

Chemical Diversity and Redox Values Change as a Function of Temporal Variations of the Essential Oil of a Tropical Forest Shrub

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Design
2.3. Essential Oil Extraction
2.4. Essential Oil Analysis
2.5. Chemical Diversity, Micromolecular Parameters and Chemometric Analysis
3. Results
3.1. Essential Oil Yields
3.2. Chemical Profile of the Essential Oil
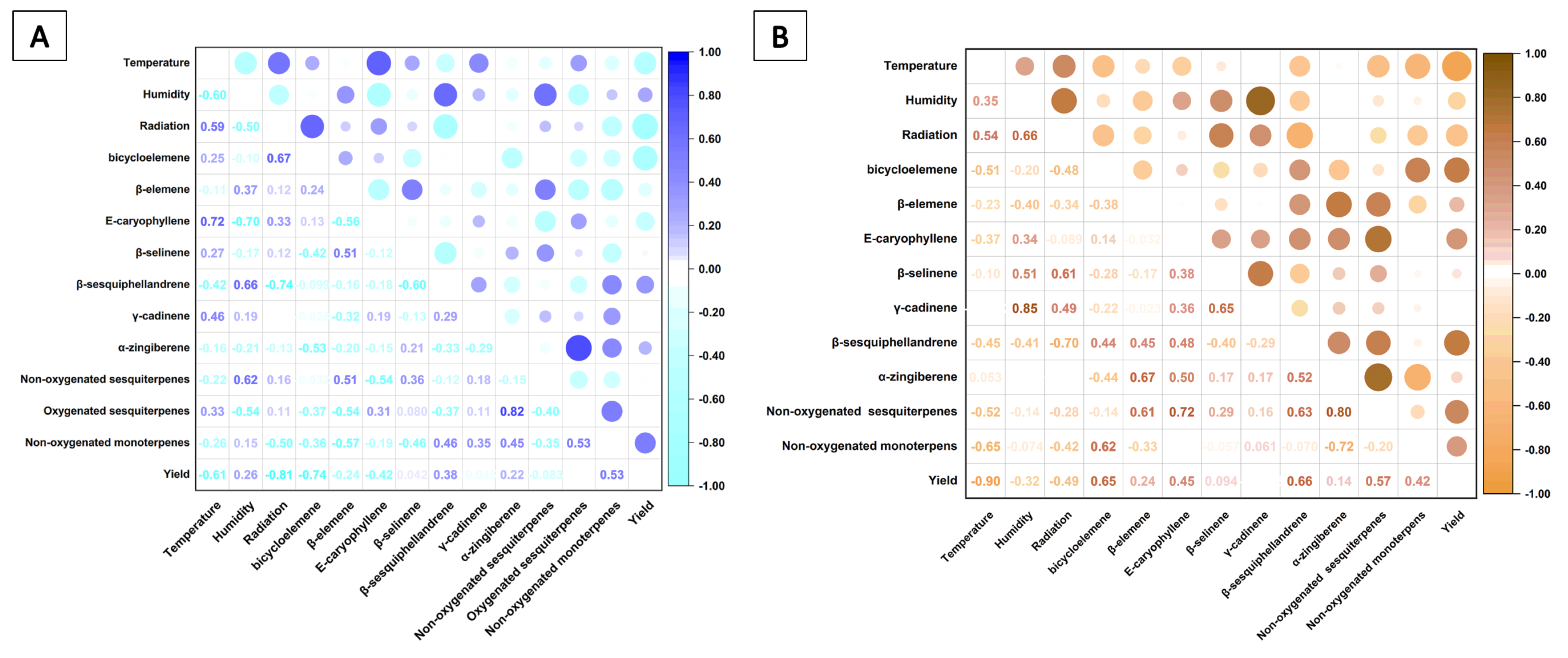
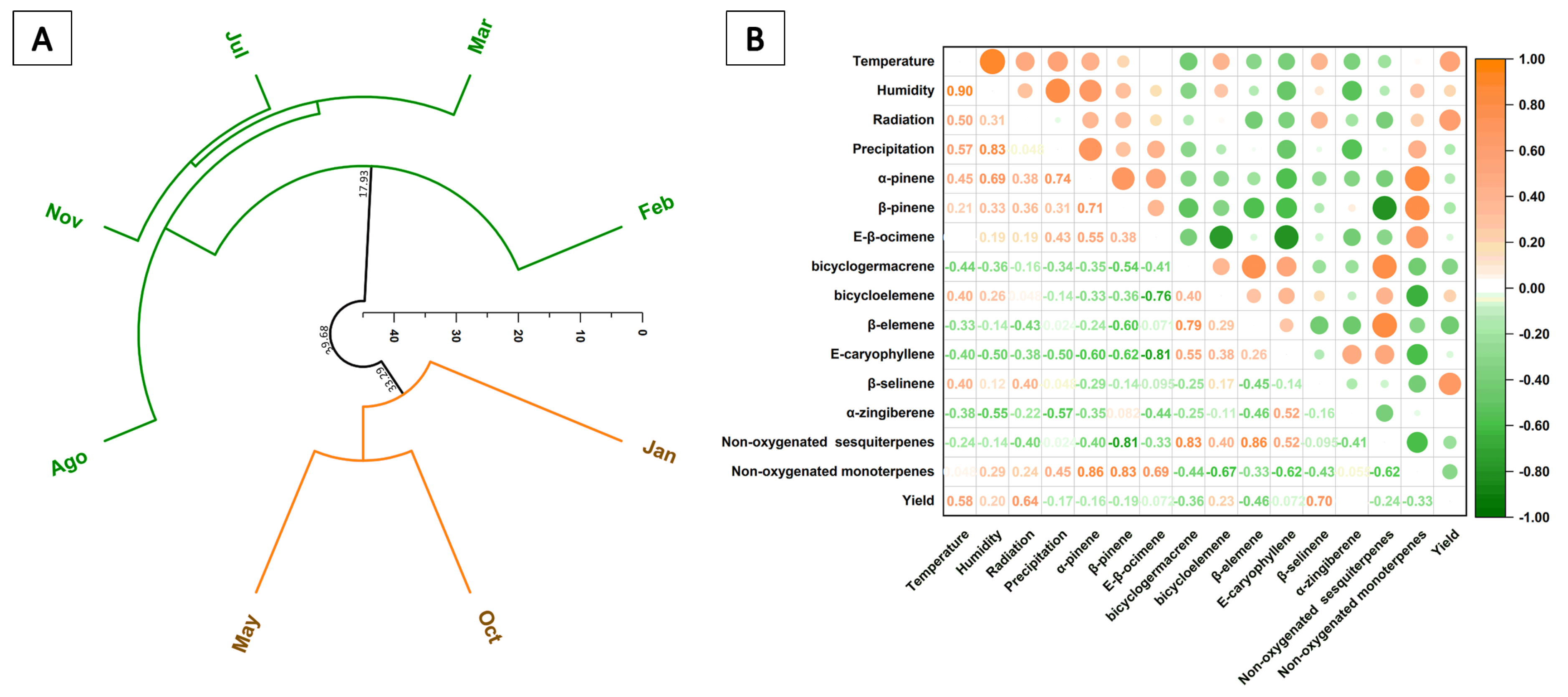
3.3. Seasonal Temporal Variation of Essential Oil Components
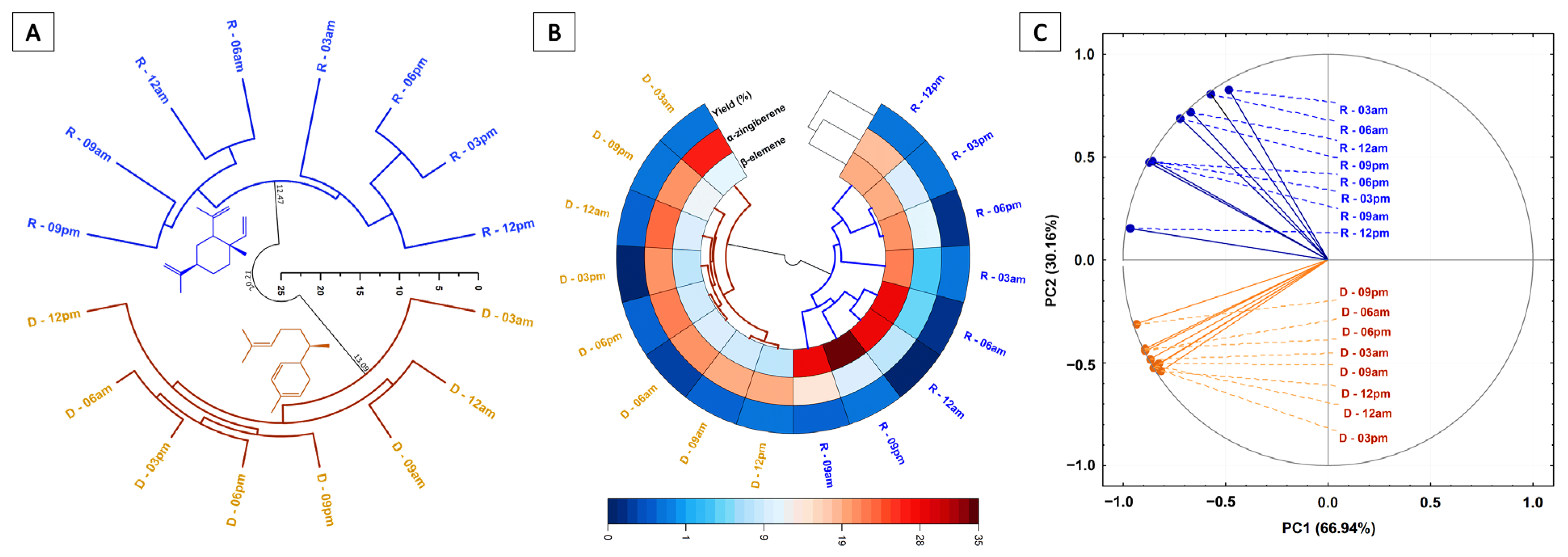
3.4. Circadian Temporal Variation of Essential Oil Components
3.5. Temporal Variation of Chemical Diversity and Redox Patterns in Essential Oils
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Theodoridis, S.; Drakou, E.G.; Hickler, T.; Thines, M.; Nogues-Bravo, D. Evaluating natural medicinal resources and their exposure to global change. Lancet Planet. Health 2023, 7, e155–e163. [Google Scholar] [CrossRef] [PubMed]
- Ramos, Y.J.; Gouvêa-Silva, J.G.; de Brito, M.D.; Felisberto, J.S.; Pereira, R.C.; Sadgrove, N.J.; de Lima Moreira, D. Chemophenetic and Chemodiversity Approaches: New Insights on Modern Study of Plant Secondary Metabolite Diversity at Different Spatiotemporal and Organizational Scales. Rev. Bras. Farmacogn. 2023, 33, 49–72. [Google Scholar] [CrossRef]
- Kessler, A.; Kalske, A. Plant secondary metabolite diversity and species interactions. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 115–138. [Google Scholar] [CrossRef]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The reemergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef]
- Costa-Oliveira, C.; Ramos, Y.J.; de Queiroz, G.A.; Guimarães, E.F.; Viçosa, A.L.; de Lima Moreira, D. Essential oils from Piper lhotzkyanum Kunth leaves from Brazilian Atlantic forest: Chemical composition and stability in different storage conditions. J. Oleo Sci. 2021, 70, 995–1005. [Google Scholar] [CrossRef]
- Brito-Machado, D.; Ramos, Y.J.; Defaveri, A.C.A.E.; de Queiroz, G.A.; Guimarães, E.F.; de Lima Moreira, D. Volatile Chemical Variation of Essential Oils and Their Correlation with Insects, Phenology, Ontogeny and Microclimate: Piper mollicomum Kunth, a Case of Study. Plants 2022, 11, 3535. [Google Scholar] [CrossRef]
- Ramos, Y.J.; Felisberto, J.S.; Gouvêa-Silva, J.G.; de Souza, U.C.; da Costa-Oliveira, C.; de Queiroz, G.A.; Moreira, D.L. Phenoplasticity of essential oils from two species of Piper (Piperaceae): Comparing wild specimens and bi-generational monoclonal cultivars. Plants 2022, 11, 1771. [Google Scholar] [CrossRef]
- Ramos, Y.J.; Machado, D.B.; Queiroz, G.A.; Guimarães, E.F.; Defaveri, A.C.A.; Moreira, D.L. Chemical composition of the essential oils of circadian rhythm and of different vegetative parts from Piper mollicomum Kunth-A medicinal plant from Brazil. Biochem. Syst. Ecol. 2020, 92, 104116. [Google Scholar] [CrossRef]
- Ramos, Y.J.; Costa-Oliveira, C.D.; Candido-Fonseca, I.; Queiroz, G.A.D.; Guimarães, E.F.; Defaveri, A.C.A.E.; Moreira, D.D.L. Advanced chemophenetic analysis of essential oil from leaves of Piper gaudichaudianum Kunth (piperaceae) using a new reduction-oxidation index to explore seasonal and circadian rhythms. Plants 2021, 10, 2116. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- Shinoda, N.; Shiga, M.; Nishimura, K. Constituents of yuzu (Citrus junos) oil. Agric. Biol. Chem. 1970, 34, 2. [Google Scholar] [CrossRef]
- Ramos, Y.J.E.; Costa-Oliveira, C.; Fonseca, I.C.; da Silva Marcelino, D.; Heggdorne-Araujo, M.; Lassounskaia, E.B.; de Lima Moreira, D. Piper multinodum C. DC. (Piperaceae) essential oils chemical variation and biological activity against Mycobacterium tuberculosis. J. Med. Plants Res. 2021, 15, 413–422. [Google Scholar]
- Hazrati, S.; Beidaghi, P.; Beyraghdar Kashkooli, A.; Hosseini, S.J.; Nicola, S. Effect of harvesting time variations on essential oil yield and composition of Sage (Salvia officinalis). Horticulturae 2022, 8, 149. [Google Scholar] [CrossRef]
- Cruz, E.D.N.S.; Peixoto, L.D.S.; da Costa, J.S.; Mourão, R.H.V.; do Nascimento, W.M.O.; Maia, J.G.S.; Figueiredo, P.L.B. Seasonal variability of a caryophyllane chemotype essential oil of Eugenia patrisii Vahl occurring in the Brazilian Amazon. Molecules 2022, 27, 2417. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Padilla-González, G.F.; Phumthum, M. Fundamental chemistry of essential oils and volatile organic compounds, methods of analysis and authentication. Plants 2022, 11, 789. [Google Scholar] [CrossRef]
- Kant, R.; Kumar, A. Review on essential oil extraction from aromatic and medicinal plants: Techniques, performance and economic analysis. Sustain. Chem. Pharm. 2022, 30, 100829. [Google Scholar] [CrossRef]
- Barbosa, Q.P.; da Câmara, C.A.; Ramos, C.S.; Nascimento, D.C.; Lima-Filho, J.V.; Guimarães, E.F. Chemical composition, circadian rhythm and antibacterial activity of essential oils of Piper divaricatum: A new source of safrole. Quím. Nova 2012, 35, 1806–1808. [Google Scholar] [CrossRef]
- Raposo, J.D.A.; Figueiredo, P.L.B.; Santana, R.L.; da Silva Junior, A.Q.; Suemitsu, C.; da Silva, R.; Maia, J.G.S. Seasonal and circadian study of the essential oil of Myrcia sylvatica (G. Mey) DC., a valuable aromatic species occurring in the Lower Amazon River region. Biochem. Syst. Ecol. 2018, 79, 21–29. [Google Scholar] [CrossRef]
- Santos, E.L.; Lima, A.M.; Moura, V.F.D.S.; Setzer, W.N.; da Silva, J.K.R.; Maia, J.G.S.; Figueiredo, P.L.B. Seasonal and circadian rhythm of a 1, 8-cineole chemotype essential oil of Calycolpus goetheanus from marajó island, Brazilian amazon. Nat. Prod. Commun. 2020, 15, 1934578X20933055. [Google Scholar] [CrossRef]
- Yakir, E.; Hilman, D.; Harir, Y.; Green, R.M. Regulation of output from the plant circadian clock. FEBS J. 2007, 274, 335–345. [Google Scholar] [CrossRef]
- Hotta, C.T. From crops to shops: How agriculture can use circadian locks. J. Exp. Bot. 2021, 72, 7668–7679. [Google Scholar] [CrossRef]
- Steed, G.; Ramirez, D.C.; Hannah, M.A.; Webb, A.A. Chronoculture, harnessing the circadian clock to improve crop yield and sustainability. Science 2021, 372, eabc9141. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Z.; Hu, H.; Shi, S.; Yuan, X.; Yan, B.; Chen, L. An update on the function, biosynthesis and regulation of floral volatile terpenoids. Horticulturae 2021, 7, 451. [Google Scholar] [CrossRef]
- Felisberto, J.E.S.; Ramos, Y.J.E.; de Queiroz, G.A.; Guimarães, E.F.; Marques, A.E.M.; Moreira, D.L. Piper rivinoides Kunth: A medicinal plant that preserves bioactive chemical substances in its essential oil throughout the seasons. J. Med. Plant Res. 2022, 16, 258–268. [Google Scholar]
- Zeng, L.; Wang, X.; Kang, M.; Dong, F.; Yang, Z. Regulation of the rhythmic emission of plant volatiles by the circadian clock. Int. J. Mol. Sci. 2017, 18, 2408. [Google Scholar] [CrossRef]
- Moraes, M.M.; da Silva, T.M.; da Silva, R.R.; Ramos, C.S.; da Câmara, C.A. Circadian variation of essential oil from Piper marginatum Jacq. Bol. latinoam. Caribe Plantas Med. Aromát 2014, 13, 270–277. [Google Scholar]
- Bonacina, C.; da Cruz, R.M.S.; Nascimento, A.B.; Barbosa, L.N.; Gonçalves, J.E.; Gazim, Z.C.; de Souza, S.G.H. Salinity modulates growth, oxidative metabolism, and essential oil profile in Curcuma longa L. (Zingiberaceae) rhizomes. S. Afr. J. Bot. 2022, 146, 1–11. [Google Scholar] [CrossRef]
- Ormeno, E.; Mévy, J.P.; Vila, B.; Bousquet-Mélou, A.; Greff, S.; Bonin, G.; Fernandez, C. Water deficit stress induces different monoterpene and sesquiterpene emission changes in Mediterranean species. Relationship between terpene emissions and plant water potential. Chemosphere 2007, 67, 276–284. [Google Scholar] [CrossRef]
- Cohen, D.A.N. Maximizing final yield when growth is limited by time or by limiting resources. J. Theor. Biol. 1971, 33, 299–307. [Google Scholar] [CrossRef]
- Mehalaine, S.; Chenchouni, H. Quantifying how climatic factors influence essential oil yield in wild-growing plants. Arab. J. Geosci. 2021, 14, 1257. [Google Scholar] [CrossRef]
- Defossez, E.; Pitteloud, C.; Descombes, P.; Glauser, G.; Allard, P.M.; Walker, T.W.; Rasmann, S. Spatial and evolutionary predictability of phytochemical diversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2013344118. [Google Scholar] [CrossRef]
- Nielsen, J.A.; Whigham, P.A.; Frew, R.D.; Callaway, R.M.; Dickinson, K.J.M. Invasion essentials: Does secondary chemistry plasticity contribute to the invasiveness of Thymus vulgaris L.? Chemoecology 2014, 24, 15–27. [Google Scholar] [CrossRef]
- Nielsen, J.A.; Grøndahl, E.; Callaway, R.M.; Dickinson, K.J.; Ehlers, B.K. Home and away: Biogeographical comparison of species diversity in Thymus vulgaris communities. Biol. Invasions 2017, 19, 2533–2542. [Google Scholar] [CrossRef]
- Müller, C.; Junker, R.R. Chemical phenotype as important and dynamic niche dimension of plants. New Phytol. 2022, 234, 1168–1174. [Google Scholar] [CrossRef]
- Chen, X.; Huang, C.; Li, K.; Liu, J.; Zheng, Y.; Feng, Y.; Kai, G.Y. Recent advances in biosynthesis and pharmacology of β-elemene. Phytochem. Rev. 2022, 22, 1–18. [Google Scholar] [CrossRef]
- Baradaran, R.V.; Askari, V.R. A mechanistic review on immunomodulatory effects of selective type two cannabinoid receptor β-caryophyllene. Biofactors 2022, 48, 857–882. [Google Scholar] [CrossRef]
- Li, Y.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Y.; Li, Y.; Lu, Z.; Li, X. Herbivore and MeJA induced volatile emissions from the redroot pigweed Amaranthus retroflexus Linnaeus: Their roles in attracting Microplitis mediator (Haliday) parasitoids. Arthropod. Plant Interac. 2018, 12, 575–589. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- Cheng, A. Seasonal variation in secondary metabolites and protective capacity against UVB radiation in leaves of Eucommia ulmoides. Plant Physiol. Biochem. 2019, 135, 16–24. [Google Scholar]
- Teles, A.M.; dos Santos, B.A.; Ferreira, C.G.; Mouchreck, A.N.; da Silva Calabrese, K.; Abreu-Silva, A.L.; Almeida-Souza, F. BoD–Books on Demand; IntechOpen: London, UK, 2019; pp. 101–108. [Google Scholar]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Moayeri, H.R.S.; Moayeri, H.R.S.; Ashouri, A.; Poll, L.; Enkegaard, A. Olfactory response of a predatory mirid to herbivore induced plant volatiles: Multiple herbivory vs. single herbivory. J. Appl. Entomol. 2007, 131, 326–332. [Google Scholar] [CrossRef]
- Pinto-Zevallos, D.M.; Strapasson, P.; Zarbin, P.H.G. Herbivore-induced volatile organic compounds emitted by maize: Electrophysiological responses in Spodoptera frugiperda females. Phytochem. Lett. 2016, 16, 70–74. [Google Scholar] [CrossRef]
- Siripoltangman, N.; Chickos, J. Vapor pressure and vaporization enthalpy studies of the major components of ginger, α-zingiberene, β-sesquiphellandrene and (−) ar curcumene by correlation gas chromatography. J. Chem. Thermodyn. 2019, 138, 107–115. [Google Scholar] [CrossRef]
- Barton, D.; Chickos, J. The vapor pressure and vaporization enthalpy of (−) β-Elemene and (−) β-Bisabolene by correlation gas chromatography. J. Chem. Thermodyn. 2020, 148, 106139. [Google Scholar] [CrossRef]
- Munné-Bosch, S. Spatiotemporal limitations in plant biology research. Trends. Plant Sci. 2022, 27, 346–354. [Google Scholar] [CrossRef]
- Costa-Oliveira, C.; Ramos, Y.J.; Queiroz, G.A.; Guimarães, E.F.; Heggdorne-Araujo, M.; Lassounskaia, E.B.; Moreira, D.L. Antimycobacterial Activity and Chemical Characterization of the Essential Oils from Reproductive Organs of Piper lhotzkyanum Kunth (Piperaceae). Rev. Virtual Quim. 2021, 5, 1196–1202. [Google Scholar] [CrossRef]
- Agrawal, A.A. Current trends in the evolutionary ecology of plant defence. Funct. Ecol. 2011, 25, 420–432. [Google Scholar] [CrossRef]
- Heil, M.; Karban, R. Explaining evolution of plant communication by airborne signals. Trends Ecol. Evol. 2010, 25, 137–144. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Gottlieb, O.R.; Kaplan, M.A.C. Phytochemical evolution: The redox theory. Nat. Prod. J. 1993, 2, 171–176. [Google Scholar] [CrossRef]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef]
- Silva, J.K.; Da Trindade, R.; Alves, N.S.; Figueiredo, P.L.; Maia, J.G.S.; Setzer, W.N. Essential oils from neotropical Piper species and their biological activities. Int. J. Mol. Sci. 2017, 18, 2571. [Google Scholar] [CrossRef]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Setzer, W.N. Piper species: A comprehensive review on their phytochemistry, biological activities and applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compound | RIcal | RIlit | Percentual Relative Area (%) * | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2019 | 2020 | |||||||||
| Autumn | Winter | Spring | Summer | |||||||
| Mar | May | Jul | Ago | Oct | Nov | Jan | Feb | |||
| α-Thujene | 920 | 924 | tr | tr | ||||||
| α-Pinene | 929 | 932 | 6.2 | 6.8 | 1.6 | 1.0 | 0.4 | 1.2 | 7.0 | 3.4 |
| Camphene | 942 | 946 | tr | tr | ||||||
| Sabinene | 969 | 969 | tr | tr | ||||||
| β-Pinene | 969 | 974 | 1.8 | 7.9 | 1.9 | 1.1 | 1.1 | 1.3 | 4.5 | 2.1 |
| 6-Methyl-5-hepten-2-one | 981 | 981 | 0.3 | 0.1 | 0.1 | tr | ||||
| Myrcene | 987 | 988 | 0.3 | tr | 0.1 | 0.2 | tr | tr | ||
| α-Phellandrene | 1000 | 1002 | 0.1 | 0.2 | tr | 0.5 | tr | tr | ||
| α-Terpinene | 1010 | 1014 | 0.2 | 0.3 | 0.6 | 0.4 | tr | |||
| ρ-Cymene | 1018 | 1022 | tr | tr | tr | |||||
| Limonene | 1022 | 1024 | 1.2 | 0.3 | 1.6 | tr | ||||
| β-Phellandrene | 1023 | 1025 | 0.8 | |||||||
| 1,8-Cineole | 1027 | 1026 | tr | tr | ||||||
| Z-β-Ocimene | 1031 | 1032 | 1.1 | 0.6 | tr | 0.6 | 0.5 | 0.8 | tr | |
| E-β-Ocimene | 1042 | 1044 | 4.7 | 5.0 | 3.1 | 5.2 | 2.3 | 3.0 | 12.6 | 3.4 |
| γ-Terpinene | 1052 | 1054 | 0.4 | 0.6 | 0.5 | 0.3 | 0.3 | tr | tr | |
| Terpinolene | 1081 | 1086 | 0.2 | 0.1 | 0.2 | tr | ||||
| Linalool | 1090 | 1095 | 0.1 | 0.1 | 0.4 | tr | tr | |||
| E-4,8-Dimethyl-1,3,7-nonatriene | 1099 | - | 0.2 | 0.3 | 0.1 | tr | ||||
| Perillene | 1108 | 1102 | 0.2 | |||||||
| Allo-ocimene | 1130 | 1128 | 1.5 | 1.0 | 0.3 | 0.2 | tr | |||
| Terpinen-4-ol | 1175 | 1174 | tr | |||||||
| α-Terpineol | 1183 | 1186 | 0.1 | tr | ||||||
| Linalyl formate | 1210 | 1214 | 1.7 | |||||||
| NI MW194 | 1265 | - | tr | tr | 0.3 | tr | tr | |||
| Bicycloelemene # | 1312 | - | 4.1 | 1.3 | 3.3 | 1.5 | 4.5 | 4.9 | 1.2 | 5.8 |
| δ-Elemene | 1335 | 1335 | 2.7 | |||||||
| α-Cubebene | 1349 | 1348 | 0.1 | 0.1 | 0.3 | 2.5 | ||||
| α-Copaene | 1370 | 1374 | 0.6 | 0.4 | 0.3 | 0.5 | ||||
| β-Cubebene | 1384 | 1387 | tr | |||||||
| β-Elemene | 1391 | 1389 | 59.6 | 24.0 | 49.4 | 66.1 | 23.0 | 62.0 | 16.1 | 31.0 |
| Cyperene | 1396 | 1398 | 0.3 | tr | tr | |||||
| Italicene | 1404 | 1405 | tr | |||||||
| α-Gurjunene | 1407 | 1409 | tr | 0.6 | 0.2 | 0.8 | ||||
| E-Caryophyllene | 1413 | 1417 | 5.7 | 4.9 | 6.3 | 5.3 | 8.5 | 6.1 | 1.8 | 4.3 |
| E-α-Ionone | 1425 | 1428 | tr | |||||||
| γ-Elemene | 1430 | 1434 | 0.4 | 0.7 | 0.4 | 0.2 | 2.0 | 0.4 | 2.9 | 0.7 |
| NI MW204 | 1436 | - | tr | |||||||
| Aromadendrene | 1438 | 1439 | 0.1 | 0.3 | 0.3 | 0.2 | 1.0 | 0.3 | 1.4 | 1.6 |
| Z-β-Farnesene | 1440 | 1440 | 1.0 | 0.2 | 2.5 | 1.6 | ||||
| cis-Muurola-3,5-diene | 1446 | 1448 | 0.3 | |||||||
| Neryl propanoate | 1449 | 1452 | 0.6 | |||||||
| α-Humulene | 1450 | 1452 | 0.2 | 1.4 | 0.1 | 0.4 | 0.1 | 0.6 | 2.9 | |
| E -β-Farnesene | 1455 | 1454 | 0.1 | tr | 0.1 | 0.9 | ||||
| Allo-aromadendrene | 1457 | 1458 | 0.4 | 0.1 | 0.1 | |||||
| 4,5-di-epi-Aristolochene | 1474 | 1471 | tr | 0.8 | 0.1 | tr | 1.1 | |||
| β-Chamigrene | 1477 | 1476 | 0.8 | 0.5 | 0.6 | 0.8 | 0.9 | 0.9 | 4.8 | 3.5 |
| γ-Muurolene | 1479 | 1478 | 0.5 | tr | ||||||
| ar-Curcumene | 1481 | 1479 | 0.4 | 0.8 | 0.8 | |||||
| Germacrene D | 1483 | 1484 | 0.1 | 0.4 | 0.03± | 1.1 | ||||
| NI MW204 | 1487 | - | tr | tr | ||||||
| β-Selinene | 1489 | 1489 | 2.6 | 1.7 | 2.8 | 3.0 | 3.9 | 3.0 | 7.5 | 3.6 |
| α-Zingiberene | 1492 | 1493 | 6.9 | 0.1 | 16.9 | |||||
| Viridiflorene | 1495 | 1496 | 0.6 | tr | tr | 4.1 | ||||
| α-Selinene | 1497 | 1498 | 5.7 | |||||||
| Z-Dihydro-apofarnesal | 1499 | 1498 | 0.3 | |||||||
| Biclogermacrene | 1500 | 1500 | 5.1 | 6.1 | 5.3 | 1.5 | 5.5 | 4.1 | ||
| α-Muurolene | 1502 | 1500 | 0.1 | |||||||
| E,E-α-Farnesene | 1504 | 1505 | 1.3 | tr | ||||||
| β-Bisabolene | 1506 | 1505 | 0.3 | 0.4 | ||||||
| Germacrene A | 1510 | 1508 | 0.1 | 1.6 | tr | |||||
| γ-Cadinene | 1512 | 1513 | 0.1 | 1.7 | 0.2 | 0.1 | 0.4 | 0.1 | tr | tr |
| Cubebol | 1516 | 1514 | tr | 0.6 | ||||||
| epi-7-α-Selinene | 1519 | 1520 | 0.6 | |||||||
| Sesquiphelandrene | 1522 | 1521 | 0.7 | 3.2 | 3.0 | |||||
| δ-Cadinene | 1524 | 1522 | 0.2 | 1.6 | 0.8 | 0.5 | 2.1 | 0.6 | tr | 0.7 |
| NI MW204 | 1525 | - | tr | |||||||
| Zonarene | 1526 | 1528 | tr | 0.4 | ||||||
| NI MW204 | 1528 | - | tr | |||||||
| cis-Calamenene | 1530 | 1528 | 0.2 | tr | tr | |||||
| NI MW204 | 1531 | - | 0.3 | tr | ||||||
| trans-Cadine-1,4-diene | 1534 | 1533 | tr | |||||||
| α-cadinene | 1536 | 1537 | 0.5 | tr | ||||||
| Selina-3,7(11)-diene | 1542 | 1545 | 0.1 | 0.6 | 0.1 | tr | 0.8 | |||
| Hedycariol | 1547 | 1546 | 0.4 | |||||||
| Germacrene B | 1553 | 1559 | 0.7 | 0.8 | 0.2 | tr | 0.2 | |||
| E-Nerolidol | 1560 | 1561 | 0.1 | 1.1 | 0.7 | 0.6 | tr | 0.1 | ||
| 3-Hexenyl-Z -benzoate | 1566 | 1565 | 0.3 | |||||||
| Spathulenol | 1575 | 1577 | 0.2 | 0.2 | 0.7 | 0.1 | tr | 0.7 | ||
| Caryophyllene oxide | 1580 | 1582 | 2.5 | 0.2 | 1.2 | 0.3 | tr | 0.6 | ||
| NI MW204 | 1581 | - | tr | |||||||
| NI MW220 | 1588 | - | tr | tr | tr | |||||
| Globulol | 1591 | 1590 | 1.2 | 0.1 | 0.4 | 0.4 | 0.2 | tr | 0.1 | |
| Viridiflorol | 1594 | 1592 | 0.1 | 0.1 | 0.2 | 0.1 | tr | |||
| Longiborneol | 1597 | 1599 | tr | tr | tr | 0.2 | 0.4 | |||
| Guayol | 1600 | 1600 | 1.3 | |||||||
| Rosifoliol | 1605 | 1600 | tr | |||||||
| Humulene II epoxide | 1607 | 1608 | tr | tr | ||||||
| NI MW220 | 1612 | - | 0.6 | tr | ||||||
| NI MW220 | 1617 | - | 0.1 | |||||||
| 10-epi-γ-Eudesmol | 1620 | 1622 | tr | |||||||
| Isolongifolanone | 1623 | 1625 | 0.2 | |||||||
| 1-epi-α-Cubenol | 1625 | 1627 | 0.1 | 0.1 | 0.8 | 0.4 | 0.3 | |||
| γ-Eudesmol | 1629 | 1630 | tr | |||||||
| NI MW220 | 1633 | - | 1.0 | |||||||
| α-Acorenol | 1635 | 1632 | 0.1 | |||||||
| Selina-1,3,7(11)-trien-8-one | 1637 | 1632 | tr | |||||||
| epi-α-Cadinol | 1637 | 1638 | 0.1 | 0.1 | 1.2 | 0.1 | ||||
| Caryophyll-4(12),8(13)-dien-5-ol | 1636 | 1639 | tr | 0.1 | ||||||
| Hinesol | 1638 | 1640 | 1.2 | |||||||
| epi-α-Muurolol | 1639 | 1640 | tr | 0.2 | 2.2 | 0.1 | ||||
| Selina-3,11-dien-6α-ol | 1641 | 1642 | 0.1 | 1.0 | 0.8 | |||||
| α-Muurolol | 1643 | 1644 | 1.9 | 2.3 | 0.6 | |||||
| NI MW220 | 1645 | - | 0.1 | |||||||
| Cubenol | 1646 | 1645 | tr | |||||||
| NI MW222 | 1647 | - | tr | |||||||
| NI MW218 | 1648 | - | tr | |||||||
| NI MW220 | 1649 | - | 1.9 | |||||||
| NI MW222 | 1650 | - | tr | 0.1 | tr | 0.1 | ||||
| Pogostol | 1652 | 1651 | 0.1 | |||||||
| Selin-11-en-4-α-ol | 1656 | 1658 | 2.2 | 1.2 | 2.6 | 0.4 | 2.3 | 1.7 | 0.1 | |
| Neo-Intermedeol | 1659 | 1658 | tr | tr | ||||||
| Intermedeol | 1662 | 1665 | 1.4 | tr | tr | tr | 1.0 | |||
| NI MW220 | 1687 | - | tr | |||||||
| NI MW220 | 1688 | - | tr | |||||||
| NI MW220 | 1690 | - | tr | |||||||
| Eudesm-7(11)-en-4-ol | 1698 | 1700 | 1.3 | tr | 0.1 | 0.1 | ||||
| NI MW220 | 1725 | - | 0.1 | |||||||
| 7,14-anyhdro-Amorpha-4,9-diene | 1750 | 1755 | 0.1 | 0.2 | ||||||
| Phytol | 1941 | 1942 | 0.1 | |||||||
| E -Phytyl acetate | 2222 | 2218 | 0.3 | |||||||
| Tricosane | 2297 | 2300 | 0.1 | |||||||
| Non-oxygenated monoterpenes | 11.6 | 22.8 | 10.0 | 8.5 | 4.9 | 6.7 | 22.5 | 9.0 | ||
| Oxygenated monoterpenes | 0.1 | 1.7 | 0.1 | 0.0 | 0.4 | 0.0 | 1.3 | 0.0 | ||
| Non-oxygenated sesquiterpenes | 81.3 | 46.7 | 75.9 | 82.8 | 72.9 | 85.1 | 57.3 | 72.3 | ||
| Oxygenated sesquiterpenes | 3.0 | 13.7 | 6.5 | 4.9 | 8.3 | 4.0 | 0.6 | 4.2 | ||
| Other chemical classes | 0.1 | 0.1 | 0.1 | 0.1 | 0.3 | 0.1 | 0.2 | 0.2 | ||
| Total identified (%) | 96.5 | 84.8 | 92.7 | 96.5 | 86.9 | 95.8 | 81.7 | 85.6 | ||
| Essential oil yeld (%) (w/w) | 0.14 | 0.10 | 0.12 | 0.14 | 0.28 | 0.10 | 0.15 | 0.22 | ||
| H′ | 2.0 | 2.9 | 2.1 | 1.5 | 3.2 | 1.6 | 2.7 | 2.6 | ||
| SRO | −150.4 | −136.4 | −146.5 | −156.0 | −138.6 | −153.1 | −129.3 | −135.6 | ||
| R&M | −3.8 | −3.4 | −2.3 | −3.6 | −3.5 | −4.1 | −2.4 | −2.2 | ||
| Compounds | RIcalc | RIlit | Percentual Relative Area (%) * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| July 2019 (Dry Season) | January 2020 (Rainy Season) | |||||||||||||||||
| 12 pm | 03 am | 06 am | 09 am | 12 am | 03 pm | 06 pm | 09 pm | 12 pm | 03 am | 06 am | 09 am | 12 am | 03 pm | 06 pm | 09 pm | |||
| α-Thujene | 920 | 924 | 0.3 | |||||||||||||||
| α-Pinene | 929 | 932 | 0.7 | 0.8 | 0.5 | 3.4 | 1.3 | 0.5 | 1.3 | 3.1 | 1.3 | 1.5 | ||||||
| β-Pinene | 970 | 974 | 1.2 | 1.2 | 1.0 | 0.5 | 2.7 | 4.8 | 1.3 | 1.2 | 2.3 | 1.0 | 1.1 | |||||
| 6-Methyl-5-hepten-2-one | 978 | 981 | 3.9 | |||||||||||||||
| Myrcene | 987 | 988 | 0.4 | tr | ||||||||||||||
| α-Phellandrene | 1000 | 1002 | 0.6 | 0.4 | 1.7 | 1.8 | tr | |||||||||||
| α-Terpinene | 1010 | 1014 | 1.7 | |||||||||||||||
| Limonene | 1022 | 1024 | 0.8 | 0.6 | 1.3 | 0.9 | tr | tr | 3.7 | 1.2 | ||||||||
| Z-β-Ocimene | 1031 | 1032 | 0.3 | 0.4 | 0.2 | tr | 0.2 | 0.5 | 3.3 | tr | 2.2 | 3.3 | ||||||
| E-β-Ocimene | 1042 | 1044 | 3.7 | 0.3 | 0.4 | 0.4 | 0.5 | |||||||||||
| γ-Terpinene | 1052 | 1054 | 0.4 | 0.7 | ||||||||||||||
| Terpinolene | 1081 | 1086 | 1.3 | |||||||||||||||
| Linalool | 1090 | 1095 | 0.1 | 0.1 | 0.1 | tr | ||||||||||||
| trans-Sabinene hydrate | 1098 | 1098 | tr | |||||||||||||||
| Allo-Ocimene | 1130 | 1128 | 0.2 | 0.2 | 0.1 | tr | tr | |||||||||||
| 2-(1Z)-Propenyl phenol | 1145 | 1146 | 0.2 | 0.1 | 0.1 | 1.1 | ||||||||||||
| NI MW194 | 1265 | 0.1 | 0.2 | 0.4 | 0.5 | 0.3 | ||||||||||||
| Bicycloelemene # | 1312 | 2.5 | 1.1 | 0.7 | 1.2 | 0.6 | 1.1 | 0.9 | 1.3 | 3.0 | 3.2 | 6.7 | 3.0 | 5.8 | 4.0 | 3.9 | 3.3 | |
| δ-Elemene | 1335 | 1335 | 0.3 | tr | 3.0 | tr | tr | 3.5 | ||||||||||
| α-Cubebene | 1349 | 1348 | 0.5 | 0.4 | 0.2 | 0.7 | 0.1 | 0.2 | tr | 0.4 | tr | tr | tr | tr | ||||
| α-Copaene | 1370 | 1374 | 0.7 | 0.8 | 0.5 | 0.8 | 0.6 | 0.7 | 0.5 | 0.7 | tr | tr | 1.4 | |||||
| β-Elemene | 1391 | 1389 | 7.7 | 11.6 | 9.8 | 8.4 | 9.7 | 8.0 | 9.2 | 11.7 | 19.5 | 22.1 | 28.4 | 29.2 | 29.2 | 19.7 | 20.2 | 35.1 |
| Cyperene | 1396 | 1398 | 0.9 | 0.4 | 0.9 | 0.5 | 0.4 | 0.5 | tr | |||||||||
| α-Gurjunene | 1407 | 1409 | 0.2 | 0.1 | 4.3 | 1.1 | 1.1 | 1.0 | ||||||||||
| E-Caryophyllene | 1413 | 1417 | 9.8 | 10.8 | 9.0 | 8.1 | 9.4 | 7.4 | 7.2 | 5.9 | 4.8 | 5.6 | 5.2 | 4.8 | 5.8 | 6.0 | 7.9 | 4.5 |
| γ-Elemene | 1430 | 1434 | 3.2 | 1.7 | 1.2 | 1.1 | 2.5 | 0.9 | 3.2 | 1.7 | 1.7 | 2.6 | 3.2 | 2.8 | ||||
| α-Guaiene | 1436 | 1437 | 0.4 | 0.5 | 0.2 | 0.3 | 0.3 | 0.3 | tr | tr | ||||||||
| Aromadendrene | 1438 | 1439 | 0.8 | 0.7 | 0.9 | 0.6 | 0.9 | 0.8 | 0.7 | 1.5 | 3.9 | 4.7 | tr | 3.8 | 1.6 | 1.2 | 2.5 | |
| 6,9-Guaiadiene | 1441 | 1442 | 0.2 | 0.2 | tr | |||||||||||||
| α-Humulene | 1450 | 1452 | 0.9 | 0.8 | ||||||||||||||
| E-β-Farnesene | 1543 | 1454 | 3.3 | 4.0 | 3.4 | 2.7 | 1.4 | 1.7 | 3.3 | 3.1 | 3.3 | 2.0 | 2.1 | 3.7 | 2.7 | 4.4 | 2.3 | 2.3 |
| Rotundene | 1456 | 1457 | 1.0 | 0.8 | 0.8 | 0.7 | 0.6 | 0.5 | 0.5 | tr | 1.0 | |||||||
| Allo-Aromadendrene | 1459 | 1458 | 0.4 | 0.3 | 0.9 | |||||||||||||
| NI MW220 | 1463 | - | 1.0 | |||||||||||||||
| γ-Gurjunene | 1474 | 1475 | 1.3 | 1.3 | 3.1 | |||||||||||||
| β-Chamigrene | 1477 | 1476 | 3.6 | 3.9 | 2.1 | 4.5 | 2.5 | 2.3 | 1.6 | |||||||||
| γ-Muurolene | 1479 | 1478 | 1.9 | |||||||||||||||
| ar-Curcumene | 1481 | 1479 | 1.4 | 1.2 | 1.7 | 1.3 | tr | 5.0 | 2.9 | 4.2 | 1.3 | 4.4 | 2.7 | tr | ||||
| Germacrene D | 1483 | 1440 | tr | 4.5 | 2.6 | 3.0 | 3.5 | 2.9 | 4.0 | |||||||||
| γ-Curcumene | 1485 | - | 2.0 | 1.1 | 1.1 | 1.1 | 2.1 | |||||||||||
| β-Selinene | 1489 | 1489 | 3.4 | 4.9 | 3.2 | 6.5 | 5.7 | 3.4 | 4.4 | 2.4 | 3.5 | 4.0 | 2.9 | 5.3 | 4.8 | 4.4 | 4.3 | 5.5 |
| α-Zingiberene | 1492 | 1493 | 18.9 | 26.1 | 21.0 | 18.8 | 23.0 | 20.6 | 21.1 | 20.6 | 17.9 | 2.8 | 4.3 | 15.1 | 7.7 | 10.4 | 11.0 | 9.5 |
| Viridiflorene | 1495 | 1496 | 1.0 | 1.9 | tr | |||||||||||||
| α-Selinene | 1497 | 1498 | 4.0 | |||||||||||||||
| α-Muurolene | 1502 | 1500 | 0.5 | |||||||||||||||
| β-Bisabolene | 1506 | 1505 | 2.3 | 3.2 | 2.2 | 2.2 | 2.0 | 1.9 | 2.3 | 2.0 | tr | 3.2 | 2.3 | 4.4 | 7.9 | 2.8 | 2.3 | 5.4 |
| α-Bulnesene | 1508 | 1509 | 0.5 | |||||||||||||||
| δ-Amorphene | 1510 | 1511 | 1.3 | 1.0 | 1.0 | 3.3 | 3.4 | 4.5 | 2.0 | 1.0 | ||||||||
| γ-Cadinene | 1512 | 1513 | 3.2 | 4.6 | 3.4 | 6.1 | 6.5 | 4.1 | 1.1 | 3.2 | 2.5 | 4.6 | 4.1 | 0.6 | 1.3 | tr | ||
| Cubebol | 1516 | 1514 | 0.9 | |||||||||||||||
| NI MW222 | 1518 | 1.5 | 0.7 | tr | ||||||||||||||
| β-Sesquiphellandrene | 1522 | 1521 | 6.6 | 8.0 | 4.5 | 2.2 | 3.5 | 3.8 | 4.0 | 5.3 | 6.3 | 8.0 | 6.0 | 1.0 | 1.0 | 0.8 | 3.4 | 4.5 |
| δ-Cadinene | 1524 | 1522 | 1.0 | 0.9 | tr | 8.1 | 3.1 | 0.8 | 4.0 | 2.2 | ||||||||
| Zonarene | 1527 | 1528 | 0.5 | tr | 0.3 | 0.4 | 0.7 | 0.4 | 0.5 | tr | tr | tr | ||||||
| NI MW220 | 1532 | - | 0.8 | 0.6 | 0.7 | 0.7 | 0.7 | 0.7 | ||||||||||
| α-Cadinene | 1536 | 1537 | tr | 0.7 | 1.2 | 0.7 | 0.7 | 1.0 | ||||||||||
| Selina-3,7(11)-diene | 1542 | 1545 | 0.9 | 0.3 | 0.6 | 1.1 | 0.9 | 0.7 | 0.7 | 0.4 | 0.3 | 1.3 | 1.0 | 0.7 | 0.7 | 0.5 | ||
| β-Vetivenene | 1552 | 1554 | 0.2 | |||||||||||||||
| Germacrene B | 1558 | 1559 | 5.1 | 0.4 | 5.1 | 0.7 | 0.8 | 1.1 | 0.3 | 0.9 | 1.0 | |||||||
| NI MW222 | 1559 | - | 0.3 | 0.3 | ||||||||||||||
| E-Nerolidol | 1560 | 1561 | 1.1 | 0.7 | 0.7 | 0.8 | 0.9 | 0.9 | 0.9 | 0.2 | 0.5 | 1.1 | 0.4 | 0.3 | 0.3 | 0.5 | ||
| Maaliol | 1565 | 1566 | 0.4 | |||||||||||||||
| NI MW220 | 1570 | - | 0.2 | 0.2 | 0.2 | 0.2 | 0.3 | |||||||||||
| Spathulenol | 1578 | 1577 | 2.7 | 0.2 | 1.1 | 2.9 | 1.2 | 1.1 | 1.3 | 1.4 | 0.3 | 0.2 | 0.3 | 0.5 | tr | 0.5 | 0.6 | |
| Caryophyllene oxide | 1580 | 1582 | 1.1 | 1.3 | 2.9 | 1.5 | 1.7 | 1.9 | 3.5 | 3.9 | 0.4 | 1.3 | 1.5 | 2.3 | 1.8 | 1.3 | 1.2 | 1.9 |
| Globulol | 1591 | 1590 | 1.0 | 0.5 | 1.3 | 0.9 | 0.9 | 0.9 | 1.1 | 0.9 | 1.5 | 0.5 | 0.4 | 0.6 | 0.4 | 0.3 | 0.4 | 0.3 |
| Viridiflorol | 1594 | 1592 | 0.5 | 0.3 | 0.2 | 0.2 | 0.2 | 0.3 | ||||||||||
| NI MW222 | 1596 | - | 0.2 | 0.2 | 0.2 | 0.2 | 0.1 | |||||||||||
| Longiborneol | 1597 | 1599 | 0.2 | 0.2 | 0.2 | 0.1 | ||||||||||||
| Guaiol | 1600 | 1600 | tr | 0.1 | ||||||||||||||
| Rosifoliol | 1601 | 1600 | 0.6 | 0.6 | 0.7 | 0.7 | 0.6 | 0.5 | ||||||||||
| Ledol | 1603 | 1602 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.2 | 0.5 | 0.2 | ||||||||
| 5-epi-7-epi-α-Eudesmol | 1606 | 1607 | 0.2 | 0.3 | 0.4 | |||||||||||||
| Humulene epoxide II | 1609 | 1608 | 0.4 | 0.4 | 0.2 | 0.3 | 0.4 | 0.5 | 0.2 | 0.2 | ||||||||
| epi-Cedrol | 1612 | 1618 | 0.7 | 0.7 | 1.4 | 1.6 | 1.7 | 1.4 | 0.1 | 0.1 | ||||||||
| 1,10-di-epi-Cubenol | 1619 | 1618 | 0.4 | 0.8 | 0.4 | 0.3 | 0.4 | 0.5 | 0.2 | |||||||||
| NI MW222 | 1620 | - | 0.2 | |||||||||||||||
| 10-epi-γ-Eudesmol | 1621 | 1622 | 4.3 | 4.8 | 4.3 | 1.2 | 3.6 | 6.0 | 4.7 | 0.4 | ||||||||
| 1-epi-Cubenol | 1625 | 1627 | 1.6 | 1.1 | 1.6 | 1.2 | 1.6 | 1.9 | 1.4 | 0.7 | 0.7 | 0.5 | 1.1 | 0.1 | ||||
| NI MW220 | 1628 | - | 0.5 | |||||||||||||||
| epi-α-Cadinol | 1633 | 1638 | 0.8 | 0.7 | 1.0 | 1.0 | 0.9 | 0.9 | tr | 0.7 | 0.3 | 0.7 | 0.6 | 0.3 | ||||
| Allo-Aromadendrene epoxide | 1635 | 1639 | 0.4 | 0.3 | 0.4 | 0.4 | 0.5 | 0.3 | 0.2 | |||||||||
| Caryophylla-4(12),8(13)-dien-5β-ol | 1637 | 1639 | 0.6 | 0.6 | 0.7 | 0.6 | ||||||||||||
| Allo-Aromadendrene epoxide | 1638 | 1639 | tr | 0.1 | ||||||||||||||
| epi-α-Muurolol | 1641 | 1640 | 0.1 | 0.1 | 0.1 | 0.1 | 0.2 | tr | ||||||||||
| α- Muurolol | 1642 | 1644 | 0.3 | 0.3 | 0.4 | 0.2 | 0.6 | 0.3 | tr | 0.7 | 1.0 | 0.2 | ||||||
| NI MW222 | 1644 | 1.1 | 1.0 | 1.4 | 1.1 | |||||||||||||
| Cubenol | 1645 | 1645 | 4.6 | 4.4 | 5.7 | 3.5 | 3.9 | 3.0 | 3.1 | |||||||||
| NI MW220 | 1646 | 0.4 | ||||||||||||||||
| Agarospirol | 1647 | 1646 | 0.1 | 0.2 | 0.2 | 0.3 | ||||||||||||
| Pogostol | 1652 | 1651 | 0.1 | 0.2 | 0.5 | 0.1 | ||||||||||||
| α-Cadinol | 1653 | 1652 | 0.1 | |||||||||||||||
| Selin-11-en-4-α-ol | 1656 | 1658 | 0.2 | tr | 0.2 | 0.2 | 0.2 | 0.1 | 0.2 | tr | 0.3 | |||||||
| E-Bisabol-11-ol | 1666 | 1667 | tr | 0.1 | tr | |||||||||||||
| 14-Hydroxy-9-epi-E-caryophyllene | 1669 | 1668 | 1.8 | 1.1 | 1.3 | 1.2 | 1.2 | 1.4 | 1.0 | 0.2 | 0.1 | 0.1 | ||||||
| NI MW220 | 1672 | 0.4 | 0.5 | 0.3 | 0.4 | 0.2 | 0.1 | |||||||||||
| α-Bisabolol | 1684 | 1685 | 0.4 | 0.2 | 0.3 | tr | tr | |||||||||||
| Germacra-4(15),5,10(14)-trien-1-α-ol | 1686 | 1685 | 0.2 | 0.2 | 0.2 | tr | ||||||||||||
| NI MW220 | 1687 | 0.4 | ||||||||||||||||
| NI MW220 | 1688 | 0.4 | 0.2 | 0.3 | 0.4 | |||||||||||||
| Shyobunol | 1689 | 1688 | 0.2 | tr | 0.2 | tr | tr | tr | ||||||||||
| 2Z,6Z-Farnesol | 1699 | 1698 | 0.3 | 0.4 | 0.3 | 0.5 | ||||||||||||
| Eudesm-7(11)-en-4-ol | 1700 | 1700 | 0.6 | 0.6 | 0.6 | 0.6 | 0.7 | 0.6 | 0.1 | |||||||||
| Amorpha-4,9-dien-2-ol | 1702 | 1700 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | tr | tr | ||||||||
| Nootkatol | 1715 | 1714 | 0.3 | 0.2 | 0.2 | 0.3 | 0.3 | tr | ||||||||||
| 2E,6E-Farnesal | 1742 | 1740 | 0.1 | 0.1 | 0.1 | |||||||||||||
| γ-Costol | 1746 | 1745 | 0.4 | 0.3 | 0.3 | 0.2 | 0.3 | 0.3 | tr | |||||||||
| 6S,7R-Bisabolone | 1750 | 1749 | 0.1 | 0.1 | tr | |||||||||||||
| Xanthorrhizol | 1752 | 1751 | 0.1 | 0.2 | 0.2 | 0.1 | 0.2 | |||||||||||
| α-Costol | 1774 | 1773 | 0.4 | 0.3 | 0.3 | 0.3 | 0.2 | |||||||||||
| 2-α-Hydroxy-amorpha-4,7(11)-diene | 1776 | 1775 | 0.1 | 0.2 | 0.3 | |||||||||||||
| Benzyl benzoate | 1760 | 1759 | 0.1 | 0.1 | 0.3 | 0.1 | 0.1 | |||||||||||
| β-Vetivone | 1823 | 1822 | 0.2 | 0.2 | ||||||||||||||
| Phytol | 1946 | 1942 | 0.5 | 0.7 | tr | |||||||||||||
| Non-oxygenated monoterpenes | 1.9 | 0.0 | 1.2 | 1.8 | 0.0 | 0.0 | 0.0 | 1.0 | 17.8 | 6.1 | 1.7 | 0.0 | 2.5 | 5.3 | 2.3 | 2.5 | ||
| Oxygenated monoterpenes | 0.3 | 0.0 | 0.0 | 0.2 | 0.0 | 0.1 | 0.0 | 1.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | ||
| Non-oxygenated sesquiterpenes | 62.2 | 77.3 | 65.1 | 60.9 | 65.8 | 53.6 | 62.2 | 60.5 | 79.5 | 87.3 | 82.7 | 92.0 | 86.9 | 78.7 | 75.5 | 82.4 | ||
| Oxygenated sesquiterpenes | 25.0 | 3.6 | 24.6 | 28.5 | 13.2 | 22.2 | 28.9 | 27.4 | 6.9 | 3.6 | 3.1 | 5.1 | 5.4 | 5.5 | 6.1 | 4.0 | ||
| Total identified (%) | 98.8 | 88.8 | 97.0 | 98.8 | 82.4 | 80.0 | 97.1 | 98.9 | 98.4 | 97.0 | 91.2 | 98.5 | 98.5 | 95.7 | 86.2 | 98.5 | ||
| Essential oil yeld (%) (w/w) | 0.22 | 0.20 | 0.08 | 0.14 | 0.11 | 0.03 | 0.11 | 0.15 | 0.20 | 0.20 | 0.06 | 0.12 | 0.01 | 0.19 | 0.06 | 0.19 | ||
| H’ | 2.54 | 2.93 | 2.97 | 2.65 | 2.59 | 2.74 | 3.10 | 2.94 | 2.40 | 3.25 | 2.40 | 3.21 | 3.29 | 2.57 | 2.92 | 3.24 | ||
| SRO | −156.69 | −157.35 | −141.78 | −154.32 | −157.44 | −131.60 | −127.32 | −151.88 | −157.35 | −156.47 | −154.43 | −144.80 | −156.03 | −157.18 | −150.76 | −137.37 | ||
| R&M | −2.49 | −2.71 | −5.67 | −2.66 | −2.46 | −4.39 | −2.83 | −2.62 | −4.25 | −2.24 | −5.33 | −4.99 | −4.22 | −4.03 | −2.43 | −3.71 | ||
| Analyzed Variable | Descriptive Statistics | Spearman Coefficient | Rho | ||||||
|---|---|---|---|---|---|---|---|---|---|
| μ | SD | RSD | T(°C) | RH(%) | R(KJ.m−2) | P(mm) | H′ | ||
| H′—S | 2.32 | 0.52 | 22.62 | 0.24 | 0.02 | 0.24 | -0.14 | - | - |
| H′—R | 2.91 | 0.34 | 11.66 | 0.47 | −0.29 | 0.10 | - | - | - |
| H′—D | 2.81 | 0.18 | 6.30 | −0.18 | 0.14 | 0.20 | - | - | - |
| SRO—S | 147.30 | 10.30 | 6.99 | 0.29 | 0.12 | 0.48 | −0.05 | 0.71 * | −14.75 |
| SRO—R | 151.80 | 5.61 | 3.70 | −0.01 | −0.05 | 0.20 | - | 0.52 * | 0.88 |
| SRO—D | 147.30 | 0.90 | 0.61 | 0.69 * | 0.29 | 0.46 | - | −0.52 * | −0.66 |
| R&M—S | −3.16 | 0.66 | −20.97 | 0.02 | −0.17 | 0.67 * | −0.45 | 0.45 | −1.65 |
| R&M—R | −3.90 | 0.83 | −21.25 | −0.43 | −0.05 | −0.17 | - | 0.48 | 0.20 |
| R&M—D | −3.23 | 0.90 | −27.89 | 0.23 | 0.26 | 0.32 | - | −0.69 * | −0.22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Oliveira, C.d.; Gouvêa-Silva, J.G.; Brito Machado, D.d.; Felisberto, J.R.S.; Queiroz, G.A.d.; Guimarães, E.F.; Ramos, Y.J.; Moreira, D.d.L. Chemical Diversity and Redox Values Change as a Function of Temporal Variations of the Essential Oil of a Tropical Forest Shrub. Diversity 2023, 15, 715. https://doi.org/10.3390/d15060715
Costa-Oliveira Cd, Gouvêa-Silva JG, Brito Machado Dd, Felisberto JRS, Queiroz GAd, Guimarães EF, Ramos YJ, Moreira DdL. Chemical Diversity and Redox Values Change as a Function of Temporal Variations of the Essential Oil of a Tropical Forest Shrub. Diversity. 2023; 15(6):715. https://doi.org/10.3390/d15060715
Chicago/Turabian StyleCosta-Oliveira, Claudete da, João Gabriel Gouvêa-Silva, Daniel de Brito Machado, Jéssica Regina Sales Felisberto, George Azevedo de Queiroz, Elsie Franklin Guimarães, Ygor Jessé Ramos, and Davyson de Lima Moreira. 2023. "Chemical Diversity and Redox Values Change as a Function of Temporal Variations of the Essential Oil of a Tropical Forest Shrub" Diversity 15, no. 6: 715. https://doi.org/10.3390/d15060715