
Habitat Loss and Biotic Velocity Response to Climate Change for Alpine Plant Species in Atlantic Oceanic Islands
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
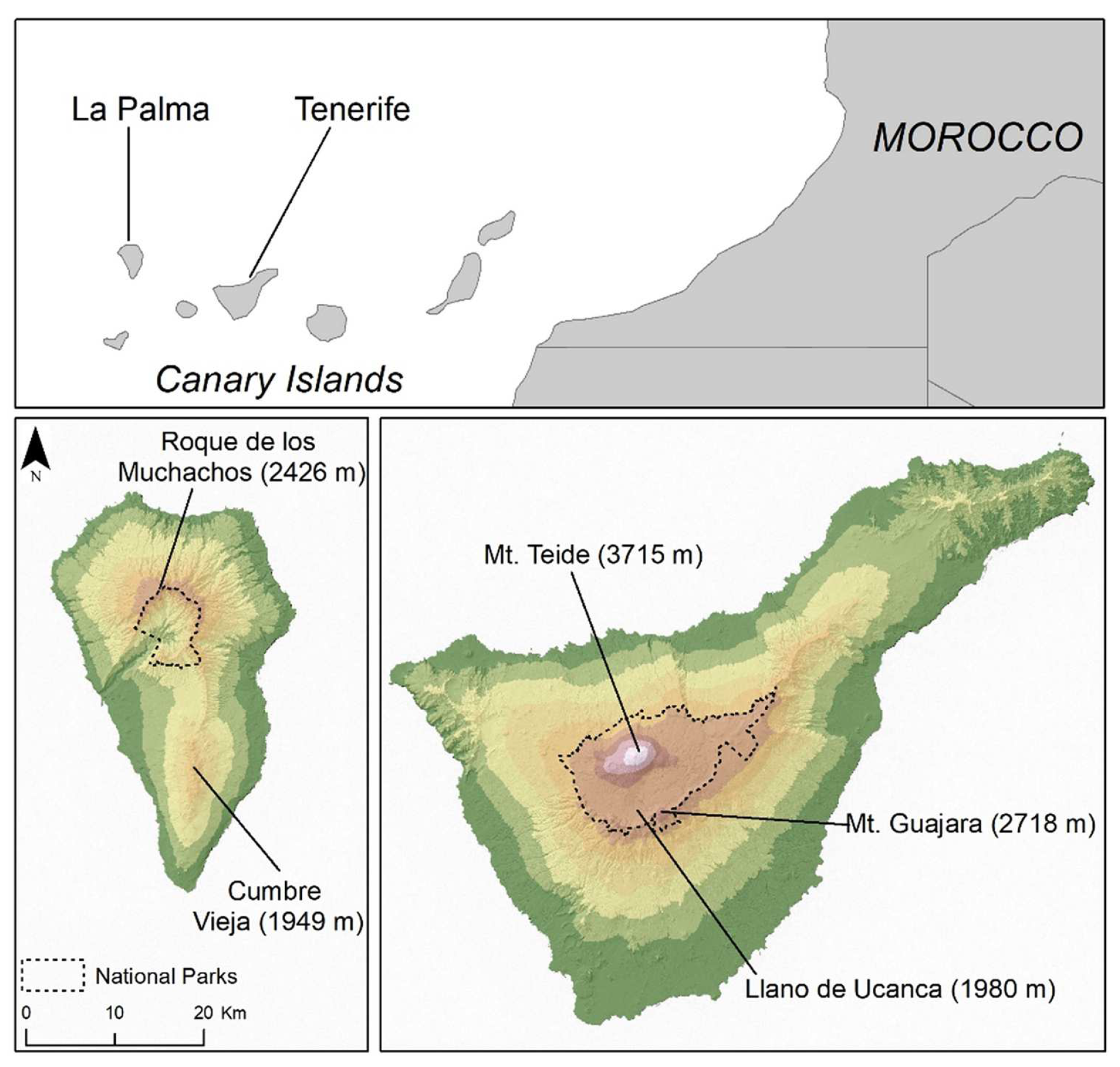
2.1. Study Area and Species Data
2.2. Climate Data
2.3. Modeling and Analysis
3. Results
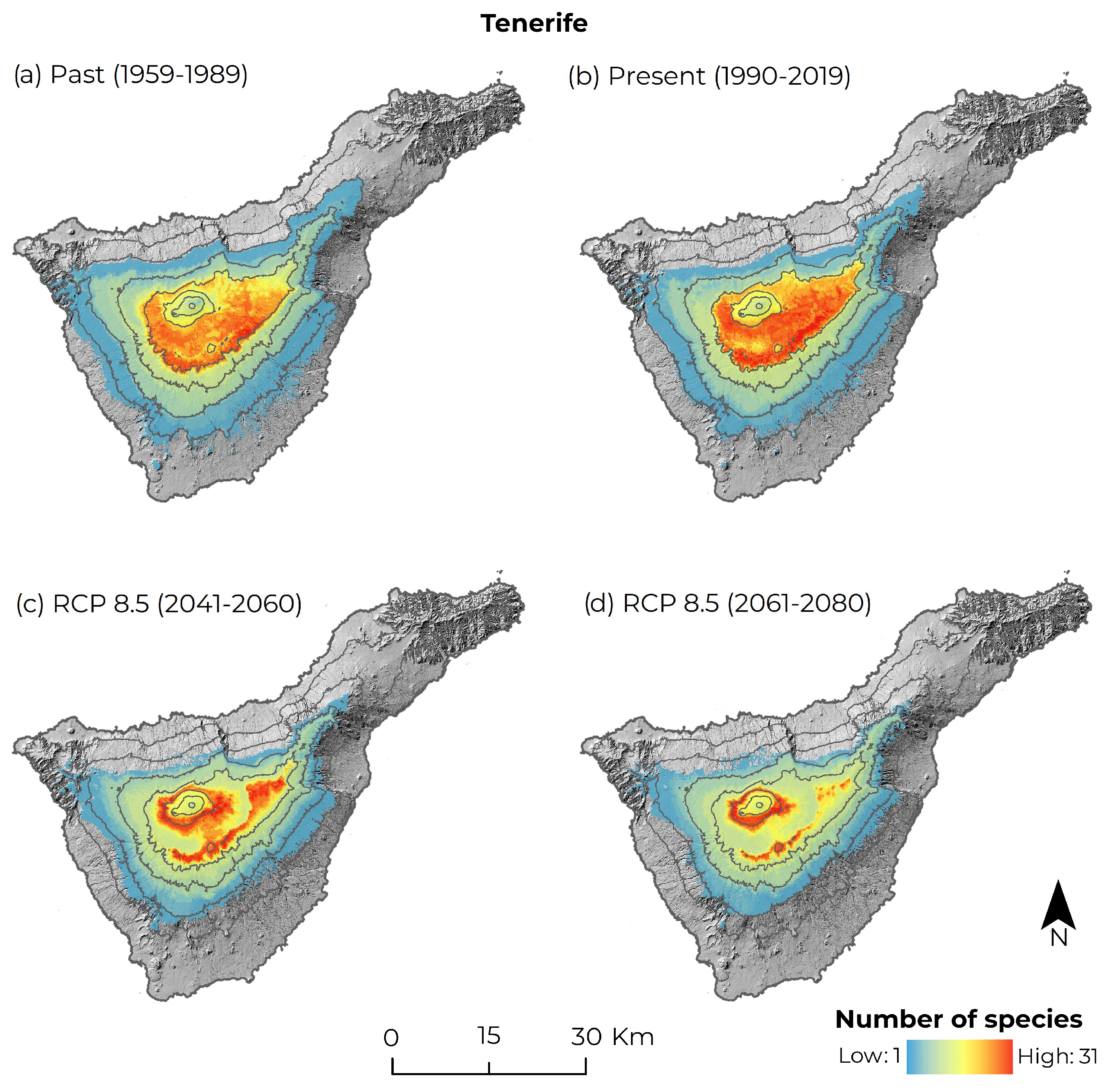
3.1. Changes in Suitable Climate Habitat
3.2. Biotic Velocity of Climate Change
3.3. Geographical Obstacles to Migration
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Island | Endemic | Status | Climatic Habitat (ha and % Change) | Biotic Velocity (m/yr) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| RP | P | 2050 | 2070 | RP-P | P-2050 | P-2070 | ||||
| Adenocarpus viscosus | LP | CAN | NT | 24,177 | 12,044 (−50.2) | 10,632 (−11.7) | 9204 (−23.6) | 29.1 | 6.8 | 9.3 |
| TF | CAN | NT | 57,108 | 52,280 (−8.5) | 47,344 (−9.4) | 43,180 (−17.4) | 15.2 | 13.5 | 12.6 | |
| Andryala pinnatifida ssp. pinnatifida | LP | CAN | NT | 17,661 | 8248 (−53.3) | 11,453 (38.9) | 11,818 (43.3) | 26.7 | 8.6 | 6.7 |
| Andryala pinnatifida subsp. teydea | TF | CAN | NT | 13,972 | 17,104 (22.4) | 11,084 (−35.2) | 6268 (−63.4) | 20 | 13 | 13.6 |
| Argyranthemum haouarytheum | LP | SI | NT | 21,104 | 12,038 (−43) | 10,919 (−9.3) | 9558 (−20.6) | 30.7 | 10.1 | 11.1 |
| Argyranthemum tenerifae | TF | SI | NT | 18,516 | 18,794 (1.5) | 8828 (−53) | 3760 (−80) | 18.9 | 16.4 | 18.6 |
| Arrhenatherum calderae | LP | CAN | NT | 5368 | 2416 (−55) | 1536 (−36.4) | 944 (−60.9) | 90.7 | 10.4 | 9.5 |
| TF | CAN | NT | 18,800 | 18,236 (−3.0) | 9708 (−46.8) | 4524 (−75.2) | 16.5 | 14.2 | 16.2 | |
| Bencomia exstipulata | LP | CAN | T | 8716 | 5128 (−41.2) | 2769 (−46) | 1632 (−68.2) | 31.8 | 13.6 | 14.8 |
| TF | CAN | T | 13,312 | 18,856 (41.6) | 17,152 (−9) | 11,396 (−39.6) | 19.9 | 14.3 | 11.5 | |
| Bethencourtia palmensis | LP | CAN | NT | 13,560 | 6877 (−49.3) | 6828 (−0.7) | 6200 (−9.8) | 36.2 | 7 | 11.9 |
| TF | CAN | NT | 12,902 | 12,288 (−4.8) | 10,301 (−16.2) | 6625 (−46.1) | 12.8 | 11.5 | 11 | |
| Carlina xeranthemoides | TF | SI | NT | 15,332 | 18,898 (23.3) | 13,359 (−29.3) | 8398 (−55.6) | 20 | 11.7 | 13 |
| Cerastium sventenii | LP | CAN | NT | 11,153 | 6092 (−45.4) | 4324 (−29) | 3288 (−46) | 37.6 | 8.5 | 7.8 |
| TF | CAN | NT | 6084 | 6145 (1) | 5369 (−12.6) | 4264 (−30.6) | 12.2 | 9.1 | 10.1 | |
| Cheirolophus teydis | LP | CAN | NT | 6696 | 2660 (−60.3) | 2604 (−2.1) | 2352 (−11.6) | 74.3 | 7.1 | 6.6 |
| TF | CAN | NT | 12,300 | 11,916 (−3.1) | 10,208 (−14.3) | 6691 (−43.9) | 12.7 | 11.7 | 10.3 | |
| Cicer canariense | LP | CAN | T | 21,290 | 19,088 (−10.3) | 14,913 (−21.9) | 12,577 (−34.1) | 17.9 | 11.3 | 9.6 |
| TF | CAN | T | 50,741 | 56,380 (11.1) | 62,620 (11.1) | 59,531 (5.6) | 19.2 | 21.7 | 24.4 | |
| Cistus osbeckiifolius | TF | SI | NT | 25,952 | 28,996 (11.7) | 27,748 (−4.3) | 24,584 (−15.2) | 25.4 | 10.2 | 9.1 |
| Cytisus supranubius | LP | CAN | T | 4292 | 2608 (−39.2) | 1112 (−57.4) | 336 (−87.1) | 52.6 | 10.5 | 15.5 |
| TF | CAN | T | 19,936 | 19,328 (−3) | 10,004 (−48.2) | 4706 (−75.7) | 18.6 | 15.2 | 17.6 | |
| Descurainia bourgeauana | LP | CAN | NT | 3051 | 1633 (−46.5) | 964 (−41) | 634 (−61.2) | 124.8 | 19.9 | 9 |
| TF | CAN | NT | 30,689 | 21,778 (−29) | 9656 (−55.7) | 5820 (−73.3) | 18.6 | 20.3 | 24 | |
| Descurainia gilva | LP | SI | NT | 11,836 | 6268 (−47) | 4340 (−30.8) | 3128 (−50.1) | 24.9 | 20.7 | 14.3 |
| Descurainia gonzalezii | TF | SI | NT | 6168 | 10,494 (70.1) | 12,660 (20.6) | 11,001 (4.8) | 17.2 | 12.1 | 9.6 |
| Echium auberianum | TF | SI | NT | 20,772 | 14,396 (−30.7) | 3560 (−75.3) | 1108 (−92.3) | 13.2 | 30.9 | 38.8 |
| Echium gentianoides | LP | SI | NT | 6109 | 3161 (−48.3) | 1884 (−40.4) | 1112 (−64.8) | 81.5 | 9.9 | 9.7 |
| Echium perezii | LP | SI | NT | 8197 | 4093 (−50.1) | 2892 (−29.3) | 2124 (−48.1) | 89.1 | 9.8 | 8.8 |
| Echium webbii | LP | SI | NT | 23,789 | 14,312 (−39.8) | 15,461 (8) | 14,253 (−0.4) | 27.8 | 9.8 | 8.4 |
| Echium wildpretii | TF | SI | NT | 7356 | 13,016 (76.9) | 11,568 (−11.1) | 7512 (−42.3) | 20 | 12.7 | 11.4 |
| Erigeron calderae | TF | SI | NT | 11,315 | 15,824 (39.9) | 9021 (−43) | 4625 (−70.8) | 20.9 | 13.1 | 16.7 |
| Erysimum scoparium | LP | CAN | NT | 16,262 | 6356 (−60.9) | 7822 (23.1) | 7545 (18.7) | 28 | 6.9 | 6 |
| TF | CAN | NT | 49,424 | 42,738 (−13.5) | 34,323 (−19.7) | 29,073 (−32) | 14.4 | 12.3 | 14.3 | |
| Genista benehoavensis | LP | SI | T | 4708 | 2608 (−44.6) | 912 (−65) | 228 (−91.3) | 63.5 | 11.9 | 15.9 |
| Helianthemum cirae | LP | SI | NT | 5158 | 2176 (−57.8) | 2792 (28.3) | 2728 (25.4) | 23.8 | 6.5 | 6 |
| Helianthemum juliae | TF | SI | T | 12,352 | 15,606 (26.3) | 8780 (−43.7) | 4088 (−73.8) | 19.6 | 13.5 | 14.8 |
| Juniperus cedrus | LP | CAN | T | 24,368 | 22,712 (−6.8) | 19,524 (−14.0) | 14,512 (−36.1) | 31 | 6.7 | 7.9 |
| TF | CAN | T | 21,365 | 20,300 (−5) | 15,714 (−22.6) | 10,593 (−47.8) | 12.7 | 8.5 | 10.3 | |
| Lactuca palmensis | LP | SI | NT | 19,241 | 11,864 (−38.3) | 8481 (−28.5) | 6597 (−44.4) | 27.1 | 10.5 | 10.1 |
| Laphangium teydeum | TF | SI | T | 13,432 | 6714 (−50.0) | 3121 (−53.5) | 2416 (−64.0) | 19.5 | 63.4 | 49.7 |
| Micromeria lachnophylla | TF | SI | NT | 19,239 | 18,544 (−3.6) | 10,226 (−44.9) | 5958 (−67.9) | 16.3 | 13.5 | 14.4 |
| Nepeta teydea | LP | CAN | NT | 2504 | 1464 (−41.5) | 268 (−81.7) | 12 (−99.2) | 21 | 17 | 29.2 |
| TF | CAN | NT | 17,398 | 15,942 (−8.4) | 9244 (−42) | 4452 (−72.1) | 12.3 | 13.7 | 15.5 | |
| Odontites hollianus | LP | CAN | T | 10,526 | 6936 (−34.1) | 4196 (−39.5) | 2618 (−62.3) | 110.8 | 11.2 | 11.9 |
| Pimpinella cumbrae | LP | CAN | NT | 10,562 | 6925 (−34.4) | 4200 (−39.4) | 2600 (−62.5) | 130.5 | 11.2 | 11.5 |
| TF | CAN | NT | 24,220 | 15,964 (−34.1) | 12,836 (−17.1) | 9572 (−40.0) | 25.4 | 11.2 | 12 | |
| Plantago webbii | LP | CAN | NT | 11,896 | 6500 (−45.4) | 5640 (−13.2) | 4677 (−28) | 45.5 | 9.7 | 9.8 |
| TF | CAN | NT | 12,820 | 12,568 (−2.0) | 8756 (−26.7) | 5064 (−59.7) | 13.4 | 12.6 | 11.6 | |
| Pterocephalus lasiospermus | TF | SI | NT | 75,386 | 66,845 (−11.3) | 60,415 (−9.6) | 56,610 (−15.3) | 14 | 10.2 | 11 |
| Pterocephalus porphyranthus | LP | SI | NT | 17,232 | 12,297 (−28.6) | 8404 (−31.7) | 6136 (−50.1) | 15.7 | 8.5 | 9.4 |
| Rhamnus integrifolia | TF | SI | T | 108,686 | 98,346 (−9.5) | 95,913 (−2.5) | 91,077 (−7.4) | 46.9 | 15.7 | 14.9 |
| Rhaponticum canariense | TF | SI | T | 17,937 | 10,397 (−42) | 4408 (−57.6) | 3040 (−70.8) | 11.2 | 21.1 | 35.5 |
| Scrophularia glabrata | LP | CAN | NT | 14,665 | 12,872 (−12.2) | 6577 (−48.9) | 4072 (−68.4) | 11.5 | 13.1 | 14.8 |
| TF | CAN | NT | 18,609 | 18,748 (0.7) | 12,021 (−35.9) | 5992 (−68) | 14.8 | 12.3 | 14.2 | |
| Silene nocteolens | TF | SI | T | 9908 | 4768 (−51.9) | 2560 (−46.3) | 2068 (−56.6) | 19 | 60.2 | 42.9 |
| Silene pogonocalyx | LP | SI | NT | 15,128 | 7505 (−50.4) | 6889 (−8.2) | 5756 (−23.3) | 33.7 | 11 | 9 |
| Teline stenopetala | LP | SI | NT | 19,505 | 10,005 (−48.7) | 8572 (−14.3) | 7092 (−29.1) | 35.2 | 9.7 | 12.1 |
| Tolpis calderae | LP | SI | NT | 16,833 | 12,808 (−23.9) | 7292 (−43.1) | 5489 (−57.1) | 16.5 | 16 | 53.9 |
| Tolpis webbii | TF | SI | NT | 17,057 | 18,254 (7) | 10,332 (−43.4) | 4904 (−73.1) | 18 | 13.8 | 15.7 |
| Viola cheiranthifolia | TF | SI | T | 7022 | 3745 (−46.7) | 2332 (−37.7) | 1860 (−50.3) | 19.6 | 40.8 | 25.3 |
| Viola guaxarensis | TF | SI | T | 7960 | 6152 (−22.7) | 4148 (−32.6) | 3132 (−49.1) | 11.4 | 15.5 | 20.6 |
| Viola palmensis | LP | SI | T | 2988 | 1784 (−40.3) | 648 (−63.7) | 148 (−91.7) | 21.1 | 11.9 | 16.7 |
References
- Evangelista, A.; Frate, L.; Carranza, M.L.; Attorre, F.; Pelino, G.; Stanisci, A. Changes in composition, ecology and structure of high-mountain vegetation: A re-visitation study over 42 years. AoB Plants 2016, 8, plw004. [Google Scholar] [CrossRef] [Green Version]
- Feeley, K.J.; Bravo-Avila, C.; Fadrique, B.; Perez, T.M.; Zuleta, D. Climate-driven changes in the composition of New World plant communities. Nat. Clim. Chang. 2020, 10, 965–970, Erratum in Nat. Clim. Chang. 2020, 10, 1062. [Google Scholar] [CrossRef]
- Palomo, I. Climate change impacts on ecosystem services in high mountain areas: A literature review. Mt. Res. Dev. 2017, 37, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Pereira, H.M.; Leadley, P.W.; Proenca, V.; Alkemade, R.; Scharlemann, J.P.W.; Fernandez-Manjarres, J.F.; Araujo, M.B.; Balvanera, P.; Biggs, R.; Cheung, W.W.L.; et al. Scenarios for global biodiversity in the 21st century. Science 2010, 330, 1496–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.J. Climate-related local extinctions are already widespread among plant and animal species. PLoS Biol. 2016, 14, e2001104. [Google Scholar] [CrossRef] [Green Version]
- Huxley, J.D.; Spasojevic, M.J. Area not geographic isolation mediates biodiversity responses of alpine refugia to climate change. Front. Ecol. Evol. 2021, 9, 633697. [Google Scholar] [CrossRef]
- Lamprecht, A.; Pauli, H.; Fernández Calzado, M.R.; Lorite, J.; Molero Mesa, J.; Steinbauer, K.; Winkler, M. Changes in plant diversity in a water-limited and isolated high-mountain range (Sierra Nevada, Spain). Alp. Bot. 2021, 131, 27–39. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated increase in plant species richness on mountain summits is linked to warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [Green Version]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F.; et al. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [Green Version]
- Nie, X.Q.; Yang, L.C.; Xiong, F.; Li, C.B.; Li, F.; Zhou, G.Y. Aboveground biomass of the alpine shrub ecosystems in Three-River Source Region of the Tibetan Plateau. J. Mt. Sci. 2018, 15, 357–363. [Google Scholar] [CrossRef]
- Nie, X.Q.; Wang, D.; Yang, L.C.; Zhou, G.Y. Controls on variation of soil organic carbon concentration in the shrublands of the north-eastern Tibetan Plateau. Eur. J. Soil. Sci. 2021, 72, 1817–1830. [Google Scholar] [CrossRef]
- Harter, D.E.V.; Irl, S.D.H.; Seo, B.; Steinbauer, M.J.; Gillespie, R.; Triantis, K.A.; Fernandez-Palacios, J.M.; Beierkuhnlein, C. Impacts of global climate change on the floras of oceanic islands—Projections, implications and current knowledge. Perspect. Plant Ecol. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Field, R.; Grytnes, J.A.; Trigas, P.; Ah-Peng, C.; Attorre, F.; Birks, H.J.B.; Borges, P.A.V.; Cardoso, P.; Chou, C.H.; et al. Topography-driven isolation, speciation and a global increase of endemism with elevation. Global Ecol. Biogeogr. 2016, 25, 1097–1107. [Google Scholar] [CrossRef]
- Flantua, S.G.A.; Payne, D.; Borregaard, M.K.; Beierkuhnlein, C.; Steinbauer, M.J.; Dullinger, S.; Essl, F.; Irl, S.D.H.; Kienle, D.; Kreft, H.; et al. Snapshot isolation and isolation history challenge the analogy between mountains and islands used to understand endemism. Global Ecol. Biogeogr. 2020, 29, 1651–1673. [Google Scholar] [CrossRef]
- Caujape-Castells, J.; Tye, A.; Crawford, D.J.; Santos-Guerra, A.; Sakai, A.; Beaver, K.; Lobin, W.; Florens, F.B.V.; Moura, M.; Jardim, R.; et al. Conservation of oceanic island floras: Present and future global challenges. Perspect. Plant Ecol. 2010, 12, 107–129. [Google Scholar] [CrossRef]
- Veron, S.; Mouchet, M.; Govaerts, R.; Haevermans, T.; Pellens, R. Vulnerability to climate change of islands worldwide and its impact on the tree of life. Sci. Rep. 2019, 9, 14471. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.L.; Bethencourt, J.; Cuevas-Agullo, E. Assessment of global warming on the island of Tenerife, Canary Islands (Spain). Trends in minimum, maximum and mean temperatures since 1944. Climatic Chang. 2012, 114, 343–355. [Google Scholar] [CrossRef]
- Del Arco, M.J. La flora y la vegetación canaria ante el cambio climático actual. In Naturaleza Amenazada por los Cambios en el Clima Actas III Semana Científica Telesforo Bravo; Instituto de Estudios Hispánicos de Canarias: Puerto de la Cruz, Spain, 2008; pp. 105–140. [Google Scholar]
- Martín-Esquivel, J.L.; Marrero-Gómez, M.V.; González Mancebo, J.M. Efectos del cambio climático en la vegetación de la alta montaña de Tenerife. Ecosistemas 2021, 30, 2189. [Google Scholar] [CrossRef]
- Bello-Rodriguez, V.; Cubas, J.; Del Arco, M.J.; Martin, J.L.; Gonzalez-Mancebo, J.M. Elevational and structural shifts in the treeline of an oceanic island (Tenerife, Canary Islands) in the context of global warming. Int. J. Appl. Earth Obs. Geoinform. 2019, 82, 101918. [Google Scholar] [CrossRef]
- Renner, S.S.; Otto, R.; Martín-Esquivel, J.L.; Marrero-Gómez, M.V.; Fernández-Palacios, J.M. Vegetation change on Mt. Teide, the Atlantic’s highest volcano, inferred by incorporating the data underlying Humboldt’s Tableau Physique des Iles Canaries. J. Biogeogr. 2023, 50, 251–261. [Google Scholar] [CrossRef]
- Martín-Esquivel, J.L.; Marrero-Gomez, M.; Cubas, J.; Gonzalez-Mancebo, J.M.; Olano, J.M.; del Arco, M. Climate warming and introduced herbivores disrupt alpine plant community of an oceanic island (Tenerife, Canary Islands). Plant Ecol. 2020, 221, 1117–1131. [Google Scholar] [CrossRef]
- Morelli, T.L.; Barrows, C.W.; Ramirez, A.R.; Cartwright, J.M.; Ackerly, D.D.; Eaves, T.D.; Ebersole, J.L.; Krawchuk, M.A.; Letcher, B.H.; Mahalovich, M.F.; et al. Climate-change refugia: Biodiversity in the slow lane. Front. Ecol. Environ. 2020, 18, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Hallfors, M.H.; Aikio, S.; Fronzek, S.; Hellmann, J.J.; Ryttari, T.; Heikkinen, R.K. Assessing the need and potential of assisted migration using species distribution models. Biol. Conserv. 2016, 196, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Baur, B. Dispersal-limited species—A challenge for ecological restoration. Basic. Appl. Ecol. 2014, 15, 559–564. [Google Scholar] [CrossRef]
- Engler, R.; Guisan, A.; Rechsteiner, L. An improved approach for predicting the distribution of rare and endangered species from occurrence and pseudo-absence data. J. Appl. Ecol. 2004, 41, 263–274. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Sousa-Silva, R.; Alves, P.; Honrado, J.; Lomba, A. Improving the assessment and reporting on rare and endangered species through species distribution models. Glob. Ecol. Conserv. 2014, 2, 226–237. [Google Scholar] [CrossRef] [Green Version]
- Burrows, M.T.; Schoeman, D.S.; Richardson, A.J.; Molinos, J.G.; Hoffmann, A.; Buckley, L.B.; Moore, P.J.; Brown, C.J.; Bruno, J.F.; Duarte, C.M.; et al. Geographical limits to species-range shifts are suggested by climate velocity. Nature 2014, 507, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Carroll, C.; Lawler, J.J.; Roberts, D.R.; Hamann, A. Biotic and climatic velocity identify contrasting areas of vulnerability to climate change. PLoS ONE 2015, 10, e0140486. [Google Scholar] [CrossRef] [Green Version]
- Serra-Diaz, J.M.; Franklin, J.; Ninyerola, M.; Davis, F.W.; Syphard, A.D.; Regan, H.M.; Ikegami, M. Bioclimatic velocity: The pace of species exposure to climate change. Divers. Distrib. 2014, 20, 169–180. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 5 June 2022).
- Guijarro, J.A. Homogeneización de Series Climáticas con Climatol. Repositorio AEMET. 2018. Available online: https://repositorio.aemet.es/bitstream/20.500.11765/12185/1/homog_climatol-es.pdf (accessed on 5 June 2023).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Karger, D.N.; Conrad, O.; Bohner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Data Descriptor: Climatologies at high resolution for the earth’s land surface areas. Sci. Data 2017, 4, 170122. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Hamann, A.; Roberts, D.R.; Barber, Q.E.; Carroll, C.; Nielsen, S.E. Velocity of climate change algorithms for guiding conservation and management. Glob. Chang. Biol. 2015, 21, 997–1004. [Google Scholar] [CrossRef]
- Vitasse, Y.; Ursenbacher, S.; Klein, G.; Bohnenstengel, T.; Chittaro, Y.; Delestrade, A.; Monnerat, C.; Rebetez, M.; Rixen, C.; Strebel, N.; et al. Phenological and elevational shifts of plants, animals and fungi under climate change in the European Alps. Biol. Rev. 2021, 96, 1816–1835. [Google Scholar] [CrossRef]
- Otto, R.; Krusi, B.O.; Delgado, J.D.; Fernandez-Palacios, J.M.; Garcia-Del-Rey, E.; Arevalo, J.R. Regeneration niche of the Canarian juniper: The role of adults, shrubs and environmental conditions. Ann. For. Sci. 2010, 67, 709. [Google Scholar] [CrossRef]
- Bello-Rodríguez, V.; Cubas, J.; Del Arco, M.J.; Martín-Esquivel, J.L.; Marrero-Gómez, M.V.; González-Mancebo, J.M. Velocidad de respuesta al cambio climático de las especies del género Viola en la alta montaña de Canarias. Conserv. Veg. 2021, 25, 16–19. [Google Scholar] [CrossRef]
- Rodriguez-Rodriguez, P.; de Castro, A.G.F.; Segui, J.; Traveset, A.; Sosa, P.A. Alpine species in dynamic insular ecosystems through time: Conservation genetics and niche shift estimates of the endemic and vulnerable Viola cheiranthifolia. Ann. Bot. 2019, 123, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Scherrer, D.; Massy, S.; Meier, S.; Vittoz, P.; Guisan, A. Assessing and predicting shifts in mountain forest composition across 25 years of climate change. Divers. Distrib. 2017, 23, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Olano, J.M.; Brito, P.; Gonzalez-Rodriguez, A.M.; Martin-Esquivel, J.L.; Garcia-Hidalgo, M.; Rozas, V. Thirsty peaks: Drought events drive keystone shrub decline in an oceanic island mountain. Biol. Conserv. 2017, 215, 99–106. [Google Scholar] [CrossRef]
- Ferreira, M.T.; Cardoso, P.; Borges, P.A.V.; Gabriel, R.; de Azevedo, E.B.; Elias, R.B. Implications of climate change to the design of protected areas: The case study of small islands (Azores). PLoS ONE 2019, 14, e0218168. [Google Scholar] [CrossRef]
- Del Arco, M.J.A.; Gonzalez-Gonzalez, R.; Garzon-Machado, V.; Pizarro-Hernandez, B. Actual and potential natural vegetation on the Canary Islands and its conservation status. Biodivers. Conserv. 2010, 19, 3089–3140. [Google Scholar] [CrossRef]
- Anderson, A.S.; Reside, A.E.; VanDerWal, J.J.; Shoo, L.P.; Pearson, R.G.; Williams, S.E. Immigrants and refugees: The importance of dispersal in mediating biotic attrition under climate change. Glob. Chang. Biol. 2012, 18, 2126–2134. [Google Scholar] [CrossRef]
- Benito, B.M.; Lorite, J.; Perez-Perez, R.; Gomez-Aparicio, L.; Penas, J. Forecasting plant range collapse in a mediterranean hotspot: When dispersal uncertainties matter. Divers. Distrib. 2014, 20, 72–83. [Google Scholar] [CrossRef] [Green Version]
- Nogales, M.; Rodriguez-Luengo, J.L.; Marrero, P. Ecological effects and distribution of invasive non-native mammals on the Canary Islands. Mammal. Rev. 2006, 36, 49–65. [Google Scholar] [CrossRef]
- Cubas, J.; Martin-Esquivel, J.L.; Nogales, M.; Irl, S.D.H.; Hernandez-Hernandez, R.; Lopez-Darias, M.; Marrero-Gomez, M.; del Arco, M.J.; Gonzalez-Mancebo, J.M. Contrasting effects of invasive rabbits on endemic plants driving vegetation change in a subtropical alpine insular environment. Biol. Invasions 2018, 20, 793–807. [Google Scholar] [CrossRef]
- Cubas, J.; Irl, S.D.H.; Villafuerte, R.; Bello-Rodriguez, V.; Rodriguez-Luengo, J.L.; del Arco, M.; Martin-Esquive, J.L.; Gonzalez-Mancebo, J.M. Endemic plant species are more palatable to introduced herbivores than non-endemics. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190136. [Google Scholar] [CrossRef] [Green Version]
- Ibarrola-Ulzurrun, E.; Marcello, J.; Gonzalo-Martin, C.; Martin-Esquivel, J.L. Temporal dynamic analysis of a mountain ecosystem based on multi-source and multi-scale remote sensing data. Ecosphere 2019, 10, e02708. [Google Scholar] [CrossRef] [Green Version]
- Willis, S.G.; Foden, W.; Baker, D.J.; Belle, E.; Burgess, N.D.; Carr, J.A.; Doswald, N.; Garcia, R.A.; Hartley, A.; Hof, C.; et al. Integrating climate change vulnerability assessments from species distribution models and trait-based approaches. Biol. Conserv. 2015, 190, 167–178. [Google Scholar] [CrossRef]
- Foden, W.B.; Young, B.E.; Akcakaya, H.R.; Garcia, R.A.; Hoffmann, A.A.; Stein, B.A.; Thomas, C.D.; Wheatley, C.J.; Bickford, D.; Carr, J.A.; et al. Climate change vulnerability assessment of species. Wires Clim. Chang. 2019, 10, e551. [Google Scholar] [CrossRef] [Green Version]
- Manes, S.; Costello, M.J.; Beckett, H.; Debnath, A.; Devenish-Nelson, E.; Grey, K.A.; Jenkins, R.; Khan, T.M.; Kiessling, W.; Krause, C.; et al. Endemism increases species’ climate change risk in areas of global biodiversity importance. Biol. Conserv. 2021, 257, 109070. [Google Scholar] [CrossRef]
- Marrero-Gómez, M.V.; Martín Esquivel, J.L.; Docoito Diaz, J.R.; Suárez Izquierdo, M. Viola guaxarensis (Violaceae): A new Viola from Tenerife, Canary Islands, Spain. Willdenowia 2020, 50, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Benito, B.; Lorite, J.; Peñas, J. Simulating potential effects of climatic warming on altitudinal patterns of key species in Mediterranean-alpine ecosystems. Clim. Chang. 2011, 108, 471–483. [Google Scholar] [CrossRef]
- Di Biase, L.; Pace, L.; Mantoni, C.; Fattorini, S. Variations in plant richness, biogeographical composition, and life forms along an elevational gradient in a Mediterranean mountain. Plants 2021, 10, 2090. [Google Scholar] [CrossRef]
- Köhler, L.; Gieger, T.; Leuschner, C. Altitudinal change in soil and foliar nutrient concentrations and in microclimate across the tree line on the subtropical island mountain Mt. Teide (Canary Islands). Flora Morphol. Distrib. Funct. Ecol. 2006, 201, 202–214. [Google Scholar] [CrossRef]




| Change in Species Habitat (%) | Biotic Velocity (m/Year) | ||||||
|---|---|---|---|---|---|---|---|
| Islands and Species Groups | Past | 2041–2060 | 2061–2080 | Past | 2041–2060 | 2061–2080 | |
| Island means | |||||||
| La Palma | −43 (±12) | −25 (±27) | −41 (±34) | 46 (±33) | 11 (±4) | 13 (±9) | |
| Tenerife | −2 (±29) | −29 (±23) | −50 (±27) | 17 (±6) | 18 (±13) | 18 (±10) | |
| La Palma comparisons | |||||||
| Single-island endemic | −43 (±9) | −26 (±26) | −43 (±33) | 38 (±24) | 11 (±4) | 14 (±12) | |
| Canary endemic | −42 (±13) | −24 (±28) | −40 (±35) | 52 (±37) | 11 (±4) | 11 (±5) | |
| Threatened | −36 (±11) | −44 (±19) | −66 (±24) | 47 (±30) | 11 (±2) | 13 (±3) | |
| Non-threatened | −45 (±11) | −19 (±27) | −34 (±33) | 46 (±34) | 11 (±4) | 12 (±10) | |
| Tenerife comparisons | |||||||
| Single-island endemic | −1 (±38) | −33 (±24) | −52 (±27) | 20 (±8) | 22 (±16) | 21 (±12) | |
| Canary endemic | −3 (±17) | −25 (±17) | −48 (±22) | 16 (±4) | 14 (±3) | 14 (±4) | |
| Threatened vs. | −14 (±31) | −30 (±22) | −47 (±25) | 20 (±9) | 26 (±19) | 24 (±13) | |
| Non-threatened | +5 (±28) | −29 (±21) | −51 (±25) | 17 (±4) | 14 (±4) | 15 (±6) | |
| Both islands | |||||||
| Single-island end. vs. | −18 (±36) | −30 (±25) | −48 (±30) | 27 (±19) | 18 (±14) | 19 (±13) | |
| Canary endemic | −24 (±25) | −25 (±24) | −44 (±30) | 35 (±32) | 12 (±4) | 13 (±5) | |
| Threatened vs. | −24 (±27) | −35 (±22) | −53 (±26) | 30 (±24) | 20 (±16) | 20 (±11) | |
| Non-threatened | −19 (±33) | −24 (±26) | −43 (±33) | 32 (±28) | 12 (±4) | 14 (±8) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bello-Rodríguez, V.; Hamann, A.; Martín-Esquivel, J.L.; Cubas, J.; Del Arco, M.J.; González-Mancebo, J.M. Habitat Loss and Biotic Velocity Response to Climate Change for Alpine Plant Species in Atlantic Oceanic Islands. Diversity 2023, 15, 864. https://doi.org/10.3390/d15070864
Bello-Rodríguez V, Hamann A, Martín-Esquivel JL, Cubas J, Del Arco MJ, González-Mancebo JM. Habitat Loss and Biotic Velocity Response to Climate Change for Alpine Plant Species in Atlantic Oceanic Islands. Diversity. 2023; 15(7):864. https://doi.org/10.3390/d15070864
Chicago/Turabian StyleBello-Rodríguez, Víctor, Andreas Hamann, Jose Luis Martín-Esquivel, Jonay Cubas, Marcelino J. Del Arco, and Juana María González-Mancebo. 2023. "Habitat Loss and Biotic Velocity Response to Climate Change for Alpine Plant Species in Atlantic Oceanic Islands" Diversity 15, no. 7: 864. https://doi.org/10.3390/d15070864