ulvellaceae, Chlorophyta), a new freshwater species from China, and an emended morphological circumscription of the genus Ulvella. Fottea 2015, 15, 95–104. [Google Scholar] [CrossRef]
- Guiry, M.; Guiry, G. AlgaeBase. World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2012. [Google Scholar]
- Goldberg, W.; Makemson, J.; Colley, S. Entocladia endozoica sp. nov., a pathogenic chlorophyte: Structure, life history, physiology, and effect on its coral host. Biol. Bull. 1984, 166, 368–383. [Google Scholar] [CrossRef]
- Bown, P.; Plumb, J.; Sánchez-Baracaldo, P.; Hayes, P.K.; Brodie, J. Sequence heterogeneity of green (Chlorophyta) endophytic algae associated with a population of Chondrus crispus (Gigartinaceae, Rhodophyta). Eur. J. Phycol. 2003, 38, 153–163. [Google Scholar] [CrossRef]
- Nielsen, R.; Gunnarsson, K.; Daugbjerg, N.; Petersen, G. Description of Ulvella elegans sp. nov. and U. islandica sp. nov.(Ulvellaceae, Ulvophyceae) from Iceland–a study based on morphology of species in culture and tufA gene sequences. Eur. J. Phycol. 2014, 49, 60–67. [Google Scholar] [CrossRef]
- Marcelino, V.R.; Verbruggen, H. Multi-marker metabarcoding of coral skeletons reveals a rich microbiome and diverse evolutionary origins of endolithic algae. Sci. Rep. 2016, 6, 31508. [Google Scholar] [CrossRef]
- Soares, L.P.; Guimaraes, S.M.; Fujii, M.T.; Batista, M.G.S.; Yoneshigue-Valentin, Y.; Yokoya, N.S. New insights on the distribution and habitat of Ulvella endozoica (Ulvellaceae, Chlorophyta) in the tropical Southwestern Atlantic, based on thallus ontogeny in culture and DNA barcoding. Mar. Biodivers. 2021, 51, 12. [Google Scholar] [CrossRef]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates 1, 2. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Famà, P.; Wysor, B.; Kooistra, W.H.; Zuccarello, G.C. Molecular phylogeny of the genus Caulerpa (Caulerpales, Chlorophyta) inferred from chloroplast tufA gene. J. Phycol. 2002, 38, 1040–1050. [Google Scholar] [CrossRef]
- Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772. [Google Scholar]
- Manolo, G.; Stéphane, G.; Olivier, G. SeaView Version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic Tree Building. Mol. Biol. Evol. 2012, 27, 221–224. [Google Scholar]
- Tamura, K. Molecular evolutionary genetics analysis using Maximum Likelihood, evolutionary distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Molecular ecology resources 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian inference of phylogeny and its impact on evolutionary biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Seibel, P.N.; Müller, T.; Dandekar, T.; Wolf, M. Synchronous visual analysis and editing of RNA sequence and secondary structure alignments using 4SALE. BMC Res. Notes 2008, 1, 91. [Google Scholar] [CrossRef]
- Friedrich, J.; Dandekar, T.; Wolf, M.; Müller, T. ProfDist: A tool for the construction of large phylogenetic trees based on profile distances. Bioinformatics 2005, 21, 2108–2109. [Google Scholar] [CrossRef] [PubMed]
- Darty, K.; Denise, A.; Ponty, Y. VARNA: Interactive drawing and editing of the RNA secondary structure. Bioinformatics 2009, 25, 1974. [Google Scholar] [CrossRef] [PubMed]
- Saunders, G.W.; Kucera, H. An evaluation of rbcL, tufA, UPA, LSU and ITS as DNA barcode markers for the marine green macroalgae. Cryptogam. Algol. 2010, 31, 487. [Google Scholar]
- Saunders, G.W.; McDevit, D.C. Methods for DNA barcoding photosynthetic protists emphasizing the macroalgae and diatoms. Methods Mol. Biol. 2012, 858, 207–222. [Google Scholar] [PubMed]
- Vanormelingen, P.; Hegewald, E.; Braband, A.; Kitschke, M.; Friedl, T.; Sabbe, K.; Vyverman, W. The systematics of a small spineless desmodesmus species, d. costato-granulatus (sphaeropleales, chlorophyceae), based on ITS2 rDNA sequence analyses and cell wall morphology 1. J. Phycol. 2007, 43, 378–396. [Google Scholar] [CrossRef]
- Bock, C.; Paoutová, M.; Krienitz, L. Phylogenetic position of Coronastrum ellipsoideum and description of Parachlorella hussii sp. nov. Biologia 2011, 66, 585–594. [Google Scholar] [CrossRef]
- Bock, C.; Pröschold, T.; Krienitz, L. Updating the genus Dictyosphaerium and description of Mucidosphaerium gen. nov. (Trebouxiophyceae) based on morphological and molecular data. J. Phycol. 2011, 47, 638–652. [Google Scholar] [CrossRef]
- Rinkel, B.E.; Hayes, P.; Gueidan, C.; Brodie, J. A molecular phylogeny of Acrochaete and other endophytic green algae (Ulvales, Chlorophyta) 1. J. Phycol. 2012, 48, 1020–1027. [Google Scholar] [CrossRef]
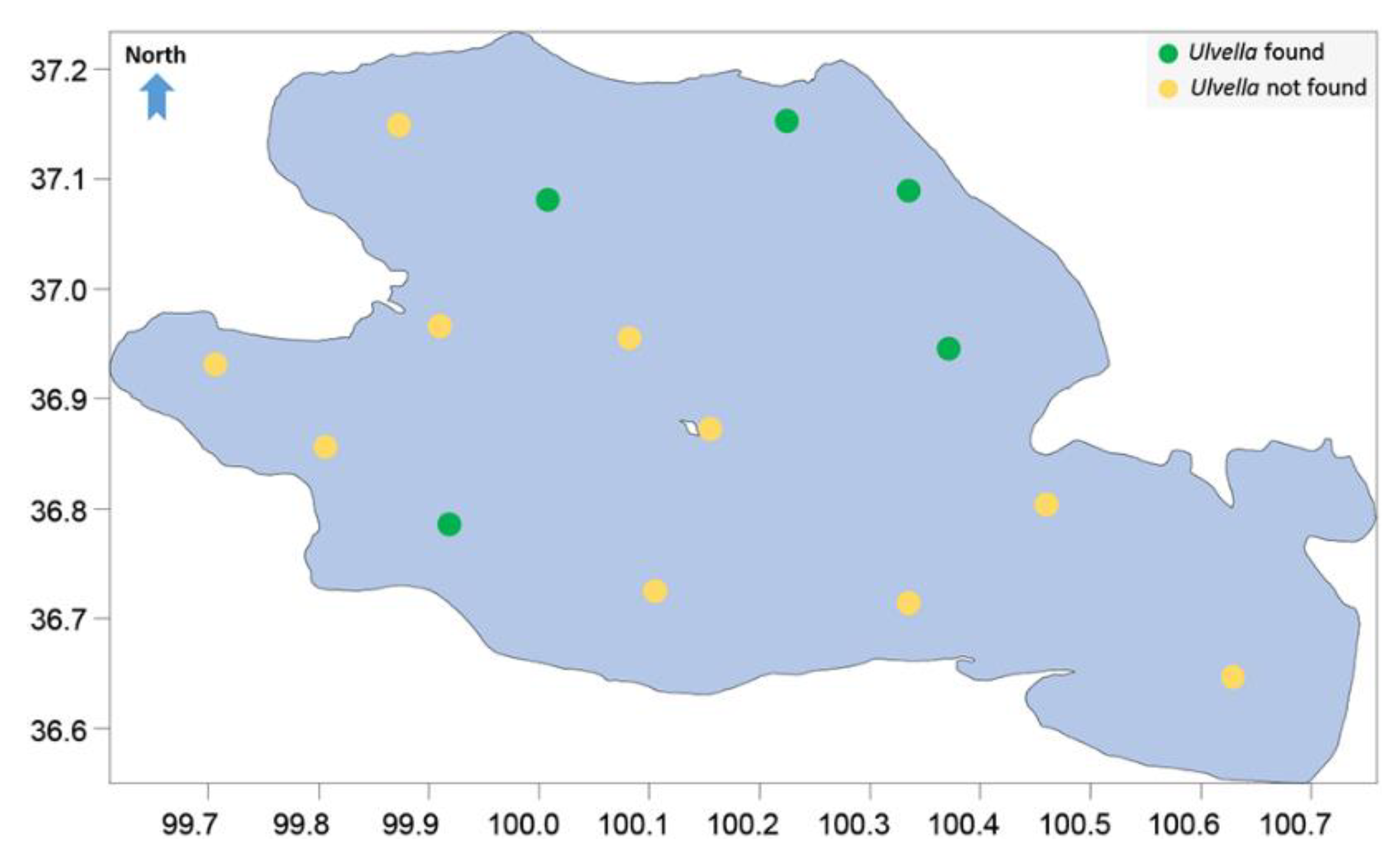
Figure 1.
The distribution of the sample points in Qinghai Lake. Average depth, 21 m.

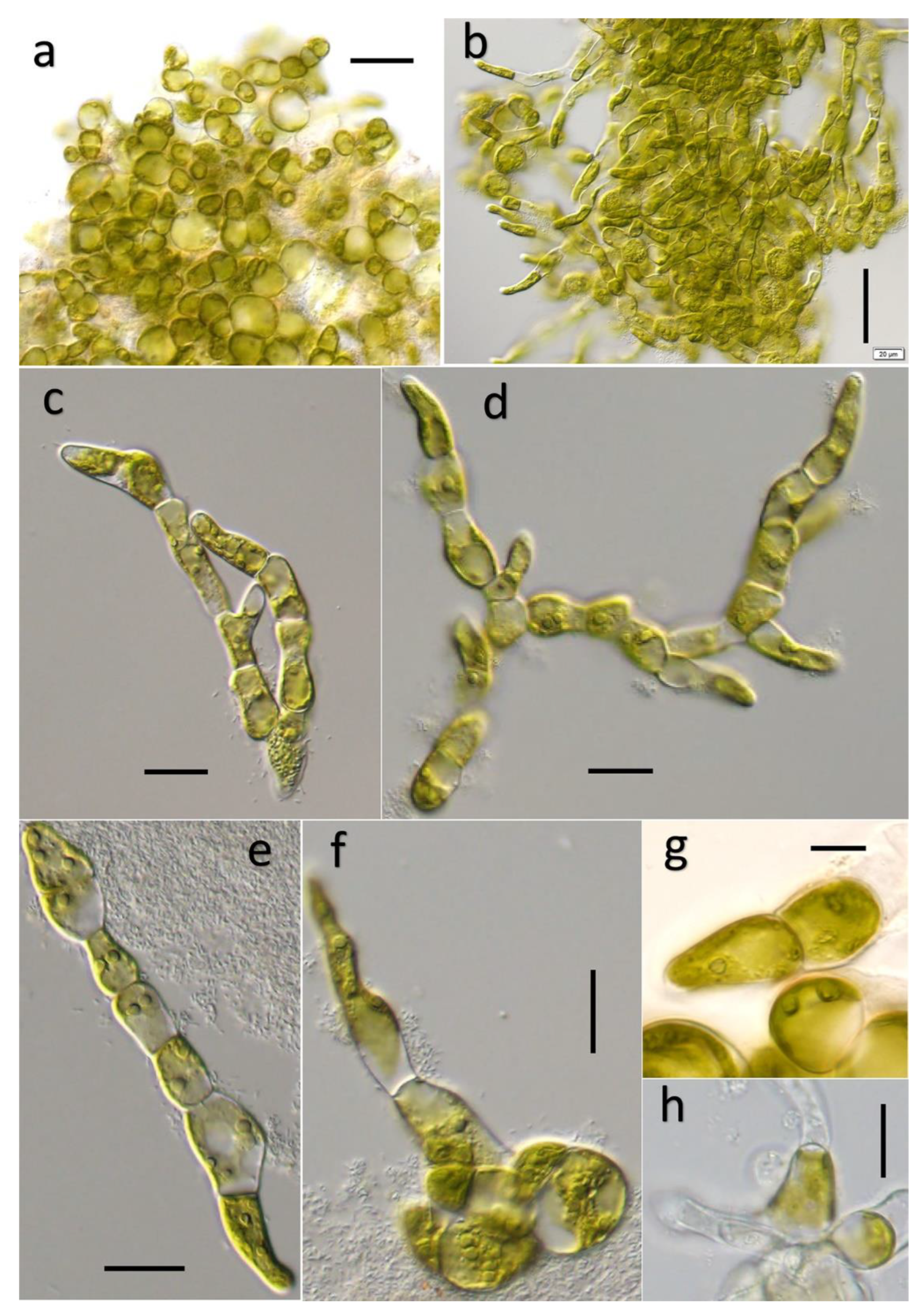
Figure 2.
Field sample morphology of Ulvella lacustris sp. nov.: (a) Habitat of Ulvella lacustris, growing on rough rock at the bottom of the lake; (b) Natural thallus on surface of rock under stereoscopy; (c) The cell arrangement was dense and the cell shape was severely compacted; (d) Single cell, with significantly thicker cell wall and integument; (e) The plant body is pseudoparenchyma-like, with thick integument and cell wall; (f) The color of young cells is green, and the gelatinous envelope is obvious; (g) The color of mature cells is yellow, and the gelatinous envelope is not obvious. Scale bar: 1 cm (a), 0.25 cm (b), 10 μm (c,f,g), 5 μm (d), 20 μm (e).
Figure 2.
Field sample morphology of Ulvella lacustris sp. nov.: (a) Habitat of Ulvella lacustris, growing on rough rock at the bottom of the lake; (b) Natural thallus on surface of rock under stereoscopy; (c) The cell arrangement was dense and the cell shape was severely compacted; (d) Single cell, with significantly thicker cell wall and integument; (e) The plant body is pseudoparenchyma-like, with thick integument and cell wall; (f) The color of young cells is green, and the gelatinous envelope is obvious; (g) The color of mature cells is yellow, and the gelatinous envelope is not obvious. Scale bar: 1 cm (a), 0.25 cm (b), 10 μm (c,f,g), 5 μm (d), 20 μm (e).

Figure 3.
Culture sample morphology of Ulvella lacustris sp. nov.: (a) Compact prostrate filaments; (b) Cultured filaments are staggered and arranged compactly; (c–e) Thallus irregular filamentous branches or unbranched branches; (f) New filaments germinating from akinetes; (g) Clearly visible pyrenoids and chloroplasts; (h) Suspected empty sporangium. Scale bar: 50 μm (a–b), 20 μm (c–f,h), 10 μm (g).
Figure 3.
Culture sample morphology of Ulvella lacustris sp. nov.: (a) Compact prostrate filaments; (b) Cultured filaments are staggered and arranged compactly; (c–e) Thallus irregular filamentous branches or unbranched branches; (f) New filaments germinating from akinetes; (g) Clearly visible pyrenoids and chloroplasts; (h) Suspected empty sporangium. Scale bar: 50 μm (a–b), 20 μm (c–f,h), 10 μm (g).

Figure 4.
Maximum likelihood phylogenetic tree of Chlorophyta based on 18S rDNA sequences. Maximum likelihood bootstrap values and Bayesian posterior probabilities are given on nodes. Values above 0.50 for BI and 50 for ML are given. The new species in this study are shaded in gray.
Figure 4.
Maximum likelihood phylogenetic tree of Chlorophyta based on 18S rDNA sequences. Maximum likelihood bootstrap values and Bayesian posterior probabilities are given on nodes. Values above 0.50 for BI and 50 for ML are given. The new species in this study are shaded in gray.

Figure 5.
Maximum likelihood phylogenetic tree of Ulvales based on tufA sequences. Maximum likelihood bootstrap values and Bayesian posterior probabilities are given on nodes. Values above 0.50 for BI and 50 for ML are given. The new species in this study are shaded in gray.
Figure 5.
Maximum likelihood phylogenetic tree of Ulvales based on tufA sequences. Maximum likelihood bootstrap values and Bayesian posterior probabilities are given on nodes. Values above 0.50 for BI and 50 for ML are given. The new species in this study are shaded in gray.

Figure 6.
(a). The phylogenetic tree reconstructed from ITS2 secondary structure and sequence. Maximum likelihood bootstrap values are given on nodes. Different species are circled with different colors as the background. (b). The ITS2 secondary structure among 8 strains in Ulvella. The darker the color, the more significant the variation.
Figure 6.
(a). The phylogenetic tree reconstructed from ITS2 secondary structure and sequence. Maximum likelihood bootstrap values are given on nodes. Different species are circled with different colors as the background. (b). The ITS2 secondary structure among 8 strains in Ulvella. The darker the color, the more significant the variation.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main habitat information and physical and chemical indicators of sampling sites. The algal strains of QHH-4, QHH-5, and QHH-15 were successfully cultured. DO: dissolved oxygen; EC: electric conductivity; TDS: total dissolved solid; SAL: salinity; SD: Secchi depth.
Table 1.
Main habitat information and physical and chemical indicators of sampling sites. The algal strains of QHH-4, QHH-5, and QHH-15 were successfully cultured. DO: dissolved oxygen; EC: electric conductivity; TDS: total dissolved solid; SAL: salinity; SD: Secchi depth.
| Latitude (°N) | Longitude (°E) | Altitude (m) | DO (mg/L) | pH | Temp (°C) | EC (ms/cm) | TDS | SAL (‰) | SD (cm) | Depth (m) | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| QHH-4 | 36.9598 | 100.3865 | 3196.10 | 7.48 | 9.12 | 13.4 | 17.55 | 9.98 | 10.4 | 400 | 22.4 |
| QHH-5 | 37.1322 | 100.2354 | 3196.25 | 7.99 | 9.13 | 11.7 | 17.67 | 10.04 | 10.45 | 370 | 17.6 |
| QHH-8 | 37.0248 | 100.0906 | 3196.09 | 7.63 | 9.19 | 12.2 | 17.42 | 10.01 | 10.43 | 455 | 28.1 |
| QHH-11 | 36.7487 | 99.8509 | 3196.18 | 6.77 | 9.1 | 16 | 17.22 | 9.76 | 10.08 | 350 | 14.6 |
| QHH-15 | 37.07059 | 100.3498 | 3196.27 | 7.42 | 9.12 | 13.1 | 17.39 | 10.05 | 10.46 | 370 | 16.5 |
Table 2.
Detailed information of alignment and nucleotide substitution in 18S and tufA concatenated phylogenies for ML analysis.
Table 2.
Detailed information of alignment and nucleotide substitution in 18S and tufA concatenated phylogenies for ML analysis.
| Dataset | 18S | tufA |
|---|---|---|
| Alignment length Number of sequences | 1655 64 | 855 58 |
| Parsimony-informative sites | 496 | 243 |
| Invariant sites | 968 | 542 |
| Best-fit model | TN93+G+I | T92+G+I |
| Base frequency (A/C/G/T) | 0.25/0.21/0.28/0.26 | 0.32/0.12/0.35/0.21 |
| Saturation test (Iss/Iss.c) | 0.123 < 0.836 | 0.080 < 0.815 |
Table 3.
CBCs and h-CBCs quantities with 8 algal sequences.
| U.lacustris OL630153 | U.heteroclada EF595353 | U.leptochaete EF595355 | U.repens EF595349 | U.viridis EF595351 | U.leptochaete MK910764 | U.leptochaete MK910760 | U.leptochaete MN510853 | |
|---|---|---|---|---|---|---|---|---|
| U.lacustris OL630153 | 0/0 | 2/2 | 1/3 | 2/3 | 0/2 | 4/2 | 4/2 | 1/2 |
| U.heteroclada EF595353 | 2/2 | 0/0 | 2/0 | 2/4 | 2/0 | 4/0 | 4/0 | 3/0 |
| U.leptochaete EF595355 | 1/3 | 2/0 | 0/0 | 1/1 | 0/0 | 0/0 | 0/0 | 1/0 |
| U.repens EF595349 | 2/3 | 2/4 | 1/1 | 0/0 | 1/1 | 3/1 | 3/1 | 2/1 |
| U.viridis EF595351 | 0/2 | 2/0 | 0/0 | 1/1 | 0/0 | 3/0 | 3/0 | 1/0 |
| U.leptochaete MK910764 | 4/2 | 4/0 | 0/0 | 3/1 | 3/0 | 0/0 | 0/0 | 1/0 |
| U.leptochaete MK910760 | 4/2 | 4/0 | 0/0 | 3/1 | 3/0 | 0/0 | 0/0 | 1/0 |
| U.leptochaete MN510853 | 1/2 | 3/0 | 1/0 | 2/1 | 1/0 | 1/0 | 1/0 | 0/0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yan, Q.; Dai, Q.; Liu, B.; Liu, G.; Zhu, H. A New Deep-Water Epilithic Green Alga, Ulvella lacustris, from an Alpine Brackish Lake in Qinghai–Tibet Plateau. Diversity 2022, 14, 594. https://doi.org/10.3390/d14080594
AMA Style
Yan Q, Dai Q, Liu B, Liu G, Zhu H. A New Deep-Water Epilithic Green Alga, Ulvella lacustris, from an Alpine Brackish Lake in Qinghai–Tibet Plateau. Diversity. 2022; 14(8):594. https://doi.org/10.3390/d14080594
Chicago/Turabian StyleYan, Qiufeng, Qingyu Dai, Benwen Liu, Guoxiang Liu, and Huan Zhu. 2022. "A New Deep-Water Epilithic Green Alga, Ulvella lacustris, from an Alpine Brackish Lake in Qinghai–Tibet Plateau" Diversity 14, no. 8: 594. https://doi.org/10.3390/d14080594
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.