

Separate and Simultaneous Effects of Silver Nanoparticles and Arsenic on the Taxonomic Diversity of Free-Living Marine Nematodes
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
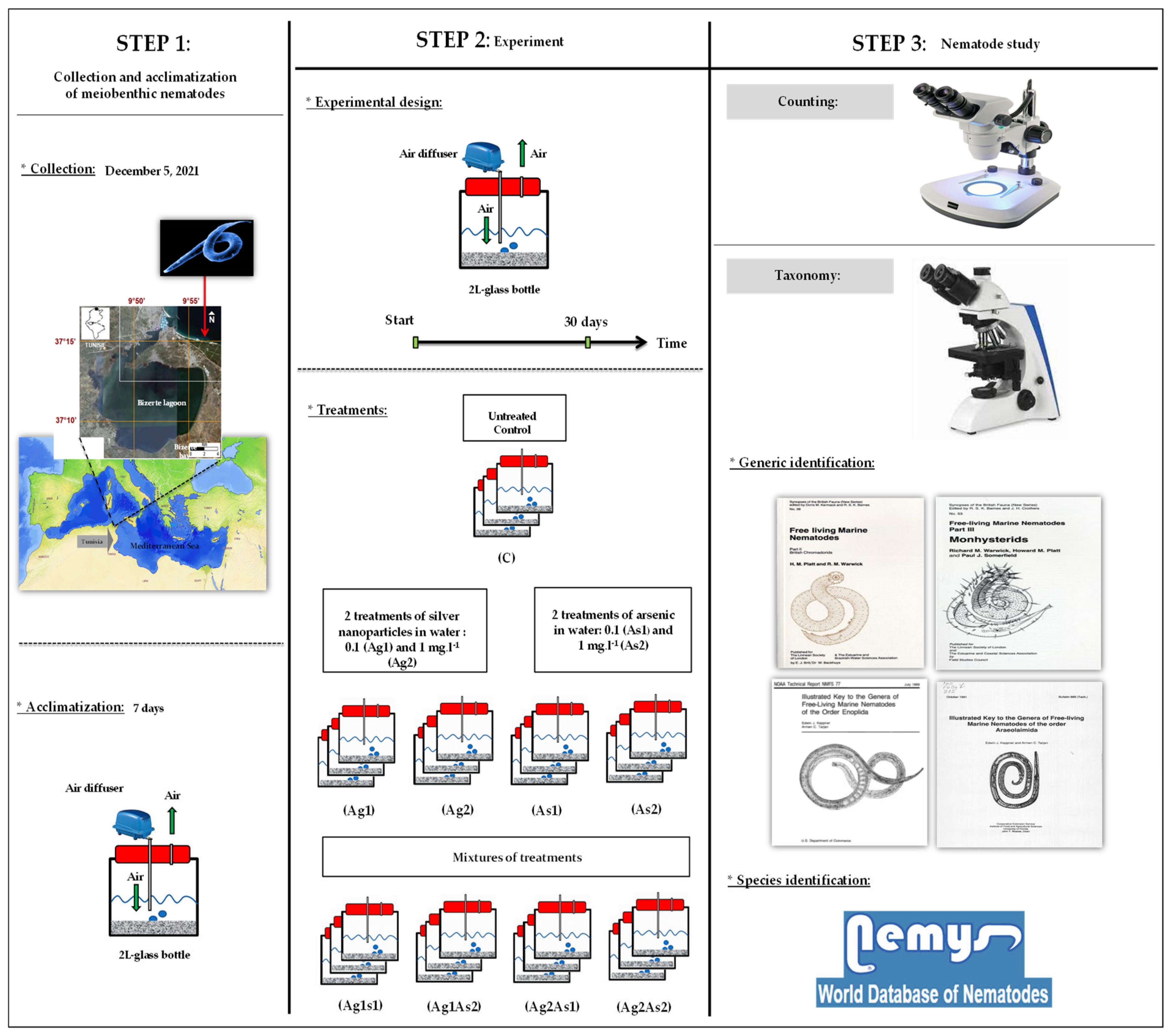
2.1. Collection Site and Laboratory Processing
2.2. Contamination with Arsenic and Silver NPs
- Stock solution was prepared from As (III), to which filtered seawater (0.7 µm pore-size Glass Microfibre GF/F, Whatman, Schnelldorf, Germany) from the collection site in Bizerte Bay (Tunisia) was added to obtain a final concentration of 100 mg·L−1. Aliquots of this solution were diluted and then added to microcosms according to the two intended concentrations: 0.1 (hereafter As1) and 1 mg·L−1 (hereafter As2).
- Ag NPs (described by the vendor as having a size < 100 nm; Sigma-Aldrich Chemical, St. Louis, MO, USA) were homogeneously dispersed in deionized water by sonication (Branson-5210 sonicator; Branson, MO, USA) for 13 h at maximum power, stirring for 7 days, then filtered through a cellulose membrane (pore size 100 nm, Advantec; Toyo Toshi Kaisha, Tokyo, Japan) to remove NP aggregates. These Ag NPs were previously characterized microscopically by [28]. The same two nominal concentrations tested for As were considered once again: 0.1 (hereafter Ag1) and 1 mg·L−1 (hereafter Ag2).
2.3. Nematode Study
2.4. Statistical Processing
3. Results
3.1. Abiotic Features of the Collection Site
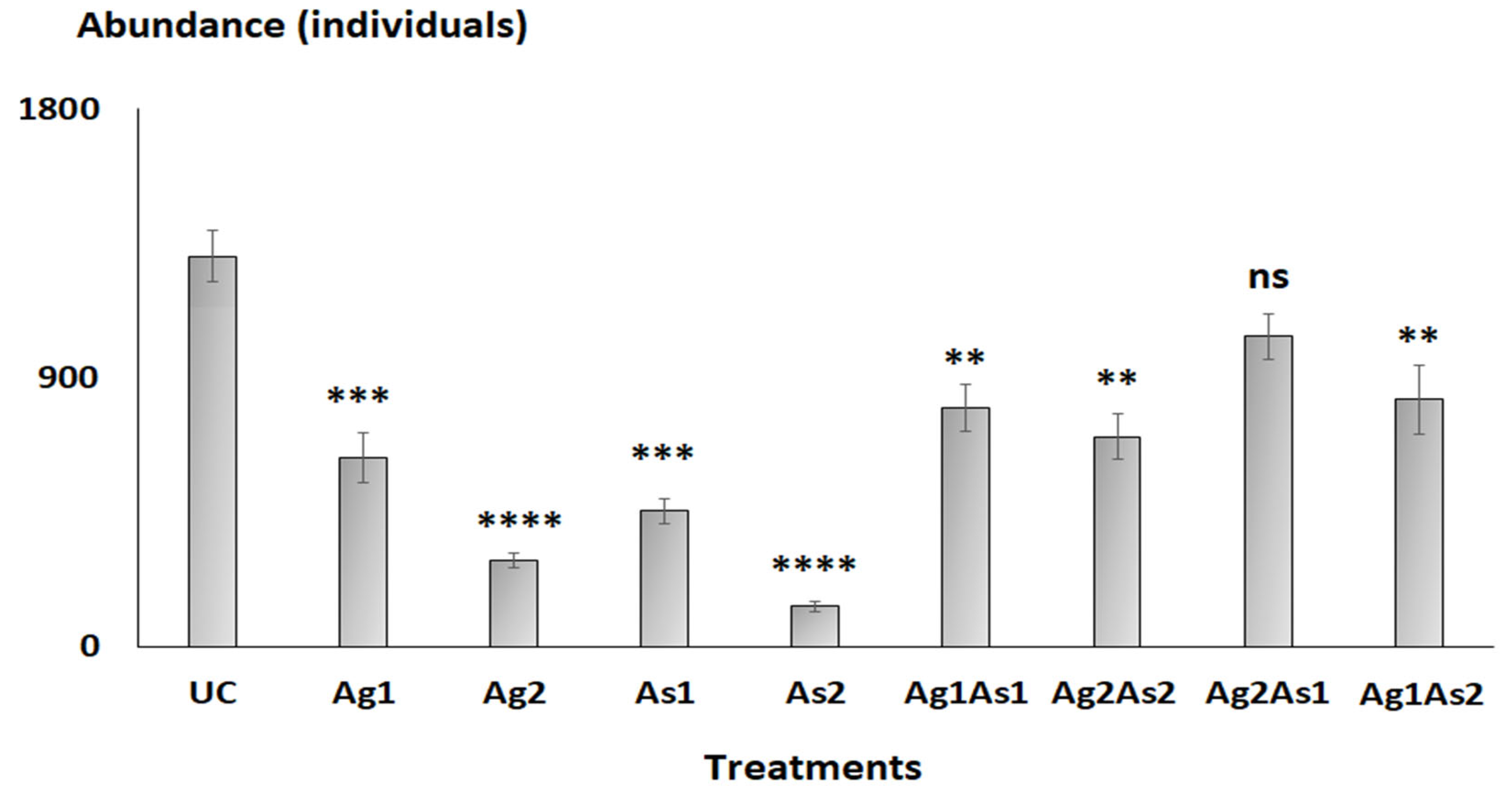
3.2. Nematode Abundances
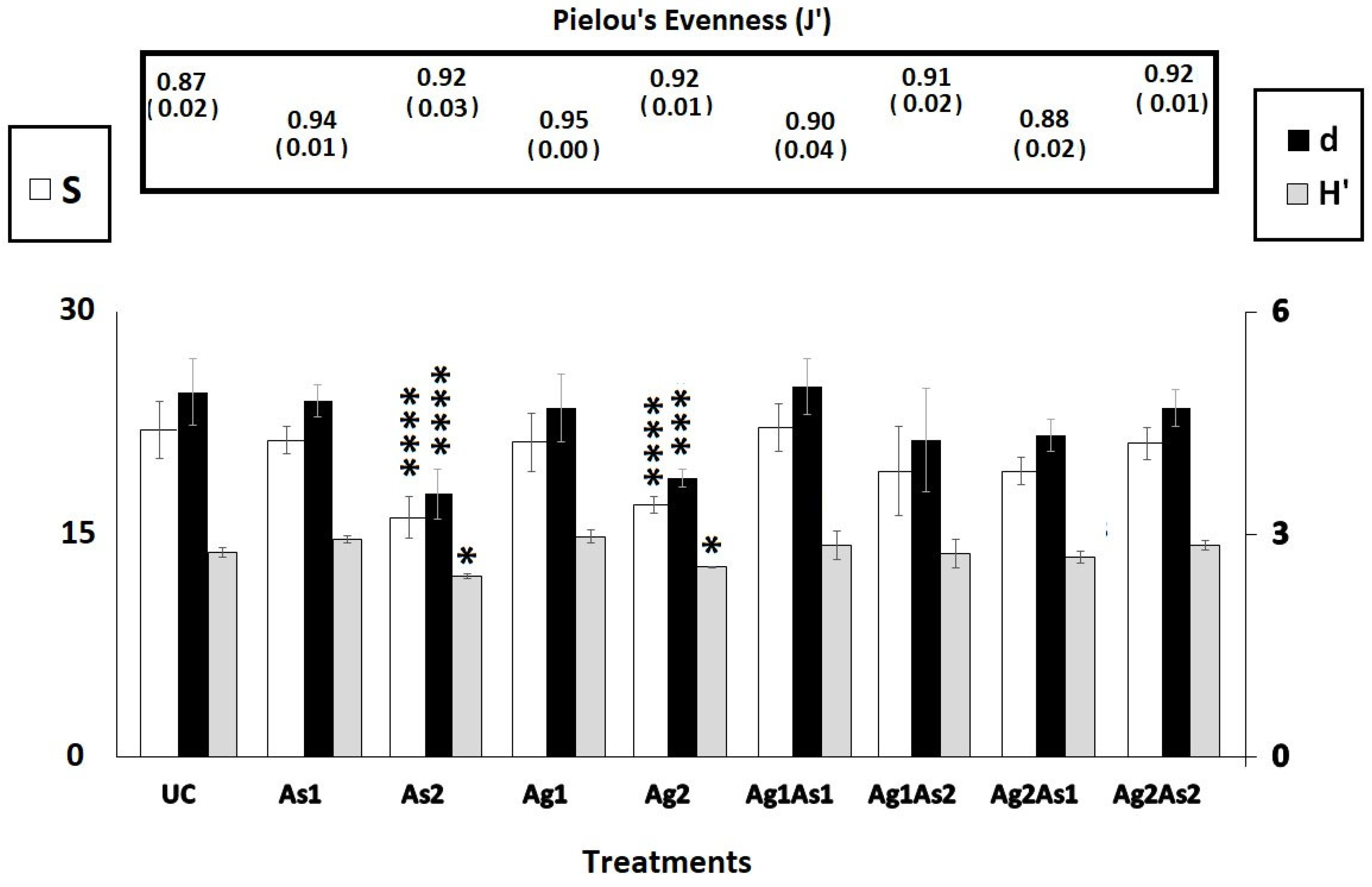
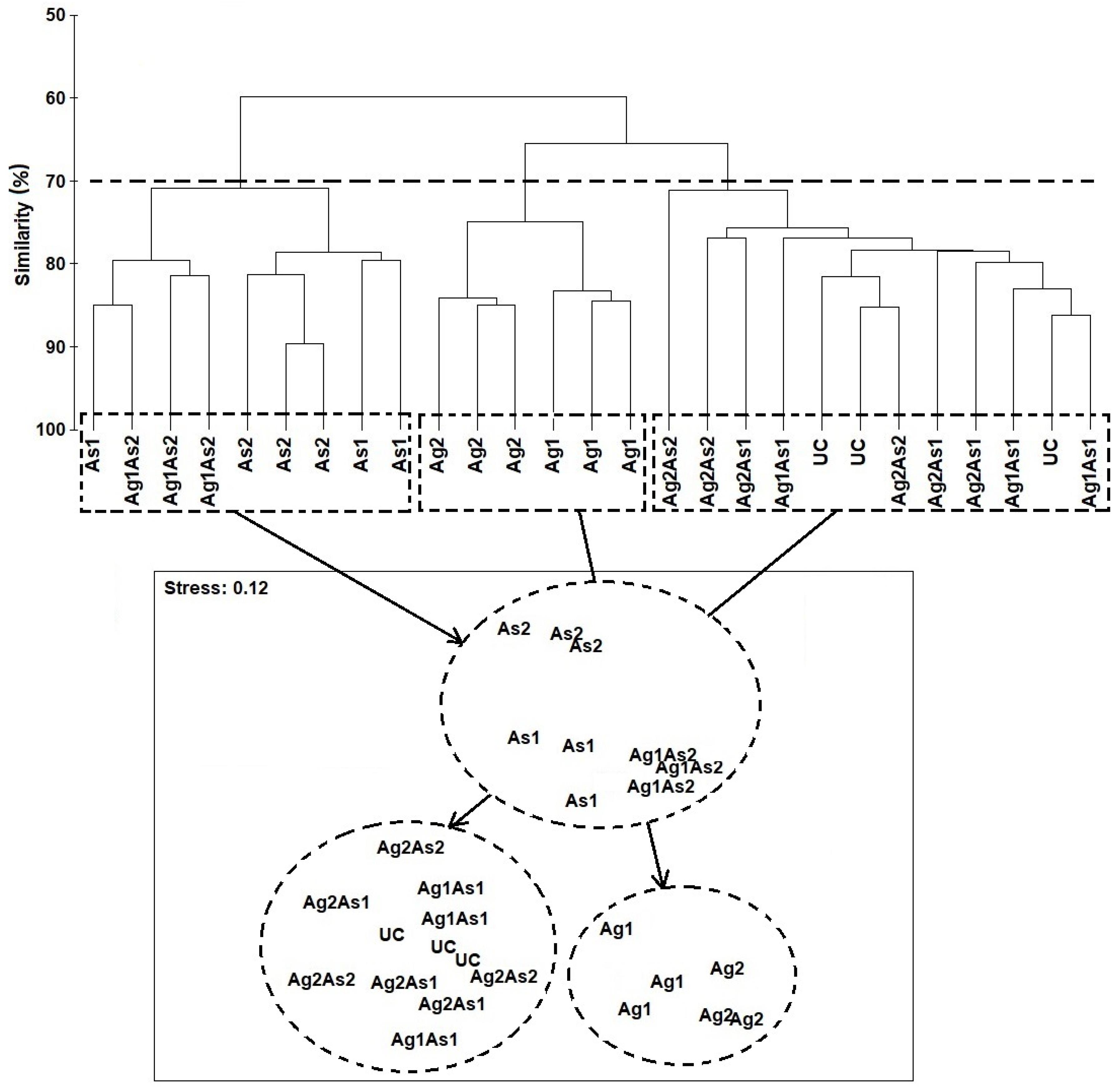
3.3. Taxonomic Census and Community Composition of Nematodes
- Comparing the species’ relative frequency among treatments, several trends were observed. First, with respect to Ag1 and Ag2 versus control, the relative abundance of the nematodes changed. The relative abundances of the species C. honestus and Halaphanolaimus sp., decreased, whereas those of M. pristiurus and D. trabeculosum increased (Table 4). The species L. longicaudatus disappeared in Ag2, whereas the frequency of O. calvadocicus increased compared to Ag1 and control.
- In As1 and As2, the relative abundances of C. honestus, Halaphanolaimus sp., and C. germanicum decreased compared to the control (Table 4); the latter species disappeared in As2. In contrast, the abundance of the species M. pristiurus, D. trabeculosum, and E. longicaudatus increased in both arsenic treatments compared to controls. Similar taxonomic changes in nematofauna were also observed after exposure to Ag1As2, namely C. honestus ↓, Halaphanolaimus sp. ↓, M. pristiurus ↑, and D. trabeculosum ↑ (Table 4).
- In Ag2As1 the relative abundances of M. pristiurus and E. longicaudatus were higher compared to control. SIMPER analyses (Table 4) revealed that the taxonomic changes compared to controls were sometimes linked to one of the two xenobiotics and sometimes to both (Table 4). Several species exhibited an opposite response in mixtures when compared to treatments applied separately (i.e., S. edax in Ag1As1 and P. paradoxus in Ag2As2).
4. Discussion
4.1. Do Meiobenthic Abundances Change When Exposed to Different Treatments?
4.2. How Do Nematodes Respond to the Treatments at the Taxonomic Level?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Avio, C.G.; Gorbi, S.; Milan, M.; Benedetti, M.; Fattorini, D.; d’Errico, G.; Pauletto, M.; Bargelloni, L.; Regoli, F. Pollutants bioavailability and toxicological risk from microplastics to marine mussels. Environ. Pollut. 2015, 198, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Menchaca, I.; Rodriguez, J.G.; Borja, A.; Jesus Belzunce-Segarra, M.; Franco, J.; Garmendia, J.M.; Larreta, J. Determination of polychlorinated biphenyl and polycyclic aromatic hydrocarbon marine regional sediment quality guidelines with the European water framework directive. Chem. Ecol. 2014, 30, 693–700. [Google Scholar] [CrossRef]
- Dhawan, A.; Sharma, V. Toxicity assessment of nanomaterials: Methods and challenges. Anal. Bioanal. Chem. 2010, 398, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Duran, N.; Marcato, P.D.; de Conti, R.; Alves, O.L.; Costa, F.T.M.; Brocchi, M. Potential use of silver nanoparticles on pathogenic bacteria, their toxicity and possible mechanisms of action. J. Braz. Chem. Soc. 2010, 21, 949–959. [Google Scholar] [CrossRef]
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef]
- Garelick, H.; Jones, H.; Dybowska, A.; Valsami-Jones, E. Arsenic pollution sources. Rev. Environ. Contam. Toxicol. 2008, 197, 17–60. [Google Scholar]
- Cubadda, F.; Aureli, F.; Ciardullo, S.; D’Amato, M.; Raggi, A.; Acharya, R.; Reddy, A.V.R.; Tejo Prakash, N. Changes in selenium speciation associated with increasing tissue concentration of selenium in wheat grain. J. Agri. Food Chem. 2010, 58, 2295–2301. [Google Scholar] [CrossRef]
- Smedley, P.L.; Kinniburgh, D.G. A review of the source, behaviour and distribution of arsenic in natural waters. Appl. Geochem. 2002, 17, 517–568. [Google Scholar] [CrossRef] [Green Version]
- Smedley, P.L.; Kinniburgh, D.G. Source and behavior of arsenic in natural waters. In United Nations Synthesis Report on Arsenic in Drinking Water; World Health Organization: Geneva, Switzerland, 2001; pp. 1–61. [Google Scholar]
- Austen, M.C.; McEvoy, A.J.; Warwick, R.M. The specificity of meiobenthiccommunity responses to different pollutants: Results from microcosm experiments. Mar. Pollut. Bull. 1994, 28, 557–563. [Google Scholar] [CrossRef]
- Suderman, K.; Thistle, D. Spills of Fuel Oil and Orimulsion Can Have Indistinguishable Effects on the Benthic Meiofauna. Mar. Pollut. Bull. 2003, 46, 49–55. [Google Scholar] [CrossRef]
- Mahmoudi, E.; Essid, N.; Beyrem, H.; Hedfi, A.; Boufahja, F.; Vitiello, P.; Aïssa, P. Effects of hydrocarbon contamination on a free-living marine nematode community: Results from microcosm experiments. Mar. Pollut. Bull. 2005, 50, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Arce, A.; De Jesús-Navarrete, A.; Leasi, F. DNA Barcoding for Delimitation of Putative Mexican Marine Nematodes Species. Diversity 2020, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Allouche, M.; Nasri, A.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Beyrem, H.; Plăvan, G.; Rohal-Lupher, M.; Boufahja, F. Do presence of gray shrimp Crangon crangon larvae influence meiobenthic features? Assessment with a focus on traits of nematodes. Environ. Sci. Pollut. Res. 2021, 28, 21303–21313. [Google Scholar] [CrossRef] [PubMed]
- Warwick, R.M. The level of taxonomic discrimination required to detect pollution effects on marine benthic communities. Mar. Pollut. Bull. 1988, 19, 259–268. [Google Scholar] [CrossRef]
- Di Toro, D.M.; Zarba, C.S.; Hansen, D.J.; Berry, W.J.; Swartz, R.C.; Cowan, C.E.; Pavlou, S.P.; Allen, H.E.; Thomas, N.A.; Paquin, P.R. Technical basis for establishing sediment quality criteria for nonionic organic chemicals using equilibrium partitioning. Environ. Toxicol. Chem. 1991, 10, 1541–1583. [Google Scholar] [CrossRef]
- Coull, B.C.; Chandler, G.T. Pollution and meiofauna: Field, laboratory and mesocosm studies. Oceanogr. Mar. Biol. 1992, 30, 191–271. [Google Scholar]
- Carman, K.R.; Todaro, M.A. Influence of polycyclic aromatic hydrocarbons on the meiobenthic-copepod community of a Louisiana salt marsh. J. Exp. Mar. Biol. Ecol. 1996, 198, 37–54. [Google Scholar] [CrossRef]
- Lotufo, G.R.; Fleeger, J.W. Effects of sediment-associated phenanthrene on survival, development and reproduction of two species of meiobenthic copepods. Mar. Ecol. Prog. Ser. 1997, 151, 91–102. [Google Scholar] [CrossRef]
- Hedfi, A.; Ben Ali, M.; Korkobi, M.; Allouche, M.; Harrath, A.H.; Beyrem, H.; Pacioglu, O.; Badraoui, R.; Boufahja, F. The exposure to polyvinyl chloride microplastics and chrysene induces multiple changes in the structure and functionality of marine meiobenthic communities. J. Hazard. Mater. 2022, 436, 12916. [Google Scholar] [CrossRef]
- Allouche, M.; Ishak, S.; Ben Ali, M.; Hedfi, A.; Almalki, M.; Karachle, P.K.; Harrath, A.H.; Abu-Zied, R.H.; Badraoui, R.; Boufahja, F. Molecular interactions of polyvinyl chloride microplastics and beta-blockers (diltiazem and bisoprolol) and their effects on marine meiofauna: Combined in vivo and modelling study. J. Hazard. Mater. 2022, 431, 128609. [Google Scholar] [CrossRef]
- Boufahja, F. Approches Communautaires et Populationnelles de Biosurveillance du Milieu Marin chez les Nématodes Libres (Lagune et Baie de Bizerte, Tunisie). Ph.D. Thesis, Biological Sciences, Faculty of Sciences of Bizerte (University of Carthage, Tunisia), Carthage, Tunisia, 2010; 300p. [Google Scholar]
- Hedfi, A.; Boufahja, F.; Ben Ali, M.; Aïssa, P.; Mahmoudi, E.; Beyrem, H. Do trace metals (chromium, copper and nickel) influence toxicity of diesel fuel for free-living marine nematodes? Environ. Sci. Pollut. Res. 2013, 20, 3760–3770. [Google Scholar] [CrossRef] [PubMed]
- Hedfi, A.; Ben Ali, M.; Noureldeen, A.; Darwish, H.; Mahmoudi, E.; Plăvan, G.; Pacioglu, O.; Boufahja, F. Effects of benzo(a)pyrene on meiobenthic assemblage and biochemical biomarkers in an Oncholaimus campylocercoides (Nematoda) microcosm. Environ. Sci. Pollut. Res. 2021, 29, 16529–16548. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, J.B. Measurement of the physical and chemical environment: Sediments. In Methods for the Study of Marine Benthos. International Biological Programme Handbook No. 16; Holme, N., McIntyre, A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1971; 334p. [Google Scholar]
- Fabiano, M.; Danovaro, R. Composition of organic matter in sediment facing a river estuary (Tyrrhenian Sea): Relationships with bacteria and microphytobenthic biomass. Hydrobiologia 1994, 277, 71–84. [Google Scholar] [CrossRef]
- Boufahja, F.; Hedfi, A.; Amorri, J.; Aïssa, P.; Beyrem, H.; Mahmoudi, E. Examination of the bioindicator potential of Oncholaimus campylocercoides (Nematoda, Oncholaimidae) from Bizerte bay (Tunisia). Ecol. Indic. 2011, 11, 1139–1148. [Google Scholar] [CrossRef]
- Lee, W.A.; Bonin, V.; Reed, M.; Graham, B.J.; Hood, G.; Glattfelder, K.; Reid, R.C. Anatomy and function of an excitatory network in the visual cortex. Nature 2016, 532, 370–374. [Google Scholar] [CrossRef] [Green Version]
- USEPA. Test methods for evaluating solid waste. In Vol. IB: Laboratory Manual Physicallchemical Methods. SW-846; USEPA: Washington, DC, USA, 1986. Available online: https://nepis.epa.gov/Exe/ZyNET.exe/ (accessed on 26 November 2022).
- ANZECC/ARMCANZ. Australian and New Zealand Guidelines for Fresh and Marine Water Quality, Vol. 1. The Guidelines; Australian and New Zealand Environment and Conservation Council and Agriculture and Resource Management Council of Australia and New Zealand: Auckland, New Zealand, 2000. [Google Scholar]
- Diniz, M.S.; Santos, H.M.; Costa, P.M.; Peres, I.; Costa, M.H.; Capelo, J.L. Metallothionein responses in the Asiatic clam (Corbicula fluminea) after exposure to trivalent Arsenic. Biomarkers 2007, 12, 589–598. [Google Scholar] [CrossRef]
- Liu, W.; Zeng, Z.; Chen, A.; Zeng, G.; Xiao, R.; Guo Zhi, Y.F.; Huang, Z.; He, K.; Hu, L. Toxicity effects of silver nanoparticles on the freshwater bivalve Corbicula fluminea. J. Environ. Chem. Eng. 2018, 6, 4236–4244. [Google Scholar] [CrossRef]
- Vitiello, P.; Dinet, A. Définition et échantillonnage du méiobenthos. Rapp. Comm. Int. Expl. Sci. Mer. Médit. 1979, 25, 279–283. [Google Scholar]
- Guo, Y.; Somerfield, P.J.; Warwick, R.M.; Zhang, Z. Large-scale patterns in the community structure and biodiversity of free-living nematodes in the Bohai Sea, China. J. Mar. Biol. Assoc. U. K. 2001, 81, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Seinhorst, J.W. A Rapid Method for the Transfer of Nematodes From Fixative To Anhydrous Glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Platt, H.M.; Warwick, R.M. Free-Living Marine Nematodes. Part I. British Enoplids. Synopses of the British Fauna No 28; Cambridge University Press: Cambridge, UK, 1983; 314p. [Google Scholar]
- Platt, H.M.; Warwick, R.M. Free-Living Marine Nematodes. Part II. British Chromadorids. Synopsis of the British Fauna (New Series) No. 38; Brill, E.J., Backhuys, W., Eds.; The Linnean Society of London: London, UK, 1988. [Google Scholar]
- Bezerra, T.N.; Decraemer, W.; Eisendle-Flöckner, U.; Hodda, M.; Holovachov, O.; Leduc, D.; Miljutin, D.; Mokievsky, V.; Peña Santiago, R.; Sharma, J.; et al. Nemys: World Database of Nematodes. 2019. Available online: http://nemys.ugent.be/ (accessed on 26 November 2022).
- Wieser, W. Die Besiehung zwischen Mundhchlen-gestalt, Ernährungsweise und. Vorkommen beifreilbenden marinen Nematoden. Ark. Zoop. Ser. 1953, 2, 439–484. [Google Scholar]
- Thistle, D.; Lambshead, P.J.D.; Sherman, K.M. Nematode tail-shape groups respond to environmental differences in the deep sea. Vie Milieu/Life Environ. 1995, 45, 107–115. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Glorley, R.N. PRIMER v5: User Manual/Tutorial; PRIMER-E: Polymouth, UK, 2001; 91p. [Google Scholar]
- Hedfi, A.; Mahmoudi, E.; Boufahja, F.; Beyrem, H.; Aïssa, P. Effects of increasing levels of nickel contamination on structure of offshore nematode communities in experimental microcosms. Bull. Environ. Contam. Toxicol. 2007, 79, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Beyrem, H.; Boufahja, F.; Hedfi, A.; Essid, N.; Aïssa, P.; Mahmoudi, E. Laboratory study on individual and combined effects of cobalt- and zinc-spiked sediment on meiobenthic nematodes. Biol. Trace Elem. Res. 2011, 144, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Wakkaf, T.; Allouche, M.; Harrath, A.H.; Mansour, L.; Alwasel, S.; Ansari, K.G.M.T.; Beyrem, H.; Sellami, B.; Boufahja, F. The individual and combined effects of cadmium, Polyvinyl chloride (PVC) microplastics and their polyalkylamines modified forms on meiobenthic features in a microcosm. Environ. Pollut. 2020, 266, 115263. [Google Scholar] [CrossRef] [PubMed]
- Bernard, G.C.; Fitch, J.; Min, B.; Shahi, N.; Egnin, M.; Ritte, I.; Collier, W.E.; Bonsi, C. Potential Nematicidial Activity of Silver Nanoparticles Against the Root-Knot Nematode (Meloidogyne incognita). Online J. Complement. Altern. Med. 2019, 2, 000531. [Google Scholar] [CrossRef]
- Ghareeb, R.Y.; Shams El-Din, N.G.E.; Maghraby, D.M.E.; Ibrahim, D.S.S.; Abdel-Megeed, A.; Abdelsalam, N.R. Nematicidal activity of seaweed-synthesized silver nanoparticles and extracts against Meloidogyne incognita on tomato plants. Sci. Rep. 2022, 12, 3841. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon nanoparticles more effectively alleviated UV-B stress than silicon in wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef]
- Wang, A.; Cui, Y.; Yang, Y.; Li, J. Capsules with silver nanoparticle enrichment subdomains and their antimicrobial properties. Chem. Asian J. 2010, 5, 1780–1787. [Google Scholar] [CrossRef]
- Jung, J.; Raghavendra, G.M.; Kim, D.; Seo, J. An investigation on the antibacterial, cytotoxic, and antibiofilm efficacy of starch-stabilized silver nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2018, 8, 916–924. [Google Scholar]
- Oukarroum, A.; Bras, S.; Perreault, F.; Popovic, R. Inhibitory effects of silver nanoparticles in two green algae, Chlorella vulgaris and Dunaliella tertiolecta. Ecotoxicol. Environ. Saf. 2012, 78, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Schirmer, K.; Bernard, L.; Sigg, L.; Pillai, S.; Behra, R. Silver nanoparticle toxicity and association with the alga Euglena gracilis. Environ. Sci. Nano 2015, 2, 594–602. [Google Scholar] [CrossRef]
- Burchardt, A.D.; Carvalho, R.N.; Valente, A.; Nativo, P.; Gilliland, D.; Garcìa, C.P.; Passarella, R.; Pedroni, V.; Rossi, F.; Lettieri, T. Effects of silver nanoparticles in diatom Thalassiosira pseudonana and cyanobacterium Synechococcus sp. Environ. Sci. Technol. 2012, 46, 11336–11344. [Google Scholar] [CrossRef]
- Navarro, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R. Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 2008, 42, 8959–8964. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Tran, I.; Agrawal, A.F. Does genetic variation maintained by environmental heterogeneity facilitate adaptation to novel selection? Am. Nat. 2016, 188, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, S.; De Francisco, P.; Olsson, S.; Aguilera, A.; González-Toril, E.; Martín-González, A. Toxicity, Physiological, and Ultrastructural Effects of Arsenic and Cadmium on the Extremophilic Microalga Chlamydomonas acidophila. Int. J. Environ. Res. Public Health 2020, 17, 1650. [Google Scholar] [CrossRef] [Green Version]
- De Francisco, P.; Martín-González, A.; Rodriguez-Martín, D.; Díaz, S. Interactions with Arsenic: Mechanisms of Toxicity and Cellular Resistance in Eukaryotic Microorganisms. Int. J. Environ. Res. Public Health 2021, 18, 12226. [Google Scholar] [CrossRef]
- Moens, T.; Vincx, M. Observations on the feeding ecology of estuarine nematodes. J. Mar. Biol. Assoc. U. K. 1997, 77, 211–227. [Google Scholar] [CrossRef]
- Skebo, J.E.; Grabinski, C.M.; Schrand, A.M.; Schlager, J.J.; Hussain, S.M. Assessment of metal nanoparticle agglomeration, uptake, and interaction using a high-illuminating system. Int. J. Toxicol. 2007, 26, 135–141. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Family | Genus | Species | FG | TS |
|---|---|---|---|---|---|
| Enoplida | Tripyloididae | Bathylaimus | sp. | 2B | co |
| Desmodorida | Microlaimidae | Calomicrolaimus | honestus | 2A | co |
| Desmoscolecida | Cyartonematidae | Cyartonema | germanicum | 1A | co |
| Monhysterida | Xyalidae | Daptonema | fallax | 1B | co |
| Monhysterida | Xyalidae | Daptonema | normandicum | 1B | cla |
| Monhysterida | Xyalidae | Daptonema | trabeculosum | 1B | cla |
| Enoplida | Thoracostomopsidae | Enoplolaimus | longicaudatus | 2B | cla |
| Enoplida | Oxystominidae | Halalaimus | gracilis | 1A | e/f |
| Plectida | Leptolaimidae | Halaphanolaimus | sp. | 1A | co |
| Chromadorida | Cyatholaimidae | Longicyatholaimus | longicaudatus | 2A | e/f |
| Chromadorida | Cyatholaimidae | Marylynnia | punctata | 2A | e/f |
| Chromadorida | Cyatholaimidae | Marylynnia | steckhoveni | 2A | e/f |
| Enoplida | Oncholaimidae | Metoncholaimus | pristiurus | 2B | cla |
| Araeolaimida | Axonolaimidae | Odontophora | wieseri | 1B | co |
| Enoplida | Oncholaimidae | Oncholaimellus | calvadocicus | 2B | cla |
| Monhysterida | Xyalidae | Paramonohystera | proteus | 1B | cla |
| Monhysterida | Sphaerolaimidae | Parasphaerolaimus | paradoxus | 2B | cla |
| Enoplida | Phanodermatidae | Phanoderma | sp. | 2A | co |
| Chromadorida | Chromadoridae | Prochromadorella | longicaudata | 2A | co |
| Araeolaimida | Comesomatidae | Sabatieria | splendens | 1B | cla |
| Araeolaimida | Comesomatidae | Sabatieria | punctata | 1B | cla |
| Desmodorida | Desmodoridae | Spirinnia | parasitifera | 2A | co |
| Chromadorida | Selachinematidae | Synonchiella | edax | 2B | co |
| Enoplida | Ironidae | Thalassironus | britannicus | 2B | co |
| Monhysterida | Xyalidae | Theristus | modicus | 1B | co |
| Enoplida | Enchelidiidae | Thoonchus | inermis | 2B | cla |
| Monhysterida | Xyalidae | Valvaelaimus | maior | 1B | co |
| Species | UC | Ag1 | Ag2 | As1 | As2 | Ag1As1 | Ag2As2 | Ag2As1 | Ag1As2 |
|---|---|---|---|---|---|---|---|---|---|
| Bathylaimus sp. | 2 (1) | 2.33 (2.52) | 4.67 (0.58) | 5.33 (1.52) | 8.67 (1.15) | 2 (1) | 3.67 (0.58) | 0.67 (1.15) | 4.33 (0.58) |
| Calomicrolaimus honestus | 14.66 (4.16) | 6 (1.73) | 1.33 (1.15) | 1.33 (1.52) | 2 (1) | 10.33 (1.53) | 11.33 (1.53) | 12.33 (2.08) | 3 (2.65) |
| Cyartonema germanicum | 7 (2.64) | 5.33 (0.58) | 0.67 (0.58) | 0.66 (0.57) | 8.33 (3.06) | 8 (1.73) | 8.67 (1.53) | 1.67 (1.15) | |
| Daptonema fallax | 2 (1) | 5.33 (0.58) | 8 (1) | 1.66 (0.57) | 2.67 (1.53) | 0.33 (0.58) | 2.67 1.15) | 2.67 (1.15) | |
| Daptonema normandicum | 7.33 (3.21) | 7.67 (1.53) | 10 (1) | 7.66 (2.08) | 1.33 (1.15) | 7 (2.65) | 7.33 (3.51) | 8.67 (3.06) | 3 (2.65) |
| Daptonema trabeculosum | 0.66 (0.57) | 3 (1) | 6.67 (1.53) | 7.66 (0.57) | 7 (2) | 3 (1) | 2.33 (2.52) | 11.33 (1.15) | |
| Enoplolaimus longicaudatus | 0.66 (1.15) | 0.33 (0.58) | 6 (3) | 11 (1) | 1 (1) | 1.33 (1.15) | 1.67 (1.15) | 2.33 (2.52) | |
| Halalaimus gracilis | 1.33 (0.57) | 1 (1) | 1.33 (1.53) | 0.66 (1.15) | 9.67 (9.87) | 2.67 (1.15) | 2 (2.65) | 0.33 (0.58) | |
| Halaphanolaimus sp. | 18 (3.6) | 7.33 (1.15) | 0.67 (1.15) | 3.66 (1.52) | 2 (1) | 10.67 (2.89) | 12 (4) | 17.67 (0.58) | |
| Longicyatholaimus longicaudatus | 3.33 (2.08) | 2.33 (1.53) | 3.33 (1.52) | 4 (2.65) | 1.67 (0.58) | 0.67 (0.58) | 2 (1) | 2.33 (1.53) | |
| Metoncholaimus pristiurus | 0.66 (1.15) | 8.33 (1.53) | 9 (1) | 5.33 (0.57) | 7 (1) | 0.67 (0.58) | 3.33 (1.15) | 1.67 (1.15) | 9.67 (1.53) |
| Marylynnia puncticaudata | 7 (1.73) | 3.33 (1.53) | 2 (2) | 6.33 (3.51) | 5.67 (2.08) | 8.67 (1.15) | 10.33 (2.89) | 7.33 (2.08) | 2.33 (2.52) |
| Marylynnia steckhoveni | 1 (1.73) | 0.67 (1.15) | 0.67 (0.58) | 2 (1) | 0.33 (0.58) | 2 (1) | 1 (1.73) | 2.33 (2.08) | 2.67 (0.58) |
| Odontophora villoti | 1.33 (0.57) | 1 (1) | 0.67 (1.15) | 5.66 (2.08) | 11.33 (3.79) | 2.67 (2.08) | 1.67 (2.08) | 1.33 (1.53) | 5.33 (0.58) |
| Oncholaimellus calvadosicus | 2 (1) | 5.67 (2.08) | 9.33 (1.15) | 1.33 (1.15) | 2.33 (1.53) | 2.33 (2.52) | 3 (1) | 0.33 (0.58) | |
| Paramonohystera proteus | 7.33 (2.08) | 6.67 (2.31) | 9.67 (1.53) | 3.66 (1.15) | 5 (2.65) | 6 (2) | 11.33 (2.52) | 8.33 (1.53) | 5 (2.65) |
| Parasphaerolaimus paradoxus | 2.33 (1.52) | 1 (1.73) | 0.33 (0.58) | 1 (1) | 1 (1.73) | 2.67 (1.53) | 2.33 (2.52) | 2 (1.73) | 0.67 (0.58) |
| Phanoderma sp. | 1.33 (1.15) | 1 (1) | 1 (1) | 2.67 (2.31) | 1.33 (0.58) | 1.67 (0.58) | 1.33 (1.53) | 2.33 (1.53) | |
| Prochromadorella longicaudata | 2.66 (1.52) | 0.33 (0.58) | 0.67 (0.58) | 10 (3.6) | 13.67 (1.53) | 1 (1) | 1.33 (1.15) | 1.67 (2.08) | 6.67 (2.52) |
| Synonchiella edax | 1.33 (1.52) | 1 (1) | 0.67 (0.58) | 5.33 (1.52) | 5.67 (1.53) | 1 (1.73) | 1 (1) | 1.33 (1.53) | 10.67 (1.53) |
| Sabatiera splendens | 1.66 (0.57) | 4 (0) | 8.33 (1.53) | 1.66 (2.08) | 2.67 (0.58) | 0.67 (1.15) | 0.67 (0.58) | 2 (1) | |
| Spirinnia parasitifera | 2.33 (1.52) | 4 (1.73) | 1 (1) | 1 (1.73) | 2 (2) | 2 (2) | 0.67 (1.15) | 3.33 (1.53) | |
| Sabatieria punctata | 2.66 (1.15) | 6.67 (0.58) | 9.67 (1.15) | 3.33 (2.08) | 3.67 (1.53) | 3 (1.73) | 2.33 (1.53) | 2.66 (2.52) | 1.67 (1.15) |
| Thalassironus britannicus | 1.66 (1.15) | 2 (1) | 2.33 (1.52) | 1.67 (1.15) | 2.33 (1.53) | 1.33 (2.31) | 1 (1.73) | 1.67 (0.58) | |
| Theristus modicus | 4 (1) | 5 (1) | 5.67 (1.15) | 4 (1) | 0.33 (0.58) | 2.33 (0.58) | 3.67 (2.08) | 3.33 (2.52) | 1.33 (1.15) |
| Thoonchus inermis | 1.33 (1.15) | 4 (0) | 3.67 (0.58) | 2.33 (0.57) | 2 (2.65) | 2.33 (1.53) | 2.67 (0.58) | ||
| Valvaelaimus maior | 2.33 (0.57) | 4.67 (1.53) | 5.33 (0.58) | 5.66 (2.08) | 6 (3) | 3 (2.65) | 2 (1.73) | 2.67 (2.08) | 10.67 (1.15) |
| Source | df | SS | MS | Pseudo-F | p (perm) | Perm | ||
|---|---|---|---|---|---|---|---|---|
| Treatment | 8 | 6257.9 | 782.23 | 253.31 | 0.001 | 998 | ||
| Residual | 18 | 55.586 | 3.0881 | |||||
| Total | 26 | 6313.4 | ||||||
| Pair-wise tests (UC vs.) | Ag1 | Ag2 | As1 | As2 | Ag1As1 | Ag2As2 | Ag2As1 | Ag1As2 |
| p | 0.0987 | 0.0385 | 0.0481 | 0.0291 | 0.6104 | 0.1941 | 0.6932 | 0.0431 |
| Comparisons | SIMPER | Species |
|---|---|---|
| AD (%) | ||
| UC vs. Ag1 | 25.34 | Metoncholaimus pristiurus (10.65%) + Halaphanolaimus sp. (6.75%) − Calomicrolaimus honestus (6.01%) − Prochromadorella longicaudata (5.54%) − Parasphaerolaimus paradoxus (5.01%) − Thoonchus inermis (4.69%) + Daptonema trabeculosum (4.62%) + Oncholaimellus calvadocicus (4.32%) + Sabatieria punctata (4.28%) + |
| UC vs. Ag2 | 41.46 | Halaphanolaimus sp. (10.8%) − Calomicrolaimus honestus (8.19%) − Metoncholaimus pristiurus (7.27%) + Cyartonema germanicum (5.57%) − Daptonema trabeculosum (5.47%) + Longicyatholamus longicaudatus (5.02%) ø Oncholaimellus calvadocicus (4.8%) + Sabatieria splendens (4.61%) + |
| UC vs. As1 | 33.38 | Calomicrolaimus honestus (9.73%) − Halaphanolaimus sp. (8.92%) − Daptonema trabeculosum (8.31%) + Cyartonema germanicum (7.56%) − Enoplolaimus longicaudatus (7.56%) + Metoncholaimus pristiurus (7.21%) + Synonchiella edax (4.62%) + |
| UC vs. As2 | 47.3 | Cyartonema germanicum (7.64%) ø Halaphanolaimus sp. (7.26%) − Enoplolaimus longicaudatus (7.25%) + Calomicrolaimus honestus (7.15%) − Odontophora villoti (6.58%) + Metoncholaimus pristiurus (6.54%) + Daptonema trabeculosum (6.37%) + Daptonema normandicum (4.4%) − |
| UC vs. Ag1As1 | 21.49 | Halalaimus gracilis (9.79%) + Daptonema trabeculosum (5.96%) + Marylynnia steckhoveni (5.86%) + Halaphanolaimus sp. (5.56%) − Thoonchus inermis (5.26%) ø Synonchiella edax (4.98%) − Prochromadorella longicaudata (4.97%) − Spirinnia parasitifera (4.33%) − Enoplolaimus longicaudatus (4.14%) + |
| UC vs. Ag2As2 | 23.85 | Metoncholaimus pristiurus (6.49%) + Longicyatholamus longicaudatus (5.24%) − Thalassironus britannicus (5.23%) − Daptonema fallax (5.17%) − Daptonema trabeculosum (4.93%) + Thoonchus inermis (4.63%) + Sabatieria splendens (4.42%) − Parasphaerolaimus paradoxus (4.25%) st Prochromadorella longicaudata (4.1%) − Omcholaimellus calvadocicus (4.04%) + Halaphanolaimus sp. (3.99%) − |
| UC vs. Ag2As1 | 19.97 | Spirinnia parasitifera (5.98%) − Marylynnia steckhoveni (5.71%) + Bathylaimus sp. (5.46%) − Thalassironus britannicus (5.4%) − Metoncholaimus pristiurus (5.22%) + Enoplolaimus longicaudatus (5.21%) + Prochromadorella longicaudata (4.65%) − Sabatieria punctata (4.57%) st Halalaimus gracilis (4.31%) + Phanoderma sp. (4.24%) st |
| UC vs. Ag1As2 | 37.92 | Halaphanolaimus sp. (12.65%) ø Daptonema trabeculosum (8.07%) + Metoncholaimus pristiurus (7.89%) + Calomicrolaimus honestus (7.1%) − Synonchiella edax (7%) + Valvaelaimus maior (5.2%) + Marylynnia puncticaudata (4.24%) − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hedfi, A.; Ishak, S.; Al Hebs, A.; Al-Hoshani, N.; Ben Ali, M.; Rudayni, H.A.; Boufahja, F. Separate and Simultaneous Effects of Silver Nanoparticles and Arsenic on the Taxonomic Diversity of Free-Living Marine Nematodes. Diversity 2023, 15, 836. https://doi.org/10.3390/d15070836
Hedfi A, Ishak S, Al Hebs A, Al-Hoshani N, Ben Ali M, Rudayni HA, Boufahja F. Separate and Simultaneous Effects of Silver Nanoparticles and Arsenic on the Taxonomic Diversity of Free-Living Marine Nematodes. Diversity. 2023; 15(7):836. https://doi.org/10.3390/d15070836
Chicago/Turabian StyleHedfi, Amor, Sahar Ishak, Abdullah Al Hebs, Nawal Al-Hoshani, Manel Ben Ali, Hassan A. Rudayni, and Fehmi Boufahja. 2023. "Separate and Simultaneous Effects of Silver Nanoparticles and Arsenic on the Taxonomic Diversity of Free-Living Marine Nematodes" Diversity 15, no. 7: 836. https://doi.org/10.3390/d15070836