

Chitons from Deep-Water Mollusk-Rich Deposits in the Southwestern Adriatic Sea (Mollusca, Polyplacophora) †

1
National Research Council (CNR-ISMAR), Institute of Marine Sciences, Via Gobetti 101, 40129 Bologna, Italy
2
Stazione Zoologica Anton Dohrn, Villa Comunale, 80121 Napoli, Italy
3
Independent Researcher, 16153 Genova, Italy
4
Independent Researcher, 16154 Genova, Italy
*
Author to whom correspondence should be addressed.
†
urn:lsid:zoobank.org:pub:C8C96D58-C4E5-4D63-AB5B-8FF26D308718.
Diversity 2023, 15(3), 359; https://doi.org/10.3390/d15030359
Submission received: 20 December 2022
/
Revised: 22 February 2023
/
Accepted: 24 February 2023
/
Published: 2 March 2023
(This article belongs to the Special Issue Diversity in 2022)
Abstract
:Sea-bottom sampling at bathyal depths off the Apulian margin (southwestern Adriatic Sea) recovered shelly biogenic sediments rich in mollusk remains. Noteworthy is the occurrence in such taphocoenoses of loose chiton valves, rarely reported in these environments. We identified four species of Polyplacophora, Leptochiton asellus (Gmelin, 1791), Belknapchiton alveolus (M. Sars MS, Lovén, 1846), Hanleya hanleyi (Bean in Thorpe, 1844), and the new species Leptochiton antondohrni. L. asellus (Gmelin, 1791) and B. alveolus are primarily of North Atlantic (boreo-celtic) affinity and thus considered to belong to last Pleistocene glacial assemblages, whose presence in the study area has long been ascertained. The same could be true for the valves of H. hanleyi and, possibly, Leptochiton antondohrni n. sp. Finally, we updated the list of mollusks found thus far in the deep-sea taphocoenoses of this area, including new records of North Atlantic-type taxa considered at present extinct in the Mediterranean basin.
1. Introduction
The deep-water environments of the southwestern Adriatic Sea (Mediterranean basin) have been the focus of intense investigation for decades. The crucial area is the Canyon Bari System, where a dramatic incision indents the Apulian margin down to the plain, cutting through terranes of different ages. In the past, part of this research was mostly devoted to collecting valuable geological and sedimentological information through sea-bottom sampling by using grab, corer and dredge gear [1,2]. With time, a less invasive approach prevailed, and priority was given to detailed geomorphological mapping and, occasionally, remotely operated vehicle observation (e.g., [3,4,5]). This area is also known to host lush charismatic cold water coral life [4,5,6,7,8] and their late Pleistocene legacy [9].
The earlier studies were pivotal in the discovery of diverse benthic faunas that encompassed both recent and pre-modern elements [10]. Besides some classic North Atlantic boreo-celtic ‘cold guests’ such as Buccinum humphreysianum, B. undatum, and Pseudamussium peslutrae [2,10,11], the occurrence of deep-water last glacial Pleistocene (ca. 115,000–11,700 years BP; defined at the time as ‘würmian’) mollusks in the submerged taphocoenoses (e.g., Puncturella noachina, Iothia fulva) was recorded for the first time, based upon the fraction residue of some dredge samples [12,13]. A significant contribution included taxonomic lists for particular submerged taphocoenoses [10,13], as extended by others to adjacent analog deposits [14].
In 2007, an additional survey was carried out in the area during cruise SETE-06 on-board RV Urania. A limited number of sea-bottom samples was taken, including also dredging the flanks of the Bari Canyon. Biogenic-rich fractions were noteworthy for their inclusion of mollusk shells representing new records for the area; these included plates from polyplacophorans (chitons) whose presence in this habitat is of ecological interest, and also included a species described here as new. This paper updates the taxonomic knowledge on the chitons and associated mollusks of this key-sector of the Mediterranean basin and discusses the paleoclimatic implications of these findings.
2. Material and Methods
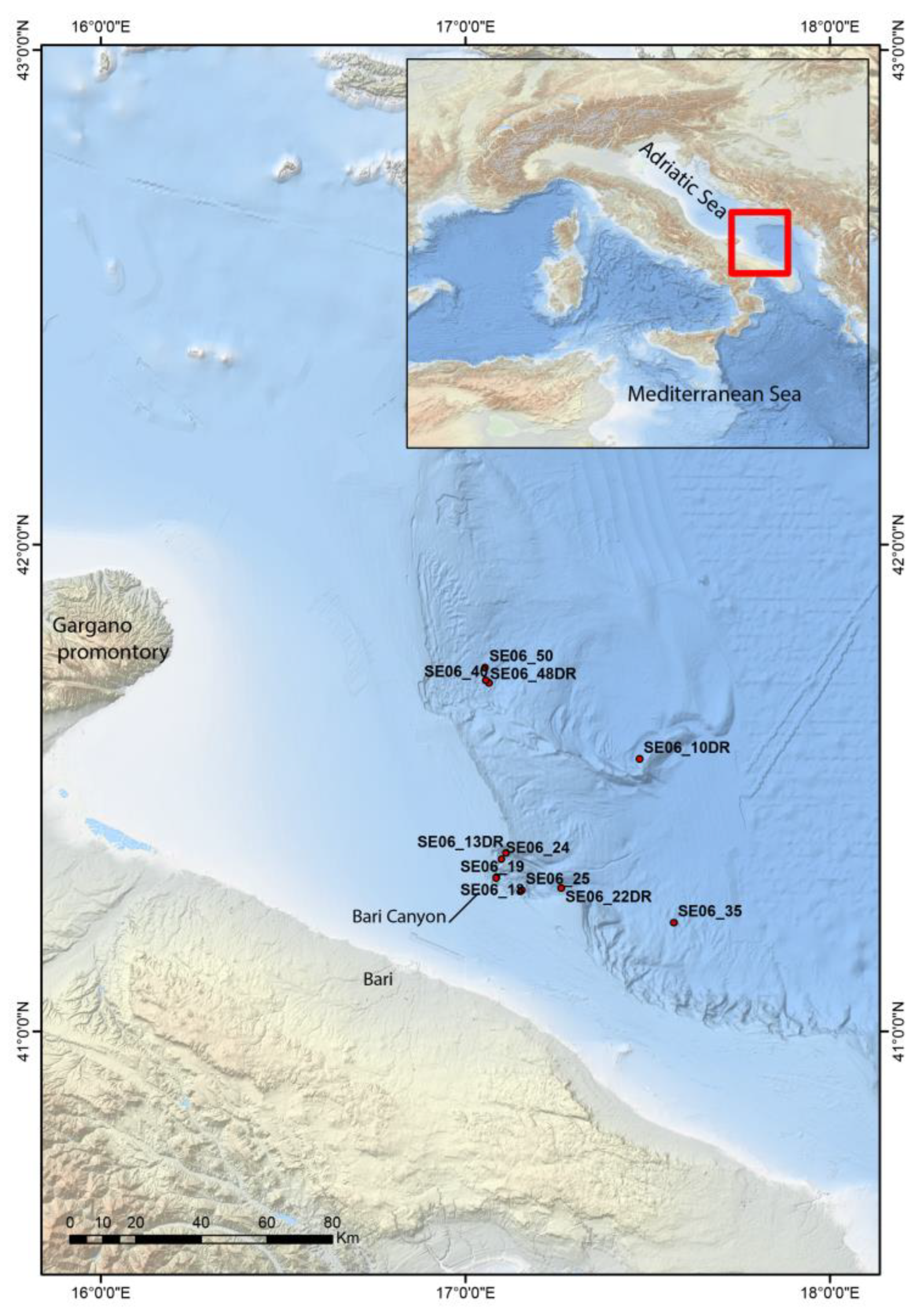
Cruise SETE-06 took place in the southwestern Adriatic Sea in spring of 2006 on-board RV Urania [15]. The sampling stations (code SE06) providing the material discussed in this paper are reported in Figure 1; their precise geographic position, depth, and type of gear are provided in Table 1. A share of the collected sediments was firstly washed onboard over 1 mm mesh, rinsed in freshwater, and dried in the oven at 30 °C. The resulting fraction was later analyzed in the laboratory under a binocular microscope and individual taxa were picked up and classified at the lowest taxonomic level whenever possible. Scanning Electron Microscopy (S.E.M.) was performed on chiton plates using the facility at the IMM-CNR, Bologna and at the BiGeA, University of Bologna. Type material and figured specimens are currently held in the malacological collection of the Museum of Evolution (formerly Zoology) of the University of Bologna (code MZUB), and voucher specimens in the Ismar-CNR, Bologna repository.
The following abbreviations are used:
AA—antemucronal area, tail valve; CA—central area, intermediate valve; H—height of the valve (mm); HV—head valve; L—length of the valve (mm); LA—lateral areas, intermediate valve; PA—postmucronal area, tail valve; W—width of the valve (mm).
3. Systematics
Class Polyplacophora Gray, 1821
Order Lepidopleurida Thiele, 1909
Family Leptochitonidae Dall, 1889
Genus Leptochiton Gray, 1847
Type species. Chiton cinereus Montagu, 1803 (misapplication of name), by subsequent designation [16], non Linnaeus, 1767 (=Chiton asellus Gmelin, 1791).
Distribution. Leptochiton is known to have existed from the Triassic to the present-day, with a living worldwide distribution, most species being from the northeastern Atlantic and the Mediterranean Sea. Fossils determined as Leptochiton have been found worldwide, and Leptochiton is one of the most ancient genera of “modern” polyplacophorans, dating back to the Triassic [17], to the Jurassic of France, Germany, Poland, Russia and Siberia [18], and possibly even to the Lower Carboniferous [18].
Leptochiton asellus(Gmelin, 1791)
Lepidopleurus (Leptochiton) asellus; Malatesta, 1962: 150, figures 5 and 6 [21]; Dell’Angelo and Palazzi, 1989: 56, plate 5 [22]; Dell’Angelo and Smriglio, 1999; 43, pls 8–9, figures 16 and 17 [23]; Dell’Angelo and Giusti, 2000: 53, figures 1–4 [24].
Leptochiton (L.) asellus; Kaas and Van Belle, 1985: 39, figure 15 [27]; Sturrock and Baxter, 1993: 49, plates 1–6 [28].
Type material. Holotype at the Zoological Museum of the University of Copenhagen, a specimen attached to a shell of Modiolus [20] (figure 26).
Type locality. Norway, Telemark, Kragerö.
Material examined.
SE06-10: 1 intermediate valve; SE06-13: 1 head valve.; SE06-18: 2 intermediate valves; SE06-19: 1 intermediate valve; SE06-22: 1 intermediate valve; SE06-24: 1 tail valve, Figure 2J–L; SE06-25: 2 intermediate valves; SE06-35: 5 valves (2 head, 2 intermediate, 1 tail); SE06-40: 1 head valve, Figure 2C,D; SE06-48: 1 intermediate valve; SE06-50: 25 valves (4 head, 20 intermediate, 1 tail), Figure 2A,B,E–I). Maximum width of the valves (head/intermediate/tail): 3.6/5.2/4.3 mm.
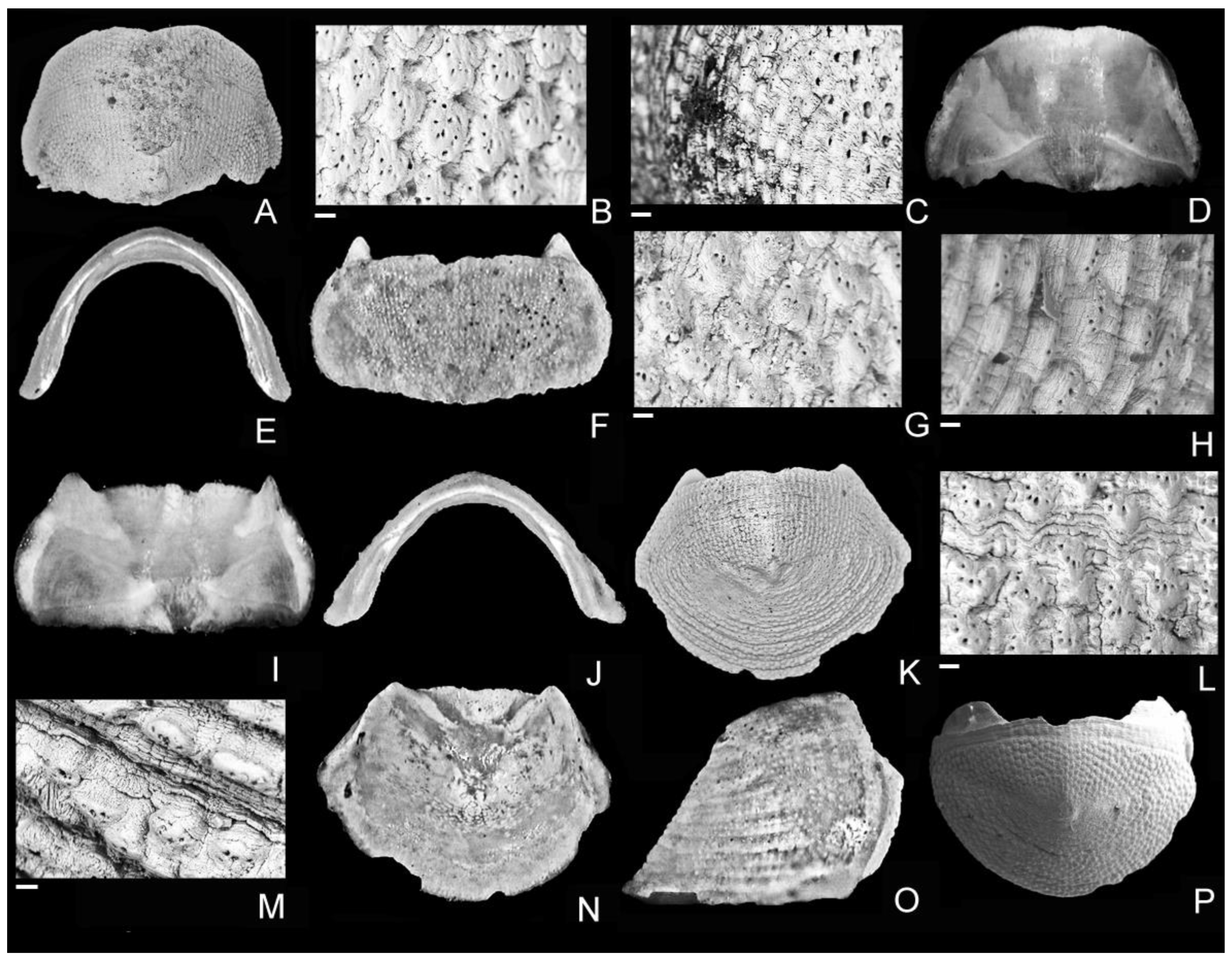
Figure 2.
Leptochiton asellus (Gmelin, 1791); Apulian margin, offshore Bari, submerged deposits, possibly late Pleistocene (last glacial). (A,B) SE06-50, MZUB 60427, head valve, width 3 mm, dorsal view (A) and close-up of surface ornamentation (B). (C,D) SE06-40, MZUB 60428, head valve, width 3.1 mm, dorsal (C) and lateral (D) views. (E–I) SE06-50, MZUB 60429, intermediate valve, width 5.2 mm, dorsal view (E), close-up of surface ornamentation of central area (F,G), ventral (H) and frontal (I) views. (J–L) SE06-24, MZUB 60430, tail valve, width 2.5 mm, dorsal view (J), close-up of surface ornamentation of antemucronal area (K) and lateral view (L). Scale bars = 100 μm (B); 80 μm (K); 20 μm (F); 10 μm (G).
Figure 2.
Leptochiton asellus (Gmelin, 1791); Apulian margin, offshore Bari, submerged deposits, possibly late Pleistocene (last glacial). (A,B) SE06-50, MZUB 60427, head valve, width 3 mm, dorsal view (A) and close-up of surface ornamentation (B). (C,D) SE06-40, MZUB 60428, head valve, width 3.1 mm, dorsal (C) and lateral (D) views. (E–I) SE06-50, MZUB 60429, intermediate valve, width 5.2 mm, dorsal view (E), close-up of surface ornamentation of central area (F,G), ventral (H) and frontal (I) views. (J–L) SE06-24, MZUB 60430, tail valve, width 2.5 mm, dorsal view (J), close-up of surface ornamentation of antemucronal area (K) and lateral view (L). Scale bars = 100 μm (B); 80 μm (K); 20 μm (F); 10 μm (G).

Remarks:Chiton asellus was originally described, although inadequately illustrated, by Chemnitz, 1785 [29] on the basis of a Norwegian specimen attached to a shell of Modiolus, “ex Museo Spengleriano”. This description is not valid, Chemnitz’s work not being strictly binomial. The taxon was validated by Gmelin [19], who added nothing new to the description, and was later better defined by Spengler [30]. This species has a complicated taxonomic history, and a thorough discussion of the interpretations given by various authors was provided by [20,25].
Leptochiton asellus (Gmelin, 1791) is characterized by the tegmentum sculpted with small granules arranged in radiating series on HV (70–80), LA (15–20), and PA, and in longitudinal series on CA (70–80) and AA, where a regular quincuncial pattern is displayed by granules of neighboring rows. Growth lines are often present in variable numbers. Aesthetes have pores of roughly the same size, one central and two to four irregularly arranged. Detailed descriptions of this species are given by [23,25,27].
The finding of Leptochiton asellus in the study area is quite significant. L. asellus was considered a ‘cold guest’ by [3,21]. This species was not considered living in the Mediterranean basin until a few years ago; if some older reports from this area have been recorded (i.e., [31,32]), there are no precise indications of locality. Lucas [33] reports: “Distribution principalement arctique mais l’espèce descend jusqu’à Gibraltar et pénètre en Méditerranée jusqu’aux Baléares et au nord de la côte espagnole”. A recent report [34] confirms the presence of L. asellus in the Mediterranean Sea (from Limnos, Greece, inside an amphora trawled from a depth of 600 m).
Fossil records for this species are scarce; historical records are doubtful and require confirmation. Leptochiton asellus is reported from the Pliocene of the U.K. [35,36,37] and Belgium [38,39] and the Pleistocene of the N. Atlantic [40,41,42], the Netherlands [26], Italy [43], and the Mediterranean Sea (off Capraia/Capo Corso [24]).
Distribution. Lower Pliocene: northeastern Atlantic: U.K. [35,36,37]; North Sea Basin, Belgium: Kallo [38,39]. Pleistocene: North Atlantic: Sweden and Norway [40,41,42], Netherlands [26]; central Mediterranean, Italy: Pezzo [43] and Capraia/Capo Corso-350/500 m [21,24]. Pleistocene, presumably last glacial: Italy: off Bari (this study). Recent: NE Atlantic Ocean: from Spitsbergen and the Barents Sea South along the Scandinavian coasts [44,45]; North Sea; all around the British Isles and Ireland, coasts of France [46], south to Spain [47] and Portugal [48]. Mediterranean Sea: Greece: Limnos Island [34].
Leptochiton antondohrni new species
Type material. Holotype: MZUB 60,418, intermediate valve, width 3.4 mm, Figure 3A–E. Paratypes: MZUB 60,419 intermediate valve, width 2.9 mm, Figure 3F–J; MZUB 60,420, tail valve, width 3.1 mm, Figure 3K–O.
Type locality. Southwestern Adriatic Sea, Apulian margin, off Bari, SE06-50.
Type stage. Late Pleistocene submerged deposits, probably late glacial epoch.
Etymology. We dedicate this new species from the Mediterranean Sea to the ‘Stazione Zoologica Anton Dohrn’ Naples, to honour the 150th of the foundation of the first marine station ever.
ZooBank code. urn:lsid:zoobank.org:pub:C8C96D58-C4E5-4D63-AB5B-8FF26D308718
Material examined. Type material, plus: SE06-10: 1 intermediate valve; SE06-35: 4 intermediate valves; SE06-40: 3 valves (1 intermediate, 2 tail); SE06-48: 2 valves (1 intermediate, 1 tail); SE06-50: 55 valves (54 intermediate, 1 tail), Figure 3P. Maximum width of the valves: --/3.4/3.1 mm.
Description. Head valve unknown. Intermediate valves broadly rectangular (W/L = 1.45–2.03), rounded in anterior profile, elevated (H/W = 0.54–0.66), anterior margin straight, side margins rounded, posterior margin straight, apex inconspicuous, lateral areas hardly or not raised. Tail valve semicircular (W/L = 1.48–1.61), anterior margin almost straight or slightly convex, mucro not prominent, in slightly anterior position, antemucronal slope slightly convex, postmucronal slope almost straight.
Tegmentum sculpture. CA and AA with well raised, rather thick but distinctly separated roundish to oval granules, extended with two or three small longitudinal varices, forming 50–55 longitudinal series with a regular quincuncial pattern displayed by granules of neighboring rows; granules with a maximum width up to 70 µm and the presence of one subcentral aesthete and up to six aesthetes irregularly disposed, with pores of the same width. LA and PA with more rectangular granules, randomly disposed along concentric lines; granules with a maximum width up to 60 µm, and the presence of 3-4 aesthetes more or less aligned, with pores of the same width. Numerous and well-marked growth lines.
Articulamentum without insertion laminae, with small, sharply triangular, widely separated apophyses.
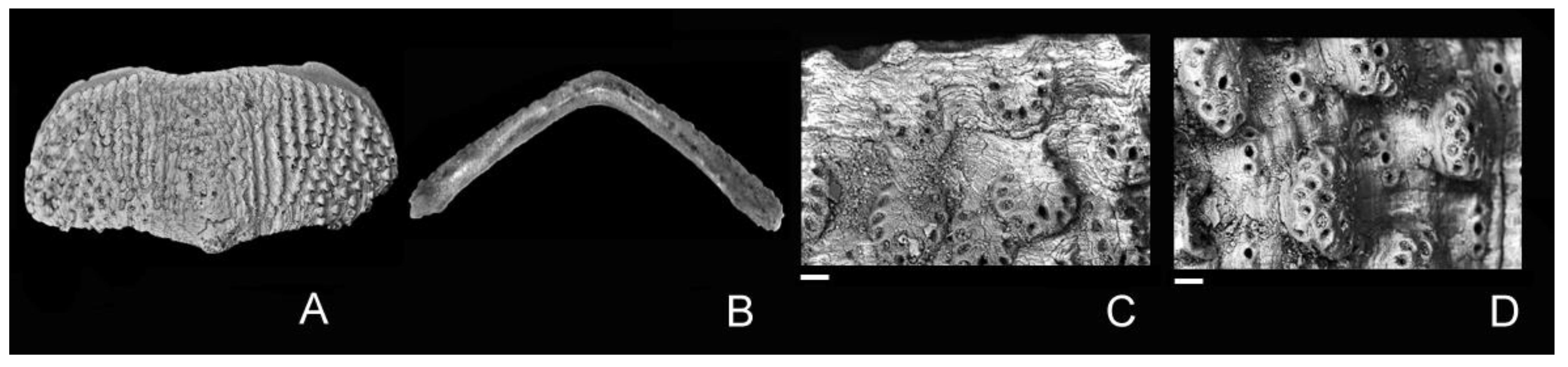
Figure 3.
Leptochiton antondohrni new species; Apulian margin, offshore Bari (SE06-19), submerged deposits, possibly late Pleistocene (last glacial). (A–E) SE06-50, Holotype MZUB 60,418, intermediate valve, width 3.4 mm, dorsal view (A), close-up of surface ornamentation of central (B) and lateral (C) areas, ventral (D) and frontal (E) views. (F–J) SE06-50, Paratype MZUB 60,419, intermediate valve, width 2.9 mm, dorsal view (F), close-up of surface ornamentation of central (G) and lateral (H) areas, ventral (I) and frontal (J) views. (K–O) SE06-50, Paratype MZUB 60,420, tail valve, width 3.1 mm, dorsal view (K), close-up of surface ornamentation of antemucronal (L) and postmucronal (M) areas, ventral (N) and frontal (O) views. (P) SE06-50, MZUB 60,425, tail valve, width 2.6 mm, dorsal view. Scale bars = 40 μm (C); 20 μm (B,G,H,L,M).
Figure 3.
Leptochiton antondohrni new species; Apulian margin, offshore Bari (SE06-19), submerged deposits, possibly late Pleistocene (last glacial). (A–E) SE06-50, Holotype MZUB 60,418, intermediate valve, width 3.4 mm, dorsal view (A), close-up of surface ornamentation of central (B) and lateral (C) areas, ventral (D) and frontal (E) views. (F–J) SE06-50, Paratype MZUB 60,419, intermediate valve, width 2.9 mm, dorsal view (F), close-up of surface ornamentation of central (G) and lateral (H) areas, ventral (I) and frontal (J) views. (K–O) SE06-50, Paratype MZUB 60,420, tail valve, width 3.1 mm, dorsal view (K), close-up of surface ornamentation of antemucronal (L) and postmucronal (M) areas, ventral (N) and frontal (O) views. (P) SE06-50, MZUB 60,425, tail valve, width 2.6 mm, dorsal view. Scale bars = 40 μm (C); 20 μm (B,G,H,L,M).

Remarks. “Leptochiton antondohrni n. sp. is characterized by a tegmentum sculptured with small granules randomly arranged on HV, LA, and PA and in longitudinal series on CA and PA. Three living Mediterranean species are known with this type of sculpture: L. cimicoides (Monterosato, 1879), L. sarsi Kaas, 1981 and L. geronensis Kaas & Van Belle, 1985”.
The more similar species is Leptochiton cimicoides (Monterosato, 1879), from which Leptochiton antondohrni n. sp. differs by a tegmentum sculpture with a greater number of longitudinal striae of granules in CA, AA (50–55 vs. 30 in L. cimicoides), and differently shaped intermediate valves (W/L = 1.45–2.03 vs. 2.73 in L. cimicoides). Leptochiton cimicoides is a recent species, known from the Mediterranean Sea and the western Sahara [23], while some fossil records attributed to this species from the Plio-Pleistocene of Italy [22,49] and Greece [50,51] need more in-depth study to confirm the specific validity of L. cimicoides in the fossil state.
Leptochiton sarsi differs from Leptochiton antondohrni n. sp. by the rougher aspect of its tegmentum and of the granules, with stems in the apical part more evident, and other small differences in the shape of the valves (e.g., the postmucronal slope of the tail valve, almost straight in Leptochiton antondohrni n. sp. versus well concave in L. sarsi).
Leptochiton geronensis differs from Leptochiton antondohrni n. sp. by the shape of the intermediate valves (less elevate and with a subcarinate anterior profile) and with fewer longitudinal series of granules in CA (40 vs. 50–55 in L. antondohrni n. sp.).
The material studied is quite well preserved, and the intermediate valves show some variability. Two types of intermediate valves are detectable, some higher [Figure 3A–E, H/W = 0.66] and slightly larger (W = 3.4 mm, W/L = 1.45–1.60), others a little less high [Figure 3F–J, H/W = 0.54–0.55] and a little smaller (W = 2.9–3 mm, W/L = 1.91–2.03). The structure of the granules also shows slight differences; the granules in CA are more oval and up to 70 µm in size in the higher valves, and more rounded and slightly smaller (up to 60 µm) in the lower valves. We provisionally consider the intermediate valves within the limits of intraspecific variation of this species, although part of the difference could likely be due to age-related variability.
Distribution. Pleistocene, presumably last glacial: Adriatic Sea, offshore Bari (this study).
Genus Belknapchiton Sirenko, Saito & Schwabe, 2022
Type species. Leptochiton belknapi Dall, 1878 by original designation.
Distribution. Belknapchiton is known to have existed from the Pliocene to the present day. There are 22 recent species known to date, most inhabiting the deep waters of the Pacific Ocean, only B. alveolus (M. Sars MS, Lovén, 1846) being known in the Atlantic Ocean. Its fossil record includes the Pliocene to Pleistocene of Italy [52,53].
Belknapchiton alveolus(M. Sars MS, Lovén, 1846)
Chiton alveolus M. Sars MS, Lovén, 1846, p. 159 [54].
Leptochiton alveolus; Ferreira, 1979, p. 152, figures 9–16,37,38 [55]; Kaas, 1981, p. 223, figures 8 and 10A–B [25]; Wu and Okutani, 1984, p. 6, pl. 1, figures 3 and 4, pl. 3, figures 9–14, pl. 4, Figure 1, pl. 5, figures 1–4 [56]; Dell’Angelo and Bonfitto, 2005, p. 5, figures 13–16 [57]; Dell’Angelo et al., 2013, p. 71, plate 2, figures A–D [52]; Dell’Angelo et al., 2021b, p. 410, figures 30–37 [53].
?Leptochiton (Leptochiton) alveolus; Squires and Goedert, 1995, p. 49, figures 3–6 [58]; Dell’Angelo et al., 2011, p. 936 [59]; Schwabe and Sellanes, 2010, table 3 [60]; Bertolaso et al., 2015, p. 9 [61].
Belknapchiton alveolus Sirenko, Saito and Schwabe, 2022, p.103–104, 115 [62].
Type material. Naturhistoriska Riksmuseet, Stockholm, type collection n. 104, one specimen in alcohol, deprived of valves (lectotype, designated by [25]).
Type locality. Bohuslän (Sweden).
Material examined. SE06-35: 1 intermediate valve; SE06-40: 18 valves (1 head, 14 intermediate, 3 tail); SE06-50: 56 valves (10 head, 40 intermediate, 6 tail), Figure 4A–L. Maximum width of the valves: 4.3/5.7/3.5 mm.
Figure 4.
Belknapchiton alveolus (M. Sars MS, Lovén, 1846); Apulian margin, offshore Bari (SE06-50), submerged late Pleistocene (last glacial) deposits. (A–D) MZUB 60421, head valve, width 4.3 mm, dorsal view (A), close-up of surface ornamentation (B), ventral (C) and lateral (D) views. (E–H) MZUB 60422, intermediate valve, width 3.5 mm, dorsal view (E), close-up of surface ornamentation of central area (F), ventral (G) and frontal (H) views. (I–L) MZUB 60423, tail valve, width 3.5 mm, dorsal view (I), close-up of surface ornamentation of antemucronal (J) and postmucronal (K) areas, and lateral (L) view. Scale bars = 20 μm (B,F,J); 10 μm (K).
Figure 4.
Belknapchiton alveolus (M. Sars MS, Lovén, 1846); Apulian margin, offshore Bari (SE06-50), submerged late Pleistocene (last glacial) deposits. (A–D) MZUB 60421, head valve, width 4.3 mm, dorsal view (A), close-up of surface ornamentation (B), ventral (C) and lateral (D) views. (E–H) MZUB 60422, intermediate valve, width 3.5 mm, dorsal view (E), close-up of surface ornamentation of central area (F), ventral (G) and frontal (H) views. (I–L) MZUB 60423, tail valve, width 3.5 mm, dorsal view (I), close-up of surface ornamentation of antemucronal (J) and postmucronal (K) areas, and lateral (L) view. Scale bars = 20 μm (B,F,J); 10 μm (K).

Remarks. The species is characterized by a tegmentum covered with well pronounced, neatly separated, rounded to oval granules, more or less distinctly quincuncially or randomly arranged on all the valves’ surfaces, the lateral areas indiscernible from the central areas, the intermediate valves arched, the tail valve with the anterior margin almost straight and the mucro at posterior 1/3 from the posterior margin. Detailed descriptions of this species are given [25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56]. The taxonomic position of B. alveolus has been recently reviewed [62] and the new genus Belknapchiton established to accommodate this taxon. This deep-water species has a very complicated synonymy, and was initially considered to be cosmopolite, but a recent study [56] demonstrated that B. belknapi Dall, 1878, considered by [55] and other authors a junior synonym of B. alveolus, shows several differential characteristics which warrant its specific separation. Belknapchiton alveolus seems to be restricted to the Atlantic Ocean, while L. belknapi is confined to the Pacific and Indian Oceans [56].
Our material fully agrees with the description of Belknapchiton alveolus, and compares well with a single intermediate valve from Sula Ridge, Norway, dredged at 215 m [57] (figures 13–16). The latter figure 16 is, to our knowledge, the only published figure of the granule structure, and the granules’ shape and dimension agree with our material (maximum diameter of oval granules in the central area of intermediate valves ca. 77 μm vs. 73 in our material, and the same structure of one posterior megaesthete and 4–5 microaesthetes upward).
The finding of Belknapchiton alveolus in the study area is quite significant. This taxon is now extinct in the Mediterranean basin, although some older reports from this area were erroneously recorded, i.e., ([63] p. 32 «Napoli, Palermo, Dalmatia», probably on the basis of records from previous authors) and [64], two specimens collected by fishers at 200–250 m off Gerona, Spain, but the identification was proven wrong, and they have been reclassified as Leptochiton geronensis Kaas & Van Belle, 1985, by the same authors.
Belknapchiton alveolus was recorded from the Ligurian Pliocene [52,53]. Specimens reported as “Leptochiton alveolus” from Eocene–Oligocene cold-seep limestones in the Olympic Peninsula, Washington [58,60] have not been considered in the distribution of B. alveolus, because their specific assignment remains an open problem [60].
Belknapchiton alveolus is similar to L. belknapi Dall, 1878, from which it differs by a few morphological and anatomical characteristics [56,62], as well as by its geographic distribution and bathymetric range (270/540 m for B. alveolus, 100/3724 m for B. belknapi). Other species with the same kind of tegmentum sculpture, fully covered in randomly or quincuncially arranged granules, are known for the fossil Neogene of the Mediterranean area: L. tavianii Dell’Angelo, Landau & Marquet, 2004 from the Pliocene of Estepona (Spain) (where, however, the granules are characterized by a fungiform section and are arranged in a beehive structure), L. salicensis Dell’Angelo & Bonfitto, 2005 from the Pleistocene of Salice (South Italy) (with more solid and dissimilar intermediate valves, and more polygonal and differently shaped granules), and L. lignatilis Dell’Angelo, Bertolaso & Sosso in Bertolaso et al., 2015 (the differences are reported in [61] (Table 1).
Distribution. Lower Pliocene: Italy, Liguria: Zinola [52], Borzoli [53]. Pleistocene, presumably last glacial: Adriatic Sea, offshore Bari (this study). Recent: North Atlantic: all along the Norwegian and Swedish west coast [25,45], Bay of Biscay [46], Spain, Galicia [47], NW Portugal [48], Gulf of Maine and Gulf of St. Lawrence [56].
Family Hanleyidae Bergenhayn, 1955
Genus Hanleya Gray, 1857
Type species. Hanleya debilis Gray, 1857 (=Chiton hanleyi Bean in Thorpe, 1844), by monotypy.
Distribution.Hanleya is known to have existed from the lower Oligocene to the present day. Currently, all known recent species of Hanleya occur only in the Atlantic Ocean and adjacent seas, from off Brazil (25.44° S) to the Barents Sea (74.27° N) [65]. Its fossil record includes the Oligocene of Germany [66], the middle Miocene of Paratethys [67,68], and the upper Miocene to the Pleistocene of Europe [38,39,69,70].
Hanleya hanleyi (Bean in Thorpe, 1844)
Chiton hanleyi Bean in Thorpe, 1844, p. 263, figure 57 [71].
Hanleya hanleyi; Malatesta, 1962, p. 153, figures 9 and 10 [21]; Sabelli, 1972, p. 97, figures 1–6 [72]; Sabelli, 1974, figures 1–13 [73]; Kaas and Van Belle, 1985, p. 193, figure 91, map 18 [27]; Dell’Angelo and Smriglio, 1999, p. 85, plate 25 figures C–D, G, pl. 26 figures I–J [23]; Sirenko et al., 2016, p. 58, figures 1–10 [65]; Dell’Angelo et al., 2018, p. 20, figure 10 [70]; Dell’Angelo et al., 2020b, p. 52, table 9 [74]; Dell’Angelo et al., 2021a, p. 125, figure 1 [75]; Dell’Angelo et al., 2022, p. 6, figure 4 [76].
Type material. Syntype, Scarborough Museums Trust, Woodend, Scarborough, U.K. (Sirenko et al. 2016).
Type locality. Scarborough (Yorkshire, England).
Material examined. SE06-19: 1 intermediate valve, width 3.3 mm, Figure 5A–D.
Figure 5.
Hanleya hanleyi (Bean in Thorpe, 1844); Apulian margin, offshore Bari (SE06-19), probably submerged late Pleistocene (last glacial) deposits. (A–D) intermediate valve, width 3.3 mm, dorsal (A) and frontal (B) views, and close-up of surface ornamentation of pleural (C) and lateral (D) areas. Scale bar = 10 μm (C,D).
Figure 5.
Hanleya hanleyi (Bean in Thorpe, 1844); Apulian margin, offshore Bari (SE06-19), probably submerged late Pleistocene (last glacial) deposits. (A–D) intermediate valve, width 3.3 mm, dorsal (A) and frontal (B) views, and close-up of surface ornamentation of pleural (C) and lateral (D) areas. Scale bar = 10 μm (C,D).

Remarks. The genus Hanleya Gray, 1857 is represented in the Mediterranean Sea by at least two living species: H. hanleyi (Bean in Thorpe, 1844) and H. mediterranea Sirenko, 2014, whose geographical distribution has been recently revised [75]. The only intermediate valve present in our material fully agrees with the description of H. hanleyi, characterized by the sculpture of the pleural areas of the intermediate valves and the antemucronal area of the tail valve consisting of longitudinal series of small, roundish to oval granules (length 75–100 μm) with narrow interstices, fine and close set on the jugum, growing larger and posteriorly converging towards the side margins. Each granule contains one subcentral aesthete surrounded by eight to twelve aesthetes with pores of practically equal width (Sirenko et al., 2016). Detailed descriptions of this species are given by [65].
Distribution. Upper Miocene: northeastern Atlantic (Tortonian): France [70]; Proto–Mediterranean Sea (Tortonian): N. Italy, Po Basin [69]. Lower Pliocene: northeastern Atlantic: U.K. [35,36,37]; Belgium: Kallo [38,39]; central Mediterranean, Italy: Liguria [52]. Lower to upper Pliocene: northeastern Atlantic: Mondego Basin, Portugal [76]. Upper Pliocene to lower Pleistocene: northeastern Atlantic: Anjou, France [70]. Pleistocene: central Mediterranean, Italy: questionable (many reports, in need of revision). Recent: North and Central Atlantic Ocean, near southern Greenland, North America, Europe [48], Canary Islands [77] and northern Africa, Madeira Arch. [78]; Mediterranean Sea: Italy [75], Aegean Sea, Turkey [65,75].
4. Discussion
Polyplacophora collected during cruise SETE-06 are represented by loose valves only, with two taxa, Leptochiton asellus and Belknapchiton alveolus, of North Atlantic (boreo-celtic) geographic origin. They belong probably to Pleistocene last glacial assemblages whose presence in this area was certified in the past [6] and which are known to host many other benthic mollusks of North Atlantic affinity, such as Puncturella noachina and Iothia fulva, at present considered extinct in the Mediterranean basin. As in the case of the many boreo-celtic index-markers [79], these polyplacophorans expanded their geographic range at the peak of the last glacial to include the Mediterranean basin but could not keep pace with the postglacial oceanographic conditions when the present homothermal ‘warm’ situation was established. In principle, these species previously entered the Mediterranean at glacial times; however, we have no confirmed records to date. The record of Belknapchiton alveolus for the Italian Pliocene is consistent with the lower than present temperatures at bathyal depths at that time. Interestingly, faunal lists from the Pleistocene to modern taphocoenoses in the southwestern Adriatic and adjacent Ionian margin do not record the presence of any Polyplacophora [14,80,81,82,83,84]. The only exception is provided by [6], which recorded Lepidochitona cinereus, but this taxonomic assignment cannot be confirmed at present. Our study of the accompanying mollusk fauna (Table 2) upgrades the list for this area, confirming the presence of some taxa already reported before [6,14], but also adding new records. For instance, we have identified from the study site another North Atlantic element, Torellia delicata (Philippi, 1844), previously cited as a glacial subfossil (as T. vestita Jeffreys, 1867) in the western Mediterranean [85].
As discussed by [2,6], the assemblages dredged from the area consist of mixed elements sourced from a variety of environments and of different ages. The deep-sea pre-modern component includes just a part of the entire taphocoenoses, but their paleoclimatic and biogeographic importance is noticeable. With respect to this component, the integration of the lists by [6,81] and our own research ascertains as putative last glacial mollusks in the southwestern Adriatic Sea the following taxa: Iothia fulva (O. F. Müller, 1776), Puncturella noachina (Linnaeus, 1771), Fissurisepta granulosa Jeffreys, 1883, Cantrainea peloritana (Cantraine, 1835), Pilus conicus (Verrill, 1884), Torellia delicata (Philippi, 1844), Cerithiella metula (Lovén, 1846), Pleuromoides packardii A. E. Verrill, 1872, Typhlomangelia nivalis (Lovén, 1846), Belknapchiton alveolus (M. Sars MS, Lovén, 1846), and Cadulus ovulum (Philippi, 1844). This is, however, a conservative figure since it is likely that other species (including H. hanleyi, Leptochiton asellus, and the new Leptochiton antondohrni species) only represented in these southwestern Adriatic Sea taphocoenoses as empty shells could be glacial subfossils.
5. Conclusions
The analysis of mollusk-rich biogenic taphocoenoses dredged from the Apulian margin in the southwestern Adriatic Sea provided four species of polyplacophorans not recorded hitherto from this sector of the Mediterranean basin, one of which (Leptochiton antondohrni) appears to be new to science. Our finding updates the number of North Atlantic-type Pleistocene (sub)fossil taxa recorded from the southwestern Adriatic Sea and the Mediterranean basin as a whole. Furthermore, our study increases the number of species previously recorded from such submerged Pleistocene taphocoenoses.
Author Contributions
Conceptualization, M.T., B.D. and M.S.; Methodology, B.D., M.S. and M.T.; Investigation, M.T.; Resources, M.T.; Writing—Original Draft Preparation, M.T. Writing—Review & Editing, B.D. and M.T.; Funding Acquisition, M.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Acknowledgments
Cruise SETE-06 (Chief Scientists Fabio Trincardi and Domenico Ridente) on-board RV Urania was organized in the frame of the E.U program Hermes; Captain, Crew and Scientific Staff are acknowledged for the naval support and sampling assistance. We thank Franco Corticelli (IMM-CNR Bologna) and Antonio Bonfitto (BIgEA, University of Bologna) for S.E.M. images, Federica Foglini (Ismar-CNR, Bologna) for the bathymetric map, and Giorgio Castellan for editorial help. Enrico Bonatti and Michael Marani kindly reviewed the English text. Thanks are due to three reviewers for their comments that helped to improve the clarity of the article. This is Ismar-CNR, Bologna scientific contribution n. 2073.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fabbri, A.; Gallignani, P. Ricerche geomorfologiche e sedimentologiche nell’Adriatico meridionale. Giorn. Geol. 1972, 38, 53–498. [Google Scholar]
- Colantoni, P.; Gallignani, P. Quaternary evolution of the continental shelf off the coast of Bari (South Adriatic Sea): Shallow seismic, sedimentological and faunal evidences. Géol. Méditer. 1978, 5, 327–338. [Google Scholar] [CrossRef]
- Trincardi, F.; Foglini, F.; Verdicchio, G.; Asioli, A.; Correggiari, A.; Minisini, D.; Piva, A.; Remia, A.; Ridente, D.; Taviani, M. The impact of cascading currents on the Bari Canyon System, SW-Adriatic Margin (Central Mediterranean). Mar. Geol. 2007, 246, 208–230. [Google Scholar] [CrossRef]
- Freiwald, A.; Beuck, L.; Rüggeberg, A.; Taviani, M.; Hebbeln, D.; R/V Meteor Cruise M70-1 participants. The white coral community in the central Mediterranean Sea revealed by ROV surveys. Oceanography 2009, 22, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Angeletti, L.; Taviani, M.; Canese, S.; Foglini, F.; Mastrototaro, F.; Argnani, A.; Trincardi, F.; Bakran-Petricioli, T.; Ceregato, A.; Chimienti, G.; et al. New deep-water cnidarian sites in the southern Adriatic Sea. Mediter. Mar. Sci. 2014, 15, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Angeletti, L.; Prampolini, M.; Foglini, F.; Grande, V.; Taviani, M. Cold-water coral habitat in the Bari Canyon System, Southern Adriatic Sea (Mediterranean Sea). In Seafloor Geomorphology as Benthic Habitat, Chapter 49, 2nd ed.; Harris, P., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 811–824, Geohab Atlas and Seafloor Geomorphic Features and Benthic Habitats. [Google Scholar]
- Taviani, M.; Angeletti, L.; Beuck, L.; Campiani, E.; Canese, S.; Foglini, F.; Freiwald, A.; Montagna, P.; Trincardi, F. Reprint of ‘On and off the beaten track: Megafaunal sessile life and Adriatic cascading processes’. Mar. Geol. 2016, 375, 146–160. [Google Scholar] [CrossRef]
- Foglini, F.; Grande, V.; Marchese, F.; Bracchi, V.A.; Prampolini, M.; Angeletti, L.; Castellan, G.; Chimienti, G.; Hansen, I.M.; Gudmundsen, M.; et al. Application of Hyperspectral Imaging to Underwater Habitat Mapping, Southern Adriatic Sea. Sensors 2019, 19, 2261. [Google Scholar] [CrossRef] [Green Version]
- Taviani, M.; Angeletti, L.; Foglini, F.; Corselli, C.; Nasto, I.; Pons-Branchu, E.; Montagna, P. U/Th dating records of cold-water coral colonization in submarine canyons and adjacent sectors of the southern Adriatic Sea since the Last Glacial Maximum. Progr. Oceanogr. 2019, 175, 300–308. [Google Scholar] [CrossRef]
- Taviani, M. Studio di Una Tanatocenosi Pleistocenica Dragata nel Basso Adriatico. Master’s Thesis (Laurea), Facoltà di Scienze Matematiche, Fisiche e Naturali, University of Bologna, Bologna, Italy; 1976. p. 145, unpublished.
- Colantoni, P.; Noto, P.; Taviani, M. Prime datazioni assolute di una fauna fossile a Pseudamussium septemradiatum dragata nel basso Adriatico. Giorn. Geol. 1975, 40, 133–140. [Google Scholar]
- Taviani, M. Nota sul ritrovamento di cinque specie di Molluschi Gastropoda, Prosobranchia poco conosciuti o nuovi per le acque del Mediterraneo. Quad. Civ. Staz. Idrobiol. Milano 1974, 5, 39–50. [Google Scholar]
- Taviani, M. Associazioni a Molluschi pleistoceniche-attuali dragate nell’Adriatico meridionale. Boll. Zool. 1978, 45, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Panetta, P.; Mastrototaro, F.; Chimienti, G.; Angeletti, L.; D’Onghia, G. Tanatocenosi wurmiana nel Canyon di Bari (Mar Adriatico). Biol. Mar. Mediter. 2013, 20, 148–149. [Google Scholar]
- Trincardi, F.; Ridente, D. SETE-06 Cruise Report R/V Urania, Monopoli, May 06—Ancona, May, 23, 2007. HERMES, Unpublished Report. Available online: https://gismarcloud.myqnapcloud.com/share.cgi?ssid=1b510bc2f0ab4ee6b3612e4aac2d441a (accessed on 15 January 2023).
- Gray, J.E. A list of the genera of Recent Mollusca, their synonyms and types. Proc. Zool. Soc. 1847, 15, 129–206. [Google Scholar]
- Laghi, G.F. Upper Triassic chitons from the Italian Dolomites. Lav. Soc. Venez. Scie. Natur. 2005, 30, 79–84. [Google Scholar]
- Sirenko, B.I. Four new species and one new genus of Jurassic chitons (Mollusca: Polyplacophora: Lepidopleurida) from the Middle Russian Sea. Proc. Zool. Inst. Russ. Acad. Sci. 2013, 317, 30–44. [Google Scholar] [CrossRef]
- Gmelin, J.F. Caroli a Linnei Systema Natura Per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Characteribus, Differentis, Synonymis, Locis etc., 13th ed.; Beer, G.E., Ed.; Lipsia, Germany, 1791; pp. 3021–4120, Editio decima tertia, aucta, reformata, cura J.F. Gmelin, 1(6). Vermes testacea. [Google Scholar]
- Kaas, P.; Knudsen, J. Lorentz Spengler’s descriptions of chitons (Mollusca: Polyplacophora). Zool. Med. 1992, 66, 49–90. [Google Scholar]
- Malatesta, A. Mediterranean Polyplacophora Cenozoic and Recent. Geol. Rom. 1962, 1, 145–171. [Google Scholar]
- Dell’Angelo, B.; Palazzi, S. Considerazioni sulla famiglia Leptochitonidae Dall, 1889 (Mollusca: Polyplacophora). III. Le specie terziarie e quaternarie europee, con note sistematiche e filogenetiche. In Atti Prima Giorn. di Studi Malacol. CISMA; Centro Italiano di Studi Malacologici: Rom, Italy, 1989; pp. 19–140. [Google Scholar]
- Dell’Angelo, B.; Smriglio, C. Chitoni Viventi del Mediterraneo; Edizioni Evolver: Rome, Italy, 1999; p. 256, (English Ed. 2001: Living chitons of the Mediterranean). [Google Scholar]
- Dell’Angelo, B.; Giusti, F. I Polyplacophora di una tafocenosi profonda del Mar Ligure meridionale. Parte II. La Conchiglia 2000, 32, 53–57. [Google Scholar]
- Kaas, P. Scandinavian species of Leptochiton Gray, 1847 (Mollusca: Polyplacophora). Sarsia 1981, 66, 217–229. [Google Scholar] [CrossRef]
- Strack, H.L. Polyplacophora (Keverslakken). In De Fossiele Schelpen van de Nederlandse Kust; Wesselingh, F.P., Moerdijk, P.W., Eds.; Nederlands Centrum voor Biodiversiteit Naturalis: Leiden, The Netherlands, 2010; Chapter 10; pp. 61–66. [Google Scholar]
- Kaas, P.; Van Belle, R.A. Monograph of Living Chitons (Mollusca: Polyplacophora). Vol. 1. Order Neoloricata: Lepidopleurina; Brill, E.J., Backuys, W., Eds.; Leiden, The Netherlands, 1985; p. 240. [Google Scholar]
- Sturrock, M.G.; Baxter, J.M. The ultrastructure of the aesthetes of Leptochiton asellus (Polyplacophora: Lepidopleurina). J. Zool. 1993, 230, 49–61. [Google Scholar] [CrossRef]
- Chemnitz, J.H. Neues Systematisches Conchylien Cabinet; Nürnberg, Germany, 1795; pp. 4–11, 4: (24) + 344 pp. pls CXXII-CLIX (1780); 5: (20) + 324 pp. pls CLXCLXXX–XIII (1781); 6: (12) + 375 pp. pls 1-36 (1782); 7: (12) + 356 pp. pls 37-69 (1784); 8: (16) + 372 pp. pls 70-102 (1785); 9(1): (12) + 151 pp. pls 103-116 (1786); 9(2): Xxvi + 194 pp. pls 117-136 (1786); 10: (20) + 376 pp. pls 137-173 (1788); 11: (20) + 310 pp. pls 174-213. [Google Scholar]
- Spengler, L. Udförlig beskrivelse over det mangeskallede konkylie-slaegt, af Linnaeus kaldet Chiton; met endeel nye Arter og Varieteter. Skrivt. Natur. Selsk. 1797, 4, 62–103. [Google Scholar]
- Leloup, E. Contributions à l’étude de la faune belge. IV. Les Polyplacophores de la côte belge. Bull. Mus. R. Hist. natur. Belg. 1934, 10, 1–23. [Google Scholar]
- Fredj, G. Essai de Stockage et d’Exploitation des Données en Ecologie Marine. Application à l’Etude Biogéographique du Benthos Mediterranéen et Perspectives. Ph.D. Thesis, University of Nice, Nice, France, 1974; pp. 1–176, Annexe A1–A164. [Google Scholar]
- Lucas, M. Les polyplacophores des cotes europeennes. Bull. Mens. Ass. Belg. Malac. Conchyl. Paleont. 1 1968. 1(2): 9–10; 1(3): 11–14; 1(4): 13–14; 1(5): 13–14; 1(6): 13–14; 1(7): 15–16; 2 (1969): 2(1): 17–18; 2(2): 17–18; 2(3): 13–14; 2(4): 15–16. [Google Scholar]
- Mifsud, C.; Ovalis, P. On the presence of Leptochiton asellus (Gmelin, 1791) (Polyplacophora: Leptochitonidae) in the Mediterranean Sea. Boll. Malacol. 2008, 44, 133–134. [Google Scholar]
- Wood, S.V. A Catalogue of Shells from the Crag. Ann. Mag. Nat. Hist. 1842, 9, 455–462, 527–544. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.V. A Monograph of the Crag Mollusca, with Descriptions of Shells from the Upper Tertiaries of the British Isles. Vol. I. Univalves; Paleontographical Society: London, UK, 1848; pp. xii + 208. [Google Scholar]
- Reid, C. Pliocene Deposits of Britain. Mem. Geolog. Surv. UK. 1890, 326. Available online: https://www.biodiversitylibrary.org/bibliography/153206 (accessed on 15 January 2023).
- Marquet, R. A remarkable molluscan fauna from the Kattendijk Formation (Lower Pliocene) at Kallo (Oost-Vlaanderen, Belgium). Bull. Soc. Belge Géol. Paléontol. Hydrol. 1984, 93, 335–345. [Google Scholar]
- Marquet, R. The Neogene Amphineura and Bivalvia (Protobranchia and Pteriomorphia) from Kallo and Doel (Oost-Vlaanderen, Belgium). Palaeontos 2002, 2, 1–100. [Google Scholar]
- Brøgger, W.C. Om de senglaciale og postglaciale nivåforandringer i Kristiania-feltet (Mollukfaunan). Norges Geol. Unders. 1901, 31, 1–731. [Google Scholar]
- Antevs, E. Post-glacial marine shell-beds in Bohuslän. Geolol. Fören. Stockholm Förhand. 1917, 39, 247–425. [Google Scholar] [CrossRef]
- Antevs, E. Shell Beds on the Skagerack. Geolol. Fören. Stockholm Förhand. 1928, 50, 479–748. [Google Scholar] [CrossRef]
- Bellomo, E.; Sabelli, B. A new addition to the Mediterranean Pleistocene “Boreal Guests”: Hanleya nagelfar (Lovén, 1864) (Mollusca, Polyplacophora) from Calabria (S. Italy). Boll. Soc. Paleontol. Ital. 1995, 34, 201–204. [Google Scholar]
- Dons, C. Norges strandfauna. V. Chitonidae. Det Kong. Norske Vidensk. Selsk. 1934, 7, 4–7. [Google Scholar]
- Hansson, H.G. NEAT (North East Atlantic Taxa): Scandinavian marine Mollusca Check-List. Internet Ed. Aug. 1998. Available online: http://www.tmbl.gu.se (accessed on 15 January 2023).
- Kaas, P. On a collection of Polyplacophora (Mollusca, Amphineura) from the Bay of Biscay. Bull. Mus. Nat. Hist. Natur. 1979, 1, 13–31. [Google Scholar]
- Rolan Mosquera, E.; Otero Schmitt, J.; Rolan Alvarez, E. Moluscos de la Ria de Vigo. II. Poliplacoforos, Bivalvos, Escafopodos, Cefalopodos. Thalassas 1990, 2, 276. [Google Scholar]
- Consolado Macedo, M.C.; Macedo, M.I.; Borges, J.P. Conchas Marinhas de Portugal; Editorial Verbo: Lisbon, Portugal, 1999; p. 516. [Google Scholar]
- Dell’Angelo, B.; Forli, M. I Polyplacophora del Pleistocene inferiore di Riparbella (Pisa), con elenco dei molluschi rinvenuti. Boll. Malacol. 1995, 30, 221–252. [Google Scholar]
- Garilli, V.; Dell’Angelo, B.; Vardala-Theodorou, E. Polyplacophora from the Pleistocene of Kyllini (NW Peloponnese, Greece). Boll. Soc. Paleontol. Ital. 2005, 44, 117–134. [Google Scholar]
- Koskeridou, E.; Vardala-Theodorou, E.; Moissette, P. Pliocene and Pleistocene shallow-water chitons (Mollusca) from Rhodes Island, Greece. N. Jahrb. Geol. Paläont—Abh. 2009, 251, 303–330. [Google Scholar] [CrossRef]
- Dell’Angelo, B.; Sosso, M.; Prudenza, M.; Bonfitto, A. Notes on Fossil Chitons. 5. Polyplacophora from the Pliocene of Western Liguria, Northwest Italy. Riv. Ital. Paleont. Strat. 2013, 119, 65–107. [Google Scholar]
- Dell’Angelo, B.; Sosso, M.; Tavano, M. Chitons (Mollusca, Polyplacophora) from the Lower Pliocene of Borzoli (Genova). Ann. Mus. Civ. St. Nat. G. Doria 2021, 114, 403–449. [Google Scholar]
- Lovén, S.L. Index molluscorum litora Scandinaviae Occidentalia habitantium. Öfversigt Af K.Sven. Vetensk.-Akad. Förhandlingar 1846, 3, 134–160. [Google Scholar]
- Ferreira, A.J. The family Lepidopleuridae (Mollusca: Polyplacophora) in the Eastern Pacific. Veliger 1979, 22, 145–165. [Google Scholar]
- Wu, S.-K.; Okutani, T. The deepsea chitons (Mollusca: Polyplacophora) collected by the R/V Soyo-Maru from Japan: I. Lepidopleuridae. Venus 1984, 43, 1–31. [Google Scholar]
- Dell’Angelo, B.; Bonfitto, A. Notes on fossil chitons. 1. A new species of Lepidopleurus (Mollusca: Polyplacophora) from the Pleistocene of Salice (Sicily, Italy). Zootaxa 2005, 821, 1–6. [Google Scholar] [CrossRef]
- Squires, R.L.; Goedert, J.L. An Extant Species of Leptochiton (Mollusca: Polyplacophora) in Eocene and Oligocene cold-seep limestones, Olympic Peninsula, Washington. Veliger 1995, 38, 47–53. [Google Scholar]
- Dell’Angelo, B.; Bonfitto, A.; Taviani, M. Chitons (Polyplacophora) from Paleogene strata in Western Washington State, U.S.A. J. Paleontol. 2011, 85, 936–954. [Google Scholar] [CrossRef]
- Schwabe, E.; Sellanes, J. Revision of Chilean bathyal chitons (Mollusca: Polyplacophora) associated with cold-seeps, including description of a n. sp. of Leptochiton (Leptochitonidae). Org. Divers. Evol. 2010, 10, 31–55. [Google Scholar] [CrossRef]
- Bertolaso, L.; Garilli, V.; Parrinello, D.; Sosso, M.; Dell’Angelo, B. A new Miocene deep-sea chiton and early evidence for Teredinidae-sustained wood-fall communities. Palaeont. Electr. 2015, 18, 1–15. [Google Scholar] [CrossRef]
- Sirenko, B.; Saito, H.; Schwabe, E. A redescription of Leptochiton belknapi Dall, 1878 (Mollusca: Polyplacophora: Leptochitonidae), the type species of the new genus Belknapchiton. Zootaxa 2022, 5205, 101–124. [Google Scholar] [CrossRef]
- Vayssière, A. Mollusques de la France et des Régions Voisines; Amphineures, Gastéropodes, Opisthobranches, Hétéropodes, Marséniades et Oncidiidés; Octave Doin et Fils: Paris, France, 1913; Volume 1, p. 420. [Google Scholar]
- Van Belle, R.A. Sur la presence en Mediterranée de Lepidopleurus alveolus (Lovén, 1846). Inform. Soc. Belge Malacol. 1975, 4, 57–58. [Google Scholar]
- Sirenko, B.I.; Sigwart, J.; Dell’Angelo, B. Hanleya hanleyi (Bean in Thorpe, 1844) (Mollusca, Polyplacophora) and the influence of the Gulf Stream System on its distribution. Ruthenica 2016, 26, 57–70. [Google Scholar]
- Janssen, R. Revision der Polyplacophora des Oligozäns in Mitteleuropa. Arch. Mollusk. 1978, 108, 215–235. [Google Scholar]
- Šulc, J. Studien über die fossilen Chitonen. I. Die fossilen Chitonen im Neogen des Wiener Beckens und der angrenzenden Gebiete. Ann. Des Nat. Mus. Wien 1934, 47, 1–31. [Google Scholar]
- Ruman, A.; Hudácková, N.H. Middle Miocene chitons (Polyplacophora) from the Slovak part of the Vienna Basin and the Danube Basin (Central Paratethys). Acta Geol. Slovaca 2015, 7, 155–173. [Google Scholar]
- Dell’Angelo, B.; Giuntelli, P.; Sosso, M.; Zunino, M. Polyplacophora from the Miocene of North Italy. Part 1: Leptochitonidae, Hanleyidae, Ischnochitonidae and Callistoplacidae. Riv. Ital. Paleont. Strat. 2015, 121, 217–242. [Google Scholar]
- Dell’Angelo, B.; Landau, B.; Van Dingenen, F.; Ceulemans, F. The upper Miocene chitons of northwest France (Mollusca: Polyplacophora). Zootaxa 2018, 4447, 62. [Google Scholar] [CrossRef] [PubMed]
- Bean, W. A supplement of new species. In British Marine Conchology; Being a Descriptive Catalogue, Arranged According to the Lamarckian System, of The Salt Water Shells in Great Britain; Thorpe, C., Ed.; Edward Lumley: London, UK, 1844; pp. 263–267. [Google Scholar]
- Sabelli, B. Sulla presenza e distribuzione di Hanleya hanleyi in Mediterraneo. Conchiglie 1972, 8, 97–100. [Google Scholar]
- Sabelli, B. Hanleya hanleyi (Bean, 1844). Schede Malacol. del Mediterr. 1974, 6. Società Malacologica Italiana, scheda n° 44. Available online: https://www.societaitalianadimalacologia.it/Schede/Hanleya%20hanleyi.pdf (accessed on 15 January 2023).
- Dell’Angelo, B.; Lesport, J.-F.; Cluzaud, A.; Sosso, M. The Oligocene to Miocene chitons (Mollusca: Polyplacophora) of the Aquitaine Basin, southwestern France, and Ligerian Basin, western France. Part 2: Lepidochitonidae, Tonicellidae, Acanthochitonidae, Cryptoplacidae and Additions to Part 1. Boll. Malacol. 2020, 56, 1–58. [Google Scholar]
- Dell’Angelo, B.; Renda, W.; Sirenko, B.I.; Sosso, M.; Giacobbe, S. The Mediterranean distribution of Hanleya hanleyi (Bean in Thorpe, 1844) and H. mediterranea Sirenko, 2014 (Polyplacophora). Boll. Malacol. 2021, 57, 124–135. [Google Scholar]
- Dell’Angelo, B.; Landau, B.M.; da Silva, C.M.; Sosso, M. Biogeography of northeastern Atlantic Neogene chitons (Mollusca, Polyplacophora): New data from the Pliocene of Portugal. J. Paleont. 2022, 96, 814–838. [Google Scholar] [CrossRef]
- Hernández, J.M.; Rolán, E. Clase Polyplacophora. In Moluscos y Conchas Marinas de Canarias; Rolán, E., Ed.; ConchBooks: Vigo, Spain, 2011; pp. 46–53, 407–411. [Google Scholar]
- Segers, W.; Swinnen, F.; de Prins, R. Marine molluscs of Madeira: The Living Marine Molluscs of the Province of Madeira (Madeira and Selvagens Archipelago); Snoeck Publishers: Zwijndrecht, Belgium, 2009; p. 612. [Google Scholar]
- Sabelli, B.; Taviani, M. The making of the Mediterranean molluscan biodiversity. In The Mediterranean Sea; Goffredo, S., Dubinsky, Z., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 285–306. [Google Scholar]
- Rosso, A.; Vertino, A.; Di Geronimo, I.; Sanfilippo, R.; Sciuto, F.; Di Geronimo, R.; Violanti, D.; Corselli, C.; Taviani, M.; Mastrototaro, F.; et al. Hard- and soft-bottom thanatofacies from the Santa Maria di Leuca deep-water coral province, Mediterranean. Deep-Sea Res. II 2010, 57, 360–379. [Google Scholar] [CrossRef]
- Panetta, P.; Mastrototaro, F.; Matarrese, A.; Tursi, A. Tanatocenosi a molluschi presenti nelle acque adriatiche albanesi. Biol. Mar. Mediter. 2003, 10, 597–601. [Google Scholar]
- Panetta, P.; Mastrototaro, F.; D’Onghia, G. Tanatocenosi a molluschi della provincia a coralli di Santa Maria di Leuca (Mar Ionio). Biol. Mar. Mediter. 2012, 19, 186–187. [Google Scholar]
- Negri, M.P.; Corselli, C. Bathyal Mollusca from the cold-water coral biotope of Santa Maria di Leuca (Apulian margin, southern Italy). Zootaxa 2016, 4186, 1–97. [Google Scholar] [CrossRef] [PubMed]
- Nasto, I.; Cardone, F.; Mastrototaro, F.; Panetta, P.; Rosso, A.; Sanfilippo, R.; Taviani, M.; Tursi, A. Benthic invertebrates associated with subfossil cold-water coral frames and hardgrounds in the Albanian deep waters (Adriatic Sea). Turk. J. Zool. 2018, 42, 360–371. [Google Scholar] [CrossRef]
- Bouchet, P.; Taviani, M. Atlantic deep sea gastropods in the Mediterranean: New findings. Boll. Malacol. 1989, 25, 137–148. [Google Scholar]
- WoRMS. World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 25 December 2022).
Figure 1.
Bathymetric map of the southwestern Adriatic Sea showing the location of each station discussed in the article (courtesy of F. Foglini).
Figure 1.
Bathymetric map of the southwestern Adriatic Sea showing the location of each station discussed in the article (courtesy of F. Foglini).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Location and characteristics of the cruise SETE-06 stations discussed in the text.
| Station | Depth (m) | Gear | Latitude N | Longitude E |
|---|---|---|---|---|
| 10 | 335 | rock dredge | 4133.67 | 1728.70 |
| 13 | 423 | rock dredge | 4122.03 | 1706.72 |
| 18 | 341 | box-corer | 4119.07 | 1705.16 |
| 19 | 308 | grab | 4118.97 | 1705.10 |
| 22 | 722 | rock dredge | 4117.73 | 1715.83 |
| 24 | 199 | box-corer | 4121.31 | 1705.98 |
| 25 | 376 | grab | 4117.47 | 1709.29 |
| 35 | 878 | grab | 4113.46 | 1734.35 |
| 40 | 764 | grab | 4143.03 | 1703.95 |
| 48 | 728 | epibenthic dredge | 4143.31 | 1703.39 |
| 50 | 755 | grab | 4144.91 | 1703.31 |
Table 2.
Taxonomic list of mollusks from SETE-06 stations discussed in the present study. The census includes both benthic and holoplanktonic shells, as well as reworked terrestrial shells of pulmonate gastropods. Regarding marine taxa, the systematic arrangement conforms to WoRMS [86].
Table 2.
Taxonomic list of mollusks from SETE-06 stations discussed in the present study. The census includes both benthic and holoplanktonic shells, as well as reworked terrestrial shells of pulmonate gastropods. Regarding marine taxa, the systematic arrangement conforms to WoRMS [86].
| Family | Genus | Species | Author | 10 | 13 | 18 | 19 | 22 | 24 | 25 | 35 | 40 | 48 | 50 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lepetidae | Propilidium | exiguum | (Thompson, 1844) | X | X | X | X | |||||||
| Lepetidae | Iothia | fulva | (Mueller, 1776) | X | X | X | ||||||||
| Fissurellidae | Puncturella | noachina | (Linnaeus, 1771) | X | X | X | X | X | X | |||||
| Fissurellidae | Emarginula | adriatica | O.G.Costa, 1829 | X | ||||||||||
| Fissurellidae | Emarginula | fissura | (Linnaeus, 1758) | X | ||||||||||
| Fissurellidae | Emarginula | rosea | Bell, 1824 | X | ||||||||||
| Fissurellidae | Emarginula | punctulum | Piani, 1980 | X | ||||||||||
| Fissurellidae | Fissurisepta | granulosa | Jeffreys, 1883 | X | ||||||||||
| Scissurellidae | Anatoma | crispata | Fleming, 1828 | X | X | X | X | X | X | X | ||||
| Scissurellidae | Anatoma | umbilicata | (Jeffreys, 1883) | X | ||||||||||
| Lepetellidae | Bogia | labronica | (Bogi, 1984) | |||||||||||
| Trochidae | Clelandella | miliaris | (Brocchi, 1814) | X | ||||||||||
| Chilodontaidae | Putzeysia | wiseri | (Calcara, 1842) | X | X | |||||||||
| Chilodontaidae | Danilia | tinei | (Calcara, 1839) | X | X | X | X | X | ||||||
| Skeneidae | Cirsonella | romettensis | (Granata Grillo, 1877) | X | X | X | X | X | ||||||
| Skeneidae | gen. | sp. | X | |||||||||||
| Bathysciadiidae | Pilus | conicus | (Verrill, 1884) | X | X | |||||||||
| Rissoidae | Alvania | cimicoides | (Forbes, 1844) | X | X | X | X | X | X | X | ||||
| Rissoidae | Alvania | testae | (Aradas e Maggiore, 1844) | X | X | X | X | X | X | X | ||||
| Rissoidae | Alvania | zetlandica | (Montagu, 1815) | X | X | X | X | |||||||
| Rissoidae | Benthonella | tenella | (Jeffreys, 1869) | X | X | X | X | X | ||||||
| Capulidae | Capulis | ungaricus | (Linnaeus, 1758) | X | X | X | ||||||||
| Capulidae | Torellia | delicata | (Philippi, 1844) | X | ||||||||||
| Ovulidae | Simnia | nicaeensis | Risso, 1826 | X | ||||||||||
| Naticidae | Euspira | cf. fusca | (Blainville, 1825) | X | ||||||||||
| Cerithiidae | Cerithidium | submammillatum | (De Rayneval & Ponzi,1854) | X | X | X | ||||||||
| Turritellidae | Turriterinella | tricarinata | (Brocchi, 1814) | X | ||||||||||
| Triphoridae | Marshallora | adversa | (Montagu, 1893) | X | ||||||||||
| Triphoridae | gen. | sp. | X | |||||||||||
| Cerithiopsidae | Cerithiopsis | atalaya | R. B. Watson, 1885 | X | X | |||||||||
| Cerithiopsidae | Cerithiopsis | sp. | X | |||||||||||
| Newtonielliidae | Cerithiella | metula | (Loven, 1846) | X | X | X | ||||||||
| Epitoniidae | Epitonium | algerianum | (Weinkauff, 1866) | X | X | |||||||||
| Epitoniidae | Epitonium | cf. algerianum | (Weinkauff, 1866) | X | ||||||||||
| Eulimidae | Aclis | attenuans | Jeffreys, 1883 | X | X | X | ||||||||
| Eulimidae | Haliella | stenostoma | (Jeffreys, 1858) | X | X | X | ||||||||
| Eulimidae | Melanella | compactilis | (Locard, 1891) | X | X | |||||||||
| Eulimidae | Melanella | petitiana | (Brusina, 1879) | X | X | |||||||||
| Eulimidae | Melanella | sp. | X | |||||||||||
| Atlantidae | Atlanta | peroni | Lesueur, 1817 | X | ||||||||||
| Atlantidae | Atlanta | sp. | X | X | ||||||||||
| Muricidae | Ocinebrina | helleri | (Brusina, 1865) | X | ||||||||||
| Muricidae | Trophon | muricatus | (Montagu, 1803) | X | X | X | X | X | X | X | X | |||
| Muricidae | Pagodula | echinata | (Kiener, 1839) | X | X | X | X | X | X | X | X | X | ||
| Nassariidae | Tyitia | lima | (Dillwyn, 1817) | X | X | X | ||||||||
| Columbellidae | Mitrella | minor | (Scacchi, 1836) | X | ||||||||||
| Columbellidae | Amphissa | acutecostata | (Philippi, 1844) | X | X | X | ||||||||
| Fasciolaridae | Pseudofusus | rostratus | (Olivi, 1792) | X | X | X | ||||||||
| Fasciolaridae | Pseudofusus | sp. | X | |||||||||||
| Raphitomidae | Cyrillia | aequalis | (Jeffreys, 1867) | X | ||||||||||
| Raphitomidae | Cyrillia | linearis | (Montagu, 1803) | X | ||||||||||
| Raphitomidae | Leufroya | sp.1 | X | |||||||||||
| Raphitomidae | Leufroya | sp.2 | X | |||||||||||
| Raphitomidae | Leufroya | concinna | (Scacchi, 1836) | X | ||||||||||
| Raphitomidae | Leufroyia | erronea | Monterosato | X | ||||||||||
| Raphitomidae | Teretia | teres | (Reeve, 1844) | X | X | X | X | X | ||||||
| Borsoniidae | Typhlomangelia | nivalis | (Lovén, 1846) | X | ||||||||||
| Borsoniidae | Drilliola | emendata | (Monterosato, 1872) | X | X | |||||||||
| Borsoniidae | Drilliola | loprestiana | (Calcara, 1841) | X | X | X | ||||||||
| Mangeliidae | Mangelia | costata | (Pennat, 1777) | X | ||||||||||
| Mangeliidae | Mangelia | sp. | X | |||||||||||
| Mangeliidae | Mangelia | costulata | Risso, 1826 | X | ||||||||||
| Architectonicidae | Heliacus | fallaciosus | (Tiberi, 1872) | X | ||||||||||
| Mathildidae | Mathilda | cochlaeformis | Brugnone, 1873 | X | ||||||||||
| Mathildidae | Mathilda | coronata | Monterosato, 1875 | X | ||||||||||
| Mathildidae | Mathilda | retusa | Brugnone, 1873 | X | ||||||||||
| Pyramidellidae | Turbonilla | micans | (Monterosato, 1865) | X | ||||||||||
| Pyramidellidae | Tibersyrnola | unifasciata | (Forbes, 1844) | X | ||||||||||
| Pyramidellidae | Parthenina | flexuosa | (Monterosato, 1874) | X | X | |||||||||
| Pyramidellidae | Tragula | fenestrata | (Jeffreys, 1848) | X | ||||||||||
| Pyramidellidae | Eulimella | scillae | (Scacchi, 1835) | X | X | X | ||||||||
| Pyramidellidae | Eulimella | unifasciata | (Forbes, 1844) | X | ||||||||||
| Pyramidellidae | Megastomia | conoidea | (Brocchi, 1814) | X | ||||||||||
| Pyramidellidae | Odostomia | acuta | Jeffreys, 1848 | X | ||||||||||
| Pyramidellidae | Odostomia | carrozzai | Van Aartsen, 1987 | X | ||||||||||
| Pyramidellidae | Ondina | cf. crystallina | Locard, 1891 | X | X | |||||||||
| Pyramidellidae | Ondina | sp. | X | |||||||||||
| Acteonidae | Crenilabium | exile | (Jeffreys, 1870) | X | ||||||||||
| Retusidae | Retusa | nitidula | (Lovén, 1846) | X | X | |||||||||
| Retusidae | Retusa | umbilicata | (Montagu, 1803) | X | ||||||||||
| Retusidae | Retusa | sp. | X | X | X | |||||||||
| Ringiculidae | Ringicula | gianninii | F. Nordsieck, 1974 | X | ||||||||||
| Ringiculidae | Ringicula | cf. gianninii | F. Nordsieck, 1974 | X | ||||||||||
| Philinidae | Hermania | scabra | (O. F. Muller, 1784) | X | ||||||||||
| Philinidae | Philine | striatula | Monterosato, 1874 | X | X | |||||||||
| Laonidae | Laona | quadrata | S.V. Wood, 1839) | X | X | |||||||||
| Laonidae | Laona | sp. | X | |||||||||||
| Cylichnidae | Scaphander | clavus | Dall, 1889 | X | ||||||||||
| Alacuppidae | Roxania | sp. | X | |||||||||||
| Cavoliniidae | Cavolinia | gibbosa | (d’Orbigny, 1835) | X | ||||||||||
| Cavoliniidae | Cavolinia | inflexa | (Lesueur, 1813) | X | X | X | X | |||||||
| Cavoliniidae | Diacria | trispinosa | (Blainville, 1821) | X | X | |||||||||
| Cliidae | Clio | pyramidata | Linnaeus, 1767 | X | X | X | X | X | X | X | X | |||
| Hyalocylidae | Hyalocylis | striata | (Rang, 1828) | X | X | |||||||||
| Creseidae | Styliola | subula | (Quoy & Gaimard, 1827) | X | X | X | X | |||||||
| Creseidae | Creseis | acicula | (Rang, 1828) | X | X | |||||||||
| Limacinidae | Limacina | bulimoides | (d’Orbigny, 1836) | X | X | X | X | X | ||||||
| Limacinidae | Limacina | retroversa | (J. Fleming, 1823) | X | X | X | ||||||||
| Heliconoididae | Heliconoides | inflatus | (d’Orbigny, 1835) | X | X | |||||||||
| Tylodinidae | Tylodina | perversa | (Gmelin, 1791) | X | ||||||||||
| Nuculidae | Ennucula | aegeensis | (Forbes, 1844) | X | X | X | X | X | X | X | ||||
| Nuculanidae | Saccella | commutata | (Philippi, 1844) | X | X | |||||||||
| Yoldiidae | Yoldiella | lucida | (Loven, 1846) | X | X | X | X | X | X | X | ||||
| Yoldiidae | Yoldiella | messanensis | (Jeffreys, 1870) | X | X | X | X | X | ||||||
| Yoldiidae | Yoldiella | philippiana | (Nyst, 1845) | X | X | X | X | X | ||||||
| Yoldiidae | Yoldiella | striolata | (Brugnone, 1876) | X | X | X | ||||||||
| Yoldiidae | Yoldiella | wareni | La Perna, 2004 | X | X | |||||||||
| Arcidae | Asperarca | nodulosa | (O.F. Muller, 1776) | X | X | X | X | X | X | X | X | |||
| Arcidae | Bathyarca | pectunculoides | (Scacchi, 1835) | X | X | X | X | X | X | X | X | X | ||
| Arcidae | Bathyarca | philippiana | (Nyst, 1848) | X | X | X | X | X | X | |||||
| Propeamussiidae | Cyclopecten | hoskynsi | (Forbes, 1844) | X | ||||||||||
| Pectinidae | Pseudamussium | clavatum | (Poli, 1795) | X | X | |||||||||
| Pectinidae | Pseudamussium | peslutrae | (Linnaeus, 1771) | X | X | X | X | |||||||
| Pectinidae | Deletopecten | vitreus | (Gmelin, 1791) | X | X | X | X | X | ||||||
| Propeamussiidae | Similipecten | similis | (Laskey, 1811) | X | X | X | ||||||||
| Propeamussiidae | Mesopeplum | fenestratum | (Forbes, 1844) | X | X | X | X | X | X | X | X | |||
| Mytiliidae | Modiolula | phaseolina | (Philippi, 1844) | X | X | X | X | X | X | X | X | |||
| Anomiidae | Heteranomia | squamula | (Linnaeus, 1758) | X | X | X | X | X | X | X | ||||
| Anomiidae | Pododesmus | patelliformis | (Linnaeus, 1761) | X | X | X | ||||||||
| Limidae | Limaea | crassa | (Forbes, 1844) | X | X | X | X | X | X | X | ||||
| Limidae | Limatula | gwyni | (Sikes, 1903) | X | X | X | X | X | X | X | X | |||
| Thyasiridae | Leptaxinus | incrassatus | (Jeffreys, 1876) | X | X | |||||||||
| Ungulinidae | Microstagon | trigonum | (Scacchi, 1835) | X | ||||||||||
| Carditidae | Centrocardita | aculeata | (Poli, 1795) | X | X | X | X | |||||||
| Astartidae | Goodallia | triangularis | (Montagu, 1803) | X | X | X | ||||||||
| Astartidae | Astarte | sulcata | (da Costa, 1778) | X | X | X | X | X | X | |||||
| Carditidae | Centrocardita | aculeata | (Poli, 1795) | X | X | X | ||||||||
| Cardiidae | Papillicardium | minimum | (Philippi, 1836) | X | X | X | X | X | X | X | ||||
| Semelidae | Abra | longicallus | (Scacchi, 1835) | X | X | X | X | X | ||||||
| Kellielliidae | Kelliella | miliaris | (Philippi, 1844) | X | X | X | X | X | X | |||||
| Trapezidae | Coralliophaga | lithophagella | (Lamarck, 1819) | X | X | |||||||||
| Veneridae | Timoclea | ovata | (Pennant, 1777) | X | X | X | X | X | X | |||||
| Corbulidae | Corbula | sp. | X | |||||||||||
| Hiatellidae | Hiatella | arctica | (Linnaeus, 1767) | X | X | X | X | X | X | X | ||||
| Xylophagaidae | Xylophaga | dorsalis | (W. Turton, 1819) | X | ||||||||||
| Cuspidariidae | Cuspidaria | jugosa | (S. V. Wood, 1857 | X | ||||||||||
| Dentaliidae | Antalis | agilis | (M. Sars, 1872) | X | X | X | X | X | X | |||||
| Dentaliidae | Antalis | dentalis | (Linnaeus, 1758) | X | ||||||||||
| Fustariidae | Fustiaria | rubescens | (Deshayes, 1826) | X | ||||||||||
| Entalinidae | Entalina | tetragona | (Brocchi, 1814) | X | X | X | X | X | X | |||||
| Pulselliidae | Pulsellum | lofotense | (M. Sars, 1865) | X | ||||||||||
| Gadilidae | Cadulus | jeffreysi | Monterosato, 1875 | X | X | X | X | X | X | |||||
| Gadilidae | Cadulus | ovulum | (Philippi, 1844) | X | ||||||||||
| Gadilidae | Cadulus | cf. subfusiformia | (M. Sars, 1865) | X | X | |||||||||
| Valloniidae | Vallonia | pulchella | (O. F. Muller, 1774) | X | ||||||||||
| Lauriidae | Lauria | sp. | X | |||||||||||
| Leptochitonidae | Leptochiton | asellus | (Gmelin, 1791) | X | X | X | X | X | X | X | X | X | X | X |
| Leptochitonidae | Leptochiton | antondohrni | new species | X | X | X | X | X | ||||||
| Leptochitonidae | Belknapchiton | alveolus | (M. Sars MS, Lovén, 1846) | X | X | X | ||||||||
| Hanleyidae | Hanleya | hanleyi | (Bean in Thorpe, 1844) | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Taviani, M.; Sosso, M.; Dell’Angelo, B. Chitons from Deep-Water Mollusk-Rich Deposits in the Southwestern Adriatic Sea (Mollusca, Polyplacophora). Diversity 2023, 15, 359. https://doi.org/10.3390/d15030359
AMA Style
Taviani M, Sosso M, Dell’Angelo B. Chitons from Deep-Water Mollusk-Rich Deposits in the Southwestern Adriatic Sea (Mollusca, Polyplacophora). Diversity. 2023; 15(3):359. https://doi.org/10.3390/d15030359
Chicago/Turabian StyleTaviani, Marco, Maurizio Sosso, and Bruno Dell’Angelo. 2023. "Chitons from Deep-Water Mollusk-Rich Deposits in the Southwestern Adriatic Sea (Mollusca, Polyplacophora)" Diversity 15, no. 3: 359. https://doi.org/10.3390/d15030359
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.