
Unciolidae of Deep-Sea Iceland (Amphipoda, Crustacea) †


1
Natural History Museum, Cromwell Road, South Kensington, London SW7 5BD, UK
2
Institut für Marine Ökosystem- und Fischereiwissenschaften, University of Hamburg, 22767 Hamburg, Germany
*
Author to whom correspondence should be addressed.
†
urn:lsid:zoobank.org:pub:D0FDBD76-6B82-4B8B-A226-3C5A7C5FF05A.
Diversity 2023, 15(4), 546; https://doi.org/10.3390/d15040546
Submission received: 23 February 2023
/
Revised: 30 March 2023
/
Accepted: 31 March 2023
/
Published: 11 April 2023
(This article belongs to the Special Issue Deep Atlantic Biodiversity)
Abstract
:An overview of North Atlantic Unciola Say, 1818 is provided along with the description of two new species, U. conchicola sp. nov. and U. icelandica sp. nov. Both deep-sea species were collected living sympatrically at an almost 2000 m depth in the Iceland Basin, southwest of Iceland, each showing a vertical distribution of more than 1 km. In addition to the wide depth range, U. icelandica sp. nov. also shows a broad geographic distribution of more than 1000 km. Unciola conchicola sp. nov. was seen to attach its tubular domicile to the inner cavity of a gastropod shell; however, no specific lifestyle information is known for the species U. icelandica sp. nov. Both species present novel characters for the genus: in U. conchicola sp. nov., the epimeral plates 1–3 have acute projections angled ventrally, while in U. icelandica sp. nov., the male antenna 2 pedunclar article 2 is developed into a distinct phalange. The large amount of unciolid material allowed for ontogentic studies. The material of U. planipes Norman, 1867 was investigated. Changes from juvenile to adult specimens of Neohela monstrosa (Boeck, 1861) are discussed. A key is provided to the eight North East Atlantic species of Unciola.
1. Introduction
During the IceAGE expeditions I and II (Icelandic Marine Animals: Genetics and Evolution), a total of 155 specimens of the family Unciolidae were recovered from ten stations with a very wide depth range, from 118 to 2749 m [1] (Brix et al., 2018).
There are two genera in the subfamily Unciolinae present in Nordic Seas, Neohela S.I. Smith, 1881 and Unciola Say, 1818. Currently, 14 species of Unciola are known worldwide [2] (Horton et al., 2023), with all species known only from the Northern Hemisphere, including eight species from the North Atlantic/subarctic sea region: (1) U. crenatipalma (Spence Bate, 1862), described from the UK, from shallow waters less than 30 m, with a southerly distribution being infrequently reported in Norwegian waters; (2) U. laticornis Hansen, 1887 and (3) U. petalocera (G.O. Sars, 1879), which have a northern distribution, with a few records within the Barents Sea [3] (Vader et al., 1997);(4) U. planipes Norman, 1867, commonly recorded in regions of Norway from Skagerrak to the north of Lofoten [3] (Vader et al., 1997) and considered a deep-water species found below 400 m on the outer parts of the Norwegian shelf [4] (Buhl-Jensen 1986); and (5) U. crassipes Hansen, 1887 and (6) U. leucopes (Krøyer, 1845), the only two Unciola species that are reported from both the eastern and western Atlantic. From the western side of the North Atlantic, (7) Unciola dissimilis Shoemaker, 1945 and (8) Unciola serrata Shoemaker, 1945 are reported.
The Unciolid genus Neohela S.I. Smith, 1881 consists of six species [2] (Horton et al., 2023) and is also known only from North Hemisphere records. One of the largest and most conspicuous species is N. monstrosa (Boeck, 1861), which reaches up to 31 mm as adults [5] (Stephensen, 1944). It is common in the cold and deep waters of the Norwegian Sea from 300 to 2000 m, where it forms dense populations in soft deep-sea sediment, where it is known to create burrows up to 10 cm deep [6] (Buhl-Mortensen et al., 2016). Here we describe a juvenile N. monstrosa from the IceAGE collections, providing additional detail on ontogenic change for material of less than 20 mm, contrasting 31 mm adults, which are well described in the literature [5] (Stephensen, 1944).
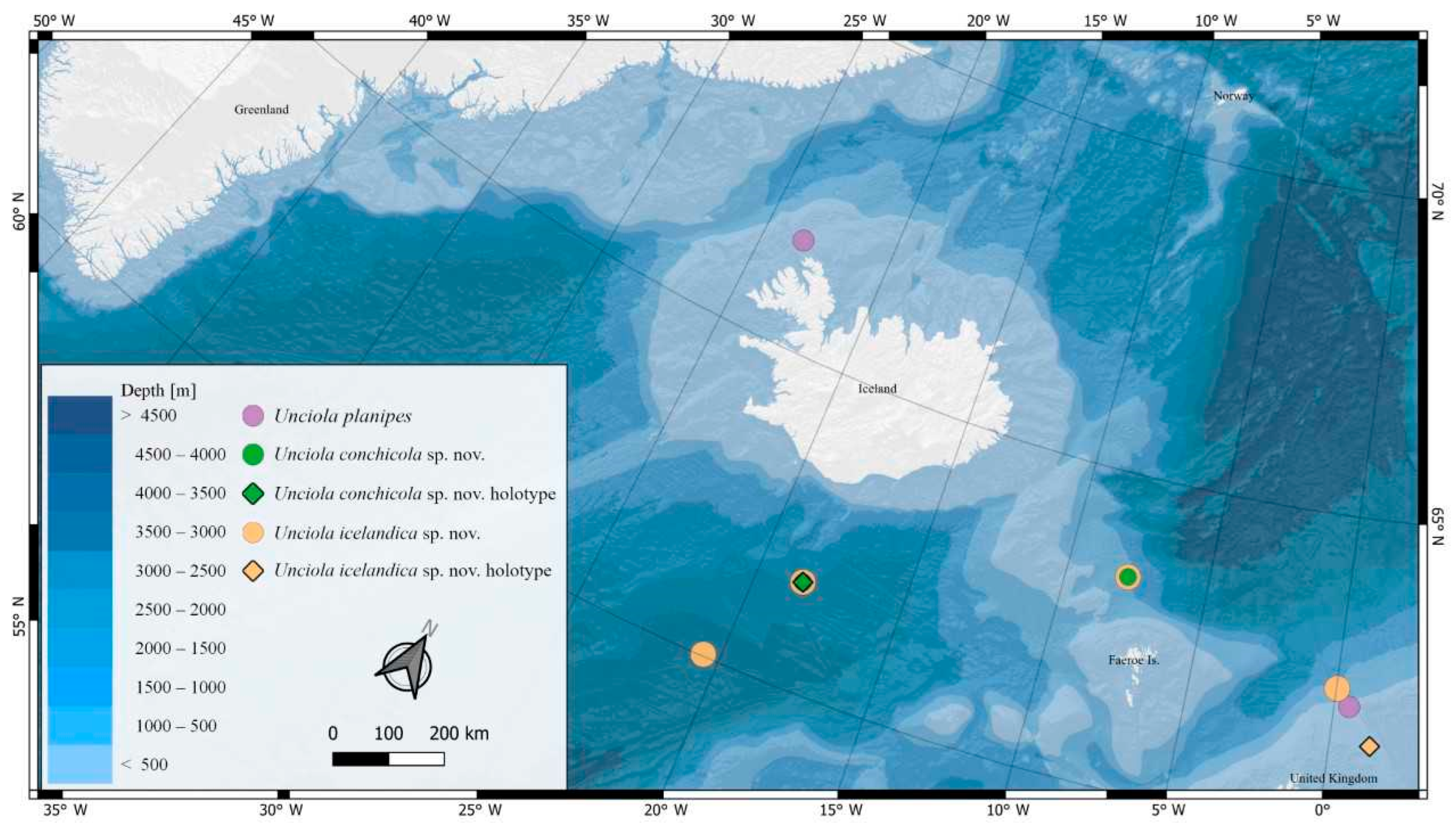
From the IceAGE material of Unciola, we described two species new to science from Icelandic waters and reported the known species U. planipes Norman, 1867 (Figure 1).
The new species, U. conchicola and U. icelandica, occur sympatrically at close to 2000 m depths. An updated key to the North Atlantic Unciola species is provided.
2. Materials and Methods
Samples were taken during the IceAGE expeditions (Icelandic Marine Animals: Genetics and Evolution [7]) via RV Meteor in 2011 and Poseidon 2013 (Table 1). The sampling gear was an epibenthic sledge [8] (Brenke, 2005). All sorting was handled according to Riehl et al. (2014) [9] using an undisturbed cooling chain protocol.
The body length of specimens was measured by tracing the dorsal length from the tip of the rostrum to the end of the telson. To ensure accuracy, telsonicsetal counts were made by mounting whole animals on slides and observing them on a stereomicrosope before returning them to ethanol. Specimens were dissected in an Euparal essence and 96% ethanol solution before being mounted in Euparal as slide preparations. The pencil drawings were conducted with a LeicaM125 and an Olympus BX53 at the laboratories of the University of Hamburg.
In preparation for scanning electron microscope (SEM) imaging, whole animal specimens and appendages were dehydrated through a graduated ethanol series from 80 to 99 percent, acetone dried, mounted on stubs and coated with gold-palladium. Stub mounted material was imaged using a SEM LEO1525.
Abbreviations are used in the following: Antenna 1 (A19, Antenna 2 (A2), Maxilliped (Mxp), Mandible (Md) Maxilla 1(Mx1), Maxilla 2 (Mx2), Gnathopod 1(G1), Gnathopod 2 (G2), Pereopod (P), Telson (T), Uropod (U), Urosome (Ur).
The material of this study is deposited at the Senckenberg Museum Frankfurt, the DZMB Hamburg and the Zoological Museum Hamburg, Germany.
3. Results
- Systematics
- Amphipoda Latreille, 1816
- Aoroidea Stebbing, 1899
- Unciolidae Myers and Lowry, 2003
- Unciolinae Myers and Lowry, 2003
- Neohela S.I. Smith, 1881
- Neohela monstrosa (Boeck, 1861)
Neohela monstrosa. —[13] S.I. Smith, 1881: 450. —[14] Hansen, 1888: 168. —[15] Della Valle, 1893: 343, pl. 55 Figure 19–24. —[16] G.O. Sars, 1894: 624, pl. 224. —[17] Stebbing, 1906: 675. —[18] Shoemaker, 1930: 129. —[19] Stephensen, 1933: 51, Figure 26, not Figure 23 (lapsus for N. maxima). —[20] Stephensen, 1942: 404 (? in part). —[21] Enequist, 1949: 381, Figure 67. —[22] Coyle and Mueller, 1981: 11. —[23] Gurjanova, 1951: 959, Figure 667. —[24] Gurjanova, 1953: 240. —[25] Barnard and Karaman, 1991: 215, Figure 39D, 42J, 44F, 45T.
Neohela phasma [13] S.I. Smith, 1881: 448; [17] Stebbing, 1906: 676; [23] Gurjanova, 1951: 959 (key). Helella monstrosa [26] G.O. Sars, 1883: 31. —[27] d’Udekemd’Acoz, 2007: 32–35, Figure 11.
Not Neohela monstrosa. —? [28] G.O. Sars, 1885: 68 (in part: deep records). —? [29] Chevreux, 1899: 147. —? [30] Chevreux 1900: 130. —? [20] Stephensen, 1942: 404, (in part: deep records). —? [31] Piepenburg et al., 1996: 439 (in part: deep records). —? [32] Węsławski et al., 2003: 81. Neohela sp. A. —? [32] Węsławski et al., 2003: 81. (accepted as Neohela lamina d’Udekem d’Acoz, 2007 following [27] d’Udekemd’Acoz, 2007).
- Material examined.
SMF 52259, one male, 19 mm; SMF 55298 (Figure 2 and Figure 3), one intersex and one juvenile, Norwegian Basin, Norwegian Sea, 1886 m, see Table 1.
Remark.
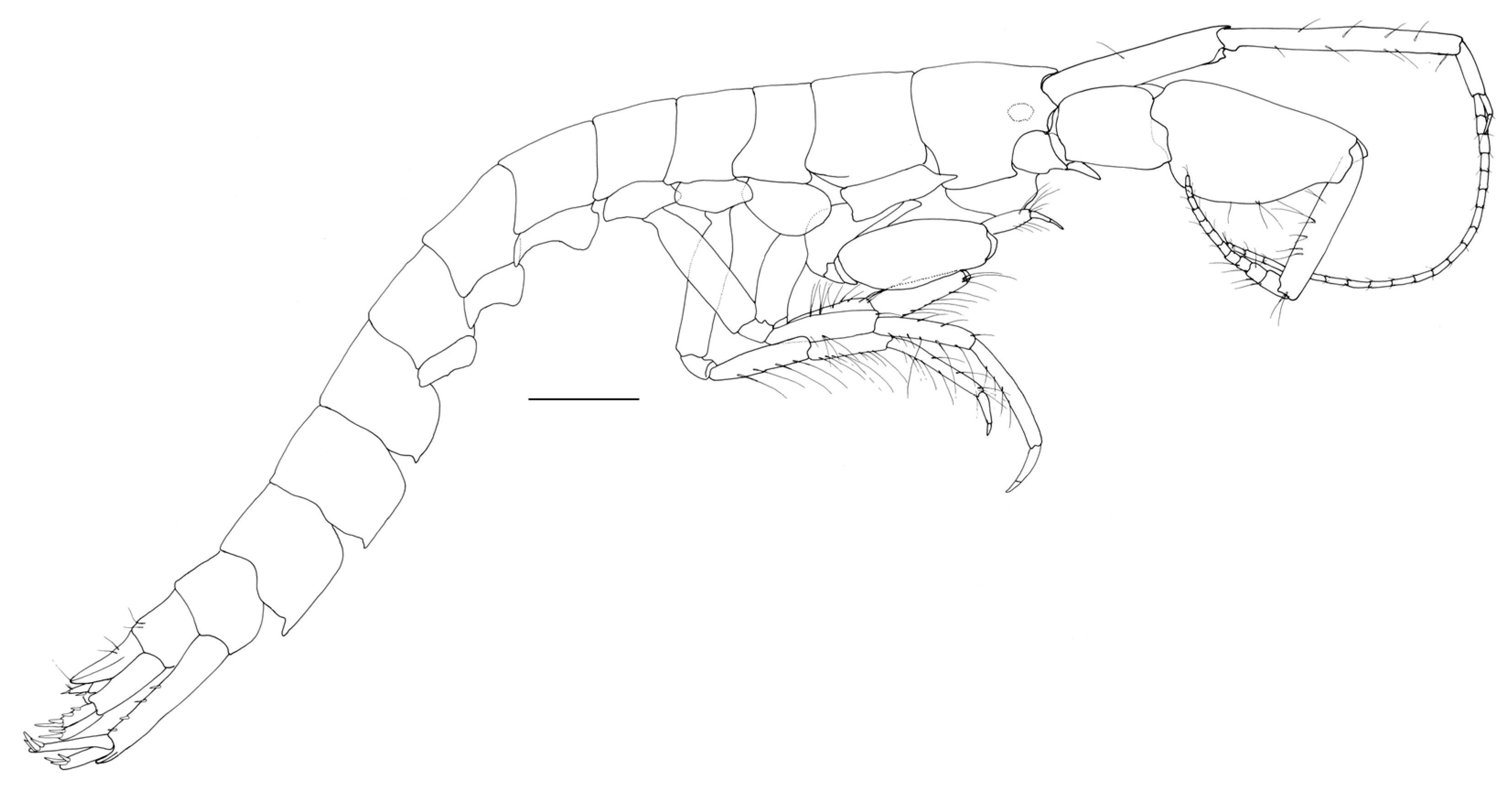
A N. monstrosa juvenile male of 19 mm is presented here via morphological investigation. Previous records document the adult specimens, which achieve a substantial adult size of 31 mm from other North Atlantic samples [5] (Stephensen, 1944). Variation include juveniles with three teeth on the gnathopod 1 palm, being a proximal, medial and distal (in the corner), adults have three teeth projecting from the palm. Other notable species-level characters which remain consistent are length to width ratio of the gnathopod 1 and 2 carpus and propodus.
- Unciola Say, 1818
- Glauconome [37] Kroyer, 1845: 491. (junior homonym)
- Dryope [38] Spence Bate, 1862: 276. (junior homonym)
- Unciola conchicola sp. nov.
- urn:lsid:zoobank.org:act:428FA701-A6FE-4728-A239-BAA5FB06772E
- Type Material
Holotype male, 10 mm, whole animal illustrated, ZMHK 62083, South Island, Iceland Basin, (61°42.630′ N, 19°32.960′ W–61°42.170′ N, 19°32.020′ W), epibenthic sledge, 1912–1921 m, 31 August 2011, expedition RV Meteor (St 85-3-989).
Paratypes, all males, same location as holotype: ZMHK 62088, ZMHK 62089, ZMHK 62090 and ZMHK-56724 on SEM stud.
Further material examined: SMF 52175, one male, one juvenile, Faroe Islands Ridge middle, 63°23.360′ N, 008°09.420′ W–63°24.620′ N, 008°11.220′ W, epibenthic sledge, 686–687 m, 31 July 2013, expedition RV Poseidon (St880-2).
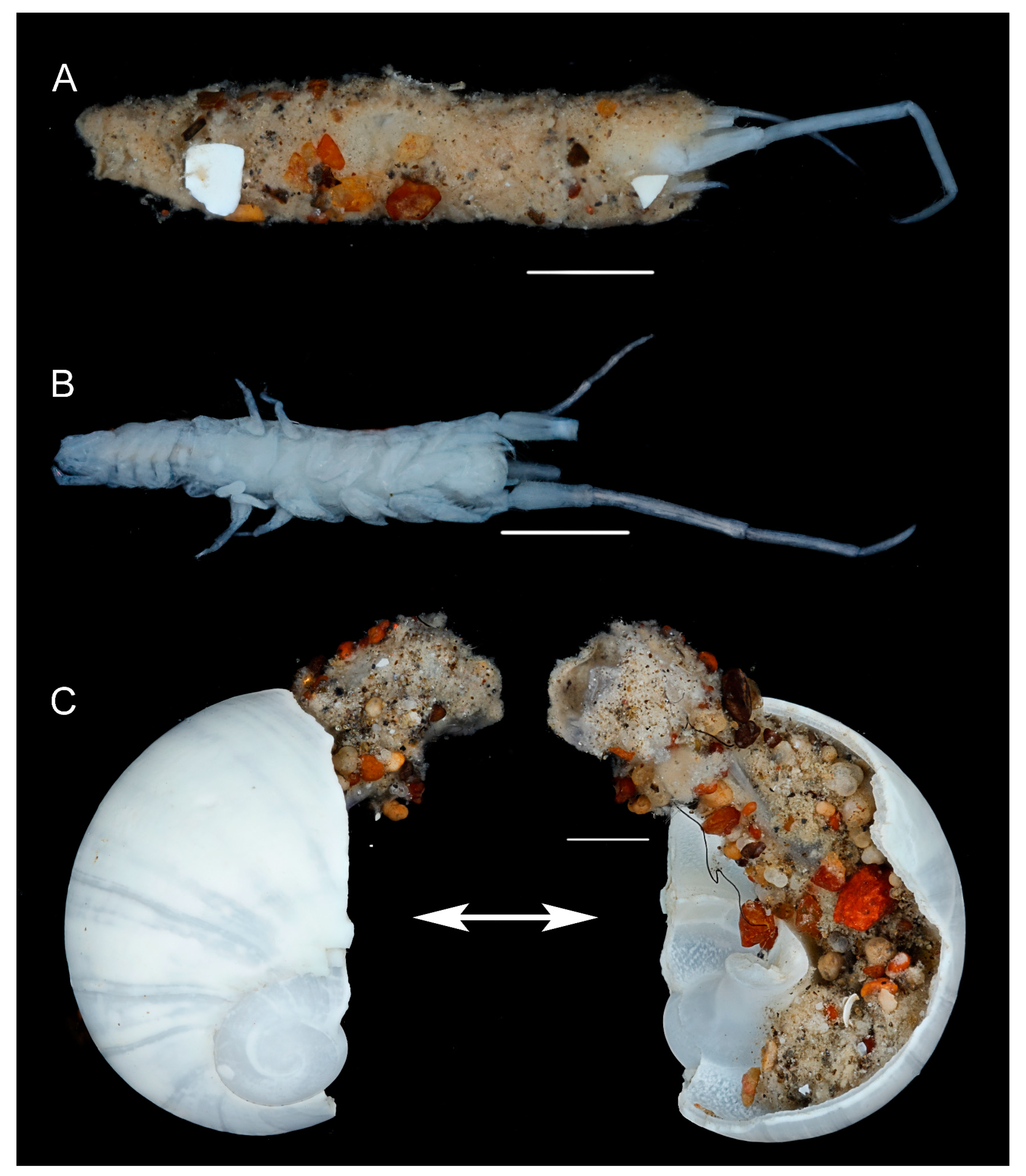
- Etymology. Named from its domicile lifestyle with molluc shells.
- Type locality. South Island, Iceland Basin.
- Description. Based on type material.
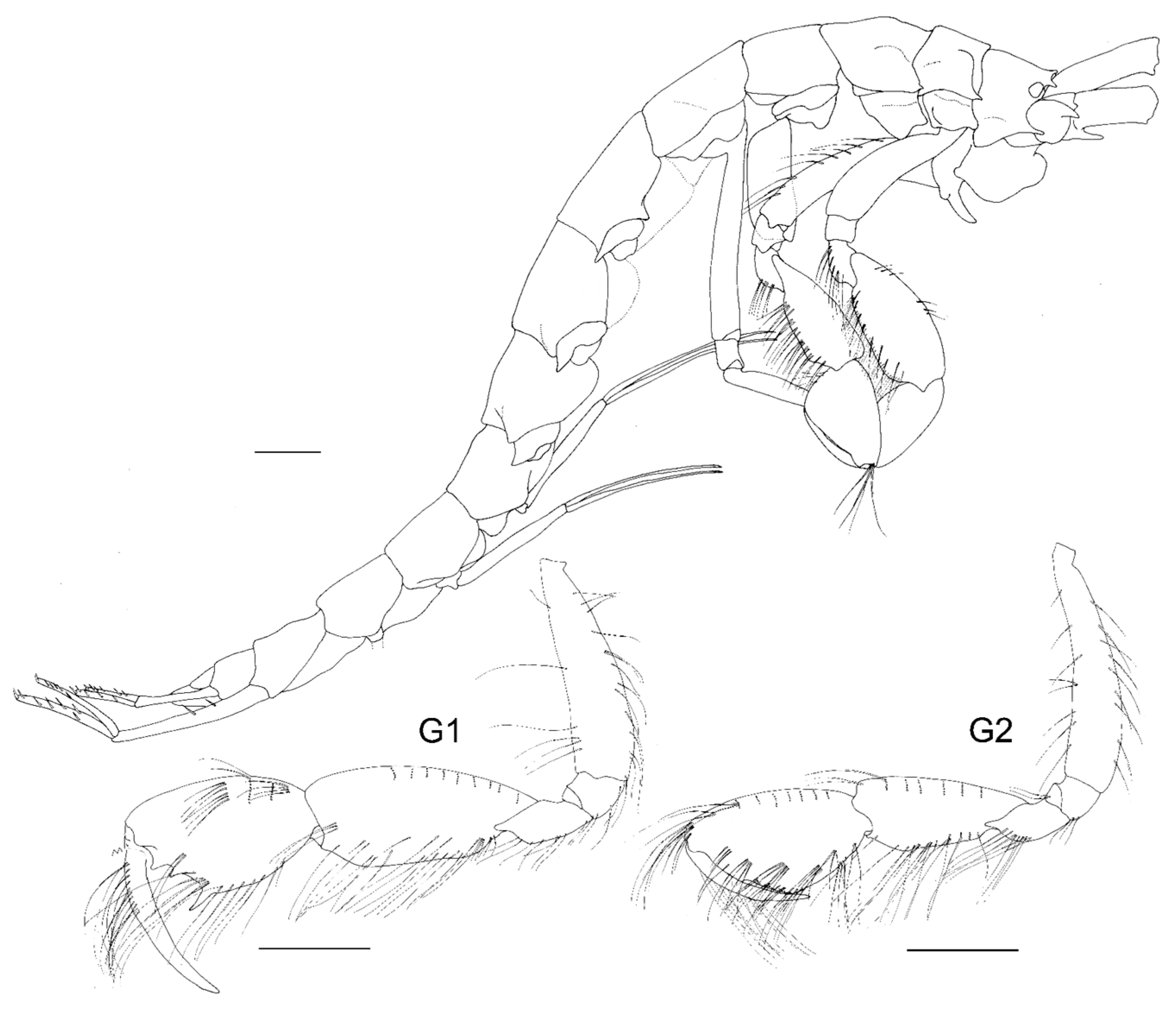
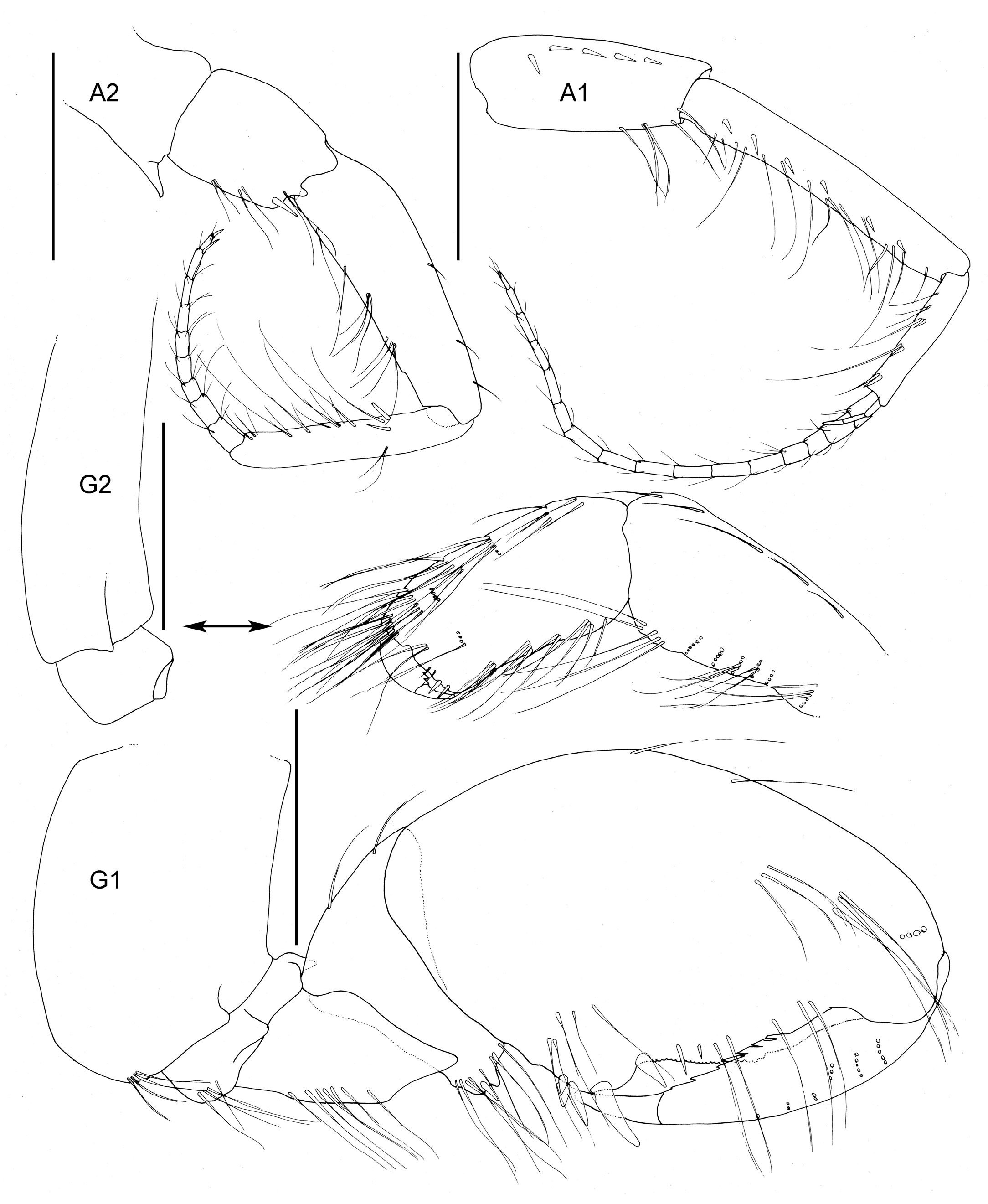
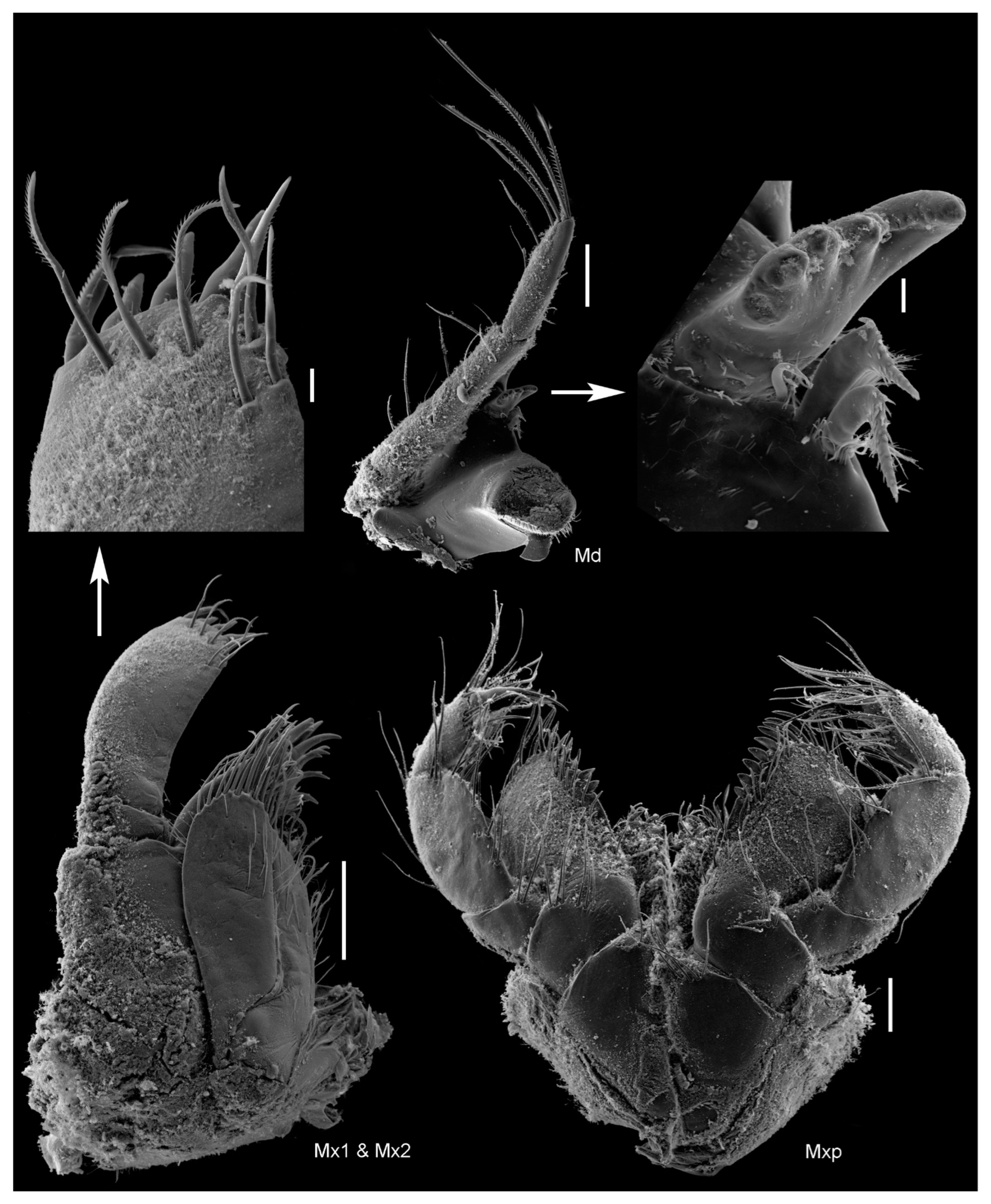
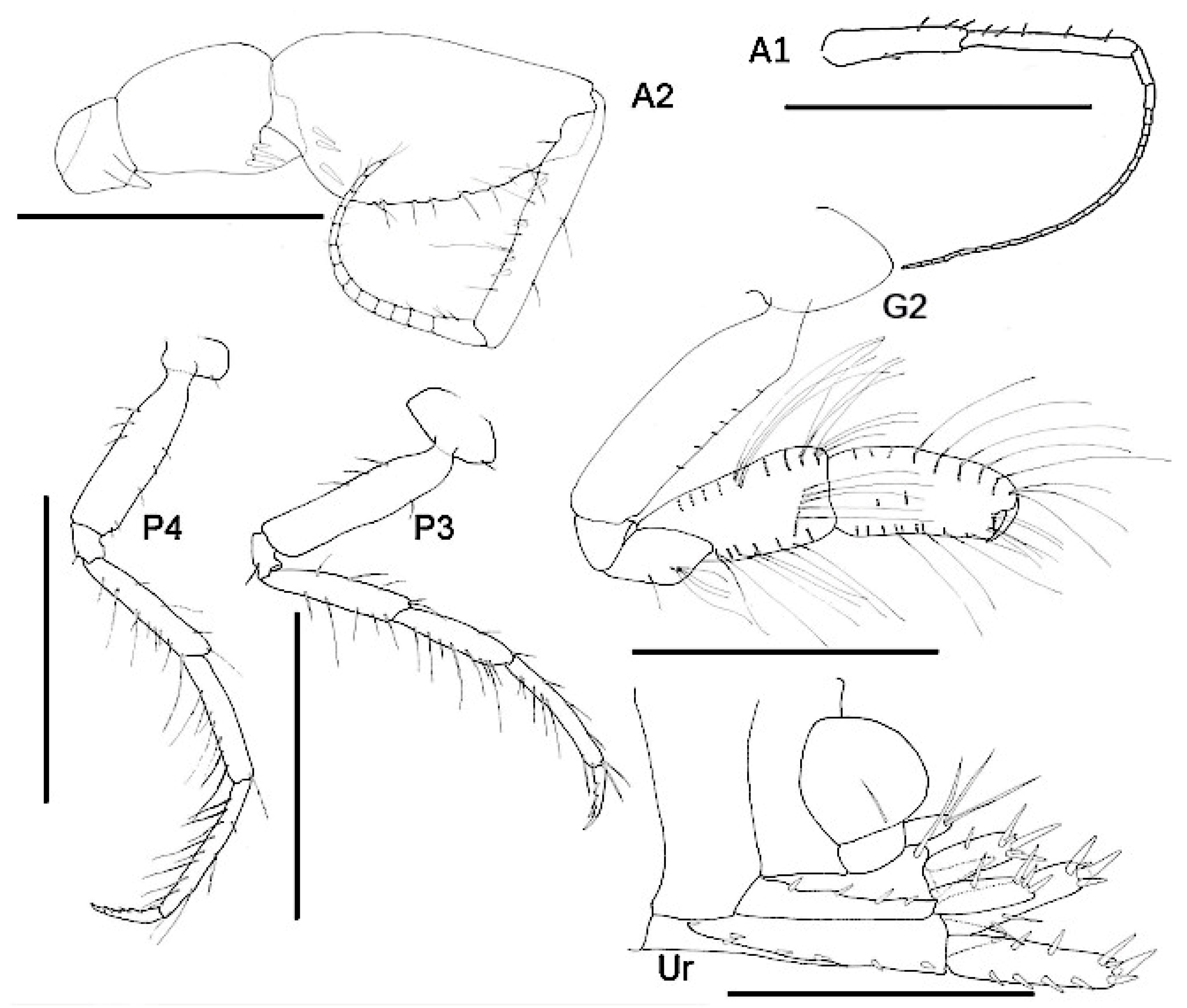
- Head. Eyes absent. Rostrum, well-developed, acute. Lateral cephalic lobes produced into flange engulfing peduncle of antenna 1, anteroventral margin subquadrate. Antenna 1 peduncle article 1 length twice the width, dorsal margin lined with robust setae; article 2 twice length of article 1, ventral margin lined with short robust and long slender setae; article 3 length five times the width, shorter than article 1, accessory flagellum 2-articulate; primary flagellum 16 articulate. Antenna 2 peduncle article 1–2 anteroventral corner acute produced; article 3 length 1.1 times as long as it was broad, anteroventral margin with robust seta, ventral margin lined with slender setae; article 5 shorter than article 4; flagellum with nine articles. Mandible molar absent; accessory setal row absent; palp article 3 shorter than 2, length 5.5 times the width, lined with long slender setae. Maxilla 1 inner plate unknown; outer plate with ten robust setae; palp 2-articlulate, article 2 with five robust and five slender setae. Maxilliped inner plate with three robust setae; outer plate apex with line with setae progressing from longer to more stout robust setae; palp article 4 proximally narrow and distally broad, dactylus with unguis present.
- Pereon. Pereonites cuticle rugosely textured. Pleonite 3 sternal process well-developed, directed anteriorly, apically acute. Pleonite 4 sternal process moderately developed, directed anteriorly, apically subacute. Pereonite 5 sternal process minutely developed, anteriorly developed, subacute. Coxa subrectangular, depth less than half of pereonites. Gnathopod 1 larger than gnathopod 2, coxa anteroventral corner acutely produced; basis stout, anterodistal margin subacute; ischium much broader than it was long; merus twice as long as it was broad, posterior margin lined with slender setae; carpus compressed between merus and propodus, posterior margin with single robust and sparse slender setae; propodus length twice the width, anterior and posterior margins with a few slender setae, posterior margin straight, palm subacute to acute, length two thirds of posterior margin, with weak distal sinus, defined by rounded corner with three robust setae; dactylus closing along palm, unguis present, anterior margin lined with rows of setae, inner margin serrate. Gnathopod 2 subchelate; basis anterior and posterior margin weakly convex without setae, anterodistal corner subacute; ischium length 1.2 times the width; merus unknown; carpus length 2.4 times the width, longer than propodus length; propodus anterior and posterior lined with rows of long slender setae, length 2.4 times the width, palm transverse, defined by subquadrate corner. Pereopods 5 to 7 basis rectilinear; posterior margins with plumose setae; dactylus unguis present.
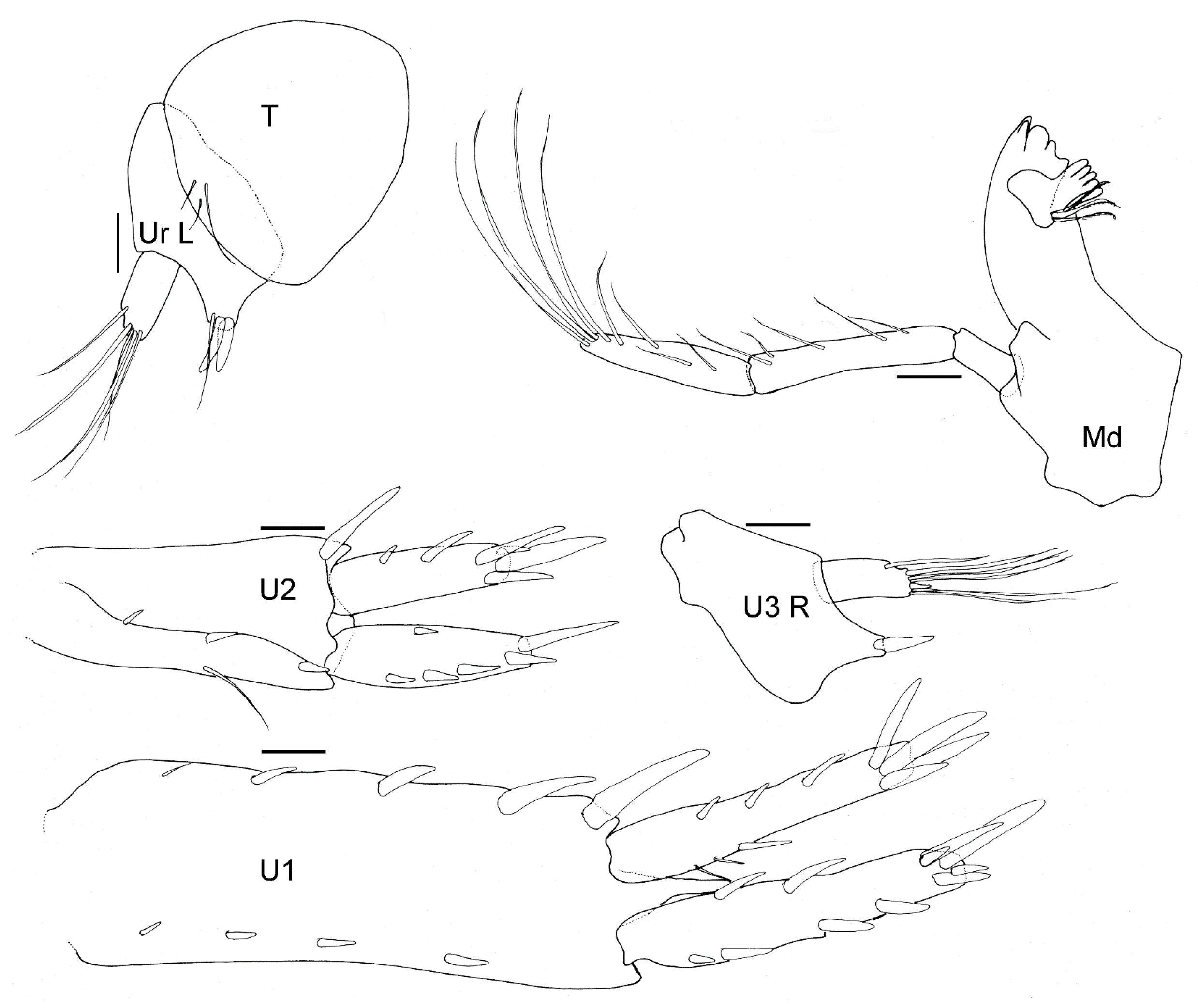
- Pleon. Epimeron 1 posteroventral margin acutely produced, lined with plumose setose. Epimera 2–3 ventral margin with acute tooth in middle of margin, posterior margin straight, corner evenly rounded. Urosomites 1–3 not coalesced. Urosomite 1 ventral margin lined with plumose setae. Uropod 1 biramous, peduncle much longer than it was broad, distoventral interramal spine shorter than peduncle; rami subequal; inner ramus with apical robust setae only. Uropod 2 biramous; peduncle with distoventral interramal spine. Uropod 3 uniramous, ramus shorter than peduncle, with apical robust setae. Telson hemiacitabulate, dorsally concave, apical margin rounded.
Remark.
Unciola conchicola sp. nov. is most similar to Unciola planipes Norman, 1867.
- Via the dorsoventrally flattened body form and rugose cuticle, these species can be readily separated by the shape of the Epimera 1 to 3, where the acute projection is directed ventrally in U. conchicola sp. nov. and posteriorly in U. planipes.
- Unciola icelandica sp. nov.
- urn:lsid:zoobank.org:act:7E1C249F-8F0A-4CF2-A238-346A316DEDF3
Type material. Holotype, 11 mm, SMF 52167, North Shetland Islands, (61°25.63′ N, 1°21.07′ E–61°25.05′ N, 21.66′ E), epibenthic sledge, 169 m, 24 July 2013, expedition RV Poseidon (st 456/3-866).
Paratype female, 6 mm, (dissected, illustrated), ZMHK-62086 South Island, Iceland Basin, (61°42.630′ N, 19°32.960′ W–61°42.170′ N, 19°32.020′ W), epibenthic sledge, 1912–1921 m, 31 August 2011, expedition RV Meteor (St 85-3-989).
The following paratypes were the same locality as the first paratype: male, ZMHK 62084, juvenile 62089, male 62087 and on SEM stud ZMHK-56725.
Paratype female, one specimen, SMF 52.171 Norwegian Channel, 62°16.200′ N, 000°01.210′ E–62°16.450′ N, 000°01.810′ E, epibenthic sledge, 846–868 m, 25 July 2013, expedition RV Poseidon (St 869-3).
Further material examined: SMF 55297, two females, two juveniles, Faroe Islands Ridge middle, 63°23.360′ N, 008°09.420′ W–63°24.620′ N, 008°11.220′ W, epibenthic sledge, 686–687 m, 31 July 2013, expedition RV Poseidon (St880-2).
SMF 55295, 24 specimens, Iceland Basin, 60°2.73′ N, 21°28.06′ W–60°2.73′ N, 21°29.88′ W, epibenthic sledge, 2568–2749 m, 28 August 2011, RV Meteor (Me85/3 st 983).
- Etymology. Named from the locality of the new species.
- Type locality. North Shetland Islands
- Description. Based on type material.
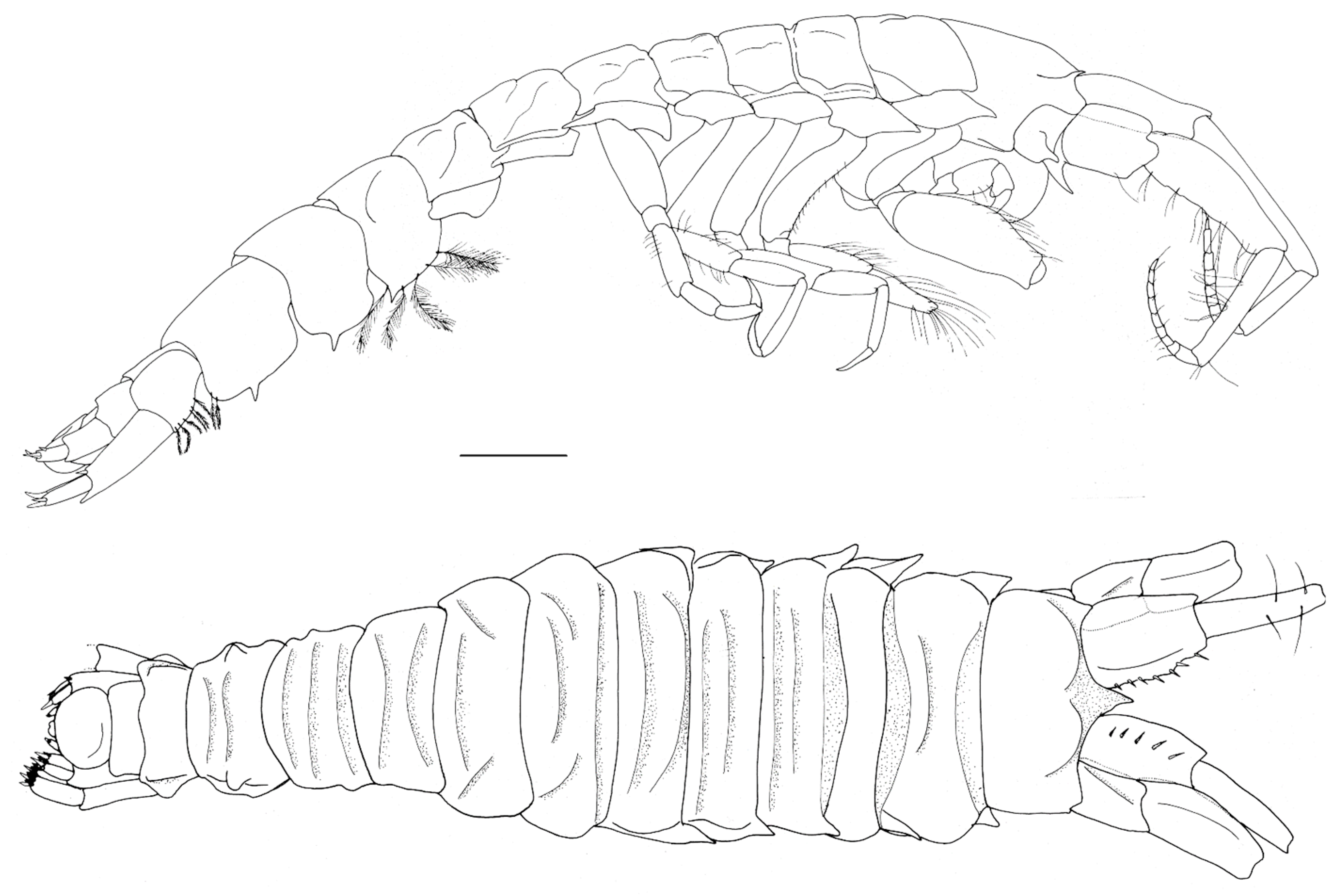
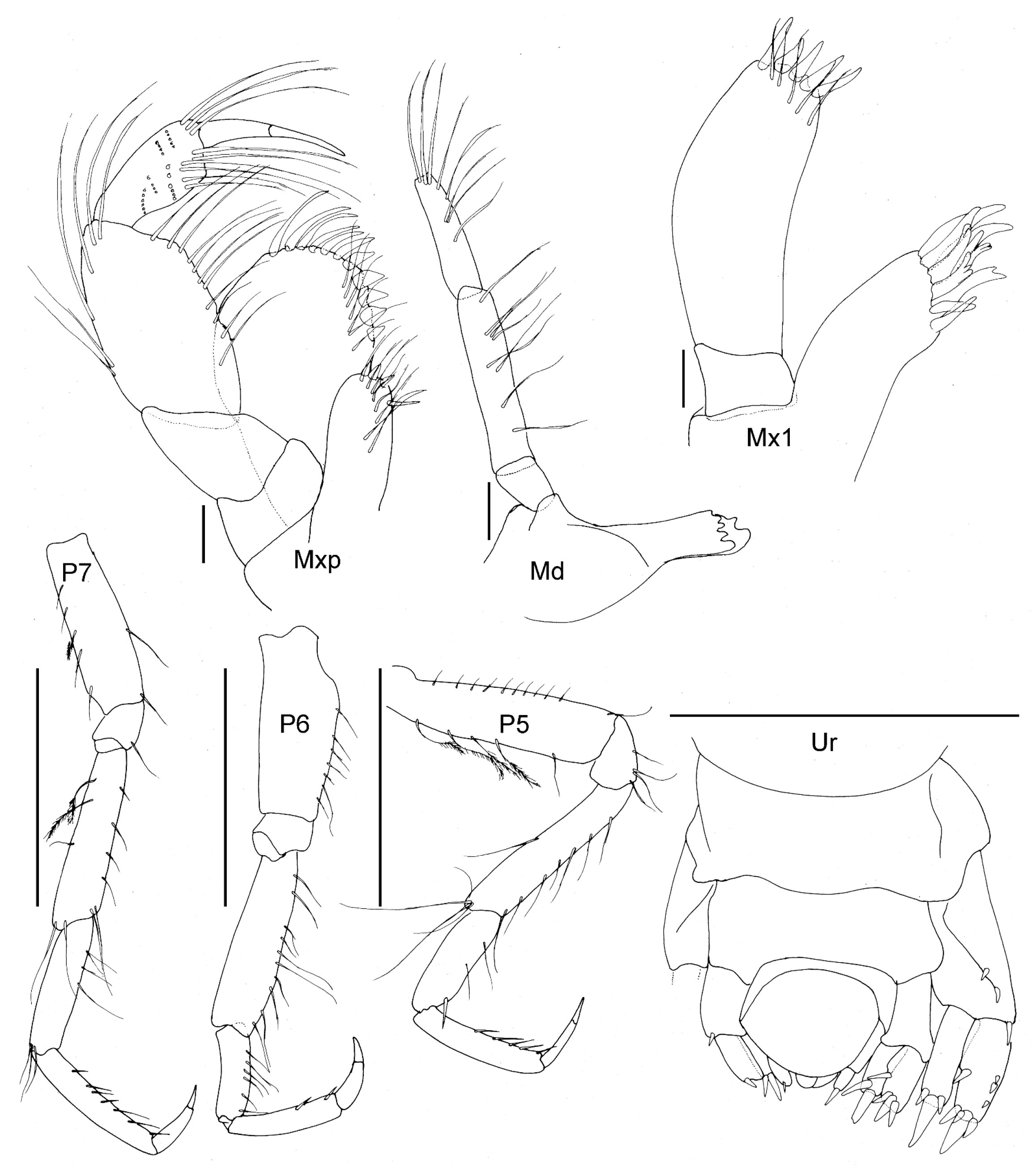
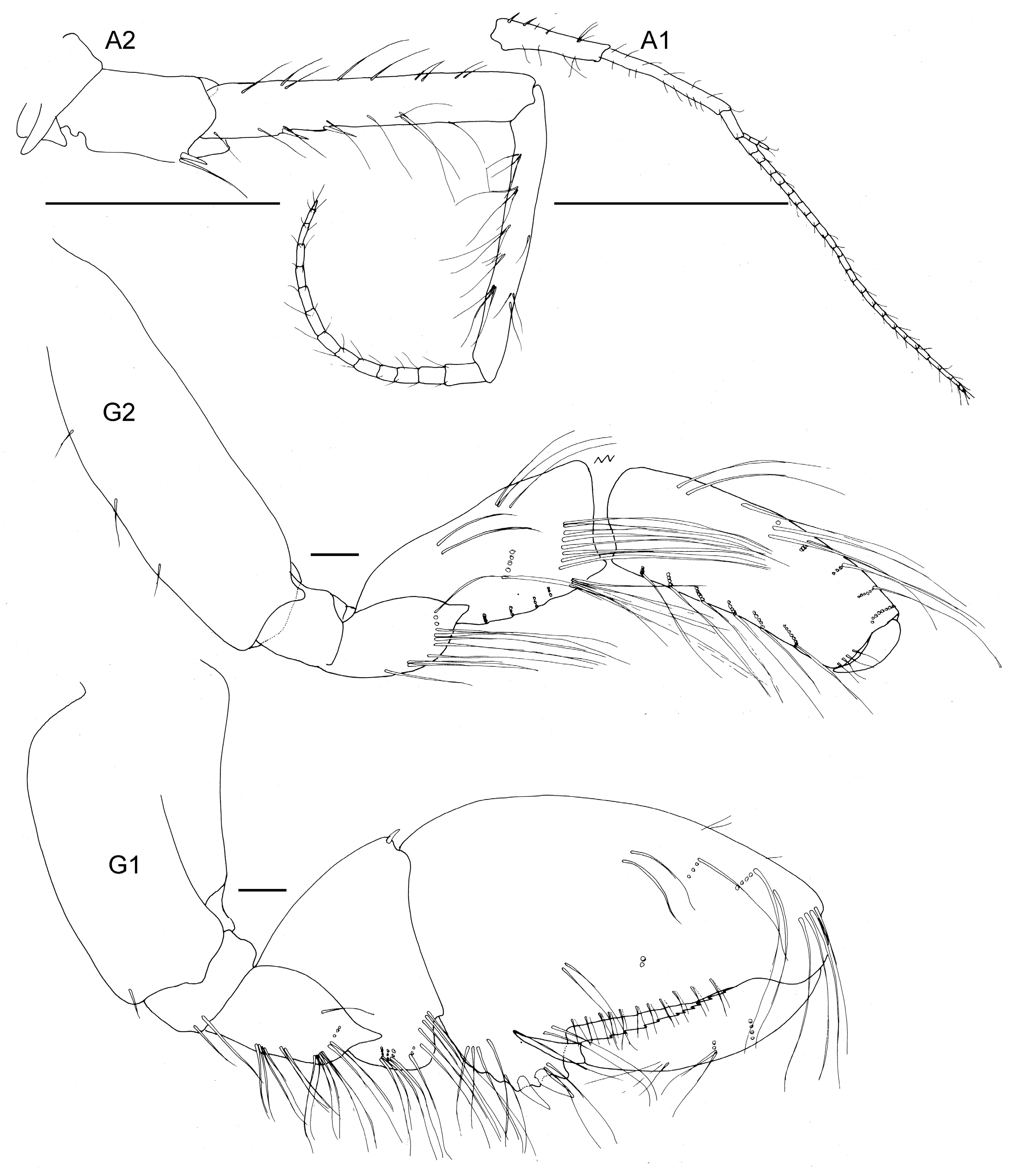
- Head. Eyes absent. Rostrum, well-developed, acute, deflexed. Lateral cephalic lobes produced, subquadrate. Antenna 1 peduncle article 1 length 5 times the width, margins lined with slender setae; article 2 twice the length of article 1; article 3 length 5 times the width, shorter than article 1, accessory flagellum 3-articulate; primary flagellum with 16 articles. Antenna 2 peduncle articles 1–2 anteroventral corner acute produced; article 3 length 1.4 times as long as it was broad, anteroventral margin with robust seta; article 4, ventral margin lined with slender setae; shorter than article 5; flagellum with 14 articles. Mandible molar absent; accessory setal row with three setae; palp article 3 shorter than 2, length 5.5 times the width, lined with long slender setae and longer apical setae. Maxilla 1 unknown. Maxilliped unknown.
- Pereon. Pereonites cuticle smooth. Pleonite 3 sternal process well-developed, directed anteriorly, apically acute. Pleonite 4 sternal process moderately developed, directed anteriorly, apically subacute. Pereonite 5 sternal process minutely developed, anteriorly developed, subacute. Coxa subrectangular to rounded, depth less than half of pereonites. Gnathopod 1 larger than gnathopod 2, coxa anteroventral corner acutely produced; basis stout, length 3 times the width, anterodistal margin rounded; ischium much broader than it was long; merus twice as long as it was broad, posterior margin lined with slender setae; carpus compressed between merus and propodus, posterior margin with sparse slender setae and without robust setae; propodus length twice the width, anterior and posterior margins with a few slender setae, posterior margin straight, palm subacute to acute, length two thirds of posterior margin, defined by rounded corner with two robust setae; dactylus closing along palm, unguis present, anterior margin lined with rows of setae, inner margin serrate. Gnathopod 2 subchelate; basis anterior and posterior margin straight without setae, anterodistal corner produced, acute; ischium length 1.2 times the width; merus length 1.5 times the width; carpus length 2.4 times the width, shorter than propodus length; propodus anterior and posterior margins lined with rows of long slender setae, length 5 times the width, palm transverse, defined by subquadrate corner. Pereopods 5 to 7 basis rectilinear; posterior margins with plumose setae; dactylus unguis present.
- Pleon. Epimeral plates 1–2 posterior margin concave, ventral margin without plumose setose, posteroventral corner acutely produced. Epimera 3 prosterior margin concave, corner produced as spine. Urosomites 1–3 not coalesced. Urosomite 1 ventral margin without plumose setae. Uropod 1 biramous, peduncle much longer than it was broad, distoventral interramal spine shorter than peduncle; rami subequal; inner ramus with marginal and apical robust setae. Uropod 2 biramous; peduncle with marginal and apical robust setae. Uropod 3 uniramous; peduncle with flange with one or two apical robust setae; ramus shorter than peduncle, with long slender apical setae. Telson dorsally concave, apical margin rounded.
Remark.
Unciola icelandica sp. nov. aligns with Unciola planipes in the shape of the Epimeron 3 with an excavate posterior margin with acute projection. These species can be separated based on the gnathopod 1 and 2. The gnathopod 1 carpus is rectilinear in U. icelandica sp. nov., in contrast to U. planipes, where the carpus is proximally broad and distally narrow. In the gnathopod 2 of U. icelandica sp. nov., the palm is straight, defined by a palm corner, while the palm in U. planipes has a proximal extension not seen in the former.
- Unciola icelandica sp. nov. has subchelate second gnathopods, while the second gnathopods of U. planipes are simple. Lastly, U. icelandica sp. nov. has subovate coxae 2 to 3, while U. leucopis has more rectilinear coxae 2 and 3.
- Unciola planipes Norman, 1867
- Unciola leucopes [43] Spence Bate and Westwood, 1868: 518.
- Material Examined
SMF 52169, four specimens; SMF 52170, eight specimens, Norwegian Channel, 61°59.830′ N, 000°30.400′ E-61°59.260′ N, 61°59.260′ N, epibenthic sledge, 290–302 m, 24 July 2013 RV Poseidon (ST867-1).
SMF 52258, four specimens, North West Iceland Denmark Strait, 66°38.600′ N, 024°31.970′ W–66°38.700′ N024°31.350′ W, epibenthic sledge, 118 m, 13 September 2011, RV Meteor (St 1104-1).
Remark.
Unciola planipes has a remarkable distribution, described from relatively shallow waters from north of Scotland to much deeper records in the Norwegian Channel. Several sampling attempts in Scotland in 2021 failed to obtain fresh material of this species for genetic comparison.
- Updated key to the North East Atlantic species of Unciola
- Epimeral plates 1 to 3 projection pointing posteriorly………………………………………………….………………..2Epimeral plates 1 to 3 projections directed ventrally………………………………………..Unciola conchicola sp. nov.
- Gnathopod 2 subchelate……………………………………………………………………………………………………..3Gnathopod 2 simple……………………………………………………………………………....U. planipes Norman, 1867
- Rostrum present, well developed …………………………………………………………………..………………………4Rostrum absent or short (not well developed) …………………………………………....U. petalocera (G.O. Sars, 1879)
- Antenna 1 shorter or same length as body……………………………………………………………..…………………..5Antenna 1 longer than body, peduncle with four robust setae, ……………………………U. laticornis Hansen, 1887
- Epimeral plate 3 posterior margin excavate, posterodistal corner with acute projections, gnathopod 2 carpus and propodus similar in length.....................................................................................................................................................6Epimeral plate 3 posterior margin entire, posterodistal corner with minute project ions, gnathopod 2 carpus shorter than propodus……….....................................................................................................….U. crassipes Hansen, 1887
- Head lateral cephalic lobe distinctly truncate…………………….......................................................U. icelandica sp. nov.Head lateral cephalic lobe rounded or acute………………………………….......................................................…………7
- Telson subovate…………………………………………………………………............................................………….……8Telson triangular…………………………………………………....................................................U. petalocera (Sars, 1876)
- Epimeral plates acute projection increasing in size; gnathopod 2 carpus length sub equal to propodus,……………………………………………..................................................................................U. leucopis (Krøyer, 1845)Epimeral plates projections subequal in size; gnathopod 2 carpus length shorter than propodus……………………………………………...............................................................U. crenatipalma (Spence Bate, 1862)
4. Discussion
While it is established that corophioid amphipods build tubes and many genera are associated with commandeering empty shells [44,45] (Myers and Lowry, 2003; Vader, 1972), the present study is the first record for the genus Unciola (Figure 14). Barnard et al. (1991) [46] distinguish 12 kinds of tube-building behaviour. Observations of how the tube building is achieved in live organisms were not possible, as these benthic amphipods were collected at a 2000 m depth. Other detailed studies of silk production in corophiideans have shown that silk is produced in the glandular systems in the third and fourth pereopods, which secrete a mucous substance that solidifies as a kind of silken thread that can trap and aggregate particles from the environment [47,48,49] (Bellan-Santini, 2015; Kroenenberger et al., 2013; Neretin, 2016).
Unciola conchicola sp. nov. specimens were removed from the tubes built within a gastropod shell (Figure 10). We conclude that Unciola conchicola sp. nov. belongs to category six of the twelve tube-building behaviours suggested by Barnard et al. (1991) [46], namely: “(6) cementation of a silk tube inside a gastropod shell or into or on the surface of other environmentally available substrates which are more or less portable and can be carried or moved by the amphipod (such as in Siphonoecetes spp.)” [38,50,51,52] (Gauthier, 1941; Just, 1977, 1984; Richter, 1978). As a secondary user dependent on shell material in the environment, these small deep-sea organisms are directly impacted by climate change, where thinning of calcium carbonate will make their domiciles a less protective barrier from predation and the amphipod may generally have less access to suitable domiciles, as thinner shells are more likely to break and fragment in the sediment system.
Author Contributions
L.E.H. and A.-N.L. contributed in equal parts to this publication. Conceptualization, A.-N.L. and L.E.H.; methodology A.-N.L. and L.E.H.; validation, A.-N.L. and L.E.H.; formal analysis, A.-N.L. and L.E.H.; investigation, A.-N.L. and L.E.H.; data curation, A.-N.L. and L.E.H.; writing—original draft preparation, A.-N.L. and L.E.H.; writing—review and editing, A.-N.L.; visualization, A.-N.L. and L.E.H. All authors have read and agreed to the published version of the manuscript.
Funding
The corresponding author was funded by the German Science Foundation LO2543/1-1.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article; new species are described based on material curated by the Senckenberg Museum Frankfurt and the Natural History Museum Hamburg. No new data were created or analyzed in this study.
Acknowledgments
Renate Walter (University Hamburg) handled the scanning electron microscope. Vivien Hartman (University Hamburg) kindly helped with creating the map. The Senckenberg Museum Frankfurt is thanked for the curation of material. Saskia Brix (DZMB Hamburg) is thanked for organizing the IceAGE expeditions. Angelika Brandt is thanked for the loan of material from the Museum of Natural History in Frankfurt. We thank four anonymous reviewers for their comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brix, S.; Lörz, A.-N.; Jażdżewska, A.M.; Hughes, L.; Tandberg, A.H.S.; Pabis, K.; Stransky, B.; Rapp-Schickel, T.; Corbe, J.C.; Hendryks, E.; et al. Amphipod family distributions around Iceland. Zookeys 2018, 731, 1–53. [Google Scholar] [CrossRef] [PubMed]
- Horton, T.; Lowry, J.; De Broyer, C.; Bellan-Santini, D.; Coleman, C.O.; Corbari, L.; Costello, M.J.; Daneliya, M.; Dauvin, J.-C.; Fišer, C.; et al. World Amphipoda Database. Unciola Say, 1818. World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=101474 (accessed on 13 March 2023).
- Vader, W.; Brattegard, T.; Buhl-Mortensen, L.; Larsen, K.M. Order Amphipoda, suborder Gammaridea (phylum Crustacea)—Gammaridean amphipods. In Distribution of Marine, Benthic Macro-Organisms in Norway; Brattegard, T., Holthe, T., Eds.; Directorate of Nature Management: Trondheim, Norway, 1997; pp. 191–213. [Google Scholar]
- Buhl-Jensen, L. The benthic amphipod fauna of the west-Norwegian continental shelf compared with the fauna of five adjacent fjords. Sarsia 1986, 71, 193–208. [Google Scholar] [CrossRef]
- Stephensen, K. Crustacea Malacostraca VIII: Amphipoda IV. In The Danish Ingolf Expedition; H. Hagerup: Copenhagen, Denmark, 1944; Volume 3, pp. 1–54. [Google Scholar]
- Buhl-Mortensen, P.; Buhl-Mortensen, L.; Purser, A. Trophic ecology and habitat provision in cold-water coral ecosystems. In Marine Animal Forests; Rossi, S., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 919–944. [Google Scholar] [CrossRef]
- Brix, S.; Meißner, K.; Kaiser, S.; Svavarsson, J.; Yasuhara, M.; Martinez Arbizu, P. The IceAGE Project—A Follow up of BIOICE. Pol. Polar Res. 2014, 35, 141–150. [Google Scholar] [CrossRef]
- Brenke, N. An epibenthic sledge for operations on marine soft bottom and bedrock. Mar. Technol. Soc. J. 2005, 39, 10–21. [Google Scholar] [CrossRef]
- Riehl, T.; Brenke, N.; Brix, S.; Driskella, A.; Kaiser, S.; Brandt, A. Field and laboratory methods for DNA studies on deep−sea isopod crustaceans. Pol. Polar Res. 2014, 35, 203–224. [Google Scholar] [CrossRef]
- Boeck, A. Bemaerkninger angaaende de ved de norske kyster forekommende Amphipoder. Forh. Ved De Skand. Nat. Mfte 1861, 8, 631–677. [Google Scholar]
- Boeck, A. Crustacea Amphipoda Borealia et Arctica; Forhandlinger i Videnskabs-Selskabet i Christiania 1870; Brøgger & Christie: Oslo, Norway, 1870; pp. 81–280. [Google Scholar] [CrossRef]
- Boeck, A. De Skandinaviske og Arktiske Amphipoder. Part 2; A.W. Brøgger: Oslo, Norway, 1876. [Google Scholar] [CrossRef]
- Smith, S.I. Preliminary notice of the Crustacea dredged, in 64 to 325 fathoms, off the south coast of New England, by the United States Fish Commission in 1880. Proc. USA Natl. Mus. 1881, 3, 413–452. [Google Scholar] [CrossRef]
- Hansen, H.J. Malacostraca marina Groenlandiae occidentalis. Oversight over det vestlige Gronlands fauna af malakostrake havkrebsdyr. Vidensk. Meddel. Naturhist. Foren. Kjøbenhavn 1888, 4, 5–226. [Google Scholar]
- Della Valle, A. Gammarini del Golfo di Napoli. Fauna Und Flora Golf. Neapel 1893, 20, 941–961. [Google Scholar]
- Sars, G.O. Crustacea Caspia. Contributions to the knowledge of the carcinological fauna of the Caspian Sea. Part III. Gammaridae (continued). Bull. Acad. Imp. Sci. St-Pétersbg. 1894, 1, 343–378. [Google Scholar] [CrossRef] [Green Version]
- Stebbing, T.R.R. Amphipoda. I. Gammaridea. In Das Tierreich; Schulze, F.E., Ed.; Verlag von R. Friedländer und Sohn: Berlin, Germany, 1906; Volume 21, pp. 1–806. [Google Scholar]
- Shoemaker, C.R. Descriptions of two new amphipod crustaceans (Talitridae) from the United States. J. Wash. Acad. Sci. 1930, 20, 107–114. [Google Scholar]
- Stephensen, K. The Godthaab expedition 1928. Amphipoda. Meddelelser om Grønland; C. A. Reitzels Forlag: Kopenhagen, Denmark, 1933; Volume 79, pp. 1–88. [Google Scholar]
- Stephensen, K. The Amphipoda of N. Norway and Spitsbergen with Adjacent Waters. Fasc. 3.; K. Karlsens Bok- & Aksidenstrykkeri: Tromso, Norway, 1942; Volume 3, pp. 363–526. [Google Scholar]
- Enequist, P. Studies on the soft-bottom amphipods of the Skagerak. Zool. Bidr. Upps. 1949, 28, 297–492. [Google Scholar]
- Coyle, K.O.; Mueller, G.J. New records of Alaskan marine Crustacea, with descriptions of two new gammaridean Amphipoda. Sarsia 1981, 66, 7–18. [Google Scholar] [CrossRef]
- Gurjanova, E.F. Bokoplavy morei SSSR i sopredelnykh vod (Amphipoda, Gammaridea). Amphipods of the seas of USSR and adjacent waters (Amphipoda, Gammaridea). Opred. Faune S.S.S.R. Akad. Nauk SSSR 1951, 41, 1–1029. [Google Scholar]
- Gurjanova, E.F. [New additions to the Far Eastern fauna of marine gammarids]. Opred. Faune S.S.S.R. Tr. Zool. Inst. 1953, 13, 216–241. [Google Scholar]
- Barnard, J.L.; Karaman, G.S. The families and genera of marine gammaridean Amphipoda (except marine gammaroids). Rec. Aust. Mus. 1991, 13, 1–866. [Google Scholar] [CrossRef] [Green Version]
- Sars, G.O. Oversigt af Norges Crustaceer med Foreløbige Bemærkninger over de nye Eller Mindre Bekjendte Arter; Chra: Oslo, Norway, 1882; Volume 18, pp. 1–124. [Google Scholar]
- d’Udekem d’Acoz, C. The genera Haliragoides and Neohela in the North Atlantic, with the description of two new deepwater species from Norway and Svalbard (Crustacea: Amphipoda). Cah. Biol. Mar. 2007, 48, 17–35. [Google Scholar]
- Sars, G.O. Den Norske Nordhavs-Expedition 1876–1878, Zoologi, Crustacea; Grøndahl & Søns Bogtrykkeri: Oslo, Norway, 1885; Volume 280, 96p. [Google Scholar]
- Chevreux, E. Sur quelques intéressantes espèces d’Amphipodes provenant de la dernière campagne du yacht Princesse-Alice. Bull. Soc. Zool. Fr. 1899, 24, 147–152. [Google Scholar]
- Chevreux, E. Amphipodes Provenant des Campagnes de l’Hirondelle (1885–1888). Résultats des Campagnes Scientifiques Accomplies sur son Yacht par Albert 1er; Prince Souverain de Monaco: Monaco, 1900; Volume 16, pp. 1–195. [Google Scholar]
- Piepenburg, D.; Chernova, N.V.; Von Dorrien, C.F.; Gutt, J. Megabenthic communities in the waters around Svalbard. Polar Biol. 1996, 16, 431–446. [Google Scholar] [CrossRef]
- Weslawski, J.M.; Wlodarska-Kowalczuk, M.; Legeżyńska, J. Occurrence of soft bottom macrofauna along the depth gradient in High Arctic, 79° N. Pol. Polar Res. 2003, 24, 73–88. [Google Scholar]
- Say, T. An account of the Crustacea of the United States. Proc. Acad. Nat. Sci. Phila. 1818, 1, 374–401. [Google Scholar]
- Barnard, J.L. Gammaridean Amphipoda of the rocky intertidal of California: Monterey Bay to La Jolla. Bull. Am. Mus. Nat. Hist. 1969, 258, 1–230. [Google Scholar] [CrossRef]
- Barnard, J.L. Revision of Corophiidae and related families (Amphipoda). Sm. C. Zool. 1973, 151, 1–27. [Google Scholar] [CrossRef]
- Karaman, G.S. Contribution to the knowledge of the Amphipoda 116. Revision of some genera of family Corophiidae with description of three new genera. Poljopr. Sumar. Titograd. 1981, 26, 3–12. [Google Scholar]
- Kroyer, H.N. Karcinologiske Bidrag. Naturhist. Tidsskr. 1845, 1, 283–345. [Google Scholar]
- Gauthier, H. Sur l’ethologie d’un amphipode qui vit dans une coquille. Bull. Soc. Hist. Nat. Afr. N. 1941, 32, 245–266. [Google Scholar]
- Norman, A.M. Report on the Crustacea. In Deep Sea Dredgings on the Coast of Northumberland and Durham, 1862–1864. Natural History Transactions of Northumberland, Durham, and Newcastle-upon-Tyne; Tyneside Naturalists’ Field Club: Newcastle-upon-Tyne, UK, 1865; Volume 1, pp. 12–29. [Google Scholar]
- Chevreux, E.; Fage, L. Amphipodes Faune de France; Paul Lechevalier: Paris, France, 1925; Volume 9, pp. 1–488. [Google Scholar]
- Schellenberg, A. Krebstiere oder Crustacea IV: Flohkrebse oder Amphipoda; Die Tierwelt Deutschlands: Jena, Germany, 1942; Volume 40, pp. 1–252. [Google Scholar]
- Lincoln, R.J. British Marine Amphipoda: Gammaridea; British Museum (Natural History): London, UK, 1979; p. 658. [Google Scholar]
- Spence Bate, C.; Westwood, J.O. A History of the British Sessile-Eyed Crustacea; John Van Voorst: London, UK, 1868. [Google Scholar]
- Myers, A.A.; Lowry, J.K. A phylogeny and a new classification of the Corophiidea Leach, 1814 (Amphipoda). J. Crust. Biol. 2003, 23, 443–485. [Google Scholar] [CrossRef] [Green Version]
- Vader, W. Associations between amphipods and molluscs. A review of published records. Sarsia 1972, 48, 13–18. [Google Scholar] [CrossRef]
- Barnard, J.L.; Sandved, K.; Thomas, J.D. Tube-building behavior in Grandidierella, and two species of Cerapus. Hydrobiologia 1991, 223, 239–254. [Google Scholar] [CrossRef]
- Bellan-Santini, D. Order Amphipoda Latreille, 1816. In Treatise on Zoology-Anatomy, Taxonomy, Biology. The Crustacea; Von Vaupel Klein, J.C., Charmantier-Daures, M., Schram, F.R., Eds.; Brill: Leiden, The Netherlands, 2015; Volume 5, pp. 93–248. [Google Scholar]
- Kronenberger, K.; Moore, P.G.; Halcrow, K.; Vollrath, F. Spinning a marine silk for the purpose of tube-building, J. Crustac. Biol. 2012, 32, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Neretin, N.Y. The morphology and ultrastructure of “amphipod silk” glands in Ampithoe rubricata (Crustacea, Amphipoda, Ampithoidae). Biol. Bull. Russ. Acad. Sci. 2016, 43, 628–642. [Google Scholar] [CrossRef]
- Just, J. A new genus and species of corophiid Amphipoda from pteropod shells of the bathyal western Atlantic, with notes on related genera (Crustacea). Steenstrupia 1977, 4, 131–138. [Google Scholar]
- Just, J. Siphonoecetinae (Crustacea, Amphipoda, Corophiidae) 2: Caribboecetes Just, 1983, with description of six new species. Steenstrupia 1984, 10, 37–64. [Google Scholar]
- Richter, G. Beobachtungen zu Entwicklung und Verhalten von Phronima sedentaria (Forskal), (Amphipoda). Senckenberg. Marit. 1978, 10, 229–242. [Google Scholar]
Figure 1.
Locations of Unciola species found in the North Atlantic based on the material collected during IceAGE expeditions.
Figure 1.
Locations of Unciola species found in the North Atlantic based on the material collected during IceAGE expeditions.

Figure 2.
Photo image of Neohela monstrosa (Boeck, 1861) male, 19 mm, SMF 52.259, Norwegian Sea, Norwegian Basin, North Atlantic, 1886 m. Scale: 1 mm.
Figure 2.
Photo image of Neohela monstrosa (Boeck, 1861) male, 19 mm, SMF 52.259, Norwegian Sea, Norwegian Basin, North Atlantic, 1886 m. Scale: 1 mm.

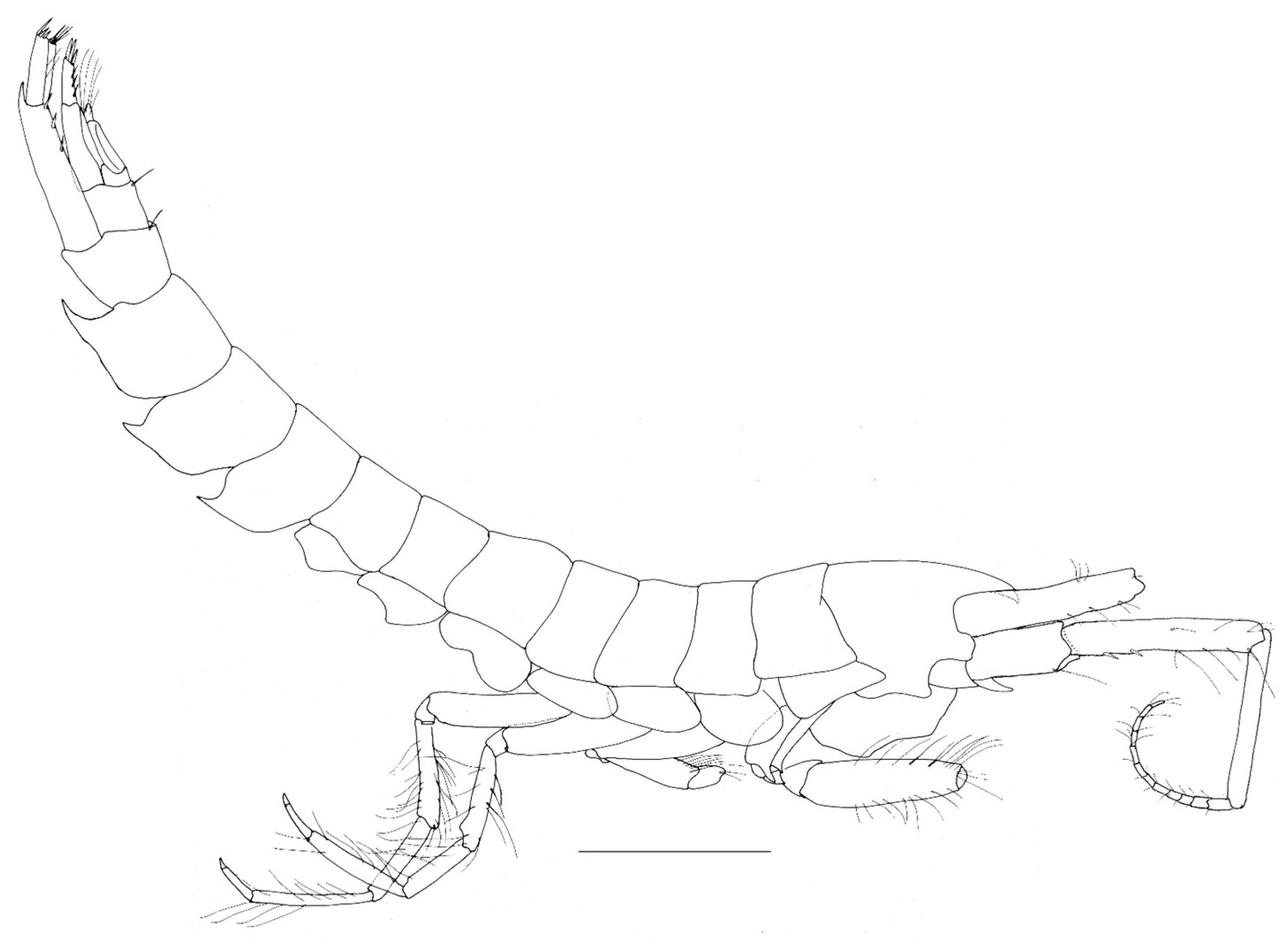
Figure 3.
Neohela monstrosa (Boeck, 1861) male, 19 mm, SMF 52.259, Norwegian Sea, Norwegian Basin, North Atlantic, 1886 m; scale habitus: 1 mm, gnathopods 1–2 scale: 0.5 mm.
Figure 3.
Neohela monstrosa (Boeck, 1861) male, 19 mm, SMF 52.259, Norwegian Sea, Norwegian Basin, North Atlantic, 1886 m; scale habitus: 1 mm, gnathopods 1–2 scale: 0.5 mm.

Figure 4.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.
Figure 4.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.

Figure 5.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.
Figure 5.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.

Figure 6.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.
Figure 6.
Unciola conchicola sp. nov. holotype male, 10 mm, ZMHK-62083, South Island, Iceland Basin, 1921 m. Scale: 0.5 mm.

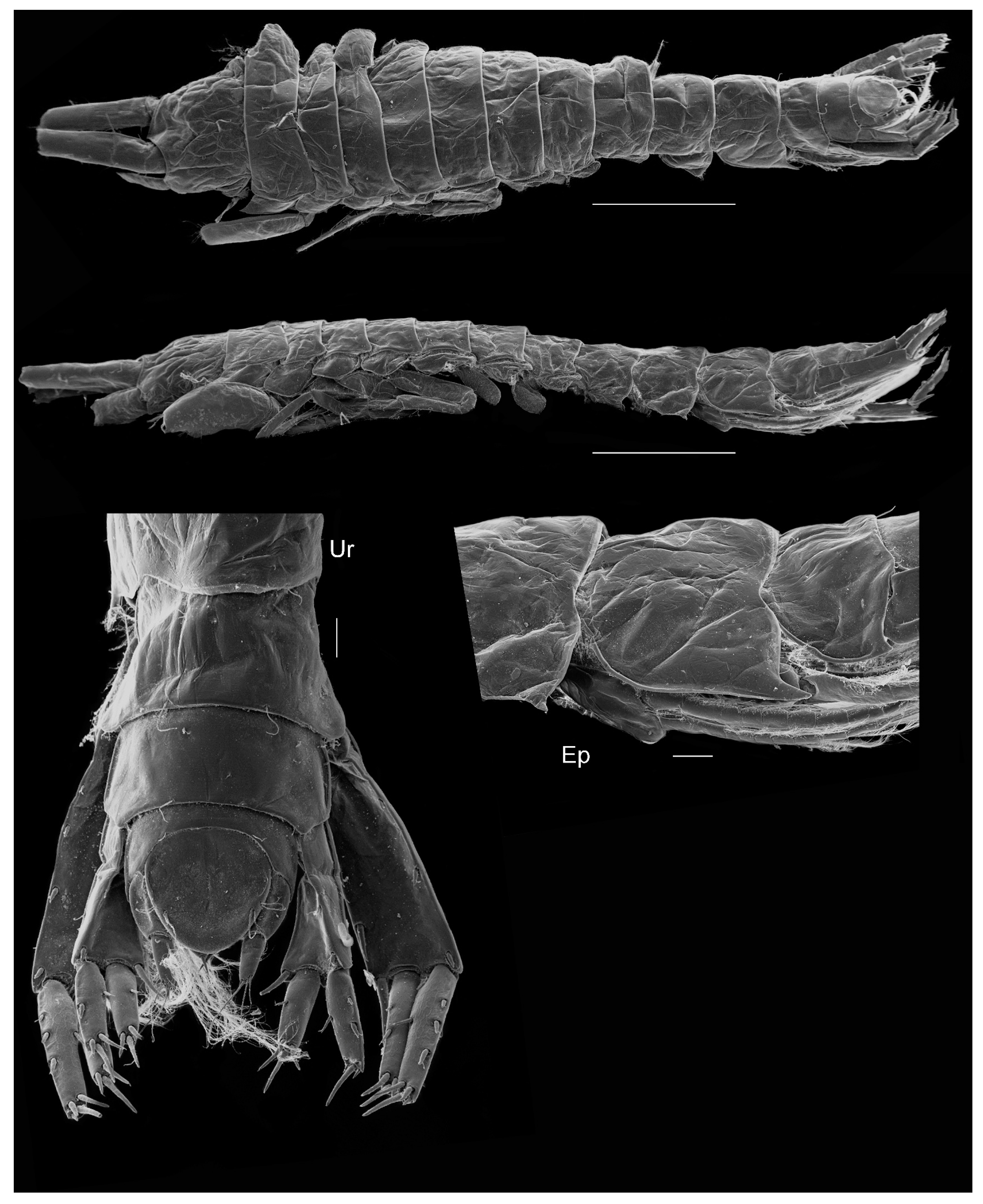
Figure 7.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Scales: habitus 1 mm, urosomites, uropod and pereopod 7 0.1 mm.
Figure 7.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Scales: habitus 1 mm, urosomites, uropod and pereopod 7 0.1 mm.

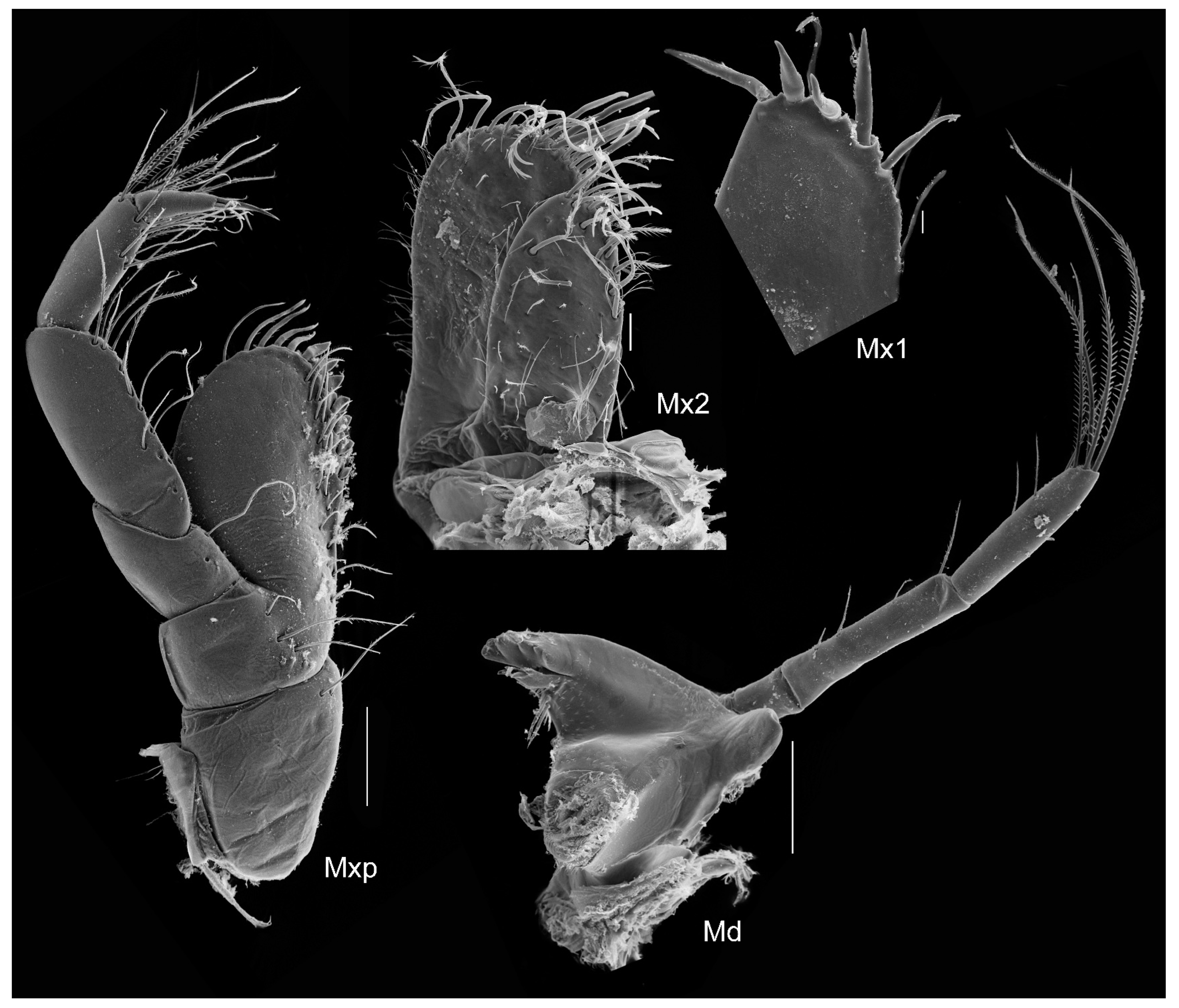
Figure 8.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Scales: 0.1 mm, details of terminal end of maxillar plate and mandibular incisor 10 µm.
Figure 8.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Scales: 0.1 mm, details of terminal end of maxillar plate and mandibular incisor 10 µm.

Figure 9.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Arrow pointing to detail of palm and dactylus. Scales: 100 µm, detail tip of dactylus 20 µm.
Figure 9.
Unciola conchicola sp. nov. SEM image, paratype male, 7 mm, ZMHK-56724, South Island, Iceland Basin, 1921 m. Arrow pointing to detail of palm and dactylus. Scales: 100 µm, detail tip of dactylus 20 µm.

Figure 10.
Unciola conchicola sp. nov., juvenile, ZMHK 62085, South Island, Iceland Basin, 1921 m. (A) in tube made of sand; (B) removed from tube; (C) tube in pteropod shell, Scale: 1 mm.
Figure 10.
Unciola conchicola sp. nov., juvenile, ZMHK 62085, South Island, Iceland Basin, 1921 m. (A) in tube made of sand; (B) removed from tube; (C) tube in pteropod shell, Scale: 1 mm.

Figure 11.
Unciola icelandica sp. nov. holotype male, 11 mm, SMF 52167, North Shetland Islands, 169 m. Scale: 1 mm.
Figure 11.
Unciola icelandica sp. nov. holotype male, 11 mm, SMF 52167, North Shetland Islands, 169 m. Scale: 1 mm.

Figure 12.
Unciola icelandica sp. nov. holotype male, 11 mm, SMF 52167, North Shetland Islands, 169 m. Scale: 0.5 mm.
Figure 12.
Unciola icelandica sp. nov. holotype male, 11 mm, SMF 52167, North Shetland Islands, 169 m. Scale: 0.5 mm.

Figure 13.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scale: 1 mm.
Figure 13.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scale: 1 mm.

Figure 14.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scales: Antennae 1 and 2 1 mm, gnathopods 1 and 2 0.1 mm.
Figure 14.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scales: Antennae 1 and 2 1 mm, gnathopods 1 and 2 0.1 mm.

Figure 15.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scales: 0.1 mm.
Figure 15.
Unciola icelandica sp. nov. paratype female, 6 mm, ZMHK 62086, South Island, Iceland Basin, 1921 m. Scales: 0.1 mm.

Figure 16.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin North 1921 m. Scales: habitus 1 mm, urosome 100 µm.
Figure 16.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin North 1921 m. Scales: habitus 1 mm, urosome 100 µm.

Figure 17.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin, 1921 m. Scales: mandible and maxilliped 100 µm, maxilla 2 20 µm, tip of maxilla 1–10 µm.
Figure 17.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin, 1921 m. Scales: mandible and maxilliped 100 µm, maxilla 2 20 µm, tip of maxilla 1–10 µm.

Figure 18.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin, 1921 m. Scales: gnathopod 1 and 2–100 µm, dactylus tip 20 µm.
Figure 18.
Unciola icelandica sp. nov. SEM image, paratype female, 6 mm, ZMHK-56725, South Island, Iceland Basin, 1921 m. Scales: gnathopod 1 and 2–100 µm, dactylus tip 20 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Examined material of unciolid genera Unciola and Neohela collected during IceAGE expeditions.
Table 1.
Examined material of unciolid genera Unciola and Neohela collected during IceAGE expeditions.
| Species | Type | Number of Specimens | Sex | Latitude [dec] | Longitude [dec] | Depth [m] | Collection ID | Station | Expedition |
|---|---|---|---|---|---|---|---|---|---|
| Unciola conchicola sp. nov. | Holotype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK62083 | 989-3 | Me85 |
| Unciola conchicola sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-62088 | 989-3 | Me85 |
| Unciola conchicola sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-62089 | 989-3 | Me85 |
| Unciola conchicola sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-62090 | 989-3 | Me85 |
| Unciola conchicola sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-56724 | 989-3 | Me85 |
| Unciola conchicola sp. nov. | juv in shell | 61.718 | −18.44 | 1921 | ZMHK-62085 | 989-3 | Me85 | ||
| Unciola conchicola sp. nov. | 2 | 1 m, 1 juv | 63.393 | −7.838 | 867 | SMF 52.175 | 880-2 | POS456 | |
| Unciola icelandica sp. nov. | Holotype | 1 | m | 61.418 | 1.352 | 169 | SMF 52.167 | 866-3 | POS456 |
| Unciola icelandica sp. nov. | Paratype | 1 | f | 61.718 | −18.44 | 1921 | ZMHK-62086 | 989-3 | Me85 |
| Unciola icelandica sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-62084 | 989-3 | Me85 |
| Unciola icelandica sp. nov. | Paratype | 2 | 1 juv | 61.718 | −18.44 | 1921 | ZMHK-62089 | 989-3 | Me85 |
| Unciola icelandica sp. nov. | Paratype | 1 | m | 61.718 | −18.44 | 1921 | ZMHK-62087 | 989-3 | Me85 |
| Unciola icelandica sp. nov. | Paratype | 1 | f | 62.272 | 0.023 | 868 | SMF 52.171 | 869-3 | POS456 |
| Unciola icelandica sp. nov. | Paratype | 1 | f | 61.718 | −18.44 | 1921 | ZMHK-56725 | 989-3 | Me85 |
| Unciola icelandica sp. nov. | 4 | 2 f, 2 juv | 63.393 | −7.838 | 687 | SMF 55.297 | 880-2 | POS456 | |
| Unciola icelandica sp. nov. | 24 | 60.035 | −20.533 | 2749 | SMF 55.295 | 983-3 | Me85 | ||
| Unciola planipes Norman, 1867 | 4 | 62.006 | 0.511 | 302 | SMF 52.169 | 867-1 | POS456 | ||
| Unciola planipes Norman, 1867 | 8 | 62.006 | 0.511 | 302 | SMF 52.170 | 867-1 | POS456 | ||
| Unciola planipes Norman, 1867 | 4 | 66.650 | −23.456 | 118 | SMF 52.258 | 1104-1 | Me85 | ||
| Neohela monstrosa G.O. Sars, 1885 | 1 | m | 67.663 | 12.233 | 1826 | SMF 52.259 | 1181-1 | Me85 | |
| Neohela monstrosa G.O. Sars, 1885 | 1 | intersex | 67.663 | 12.233 | 1826 | SMF 52298 | 1181-1 | Me85 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hughes, L.E.; Lörz, A.-N. Unciolidae of Deep-Sea Iceland (Amphipoda, Crustacea). Diversity 2023, 15, 546. https://doi.org/10.3390/d15040546
AMA Style
Hughes LE, Lörz A-N. Unciolidae of Deep-Sea Iceland (Amphipoda, Crustacea). Diversity. 2023; 15(4):546. https://doi.org/10.3390/d15040546
Chicago/Turabian StyleHughes, Lauren E., and Anne-Nina Lörz. 2023. "Unciolidae of Deep-Sea Iceland (Amphipoda, Crustacea)" Diversity 15, no. 4: 546. https://doi.org/10.3390/d15040546
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.