

Diversity and Phylogeny of Gyrodactylus spp. (Monogenea: Gyrodactylidae) across the Strait of Gibraltar: Parasite Speciation and Historical Biogeography of West Mediterranean Cyprinid Hosts

Abstract
:1. Introduction
2. Material and Methods
2.1. Collection of Cyprinid Host Specimens and Their Gyrodactylid Monogeneans
2.2. Morphological Characterization and Multivariate Analysis of West Mediterranean Gyrodactylus
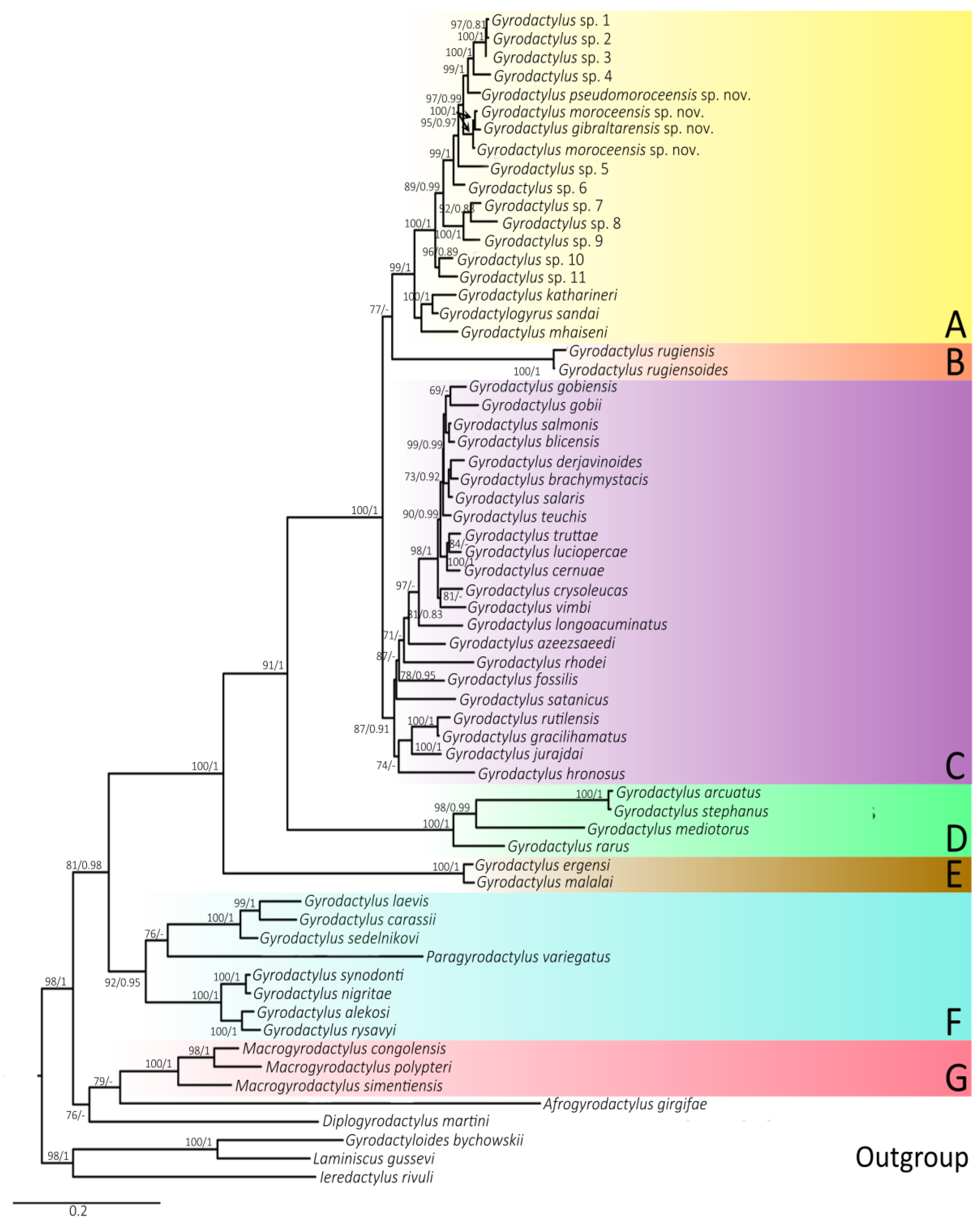
2.3. Phylogenetic Reconstruction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gyrodactylid Species | Host Species | Host Family | Geographical Locality | 18S rDNA | ITS Regions | Reference |
|---|---|---|---|---|---|---|
| Afrogyrodactylus girgifae Přikrylová et Luus-Powell, 2014 | Brycinus nurse (Rüppell, 1832) | Alestidae | Africa | HF548672 | HF548671 | [75] |
| Diplogyrodactylus martini Přikrylová, Matějusová, Musilová, Gelnar and Harris, 2009 | Polypterus senegalus Cuvier, 1829 | Polypteridae | Africa | HE858426 | AM943008 | [75] |
| Gyrodactyloides bychowskii Albova, 1948 | Salmo salar Linnaeus, 1758 | Salmonidae | Europe | AJ566379 | AJ249348 | [76] |
| Gyrodactylus alekosi Prikrylova, Blazek and Vanhove, 2011 | Clarias gariepinus (Burchell, 1822) | Clariidae | Africa | FR850683 | FR850682 | [77] |
| Gyrodactylus arcuatus Bychowsky, 1933 | Gasterosteus aculeatus Linnaeus, 1758 | Gasterosteidae | North America | JF836088 | AF156668 | [61,78] |
| Gyrodactylus blicensis Glaser, 1974 | Gymnocephalus cernua (Linnaeus, 1758) | Percidae | Europe | AJ407896 | AJ407869 AJ407919 | [62] |
| Gyrodactylus brachymystacis Ergens, 1978 | Brachymystax lenok (Pallas, 1773) | Salmonidae | Asia | JF836109 | KP325622 * | [78] |
| Carassius auratus (Linnaeus, 1758) | Cyprinidae | |||||
| Oncorhynchus mykiss (Walbaum, 1792) | Salmonidae | |||||
| Gyrodactylus carassii Malmberg, 1957 | Alburnus alburnus (Linnaeus, 1758) | Leuciscidae | Europe | AJ566377 | AY278033 | [62,79] |
| Carassius carassius (Linnaeus, 1758) | Cyprinidae | |||||
| Gyrodactylus cernuae Malmberg, 1957 | Gymnocephalus cernua (Linnaeus, 1758) | Percidae | Europe | AJ407897 | AJ407869 AJ407919 | [62] |
| Gyrodactylus crysoleucas Mizelle and Kritsky, 1967 | Notemigonus crysoleucas (Mitchill, 1814) | Leuciscidae | North America | KT149283 | KT149287 | [80] |
| Gyrodactylus derjavinoides Malmberg, Collins, Cunningham and Jalali, 2007 | Salmo salar Linnaeus, 1758 | Salmonidae | Europe | JF836110 | GQ368236 | [78] |
| Gyrodactylus ergensi Přikrylová, Matějusová, Musilová et Gelnar, 2009 | Sarotherodon galilaeus (Linnaeus, 1758) | Cichlidae | Africa | HF548668 | FN394985 | [75] |
| Gyrodactylus fossilis Lupu and Roman, 1956 | Misgurnus fossilis (Linnaeus, 1758) | Cobitidae | Europe | AJ407898 | AJ407871 AJ407921 | [76] |
| Gyrodactylus gibraltarensis Rahmouni sp. nov. | Luciobarbus graellsii (Steindachner, 1866) | Cyprinidae | Europe | OR773531 | OR773480 | This study |
| Gyrodactylus gobiensis Gloser, 1974 | Gobio gobio (Linnaeus, 1758) | Gobionidae | Europe | AJ566375 | AY278041 | [76] |
| Gyrodactylus gobii Schulman, 1953 | Gobio gobio (Linnaeus, 1758) | Gobionidae | Europe | AJ407900 | AJ407873 | [62] |
| Gyrodactylus gracilihamatus Malmberg, 1964 | Abramis brama (Linnaeus, 1758) Alburnus alburnus (Linnaeus, 1758) | Leuciscidae | Europe | AJ407901 | AF484531 | [76,81] |
| Gyrodactylus azeezsaeedi Rahmouni, 2023 | Squalius berak Heckel, 1843 | Leuciscidae | Middle East | OR777687 | OR773093 | [82] this study |
| Gyrodactylus hronosus Žitňan, 1964 | Alburnoides bipunctatus (Bloch, 1782) | Leuciscidae | Europe | AJ407902 | AJ407876 AJ407924 | [62] |
| Gyrodactylus jurajdai Rahmouni, 2023 | Chondrostoma regium (Heckel, 1843) | Leuciscidae | Middle East | OR777685 | OR773088 | [82], this study |
| Gyrodactylus katharineri Malmberg, 1964 | Barbus barbus (Linnaeus, 1758) | Cyprinidae | Europe | AJ407903 | AJ407878 AJ407926 | [62] |
| Gyrodactylus laevis Malmberg, 1957 | Alburnoides bipunctatus (Bloch, 1782) | Leuciscidae | Europe | AJ407904 | AY278036 | [76] |
| Phoxinus phoxinus (Linnaeus, 1758) | ||||||
| Gyrodactylus longoacuminatus Žitňan, 1964 | Carassius auratus (Linnaeus, 1758) | Cyprinidae | Europe | AJ407906 | AJ407883 AJ407930 | [76] |
| Gyrodactylus luciopercae Gussev, 1962 | Perca fluviatilis Linnaeus, 1758 | Percidae | Europe | AJ407907 | AJ407885 | [76] |
| Gyrodactylus malalai Přikrylová, Blažek et Gelnar, 2012 | Oreochromis niloticus (Linnaeus, 1758) | Cichlidae | Africa | FR695485 | FR695484 | [75] |
| Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013 | Notropis texanus (Girard, 1856) | Gasterosteidae | North America | MW666777 | MW666182 | [83] |
| Gyrodactylus moroccensis Rahmouni sp. nov. | Luciobarbus rabatensis Doadrio, Perea and Yahyaoui, 2015 | Cyprinidae | North Africa | OR773529 | OR773478 | This study |
| Luciobarbus rifensis Doadrio, Casal-Lopez and Yahyaoui, 2015 | OR773528 | OR773477 | ||||
| Gyrodactylus mhaiseni Rahmouni, 2023 | Alburnus sellal Heckel, 1843 | Leuciscidae | Middle East | OR777688 | OR773082 | [82], this study |
| Gyrodactylus nigritae Přikrylová, Blažek and Vanhove, 2012 | Synodontis nigrita Valenciennes, 1840 | Mochokidae | Africa | FR850687 | FR850686 | [77] |
| Gyrodactylus pseudomoroccensis Rahmouni sp. nov. | Luciobarbus ksibi (Boulenger, 1905) | Cyprinidae | North Africa | OR773530 | OR773479 | This study |
| Gyrodactylus rarus Wegener, 1910 | Spinachia spinachia (Linnaeus, 1758) | Gasterosteidae | Europe | AY339776 | AY338445 | [84] |
| Gyrodactylus rhodei Žitňan, 1964 | Rhodeus sericeus (Pallas, 1776) | Acheilognathidae | Europe | AJ567670 | AJ407889 AJ407933 | [76] |
| Gyrodactylus rugiensis Glaser, 1974 | Pomatoschistus microps (Krøyer, 1838) | Gobiidae | Europe | AY339762 | AY338446 | [84] |
| Gyrodactylus rugiensoides Huyse and Volckaert, 2002 | Pomatoschistus minutus (Pallas, 1770) | Gobiidae | Europe | AY339763 | AJ427414 | [84,85] |
| Gyrodactylus rutilensis Glaser, 1974 | Rutilus rutilus (Linnaeus, 1758) | Leuciscidae | Europe | AJ566376 | AJ407890 AJ407934 | [76] |
| Gyrodactylus rysavyi Ergens, 1973 | Clarias anguillaris (Linnaeus, 1758) | Clariidae | Africa | FR850680 | FR850679 | [77] |
| Gyrodactylus salaris Malmberg, 1957 | Salmo salar Linnaeus, 1758 | Salmonidae | Europe | JF836111 | AJ515912 | [86] |
| Oncorhynchus mykiss (Walbaum, 1792) | ||||||
| Gyrodactylus salmonis Yin and Sproston, 1948 | Oncorhynchus mykiss (Walbaum, 1792) | Salmonidae | Europe | JF836097 | MN850542 | [78,87] |
| Gyrodactylus sandai Rahmouni, 2023 | Capoeta umbla (Heckel, 1843) | Cyprinidae | Middle East | OR777689 | OR773089 | [82], this study |
| Gyrodactylus satanicus Rahmouni, 2023 | Garra rufa Heckel, 1843 | Cyprinidae | Middle East | OR777686 | OR773091 | [82], this study |
| Gyrodactylus sedelnikovi Gvozdev, 1950 | Barbatula barbatula (Linnaeus, 1758) | Nemacheilidae | Europe | AJ407911 | AJ407891 AJ407935 | [62] |
| Gyrodactylus stephanus Müller, 1937 | Fundulus heteroclitus (Linnaeus, 1766) | Gasterosteidae | North America | JF836099 | FJ845515 | [78,88] |
| Gyrodactylus synodonti Přikrylová, Blažek, Maarten and Vanhove, 2012 | Synodontis nigrita Valenciennes, 1840 | Mochokidae | Africa | FR850685 | FR850684 | [77] |
| Gyrodactylus teuchis Lautraite, Blanc, Thiery, Daniel and Vigneulle, 1999 | Salmo trutta Linnaeus, 1758 | Salmonidae | Europe | AJ407912 | AJ249350 | [76] |
| Gyrodactylus truttae Gläser, 1974 | Salvelinus fontinalis (Mitchill, 1814) | Salmonidae | Europe | JF836112 | AJ132260 | [65,78] |
| Gyrodactylus vimbi Shulman, 1954 | Squalius cephalus (Linnaeus, 1758) | Leuciscidae | Europe | AJ407914 | AJ407892 AJ407936 | [62] |
| Ieredactylus rivuli Schelkle et al., 2011 | Anablepsoides hartii (Boulenger, 1890) | Cyprinodontiformes | Neotropical | JX840358 | HQ738514 | [89,90] |
| Laminiscus gussevi (Bychovsky et Polyansky, 1953) | Mallotus villosus (Müller, 1776) | Osmeridae | Europe | HF548679 | HF548678 | [75] |
| Macrogyrodactylus congolensis (Prodhoe, 1957) | Clarias gariepinus (Burchell, 1822) | Clariidae | Africa | HF548680 | GU252716 | [75,91] |
| Macrogyrodactylus polypterid Malmberg, 1957 | Polypterus senegalus Cuvier, 1829 | Polypteridae | Africa | AJ567671 | AJ567672 | [76] |
| Macrogyrodactylus simentiensis Přikrylová and Gelnar, 2008 | Polypterus senegalus Cuvier, 1829 | Polypteridae | Africa | HF548682 | HF548681 | [75] |
| Paragyrodactylus variegatus You, King, Ye and Cone, 2014 | Homatula variegata (Dabry de Thiersant, 1874) | Nemacheilidae | Asia | KF680220 | KF680221 | [19] |
| Gyrodactylus sp. 1 | Luciobarbus bocagei (Steindachner, 1864) | Cyprinidae | Europe | OR807824 | OR807835 | This study |
| Gyrodactylus sp. 2 | Luciobarbus sclateri (Günther, 1868) | Cyprinidae | Europe | OR807825 | OR807836 | This study |
| Gyrodactylus sp. 3 | Luciobarbus comizo (Steindachner, 1864) | Cyprinidae | Europe | OR807826 | OR807837 | This study |
| Gyrodactylus sp. 4 | Luciobarbus bocagei (Steindachner, 1864) | Cyprinidae | Europe | OR807827 | OR807838 | This study |
| Gyrodactylus sp. 5 | Labeobarbus maroccanus (Günther, 1902) | Cyprinidae | North Africa | OR807828 | OR807839 | This study |
| Gyrodactylus sp. 6 | Luciobarbus zayanensis Doadrio, Casal-López & Yahyaoui, 2016 | Cyprinidae | North Africa | OR807829 | OR807840 | This study |
| Gyrodactylus sp. 7 | Luciobarbus rabatensis Doadrio, Perea and Yahyaoui, 2015 | Cyprinidae | North Africa | OR807830 | OR807841 | This study |
| Gyrodactylus sp. 8 | Luciobarbus massaensis (Valenciennes, 1842) | Cyprinidae | North Africa | OR807831 | OR807842 | This study |
| Gyrodactylus sp. 9 | Luciobarbus rifensis Doadrio, Casal-Lopez and Yahyaoui, 2015 | Cyprinidae | North Africa | OR807832 | OR807843 | This study |
| Gyrodactylus sp. 10 | Luciobarbus yahyaouii Doadrio, Casal-Lopez & Perea, 2016 | Cyprinidae | North Africa | OR807833 | OR807844 | This study |
| Gyrodactylus sp. 11 | Luciobarbus massaensis (Valenciennes, 1842) | Cyprinidae | North Africa | OR807834 | OR807845 | This study |
3. Results
3.1. Systematics and Molecular Characterization
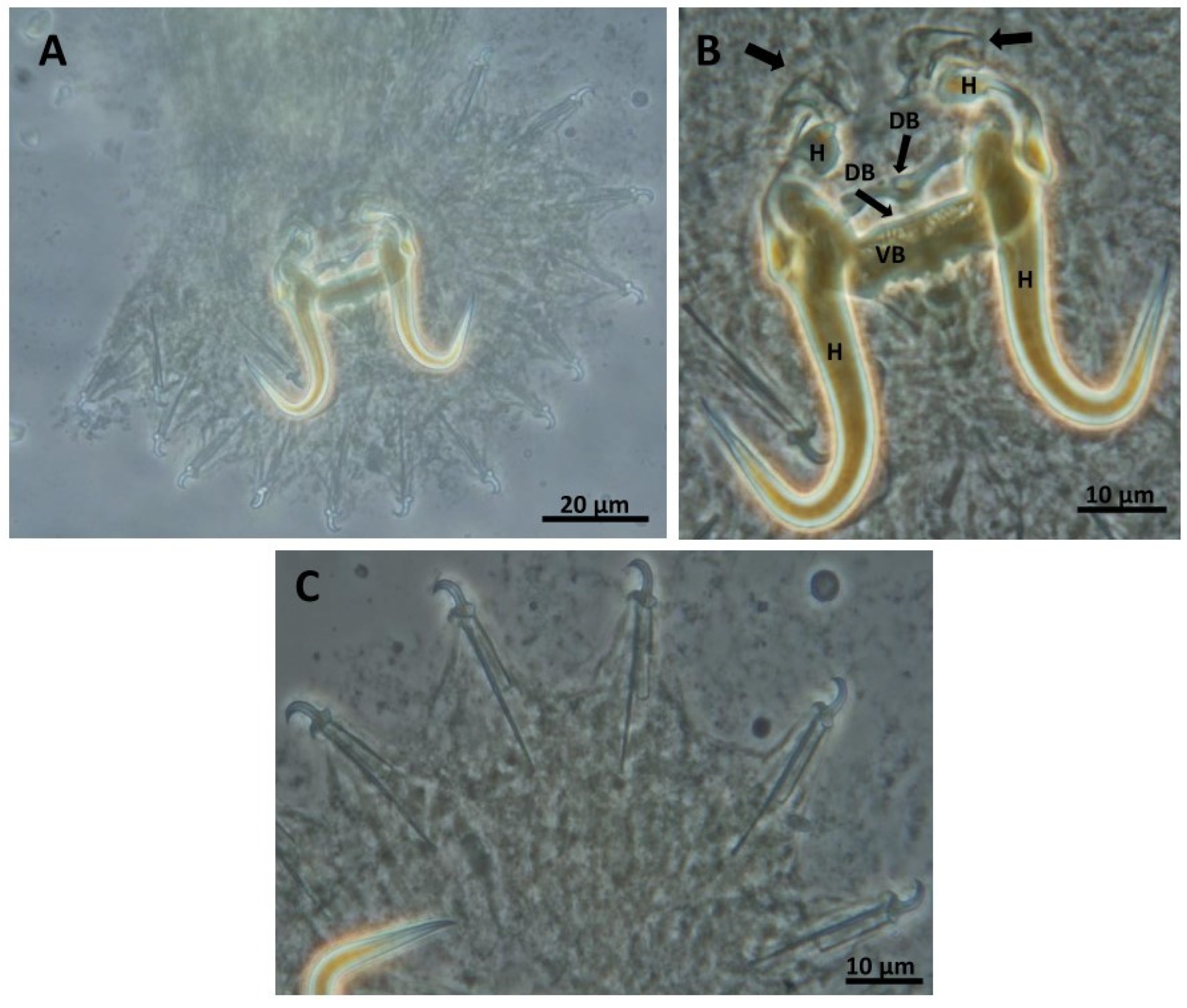
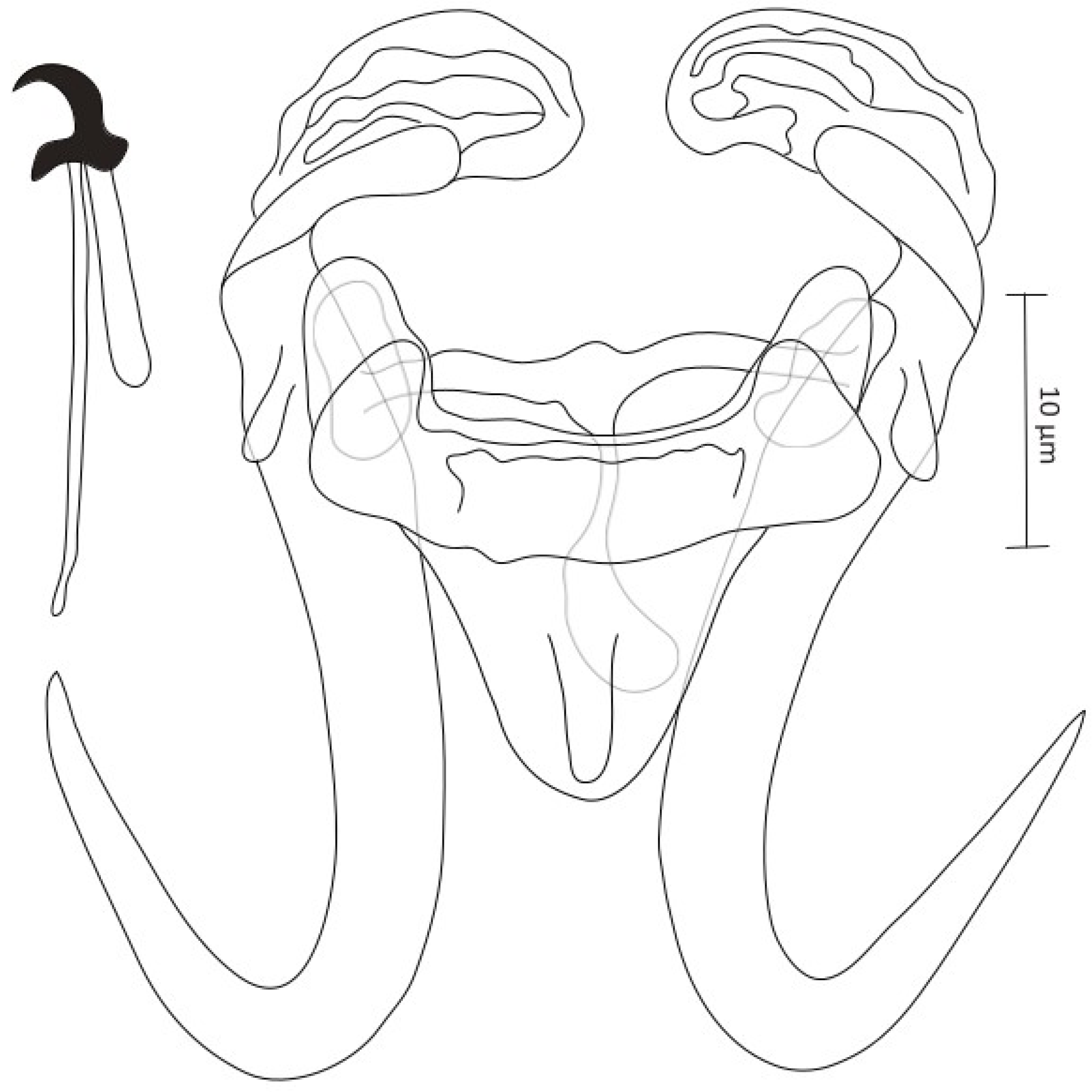
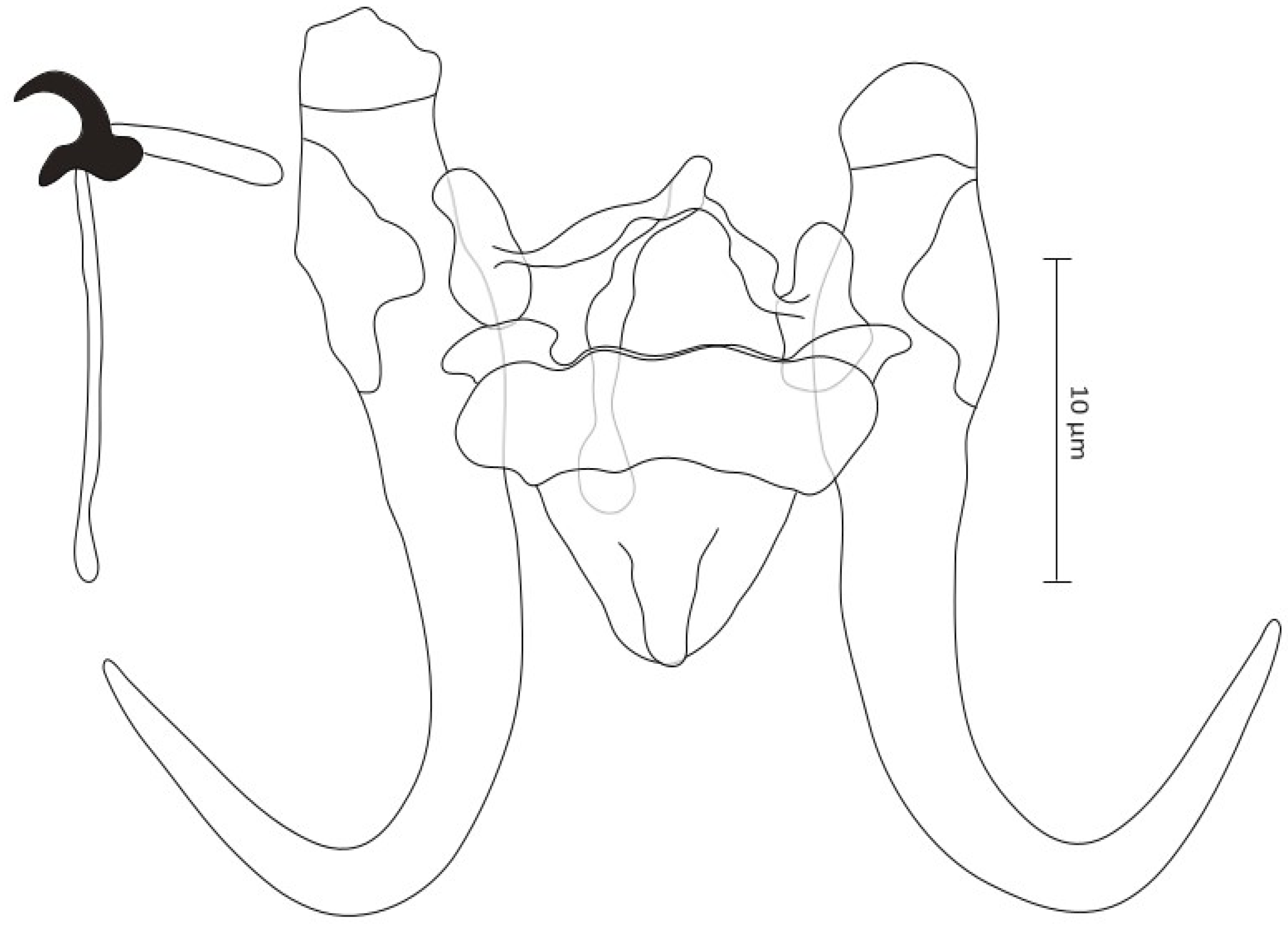
- Gyrodactylus gibraltarensis Rahmouni sp. nov. (Figure 3)

- Zoobank registration: urn:lsid:zoobank.org:act:88464789-AB95-4F25-99D2-44EA94E81F59
- Type–host: Iberian barbel Luciobarbus graellsii (Steindachner, 1866) (Cyprinidae)
- Type–material: one holotype and one paratype under the accession number IPCAS M-779
- Site on the host: gill filaments
- Etymology: The epithet “gibraltarensis” of the type species refers to the Strait of Gibraltar, which played an important role in the dispersal of freshwater fish to the Iberian Peninsula
- DNA sequences: 18S rDNA: OR773531, ITS regions: OR773480Description (morphometric measurements are given in Table 3)
| Gyrodactylus moroccensis sp. nov. | Gyrodactylus gibraltarensis sp. nov. | Gyrodactylus pseudomoroccensis sp. nov. | |||||
|---|---|---|---|---|---|---|---|
| Cyprinid fish hosts | Luciobarbus maghrebensis | Luciobarbus yahyaouii | Luciobarbus zayanensis | Luciobarbus rabatensis * | Luciobarbus rifensis | Luciobarbus graellsii * | Luciobarbus ksibi * |
| Hamuli | |||||||
| Length | 47.6 (45.9–50.9)10 | 48 (45.1–49.7)3 | 51.8 (49.9–54)3 | 47.6 (45.9–50.9)10 | 44.9 (43.8–46)2 | 36.9 (35.3–40.8)4 | 37.1 (36.1–38.2)5 |
| Outer root length | 10.6 (8.8–12.5)10 | 11.7 (9.5–13.9)3 | 12.3 (11.1–13.4)3 | 10.6 (8.8–12.5)10 | 8.4 (8–8.9)2 | 8.5 (7.6–9.8)4 | 11.4 (10.6–12)5 |
| Shaft length | 37.6 (35.5–39)10 | 37.2 (36.5–37.9)3 | 40.5 (38.6–41.9)3 | 37.6 (35.5–39)10 | 36.7 (35.7–37.6)2 | 28.5 (27.1–30.7)4 | 27.3 (26–28.9)5 |
| Point length | 23 (20.1–25.1)10 | 22.5 (21.9–23.4)3 | 23.2 (22.6–23.6)3 | 23 (20.1–25.1)10 | 24.2 (22.8–25.5)2 | 17.4 (15.5–19.3)4 | 14.9 (14.3–15.4)5 |
| Ventral bar | |||||||
| Length | 28.8 (28.6–29.4)10 | 29.5 (27.6–31.9)3 | 30.9 (29.1–33.6) 3 | 28.8 (28.6–29.4)10 | 27.9 (27.6–28.3)2 | 21.2 (19.2–25.6)4 | 15.3 (14.7–16)5 |
| Width | 29.7 (27.7–31)10 | 26.7 (26.1–27.2)3 | 32 (31–32.5)3 | 29.7 (27.7–31)10 | 26.8 (26.2–27.3)2 | 21.6 (19.8–24.6)4 | 17.4 (16.5–18.4)5 |
| Tips length | 8 (6.3–9.4)10 | 6.1 (5.7–6.5)3 | 7.2 (6.9–7.6)3 | 8 (6.3–9.4)10 | 7.1 (5.9–8.3)2 | 2.9 (2.7–3.2)4 | 3.2 (2.4–4.1)5 |
| Distance between tips | 31.1 (29.2–32.4)10 | 27.9 (26.7–28.8)3 | 31.5 (30.8–32.2)3 | 31.1 (29.2–32.4)10 | 27.3 (26.7–27.9)2 | 23.2 (21.9–24)4 | 17.4 (16–18.1)5 |
| Median width | 7.5 (6.5–8.2)10 | 5.9 (5.4–6.2)3 | 7.5 (7–7.9)3 | 7.5 (6.5–8.2)10 | 5.3 (4.8–5.8)2 | 4.2 (3.6–5.4)4 | 5.3 (5.1–5.6)5 |
| Membrane length | 12.3 (11.1–13.5)10 | 14.7 (14.1–15.6)3 | 14.2 (13.7–14.5)3 | 12.3 (11.1–13.5)10 | 13 (12.5–13.5)2 | 11.6 (10.6–12.8)4 | 9 (8.3–9.6)5 |
| Membrane width | 15.3 (14.1–16.6)10 | 13.3 (11.2–15.2) 3 | 18.7 (17.2–20.3)3 | 15.3 (14.1–16.6)10 | 15.8 (15.1–16.4)2 | 13.7 (13–15.2)4 | 10.4 (9.4–11.2)5 |
| Dorsal bar | |||||||
| Length | 27.9 (26.7–28.5)10 | 27.9 (26.8–29.1)3 | 31.5 (31.2–32)3 | 27.9 (26.7–28.5)10 | 27.3 (25.4–29.1)2 | 22.3 (20.5–24.2)4 | 19.9 (18.7–21.5)5 |
| Median width | 17.9 (16–19.4)10 | 16.9 (15.8–17.7)3 | 20 (19.7–20.6)3 | 17.9 (16–19.4)10 | 15.4 (14.3–16.5)2 | 10.8 (8.6–14)4 | 16.2 (15–17)5 |
| Marginal hooks | |||||||
| Length | 29.5 (28.1–31.8)10 | 27.1 (26.7–27.5)3 | 31.3 (30.5–32)3 | 29.5 (28.1–31.8)10 | 25.5 (25.3–25.7)2 | 22.8 (21.5–23.4)4 | 22.1 (20.5–23.6)5 |
| Filament loop length | 10.6 (9.2–12.7)10 | 8.9 (8.3–10)3 | 13.2 (12.4–14.3)3 | 10.6 (9.2–12.7)10 | 11.5 (10.9–12)2 | 10.1 (9.6–10.6)4 | 8.2 (7.5–8.8)5 |
| Handle length | 24.1 (22.6–26.6)10 | 21.2 (20.4–21.8)3 | 24.9 (24.2–25.4)3 | 24.1 (22.6–26.6)10 | 20.4 (20.4–20.5)2 | 17.8 (17.3–18.6)4 | 17.6 (16.8–18.6)5 |
| Sickle length to shaft attachment | 6 (5–6.9)10 | 5.4 (5.2–5.5)3 | 6.2 (5.9–6.4)3 | 6 (5–6.9)10 | 5.1 (5–5.2)2 | 4.6 (4.2–5) 4 | 4.6 (4.4–4.8)5 |
| Sickle proximal width | 5.1 (4.6–5.5)10 | 4.5 (4.2–4.8)3 | 5.1 (4.7–5.4)3 | 5.1 (4.6–5.5)10 | 4.2 (4.1–4.3)2 | 3.8 (3.3–4.5)4 | 3.7 (3.5–4)5 |
| Sickle distal width | 4.9 (4.4–5.2)10 | 5.3 (4.7–5.6)3 | 6.1 (5.6–6.4)3 | 4.9 (4.4–5.2)10 | 4.3 (4.2–4.4)2 | 4.2 (3.9–4.4)4 | 4.1 (4–4.4)5 |
| Shaft length of sickle | 4.9 (4.6–5.1)10 | 4.7 (4.1–5.2)3 | 5.3 (4.7–5.8)3 | 4.9 (4.6–5.1)10 | 4 (3.9–4.1)2 | 4.4 (3.7–5.5)4 | 3.7 (3.5–3.8)5 |
| Point length of sickle | 2.5 (2.1–2.7)10 | 2.6 (2.4–2.8)3 | 3.7 (3.4–3.9)3 | 2.5 (2.1–2.7)10 | 2.2 (2.2–2.3)2 | 1.8 (1.3–2.8)4 | 1.9 (1.8–2)5 |
| Male copulatory organ (MCO) | |||||||
| Length | 16 (14.7–19.4)6 | Not observed | 17.71 | 16 (14.7–19.4)10 | 12.71 | Not observed | Not observed |
| Width | 17.2 (15.9–19.2)6 | Not observed | 17.61 | 17.2 (15.9–19.2)10 | 13.51 | Not observed | Not observed |
| Pharynx | |||||||
| Length | 46.8 (43.5–54.9)10 | 38.8 (35.2–44.2)3 | 40.1 (38.6–42.9)10 | 40.1 (38.6–42.9)10 | 36.1 (35.7–36.5)2 | – | – |
| Width | 39.3 (37.6–44.5)10 | 38.7 (36–41.8)3 | 36.5 (30.6–40.3)10 | 36.5 (30.6–40.3)10 | 27 (25–4–28.7)2 | – | – |
- Diagnosis
- Molecular taxonomy
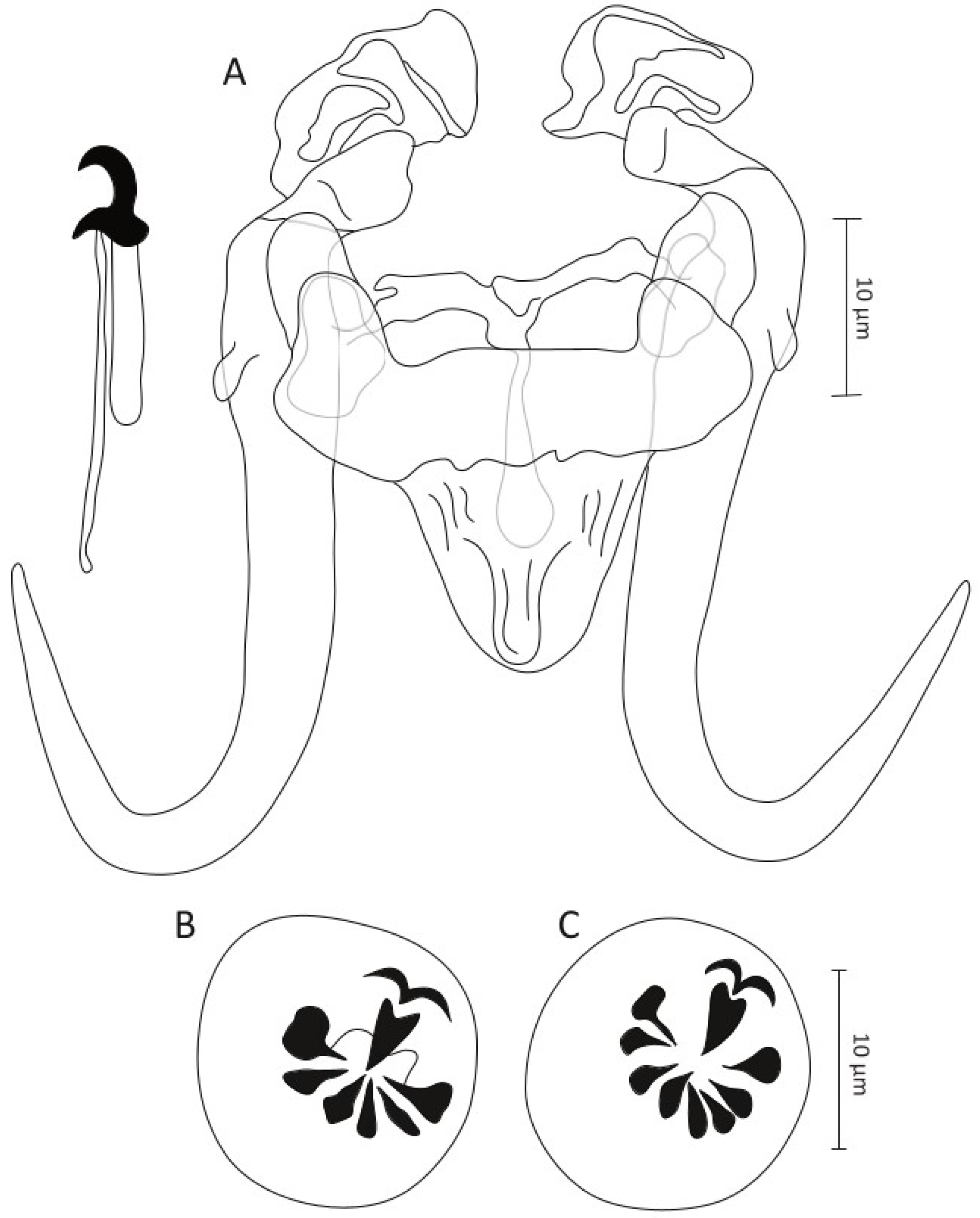
- Gyrodactylus moroccensis Rahmouni sp. nov. (Figure 4)

- Zoobank registration: urn:lsid:zoobank.org:act:D58EB381-CAB9-4DF5-9157-FCC55EB0EE18
- Type–host: Rabat barbel Luciobarbus rabatensis Doadrio, Perea and Yahyaoui, 2015 (Cyprinidae)
- Additional hosts: Yahyaoui barbel Luciobarbus yahyaouii Doadrio, Casal-Lopez & Perea, 2016, Zayan Barbel Luciobarbus zayanensis Doadrio, Casal-López & Yahyaoui, 2016, Maghreb barbel Luciobarbus maghrebensis Doadrio, Perea and Yahyaoui, 2015, and Rifian barbel Luciobarbus rifensis Doadrio, Casal-Lopez & Yahyaoui, 2015 (all Cyprinidae)
- Type–material: one holotype and three paratypes under the accession number IPCAS M-779
- Site on the host: fins for L. maghrebensis, L. yahyaouii, L. zayanensis and L. rifensis; gill filaments for L. rabatensis
- Etymology: The epithet “moroccensis” of the type–species refers to its country of origin (Morocco)
- DNA sequences: 18S rDNA: OR773528-29, ITS regions: OR773477-78
- Description (morphometric measurements are given in Table 3)
- Diagnosis
- Molecular taxonomy
- Gyrodactylus pseudomoroccensis Rahmouni sp. nov. (Figure 5)

- Zoobank registration: urn:lsid:zoobank.org:act:45ED3EAA-7F91-4370-ACDE-7B5AEA155363
- Type–material: one holotype and one paratype under the accession number IPCAS M-781Site on the host: fins
- Etymology: The epithet “pseudomoroccensis” of the type–species refers to the morphological similarity to specific elements of the attachment organ (T-shaped dorsal bar) in the two new Gyrodactylus spp. described from Morocco.
- DNA sequences: 18S rDNA: OR773530, ITS regions: OR773479
- Description (morphometric measurements are given in Table 3)
- Diagnosis
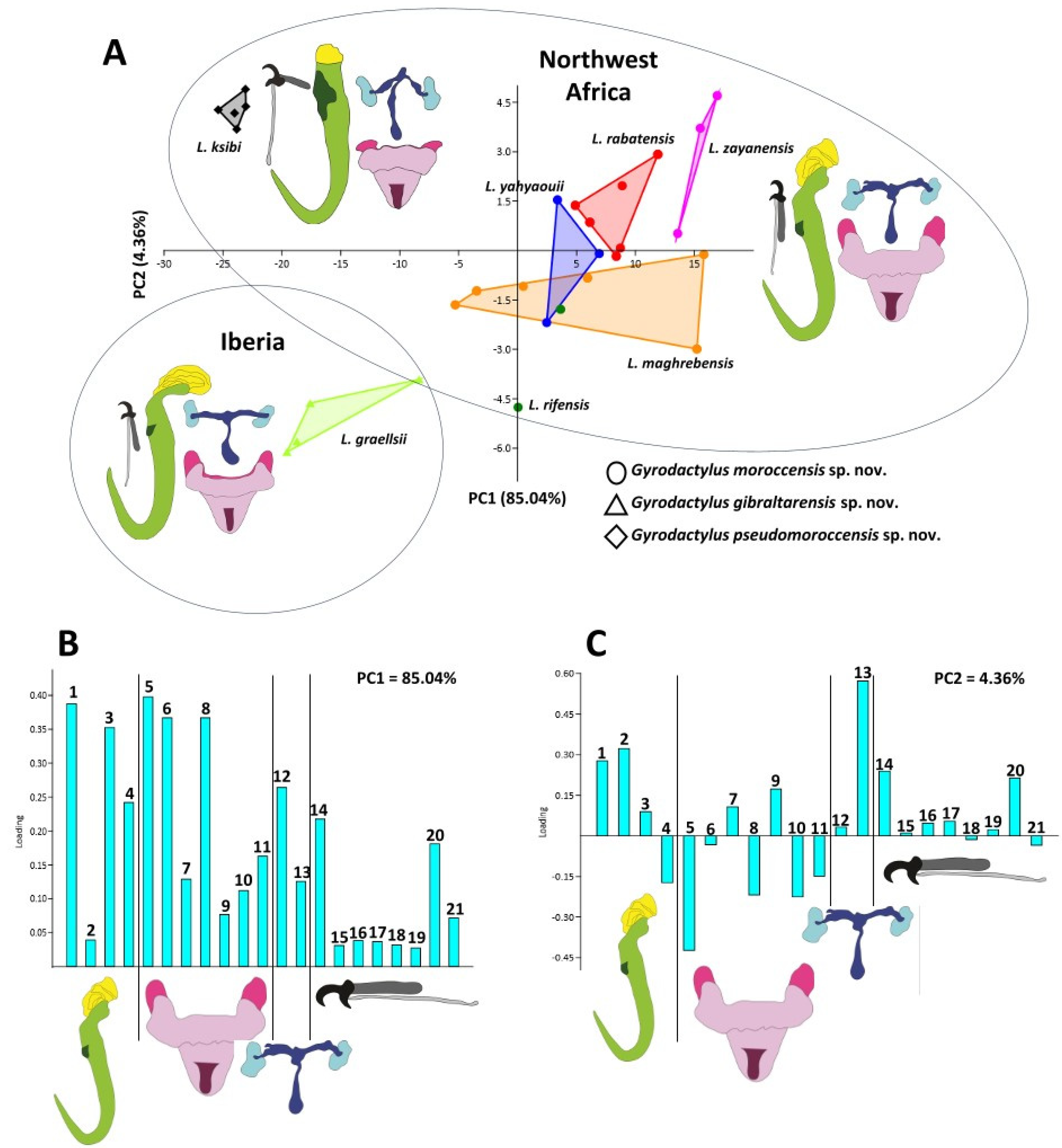
3.2. Morphological Delineation among Northwest African and Iberian Gyrodactylus
3.3. Phylogenetic Relationships
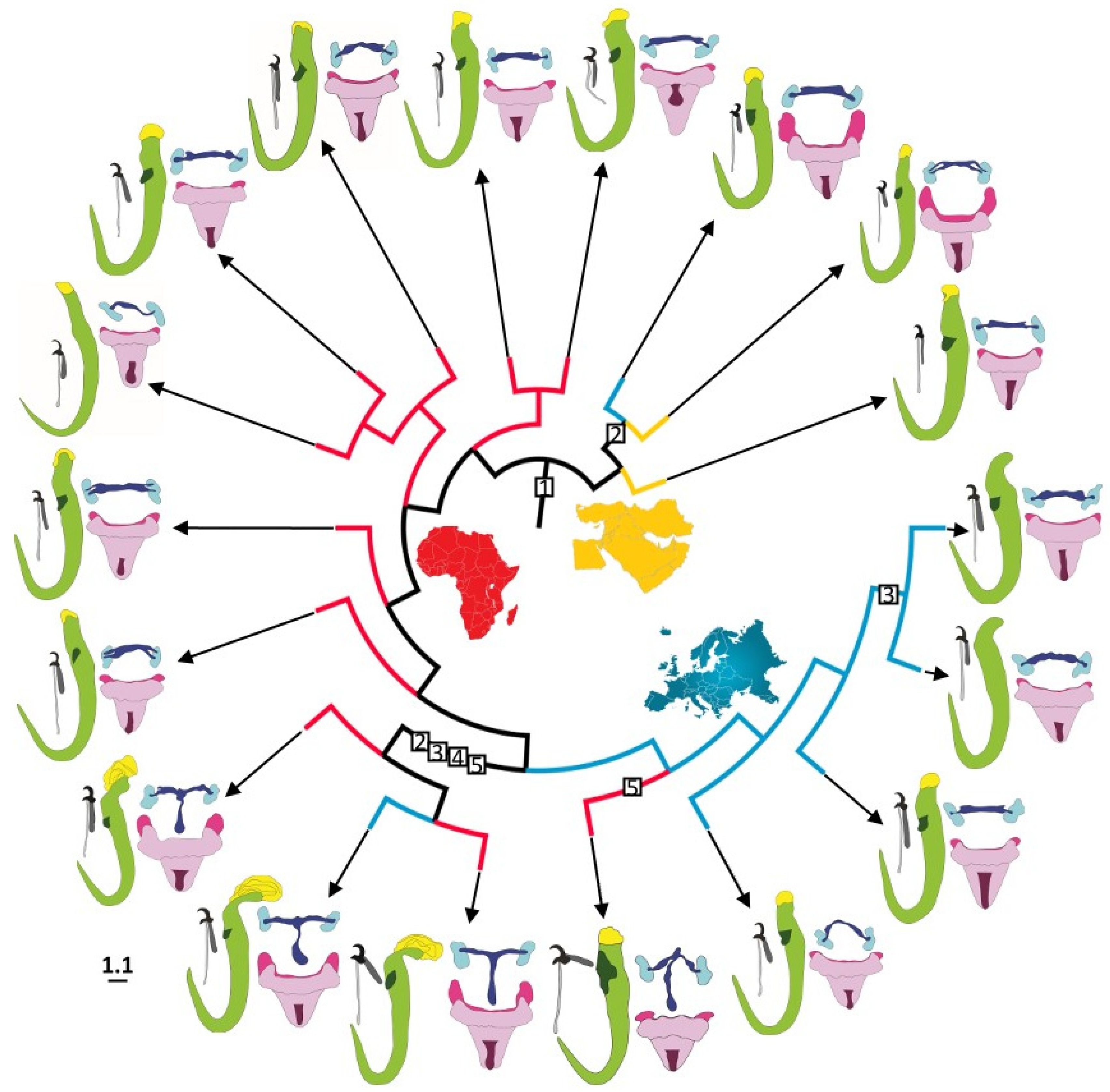
3.4. Morphological Evolution of Gyrodactylus in the West Mediterranean Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huyse, T.; Poulin, R.; Théron, A. Speciation in parasites: A population genetics approach. Trends Parasitol. 2005, 21, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, S.B.; Kuris, A.M. Independent origins of parasitism in Animalia. Biol. Lett. 2016, 12, 20160324. [Google Scholar] [CrossRef] [PubMed]
- Rahmouni, C.; Vanhove, M.P.M.; Šimková, A.; Van Steenberge, M. Morphological and genetic divergence in a gill monogenean parasitizing distant cichlid lineages of Lake Tanganyika: Cichlidogyrus nshomboi (Monogenea: Dactylogyridae) from representatives of Boulengerochromini and Perissodini. Evol. Biol. 2022, 49, 221–238. [Google Scholar] [CrossRef]
- McCoy, K.D. Sympatric speciation in parasites—What is sympatry? Trends Parasitol. 2003, 19, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Šimková, A.; Serbielle, C.; Pariselle, A.; Vanhove, M.P.M.; Morand, S. Speciation in Thaparocleidus (Monogenea: Dactylogyridae) parasitizing Asian Pangasiid catfishes. Biomed Res. Int. 2013, 2013, 353956. [Google Scholar] [CrossRef]
- Řehulková, E.; Seifertová, M.; Přikrylová, I.; Francová, K. Monogenea. In A Guide to the Parasites of African Freshwater Fishes; Scholz, T., Vanhove, M.P.M., Smit, N., Jayasundera, Z., Gelnar, M., Eds.; AbcTaxa: Brussels, Belgium, 2018; pp. 185–243. [Google Scholar]
- Poulin, R.; Morand, S. The diversity of parasites. Q Rev. Biol. 2000, 75, 277–293. [Google Scholar] [CrossRef]
- Šimková, A.; Morand, S.; Jobet, E.; Gelnar, M.; Verneau, O. Molecular phylogeny of congeneric monogenean parasites (Dactylogyrus): A case of intrahost speciation. Evolution 2004, 58, 1001–1018. [Google Scholar] [CrossRef]
- Vanhove, M.P.M.; Pariselle, A.; Van Steenberge, M.; Raeymaekers, J.A.M.; Hablützel, P.I.; Gillardin, C.; Hellemans, B.; Breman, F.C.; Koblmüller, S.; Sturmbauer, C.; et al. Hidden biodiversity in an ancient lake: Phylogenetic congruence between Lake Tanganyika tropheine cichlids and their monogenean flatworm parasites. Sci. Rep. 2015, 5, 13669. [Google Scholar] [CrossRef]
- Rahmouni, C.; Vanhove, M.P.M.; Šimková, A. Underexplored diversity of gill monogeneans in cichlids from Lake Tanganyika: Eight new species of Cichlidogyrus Paperna, 1960 (Monogenea: Dactylogyridae) from the northern basin of the lake, with remarks on the vagina and the heel of the male copulatory. Parasite Vectors 2017, 10, 591. [Google Scholar] [CrossRef]
- Rahmouni, C.; Vanhove, M.P.M.; Koblmüller, S.; Šimková, A. Molecular phylogeny and speciation patterns in host-specific monogeneans (Cichlidogyrus, Dactylogyridae) parasitizing cichlid fishes (Cichliformes, Cichlidae) in Lake Tanganyika. Int. J. Parasitol. 2022, 52, 359–375. [Google Scholar] [CrossRef]
- Meinilä, M.; Kuusela, J.; Ziȩtara, M.S.; Lumme, J. Initial steps of speciation by geographic isolation and host switch in salmonid pathogen Gyrodactylus salaris (Monogenea: Gyrodactylidae). Int. J. Parasitol. 2004, 34, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Bakke, T.A.; Cable, J.; Harris, P.D. The biology of gyrodactylid monogeneans: The “Russian-doll killers”. Adv. Parasitol. 2007, 64, 161–376. [Google Scholar] [CrossRef] [PubMed]
- Cable, J.; Harris, P.D.; Tinsley, R.C. Melanin deposition in the gut of the monogenean Macrogyrodactylus polypteri Malmberg 1957. Int. J. Parasitol. 1997, 27, 1323–1331. [Google Scholar] [CrossRef] [PubMed]
- Malmberg, G. The excretory systems and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Ark. Zool. 1970, 23, 1–235. [Google Scholar]
- Boeger, W.A.; Kritsky, D.C.; Patella, L.; Bueno-Silva, M. Phylogenetic status and historical origins of the oviparous and viviparous gyrodactylids (Monogenoidea, Gyrodactylidea). Zool. Scr. 2021, 50, 112–124. [Google Scholar] [CrossRef]
- Přikrylová, I.; Barson, M.; Shinn, A.P. Description of Tresuncinidactylus wilmienae gen. et sp. n. (Monogenea: Gyrodactylidae), from the gills of the bulldog, Marcusenius macrolepidotus (Peters) from Lake Kariba, Zimbabwe. Folia Parasitol. 2021, 68, 25. [Google Scholar] [CrossRef]
- Pugachev, O.N.; Gerasev, P.I.; Gussev, A.V.; Ergens, R.; Khotenowsky, I. Guide to Monogenoidea of Freshwater Fish of Palaearctic and Amur Regions; Ledizioni: Milan, Italy, 2010; pp. 361–363. [Google Scholar]
- You, P.; King, S.D.; Ye, F.; Cone, D.K. Paragyrodactylus variegatus n. sp. (Gyrodactylidae) from Homatula variegata (Dabry De Thiersant, 1874) (Nemacheilidae) in central China. J. Parasitol. 2014, 100, 350–355. [Google Scholar] [CrossRef]
- Pálsson, J.; Beverley-Burton, M. Laminiscus n.g. (Monogenea: Gyrodactylidae) from capelin, Mallotus villosus (Müller), (Pisces: Osmeridae) in the northwest Atlantic with redescriptions of L. gussevi n. comb., Gyrodactyloides petruschewskii, and G. andriaschewi. Can. J. Zool. 1983, 61, 298–306. [Google Scholar] [CrossRef]
- Ahyong, S.; Boyko, C.B.; Bailly, N.; Bernot, J.; Bieler, R.; Brandão, S.N.; Daly, M.; De Grave, S.; Gofas, S.; Hernandez, F.; et al. World Register of Marine Species (WoRMS). Available online: https://www.marinespecies.org/ (accessed on 12 March 2023).
- Vercammen Grandjean, P.H. Les trematodes du Lac Kivu Sud (Vermes). Ann. Mus. R. Afr. Centr. Scien. Zool. 1960, 5, 1–171. [Google Scholar]
- Bakke, T.A.; Harris, P.D.; Cable, J. Host specificity dynamics: Observations on gyrodactylid monogeneans. Int. J. Parasitol. 2002, 32, 281–308. [Google Scholar] [CrossRef]
- Boeger, W.A.; Kritsky, D.C.; Pie, M.R. Context of diversification of the viviparous Gyrodactylidae (Platyhelminthes, Monogenoidea). Zool. Scr. 2003, 32, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, M.P.M.; Boeger, W.; Bukinga, F.; Volckaert, F.; Huyse, T.; Pariselle, A. A new species of Gyrodactylus (Monogenea, Gyrodactylidae), an ectoparasite from the endemic Iranocichla hormuzensis (Teleostei, Cichlidae), the only Iranian cichlid. Eur. J. Taxon. 2012, 30, 1–10. [Google Scholar] [CrossRef]
- Benovics, M.; Nejat, F.; Abdoli, A.; Šimková, A. Molecular and morphological phylogeny of host-specific Dactylogyrus parasites (Monogenea) sheds new light on the puzzling Middle Eastern origin of European and African lineages. Parasite Vectors 2021, 14, 372. [Google Scholar] [CrossRef] [PubMed]
- Šimková, A.; Řehulková, E.; Choudhury, A.; Seifertová, M. Host-specific parasites reveal the history and biogeographical contacts of their hosts: The Monogenea of Nearctic cyprinoid fishes. Biology 2022, 11, 229. [Google Scholar] [CrossRef]
- Šimková, A.; Benovics, M.; Rahmouni, I.; Vukić, J. Host-specific Dactylogyrus parasites revealing new insights on the historical biogeography of Northwest African and Iberian cyprinid fish. Parasite Vectors 2017, 10, 589. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Genera, species, references Electronic Version. Available online: https://researcharchive.calacademy.org/ (accessed on 12 March 2023).
- Tan, M.; Armbruster, J.W. Phylogenetic classification of extant genera of fishes of the order Cypriniformes (Teleostei: Ostariophysi). Zootaxa 2018, 4476, 6–39. [Google Scholar] [CrossRef]
- Stout, C.C.; Tan, M.; Lemmon, A.R.; Lemmon, E.M.; Armbruster, J.W. Resolving Cypriniformes relationships using an anchored enrichment approach. BMC Evol. Biol. 2016, 16, 244. [Google Scholar] [CrossRef]
- Skelton, P.H.; Tweddle, D.; Jackson, P.B.N. Cyprinids of Africa. In Cyprinid Fishes: Systematics, Biology and Exploitation; Winfield, I.J., Nelson, J.S., Eds.; Springer: Dordrecht, The Netherlands, 1991; pp. 211–239. ISBN 978-94-011-3092-9. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; p. 601, ISBN 9781119174844. [Google Scholar]
- Vreven, E.J.W.M.N.; Musschoot, T.; Snoeks, J.; Schliewen, U.K. The African hexaploid Torini (Cypriniformes: Cyprinidae): Review of a tumultuous history. Zool. J. Linn. Soc. 2016, 177, 231–305. [Google Scholar] [CrossRef]
- Yang, L.; Sado, T.; Vincent Hirt, M.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef]
- Borkenhagen, K.; Esmaeili, H.R.; Mohsenzadeh, S.; Shahryari, F.; Gholamifard, A. The molecular systematics of the Carasobarbus species from Iran and adjacent areas, with comments on Carasobarbus albus (Heckel, 1843). Environ. Biol. Fishes 2011, 91, 327–335. [Google Scholar] [CrossRef]
- Borkenhagen, K.; Krupp, F. Taxonomic revision of the genus Carasobarbus Karaman, 1971 (Actinopterygii, Cyprinidae). Zookeys 2013, 339, 1–53. [Google Scholar] [CrossRef] [PubMed]
- Froese, R.; Pauly, D. FishBase World Wide Web Electronic Publication. Available online: http://www.fishbase.org/search.php (accessed on 20 March 2023).
- Tsigenopoulos, C.S.; Kasapidis, P.; Berrebi, P. Phylogenetic relationships of hexaploid large-sized barbs (genus Labeobarbus, Cyprinidae) based on mtDNA data. Mol. Phylogenet. Evol. 2010, 56, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Tsigenopoulos, C.; Durand, J.-D.; Unlu, E.; Berrebi, P. Rapid radiation of the Mediterranean Luciobarbus species (Cyprinidae) after the Messinian salinity crisis of the Mediterranean Sea, inferred from mitochondrial phylogenetic analysis. Biol. J. Linn. Soc. 2003, 80, 207–222. [Google Scholar] [CrossRef]
- Doadrio, I. Freshwater fish fauna of North Africa and its biogeography. Ann. Mus. R. Cent. Afr. Zool. 1994, 275, 21–34. [Google Scholar]
- Krijgsman, W.; Hilgen, F.J.; Raffi, I.; Sierro, F.J.; Wilson, D.S. Chronology, causes and progression of the Messinian salinity crisis. Nature 1999, 400, 652–655. [Google Scholar] [CrossRef]
- Rosenbaum, G.; Lister, G.; Duboz, C. Reconstruction of the tectonic evolution of the Western Mediterranean since the Oligocene. J. Virtual Explor. 2001, 8, 107–130. [Google Scholar] [CrossRef]
- Machordom, A.; Doadrio, I. Evidence of a Cenozoic Betic–Kabilian connection based on freshwater fish phylogeography (Luciobarbus, Cyprinidae). Mol. Phylogenet. Evol. 2001, 18, 252–263. [Google Scholar] [CrossRef]
- Mesquita, N.; Cunha, C.; Carvalho, G.R.; Coelho, M.M. Comparative phylogeography of endemic cyprinids in the south-west Iberian Peninsula: Evidence for a new ichthyogeographic area. J. Fish Biol. 2007, 71, 45–75. [Google Scholar] [CrossRef]
- Casal-López, M.; Doadrio, I. The Messinian imprint on the evolution of freshwater fishes of the genus Luciobarbus Heckel, 1843 (Teleostei, Cyprinidae) in the western Mediterranean. J. Biogeogr. 2018, 45, 1593–1603. [Google Scholar] [CrossRef]
- Machordom, A.; Doadrio, I.; Berrebi, P. Phylogeny and evolution of the genus Barbus in the Iberian Peninsula as revealed by allozyme electrophoresis. J. Fish Biol. 1995, 47, 211–236. [Google Scholar] [CrossRef]
- Zardoya, R.; Doadrio, I. Phylogenetic relationships of Iberian cyprinids: Systematic and biogeographical implications. Proc. Biol. Sci. 1998, 265, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Doadrio, I.; Casal-López, M.; Perea, S.; Yahyaoui, A. Taxonomy of rheophilic Luciobarbus Heckel, 1842 (Actinopterygii, Cyprinidae) from Morocco with the description of two new species. Graellsia 2016, 72, e039. [Google Scholar] [CrossRef]
- Casal-López, M.; Perea, S.; Yahyaoui, A.; Doadrio, I. Taxonomic review of the genus Luciobarbus Heckel, 1843 (Actinopterygii, Cyprinidae) from Northwestern Morocco with the description of three new species. Graellsia 2015, 71, e027. [Google Scholar] [CrossRef]
- Brahimi, A.; Libois, R.; Henrard, A.; Freyhof, J. Luciobarbus Lanigarensis and L. Numidiensis, two new species of barbels from the Mediterranean Sea Basin in North Africa (Teostei: Cyprinidae). Zootaxa 2018, 4433, 542–560. [Google Scholar] [CrossRef]
- Crespo-Blanc, A.; Comas, M.; Balanyá, J. Clues for a Tortonian reconstruction of the Gibraltar Arc: Structural pattern, deformation, diachronism and block rotations. Tectonophysics 2016, 683, 308–324. [Google Scholar] [CrossRef]
- Benovics, M.; Vukić, J.; Šanda, R.; Rahmouni, I.; Šimková, A. Disentangling the evolutionary history of peri-Mediterranean cyprinids using host-specific gill monogeneans. Int. J. Parasitol. 2020, 50, 969–984. [Google Scholar] [CrossRef]
- Shigoley, M.; Rahmouni, I.; Louizi, H.; Pariselle, A.; Vanhove, M.P.M. First study on Gyrodactylus (Monogenea: Gyrodactylidae) in Morocco, with description of a New species from cyprinids (Actinopterygii: Cyprinidae). Animals 2023, 13, 1624. [Google Scholar] [CrossRef]
- Gutiérrez-Galindo, J.F.; Lacasa-Millán, M.I. Study of the Monogenea of Cyprinidae in the Llobregat River, Northeastern Spain. 11. Species composition on Barbus graellsii Steindachner, 1866. Rev. Ibérica Parasitol. 2001, 61, 91–96. [Google Scholar]
- Malmberg, G. On the Occurrence of Gyrodactylus on Swedish Fishes. Skr. Utgivna Av Södra Sver. Fisk (In Swedish). 1957, 19–76. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Ondračková, M.; Seifertová, M.; Bryjová, A.; Leis, E.; Jurajda, P. Morphometric and genetic evidence for cryptic diversity in Gyrodactylus (Monogenea) infecting non-native European populations of Ameiurus nebulosus and A. melas. Parasitology 2020, 147, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Ø.; Harper, D.; Ryan, P. Past: Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 9. [Google Scholar] [CrossRef]
- Rahmouni, C.; Seifertová, M.; Simková, A. Revealing the hidden diversity of Gyrodactylus communities (Monogenea: Gyrodactylidae) from Nearctic Catostomidae and Leuciscidae fish hosts (Teleostei: Cypriniformes), with description of ten new species. Parasite 2023, 30, 40. [Google Scholar] [CrossRef]
- Ziętara, M.S.; Arndt, A.; Geets, A.; Hellemans, B.; Volckaert, F.A.M. The Nuclear rDNA Region of Gyrodactylus arcuatus and G. branchicus (Monogenea: Gyrodactylidae). J. Parasitol. 2000, 86, 1368–1373. [Google Scholar] [CrossRef]
- Matějusova, I.; Gelnar, M.; McBeath, A.J.; Collins, C.M.; Cunningham, C.O. Molecular markers for gyrodactylids (Gyrodactylidae: Monogenea) from five fish families (Teleostei). Int. J. Parasitol. 2001, 31, 738–745. [Google Scholar] [CrossRef]
- Cone, D.; Abbott, C.; Gilmore, S.; Burt, M. A new genus and species of gyrodactylid (Monogenea) from silver hake, Merluccius bilinearis, in the bay of fundy, New Brunswick, Canada. J. Parasitol. 2010, 96, 681–684. [Google Scholar] [CrossRef]
- Rokicka, M.; Lumme, J.; Ziętara, M.S. Identification of Gyrodactylus ectoparasites in Polish salmonid farms by PCR-RFLP of the nuclear ITS segment of ribosomal DNA (Monogenea, Gyrodactylidae). Acta Parasitol. 2007, 52, 185–195. [Google Scholar] [CrossRef]
- Cunningham, C.O. Species variation within the internal transcribed spacer (ITS) Region of Gyrodactylus (Monogenea:Gyrodactylidae) ribosomal RNA genes. J. Parasitol. 1997, 83, 215–219. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.4. Molecular Evolution, Phylogenetetics and Epidemiology. Available online: https://tree.bio.ed.ac.uk/ (accessed on 16 July 2023).
- Müller, J.; Müller, K. TREEGRAPH: Automated drawing of complex tree figures using an extensible tree description format. Mol. Ecol. Notes 2004, 4, 786–788. [Google Scholar] [CrossRef]
- Přikrylová, I.; Vanhove, M.P.M.; Janssens, S.B.; Billeter, P.A.; Huyse, T. Tiny worms from a mighty continent: High diversity and new phylogenetic lineages of African monogeneans. Mol. Phylogenet. Evol. 2013, 67, 43–52. [Google Scholar] [CrossRef]
- Matějusová, I.; Gelnar, M.; Verneau, O.; Cunningham, C.O.; Littlewood, D.T.J. Molecular phylogenetic analysis of the genus Gyrodactylus (Platyhelminthes: Monogenea) inferred from rDNA ITS region: Subgenera versus species groups. Parasitology 2003, 127, 603–611. [Google Scholar] [CrossRef]
- Přikrylová, I.; Blažek, R.; Vanhove, M.P.M. An overview of the Gyrodactylus (Monogenea: Gyrodactylidae) species parasitizing African catfishes, and their morphological and molecular diversity. Parasitol. Res. 2012, 110, 1185–1200. [Google Scholar] [CrossRef]
- Gilmore, S.R.; Cone, D.K.; Lowe, G.; King, S.K.; Jones, S.R.M.; Abbott, C.L. Molecular phylogeny of Gyrodactylus (Monogenea) parasitizing fishes in fresh water, estuarine, and marine habitats in Canada. Can. J. Zool. 2012, 90, 776–786. [Google Scholar] [CrossRef]
- Ziętara, M.; Lumme, J. Comparison of molecular phylogeny and morphological systematics in fish parasite genus Gyrodactylus Nordmann, 1832 (Monogenea, Gyrodactylidae). Zool. Pol. 2004, 49, 5–28. [Google Scholar]
- Leis, E.; King, S.; Leis, S.; Cone, D. Infections of Gyrodactylus Crysoleucas and Gyrodactylus sp. (Monogenea) at a Golden Shiner (Notemigonus Crysoleucas) Farm in Minnesota. Comp. Parasitol. 2016, 83, 105–110. [Google Scholar] [CrossRef]
- Ziętara, M.S.; Lumme, J. Speciation by host switch and adaptive radiation in a fish parasite genus Gyrodactylus (Monogenea, Gyrodactylidae). Evolution 2002, 56, 2445–2458. [Google Scholar] [CrossRef] [PubMed]
- Benovics, M.; Rahmouni, C.; Rehulková, E.; Nejat, F.; Simková, A. Uncovering the monogenean species diversity of cyprinoid fish (Cypriniformes: Cyprinidae) in Iraq using integrative approach. Parasitology 2023, in press. [Google Scholar]
- Leis, E.; Easy, R.; Cone, D. A variant of Gyrodactylus mediotorus King, Marcogliese, Forest, McLaughlin and Bentzen, 2013 (Monogenea) identified from weed shiner (Notropis texanus) in Wisconsin portions of the upper Mississippi River. Syst. Parasitol. 2021, 98, 247–253. [Google Scholar] [CrossRef]
- Huyse, T.; Audenaert, V.; Volckaert, F.A.M. Speciation and host-parasite relationships in the parasite genus Gyrodactylus (Monogenea, Platyhelminthes) infecting gobies of the genus Pomatoschistus (Gobiidae, Teleostei). Int. J. Parasitol. 2003, 33, 1679–1689. [Google Scholar] [CrossRef]
- Huyse, T.; Volckaert, F.A.M. Identification of a host-associated species complex using molecular and morphometric analyses, with the description of Gyrodactylus Rugiensoides n. sp. (Gyrodactylidae, Monogenea). Int. J. Parasitol. 2002, 32, 907–919. [Google Scholar] [CrossRef]
- Cunningham, C.O.; McGillivray, D.M.; MacKenzie, K.; Melvin, W.T. Discrimination between Gyrodactylus Salaris, G. Derjavini and G. Truttae (Platyhelminthes: Monogenea) using restriction fragment length polymorphisms and an oligonucleotide probe within the small subunit ribosomal RNA gene. Parasitology 1995, 111, 87–94. [Google Scholar] [CrossRef]
- Leis, E.; Chi, T.K.; Lumme, J. Global phylogeography of salmonid ectoparasites of the genus Gyrodactylus, with an emphasis on the origin of the circumpolar Gyrodactylus Salmonis (Platyhelminthes: Monogenea). Comp. Parasitol. 2021, 88, 130–143. [Google Scholar] [CrossRef]
- King, S.D.; Cone, D.K. Morphological and molecular taxonomy of a new species of Fundulotrema and comments on Gyrodactylus Stephanus (Monogenea: Gyrodactylidae) from Fundulus Heteroclitus (Actinopterygii: Cyprinodontiformes) in Nova Scotia, Canada. J. Parasitol. 2009, 95, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Kritsky, D.C.; Boeger, W.A.; Mendoza-Franco, E.F.; Vianna, R.T. Neotropical Monogenoidea. 57. Revision and phylogenetic position of Scleroductus Jara & Cone, 1989 (Gyrodactylidae), with descriptions of new species from the Guatemalan chulin Rhamdia Guatemalensis (Günther) (Siluriformes: Heptapteridae) in Mexico and the barred sorubim Pseudoplatystoma fasciatum (Linnaeus) (Siluriformes: Pimelodidae) in Brazil. Syst. Parasitol. 2013, 84, 1–15. [Google Scholar] [CrossRef]
- Schelkle, B.; Paladini, G.; Shinn, A.P.; King, S.; Johnson, M.; van Oosterhout, C.; Mohammed, R.S.; Cable, J. Ieredactylus Rivuli Gen. et Sp. Nov. (Monogenea, Gyrodactylidae) from Rivulus Hartii (Cyprinodontiformes, Rivulidae) in Trinidad. Acta Parasitol. 2011, 56, 360–370. [Google Scholar] [CrossRef]
- Barson, M.; Přikrylová, I.; Vanhove, M.P.M.; Huyse, T. Parasite hybridization in African Macrogyrodactylus spp. (Monogenea, Platyhelminthes) signals historical host distribution. Parasitology 2010, 137, 1585–1595. [Google Scholar] [CrossRef]
- Harris, P.D.; Shinn, A.P.; Cable, J.; Bakke, T.A. Nominal species of the genus Gyrodactylus von Nordmann 1832 (Monogenea: Gyrodactylidae), with a list of principal host species. Syst. Parasitol. 2004, 59, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Ergens, R. Zwei weitere befunde der Gyrodactylus-Art (Monogenoidea) aus der Tschechoslowakei. Věstník Ceskoslov. Společnosti Zool. 1961, 25, 25–27. [Google Scholar]
- Ziȩtara, M.S.; Lumme, J. The crossroads of molecular, typological and biological species concepts: Two new species of Gyrodactylus Nordmann, 1832 (Monogenea: Gyrodactylidae). Syst. Parasitol. 2003, 55, 39–52. [Google Scholar] [CrossRef]
- Huyse, T.; Malmberg, G.; Volckaert, F.A.M. Four new species of Gyrodactylus von Nordmann, 1832 (Monogenea, Gyrodactylidae) on gobiid fishes: Combined DNA and morphological analyses. Syst. Parasitol. 2004, 59, 103–120. [Google Scholar] [CrossRef]
- Cable, J.; Harris, P.D.; Tinsley, R.C.; Lazarus, C.M. Phylogenetic analysis of Gyrodactylus spp. (Platyhelminthes: Monogenea) using ribosomal DNA sequences. Can. J. Zool. 1999, 77, 1439–1449. [Google Scholar] [CrossRef]
- Gettová, L.; Gilles, A.; Šimková, A. Metazoan parasite communities: Support for the biological invasion of Barbus Barbus and its hybridization with the endemic Barbus meridionalis. Parasit. Vectors 2016, 9, 588. [Google Scholar] [CrossRef]
- Rahmouni, I.; Řehulková, E.; Pariselle, A.; Rkhami, O.B.; Šimková, A. Four new species of Dactylogyrus Diesing, 1850 (Monogenea: Dactylogyridae) parasitising the gills of northern Moroccan Luciobarbus Heckel (Cyprinidae): Morphological and molecular characterisation. Syst. Parasitol. 2017, 94, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Benovics, M.; Desdevises, Y.; Šanda, R.; Vukić, J.; Scheifler, M.; Doadrio, I.; Sousa-Santos, C.; Šimková, A. High diversity of fish ectoparasitic monogeneans (Dactylogyrus) in the Iberian Peninsula: A case of adaptive radiation? Parasitology 2020, 147, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Řehulková, E.; Rahmouni, I.; Pariselle, A.; Šimková, A. Integrating morphological and molecular approaches for characterizing four species of Dactylogyrus (Monogenea: Dactylogyridae) from Moroccan cyprinids, with comments on their host specificity and phylogenetic relationships. PeerJ 2021, 9, e10867. [Google Scholar] [CrossRef] [PubMed]
- Truter, M.; Smit, N.J.; Malherbe, W.; Přikrylová, I. Description of Gyrodactylus paludinosus sp. nov. (Monogenea: Gyrodactylidae) from the Straightfin Barb, Enteromius paludinosus (Peters, 1852), in South Africa. Acta Parasitol. 2021, 67, 446–453. [Google Scholar] [CrossRef]
- Přikrylová, I.; Shinn, A.P.; Paladini, G. Description of Citharodactylus gagei n. gen. et n. sp. (Monogenea: Gyrodactylidae) from the moon fish, Citharinus citharus (Geoffroy Saint-Hilaire), from Lake Turkana. Parasitol. Res. 2016, 116, 281–292. [Google Scholar] [CrossRef]
- Bueno-Silva, M.; Boeger, W.A.; Pie, M.R. Choice matters: Incipient speciation in Gyrodactylus corydori (Monogenoidea: Gyrodactylidae). Int. J. Parasitol. 2011, 41, 657–667. [Google Scholar] [CrossRef]
- Benovics, M.; Vukić, J.; Šanda, R.; Nejat, F.; Charmpila, E.A.; Buj, I.; Shumka, S.; Porcelloti, S.; Tarkan, S.A.; Aksu, S.; et al. Monogeneans and chubs: Ancient host-parasite system under the looking glass. Mol. Phylogenet. Evol. 2023, 179, 107667. [Google Scholar] [CrossRef]
- Cunningham, C.O.; Mo, T.A.; Collins, C.M.; Buchmann, K.; Thiery, R.; Blanc, G.; Lautraite, A. Redescription of Gyrodactylus teuchis Lautraite, Blanc, Thiery, Daniel & Vigneulle, 1999 (Monogenea: Gyrodactylidae); a species identified by ribosomal RNA sequence. Syst. Parasitol. 2001, 48, 141–150. [Google Scholar] [CrossRef]
- Dmitrieva, E.; Sanna, D.; Vodiasova, E.; Prokhorova, D.; Casu, M.; Burreddu, C.; Piras, M.C.; Garippa, G.; Merella, P. Morphological and genetic variability of the cryptic Gyrodactylus sphinx and Gyrodactylus gerasevi n. sp. (Platyhelminthes: Monogenea) from the Mediterranean Sea and Black Sea: Two new members of the cross-ocean distributed Gyrodactylus orecchiae species. J. Helminthol. 2022, 96, e9. [Google Scholar] [CrossRef]
- Ouassal, K.; Doadrio, I.; Perea, S.; Casal-López, M.; Yahyaoui, A.; Jaziri, H. Mitochondrial genetic diversity, phylogeography and historical demography of Moroccan native freshwater fishes: A case study of the genus Luciobarbus Heckel, 1843. Biologia 2022, 77, 2183–2205. [Google Scholar] [CrossRef]
- Vanhove, M.P.M.; Briscoe, A.G.; Jorissen, M.W.P.; Littlewood, D.T.J.; Huyse, T. The first next-generation sequencing approach to the mitochondrial phylogeny of African monogenean parasites (Platyhelminthes: Gyrodactylidae and Dactylogyridae). BMC Genom. 2018, 19, 520. [Google Scholar] [CrossRef] [PubMed]





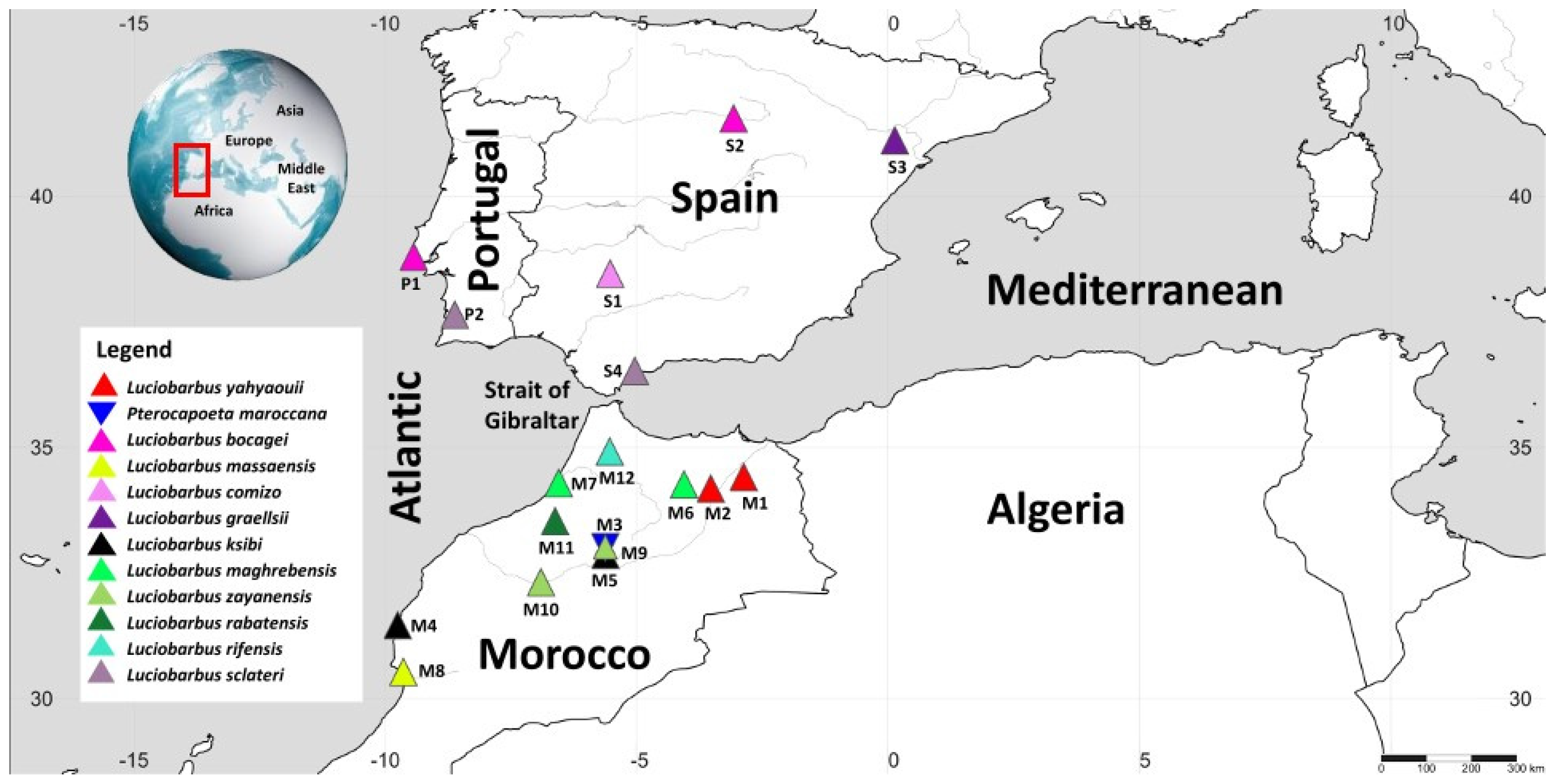
| Host Subfamily | Host Species | Total Hosts | Total Worms | Sampling Locality | Abbreviation on the Map | GPS Coordinates | Country | Gyrodactylus spp. | Prevalence (%) | Intensity of Infection |
|---|---|---|---|---|---|---|---|---|---|---|
| Torinae Karaman, 1971 | Pterocapoeta maroccana Günther, 1902 | 03 | 14 | Oum Er-Rbia River (El Borj) | M3 | 33°00′58.07″ N 05°37′48.06″ W | Morocco | Gyrodactylus sp. 5 | 33 | 1–6 |
| Barbinae Bleeker, 1859 | Luciobarbus bocagei (Steindachner, 1864) | 15 | 03 | Colares | P1 | 38°47′53.37″ N 09°26′14.16″ W | Portugal | Gyrodactylus sp. 1 | 7 | 3 |
| 05 | Gyrodactylus sp. 4 | 13 | 2–3 | |||||||
| 10 | 08 | Rio Ucera | S2 | 41°32′49.11″ N 03°04′32.50″ W | Spain | Gyrodactylus sp. 1 | 30 | 2–4 | ||
| 11 | Gyrodactylus sp. 4 | 60 | 1–3 | |||||||
| Luciobarbus comizo (Steindachner, 1864) | 11 | 25 | Peraleda de Zancejo, Rio Zujar | S1 | 38°27′12.02″ N 05°31′59.67″ W | Spain | Gyrodactylus sp. 3 | 27 | 5–14 | |
| Luciobarbus graellsii (Steindachner, 1866) | 12 | 10 | upstream Mella, tributary of Materraña | S3 | 41°06′41.00″ N 00°08′05.00″ E | Spain | G. gibraltarensis sp. nov. | 16 | 1–9 | |
| Luciobarbus yahyaouii Doadrio, Casal-Lopez & Perea, 2016 | 09 | 04 | Za River | M1 | 34°24′38.09″ N 02°52′29.1″ W | Morocco | G. moroccensis sp. nov. | 11 | 1–2 | |
| 05 | 04 | Meloulou River | M2 | 34°10′51.07″ N 03°31′59.06″ W | Gyrodactylus sp. 10 | 60 | 1–2 | |||
| Luciobarbus sclateri (Günther, 1868) | 10 | 0 | Torgal river, Mira basin | P2 | 37°38′16.76″ N 08°37′10.58″ W | Portugal | - | - | - | |
| 10 | 10 | Benahavis, Rio Guadalmina | S4 | 36°31′03.45″ N 05°02′25.07″ W | Spain | Gyrodactylus sp. 2 | 20 | 4–6 | ||
| Luciobarbus ksibi Doadrio, Perea and Yahyaoui, 2015 | 6 | 5 | Ksob | M4 | 31°27′50.07″ N 09°45′25.03″ W | Morocco | G. pseudomoroccensis sp. nov. | 45 | 1–3 | |
| 9 | 0 | Oum Er-Rbia River (Chakouba) | M5 | 32°51′32.09″ N 05°37′18.09″ W | - | - | - | |||
| Luciobarbus maghrebensis Doadrio, Perea and Yahyaoui, | 05 | 02 | Lahdar River | M6 | 34°15′30.01″ N 04°03′52.01″ W | Morocco | G. moroccensis sp. nov. | 20 | 2 | |
| 05 | 28 | Sebou River | M7 | 34°17′14.2″ N 06°33′14.08″ W | 20 | 1–23 | ||||
| Luciobarbus massaensis (Pellegrin, 1922) | 11 | 4 | Tamrhakht | M8 | 30°31′33.06″ N 09°38′53.06″ W | Morocco | Gyrodactylus sp. 11 | 45 | 2 | |
| 4 | Gyrodactylus sp. 8 | 75 | 2 | |||||||
| Luciobarbus rabatensis Doadrio, Perea and Yahyaoui, 2015 | 11 | 18 | Maleh River | M11 | 33°31′58.00″ N 06°37′39.06″ W | Morocco | G. moroccensis sp. nov. | 72 | 1–9 | |
| 22 | Gyrodactylus sp. 7 | 27 | 1–17 | |||||||
| Luciobarbus rifensis Doadrio, Casal-Lopez & Yahyaoui, 2015 | 10 | 02 | Tributary of Loukkos | M12 | 34°54′57.02″ N 05°32′17.02″ W | Morocco | G. moroccensis sp. nov. | 10 | 2 | |
| 06 | Gyrodactylus sp. 9 | 50 | 2 | |||||||
| Luciobarbus zayanensis Doadrio, Casal-López & Yahyaoui, 2016 | 06 | 03 | Oum Er-Rbia River (El Borj) | M9 | 33°00′58.07″ N 05°37′48.06″ W | Morocco | G. moroccensis sp. nov. | 13 | 2 | |
| 04 | Oum Er-Rbia River (El Borj) | M9 | 33°00′58.07″ N 05°37′48.06″ W | Gyrodactylus sp. 6 | 50 | 2 | ||||
| 09 | 0 | Oum Er-Rbia River (Dar Oul Zidouh) | M10 | 32°18′54.00″ N 06°54′28.07″ W | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahmouni, C.; Seifertová, M.; Benovics, M.; Šimková, A. Diversity and Phylogeny of Gyrodactylus spp. (Monogenea: Gyrodactylidae) across the Strait of Gibraltar: Parasite Speciation and Historical Biogeography of West Mediterranean Cyprinid Hosts. Diversity 2023, 15, 1152. https://doi.org/10.3390/d15111152
Rahmouni C, Seifertová M, Benovics M, Šimková A. Diversity and Phylogeny of Gyrodactylus spp. (Monogenea: Gyrodactylidae) across the Strait of Gibraltar: Parasite Speciation and Historical Biogeography of West Mediterranean Cyprinid Hosts. Diversity. 2023; 15(11):1152. https://doi.org/10.3390/d15111152
Chicago/Turabian StyleRahmouni, Chahrazed, Mária Seifertová, Michal Benovics, and Andrea Šimková. 2023. "Diversity and Phylogeny of Gyrodactylus spp. (Monogenea: Gyrodactylidae) across the Strait of Gibraltar: Parasite Speciation and Historical Biogeography of West Mediterranean Cyprinid Hosts" Diversity 15, no. 11: 1152. https://doi.org/10.3390/d15111152