
Back to Linnaeus: Proper Botanical Naming of the Tetraploid Indian Acorus (Acoraceae), an Important Medicinal Plant

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Leaf Anatomy
- (1)
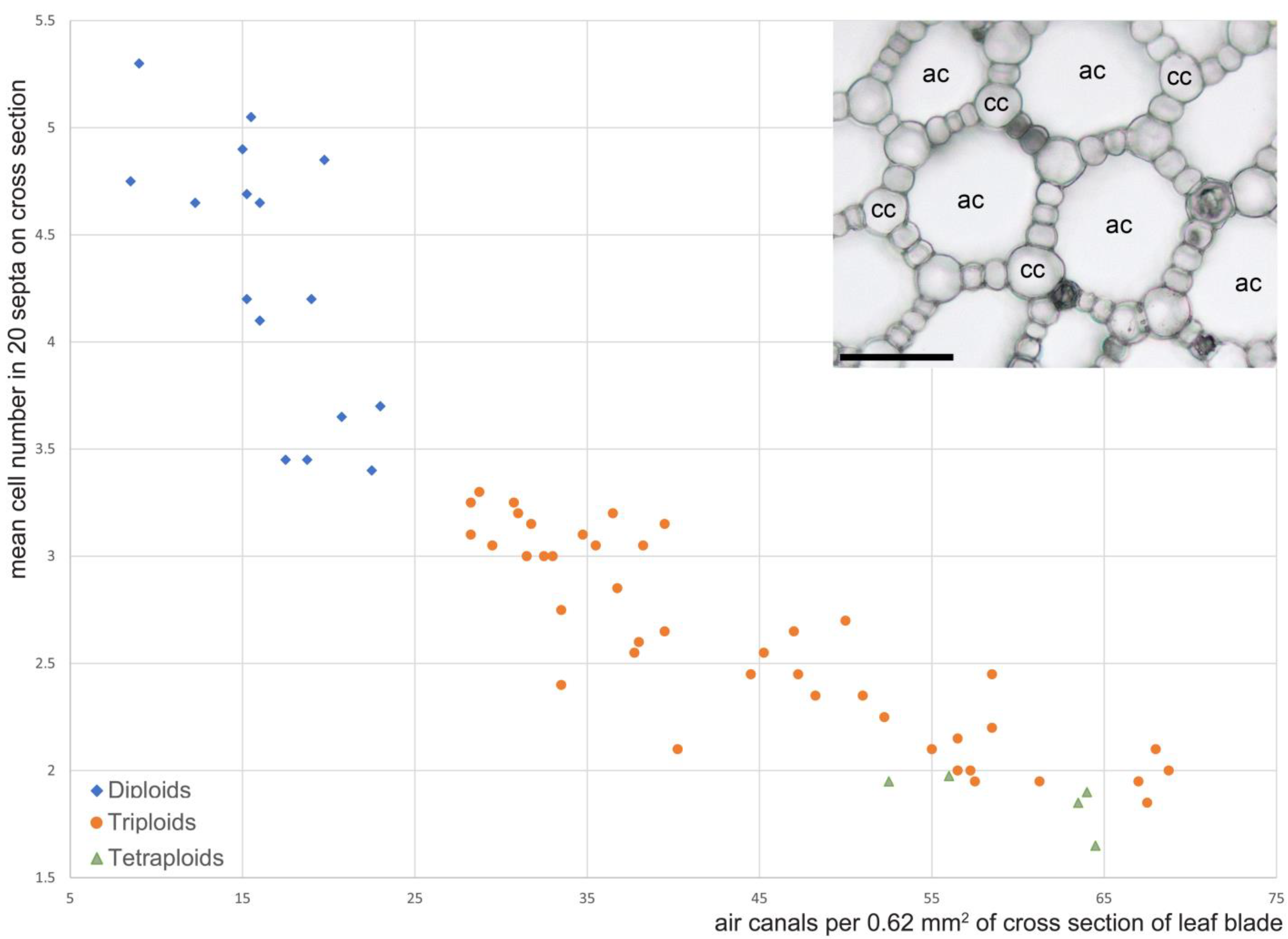
- The number of air canals per 0.62 mm2 of cross-section. We counted the number of the canals in each leaf in two circular fields of view situated in each leaf side within the secondary midrib as close as possible to the leaf margin, but avoiding the presence of chlorenchyma and peripheral vascular bundles. The number of canals per area was calculated as the total of completely visible canals plus ½ of the total of incompletely visible canals. The mean of the canal numbers in the two fields was calculated and used as a metric.
- (2)
- Mean number of cells in the septa between the air canals visible in cross-section of the leaf blade. Cells belonging to a septum were defined as those that are exposed into two adjacent canals. Cells situated at junctions of three or more septa and therefore exposed into more than two air canals (corner cells) were not counted. For each examined leaf, cell number was counted in 20 randomly selected septa in the fields of view described above, and then mean cell number per septum was calculated.
2.3. Chromosome Numbers
2.4. Nomenclatural Analysis
3. Results and Discussion
3.1. Nomenclatural Status and Taxonomic Identity of Acorus calamus var. angustatus Engl.
3.2. Taxonomic Identity of Acorus calamus var. verus L.
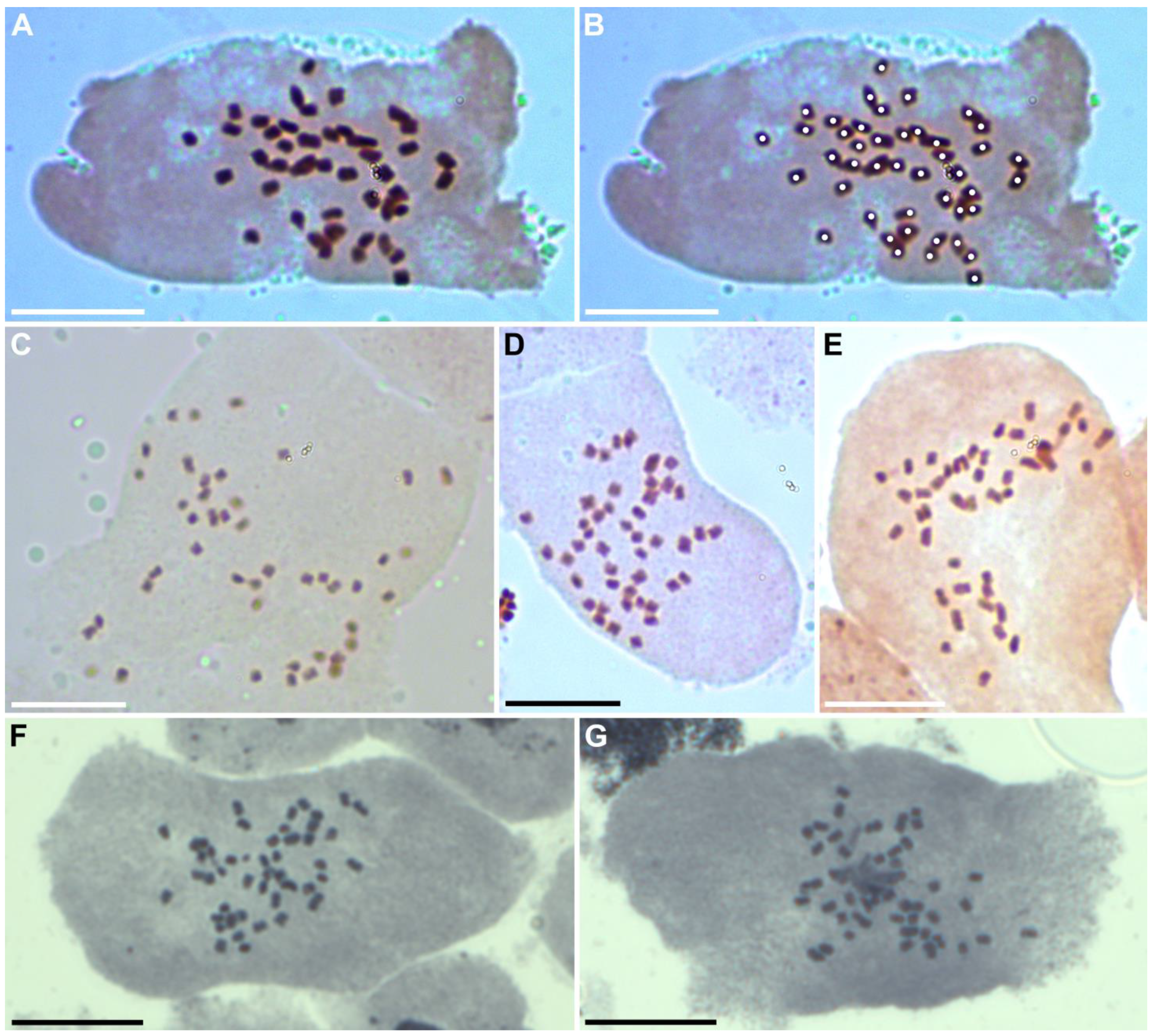
3.3. Chromosome Number of the Epitype Accession of Acorus calamus var. verus: A Possibility of Aneuploidy and Mixoploidy
3.4. Leaf Anatomy and Ploidy Level of the Epitype Accession
3.5. Flowering and Growth Pattern of the Tetraploid Indian Acorus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Röst, L.C.M. Biosystematic investigations with Acorus—4. Communication. A synthetic approach to the classification of the genus. Planta Med. 1979, 37, 289–307. [Google Scholar] [CrossRef]
- Li, H.; Zhu, G.; Bogner, J. Acoraceae. In Flora of China; Wu, Z., Raven, P., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2010; Volume 2, pp. 1–2. [Google Scholar]
- Bogner, J. Acoraceae. In Flora Malesiana, Ser. 1; Noteboom, H.P., Ed.; Nationaal Herbarium Nederland: Leiden, The Netherlands, 2011; Volume 20, pp. 1–13. [Google Scholar]
- Boyce, P. Acoraceae. In Flora of Thailand; Santisuk, T., Larsen, K., Eds.; The Forest Herbarium, Department of National Parks, Wildlife and Plant Conservation: Bangkok, Thailand, 2012; Volume 11, pp. 322–325. [Google Scholar]
- Vinogradova, Y.K. Variability of Acorus calamus L. biological characteristics in natural and invasive populations. Byull. Glavn. Bot. Sada 2004, 187, 23–31. (In Russian) [Google Scholar]
- Sokoloff, D.D.; Skaptsov, M.V.; Vislobokov, N.A.; Smirnov, S.V.; Shmakov, A.I.; Remizowa, M.V. Morphological characterization of diploid and triploid Acorus calamus (Acoraceae) from southern Western Siberia, parthenocarpy in sterile plants and occurrence of aneuploidy. Bot. J. Linn. Soc. 2021, 195, 189–215. [Google Scholar] [CrossRef]
- Packer, J.G.; Ringius, G.S. The distribution and status of Acorus (Araceae) in Canada. Can. J. Bot. 1984, 62, 2248–2252. [Google Scholar] [CrossRef]
- Thompson, S.A. Systematics and Biology of the Araceae and Acoraceae of Temperate North America. Ph.D. Thesis, University of Illinois at Urbana-Champaign, Urbana, IL, USA, 1995. [Google Scholar]
- Thompson, S.A. Acoraceae. In Flora of North America North of Mexico. Magnoliophyta: Alismatidae, Arecidae, Commelinidae (in Part), and Zingiberidae; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2000; Volume 22, pp. 124–127. [Google Scholar]
- Rajput, S.B.; Tonge, M.B.; Karuppayil, S.M. An overview on traditional uses and pharmacological profile of Acorus calamus Linn. (Sweet flag) and other Acorus species. Phytomedicine 2014, 21, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Shu, H.; Zhang, S.; Lei, Q.; Zhou, J.; Ji, Y.; Luo, B.; Hong, L.; Li, F.; Liu, B.; Long, C. Ethnobotany of Acorus in China. Acta Soc. Bot. Polon. 2018, 87, 3585. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Shu, H.; Zhang, S.; Luo, B.; Gu, R.; Zhang, R.; Ji, Y.; Li, F.; Long, C. From folk taxonomy to species confirmation of Acorus (Acoraceae): Evidences based on phylogenetic and metabolomic analyses. Front. Plant Sci. 2020, 11, 965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W. Pharmacognostic Study of Acorus—From a Phylogenetic Perspective. Ph.D. Thesis, Hong Kong Baptist University, Hong Kong, China, 2005. [Google Scholar]
- Motley, T.J. The ethnobotany of sweet flag, Acorus calamus (Araceae). Econ. Bot. 1994, 48, 397–412. [Google Scholar] [CrossRef]
- Imam, H.; Riaz, Z.; Azhar, M.; Sofi, G.; Hussain, A. Sweet flag (Acorus calamus Linn.): An incredible medicinal herb. Int. J. Green Pharm. 2013, 7, 288–296. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, P.K.; Malviya, R. Pharmacological properties and ayurvedic value of Indian buch plant (Acorus calamus): A short review. Adv. Biol. Res. 2011, 5, 145–154. [Google Scholar]
- Röst, L.C.M.; Bos, R. Biosystematic investigations with Acorus L. 3. Communication. Constituents of essential oils. Planta Med. 1979, 36, 350–361. [Google Scholar] [CrossRef]
- Wulff, H.D. Ölgehalt und Chromosomenzahl des nordamerikanischen Kalmus (Acorus calamus L.). Arch. Pharm. 1950, 283, 155–161. [Google Scholar] [CrossRef]
- Mittal, N.; Varshney, V.K.; Song, B.H.; Ginwal, H.S. High levels of diversity in the phytochemistry, ploidy and genetics of the medicinal plant Acorus calamus L. Med. Aromat. Plants 2015, S1, 002. [Google Scholar] [CrossRef]
- Ahlawat, A.; Katoch, M.; Rama, G.; Ahuja, A. Genetic diversity in Acorus calamus L. as revealed by RAPD markers and its relationship with β-asarone content and ploidy level. Sci. Hortic. 2010, 124, 294–297. [Google Scholar] [CrossRef]
- Ogra, R.K.; Mohanpuria, P.; Sharma, U.K.; Sharma, M.; Sinha, A.K.; Ahuja, P.S. Indian calamus (Acorus calamus L.): Not a tetraploid. Curr. Sci. 2009, 97, 1644–1647. [Google Scholar]
- Wulff, H.D. Zur Zytologie, geographischen Verbreitung und Morphologie des Kalmus. Arch. Pharm. 1954, 287, 529–541. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2023. Available online: http://www.plantsoftheworldonline.org/ (accessed on 29 March 2023).
- Sokoloff, D.D.; Remizowa, M.V.; Nuraliev, M.S.; Averyanov, L.V.; Sennikov, A.N. The first genome from the basal monocot family has been misnamed: Taxonomic identity of Acorus tatarinowii (Acoraceae), a source of numerous chemical compounds of pharmaceutical importance. Diversity 2023, 15, 176. [Google Scholar] [CrossRef]
- Engler, A. Araceae-Pothoideae. In Das Pflanzenreich, 21 (IV. 23B); Engler, A., Ed.; Engelmann: Leipzig, Germany, 1905; pp. 1–330. [Google Scholar]
- Besser, W. Ueber die Flora des Baikals. Flora 1834, 17, 1–30. [Google Scholar]
- Turczaninow, N.S. List of phanerogams and ferns of the Baikal flora. Ukaz. Otkrytij Po Fiz. Khimii Estestv. Istor. I Tekhnologii Izd. N. Shcheglovym 1831, 8, 394–415. (In Russian) [Google Scholar]
- Schott, H.W. Prodromus systematis Aroidearum; Typis Congregationis Mechitharisticae: Vienna, Austria, 1860. [Google Scholar]
- Wein, K. Die älteste Einführungs- und Ausbreitungsgeschichte von Acorus calamus. Hercynia 1939, 1, 367–460. [Google Scholar]
- Linnaeus, C. Species plantarum. L.; Salvius: Holmia, Sweden, 1753. [Google Scholar]
- Smirnov, J.A. Accelerated method of study of somatic chromosomes of fruit plants. Tsitologiya 1968, 10, 1601–1602. (In Russian) [Google Scholar]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code): Adopted by the Nineteenth International Botanical Congress, Shenzhen, China, July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- Chepinoga, V.V.; Gnutikov, A.A.; Enushchenko, I.V.; Chepinoga, A.V. IAPT/IOPB chromosome data 6. Taxon 2008, 57, 1267–1273. [Google Scholar] [CrossRef]
- Krivenko, D.A.; Kotseruba, V.V.; Kazanovsky, S.G.; Verkhozina, A.V.; Chernova, O.D. IAPT/IOPB chromosome data 16. Taxon 2013, 62, 1355–1360. [Google Scholar]
- Probatova, N.S.; Gnutikov, A.A.; Rudyka, E.G.; Chepinoga, V.V. Chromosome numbers of some plant species from Baikal Siberia. Bot. Zhurn. 2008, 93, 162–181. (In Russian) [Google Scholar]
- Chepinoga, V.V. Chromosome Numbers of Plant Species from Baikal Siberia; Nauka: Novosibirsk, Russia, 2014. (In Russian) [Google Scholar]
- Engler, A. Araceae. In Monographiae Phanerogamarum Prodromi Nunc Continuatio, Nunc Revisio; de Candolle, A., de Candolle, C., Eds.; G. Masson: Paris, France, 1879; Volume 2. [Google Scholar]
- Acorus calamus var. angustatus Besser in GBIF Secretariat. GBIF Backbone Taxonomy. Checklist Dataset; GBIF Secretariat: Copenhagen, Denmark, 2022.
- World Flora Online. Available online: https://www.worldfloraonline.org/ (accessed on 13 April 2023).
- Schott, H.W. Araceae (pars altera). Ann. Mus. Bot. Lugduno-Batavi 1864, 1, 278–286. [Google Scholar]
- van Steenis, M.J. Contributions to the history of botany and exploration in Malaysia. 8—9. Blumea 1962, 11, 495–508. [Google Scholar]
- Sokoloff, D.D.; Remizowa, M.V.; Severova, E.E.; Sennikov, A.N. Inference of ploidy level in 19th-century historical herbarium specimens reveals the identity of five Acorus species described by Schott. Diversity 2023, 15, 766. [Google Scholar] [CrossRef]
- Bauhin, C. Pinax Theatri Botanici; Sumptibus & typis Ludovici Regis: Basel, Switzerland, 1623. [Google Scholar]
- Hermann, P. Horti Academici Lugduno-Batavi Catalogus; C. Boutesteyn: Leiden, The Netherlands, 1687. [Google Scholar]
- Rheede tot Drakenstein, H. van Hortus Malabaricus; I. van Someren et al.: Amsterdam, The Netherlands, 1692; Volume 11. [Google Scholar]
- Linnaeus, C. Flora zeylanica. L.; Salvius: Holmia, Sweden, 1747. [Google Scholar]
- Burman, N.L. Flora Indica; C. Haek & J. Schreuder: Leiden, The Netherlands, 1768. [Google Scholar]
- Nicolson, D.; Suresh, C.R.; Manilal, K.S. An Interpretation of Van Rheede’s Hortus Malabaricus; Koeltz Scientific Books: Königstein, Germany, 1988. [Google Scholar]
- Röst, L.C.M. Biosystematic investigations with Acorus L. (Araceae). 1. Communication. Cytotaxonomy. Proc. K. Ned. Akad. Wet. 1978, 81, 428–441. [Google Scholar]
- Manilal, K.S. Van Rheede’s Hortus Malabaricus (Malabar Garden); English edition; University of Kerala: Thiruvananthapuram, India, 2003; Volume 11. [Google Scholar]
- Petersen, G. Cytology and systematics of Araceae. Nord. J. Bot. 1989, 9, 119–166. [Google Scholar] [CrossRef]
- Oganezova, G.H.; Barsegian, N.A. Some peculiarities of the generative organs of Acorus calamus L. from Armenian populations. Flora Rastit. Rastit. Resur. Armen. 1999, 12, 39–41. [Google Scholar]
- Krahulcová, A. Chromosome numbers in selected monocotyledons (Czech Republic, Hungary and Slovakia). Preslia 2003, 75, 97–113. [Google Scholar]
- Larsen, K. Cytology of vascular plants III. A study of Thai aroids. Dansk Bot. Ark. 1969, 27, 39–59. [Google Scholar]
- Wulff, H.D.; Fritz, E. Untersuchungen an einem variegaten Kalmus (Acorus calamus L.). Flora 1958, 146, 328–339. [Google Scholar] [CrossRef]
- Wang, H.; Li, W.L.; Gu, Z.J.; Chen, Y.Y. Cytological study on Acorus L. in southwestern China, with some cytogeographical notes on A. calamus. Acta Bot. Sin. 2001, 43, 354–358. [Google Scholar]
- Ramachandran, K. Cytological studies on South Indian Araceae. Cytologia 1978, 43, 289–303. [Google Scholar] [CrossRef] [Green Version]
- Wulff, H.D. Über die Ursache der Sterilität des Kalmus (Acorus calamus L.). Planta 1940, 31, 478–491. [Google Scholar] [CrossRef]
- Kumar, K.; Singh, M. Chromosomal diversity among different ecotypes of Acorus calamus L. reported from Ranchi Jharkhand, India. Int. J. Bioassays 2015, 4, 3656–3658. [Google Scholar]
- Murata, J. Acoraceae. In Flora of Japan; Iwatsuki, K., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 2016; Volume IVb, p. 5. [Google Scholar]
- Seregin, A.P. (Ed.) Moscow Digital Herbarium: Electronic Resource; Moscow State University: Moscow, Russia, 2023; Available online: https://plant.depo.msu.ru/ (accessed on 11 April 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diploid | Triploid | Tetraploid | Hexaploid | |
|---|---|---|---|---|
| Chromosome numbers | 24, ?sometimes 22 | 36, sometimes 33–35 | 44–48 | 66, 72 |
| Distribution | Asia, N America | Asia, naturalized in Europe and N America | Asia | Rare in Asia (Kashmir and Yunnan) [19,53] |
| Name with nomenclatural priority at the rank of variety | Acorus calamus var. americanus Raf. according to [1] | Acorus calamus var. calamus | Acorus calamus var. verus L. | Unknown |
| Name with nomenclatural priority at the rank of species | Acorus americanus (Raf.) Raf. according to [9] | Acorus calamus L. s.str. | Acorus verus (L.) Burm. f. | Unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokoloff, D.D.; Remizowa, M.V.; Skaptsov, M.V.; Yadav, S.R.; Sennikov, A.N. Back to Linnaeus: Proper Botanical Naming of the Tetraploid Indian Acorus (Acoraceae), an Important Medicinal Plant. Diversity 2023, 15, 785. https://doi.org/10.3390/d15060785
Sokoloff DD, Remizowa MV, Skaptsov MV, Yadav SR, Sennikov AN. Back to Linnaeus: Proper Botanical Naming of the Tetraploid Indian Acorus (Acoraceae), an Important Medicinal Plant. Diversity. 2023; 15(6):785. https://doi.org/10.3390/d15060785
Chicago/Turabian StyleSokoloff, Dmitry D., Margarita V. Remizowa, Mikhail V. Skaptsov, Shrirang R. Yadav, and Alexander N. Sennikov. 2023. "Back to Linnaeus: Proper Botanical Naming of the Tetraploid Indian Acorus (Acoraceae), an Important Medicinal Plant" Diversity 15, no. 6: 785. https://doi.org/10.3390/d15060785