

Two New Shellear Species (Gonorhynchiformes: Kneriidae), from the Luansa River (Upper Congo Basin): Hidden Diversity Revealed by Integrative Taxonomy

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Material and Methods
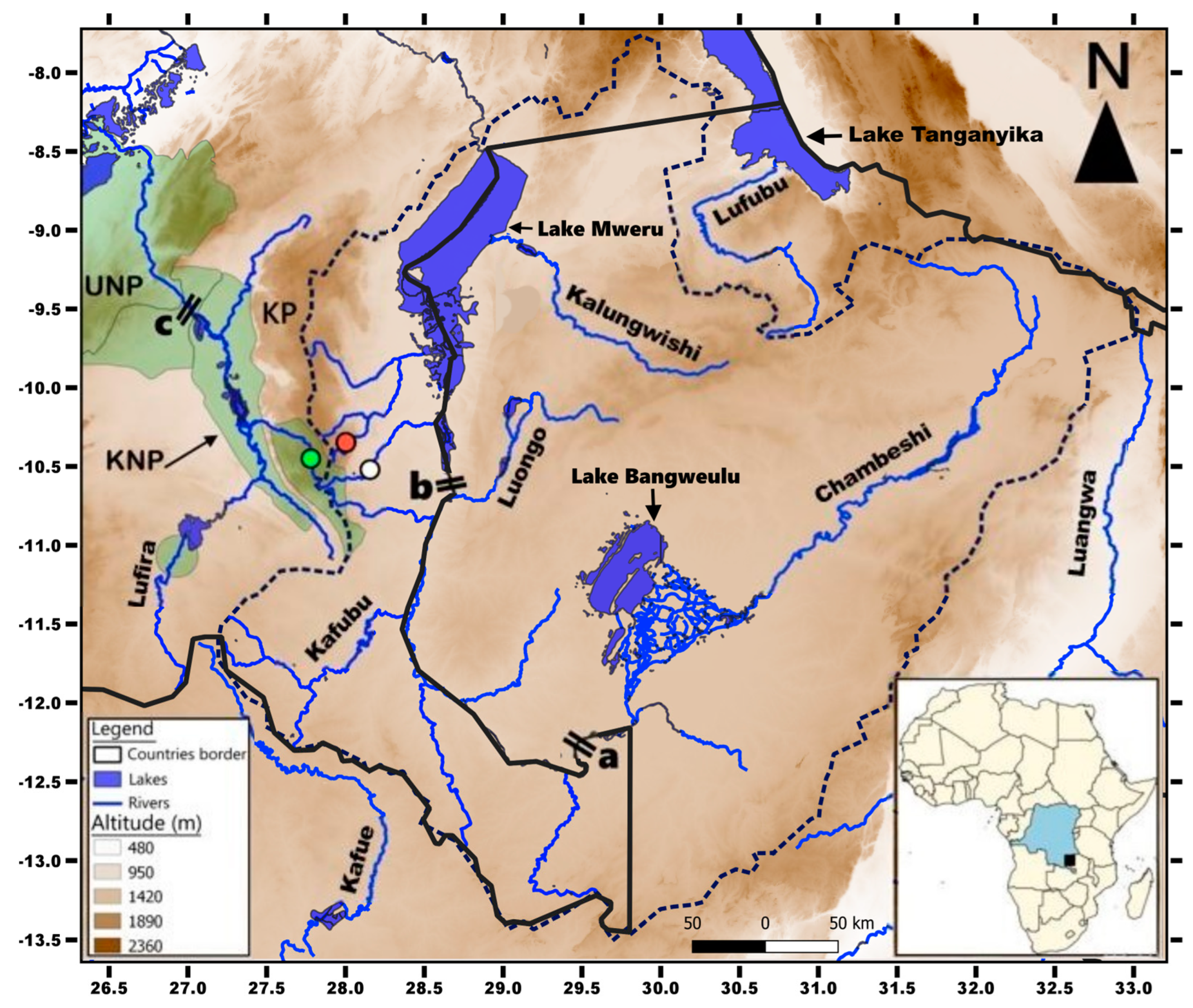
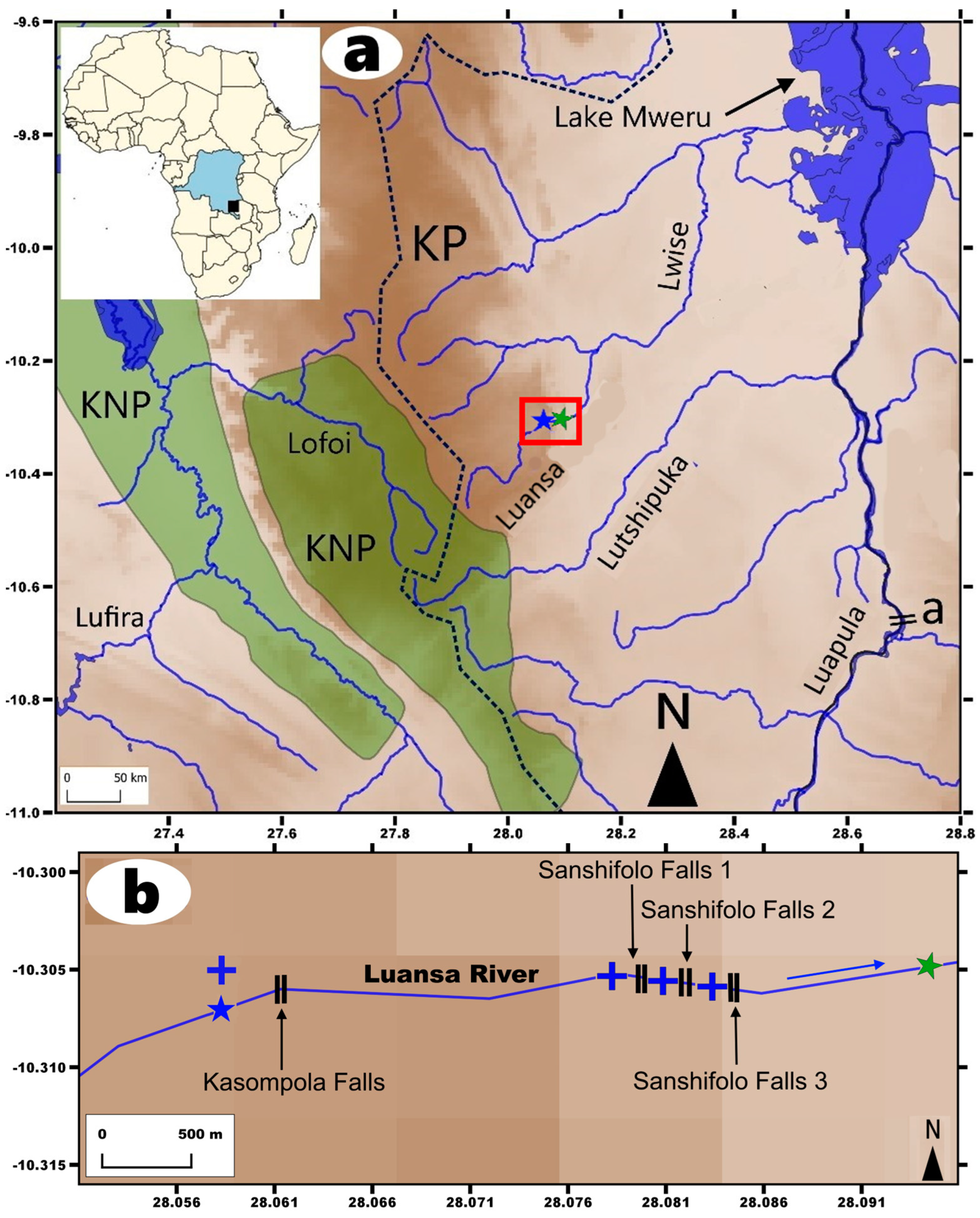
2.1. Sampling Methods
2.2. Morphological Approach
2.3. Genetic Approach
2.3.1. Taxonomic and Nucleotide Sampling
2.3.2. Molecular Methods
2.3.3. DNA–Sequence Edition and Phylogenetic Analysis
2.4. Nomenclatural Acts
3. Results
3.1. Morphological Explorations
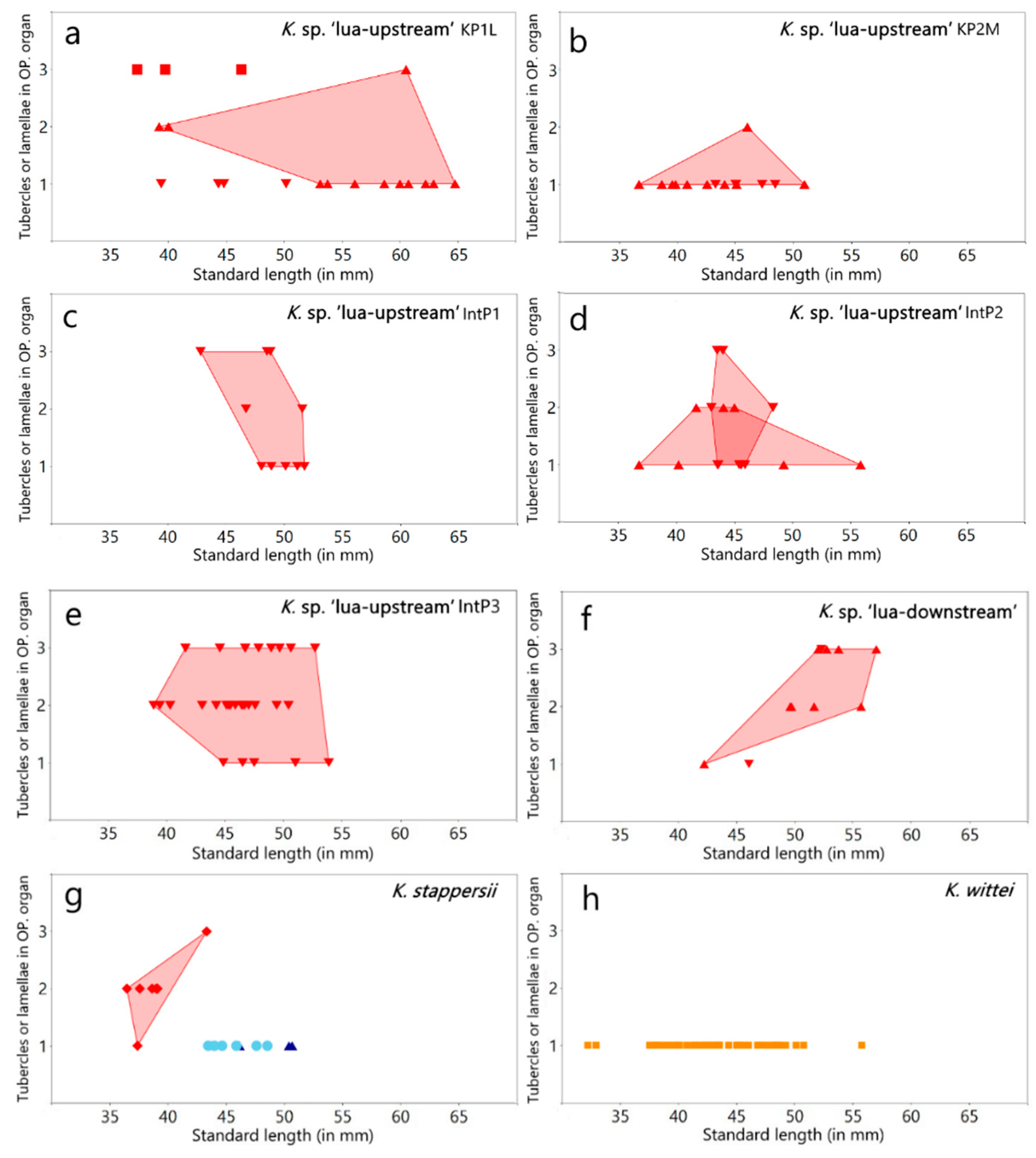
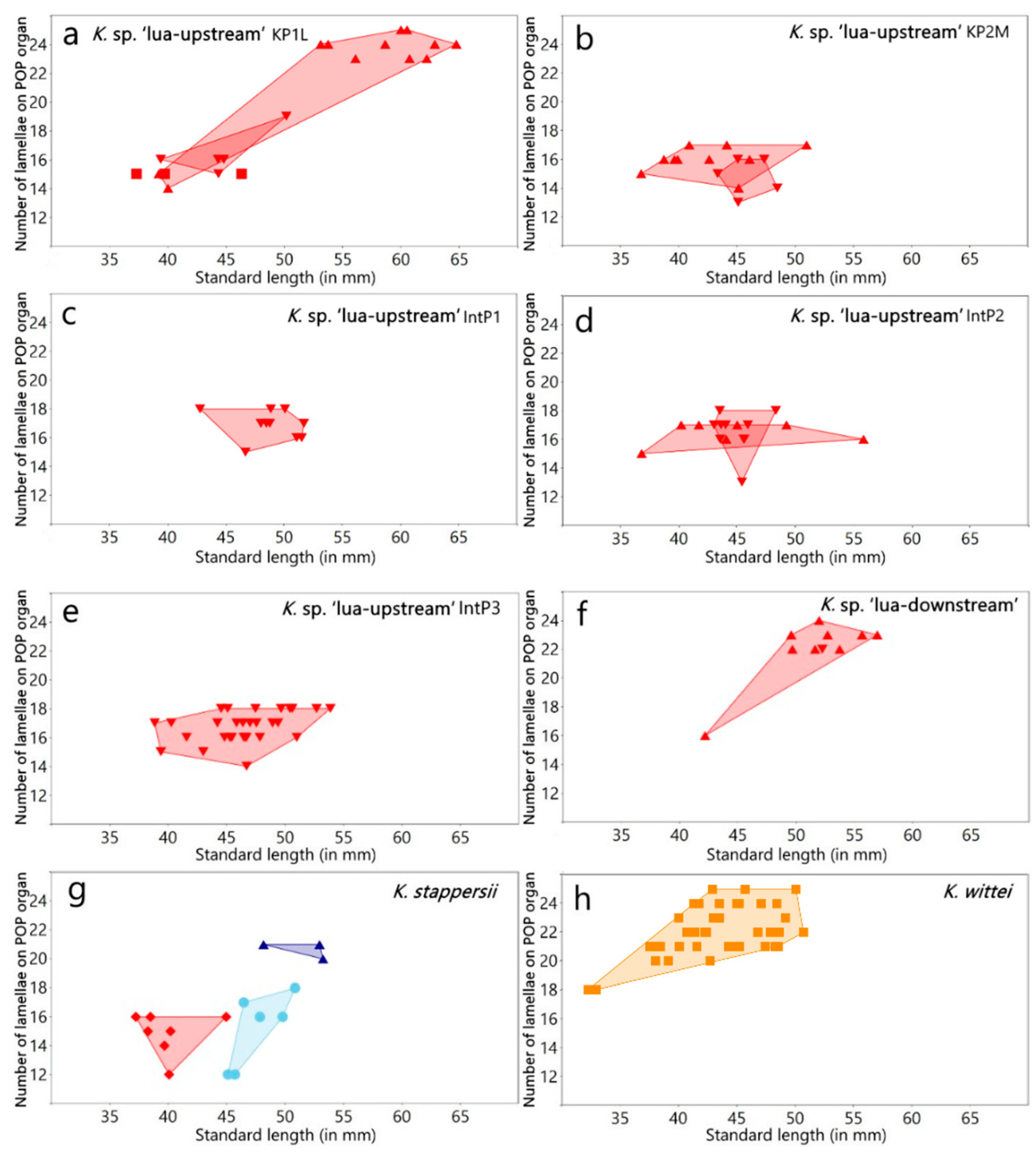
3.1.1. Variation in the Lamellar Structure of the Opercular (OP) and Postopercular (POP) Organ in Males
3.1.2. Qualitative Observations
Colouration and colour pattern variation within a single population
Colouration and colour pattern differences between (potential) species
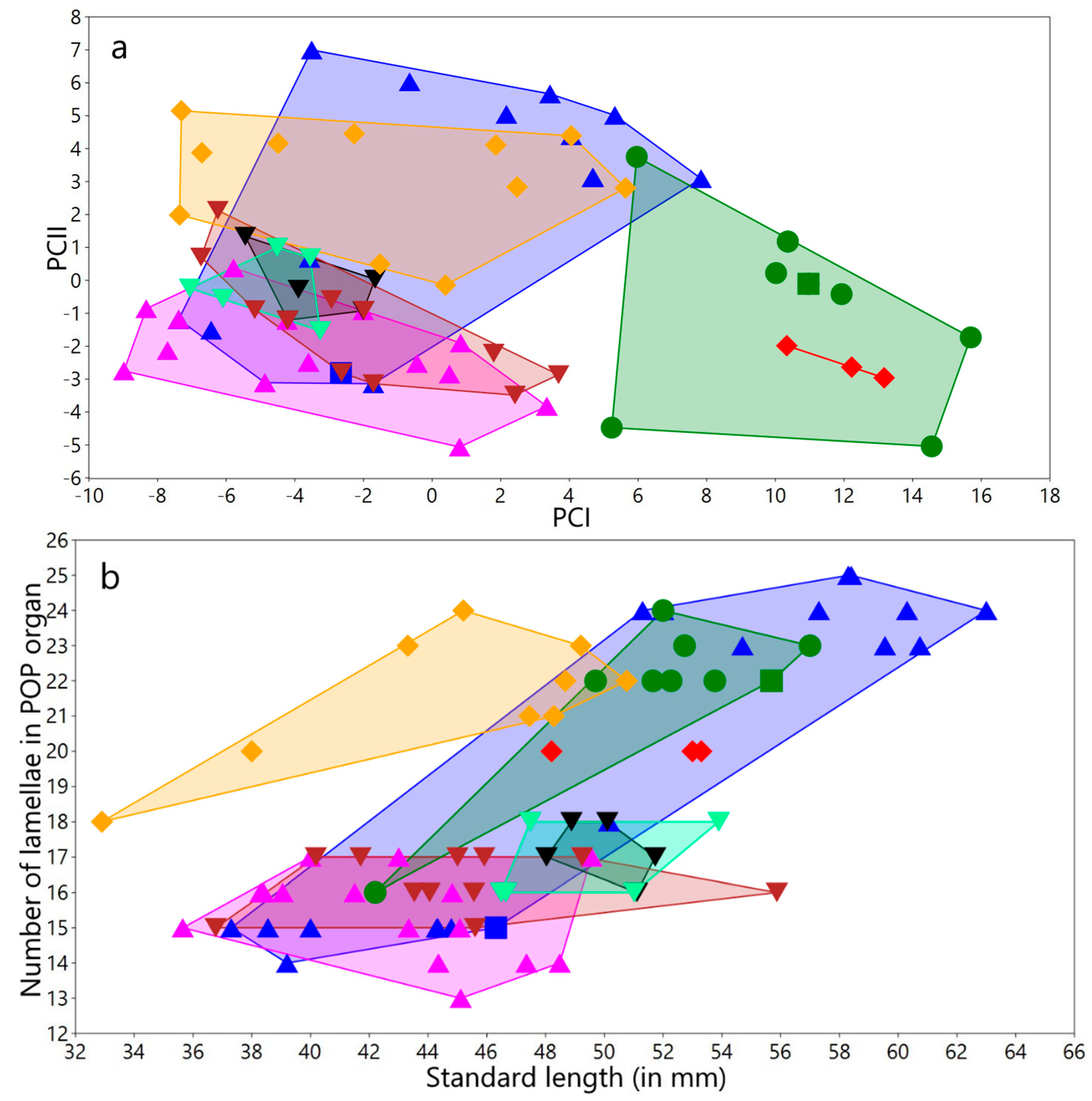
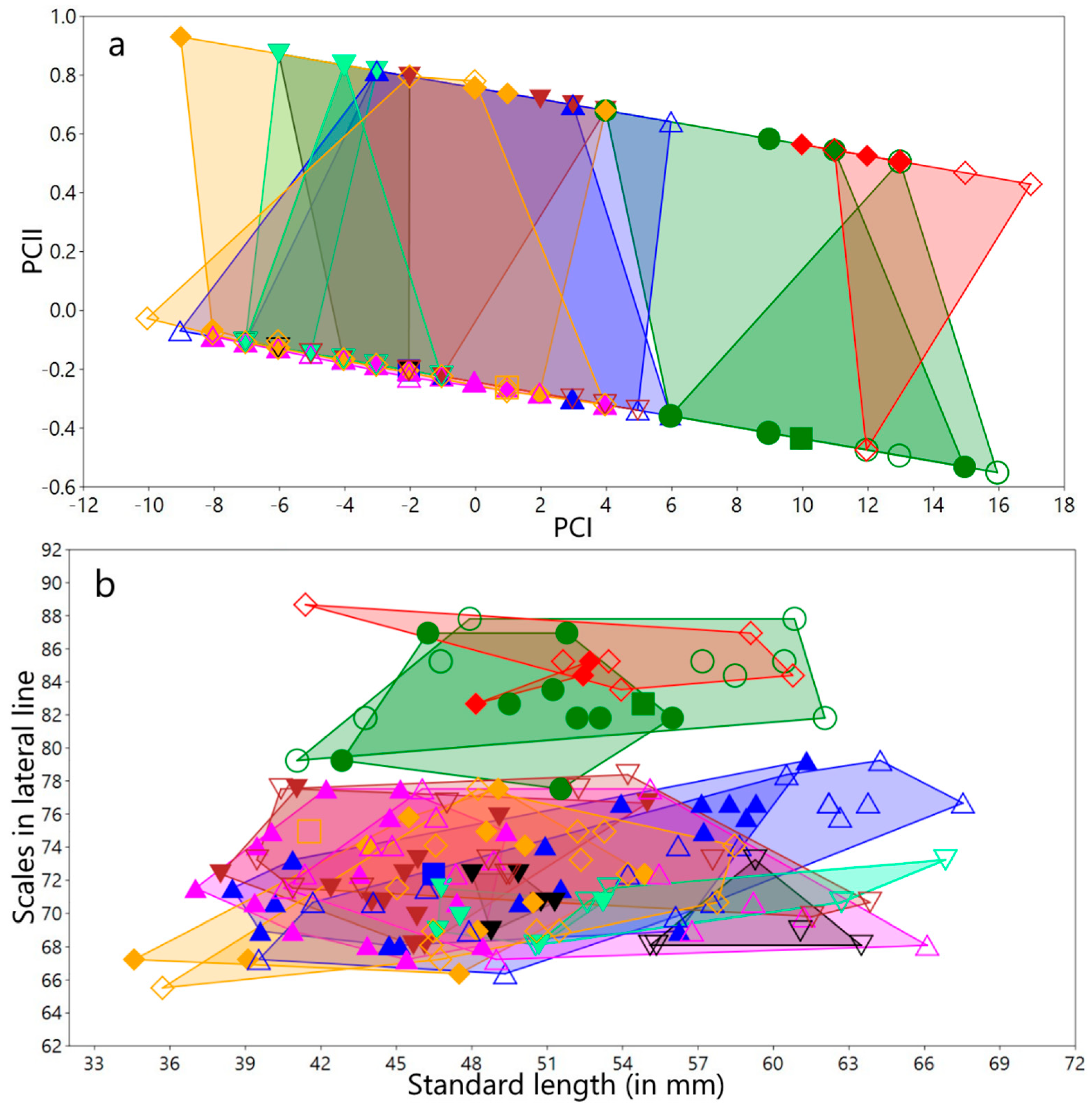
3.1.3. Meristic Variation
Meristic variation in females
Meristic variation in males
Meristic variation in both sexes and for all populations/species
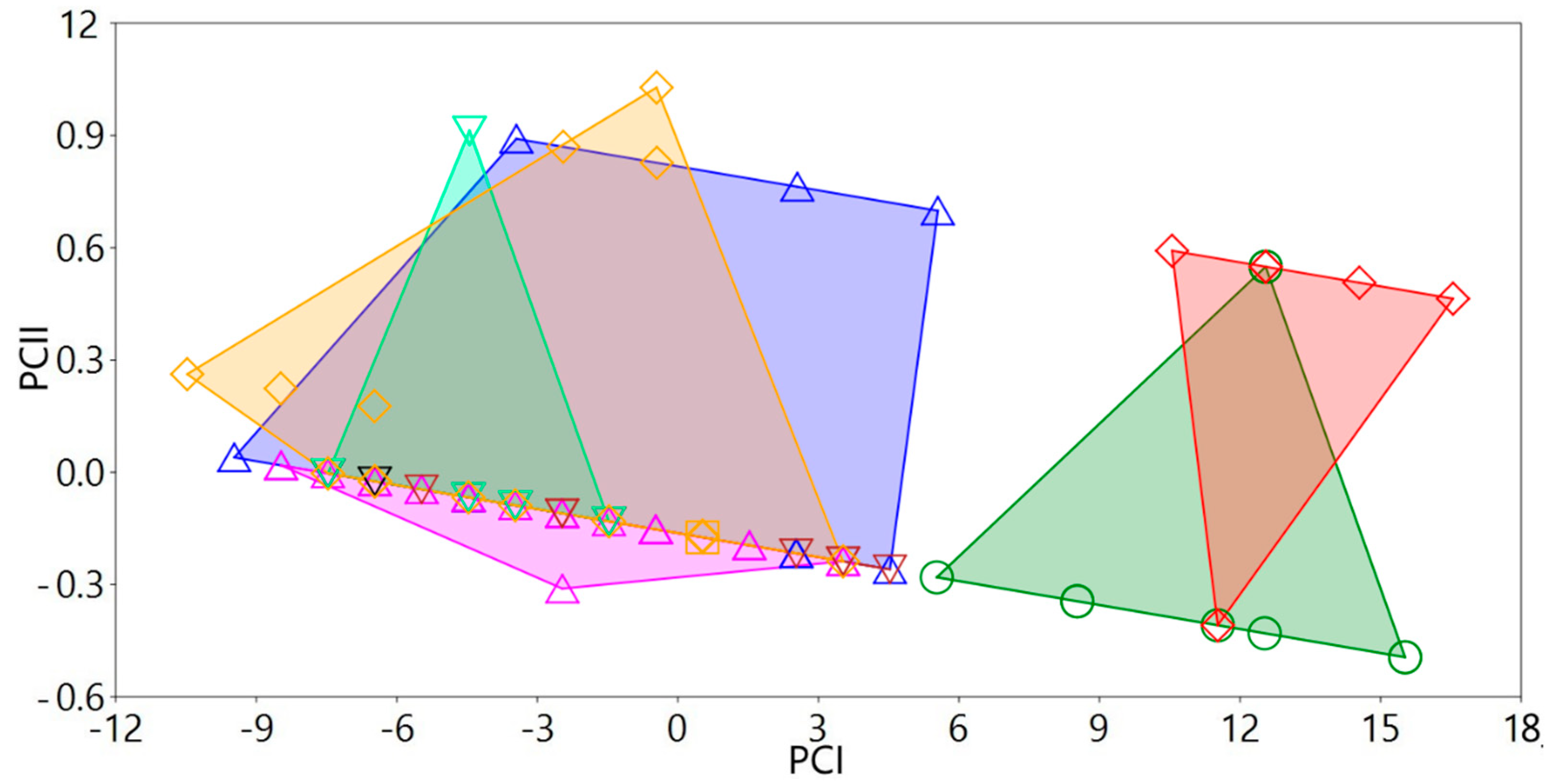
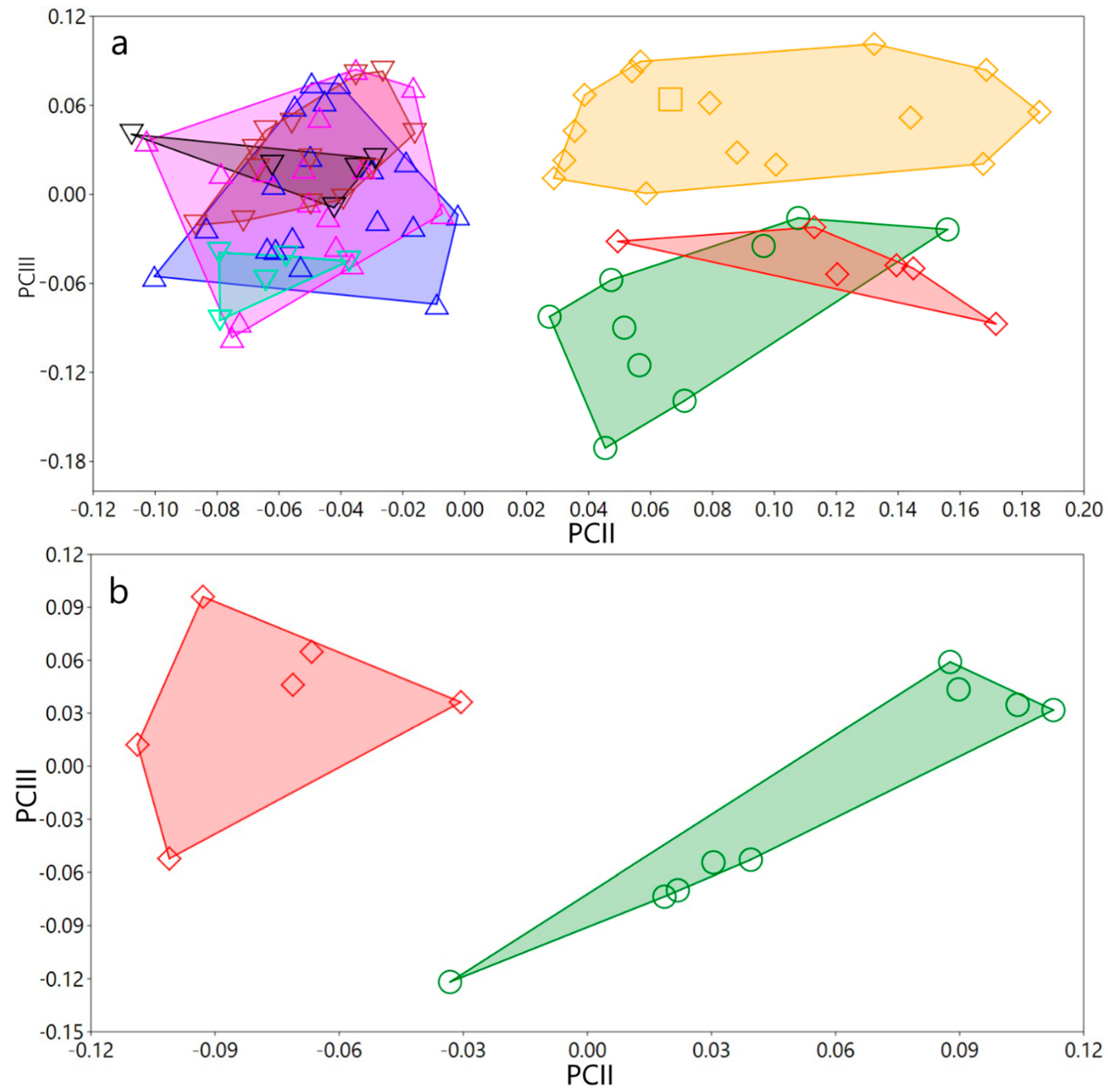
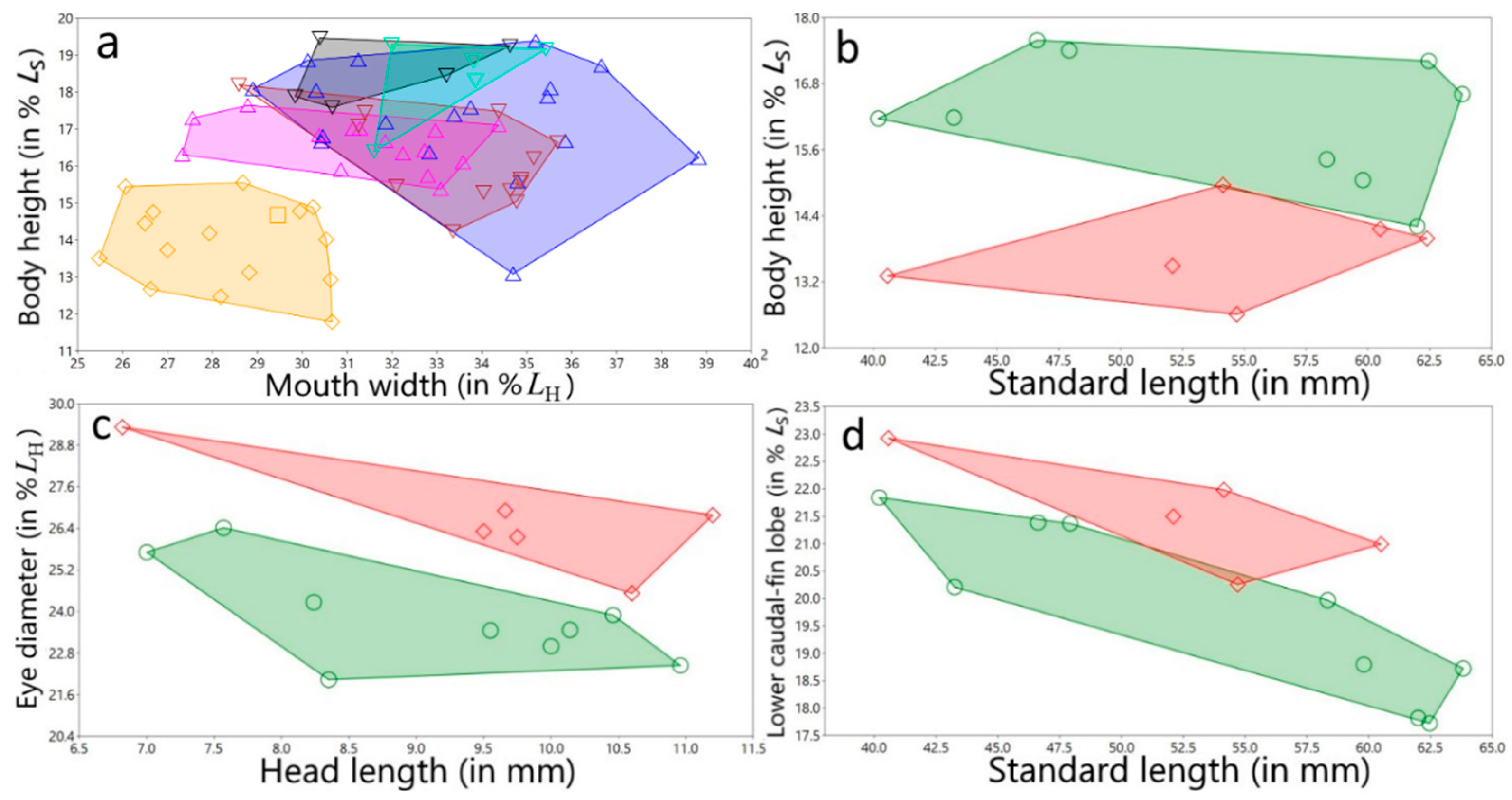
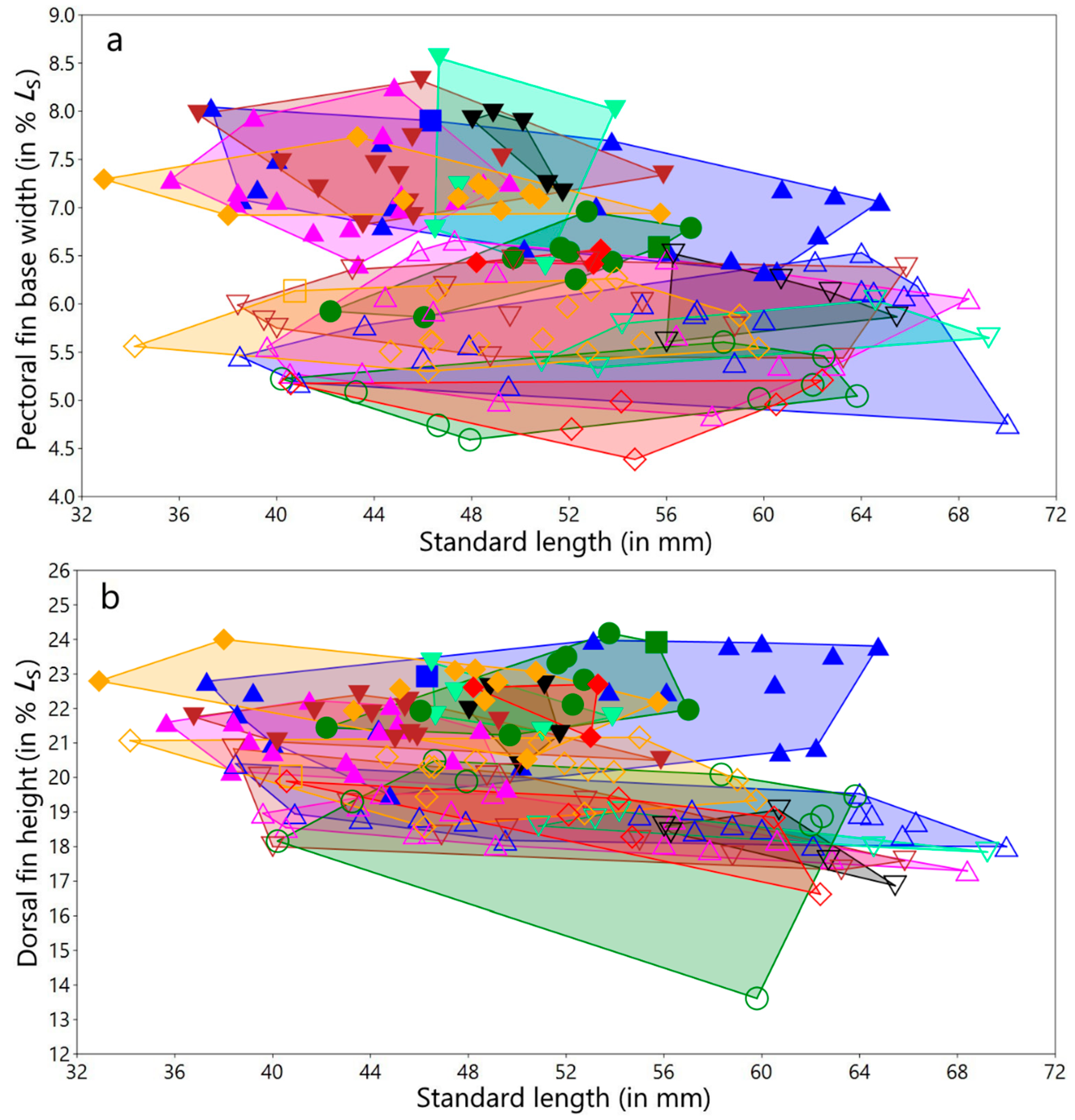
3.1.4. Variation in Measurements
Female variation in measurements
Male variation in measurements
Variation in measurements of both sexes and for all populations/species studied
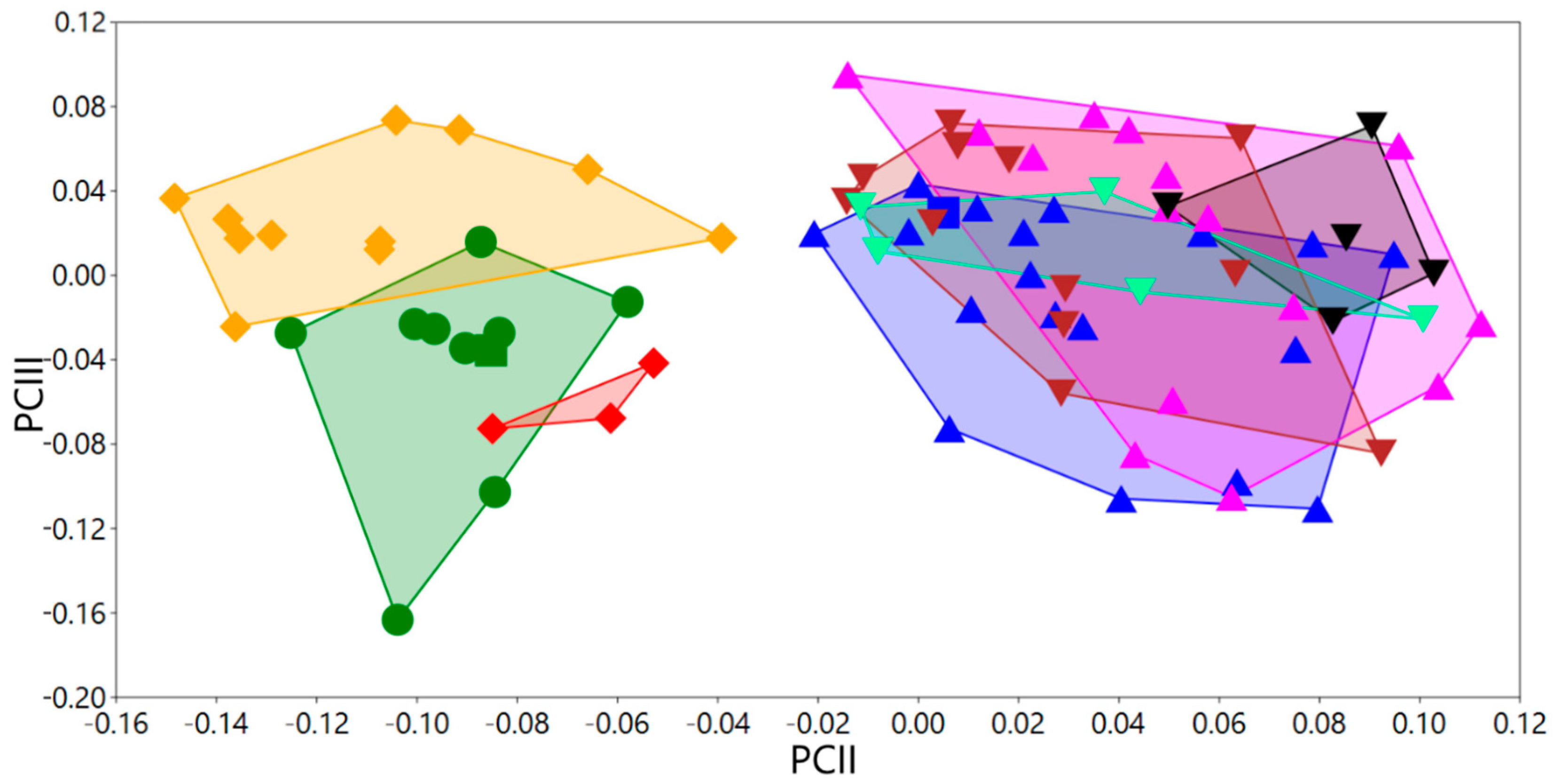
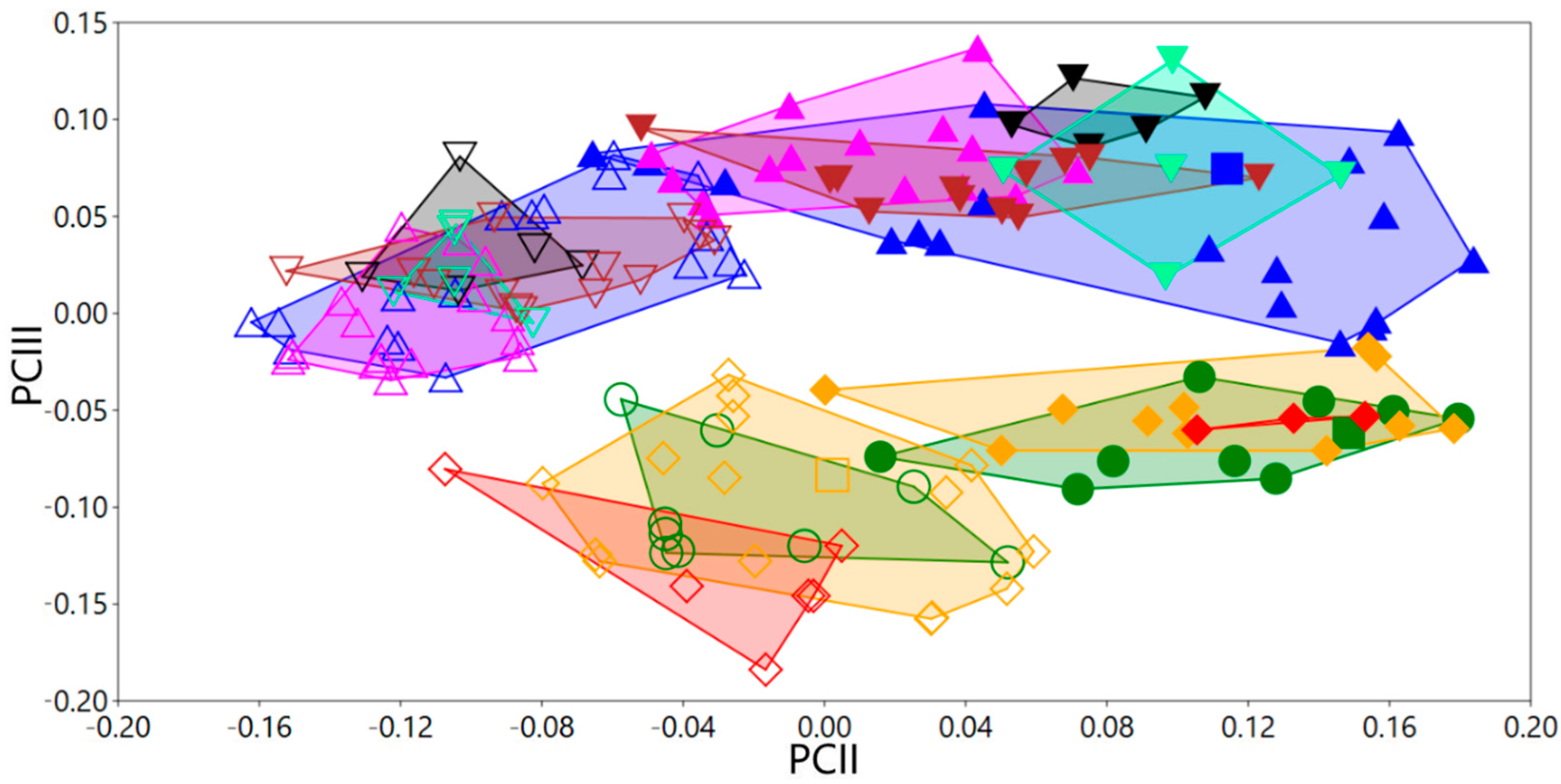
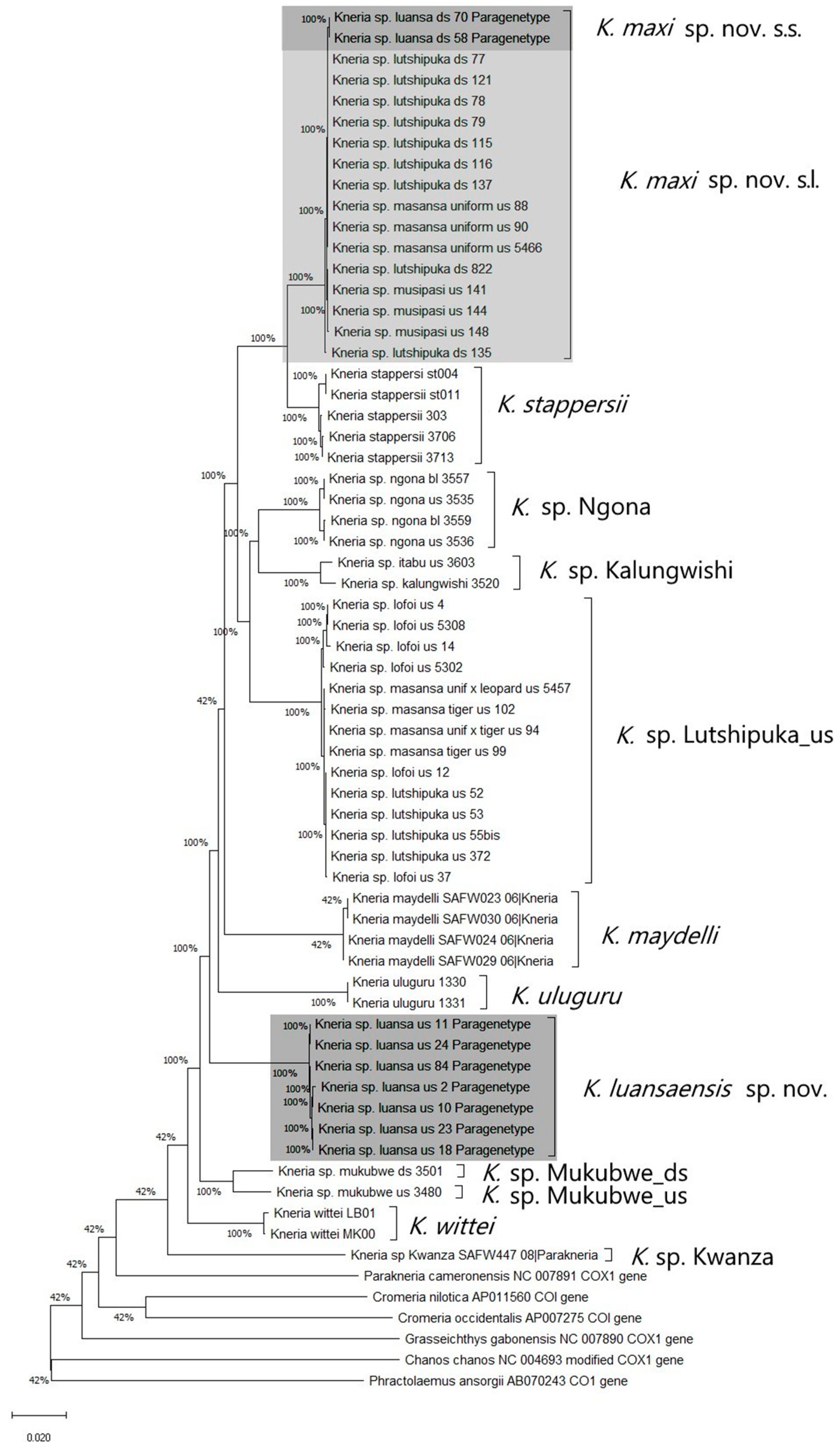
3.2. Phylogenetic Exploration: Mitochondrial COI DNA–Sequence Analyses
3.3. Taxonomic Decisions Based on the Integrative Approach
3.4. Species Descriptions
3.4.1. Kneria luansaensis sp. nov.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | K. luansaensis sp. nov. | K. maxi sp. nov. | K. wittei | K. stappersii | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| h (M) | Males (n = 56) | Females (n = 57) | h (M) | Males (n = 10) | Females (n = 9) | h (F) | Males (n = 8) | Females (n = 16) | Males (n = 3) | Females (n = 6) | |||||||||
| Range | m | Range | m | Range | m | Range | m | Range | m | Range | m | Range | m | Range | m | ||||
| Standard length LS (mm) | 46.3 | 35.6–63.0 | 47.1 | 35.0–70.0 | 53.7 | 55.7 | 42.2–57.0 | 51.3 | 40.2–63.8 | 53.8 | 40.7 | 32.9–55.8 | 46.4 | 34.2–59.8 | 48.2–53.3 | 51.5 | 40.6–62.4 | 54.1 | |
| Head length (LH) | 7.7 | 6.7–9.3 | 7.9 | 5.7–12.3 | 9.2 | 9.2 | 7.2–9.2 | 8.5 | 7.0–11.0 | 9.1 | 7.7 | 6.8–9.3 | 8.32 | 7.0–10.7 | 9.0 | 8.3–8.9 | 8.7 | 6.8–11.2 | 9.6 |
| Head measurements (%LH) | |||||||||||||||||||
| Post–orbital distance | 43.2 | 35.1–47.6 | 42.1 | 37.6–48.3 | 43.1 | 44.9 | 41.9–48.8 | 45.6 | 40.2–50.0 | 45.6 | 41.8 | 39.7–50.7 | 44.4 | 39.7–49.5 | 45.0 | 42.2–45.2 | 43.7 | 44.2–47.4 | 45.5 |
| Interorbital distance | 42.8 | 28.9–47.1 | 38.5 | 35.9–48.9 | 40.7 | 37.2 | 31.9–41.0 | 38.2 | 35.8–41.0 | 38.9 | 40.5 | 36.8–40.0 | 38 | 36.8 | 39.1 | 37.4–42.1 | 40.2 | 37.2–39.6 | 38.9 |
| Head height | 61.0 | 51.8–67.1 | 57.6 | 51.5–66.2 | 57.3 | 56.7 | 51.4–62.3 | 58.2 | 52.6–64.2 | 58.2 | 55.6 | 52.9–66.8 | 58.3 | 51.9–62.3 | 57.3 | 54.0–55.9 | 54.7 | 51.1–61.3 | 54.7 |
| Head width | 60.7 | 51.3–67.6 | 59.0 | 57.0–71.2 | 62.7 | 55.2 | 50.0–60.3 | 56.7 | 49.1–60.3 | 55.9 | 58.2 | 51.8–62.9 | 56.6 | 52.1–67.3 | 58.8 | 50.6–54.2 | 52.5 | 42.6–56.5 | 52.0 |
| Snout length | 29.5 | 26.2–38.6 | 32.2 | 26.3–37.7 | 32.5 | 29.2 | 27.5–29.9 | 28.9 | 24.8–31.6 | 28.6 | 28.4 | 23.7–32.9 | 28.9 | 24.3–34.2 | 29.2 | 29.0–30.1 | 29.7 | 29.3–35.2 | 31.6 |
| Mouth width | 36.2 | 27.1–40.0 | 34.1 | 27.0–38.8 | 32.7 | 29.0 | 24.1–30.8 | 28.5 | 23.5–30.0 | 27.7 | 29.4 | 26.4–31.3 | 28.5 | 25.4–30.6 | 28.3 | 27.1–27.7 | 27.4 | 22.6–28.6 | 26.5 |
| Eye diameter | 24.6 | 22.0–34.0 | 27.1 | 21.9–31.0 | 25.0 | 24.0 | 22.3–26.5 | 24.2 | 22.0–26.4 | 23.7 | 26.1 | 20.6–27.9 | 23.8 | 21.0–26.1 | 22.7 | 27.1–27.7 | 27.4 | 24.5–29.3 | 26.7 |
| Outer opercular organ diameter | 38.0 | 28.8–48.2 | 40.8 | − | − | 43.6 | 34.0–47.6 | 41.3 | − | − | 32.0–43.2 | 38.3 | − | − | 34.9–38.6 | 36.6 | − | − | |
| Inner opercular organ diameter | 22.0 | 16.1–23.7 | 20.5 | − | − | 19.6 | 18.3–21.9 | 20.2 | − | − | 16.2–25.0 | 21.1 | − | − | 19.8–22.9 | 21.4 | − | − | |
| Body measurements (%LS) | |||||||||||||||||||
| Head length | 16.7 | 15.4–19.9 | 17.1 | 15.3–19.7 | 17.5 | 16.5 | 15.4–18.2 | 16.6 | 15.4–17.9 | 17.1 | 18.8 | 16.4–20.7 | 17.9 | 17.1–20.3 | 18.3 | 16.5–17.2 | 16.8 | 16.8–18.2 | 17.7 |
| Postopercular organ length | 11.5 | 9.1–13.2 | 11.5 | − | − | 10.8 | 10.7–12.7 | 11.4 | − | − | 10.2–13.7 | 12.4 | − | − | 10.4–11.3 | 11.0 | |||
| Pre–dorsal distance | 49.3 | 45.4–51.4 | 48.6 | 48.1–54.1 | 50.2 | 49.0 | 48.2–50.3 | 49.0 | 50.4–53.5 | 51.4 | 50.8 | 47.2–50.2 | 48.8 | 48.4–50.8 | 49.6 | 48.5–49.8 | 49.3 | 50.4–53.8 | 51.6 |
| Pectoral–pelvic distance | 34.7 | 32.3–37.7 | 34.5 | 31.5–37.9 | 34.4 | 30.6 | 30.6–34.7 | 32.1 | 30.4–34.5 | 32.2 | 34.1 | 31.1–33.9 | 33.0 | 31.1–34.1 | 32.8 | 31.5–32.5 | 32.0 | 31.4–35.0 | 32.3 |
| Pectoral–anal distance | 57.7 | 54.2–62.1 | 58.5 | 54.5–61.8 | 58.4 | 56.5 | 55.6–59.2 | 56.9 | 54.7–59.1 | 57.2 | 55.7 | 54.8–57.7 | 56.0 | 54.5–57.4 | 56.1 | 55.4–57.0 | 56.4 | 53.8–58.7 | 56.3 |
| Post–dorsal distance | 52.3 | 49.8–54.7 | 52.5 | 47.8–52.9 | 51.1 | 52.8 | 50.2–52.8 | 51.3 | 48.5–51.0 | 49.6 | 49.1 | 50.0–53.5 | 51.6 | 49.1–52.4 | 51.1 | 50.2–52.3 | 51.3 | 48.1–50.7 | 49.3 |
| Pre–pelvic distance | 49.3 | 45.5–51.7 | 48.4 | 46.8–52.0 | 49.4 | 46.6 | 45.7–50.2 | 47.2 | 45.3–49.4 | 47.9 | 51.6 | 48.1–50.3 | 49.1 | 47.1–51.6 | 49.4 | 46.3–47.3 | 46.9 | 46.5–50.5 | 48.2 |
| Pre-anal distance | 72.0 | 68.1–75.0 | 71.6 | 70.2–75.7 | 72.5 | 71.9 | 71.2–73.5 | 72.0 | 70.5–73.4 | 72.0 | 72.9 | 69.8–72.6 | 71.1 | 70.9–72.9 | 71.9 | 70.8–72.8 | 71.6 | 71.5–73.3 | 72.2 |
| Pre–pectoral distance | 15.5 | 13.1–18.2 | 14.9 | 14.54–18.1 | 16.0 | 14.5 | 13.6–16.9 | 14.9 | 14.4–16.3 | 15.5 | 18.9 | 14.5–17.7 | 16.0 | 15.5–18.9 | 17.0 | 15.1–16.1 | 15.5 | 16.2–18.5 | 17.1 |
| Body height | 15.7 | 13.4–19.2 | 16.0 | 13.1–19.6 | 17.1 | 14.7 | 14.7–16.8 | 15.3 | 14.2–17.6 | 16.2 | 14.7 | 13.0–15.7 | 14.4 | 11.8–15.6 | 14.0 | 13.7–15.4 | 14.6 | 12.6–15.0 | 13.8 |
| Body width | 10.2 | 9.7–12.3 | 10.6 | 9.9–12.0 | 10.7 | 8.0 | 8.0–9.0 | 8.8 | 8.7–9.1 | 8.8 | 10.6 | 9.6–10.8 | 10.1 | 9.7–11.7 | 10.4 | 8.4–9.2 | 8.9 | 8.6–9.1 | 8.9 |
| Caudal peduncle height | 9.9 | 8.1–11.7 | 9.8 | 8.1–12.1 | 9.6 | 10.9 | 9.5–10.9 | 10.2 | 8.8–10.4 | 9.6 | 10.1 | 9.5–12.0 | 10.8 | 9.2–11.5 | 10.2 | 10.3–10.9 | 10.6 | 9.2–10.6 | 10.0 |
| Caudal peduncle length | 22.5 | 18.7–22.5 | 20.9 | 18.2–22.9 | 20.2 | 19.03 | 18.3–20.2 | 19.3 | 18.2–21.4 | 19.2 | 18.4 | 18.2–20.6 | 19.4 | 18.2–20.7 | 19.7 | 20.8–21.5 | 21.0 | 19.6–22.1 | 20.6 |
| Dorsal fin height | 22.9 | 19.5–24.0 | 21.7 | 16.9–20.8 | 18.7 | 23.9 | 21.2–24.2 | 22.6 | 13.6–20.5 | 18.7 | 20.0 | 20.5–24.0 | 22.6 | 18.6–21.2 | 20.1 | 21.2–22.7 | 22.2 | 16.6–19.9 | 18.7 |
| Pelvic fin length | 21.0 | 15.5–21.0 | 18.5 | 15.3–19.1 | 17.0 | 17.7 | 15.8–18.9 | 17.7 | 14.8–19.1 | 17.1 | 18.7 | 17.9–20.9 | 19.1 | 16.1–20.5 | 18.3 | 18.1–18.8 | 18.5 | 15.4–18.1 | 16.9 |
| Pectoral fin length | 16.5 | 15.7–21.1 | 18.0 | 15.7–21.8 | 18.9 | 16.2 | 13.3–18.2 | 15.9 | 14.4–20.1 | 17.4 | 20.9 | 15.9–20.2 | 18.1 | 17.7–20.9 | 19.0 | 16.0–17.2 | 16.6 | 16.2–18.0 | 17.3 |
| Upper caudal fin lobe | 22.3 | 17.2–3.2 | 20.9 | 16.8–22.0 | 19.3 | 19.6 | 18.6–22.5 | 20.2 | 17.7–21.6 | 19.7 | − | 18.3–23.0 | 21.0 | 15.1–21.7 | 19.3 | 20.8–21.4 | 21.0 | 14.3–21.2 | 19.3 |
| Lower caudal fin lobe | 24.8 | 19.6–26.0 | 22.8 | 14.2–22.0 | 18.7 | 20.1 | 19.7–22.3 | 21.1 | 17.7–21.8 | 19.8 | − | 22.1–24.8 | 23.1 | 17.6–21.8 | 19.7 | 24.3–24.9 | 24.6 | 20.3–22.9 | 21.5 |
| Dorsal fin base width | 9.7 | 7.7–10.2 | 9.0 | 7.3–9.4 | 8.4 | 9.9 | 8.5–10.2 | 9.4 | 8.2–9.8 | 8.8 | 9.8 | 9.2–11.6 | 10.4 | 8.5–10.4 | 9.4 | 9.4–9.6 | 9.5 | 7.9–8.7 | 8.3 |
| Pelvic fin base width | 2.9 | 2.6–3.5 | 3.1 | 2.5–3.7 | 3.1 | 3.6 | 3.3–3.7 | 3.6 | 3.1–3.6 | 3.4 | 3.1 | 3.0–3.7 | 3.3 | 2.9–3.7 | 3.44 | 3.3–3.4 | 3.4 | 3.1–3.7 | 3.5 |
| Pectoral fin base width | 7.9 | 6.3–8.6 | 7.3 | 4.8–6.7 | 5.8 | 6.6 | 5.9–7.0 | 6.5 | 4.6–5.6 | 5.1 | 6.1 | 6.9–7.7 | 7.2 | 5.3–6.3 | 5.8 | 6.9–7.0 | 7.0 | 4.4–5.2 | 4.9 |
| Anal fin base width | 6.9 | 5.8–9.6 | 7.7 | 6.0–8.6 | 7.1 | 7.4 | 6.8–8.8 | 7.7 | 6.10–8.06 | 6.8 | 8.6 | 7.7–8.7 | 8.2 | 6.3–8.8 | 8.0 | 7.4–7.9 | 7.7 | 7.2–8.4 | 7.7 |
| Pelvic-anal distance | 24.2 | 22.1–28.2 | 25.0 | 21.1–27.3 | 24.9 | 24.6 | 23.9–26.0 | 25.0 | 22.6–27.9 | 25.3 | 22.1 | 22.1–24.5 | 23.0 | 22.1–25.1 | 23.4 | 25.2–26.3 | 25.7 | 24.0–26.5 | 25.2 |
| Anal–caudal distance | 30.5 | 26.2–31.8 | 29.5 | 26.4–30.9 | 28.3 | 28.7 | 26.8–29.2 | 28.5 | 27.2–29.0 | 28.1 | 26.8 | 27.1–30.8 | 29.0 | 26.8–29.8 | 28.4 | 28.8–30.2 | 29.5 | 27.3–29.9 | 28.7 |
| Meristics | h | range | me | range | me | h | range | me | range | me | h | range | me | range | me | range | me | range | me |
| Simple dorsal fin rays | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | − | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Branched dorsal fin rays | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 | − | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| Simple caudal fin | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Branched caudal fin rays | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 | 17 |
| Simple anal fin rays | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Branched anal fin rays | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 | − | 6 | 6 | 5.0–6 | 6 | 6 | 6 | 6 | 6 |
| Total pelvic fin rays | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 7.0–8 | 8 | 7.0–8 | 8 | 8 | 8 | 8 | 8 |
| Total pelvic fin rays | 15 | 15–16 | 15 | 15–16 | 15 | 15 | 15–16 | 15 | 15–16 | 15 | 15 | 16 | 16 | 15–16 | 15 | 16 | 16 | 15–16 | 16 |
| Lateral line scales | 72 | 66–80 | 71 | 65–80 | 72 | 84 | 78–89 | 83.5 | 80–90 | 87 | 75 | 61–78 | 72 | 64–78 | 71.5 | 84–87 | 86 | 85–91 | 87 |
| Lamellae in the postopercular organ | 15 | 13–25 | 16 | − | − | 22 | 22–24 | 22 | − | − | − | 18–24 | 22 | − | − | 20 | 20 | − | − |
3.4.2. Kneria maxi sp. nov.


4. Discussion
4.1. The External Morphological Characterisation of Sexual Maturity
4.1.1. The Opercular Organ
4.1.2. The Postopercular Organ
4.1.3. Body Tubercles on Males
4.1.4. Sexual Dichromatism
4.2. Sexual Dimorphism beyond the Presence/Absence of the Opercular and Postopercular Organ
4.3. Species Diversity in Kneria: A Concise Overview of an Underestimated Phenomenon
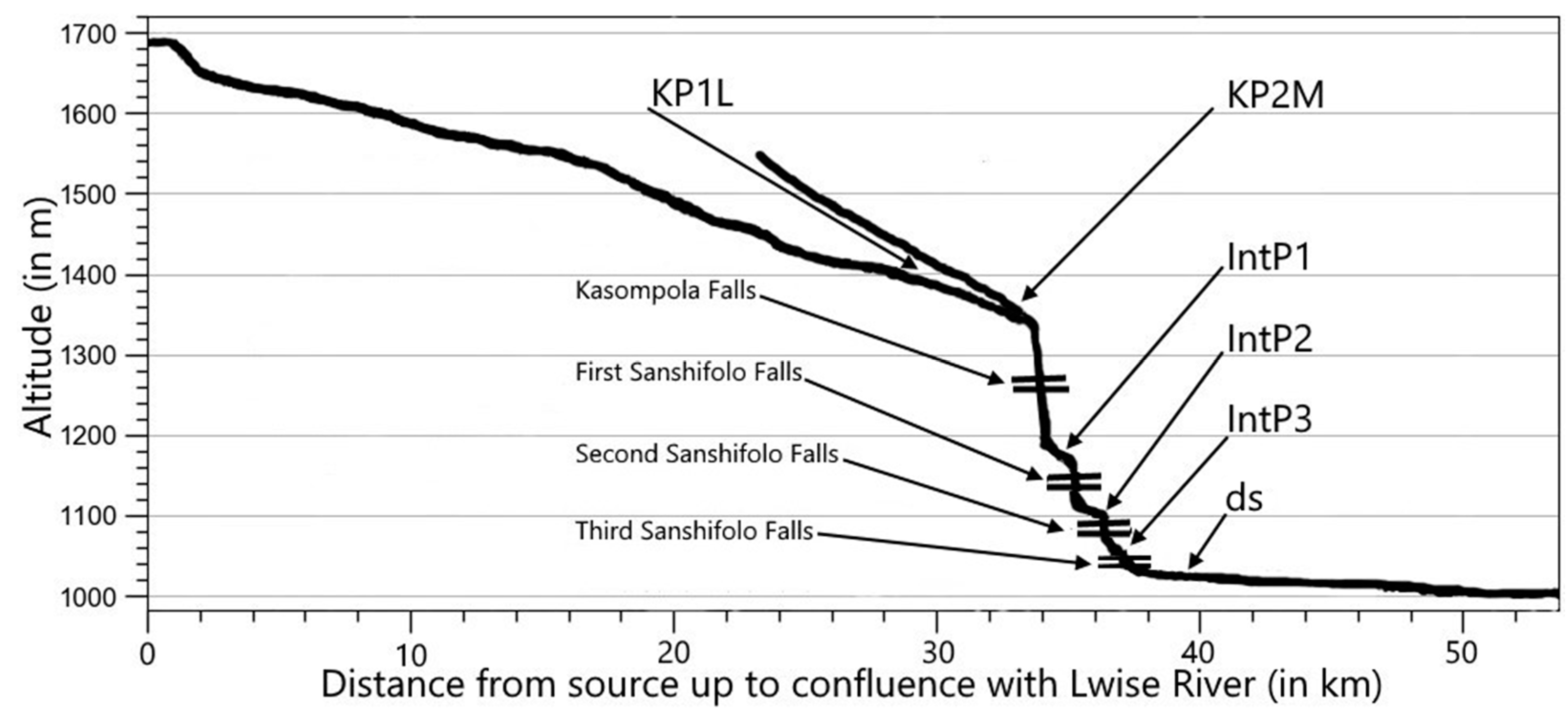
4.4. On the Possible Influence of Waterfalls on the Species Diversity in Kneria
4.5. Protection of the Fishes of the Luansa River, the KP, and Its Surroundings
5. Comparative Material Examined
5.1. Kneria ansorgii
5.2. Kneria auriculata
5.3. Kneria katangae
5.4. Kneria paucisquamata
5.5. Kneria polli
5.6. Kneria stappersii
5.7. Kneria wittei
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Poll, M. Nombre et distribution géographique des poissons d’eau douce africains. Bull. Muséum Natl. D’histoire Nat. 1973, 150, 113–128. [Google Scholar]
- Poll, M. Kneriidae (129–133), Cromeriidae (p. 134), Grasseichthyidae (p. 135). In Check List of the Freshwater Fishes of Africa; Daget, J., Gosse, J.P., van den Audenaerde, D.F.E.T., Eds.; Royal Museum for Central Africa: Tervuren, Belgium, 1984; Volume I, p. 410. [Google Scholar]
- Lavoué, S.; Miya, M.; Inoue, J.G.; Saitoh, K.; Ishiguro, N.B.; Nishida, M. Molecular systematics of the gonorynchiform fishes (Teleostei) based on whole mitogenome sequences: Implications for higher-level relationships within the Otocephala. Mol. Phylogenetics E 2005, 37, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Poyato-Ariza, F.J.; Grande, T.; Diogo, R. Gonorynchiform interrelationships: Historic overview, analysis, and revised systematics of the group. In Origin and Phylogenetic Interrelationships of Teleosts. Honoring Gloria Arratia; Nelson, J.S., Wilson, H.P., Schultze, M.V.H., Eds.; CRC Press: München, Germany, 2010; pp. 269–293. [Google Scholar]
- Berra, T.M. Freshwater Fish Distribution; University of Chicago Press: Chicago, IL, USA, 2007. [Google Scholar]
- Britz, R.; Moritz, T. Reinvestigation of the osteology of the miniature African freshwater fishes Cromeria and Grasseichthys (Teleostei, Gonorynchiformes, Kneriidae), with comments on kneriid relationships. Zoosystematics Evol. 2007, 83, 3–42. [Google Scholar] [CrossRef]
- Lavoué, S.; Miya, M.; Moritz, T.; Nishida, M. A molecular timescale for the evolution of the African freshwater fish family Kneriidae (Teleostei: Gonorynchiformes). Ichthyol. Res. 2011, 59, 104–112. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.; Fong, J. Catalog of Fishes: Genera, Species, References; California Academy of Sciences: San Francisco, CA, USA, 2017. [Google Scholar]
- Schelly, R.C. KNERIIDAE. Poisson. D’eaux Douces Saumâtres Basse Guinée Ouest L’afrique Cent. 2007, 1, 335–342. [Google Scholar]
- Steindachner, F. Ichthyologische Mittheilungen. (IX.). Verh. Zool. Bot. Ges. Wien 1866, 16, 761–771. [Google Scholar]
- Giltay, L. Notes ichthyologiques. IX. Contribution à l’étude du genre Xenopomatichthys (Kneriidae). Bull. Musée R. D’histoire Nat. De Belg. 1934, 10, 1–22. [Google Scholar]
- Pellegrin, J. Poissons nouveaux du Mozambique Bull. Muséum Natl. D’histoire Nat. 1905, 11, 145–146. [Google Scholar]
- Poll, M. Contribution à la Faune Ichthyologique du Katanga. Ann. Musée Congo Belg. 1933, 1, 52. [Google Scholar]
- Trewavas, E. DR. KARL JORDAN’S Expedition to South-West Africa and Angola: The fresh-water fishes. Novit. Zool. 1936, 40, 63–74. [Google Scholar]
- Lenglet, G. Contribution à l’étude de l’anatomie viscérale des Kneriidae. Ann. Société R. Zool. Belg. 1973, 103, 239–270. [Google Scholar]
- Seegers, L. Revision of the Kneriidae of Tanzania with description of three new Kneria-species (Teleostei: Gonorhynchiformes). Ichthyol. Explor. Freshw. 1995, 6, 97–128. [Google Scholar]
- Lenglet, G. Note sur la morphologie des Kneriidae (Pisces, Gonorhynchiformes). Les Nat. Belg. 1973, 54, 170–174. [Google Scholar]
- Poll, M. Descriptions de poissons nouveaux recueillis dans la région d’Albertville (Congo Belge) par le Dr G. Pojer. Bull. Du Musée R. D’histoire Nat. Belg. 1944, 20, 1–12. [Google Scholar]
- Norris, S.M.; Skelton, P.H. A Complete Guide to the Freshwater Fishes of Southern Africa. Copeia 1996, 1996, 755. [Google Scholar] [CrossRef]
- Poll, M. Exploration du Parc National de l’Upemba; Poissons. Fondation pour favoriser les Recherches Scientifiques en Afrique, Bruxelles: Brussels, Belgium, 1976; p. 127. [Google Scholar]
- Poll, M.; Gosse, J.P. Genera des poissons d’eau douce de l’Afrique. Mémoire de la Classe des Sciences. Acad. R. Belg. 1995, 9, 324p. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. Available online: www.fishbase.org (accessed on 16 March 2023).
- Runge, J. The Congo River, Central Africa. In Large Rivers: Geomorphology and Management; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008; pp. 293–309. [Google Scholar]
- Malaisse, F. Ecologie de la Riviѐre Luanza = Ecology of the Luanza River; Cercle Hydrobiologique de Bruxelles; Séries Exploration Hydrobiologique du lac Bangweolu et du Luapula: Brussels, Belgium, 1976; p. 151. [Google Scholar]
- Malaisse, F. La pêche collective par empoisonnement au ‘Buba’ (Tephrosia vogelii Hook). Son utilisation dans l’étude des populations de poissons. Nat. Belg. Bull. Mens. 1969, 50, 481–500. [Google Scholar]
- Malaisse, F. Se nourrir en Foret Claire Africaine: Approche Écologique et Nutritionnelle; CTA: Chicago, IL, USA, 1997. [Google Scholar]
- Van Steenberge, M.; Vreven, E.; Snoeks, J. The fishes of the Upper Luapula area (Congo basin): A fauna of mixed origin. Ichthyol. Explor. Freshw. 2014, 24, 329–345. [Google Scholar]
- Abwe, E. The Fish Fauna of the Kundelungu National Park (DR Congo): Diversity and Conservation; KU Leuven: Leuven, Belgium, 2022. [Google Scholar]
- Abel, R.; Thieme, M.L.; Revenga, C.; Bryer, M.; Kottelat, M.; Bogutskaya, N.; Coad, B.; Mandrak, N.; Balderas, S.C.; Bussing, W.; et al. Freshwater ecoregions of the world: A new map of biogeographic units for freshwater biodiversity conservation. BioScience 2008, 58, 403–414. [Google Scholar] [CrossRef]
- Neumann, D.A. Kinesiology of the Hip: A Focus on Muscular Actions. J. Orthop. Sports Phys. Ther. 2010, 40, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Somers, K.M. Multivariate Allometry and Removal of Size with Principal Components Analysis. Syst. Zool. 1986, 35, 359. [Google Scholar] [CrossRef]
- Bookstein, F.L.; Chernoff, B.; Elder, R.L. Morphometrics in Evolutionary Biology: The Geometry of Size and Shape Change, with Examples from Fishes; Academy of Natural Sciences of Philadelphia: Philadelphia, PA, USA, 1985; p. 15. [Google Scholar]
- Hammer. PAleontological STatistics (PAST), Version 3.10, Reference Manual; Natural History Museum, University of Oslo: Oslo, Norway, 2015. [Google Scholar]
- Snoeks, J. The Cichlid Diversity of Lake Malawi/Nyasa/Niassa: Identification, Distribution and Taxonomy; Cichlid Press: El Paso, TX, USA, 2004; p. 360. [Google Scholar]
- Vreven, E.J.; Teugels, G.G. Redescription of Mastacembelus liberiensis Boulenger, 1898 and description of a new West African spiny-eel (Synbranchiformes: Mastacembelidae) from the Konkoure River basin, Guinea. J. Fish Biol. 2005, 67, 332–369. [Google Scholar] [CrossRef]
- Vreven, E.J.; Snoeks, J. A new mastacembelid species from Lake Tanganyika: A case of complex evolutionary history. J. Fish Biol. 2009, 75, 1018–1047. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.R. Analyzing Tables of Statistical Tests. Evolution 1989, 43, 223. [Google Scholar] [CrossRef]
- Moelants, T.; Mbadu, Z.V.; Snoeks, J.; Vreven, E. A review of the Distichodus antonii assemblage (Characiformes: Distichodontidae) from the Congo basin. J. Nat. Hist. 2014, 48, 1707–1735. [Google Scholar] [CrossRef]
- UNESCO. The Miombo ecosystem. In Tropical Forest Ecosystems, a State of Knowledge; UNESCO: Paris, France, 1978. [Google Scholar]
- Mutambala, P.K.; Abwe, E.; Schedel, F.D.B.; Manda, A.C.; Schliewen, U.K.; Vreven, E.J.W.M.N. A new Parakneria Poll 1965 (Gonorhynchiformes: Kneriidae), ‘Mikinkidi’ from the Upper Lufira Basin (Upper Congo: DRC): Evidence from a morphologic and DNA barcoding integrative approach. J. Fish Biol. 2022, 102, 4–26. [Google Scholar] [CrossRef]
- Near, T.J.; Dornburg, A.; Friedman, M. Phylogenetic relationships and timing of diversification in gonorynchiform fishes inferred using nuclear gene DNA sequences (Teleostei: Ostariophysi). Mol. Phylogenetics E 2014, 80, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.R. Geographical distribution of African freshwater fishes. Zoöl. J. Linn. Soc. 1975, 57, 249–319. [Google Scholar] [CrossRef]
- Chakrabarty, P. Genetypes: A concept to help integrate molecular phylogenetics and taxonomy. Zootaxa 2010, 2632, 67–68. [Google Scholar] [CrossRef]
- Knebelsberger, T.; Stöger, I. DNA Extraction, Preservation, and Amplification. In DNA Barcodes: Methods and Protocols; Kress, J.W., Erickson, D.L., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 311–338. [Google Scholar]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Heled, J.; Kearse, M.; Moir, R.; Stones-Havas, S.; Sturrock, S.; et al. 2010 Geneious v5.5. Available online: http://www.geneious.com (accessed on 16 March 2023).
- Schedel, F.D.B.; Musilova, Z.; Schliewen, U.K. East African cichlid lineages (Teleostei: Cichlidae) might be older than their ancient host lakes: New divergence estimates for the east African cichlid radiation. BMC E Biol. 2019, 19, 94. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. E 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. E 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Srivathsan, A.; Meier, R. On the inappropriate use of Kimura-2-parameter (K2P) divergences in the DNA-barcoding literature. Cladistics 2011, 28, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Bektas, O.; Jones, J.A.; Sankararaman, S.; Roychoudhury, I.; Goebel, K. Reconstructing secondary test database from PHM08 challenge data set. Data Brief 2018, 21, 2464–2469. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.A.; Simonov, E.; Matveyev, M.P.; Artaev, O.N.; Mustafayev, N.J.; Pashkov, A.N.; Roubenyan, H.R. DNA barcoding of the fishes of the genus Alburnoides (Actinopterygii, Cyprinidae) from Caucasus. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2018, 29, 49–55. [Google Scholar] [CrossRef]
- Peters, N. Opercular-und Postopercularorgan (Occipitalorgan) der Gattung Kneria (Kneriidae, Pisces) und ein Vergleich mit verwandten Strukturen. Z. Für Morphol. Und Okol. Der Tiere 1967, 59, 381–435. [Google Scholar] [CrossRef]
- Wiley, M.L.; Collette, B.B. Breeding tubercles and contact organs in fishes: Their occurrence, structure, and significance. Bull. Am. Mus. Nat. Hist. 1970, 143, 145–216. [Google Scholar]
- Mittal, A.K.; Whitear, M. Keratinization of fish skin with special reference to the catfish Bagarius bagarius. Cell Tissue Res. 1979, 202, 213–230. [Google Scholar] [CrossRef]
- Roberts, T.R. Unculi (Horny Projections Arising from Single Cells), an Adaptive Feature of the Epidermis of Ostariophysan Fishes. Zoöl. Scr. 1982, 11, 55–76. [Google Scholar] [CrossRef]
- Manda, B.K.; Snoeks, J.; Decru, E.; Bills, R.; Vreven, E. Enteromius thespesios (Teleostei: Cyprinidae): A new minnow species with a remarkable sexual dimorphism from the south-eastern part of the Upper Congo River. J. Fish Biol. 2019, 96, 1160–1175. [Google Scholar] [CrossRef] [PubMed]
- Ahnelt, H.; Keckeis, H. Breeding tubercles and spawning behavior in Chondrostoma nasus (Teleostei: Cyprinidae): A correlation? Ichthyol. Explor. Freshw. 1994, 5, 321–330. [Google Scholar]
- Kortet, R.; Taskinen, J.; Sinisalo, T.; Jokinen, I. Breeding-related seasonal changes in immunocompetence, health state and condition of the cyprinid fish, Rutilus rutilus L. Biol. J. Linn. Soc. 2003, 78, 117–127. [Google Scholar] [CrossRef]
- Kleynhans, C.J. Aspects of the ecology of kneria auriculata (Pellegrin, 1905) (pisces: Kneriidae) from the eastern transvaal, south africa. J. Limnol. Soc. South Afr. 1988, 14, 108–118. [Google Scholar]
- Kalumba, L.N.; Abwe, E.; Frederic, S.; Manda, A.C.; Schliewen, U.; Vreven, E.J.W.M.N. Exploring the Kneria Steindachner, 1866 (Gonorhynchiformes: Kneriidae) «Miringiri» diversity of the Luanza and the Luongo rivers (Bangweulu-Mweru basin; DR Congo & Zambia). In Proceedings of the Sixth International Conference of the Pan African Fish and Fisheries, PAFFA, Malawi, 24–28 September 2018, African Fish and Fisheries: Diversity, Conservation and Sustainable Management, 6th ed.; PAFFA: Mangochi, Malawi, 2018; pp. 28–29. [Google Scholar]
- Levêque, C.; Paugy, D. Geographical distribution and affinities of African freshwater fishes. In The Inland Water Fishes of Africa: Diversity, Ecology and Human Use; Paugy, D., Levêque, C., Otero, O., Eds.; RMCA: Tervuren, Belgium, 2017; pp. 97–114. [Google Scholar]
- Boulenger, G.A. Catalogue of the Fresh-Water Fishes of Africa in the British Museum (Natural History); Taylor and Francis: London, UK, 1909. [Google Scholar]
- Poll, M. Revision des Characidae nains Africans. Ann. Mus. R. De L’afrique Cent. Zool. 1967, 8, 158p. [Google Scholar]
- David, L.R.; Poll, M. Contribution à la faune ichthyologique du Congo Belge collections du Dr. H. Schouteden (1924–1926). Ann. Musée Congo Belg. Zool. 1937, 3, 189–294. [Google Scholar]
- Thieme, M.L.; Abell, R.; Burgess, N.; World Wildlife Fund; Lehner, B.; Dinerstein, E.; Olson, D.; Teugels, G.; Kamdem-Toham, A.; Stiassny, M.L.J.S.; et al. Freshwater Ecoregions of Africa and Madagascar: A Conservation Assessment; Mendeley: Washington, DC, USA, 2005. [Google Scholar]
- Daget, J. Evolution et spéciation dans les milieux autres que les grands lacs. In Biologie et Écologie des Poissons D’eau Douce Africains; Leveque, C., Bruton, M.N., Ssentongo, G.W., Eds.; ORSTOM: Paris, France, 1988; pp. 53–63. [Google Scholar]
- Lévêque, C.; Agnèse, J.-F. La diversité des poissons africains: L’héritage de l’évolution. In Les Poissons des Eaux Continentales Africaines: Diversité, Écologie, Utilisation par L’homme; Lévêque, C., Paugy, D., Eds.; IRD: Paris, France, 2006; pp. 89–104. [Google Scholar]
- Roberts, T.R.; Stewart, D.J. An Ecological and Systematic Survey of Fishes in the Rapids of the Lower Zaire or Congo River. Bull. Mus. Comp. Zool. Harv. Coll. 1976, 147, 239–317. [Google Scholar]
- Worthington, E.B., III. The fishes (other than Cichlidæ) of Lake Bangweulu and adjoining regions, including descriptions of three new species. Ann. Mag. Nat. Hist. 1933, 12, 34–52. [Google Scholar] [CrossRef]
- PJackson, B.N. The Fishes of Northern Rhodesia: A Check List of Indigenous Species; Lusaka, Govt.: Printer, Zambia, 1961.
- Poll, M.; Stewart, D. Un Mochocidae et un Kneriidae nouveaux de la rivière Luongo (Zambia), affluent du bassin du Congo (Pisces). Rev. Zool. Afr. 1975, 89, 151–159. [Google Scholar]
- Balon, E.K.; Stewart, D.J. Fish assemblages in a river with unusual gradient (Luongo, Africa-Zaire system), reflections on river zonation, and description of another new species. Environ. Biol. Fishes 1983, 9, 225–252. [Google Scholar] [CrossRef]
- Romero, A.; Paulson, K.M. It’s a wonderful hypogean life: A guide to the troglomorphic fishes of the world. Biol. Hypogean Fishes 2001, 62, 13–41. [Google Scholar] [CrossRef]
- White, T.A.; Perkins, S.E. The ecoimmunology of invasive species. Funct. Ecol. 2012, 26, 1313–1323. [Google Scholar] [CrossRef]
- Boulenger, G.A. Mission Stappers au Tanganika-Moero. Diagnoses de poissons nouveaux. II. Mormyrides, Kneriides, Characinides, Cyprinides, Silurides. Rev. Zool. Afr. 1915, 4, 162–171. [Google Scholar] [CrossRef]
- Daly, M.C.; Green, P.; Watts, A.B.; Davies, O.; Chibesakunda, F.; Walker, R. Tectonics and Landscape of the Central African Plateau and their Implications for a Propagating Southwestern Rift in Africa. Geochem. Geophys. Geosystems 2020, 21, e2019GC008746. [Google Scholar] [CrossRef]
- De Dapper, M. The micro-relief of the sand-covered plateaux near Kolwezi (Shaba, Zaire). 1. The micro-relief of the over-all dilungu. Rev. Int. Géologie Géographie D’écologie Trop. 1979, 3, 1–18. [Google Scholar]
- Cotterill, F.P.D. The Upemba lechwe, Kobus anselli: An antelope new to science emphasizes the conservation importance of Katanga, Democratic Republic of Congo. J. Zool. 2005, 265, 113–132. [Google Scholar] [CrossRef]
- Davis, M.P.; Arratia, G.; Kaiser, T.M. The first fossil shellear and its implications for the evolution and divergence of the Kneriidae (Teleostei: Gonorynchiformes). Mesoz. Fishes 5-Glob. Divers. Evol. 2013, 325–362. [Google Scholar]
- Lavoué, S. Origins of Afrotropical freshwater fishes. Zool. J. Linn. Soc. 2019, 188, 345–411. [Google Scholar] [CrossRef]
- Chabwela, H.N. Current threats to the wetlands of Zimbabwe. In Wetlands Ecology and Priorities for Conservation in Zimbabwe: Proceedings of a Seminar on Wetlands Ecology and Priorities for Conservation in Zimbabwe; Matiza, T., Crafter, S.A., Programme, I.W., Eds.; IUCN: Gland, Switzerland, 1994; pp. 13–15. [Google Scholar]
- Toham, A.K. A Vision for Biodiversity Conservation in Central Africa: Biological Priorities for Conservation in the Guinean-Congolian Forest and Freshwater Region; World Wildlife Fund: Gland, Switzerland, 2006. [Google Scholar]
- Brooks, E.G.E.; Allen, D.J.; Darwall, W.R.T. The Status and Distribution of Freshwater Biodiversity in Central Africa; IUCN (International Union for the Conservation of Nature and Natural Resources): Gland, Switzerland; Cambridge, UK, 2011. [Google Scholar]
- Brooks, R.P.; Serfass, T.L. Aquatic Habitat Classifications: Implications for Otter Conservation. In Proceedings of the 14th International Otter Congress: Sichuan, China, 8–19 April 2019; Volume 31, pp. 3–12. [Google Scholar]
- Holland, R.A.; Garcia, N.; Brooks, E.G.E.; Juffe, D. Synthesis for all taxa. In The diversity of Life in African Freshwaters: Underwater, Underthreat. An Analysis of the Status and Distribution of Freshwater Species throughout Mainland Africa; Darwall, W., Smith, K., Allen, D., Holland, R., Brooks, I., Harrison, E., Eds.; IUCN: Gland, Switzerland; Cambridge, UK, 2011; pp. 228–269. [Google Scholar]
- Brooks, P.D.; Chorover, J.; Fan, Y.; Godsey, S.E.; Maxwell, R.M.; McNamara, J.P.; Tague, C. Hydrological partitioning in the critical zone: Recent advances and opportunities for developing transferable understanding of water cycle dynamics. Water Resour. Res. 2015, 51, 6973–6987. [Google Scholar] [CrossRef]
- Luther, H.; Rzo, J. Project Aqua: A Source Book of Inland Waters Proposed for Conservation; Blackwell Scientific: Oxford, UK, 1971. [Google Scholar]
















| Populations/Species | a. Inter-Group Distance | b. Intra-Group Distance | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | K2P | Uncor P | ||
| K. luansaensis sp. nov. | 1 | 7.0 | 8.8 | 8.2 | 9.5 | 7.9 | 6.6 | 6.7 | 7.5 | 8.2 | 8.5 | 7.1 | 0.2 | 0.2 | |
| K. maxi sp. nov. | 2 | 7.9 | 7.8 | 6.5 | 11.6 | 6.2 | 6.9 | 7.2 | 6.2 | 2.8 | 8.2 | 7.7 | 0.1 | 0.1 | |
| K. maydelli | 3 | 9.6 | 8.3 | 9.0 | 11.3 | 8.1 | 7.4 | 8.1 | 7.4 | 7.5 | 8.5 | 7.8 | 0.1 | 0.1 | |
| K. sp. kalungwishi | 4 | 8.7 | 7.0 | 9.8 | 10.7 | 5.6 | 6.8 | 6.4 | 5.0 | 6.1 | 7.8 | 6.7 | 1.0 | 1.0 | |
| K. sp. kwanza | 5 | 10.5 | 12.9 | 12.5 | 11.8 | 10.8 | 10.4 | 10.1 | 10.7 | 11.0 | 11.2 | 8.7 | n/c | n/c | |
| K. sp. lutshipuka | 6 | 8.6 | 6.6 | 8.7 | 5.9 | 12.0 | 6.7 | 6.1 | 5.3 | 6.1 | 7.1 | 7.1 | 0.2 | 0.2 | |
| K. sp. mukubwe ds | 7 | 7.1 | 7.4 | 8.0 | 7.2 | 11.5 | 7.2 | 2.8 | 6.8 | 6.5 | 7.4 | 5.2 | n/c | n/c | |
| K. sp. mukubwe us | 8 | 7.2 | 7.7 | 8.7 | 6.8 | 11.1 | 6.5 | 2.9 | 6.3 | 6.7 | 7.4 | 5.2 | n/c | n/c | |
| K. sp. ngona | 9 | 8.1 | 6.5 | 7.9 | 5.2 | 11.8 | 5.6 | 7.3 | 6.8 | 6.1 | 7.7 | 6.7 | 0.3 | 0.3 | |
| K. stappersii | 10 | 8.8 | 2.8 | 8.0 | 6.5 | 12.1 | 6.5 | 6.9 | 7.2 | 6.4 | 8.3 | 7.7 | 0.3 | 0.3 | |
| K. uluguru | 11 | 9.2 | 8.9 | 9.2 | 8.4 | 12.4 | 7.5 | 7.9 | 7.9 | 8.3 | 9.0 | 8.2 | 0 | 0 | |
| K. wittei | 12 | 7.6 | 8.3 | 8.4 | 7.2 | 9.4 | 7.6 | 5.4 | 5.4 | 7.2 | 8.3 | 8.8 | 0.3 | 0.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalumba, L.N.; Abwe, E.; Schedel, F.D.B.; Chocha Manda, A.; Schliewen, U.K.; Vreven, E.J.W.M.N. Two New Shellear Species (Gonorhynchiformes: Kneriidae), from the Luansa River (Upper Congo Basin): Hidden Diversity Revealed by Integrative Taxonomy. Diversity 2023, 15, 1044. https://doi.org/10.3390/d15101044
Kalumba LN, Abwe E, Schedel FDB, Chocha Manda A, Schliewen UK, Vreven EJWMN. Two New Shellear Species (Gonorhynchiformes: Kneriidae), from the Luansa River (Upper Congo Basin): Hidden Diversity Revealed by Integrative Taxonomy. Diversity. 2023; 15(10):1044. https://doi.org/10.3390/d15101044
Chicago/Turabian StyleKalumba, Lewis Ngoy, Emmanuel Abwe, Frederic D. B. Schedel, Auguste Chocha Manda, Ulrich K. Schliewen, and Emmanuel J. W. M. N. Vreven. 2023. "Two New Shellear Species (Gonorhynchiformes: Kneriidae), from the Luansa River (Upper Congo Basin): Hidden Diversity Revealed by Integrative Taxonomy" Diversity 15, no. 10: 1044. https://doi.org/10.3390/d15101044