1. Introduction
The skates are currently considered to contain about 304 species in two highly diverse (the Arhynchobatidae, or softnose skates, and the Rajidae, or hardnose skates) and two small (the Anacanthobatidae, or legskates, and the Gurgesiellidae, or pygmy skates) families [
1,
2,
3,
4]. The Arhynchobatidae comprise 13 genera with about 109 species, reaching maximum sizes of 28–203 cm in total length (TL) and occurring in 0–3280 m of depth in all oceans, with the center of distribution in polar and cool temperate regions. The family Rajidae comprises 16 genera with about 160 species, attaining maximum sizes of 33–264 cm TL and occurring at 0–4156 m depth in all oceans, with the center of distribution in cold water on continental slopes and abyssal plains. The Anacanthobatidae include five genera with 16 species, reach maximum sizes of 29–75 cm TL and occur in 150–1725 m of depth on continental and insular slopes of the Central-West Atlantic Ocean, Southwest Indian Ocean and Indo-West Pacific Ocean. The Gurgesiellidae contain 3 genera with 19 species, attain 23–59 cm TL and occur in 39–1096 m of depth in warm temperate and tropical seas of the Atlantic, Indian and Pacific Oceans. All data in this paragraph were updated from [
5,
6] while considering [
1,
2,
3,
4]. The genus
Bathyraja Ishiyama, 1958 is by far the most speciose genus of softnose skates, containing 52 valid species [
7]. Additionally, the four valid species currently assigned to
Rhinoraja Ishiyama, 1952 likely belong to the same genus due to a lack of pronounced morphological and molecular differences [
5,
7], with species of
Rhinoraja being deeply nested within the genus
Bathyraja [
7].
An overview of the characteristics and distributions of softnose skates in general has been provided by Last et al. [
2], and for the Southern Ocean by Weigmann et al. [
8]. Currently, six described and one undescribed species belonging to the genus
Bathyraja are considered to occur in the Southern Ocean, representing the only softnose skate genus occurring in this area. The species of
Bathyraja can be found throughout Antarctic continental waters and around sub-Antarctic islands, where they are the dominant rajiform group [
8]. However, the center of distribution of the two closely related deep-water genera
Bathyraja and
Rhinoraja together lies in the northwestern Pacific, where almost half of the species occur [
5]. In general, three morphotypes of shallow-water, transitional and deep-water species have been characterized [
9], the latter so far not having been reported from the Southern Ocean [
8]. The three morphotypes differ in size, squamation, color pattern and depth range [
9]. In the Southern Ocean, species of
Bathyraja range from small (to about 60 cm TL) to rather large (to about 160 cm TL) in size and have been reported from the Ross Sea, around Kerguelen and Heard islands, from the continental margin in the Indian Ocean sector, the Weddell Sea, the Lazarev Sea, around the Antarctic Peninsula, South Shetland and South Orkney islands, and at the deep slope of South Georgia [
8].
Bathyraja arctowskii is the smallest known species of the genus, reaching only about 61 cm total length (TL) and maturing at around 36–50 cm TL [
7]. It is a species of transitional morphotype [
9] that can be further differentiated from its congeners by the, at least partial, dark pigmentation of its mouth cavity and underside of its nasal curtain, which are light in other species [
7].
B. arctowskii is also an exceptional case in skate taxonomy, as it was named by Dollo in 1904 based only on three empty egg cases of tiny size from off the Antarctic Peninsula [
10]. As such, the species remained undescribed for more than 100 years. The reason was the lack of specimens that could be assigned to Dollo’s small egg cases. While examining a large collection of 276 skates and 40 egg cases from Antarctic waters stored in the Zoological Museum Hamburg, Germany (ZMH), SW found an egg case that contained a near-term embryo. A comparison of the embryo with other specimens of different maturity stages and sexes proves their conspecificity [
7]. Previously,
B. arctowskii had been treated as a
nomen dubium, and skates belonging to this species were confused with the much larger
B. griseocauda (Norman, 1937) by Bigelow & Schroeder [
11] and Springer [
12]. Although finally the identity of
B. arctowskii was unambiguously clarified by Stehmann et al. [
7], the syntype egg cases could not be examined at that time due to the COVID-19 pandemic. This has now become possible, and detailed data of the syntypes are provided in the present paper for the first time, together with hitherto unknown details of the tooth, dermal denticle and thorn morphology. Furthermore, a lectotype and two paralectotypes are designated from the syntypes. The detailed comparison of the type egg cases with the egg cases stored in the ZMH (including the aforementioned egg case with a near-term embryo) provides further evidence of their conspecificity.
2. Materials and Methods
One adult male (ZMH 115243, 502 mm TL) and one adult female (ZMH 115243, 510 mm TL) were sampled for teeth by cutting a strip of tissue (with several teeth) from the symphyseal, lateral and commissural regions of the upper (palatoquadrate) and lower jaw (mandible). To obtain individual, clean teeth for photographic documentation, the tissue enclosing the roots was weakened and partly destroyed in a strongly alkalic solution prepared by dissolving 2–3 solid KOH pastilles in ca. 3 mL water (for each strip) in a small glass or PE vessel. The process of weakening the cohesion and releasing the teeth from the tissue was monitored under binoculars and took 45–120 min. Any adhering organic material was removed under the microscope using tweezers and a thin needle. After rinsing in water, the teeth were cleaned in an ultrasonic bath for 10–15 s.
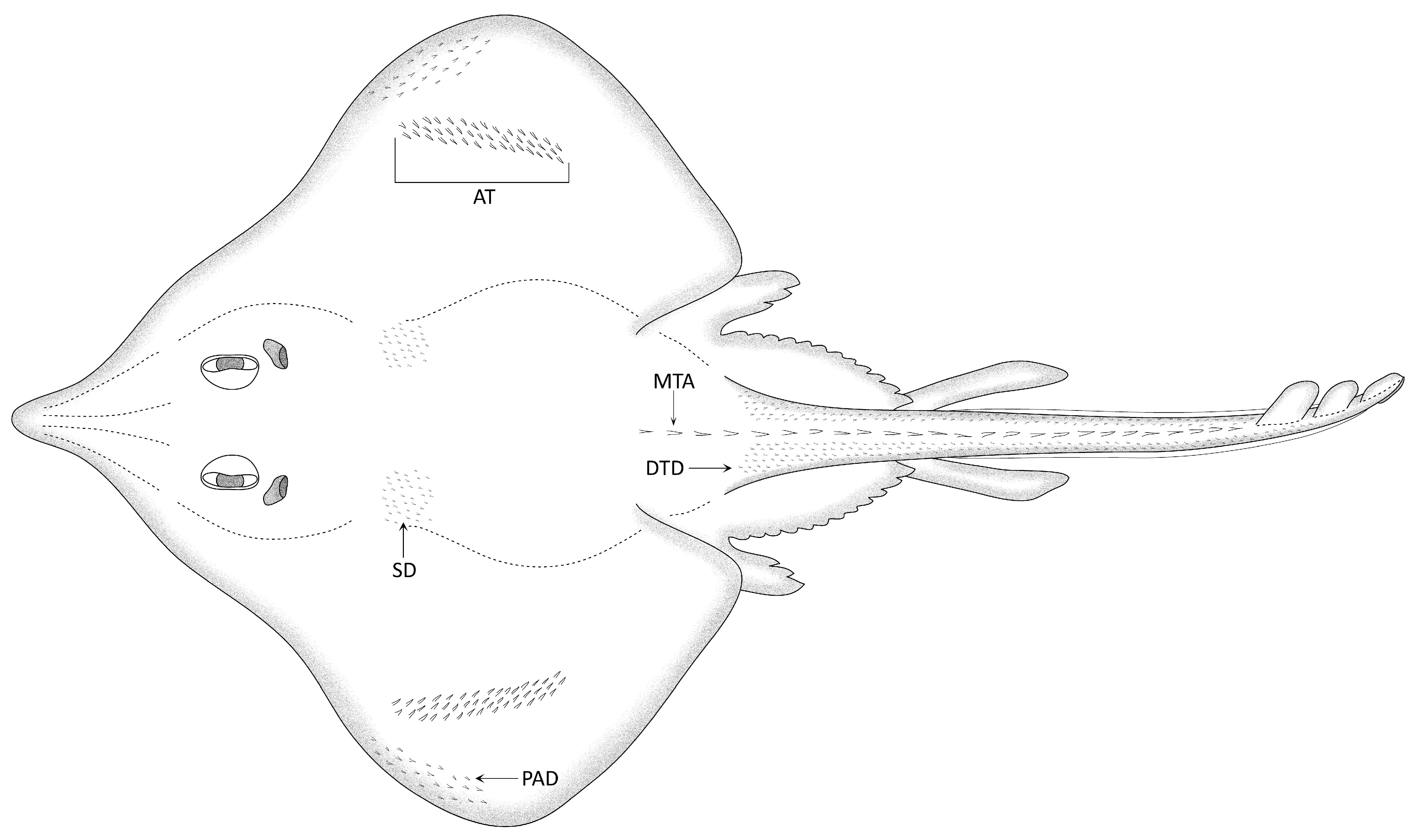
Samples of the squamation were cut from five positions on the dorsal sides, which were equivalent in the female and male individuals but with one additional alar thorn sample in the male. The schematic drawing in
Figure 1 illustrates the terminology and position of the different pattern components of thorns/thornlets and denticles composing the squamation. The process for separating the denticles/thornlets/thorns from the skin was the same as described above for the teeth, but special care had to be taken to avoid damaging the delicate basal root extensions of the denticles.
The dry teeth and denticles/thorns were fixed with colorless fast-drying nail varnish on the tips of metal or glass fixing pins, previously glued in small brass tubes for easier manipulation under the microscope. If the fixation point is chosen correctly, usually the outermost tip of a root lobe, four to five standard directions (occlusal, basal, labial, lingual and profile views) can be photographed in a single pass. Before imaging, the teeth were stained in a solution of red nail varnish in acetone (1:6 to 1:10 ratios), kept in a small glass vessel and protected against evaporation by a ground-in glass lid. This treatment improves the spatial depth and contrast of the images, in particular for smooth or poorly textured crown surfaces occurring in many teeth. The teeth and denticles/thorns were photographed with a Keyence VHX-950F digital microscope, equipped with a VHX-6020 camera, VH-Z20T zoom objective (20–200× magnification) and a strong diffusor.
Descriptions of the methodology used for taking external and skeletal morphometric measurements, meristics and egg case measurements can be found in the study by Stehmann et al. [
7]. The morphological description of the egg cases follows [
7,
13,
14,
15,
16,
17]. The maturity stages are according to [
18,
19]. The terminology of rajoid tooth morphology follows Ward [
20] (text-fig. 1), with additions by Herman et al. [
21] (text- fig. 1). In contrast to Herman et al. [
21] and in agreement with Ward [
20], we prefer the terms “labial” and “lingual” instead of “external” and “internal”, respectively (in conjunction with crown/root surface, view, etc.). Definitions of heterodonty are derived from Cappetta [
22] (p. 13). The terminology of denticle morphology follows Gravendeel et al. [
23], while the terminology of thorn morphology follows Bustamante et al. [
24]. The superordinate term “dermal denticles” is used in our study for all morphological types, independent of their size. Differentiation between denticles, thornlets, and thorns is based on their size, following Bustamante et al. [
24]: denticles are less than about 2 mm in height (in subadults and adults), whereas thornlets are similar in shape but larger and more robust than denticles and usually more than 3 mm high (subadults and adults). Thorns are doubling thornlets in size and height (usually 6–8 mm in height in subadults and adults). Bustamante et al. [
24] reported the size of thorns erroneously as 60–80 mm. Although the median tail thorns of
Bathyraja arctowskii are smaller than the tail thorns of many other
Bathyraja spp., they are still called thorns rather than thornlets due to their equivalence. Institutional acronyms follow Sabaj [
25].
Nomenclatural Acts: the electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature. Hence, the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved, and the associated information viewed through any standard web browser by appending the LSID to the prefix “
https://zoobank.org/”. The LSID for this publication is urn:lsid:zoobank.org:pub:90806841-080A-4F73-B810-8960BCFDF526. The electronic edition of this work was published in a journal with an ISSN and was archived. It is now available from the following digital repositories: CLOCKSS, Swiss National Library (Helveticat).
3. Results
Systematic account
Family Arhynchobatidae Fowler, 1934
Genus Bathyraja Ishiyama, 1958
Bathyraja arctowskii (Dollo, 1904)
English vernacular name: Antarctic dark-mouth skate
Raja arctowskii Dollo, 1904―Poissons in: Expédition Antarctique Belge 1897–99 Zoologie, Antwerpen 1904: 11, 51, 52, plate IX, fig. 10. Types: three empty egg cases IRSNB 25 [orig. 3005], IRSNB 26 [orig. 3006], IRSNB 27 [orig. 3007]; Norman [
26]: 5; Weigmann [
5]: 953; Last et al. [
27]: 17.
Bathyraja arctowskii (Dollo, 1904)―Stehmann et al. [
7]: 1–27; Franzese et al. [
28]: 92, 99, 105.
Bathyraja griseocauda (not Norman, 1937)―Springer [
12]: 5–7, figs. 2 and 4b; Iwami & Abe [
29]: 58, 60, 61.
Bathyraja n. sp. (dwarf)―Hanchet et al. [
30]: 621, 631.
Bathyraja sp. 2―Stehmann [
31]: 209; Stehmann [
9]: 263; Jones et al. [
32]: 53, 54, 63 [erroneously as
Bathydraco sp. 2], 64; Kalisz [
33]: 4–24.
Bathyraja sp.―Stehmann & Bürkel [
34]: 88, 94.
Bathyraja sp. (dwarf)―Smith et al. [
35]: 1170–73, 1175–78.
Breviraja griseocauda (not Norman, 1937)―Bigelow & Schroeder [
11]: R43–R46, fig. 4.
We herein designate syntype IRSNB 25 [orig. 3005], housed in the collection of the Institut royal des Sciences naturelles de Belgique, as the lectotype of Bathyraja arctowskii (Dollo, 1904) and the two other syntypes, IRSNB 26 [orig. 3006] and IRSNB 27 [orig. 3007], as the paralectotypes of B. arctowskii (Dollo, 1904).
Lectotype IRSNB 25 [orig. 3005], 71°19′ S, 87°37′ W, 27–28 May 1898, 435 m depth; paralectotype IRSNB 26 [orig. 3006], 70°23′ S, 82°47′ W, 7–8 October 1898, 400 m depth; paralectotype IRSNB 27 [orig. 3007], 70°15′ S, 84°06′ W, 19–20 December 1898, 569 m depth; ZMH 9014, one empty egg case, 77.6 mm egg case length (ECL), one near-term male embryo, 120 mm TL (removed from egg case, 76.1 mm ECL, also catalogued under ZMH 9014), 61°13.9′ S, 56°25.4′ W, 21.11.96, 403–415 m depth; ZMH 115243 (ex ISH 689-1978), one adult male, 502 mm TL, and one adult female, 510 mm TL, taken together with two juvenile males, 299 and 346 mm TL, one adult males, 520 mm TL, two adult females, 492 and 535 mm TL (all damaged), 60°53′ S, 55°19′ W, 24.01.78, 345–400 m depth; ZMH 121822 (ex ISH 37-1984), one juvenile female, 236 mm TL, and one subadult male, 424 mm TL, taken together with 92 specimens, males 141–432 mm TL, females 135–475 mm TL, 61°11′ S, 56°12′ W, 13.11.83, 375 m depth; ZMH 120216 (ex ISH 489-1981), one adult male, 533 mm TL, taken together with one female, 347 mm TL, one juvenile male, 327 mm TL, 60°51′ S, 55°34′ W, 18.03.81, 280–294 m depth.
Stehmann et al. [
7] already provided a detailed diagnosis and description of
B. arctowskii. An updated diagnosis and description of
B. arctowskii is provided in the present paper, based on the same specimens but considering new information on the lectotype and paralectotype egg cases and the morphology of teeth, dermal denticles and thorns.
3.1. Diagnosis (Updated from Stehmann et al. [7])
Bathyraja arctowskii is the smallest known species of the species-rich genus Bathyraja, attaining only about 61 cm TL based on more than 300 specimens examined. It is characterized by an evenly inverse heart-sharped disc (with distinctly undulated margins in adult males) with broadly rounded outer corners and with body length to mid-cloaca shorter than or equal to tail length from mid-cloaca. Preorbital snout length 8.1–16.2% TL, depending on ontogenetic stage, and distance between first gill slits 14.2–15.9% of TL. Orbits moderately large, horizontal diameter 1.3–1.8 times interorbital width. Jaws weakly angled, with 22–29 upper and 21–29 lower tooth rows, arranged in parallel rows in adult males but in quincunx pavement pattern in females and juvenile males. Sexual dimorphism also apparent in individual teeth, which have a pronounced triangular cusp in adult males but a low cusp in females and juvenile males. Upper side of disc and tail entirely rough prickly with dermal denticles, underside smooth. Individual denticles of star or cross type and mostly with a pronounced, hook-like cusp. Except for non-retractable, hook-like alar thorns of mature males, no thorns on disc but only a median row of 19–30 small thorns along tail to first dorsal fin. The blunt-tipped, circular-based tail thorns become smaller and more widely spaced posteriorly in subadults and adults and are generally wider-spaced in large juveniles to adults as compared to small juveniles. Bases of low, more or less equal-sized dorsal fins confluent or with short interspace. Postdorsal tail section very short, 1.7–5.2% TL, with low epichordal caudal lobe which is confluent with second dorsal fin. Dorsal ground color plain dark to medium grayish-brown, often with mostly indistinct scattered pale and dusky spots on disc, posterior pelvic lobes and on sides of tail, occasionally with transverse white pseudocellus-stripe on inner posterior pectorals. Underside mostly plain white or pale, often with gray marked cloaca and gray spots on belly to gill region, occasionally gray spotted posterior pectoral margins, origin and sides of tail, or tail partly dark. Mouth cavity and underside of nasal curtain at least partly, usually completely pigmented medium to dark grayish from very small juvenile stages onwards. Opened clasper tip shows all components typical for Bathyraja species, of which most apparent a long and deep pseudosiphon (ps) along outer edge of dorsal lobe, as well as a massive projection (pj) over entire length of the inner ventral lobe. Clasper terminal skeleton with distal processes of dorsal marginal cartilage (forming external pseudorhipidion) and ventral marginal cartilage (forming external projection), three dorsal terminal cartilages (with dt1 very large and encapsulating terminal skeleton), ventral terminal and one accessory terminal cartilage. Sexual dimorphism apparent in scapulocoracoid, with post-mesocondyle length longer in females than in males. Both surfaces of blackish-brown egg cases smooth, with coverage of long, fine anchoring fibers. Extension of aprons two-thirds onto horns. Upper side of egg cases strongly convex, lower side moderately convex. Cross-section (anterior and posterior horns) depressed. B. arctowskii differs from its congeners in the small adult size, dark-pigmented mouth cavity and underside of nasal curtain, lack of thorns on upper disc, as well as small and slender egg cases.
3.2. Description
Based on Stehmann et al. [
7], a condensed and updated description of the external and skeletal morphology of
B. arctowskii is provided in the present paper, based on the same specimens as in Stehmann et al. [
7], but considering new information on the morphology of teeth, dermal denticles and thorns. For more details on other descriptive aspects, the reader is referred to the description section in Stehmann et al. [
7]. In addition, detailed descriptions of the lectotype and paralectotype egg cases, as well as ontogenetic changes and sexual dimorphism are given.
3.2.1. External Morphology
Disc evenly inverse heart-shaped, anterior margins not to strongly undulated, outer corners of disc broadly rounded. Disc width 1.2–1.4 times disc length, markedly wide. Axis of maximum disc width at 24.2–31.9% TL, or 54.1–61.0% of disc length, and distinctly posterior to shoulder girdle. Dorsal head length 14.2–21.6% TL. Snout tip somewhat pronounced, narrowly rounded; snout moderately elongated and pointed at 110–140°, with preorbital length 8.1–16.2% TL and 2.2–5.3 times the narrow interorbital width. Orbit horizontal diameter 1.3–1.8 times interorbital width and 1.5–2.0 times length of spiracle depression; interspiracular space wide, 2.2–2.6 times interorbital space; about 10 pseudobranchial folds in each spiracle. Tail slender, gradually tapering towards tip; a low triangle in cross-section, width at level of pelvic tips 1.1–1.8 times height, width at first dorsal-fin origin 1.1–2.6 times height; tail length from mid-cloaca 1.0–1.5 times distance snout tip to mid-cloaca. Both dorsal fins parallelogram-shaped with broadly rounded upper margin and somewhat frayed tip widely overhanging base end; posterior margin strongly inclined forward and concave; both dorsal fins about as long as high, base of first dorsal fin 0.6–1.6 times that of second dorsal fin. Bases of both dorsal fins confluent or with very small interspace. Postdorsal tail short, 47.4–155.6% of second dorsal-fin base length, with a low and indistinct epichordal caudal fold confluent with second dorsal fin, height of epichordal lobe 0 to ~30% of second dorsal-fin height; hypochordal caudal fold absent. Lateral tail folds along posterior two thirds to full tail length.
Ventral head length 25.0–28.0% TL. Preoral snout length 1.5–2.3 times internarial width and 1.2–2.4 times mouth width, the latter 22.9–36.7% of ventral head length and 97.5–129.4% of internarial space; ventral head length 3.2–4.3 times internarial space; distance between fifth gill slits 58.5–81.9% of distance between first gill slits, the latter 2.1–2.4 times internarial space. Anterior nasal flaps rather small and smooth-edged. Outer edges of nasal curtain smooth and not notched, apices rounded, their outer margin smooth; rear margin of curtain weakly fringed by broad and fleshy fringes; isthmus deeply arc-shaped. Anterior and posterior pelvic-fin lobes separated by a deep notch, posterior lobe with angular outer margin and rounded tip 1.1–1.6 times longer than solid anterior lobe tapering to a blunt tip. Fully developed claspers long and slender, with terminal region only somewhat widened; distal half of clasper stem plus terminal region exceeding tips of posterior pelvic lobes; clasper postcloaca length about half of tail length from mid-cloaca.
Jaws weakly angled, with 22–29 upper and 21–29 lower tooth rows. As typical for the genus (and many other rajoid genera), dentitions of B. arctowskii show gradient monognathic heterodonty and lack dignathic heterodonty. Sexual heterodonty is apparent by the presence of a well-developed crown with more or less triangular, pointed cusp in teeth of adult males, whereas teeth of adult females have a rather low crown with lingually displaced cusp. Furthermore, the teeth are arranged in parallel rows in adult males but in quincunx pavement pattern in females and juvenile males. Only teeth of males present ontogenetic heterodonty, bearing a low crown with small cone missing cutting edges in juveniles that grows to a distinct cusp with maturing.
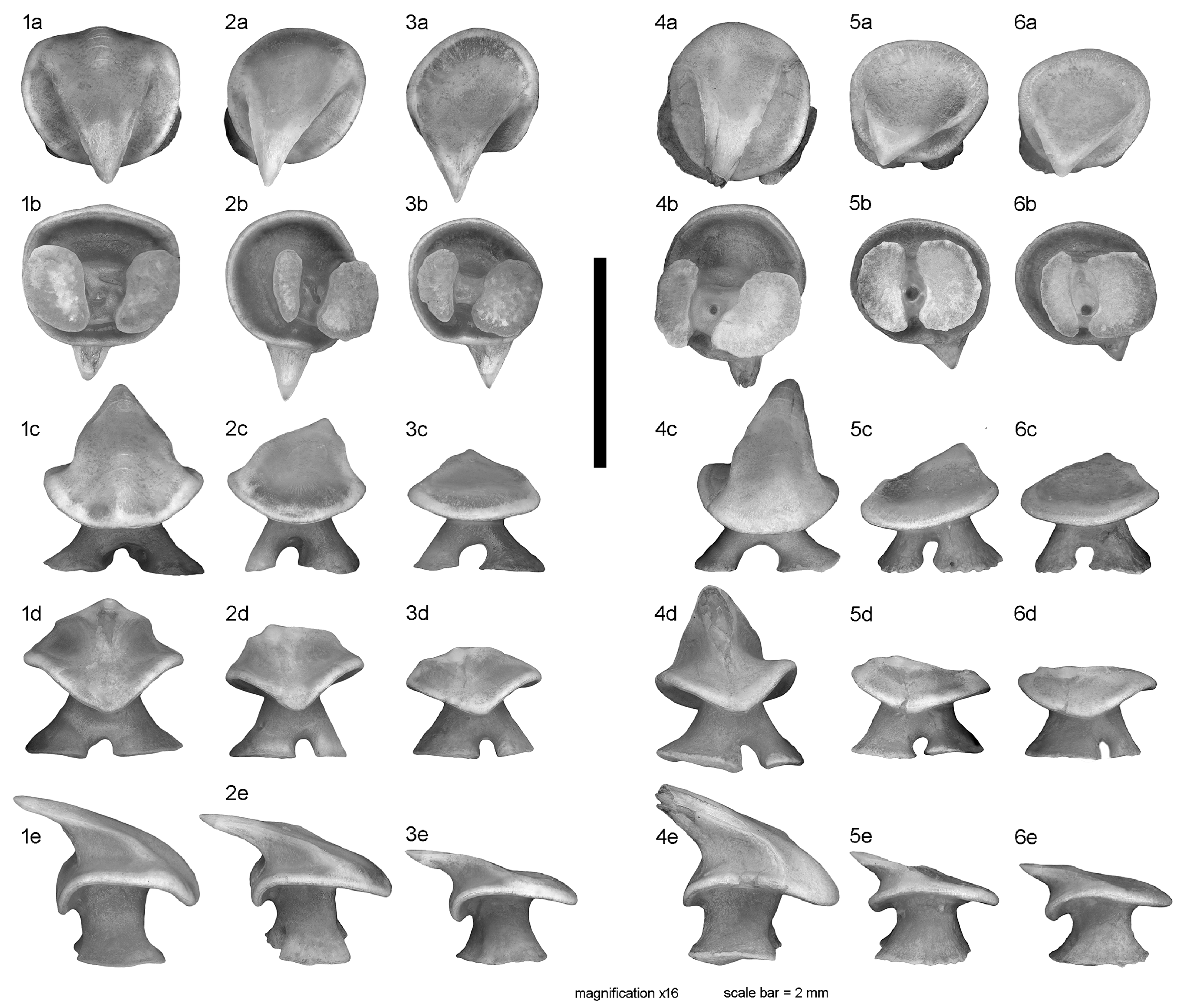
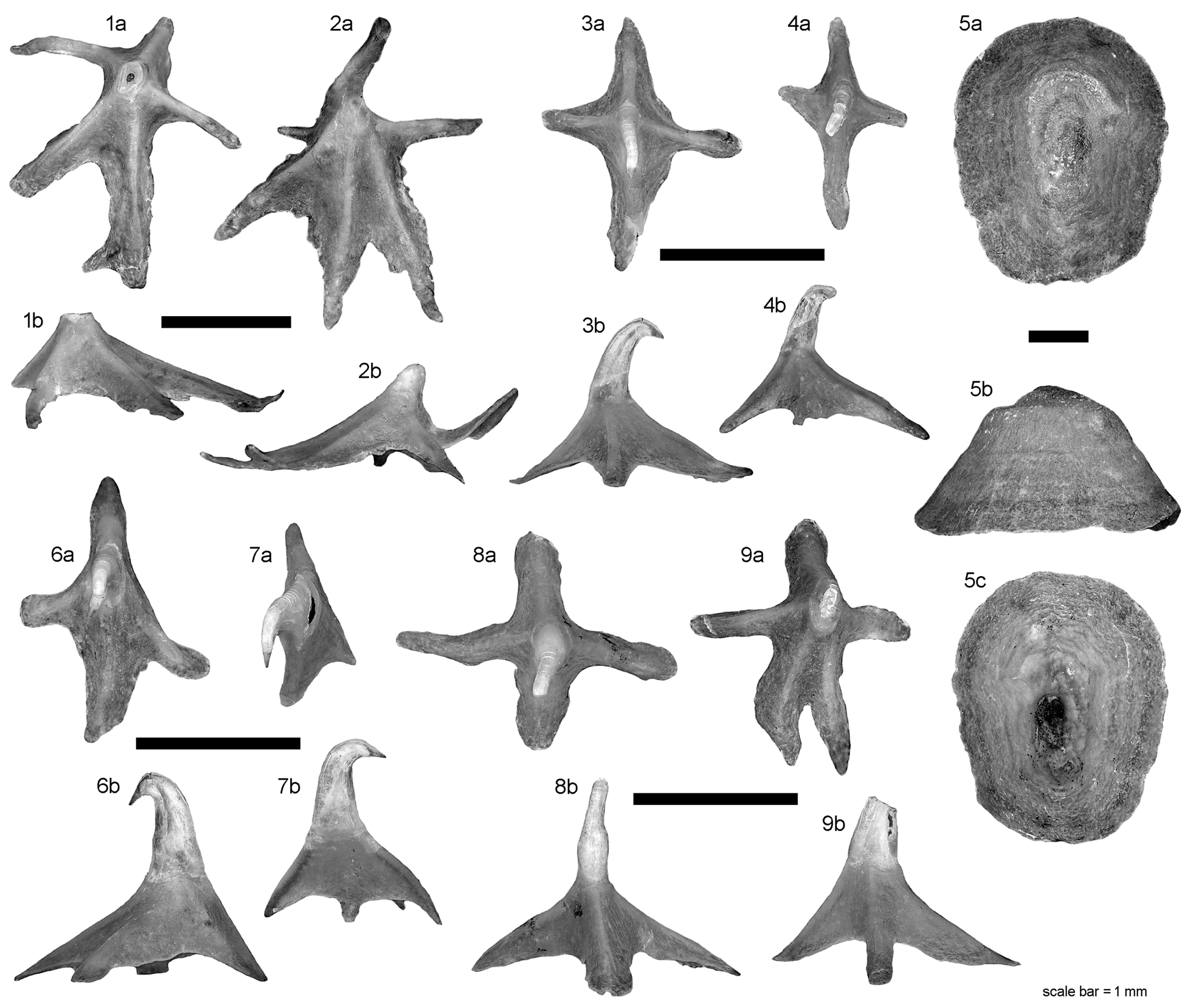
Individual teeth of adult males (
Figure 2) with suboval crown base, being almost as wide as long (in occlusal view), and curved basally at labial and lingual crown margins (in lingual and profile view). Labial visor rather thin (in profile view), with a weak to absent apron. Uvula (= medial lingual crown base) slightly angular and warped in basal direction. Crown of symphyseal/anterior teeth moderately high with symmetrical to oblique cusp, strongly inclined lingually and thereby projecting the lingual crown base (in profile/basal view). Transverse cutting edge of the cusp extending continuously from near the crown base to the crown apex. Labial cutting edge absent. Labial and lingual crown surfaces smooth. Labial surface mesio-distally rather flat, but apico-basally curved in symphyseal to anterior teeth and changing to almost straight in lateral to commissural teeth. Holaulacorhizid root slightly lower (in symphyseal/anterior teeth) or higher than crown (in lateral/commissural teeth). Root stem well developed, oval shaped in cross-section, strongly widening in basal direction and giving way to two, often differently thick root lobes that are separated by a deep median groove with a single large foramen. Basal root surfaces flat to slightly concave.
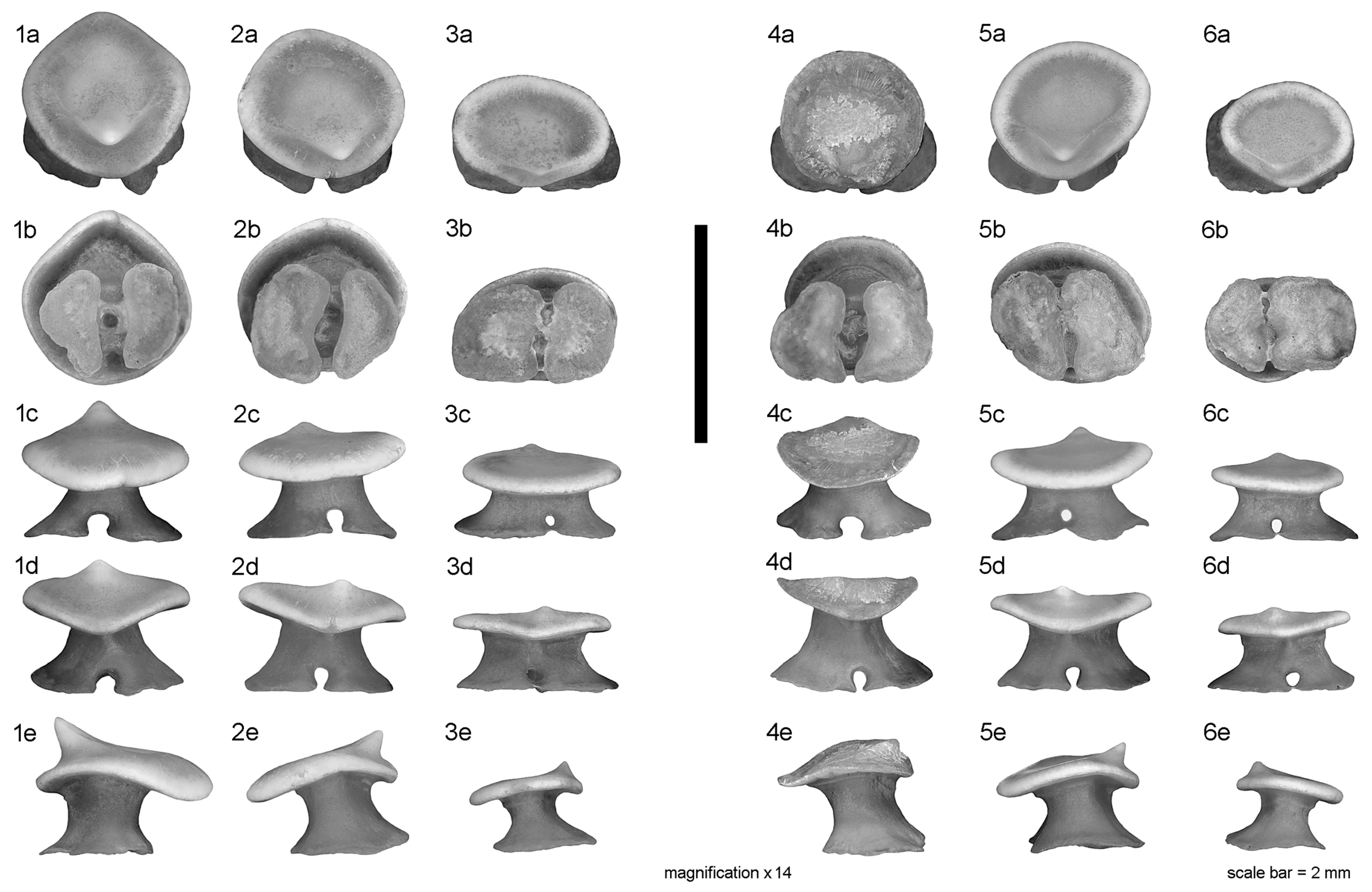
Teeth of adult females (
Figure 3) with sub-circular to oval crown base, nearly as wide as long to wider than long in posterior/commissural teeth (in occlusal view). Crown lower or much lower than root (in profile view). Cusp pointed but low, strongly displaced lingually (in occlusal/profile view), and commonly slightly oblique in distal direction (except in symphyseal/anterior-most teeth). Cusp in profile view lingually directed except in posterior/commissural teeth which have an upright cusp. Uvula, apron and ornamentation absent. Lingual face in profile view concave, labial one flat to slightly convex. Root characteristics very similar to those of male teeth.
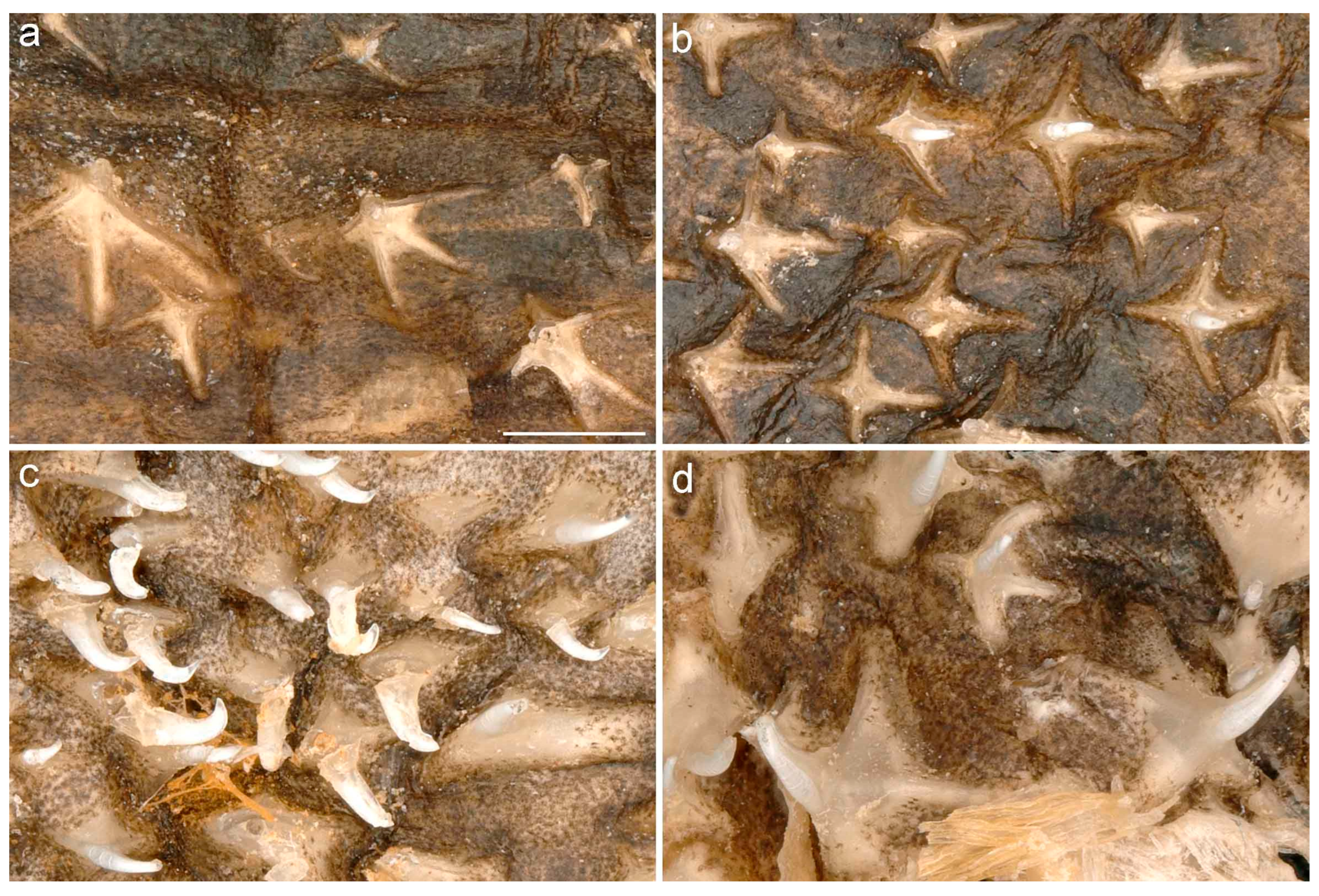
No thorns on upper disc except for very elongated field of alar thorns across outer pectoral corners and parallel to anterior half of posterior margins; alar thorns hook-like and of non-retractable, permanently erect type; alar thorns set in three to five longitudinal and about 20 transverse rows. Tail with 19–30 median thorns from between pelvic and pectoral insertions to somewhat anterior to first dorsal-fin base. The tail thorns become smaller and more widely spaced posteriorly in subadults and adults and are generally wider-spaced in large juveniles to adults as compared to small juveniles. Usually no, occasionally one interdorsal thorn.
Individual tail thorns of males and females, up to ca. 5 mm long, consisting of a basal plate capped with a low enamelled crown which became lost in the specimens illustrated in
Figure 4.5 and
Figure 5.5 (compare with Stehmann et al. [
7]: fig. 13). Outline of well-rounded suboval basal plate longer than wide. Pulp cavity large und ventrally open. Alar thorns of male (
Figure 4.1) up to ca. 8 mm long, strongly asymmetrical in dorsal view and curved along maximum extent. Crown positioned slightly posterior to the center of the basal plate, with posteriorly curved, long and slender cusp lacking barb(s) (fractured in
Figure 4.1). Basal plate anterior to crown longer than posterior part. Posterior part of basal plate unilaterally expanded, with strongly lobed basal lateral outline (
Figure 4.1). Anterior part of basal plate elongated and tubular, tapering at the extremity to a sharp tip. Pulp cavity ventrally open in posterior part of the basal plate only.
The dorsal surface is almost completely and densely set with fine dermal denticles, including the caudal and dorsal fins, as well as partially the integument covering the eyeballs dorsally, but becomes more loosely set on pectoral centers and axils with growth. Posterior pelvic-fin lobes are smooth in most specimens, outer and inner surfaces of the clasper are completely smooth, like also the whole ventral surface.
Denticles in equivalent positions (SD, DTD, PAD in
Figure 1) of males and females generally have similar or even the same morphologies and dimensions. Denticles from all sampled positions (except the alar thorns) are of the “spiny crown” type, whereas the “myrmecoid crown” type is absent (compare with fig. 2 in Gravendeel et al. [
23]). In all denticles, the pulp cavity is ventrally open (
Figure 4.6c–4.9c). Scapular denticles (SD, ca. 0.8–1.5 mm in diameter) typically belong to the “cross type” with four limbs (= “dorsal ridges” of Gravendeel et al. [
23]: fig. 3;
Figure 4.4 and
Figure 5.4) or rarely the “star type” with five limbs of the basal plate. Dorsolateral tail denticles (DTD) tend to be slightly larger in dimensions than SD denticles, and have more variable shapes of the basal plate, ranging from three- to four-limbed (cross type) or to irregular five-limbed shapes (
Figure 4.6–4.9 and
Figure 5.6–5.9). All SD and DTD denticles examined have a dome-shaped basal plate that merges apically into the upright lower crown and the strongly hooked cusp. The enamel-dentine junction typically occurs at the middle of the denticle’s height. In contrast to these, the PAD denticles have a low, five-limbed basal plate of the "leaf-type" or modified "star-type" and a low crown that appears to lack a hooked cusp (
Figure 4.2–4.3 and
Figure 5.1–5.2). However, inspection of the PAD fields in the male shows that a low cusp was present in most of the denticles before abrasion. The thin dentine lamellae between the dorsal ridges may be irregularly widened and then show crenated to lobed edges.
Coloration. Dorsal ground color plain dark to medium greyish-brown, often with mostly indistinct scattered pale and dusky spots on disc, posterior pelvic lobes and on sides of tail, occasionally with transverse white pseudocellus-stripe on inner posterior pectorals. Underside mostly plain white or pale, often with grey marked cloaca and grey spots on belly to gill region, occasionally grey spotted posterior pectoral margins, origin and sides of tail, or tail partly dark.
Clasper external morphology. Clasper stem a solid rod of equal width, with the glans only little widening and short. The most obvious component of the dorsal lobe is a deep and long pseudosiphon (ps) along the outer margin extending over three fourths of glans length. A cleft (cf) is found in distal half of the glans between the axial cartilage ridge and the inner wall of dorsal lobe. A narrow cartilage rod is situated medially in proximal glans, the pseudorhipidion (pr), which covers proximally the axial cartilage (ax). The latter continues visible on deeper level as a ridge from underneath tip of pr to the extreme tip of the glans. Very prominent component on entire length of ventral lobe is the somewhat twisted and long projection (pj), a firm cartilage covered by integument and connected to outer ventral lobe margin with integument. A deep and wide sentina hollow (sn) underneath the pj and the integument spanning to outer ventral lobe margin runs out with its proximal wall as a narrow integument ridge diagonally across the axial cartilage to the proximal end of the cleft. Within the sentina, externally not visible, is the ax-blade-like tip of the sentinel (st) found as a sharp cutting tip edge of the at1 cartilage.
3.2.2. Skeletal Morphology
Clasper skeleton. Clasper terminal skeleton with distal processes of dorsal marginal cartilage (forming external pseudorhipidion) and ventral marginal cartilage (forming external projection), three dorsal terminal cartilages (with dt1 very large and encapsulating terminal skeleton), drop-shaped ventral terminal (not appearing as an external component) and one accessory terminal cartilage.
Cranium. Cranium with short, broadly bullet- or spearhead-shaped anterior fontanelle and elongated, narrowly triangular posterior fontanelle. Ovoid nasal capsules large and strongly forwardly inclined at a 54–75° angle of their straight rear edges to longitudinal axis of cranium. Orbital region evenly deeply concave; otic region broadly bulky, with short occipital condyles and small jugal arches not or hardly exceeding the contour of the occiput rearward or laterally. Snout supported by forward extension of pectoral propterygia and radials extended to nearly snout tip and rostral node. Rostral shaft length 44.5–53.8% of cranium total and 80.9–116.5% of nasobasal length; maximum ethmoidal width 45.1–56.9% of cranium total length, 95.8–103.7% of nasobasal length, 2.9–3.4 times minimum dorsal interorbital width, and 1.7–1.9 times maximum width of otic region. Anterior fontanelle length 18.7–38.9% of rostral shaft and 10.1–17.9% of cranium total length, its length 0.9–1.8 times its maximum width, and its tip hardly extending into rostral shaft length.
Pelvic girdle. Pelvic girdle with massive ischiopubic bar, anterior contour nearly straight, posterior contour slightly to deeply concave; prepelvic processes moderately long, solid, conical, and straight to somewhat inclined inwards, their length 3.2–4.4 times median thickness of ischiopubic bar. Iliac processes massive and curving inwards, with broad, quadrangular tip. Each iliac region with two foramina.
Scapulocoracoid. Scapulocoracoid elongated rectangular in lateral view, with very asymmetrical anterior position of the mesocondyle. Overall shape and number, shape and arrangement of postdorsal and postventral foramina variable, but scapulocoracoid always with a solid horizontal anterior bridge separating the anterior fenestra into a smaller anterior dorsal and an anterior ventral fenestra. Overall shape moderately short to elongated rectangular, with rounded to sharply marked rear corner and nearly straight to deeply concave posterior margin sloping to the metacondyle. Anterior dorsal and ventral fenestrae large, postdorsal fenestrae large or a combination of a large anterior fenestra and several small foramina, postventral fenestrae mostly small and arranged in a chain along the ridge between meso- and metacondyle. Pre-mesocondyle length 25.6–38.0% of maximum length and 35.7–58.7% of post-mesocondyle length. Maximum length 1.3–1.6 times maximum height at scapular process. Height at rear corner 83.3–85.3% of maximum height. Combined height of both anterior fenestrae 37.4–43.3% of maximum height at scapular process and 26.8–28.8% of the element’s maximum length.
Skeletal meristics. Trunk vertebrae (Vtr): 30–37; predorsal tail vertebrae (Vprd): 66–80; total predorsal vertebrae: 97–113; terminal tail vertebrae (Vterm, approximately): 23–40; total vertebrae (Vtotal, approximately): 120–151; pectoral radials, left: 70–89, right: 70–90; pelvic radials, left: 4+16–5+20, right: 4+17–5+19.
3.2.3. Egg Case Morphology
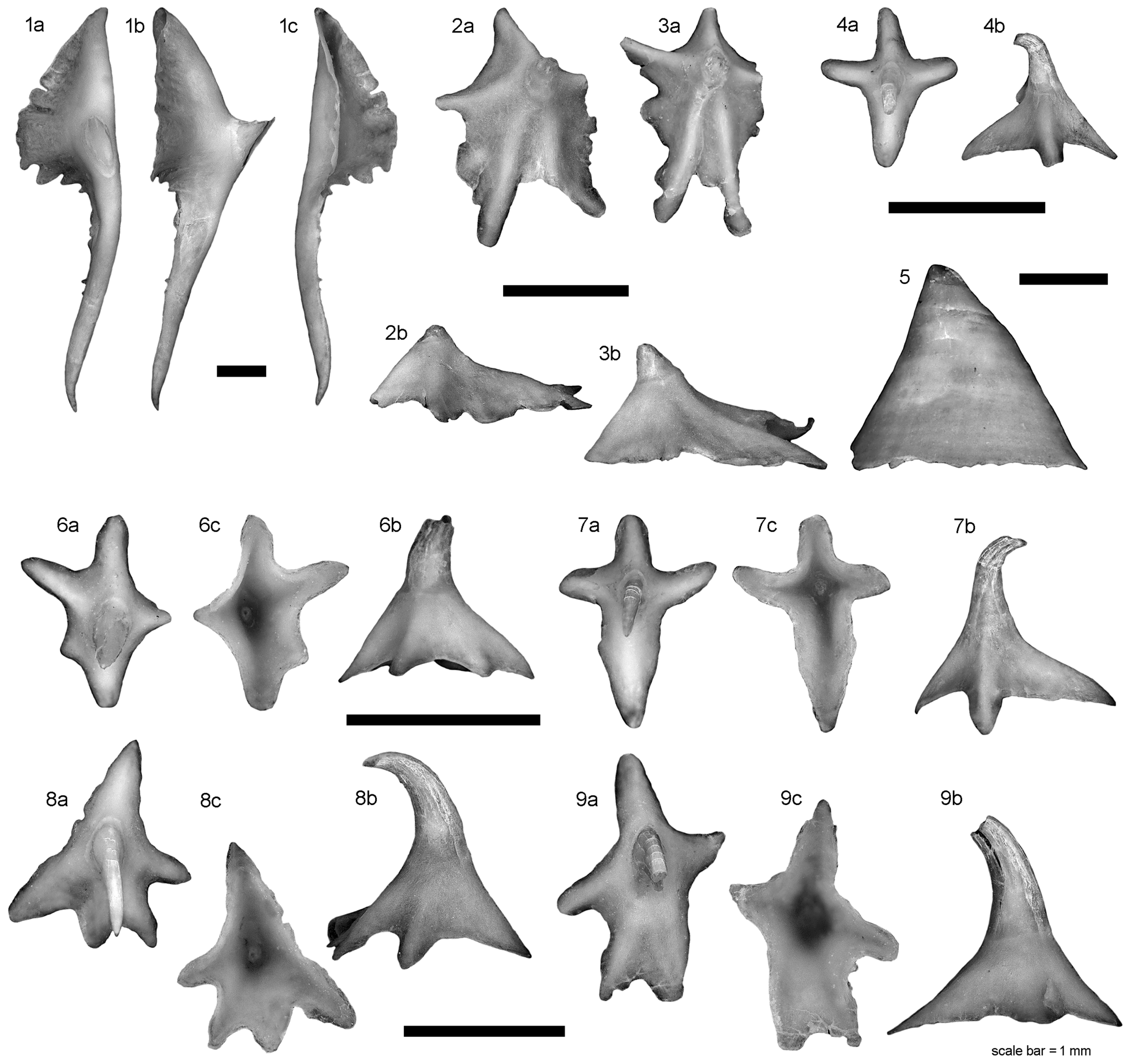
The description of the egg case morphology is based on the lectotype (IRSNB 25 [orig. 3005]) and paralectotypes (IRSNB 26 [orig. 3006] and IRSNB 27 [orig. 3007]), as well as two egg cases catalogued under ZMH 9014).
Figure 6 shows the lectotype and paralectotype egg cases in dorsal and ventral views,
Table 1 provides morphometrics of all five egg cases.
The lectotype and paralectotype egg cases generally correspond well with the two egg cases catalogued under ZMH 9014. Several differences can be ascribed to the generally worse condition of the types compared to the more recent egg cases at ZMH. Such damages are particularly found in the horns, leading to unnaturally small values in several measurements of the types (marked with “>”). Deviations due to the condition of egg cases are most pronounced in paralectotype IRSNB 27, which has the capsule body opened and strongly distorted. Apart from these artificial differences, all five egg cases agree well and only exhibit differences indicative of natural variation. The generally somewhat smaller values found in the type egg cases can be explained by shrinkage caused by the long-term storage of those egg cases. Skate egg cases are generally subject to shrinkage processes, Stehmann & Merrett [
36], e.g., found a shrinkage between ~20–33% when comparing dry and wet egg cases of
B. richardsoni (Garrick, 1961).
The egg cases of B. arctowskii are blackish-brown, the surface is smooth, with coverage of long, fine anchoring fibers. The aprons extend two-thirds onto the horns. The upper side of the egg case is strongly convex, the lower side moderately convex. The cross-section (anterior and posterior horns) is depressed. The length of the egg cases is about 120 mm including the horizontally measured length of the bent horns and ~63–78 mm measured longitudinally between the anterior and posterior apron borders. The latter measurement is likely influenced by shrinkage in the three type egg cases (62.6–67.5 mm) and the fresh measurement is probably closer to the measurements of the two non-type egg cases (76.1–77.6 mm). The maximum case width (MAW), measured as the transverse case width in its lateral plane at its widest part, is ~37.4–42.3 mm and ~54–62% of egg case length (ECL). Again, the values of the type egg cases tend to be smaller possibly due to shrinkage and the measurements of paralectotype IRSNB 27 have been excluded due to its strong distortion. Lateral keels very narrow, ~2–4% of MAW. Anterior apron border broad, deeply concave. Posterior apron nearly straight, broad, and transverse. Anterior border width similar to that of posterior one, but posterior apron ~2.5–3.5 times longer than anterior one. Anterior horns short, their length along the curve (AHL_1) ~44–56% of ECL, curved inwards, tapering towards tips, becoming thin and filamentous. Posterior horns short but ~1.3–1.4 times longer than anterior ones and ~59–64% of ECL, curved inwards, tapering towards tips with tendrils coiling towards filamentous tips.
3.2.4. Ontogenetic Changes and Sexual Dimorphism
Bathyraja arctowskii shows pronounced ontogenetic changes and sexual dimorphism (
Figure 7). As typical for skates, adult males have strongly undulated disc margins showing concavity at anterior sides of snout and at level of nape (vs. not to slightly undulated in females and juvenile males), alar thorns are present, and the teeth have longer cusps and are arranged in parallel rows (vs. quincunx pavement pattern).
Sexual dimorphism is particularly pronounced in the skeleton with the pelvic girdle being narrower with deeply (vs. slightly) concave posterior contour of ischiopubic bar as also typical for skates. Accordingly, the maximum width of the shoulder girdle and the maximum width of the pelvic girdle show sexual dimorphism, with dimorphism of shoulder girdle maximum width evidenced by ratio shoulder girdle maximum width/dorsal interorbital width (4.0–4.5 in males vs. ~5.4 in females) and dimorphism of pelvic girdle shown by ratio maximum width of shoulder girdle/maximum width of pelvic girdle (1.4–1.5 vs. ~1.8). As typical for members of the genus Bathyraja, the degree of intraspecific variation of scapulocoracoid features is very high, referable to sexual dimorphism in the scapulocoracoid’s overall shape and pronounced intraspecific natural variation in the number, shape and arrangement of postdorsal and postventral foramina, the latter probably being influenced by ontogenetic changes. The sexual dimorphism in the scapulocoracoid’s general appearance is reflected in differences in the pre-mesocondyle length (~38.0% of maximum length in males vs. ~25.6% in females and ~58.7% vs. ~35.7% of post-mesocondyle length) and the ratio maximum length/maximum height at scapular process (~1.3 vs. ~1.6).
Apart from these differences, a very high number of ontogenetic changes can be found in
B. arctowskii, including median tail thorns that become smaller and more widely spaced posteriorly in subadults and adults and are generally wider-spaced in large juveniles to adults as compared to small juveniles. Furthermore, the disc of embryos is narrower with disc width only ~51.7% TL (vs. 61.3–66.0% TL), the head is longer in larger specimens (dorsal length 17.6–21.6% TL in subadults and adults vs. 16.8–18.7% TL in juveniles vs. ~14.2% TL in embryos; ventral length 25.0–28.0% TL vs. 21.5–25.9% TL vs. not measurable), the snout is more acutely angled (snout angle 110–120° vs. 125–133° vs. ~140°) and proportionally longer (preorbital length 11.4–16.2% TL vs. 9.6–11.6% TL vs. ~8.1% TL and 3.7–5.3 vs. 2.5–3.6 vs. ~2.2 times interorbital width; preoral length 11.1–15.2% TL vs. 9.0–11.3% TL vs. ~8.8% TL; prenasal length 9.1–12.9% TL vs. 7.4–9.4% TL vs. ~7.4% TL), the eyes are smaller (orbit horizontal diameter 4.1–4.8% TL vs. 4.6–5.8% TL vs. ~5.4% TL; eyeball horizontal diameter 3.4–3.7% TL vs. 3.9–4.6% TL vs. ~4.4% TL), the spiracles are positioned closer together (interspiracular width 7.0–7.9% TL vs. 7.9–8.8% TL vs. ~8.7% TL), the nasal curtain is longer (length 3.4–4.1% TL vs. 2.8–3.4% TL vs. ~2.4% TL) and its lobes are set farther apart (interspace 4.0–5.3% TL vs. 2.8–4.0% TL vs. ~3.3% TL), the tail is wider and lower (tail height at pelvic tips 2.0–2.2% TL vs. 2.3–3.0% TL vs. ~4.3% TL; tail width 1.5–1.8 times vs. 1.1–1.5 times vs. ~1.1 times height at pelvic tips; tail height at first dorsal-fin origin 0.7–0.9% TL vs. 0.9–1.3% TL vs. ~1.3% TL; tail width 1.7–2.6 times vs. 1.1–1.8 times vs. ~1.4 times height at first dorsal-fin origin) and has longer lateral folds (length 44.3–47.7% TL vs. 35.0–47.6% TL vs. ~32.8% TL and along full tail length vs. along posterior two thirds to almost full length vs. along posterior two thirds), the trunk is longer (snout tip to mid-cloaca 45.6–49.6% TL vs. 41.9–45.6% TL vs. ~41.3% TL; snout tip to first hemal spine 48.0–51.5% TL vs. 43.8–48.2% TL vs. not measurable), and the tail is shorter (mid-cloaca to first dorsal-fin origin 38.4–42.8% TL vs. 44.1–46.4% TL vs. ~46.7% TL; mid-cloaca to second dorsal-fin origin 42.5–47.4% TL vs. 48.2–50.4% TL vs. ~52.3% TL; mid-cloaca to tail tip 49.4–52.7% TL vs. 53.2–59.3% TL vs. ~60.2% TL; mid-cloaca to tail tip 1.0–1.2 times vs. 1.2–1.4 times vs. ~1.5 times snout tip to mid-cloaca). Additionally, the dorsal dermal denticle coverage becomes more loosely set on pectoral centers and axils with growth and the dorsal color pattern of light dots and streaks is generally more pronounced in small specimens albeit variable individually. Intraspecific variation in color pattern has been reported regularly within the Arhynchobatidae (see, e.g., Weigmann [
6]).
3.2.5. Size and Maturity
Bathyraja arctowskii is the smallest known species of the genus, attaining about 61 cm TL based on a 61 cm TL specimen from the Ross Sea (NMNZ P.037551), maturing around 36–50 cm TL (based on the largest examined juvenile male of 38.4 cm TL, examined subadult males from 36.1–45.0 cm TL, examined late subadult males from 46.1–49.5 cm TL, and the smallest examined adult male of 49.0 cm TL), hatching at about 12–13 cm TL (based on male embryo ZMH 9014 and the smallest juvenile female examined in detail and catalogued under ZMH 121822), and laying egg cases 6.3–7.8 cm long and 3.7–4.2 cm wide (based on the three types and non-type egg cases examined).
3.2.6. Distribution
Bathyraja arctowskii appears to be a circum-Antarctic species, with its center of distribution in the Atlantic sector of the Southern Ocean. It is known from off the South Shetland, Brabant, and Biscoe Islands to the South-East Weddell Sea, from the Ross Sea in the Pacific sector, and two specimens from Prydz Bay in the Indian Ocean sector of the Southern Ocean. The known depth distribution is 0–1685 m but the species is mainly found between 126 and 810 m depth based on 270 specimens examined in the ZMH collection. Records shallower than that are based only on one lot with six specimens from 0–140 m depth (ZMH 120223) and records deeper than 810 m depth are based solely on 61 specimens from the NMNZ collection. So far, no maturity-related differences in preferred depth areas have been detected, with both juveniles and adults occurring in shallower as well as deeper zones of the known depth range.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}