
Phylogenetic Position of the Genus Manunema (Nematoda, Plectida, Leptolaimidae)

1
Center for Creative Convergence Education, Hanyang University, Seoul 04763, Republic of Korea
2
Research Institute of Natural Science, Hanyang University, Seoul 04763, Republic of Korea
3
Department of Environmental Science, Hanyang University, Seoul 04763, Republic of Korea
4
Department of Invertebrate Zoology, Faculty of Biology, Moscow State University, 119991 Moscow, Russia
*
Author to whom correspondence should be addressed.
Diversity 2023, 15(8), 914; https://doi.org/10.3390/d15080914
Submission received: 30 May 2023
/
Revised: 31 July 2023
/
Accepted: 5 August 2023
/
Published: 7 August 2023
(This article belongs to the Special Issue The Taxonomy, Evolution, and Phylogeography of Marine Invertebrates)
Abstract
:During an investigation of a natural sandy beach on Jeju Island, South Korea, a species of marine nematode belonging to the rare genus Manunema was discovered and is now being reported. Up until now, no molecular sequences of this genus have been available, and its position within the nematode system has been a subject of dispute. Currently, Manunema is classified within the order Plectida, either as the only genus of the family Peresianidae in the superfamily Haliplectoidea or directly as a genus of the family Leptolaimidae. Here, we report on the discovery of Manunema cf. proboscidis Gerlach, 1957, which was found off Jeju Island, South Korea. We also conduct an in-depth investigation of its peculiar morphology, with a particular focus on the reproductive system type observed within the species. Additionally, we examine the phylogenetic relationship of the genus using 18S rRNA sequences. Partial mtCOI, nearly complete sequences of the 18S ribosomal RNA gene as well as a partial 28S (D2–D3 region) gene are provided. Morphologically, the presence of tubular precloacal supplements (shared by superfamily Leptolaimoidea and not Haliplectoidea) and antidromously reflected ovaries of females in Manunema (shared by the order Plectida and not Desmoscolecida) may have taxonomic significance. Our phylogenetic trees based on 18S rRNA also reflected this, including Manunema in a well-supported monophyletic clade of the family Leptolaimidae.
1. Introduction
Species of Manunema are rare marine nematodes distinguished by an unusual appearance, and they currently hold a disputable position in the nematode system. The discovery of Manunema specimens provides an opportunity to enhance our understanding of the anatomy and phylogeny of this taxon. During an investigation of free-living marine nematodes on the intertidal sandy littoral beach of Jeju Island (South Korea), several new nematode species have been discovered and reported to date [1,2,3,4,5]. From the same location, we have obtained several specimens of Manunema proboscidis Gerlach, 1957, and conducted both morphological and molecular analyses.
The establishment of the new genus, along with the description of the first species Manunema proboscidis, was published by Gerlach (1957), based on one male and one female specimen from the Brazilian coast [6]. As the principal genus characters, the anteriormost, very agile, and narrow proboscis-like body region, strong cuticular lining of the posterior muscular pharynx, and precloacal tubular supplementary organs were emphasized. Gerlach assigned the new genus to the family Halaphanolaimidae (now a junior synonym of Leptolaimidae). The second species, under the name Peresiana annulata, was discovered by Vitiello and De Coninck (1968) in the Mediterranean [7]. The authors did not mention Gerlach’s paper, and instead, they associated Peresiana with the family Meyliidae, which was then affiliated with Desmoscolecoidea. To accommodate Peresiana, a monotypic subfamily Peresianinae Vitiello and De Coninck, 1968, within Meyliidae, Desmoscolecoidea, was established. Independently, Andrássy (1973) created a subfamily Manunematinae to accommodate Manunema and Anomonema, Hopper, 1963, within Leptolaimidae [8,9]. Later, Lorenzen (1981) synonymized Manunematinae with Peresianinae and elevated the latter taxon up to the family level [10]. The next significant contribution to our understanding of Manunema was made by Riemann et al. (1971), who conducted a study on both known species newly discovered in the North Sea [11]. They considered Peresiana, Vitiello and De Coninck, 1968, as a junior synonym of Manunema Gerlach, 1957, and suspected a possible identity of M. proboscidis and M. annulatum because the only difference between the two species was the presence or absence, respectively, of precloacal tubular supplementary organs. Furthermore, considering the setae on the socles and the position of the anus on a papilla, Riemann et al. (1971) suggested a possible relation of Manunema to Desmoscolecida and assumed the genus Manunema, along with Anomonema (a genus within Leptolaimidae), and Tricoma mirabilis (previously regarded as an atypical species of the desmoscolecid genus Tricoma but now elevated to its own genus Desmotimmia) serve as intermediate links connecting Leptolaimidae and Desmoscolecidae [11]. Lorenzen (1981, 1994) appears to have been the first to recognize the outstretched ovaries in Manunema, an important feature of the orders Monhysterida, Araeolaimida, and Desmoscolecida [10,12]. However, he classified the family Peresianidae within Leptolaimina, Lorenzen (1981) [10], based on precloacal tubules, a single posterior testis, and the structure of the pharynx. Holovachov and Boström (2004), based on their phylogenetic analysis of a set of morphological data, placed Manunema within the family Leptolaimidae, closely related to Anomonema [13]. The discovery of additional new species made by Stewart and Nicholas (1995) and Barnes and Ferrero (2009) increased the diversity of Manunema up to a total of five species [14,15]. Currently, the genus Manunema is positioned based on the morphological traits within the order Plectida, either as an only genus of the family Peresianidae in the superfamily Haliplectoidea [16,17] or directly as a genus of the family Leptolaimidae [18].
Until now, no molecular sequences have been associated with any reports of Manunema species. Speculations regarding the position of this genus have been solely based on morphological comparisons. Despite the existence of only five valid species within the genus, the position of Manunema (and the validity of the family Peresianidae) has not been firmly established. Manunema remains as one of the very few remaining nematode genera that still lacks any molecular data. With the discovery of M. cf. proboscidis in Korean marine waters, we are conducting a taxonomic review of the genus Manunema, incorporating both morphological and molecular analysis, to discuss and determine its taxonomic position within the phylum Nematoda.
This study aims to provide a morphological and molecular characterization of the species M. cf. proboscidis, by including a description, morphometric measurements, and illustrative depictions of both male and female individuals (including their reproductive organs), as well as the first scanning electron microscopy (SEM) image of the species. Partial and nearly complete sequences of mtCOI, 18S, and 28S rRNA regions are provided, along with a phylogenetic tree constructed using the obtained and databased 18S rRNA sequences, to determine the taxonomic position of the species.
2. Materials and Methods
2.1. Sampling and Morphological Analysis
The nematodes were sampled at Sinyang Beach, a sandy beach located southeast of Jeju Island (33°26′04.4″ N 126°55′23.6″ E), in July 2019. During low tide, qualitative sediment samples of mid and upper horizons (levels) of the intertidal zone were taken with a plastic acryl corer, and the entire sediment samples were fixed in neutralized 5% formol with filtered sea water. Samples were brought back to the laboratory, where meiofauna, including nematodes, were extracted using the Ludox method [19]. Extracted nematodes were postfixed with 70% ethanol stained with rose bengal. Sorted specimens were then transferred to a dish containing a mixture of 5% glycerine and 95% ethanol, and placed in a dry oven maintained at 40 °C. After two to three days of evaporation which left specimens bathed in pure glycerin [20], permanent slides were prepared using beeswax-paraffin rings [21] with glass beads as separators. Specimens were observed, measured, and photographed using an optical microscope, Leica DM5000 (Leica, Wetzlar, Germany), equipped with IC measure v.2.0.0.161 software, and a digital camera, Leica DFC 425C (Leica, Wetzlar, Germany). For scanning electron microscopy (SEM), specimens were dehydrated in a graded series of ethanol–acetone solutions, when critical point dried with carbon dioxide. Prepared specimens were mounted on a stub and were then sputter-coated with gold and examined with a CAMSCan S-2 (Cambridge Instruments, London, UK).
2.2. Molecular Analysis
2.2.1. Sample Preparation and Species Verification
The same locality was revisited in July 2022 to collect fresh specimens for molecular analysis. During low tide, qualitative sediment samples of mid and upper horizons (levels) of the intertidal zone were scraped and transferred to a bucket, then filtered with freshwater to extract meiofaunal specimens via decantation. Samples were immediately fixed in 99% Et-OH (ethanol) and stored in a freezer maintained at −24 °C. Specimens of interest were picked out and sorted into individual wells (of a 24-well dish) containing ultrapure water. Specimens were then transferred to a droplet of ultrapure water on a concave glass slide to make a temporary slide. A cover glass was placed over the droplet containing the specimen, and they were quickly viewed under a stereo microscope at varying magnifications to verify species identification. Upon verification, the temporary slides were disassembled, and the specimens were transferred back to a well containing ultrapure water to initiate the steps for DNA extraction and amplification.
2.2.2. DNA Extraction and Amplification
The specimens were left to soak in a well filled with ultrapure water for 20 min to remove any excess ethanol and debris. The DNA template of each specimen was obtained following the hotshot protocol, as described by Meeker et al., 2007 [22]. Each specimen was picked and transferred to a microcentrifuge tube containing 95 µL of ultrapure water and 5 µL of 1M NaOH. Microcentrifuge tubes were placed in a thermocycler set to run at 95 °C for 20 min and 4 °C for 5 min. Following this step, tubes were centrifuged, and 10 µL of 1M Tris-HCl was added to each tube containing the solution to yield a total of 110 µL of DNA template [22]. The DNA templates were immediately used for DNA amplification or stored for long-term storage in a freezer maintained at −24 °C. For DNA amplification, IP-Taq PCR premix (COSMOgenetech, Seoul, Republic of Korea) was used with 4 µL DNA template, 4 µL ultrapure water, and 1 µL of each primer (0.33–0.62 µM of each primer final concentration, depending on the primer set) for a total reaction volume of 20 µL. The MtCOI gene was amplified using JB3 and JB5 primers, with the amplification protocol set to an initial denaturation at 94 °C for 5 min, 5 cycles of 94 °C for 30 s, 54 °C for 30 s, and 72 °C for 30 s, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 50 °C for 30 s, and an extension at 72 °C for 30 s, followed by a final extension step at 72 °C for 10 min [23]. A partial 18S rRNA gene was amplified using MN18F and 22R primers, with the amplification protocol set to an initial denaturation at 95 °C for 5 min followed by 37 cycles of denaturation at 95 °C for 30 s, annealing at 56 °C for 1 min, and extension at 72 °C for 1 min 30 s, followed by a final extension step at 72 °C for 5 min [24]. Nearly full 18S rRNA genes were attempted for amplification using primers (18S-CL-F, 18S-CL-F3, 18S-CL-R1, 18S-CL-R2, 18S-CL-R7, 530R and 1912R for PCR; 530F, 18S-CL-R2, 1912R, and 530R for sequencing), with the amplification protocol set to an initial denaturation at 95 °C for 3 min, followed by 36 cycles of denaturation at 95 °C for 30 s, annealing at 50 °C for 45 s, and extension at 72 °C for 3 min, followed by a final extension step at 72 °C for 7 min [25,26]. The D2-D3 region was amplified using D2A and D3B primers, with amplification protocol set to an initial denaturation at 95 °C for 5 min, followed by 37 cycles of denaturation at 95 °C for 30 s, annealing at 56 °C for 1 min, and extension at 72 °C for 1 min 30 s, followed by a final extension step at 72 °C for 5 min [27]. Additional primer-related information is provided below (Table 1). Amplification success was confirmed by visualizing the PCR products with 1% Agarose gel electrophoresis. PCR products were sent to BIONICS (Seoul, Korea) to be purified and sequenced. All sequences obtained through this study were uploaded to GenBank (accession number listed in Table 2).
2.2.3. Data Analysis
All obtained sequences were visualized using FinchTV (v. 1.4.0), and low-quality peaks were examined by comparing both forward and reverse strands. Two strands were aligned with ClustalW [29] from MEGA (v. 11.0.13) [30]. Built sequences were subsequently used with the BLAST search [31] from GenBank to verify closely related species. Pairwise distance between its respective mtCOI, 18S, and 28S rRNA sequences was calculated using MEGA 11 with the K2P model [32]. Phylogenetic trees with mtCOI and 28S genes were not constructed, as only a few mtCOI and 28S rRNA sequences of related species were available on GenBank. To construct a phylogenetic tree, relative sequences were retrieved from GenBank and compiled to be aligned using MEGA11 (v. 7.245). Phylogenetic analyses were performed using maximum likelihood (ML) and Bayesian inference (BI) approaches. Of the five 18S rRNA sequences of Manunema obtained from this study, only sequences longer than 900 bp were used to construct a phylogenetic tree to ensure higher resolution. For ML analysis, IQ-TREE (multicore version 2.0.3) and its implemented ModelFinder module were utilized to determine the best-fit model using the Akaike information criterion (AIC). The GTR + F + I + G4 model was selected for constructing a ML tree with 1000 bootstrap replicates using IQTree webservers. For BI analysis, the best model was determined using jModelTest software (v. 2.1.7). The BI tree was constructed using MrBayes software (v. 3.2.6) with the following model parameters: Lset base = (0.2482 0.2240 0.2746 0.2532) nst = 6 rmat = (1.3253 1.8817 1.4838 0.7934 3.5530) rates = gamma shape = 0.4100 ncat = 4 pinvar = 0 with Enoplus sp. (GenBank accession: KR265036) set as an outgroup. Markov chain Monte Carlo (MCMC) was run with the following parameters: ngen = 1,000,000, nchains = 4, samplefreq = 100, savebrlens = yes, and printfreq = 1000. The “sump” and “sumt” commands were used with burnin of 250 to summarize the tree. All generated trees were exported to FigTree (v. 1.4.4) for visualization and adjustments. Resulting trees were visualized using FigTree (v. 1.4.4), and adjustments were made using Adobe photoshop (Adobe Inc., San Jose, CA, USA).
3. Results
3.1. Systematics
Class Chromadorea Inglis, 1983
Order Plectida Gadea, 1973
Family Leptolaimidae Örley, 1880
Subfamily Leptolaiminae Örley, 1880
Genus Manunema Gerlach, 1957
3.2. Diagnosis (Amended and Updated after Gerlach 1957, Vitiello and De Coninck 1968, Lorenzen 1996, and Holovachov 1914)
Leptolaimidae. Anterior body attenuated as a slender proboscis bent ventrally, followed by an inflated region. Cuticle coarsely annulated, with lateral alae absent or present in only one species. Anterior sensilla presented by only four simple cephalic setae without socles. Somatic setae on socles, arranged in two latero-median rows on each side of the body. Buccal cavity tight tubular, smoothly continued to the lumen of the slender anterior pharynx. Posterior pharynx as double muscular bulb with a median constriction and thickened internal lining. Nerve ring at the constriction of the double bulb. Anus on the prominent anal papilla. Secretory–excretory gland not evident. Tail elongated conical, with spinneret and caudal glands. Female reproductive system didelphic, with outstretched or antidromously reflected ovaries. Male reproductive system variable or unclear. Spicules simple and curved. Precloacal tubular supplements one to three in number, or absent.
Type species: Manunema proboscidis Gerlach, 1957.
3.3. Annotated List of Manunema Species
Manunema annulatum (Vitiello and De Coninck, 1968), Riemann, von Thun, and Lorenzen, 1971. Vitiello and De Coninck, 1968: 201–204, figs. 1–4 (males, female, juveniles) (as Peresiana annulata); Mediterranean, French coast, 8–20 m deep, “Amphioxus sand”. Riemann et al., 1971: 148, Abb. 3 (male); North Sea, Römö Island, 16 m deep, coarse sand [7,11].
Manunema kithara Barnes and Ferrero, 2009. Barnes and Ferrero, 2009: 47–49, figs. 1–3 (males, females, juveniles); Arabian (Persian) Gulf, Kuwait Bay and Kazima Bay, intertidal sandy beach [15].
Manunema kuwaitiensis Barnes and Ferrero, 2009. Barnes and Ferrero, 2009: 51–54, figs. 4 and 5 (males, females, juveniles); Arabian (Persian) Gulf, Kuwait coast, 6–20 m deep, subtidal medium sand [15].
Manunema pectenophora Stewart and Nicholas, 1995. Stewart and Nicholas, 1995: 163–169, figs. 1–11 (males, females); Australia, Northern Territory, intertidal sandy beach [14].
Manunema proboscidis Gerlach, 1957. Gerlach, 1957: 451, Abb. 14a–c (male and female); Brazil coast, Espirito Santo, 2 m deep, coarse sand. Riemann et al., 1971: 148, Abb. 1, 2 (male); North Sea, Helgoland and Römö Islands, 16–30 m deep, coarse sand. Tchesunov, 2006, fig. 2.29 (male); White Sea (Northern Russia), Kandalaksha Bay, 12 m deep, coarse sand [6,11,33].
3.4. Description
Manunema cf. proboscidis Gerlach, 1957
Materials:
Thirty specimens collected in July 2019 from the intertidal zone on the coast of Jeju Island, Seopji Beach, South Korea (33°26′04.4″ N 126°55′23.6″ E). Twenty-four adult specimens (nine males and 15 females) were measured. Three female individuals were studied using a scanning electron microscope. Individual morphometric measurements are provided in Tables S1 and S2.
Description:
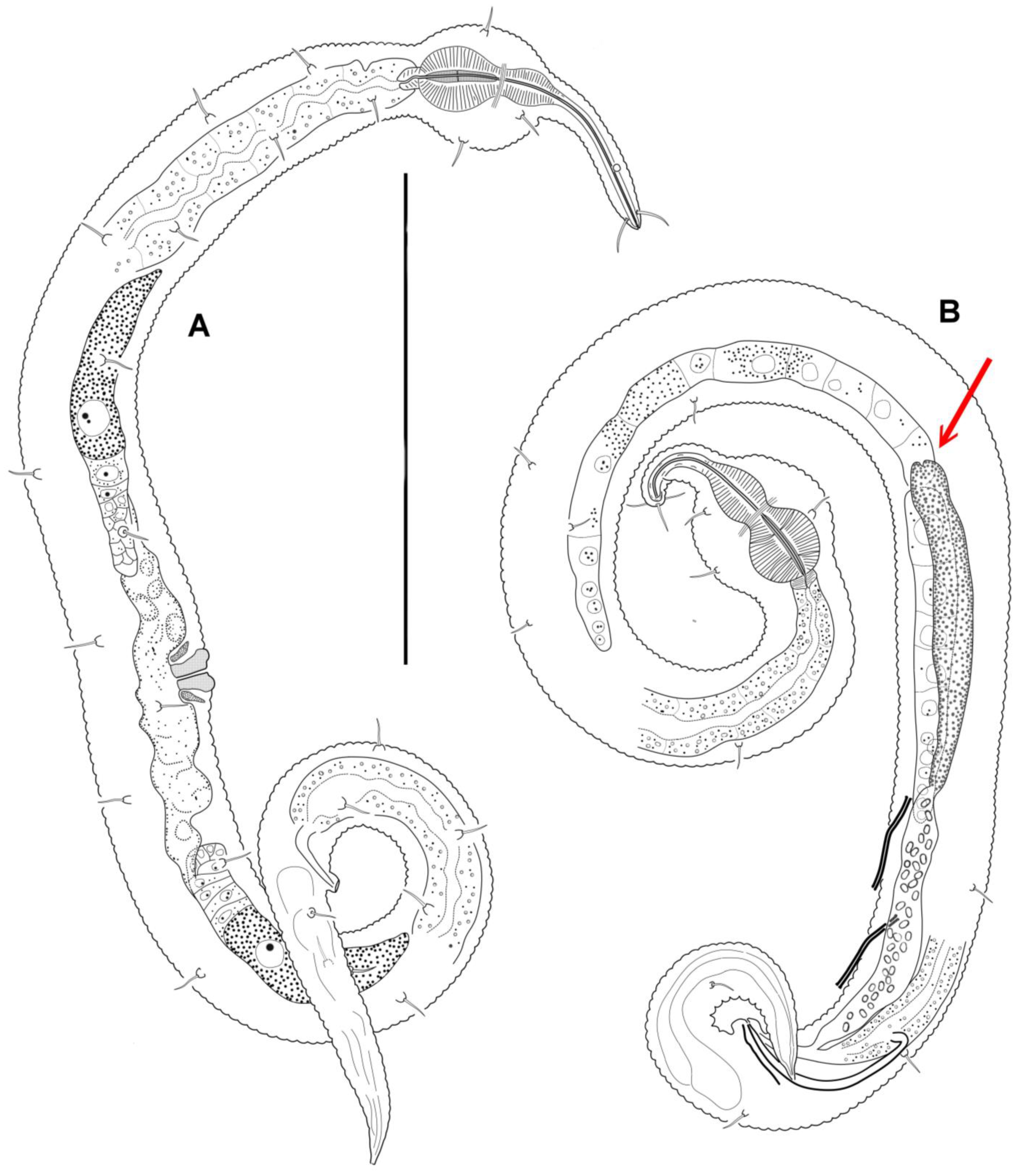
General: Body shape is very peculiar. Anterior half of the pharyngeal region of the body is shaped like a thin and apparently very flexible and mobile proboscis (Figure 1). In all the specimens, the proboscis is tucked under ventrally. The posterior half of the pharyngeal region of the body, on the contrary, is spherically inflated and set off from the rest of the body by a constriction. The body cuticle is distinctly annulated throughout the body, except for the tip of the anterior end and the tail tip, which have a smoothed cuticle; there is no lateral ridge or lateral differentiation. The number of annules within 20 μm in a male is as follows: 13–14 on the proboscis, dorsally 9 and ventrally 19 on the spherical body inflation, 14 on the midbody, dorsally 19 and ventrally 25 on the midtail.
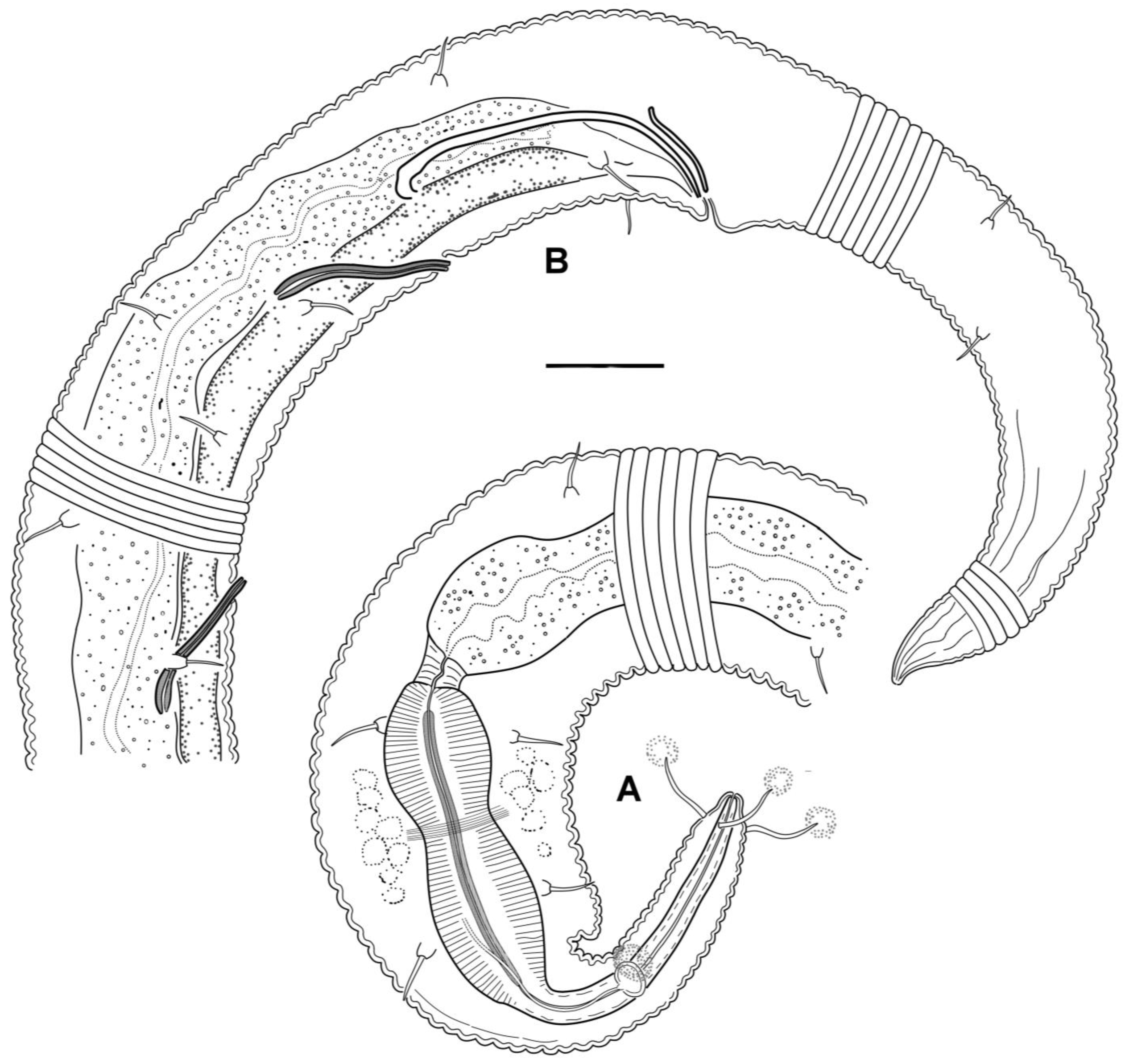
There is no discernable inner or outer labial sensilla. Four cephalic setae appear as thin, rather long setae without basal socles. Cephalic setae are provided with apical pores (Figure 4D), and the cephalic setae of many specimens are marked with clots of secretions issued from the apical pores. Amphideal fovea is situated on the proboscis, about two-thirds of its length from the apex; the fovea is rounded or slightly transversally oval and slightly elevated above the body cuticle surface, with the cuticular rim interrupted posteriorly; the amphideal nerve, which descends from the site of interruption, appears slightly cuticularized and can be followed to the posterior inflated pharyngeal body region. Somatic setae are thin and are inserted on basal socles, 4–7 μm long with socles; the setae are arranged in two rows, with 10–14 latero-dorsal setae and 9–13 latero-ventral, often accompanied by one lateral seta at the anterior edge of the spheric inflation on either side of the body.
The mouth opening is located on the apex of the mouth cone and is very small. The stoma is not well developed. The anterior pharynx within the proboscis region is very slender, lacking obvious radial striations but having a distinct cuticularized inner lumen (Figure 1 and Figure 2A). The posterior pharynx within the spheric body inflation is broad and guitar-shaped, exhibiting radial muscular striations and a thickened internal cuticular lining. The nerve ring is situated at the narrowing (isthmus) in the middle of the posterior muscular pharynx section. The cardia is small and conical. The intestine possesses a distinct internal lumen.
No ventral pore or ventral gland (renette) is found.
The tail is elongated and conical, with a smooth-cuticular tip that is turned to the left. Caudal glands are present within the tail region.
Males: It is difficult to maintain that males are either monorchic or diorchic because of the minute size of the nematodes and the poor visibility of male internal reproductive organs. In some males, a seemingly solitary sausage-shaped gonad without a constriction in the middle may appear to have two germinative zones at opposite ends of the gonad. In other males, a germinative zone is indistinct, either at the posterior end or at both ends of the gonad. The attachment of the vas deferens (gonoduct) to the gonad is also mostly unclear. The only satisfactorily discernible variant depicted (Figure 1B) appears to be diorchic: two testes, with the anterior one outstretched and the posterior one reflexed, and the vas deferens attached at the site where the testes diverge, stretching backward to the left of the posterior gonad. The gonads may be filled with spherical spermatozoa, approximately 1 μm in diameter. Spicules are paired, equal in size, rather weak, and arcuate, with ventrally bent knobs at the anterior end pointed posteriorly. The gubernaculum appears as an indistinct bar and may be indiscernible in some males (Figure 2). There are two subsequent precloacal, straight, rod-like tubular supplementary organs, which exhibit light refraction and are cuticularized. These organs slant anteriorly and bend at the side of the vas deferens. No postcloacal supplementary organs were observed.
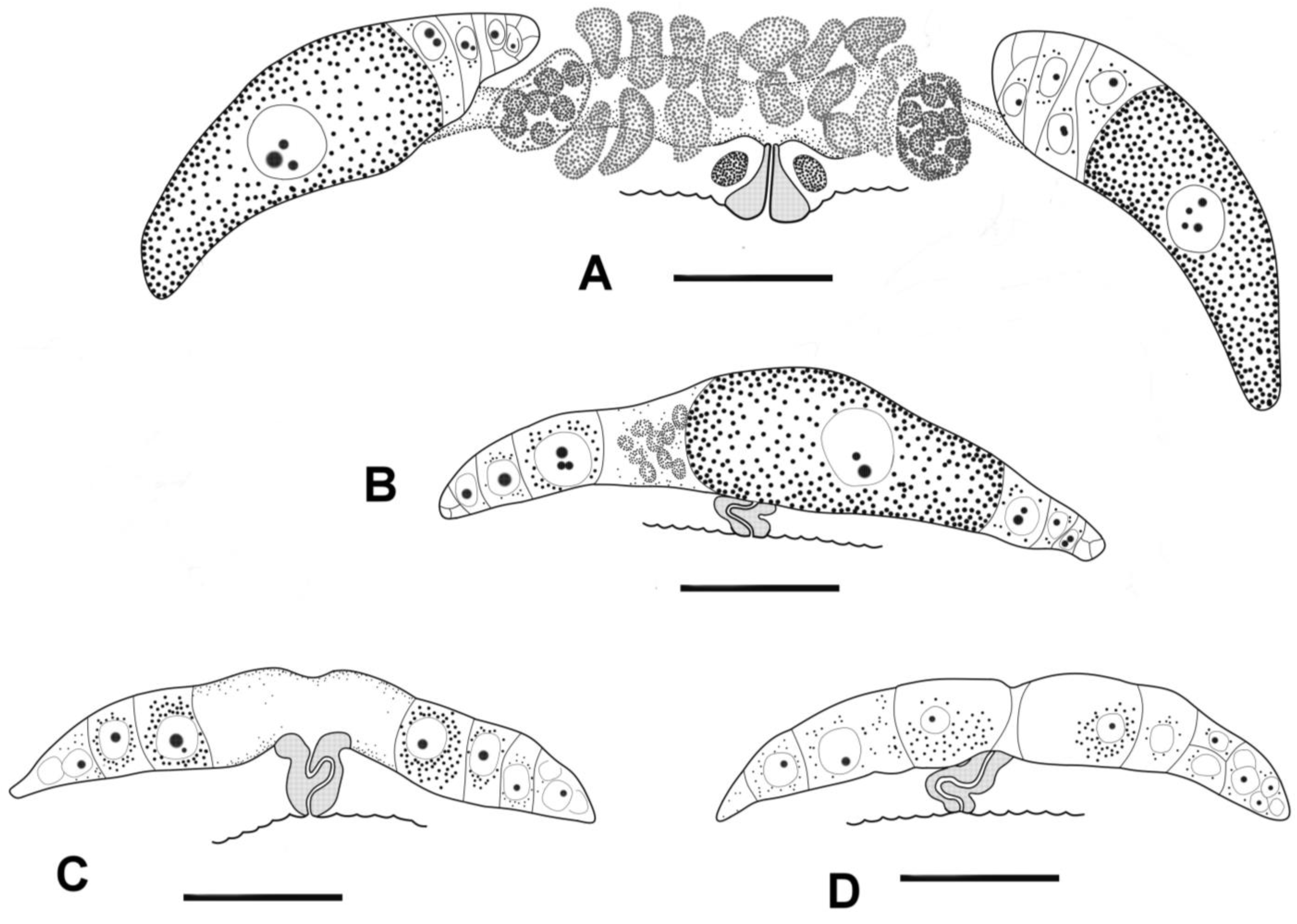
Females: The reproductive system is didelphic. The ovaries are short and may be either antidromously reflexed or outstretched in females of approximately similar body size (Figure 1A and Figure 3). In the case of reflected gonads, the anterior ovary is situated ventrally and to the left of the intestine, while the posterior ovary is situated ventrally and to the right of the intestine. The vagina is short, with thick cuticular walls. In females with reflexed ovaries, the vaginal canal is straight, and the prominent vulva projects slightly beyond the body contour. In the case of outstretched gonads, both ovaries are situated ventrally to the intestine. In females with outstretched ovaries, the vagina appears vestigial, the vaginal canal is strongly convoluted, and the vulva does not project beyond the body contour.
Remarks:
Manunema proboscides was first discovered on the coast of Brazil [6] and then recorded from the North Sea [11]. Despite the great distance and climatic disparity between those two localities, Riemann and co-workers have found neither morphological nor morphometric differences between the two populations and defined them both as the same species. A single male found in the White Sea [33] also corresponds to both descriptions. Our specimens conform to all the cited descriptions, figures, and morphometrics; hence, they are treated here as the same morphospecies, Manunema proboscidis. Due to the morphological and molecular variability revealed within our sample (see also Discussion below), we designate our Manunema population as M. cf. proboscidis instead of M. proboscidis. The term “cf.” (conformis) signifies that assigning the specimens to a specific species is plausible but inauthentic due to limitations in the available material. An important discrepancy is the structure of ovaries and testes, but this is probably a result of new observations and interpretations not interspecific differences. The reproductive system of Manunema is discussed below.
Specimens within our sample have some differences between them, which may exceed individual variation and thus can indicate some heterogeneity and possibly the existence of cryptic species. For instance, the length of precloacal supplementary organs may differ twofold in different males (cf. Figure 1B and Figure 2B, Table S1). It is unknown yet whether the morphometric differences may correspond to the molecular distances between individuals revealed. At the moment, we are not able to elaborate on this issue because the small number of individuals does not allow us to conduct a relevant morphometric analysis.
3.5. Molecular Phylogenetics
3.5.1. Pairwise Distance
Pairwise distances among the five specimens of Korean Manunema cf. proboscidis were zero for the partial 18S rRNA (MN18F/22R region used for comparison) and the 28S large subunit (D2–D3 region) (Tables S3 and S4). The pairwise distance ranged from 2.9% to 13.0% among the five specimens of Korean Manunema cf. proboscidis for the mtCOI region (Table S5). As this is the first sequence of Manunema available, there were no congeners available for comparing interspecific variations. In the absence of congeneric or confamilial sequences, species belonging to the same order, Plectida (e.g., Plectus), were compared. In terms of the 18S rRNA interspecific K2P distance, when compared to species like Amononema, Leptolaimus, and Plectus (belonging to Plectida), Manunema species exhibited a range of 5.1% to 13.9% variation, whereas Desmoscolecids showed a range of 19.9% to 20.8% variation (Table S3). For the 28S rRNA interspecific K2P distance, when compared to other plectids like Haliplectus major and Plectus velox, they differed by 20.3% and 10.3%, respectively (Table S4). Lastly, for the mtCOI interspecific K2P distance, Manunema exhibited a range of 37.5% to 42.9% variation when compared to other plectids such as Deontolaimus and Plectus (Table S5). Two mtCOI sequences of Leptolaimus hormozganensis (GenBank accession: MT001448, MT001449) were available online, but they did not align with our mtCOI sequences and were omitted from further comparative analysis.
3.5.2. Phylogenetic Tree
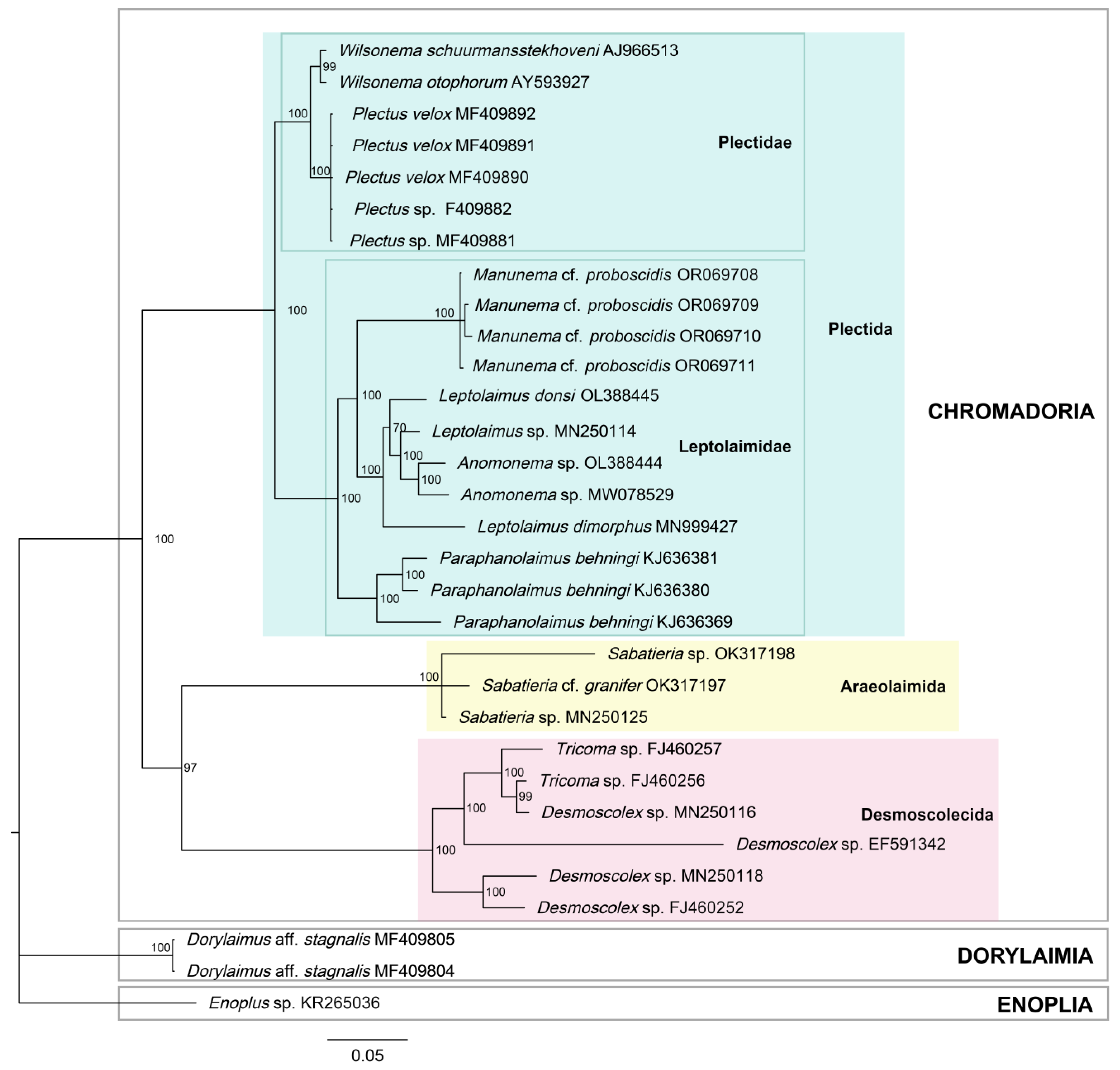
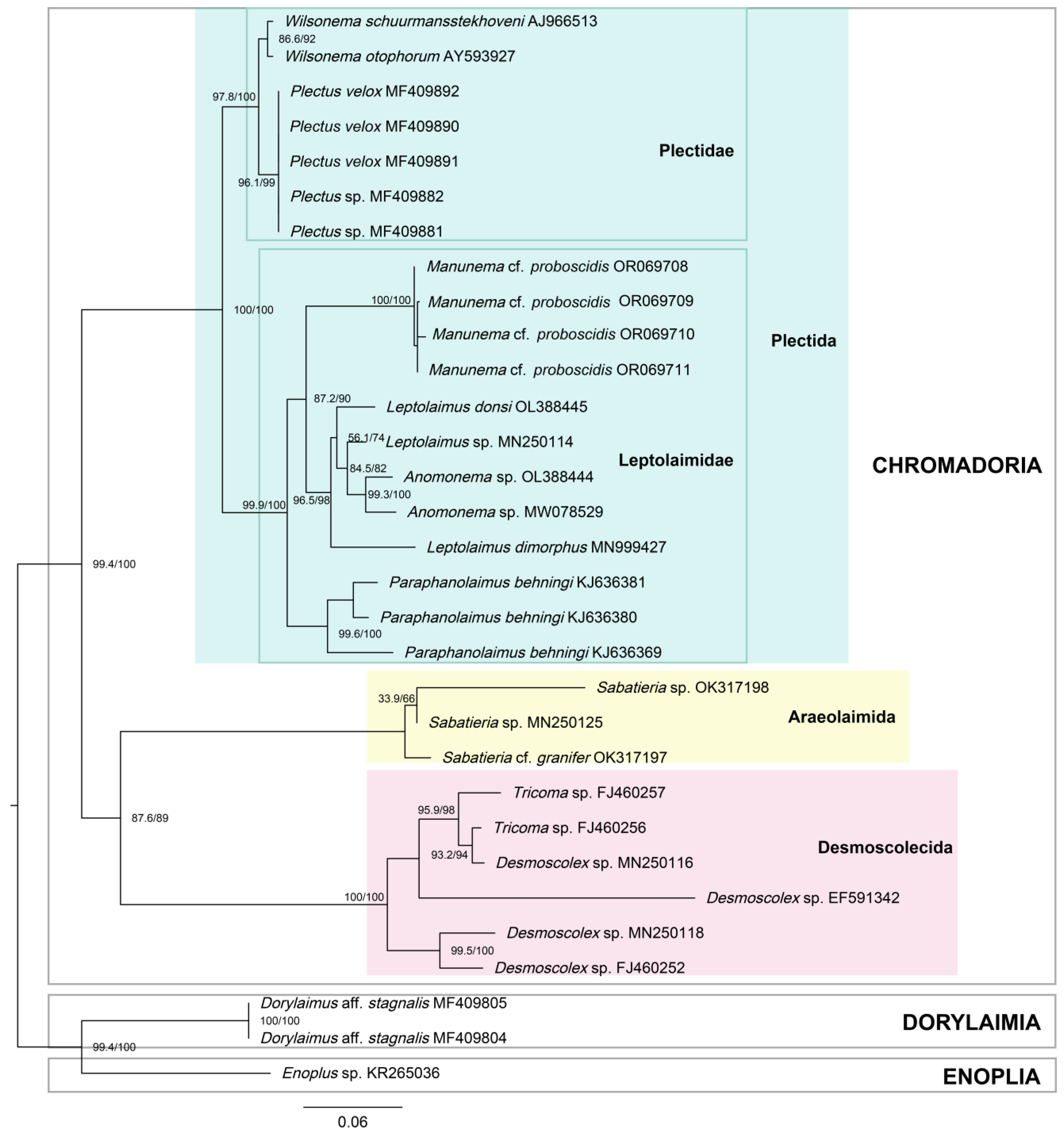
In the Bayesian inference (BI) and maximum likelihood (ML) trees of the 18S rRNA (Figure 5 and Figure 6), the topology was identical between the two, with relatively high support. All branches in the BI tree had posterior probabilities of 97% or 100%, except for the branch including Leptolaimus, which will be discussed later (Figure 5). The ML tree also demonstrated strong support for the Plectida clade (UFboot = 100), as well as families within it (Leptolaimidae and Plectidae with UFboot values of 99.9 and 97.8, respectively) (Figure 6). With Enoplus sp. used as the outgroup and root, species belonging to class Chromadoria formed a monophyletic group. The orders Desmoscolecida and Araeolaimida formed a sister clade, and species belonging to order Plectida also formed a well-supported monophyletic group. Within this clade, the families Plectidae and Leptolaimidae formed sister monophyletic groups. Within Leptolaimidae, Leptolaimus formed a paraphyly with Anonema species. Manunema was placed within the family Leptolaimidae with a posterior probability of 100 (Figure 5).
4. Discussion
4.1. Female Reproductive System
The most striking newly observed feature in our specimen of Manunema cf. proboscidis is the type of female gonads. They are antidromously reflected and not outstretched, contrary to the common statements of Lorenzen (1981, 1994), Tchesunov (2006), and Holovachov (2014) [10,12,34]. In antidromously reflexed ovaries, the entire ovary (germinal and growth zone) bends backwards against the oviduct towards the vulva, and the ova moves to the point of flexure with one pole and to the vulva with the other pole. In outstretched ovaries, the germinal and growth zones are a straight continuation of the oviduct. The type of the ovaries has been considered by Filipjev (1934) [35] and especially by Lorenzen (1978, 1981, 1994) [10,12,34], as a fundamental structural trait characterizing major taxa of free-living, particularly aquatic nematodes. Therefore, Enoplida (clade Enoplia), Dorylaimida and Mononchida (clade Dorylaimia), and Chromadorida, Desmodorida, and Plectida (clade Chromadoria) are characterized by antidromously reflected ovaries, while Monhysterida, Araeolaimida, and Desmoscolecida (clade Chromadoria) have outstretched ovaries. Mostly terrestrial and all parasitic nematodes have a third type, elongated homodromously reflected ovaries adopted for producing large quantities of relatively small eggs. There are quite a few exceptions concerning ovary types in certain free-living orders of Enoplia and Chromadoria (reversing from reflected to outstretched types), which are considered as isolated secondary peculiarities.
In our material, the ovaries of some seemingly adult females of Manunema cf. proboscidis are clearly reflected, while those of others are outstretched (Figure 3). Females with different ovary types may have a similar body size. If the ovaries are outstretched, they are rather short, and the largest and ripest oocyte in one of the two ovaries may exceed the size of another nearest oocyte and be characterized by coarse granulation. The vagina in such females is tightly sinusoidally bent, and the vulva does not protrude beyond the ventral body surface, suggesting it may be non-functional. If reflected, the ovaries turn with the germinal zone towards the vagina, and the largest and ripest oocyte is located at the end of the ovary opposite the vulva. The vagina of such females is straight and perpendicular to the longitudinal body axis, while the vulva bulges beyond the ventral body surface. We interpret these two ovary types in Manunema proboscidis as two stages of ontogenetic development, where the preceding stage of outstretched ovaries transforms into the subsequent stage of antidromously reflected ovaries. Since female specimens with different ovary types do not differ in body size, we assume that this transformation can occur within the same terminal adult stage, after the fourth molting. Our material does not provide a full series of intermediate stages of the ovaries of Manunema from the outstretched to the reflexed phase, probably because the transformation occurs rapidly. A review of the literature reveals that the transformation from the outstretched rudiment to the definitive reflexed state does not occur through bending but rather by the formation of a germinative zone closer to the vagina [36,37,38,39,40,41].
As for published original morphological observations on females, the type of ovaries is indicated either as definitely outstretched (Lorenzen, 1994—Manunema annulatum and M. proboscidis [12]; Stewart and Nicholas, 1995—Manunema pectenophora [14]; Barnes and Ferrero, 2009—Manunema kithara and M. kuwaitiensis [15]), not mentioned, or unclear (Gerlach, 1957—Manunema proboscides [6]; Vitiello and De Coninck, 1968—Manunema annulatum [7]). However, Figure 3 and Figure 5 of Barnes and Ferrero (2009) showing females of M. kithara and M. kuwaitiensis, respectively, allow for another interpretation: The terminal ends of ovaries appear as occupied by the largest granulated oocyte not by a germinative zone [15].
4.2. Male Reproductive System
The construction of male gonads in our specimen of Manunema proboscidis is less understandable than that of female gonads because the male reproductive system is hardly visible in such tiny nematodes. The seemingly clearest variant is depicted in Figure 1B: two opposed testes with terminal germinative zones at both ends and a seminal duct attached at the left at the site of the junction (marked by an arrow in Figure 1B). The testes are not narrowed at the site of the junction. The seminal duct is divided into a granular anterior part and a posterior part containing some number of spermatozoa. The gonads of other males are either torn off and dislodged from the seminal duct or have scarcely discernible germinative ends, making it difficult to qualify them with certainty. In previous taxonomic works, the male reproductive system of Manunema has been described as monorchic, with a single hind reflected testis (Lorenzen, 1994—Manunema annulatum and M. proboscidis [12]), monorchic, with a single headward outstretched testis (Stewart and Nicholas, 1995—Manunema pectenophora [14]) or diorchic (Barnes and Ferrero, 2009—Manunema kithara) and monorchic, with single posterior testis (Barnes and Ferrero, 2009—Manunema kuwaitiensis) [15]. We suspect that this wide range of variations is due to the difficulty of observation rather than actual diversity.
4.3. Phylogeny and Position in the Nematode System
As mentioned above, the main morphological traits that indicate the desmoscolecide relation of Manunema are as follows: (1) setae on socles and (2) outstretched ovaries. In Desmoscolecida, cephalic setae are typically found on socles, while the socles of body setae are not clearly visible or may be concealed by desmal concretions. Manunema lacks socles for cephalic setae, but the body setae are positioned on noticeable socles. The type of ovaries is an important character used to distinguish between major nematode taxa [10,12]. Desmoscolecida is characterized by outstretched ovaries, whereas the order Plectida is characterized by antidromously reflected ovaries. Our discovery of antidromous ovaries in Manunema proboscidis resolves the main objection against classifying Manunema within the order Plectida. Within the Plectida, Manunema shares tubular precloacal supplementary organs with members of the superfamily Leptolaimoidea rather than Haliplectoidea. Among genera of Leptolaimidae, the closest relative to Manunema is the genus Anomonema. Anomonema resembles an elongated and attenuated version of Manunema with a transversally oval amphid, located towards the posterior on the slender neck. The pharynx of Anomonema consists of narrow precorpus, an expanded corpus, a narrowed isthmus with a nerve ring, and a terminal bulb. However, unlike Manunema, Anomonema lacks socled body setae [9,42]. The relationship between Manunema and Anomonema has already been supported on morphological grounds by Riemann et al. (1971), Andrássy (1973), and Stewart and Nicholas (1995) [8,11,14]. However, both known species of Anomonema have only been described based on males, and therefore, the female reproductive system of this genus is unknown.
Our morphological data align well with the molecular genetic data in determining the position of Manunema within the family Leptolaimidae, order Plectida, closely related to the genus Anomonema. Both our BI and ML trees of 18S rRNA (Figure 5 and Figure 6) include Manunema as a constituent of the monophyletic group formed around the family Leptolaimidae, in close relation to Leptolaimus and Anomonema. This finding is consistent with the observations of Holovachov (2006), who synonymized Peresianidae with Leptolaimidae after observing a closer alliance to Leptolaimus in his phylogenetic analysis [43]. Holovachov (2006) noted the structural similarities in the digestive system, reproductive system, and peculiar structure of tubular supplements between Manunema and Leptolaimus. Our findings strongly indicate that Manunema rightfully belongs to the family Leptolaimidae. It is still insufficient to determine whether the genus Manunema serves as an intermediate link between leptolaimid and desmoscolecids. To address this, a more comprehensive accumulation of molecular data is required for in-depth comparison.
Among the three molecular markers utilized in this study, 18S rRNA had the most available resources. Often, there was a lack of comparable sequences for mtCOI and 28S rRNA, which limited the extraction of meaningful findings. In terms of K2P distances, 18S and 28S rRNA showed no intraspecific differences, while mtCOI exhibited variation ranging from 2.9% to 13.0%. The usual K2P threshold to discern interspecies differences for mtCOI genes is >5% [23,44], which our Manunema specimens seemed to exceed intraspecifically. Morphological observation and morphometric comparison may also allude to the existence of cryptospecies within our sample. This intraspecific distance observed among Manunema specimens found in Korea could indicate a cryptic species complex, consisting of one or more nearly morphologically identical species. Although it is true that mtCOI is often preferred for discerning closely related or cryptic species, due to its relatively short and fast-evolving genes, it is still common practice to compare different marker regions. This is because some taxa tend to show higher variability in certain regions than others, making them less suitable for species identification. The 18S region (MN18F/22R) and 28S region (D2A/D3B) are widely used for rapid species identification of nematodes. Considering that the 18S and 28S regions of Manunema showed zero intraspecific distance, it is more likely that the mtCOI region of this group is simply more variable than other regions. This is further supported by the high interspecific distance between Manunema and other plectids, with approximately a 40% difference (Table S5). Since there are currently no congener sequences available to establish a suitable threshold for determining interspecies/cryptic species, relying solely on the genetic distance of mtCOI is inadequate to determine the cryptic nature of Manunema. It will require additional molecular data from closely related species to further investigate this matter.
From this study, we were able to determine that Manunema belongs to the family Leptolaimidae based on both morphological and molecular evidence. The presence of tubular precloacal supplements in males and antidromously reflected ovaries in females appears to be taxonomically significant for Manunema species. Although our molecular analyses were based on a limited number of sequences, our phylogenetic tree, constructed using the 18S rRNA sequences, placed Manunema specimens within a well-supported clade of the family Leptolaimidae. While the intraspecific distance of mtCOI suggests high variability and a possibly cryptic nature of the species, further accumulation of molecular data is necessary to resolve this issue.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15080914/s1, Table S1. Morphometric measurements of individual male specimens observed; Table S2. Morphometric measurements of individual female specimens observed; Table S3. Kimura 2-parameter distance between closely related species based on 18S rRNA alignment with 1000 bootstrap. Standard deviation marked in blue; Table S4. Kimura 2-parameter distance between closely related species based on 28S rRNA alignment with 1000 bootstrap. Standard deviation marked in blue, Table S5. Kimura 2-parameter distance between closely related species based on mtCOI alignment with 1000 bootstrap. Standard deviation marked in blue.
Author Contributions
Conceptualization, R.J. and A.V.T.; methodology, R.J. and A.V.T.; software, R.J.; validation, R.J. and A.V.T.; formal analysis, R.J. and A.V.T.; investigation, R.J. and A.V.T.; resources, R.J. and A.V.T.; data curation, R.J. and A.V.T.; writing—original draft preparation, R.J. and A.V.T.; writing—review and editing, R.J. and A.V.T.; visualization, R.J. and A.V.T.; supervision, A.V.T.; project administration, R.J. and A.V.T.; funding acquisition, R.J. and A.V.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was prepared within the framework of the state assignment of M.V. Lomonosov Moscow State University No. 121032300117-3 and supported by a grant (NRF-2021R1I1A1A01040377) from the National Research Foundation of Korea, and a grant (NIBR 202102108) from the National Institute of Biological Sciences (NIBR) funded by the Ministry of Environment (MOE) of the Republic of Korea.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The voucher specimens examined within this study were deposited in the National Institute of Biological Resources (NIBR), Korea. Partial sequences of mtCOI, 18S, and 28S rRNA were deposited in GenBank. The GenBank accession numbers are listed in Table 2.
Acknowledgments
We would like to thank Wonchoel Lee for providing access to materials required for the study. We are grateful to all the reviewers whose valuable remarks allowed us to find some mistakes and improve the paper.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Jeong, R.; Tchesunov, A.V.; Lee, W. Two species of Thoracostomopsidae (Nematoda: Enoplida) from Jeju Island, South Korea. PeerJ 2020, 8, e9037. [Google Scholar] [CrossRef]
- Jeong, R.; Tchesunov, A.V.; Lee, W. Bibliographic revision of Mesacanthion Filipjev, 1927 (Nematoda: Thoracostomopsidae) with description of a new species from Jeju Island, South Korea. PeerJ 2019, 7, e8023. [Google Scholar] [CrossRef]
- Tchesunov, A.; Jeong, R.; Lee, W. Two New Marine Free-Living Nematodes from Jeju Island Together with a Review of the Genus Gammanema Cobb 1920 (Nematoda, Chromadorida, Selachinematidae). Diversity 2020, 12, 19. [Google Scholar] [CrossRef] [Green Version]
- Tchesunov, A.V.; Jeong, R.; Lee, W. Onyx disparamphis sp. n. (Nematoda, Desmodorida) from South Korea with a taxonomic review of the genus. PeerJ 2022, 10, e13010. [Google Scholar] [CrossRef] [PubMed]
- Tchesunov, A.V.; Jeong, R.; Lee, W. A new genus and species of the family Microlaimidae (Nematoda: Chromadorea) from intertidal sand of the Jeju Island, South Korea. Zootaxa 2021, 5020, 130–140. [Google Scholar] [CrossRef]
- Gerlach, S.A. Die Nematodenfauna des Sandstrandes an der Küste von Mittelbrasilien (Brasilianische Meeres-Nematoden IV). Mitteilungen Aus Dem Zool. Mus. Berl. 1957, 33, 411–459. [Google Scholar] [CrossRef]
- Vitiello, P.; De Coninck, L. Peresiana annulata n. gen., n. sp., type intéressant de Desmoscolecida. Rapp. Comm. Int. Mer Médit. 1968, 19, 201–204. [Google Scholar]
- Andrássy, I. Nematoden aus Strand- und Hohlenbiotopen von Kuba. Acta Zool. Acad. Sci. Hung. 1973, 19, 233–270. [Google Scholar]
- Hopper, B.E. Marine nematodes from the coast line of the Gulf of Mexico. III. Additional species from Gulf shores, Alabama. Can. J. Zool. 1963, 41, 841–863. [Google Scholar] [CrossRef]
- Lorenzen, S. Entwurf eines phylogenetischen Systems der freilebenden Nematoden. In Veröffentlichungen des Instituts für Meeresforschung in Bremerhaven; Kommissionsverlag F. Leuwer: Bremerhaven, Germany, 1981; Volume 7, 472p. [Google Scholar]
- Riemann, F.; von Thun, W.; Lorenzen, S. Über den phylogenetischen Zusammenhang zwischen Desmoscolecidae und Leptolaimidae (freilebende Nematoden). In Veröffentlichungen des Instituts für Meeresforschung Bremerhaven; Kommissionsverlag F. Leuwer: Bremerhaven, Germany, 1971; Volume 13, pp. 147–152. [Google Scholar]
- Lorenzen, S. The Phylogenetic Systematics of Freeliving Nematodes (Translation of the 1981 Paper); Ray Society: Andover, UK, 1994; Volume 162, pp. 1–383. [Google Scholar]
- Holovachov, O.; Boström, S. Morphology and systematics of the superfamilies Leptolaimoidea Örley, 1880 and Camacolaimoidea Micoletzky, 1824 (Nematoda: Plectida). J. Nematode Morphol. Syst. 2004, 7, 1–49. [Google Scholar]
- Stewart, A.C.; Nicholas, W.L. Manunema pectenophora sp. nov. (Peresianidae, Leptolaimina), a nematode possessing unusual male supplementary organs. Trans. R. Soc. S. Aust. 1995, 119, 163–169. [Google Scholar]
- Barnes, N.; Ferrero, T.J. Two new species of Manunema (Plectida: Peresianidae) from the Arabian Gulf, with notes on the phylogeny of the genus. Zootaxa 2009, 2053, 43–58. [Google Scholar] [CrossRef]
- De Ley, P.; Blaxter, M.L. A new system for Nematoda: Combining morphological characters with molecular trees, and translating clades into ranks and taxa. In Proceedings of the Fourth International Congress of Nematology, Tenerife, Spain, 8–13 June 2002; Brill: Leiden, The Netherlands, 2004; Volume 2, pp. 633–653. [Google Scholar]
- Hodda, M. Phylum Nematoda: A classification, catalogue and index of valid genera, with a census of valid species. Zootaxa 2022, 5114, 1–289. [Google Scholar] [CrossRef]
- Holovachov, O. 7.16 Order Plectida Gadea, 1973. In Handbook of Zoology Gastrotricha, Cycloneuralia and Gnathifera. Volume 2: Nematoda; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2014; pp. 487–535. [Google Scholar] [CrossRef]
- Burgess, B. An improved protocol for separating meiofauna from sediments using colloidal silica sols. Mar. Ecol. Prog. Ser. 2001, 214, 161–165. [Google Scholar] [CrossRef] [Green Version]
- Seinhorst, J. A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 1959, 4, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Hopper, D. Drawing and Measuring Nematodes; Ministry of Agriculture, Fisheries and Food: Her Majesty’s Stationery Office: London, UK, 1970.
- Meeker, N.D.; Hutchinson, S.A.; Ho, L.; Trede, N.S. Method for isolation of PCR-ready genomic DNA from zebrafish tissues. Biotechniques 2007, 43, 610–614. [Google Scholar] [CrossRef]
- Derycke, S.; Vanaverbeke, J.; Rigaux, A.; Backeljau, T.; Moens, T. Exploring the use of cytochrome oxidase c subunit 1 (COI) for DNA barcoding of free-living marine nematodes. PLoS ONE 2010, 5, e13716. [Google Scholar] [CrossRef] [Green Version]
- Bhadury, P.; Austen, M.C.; Bilton, D.T.; Lambshead, P.J.D.; Rogers, A.D.; Smerdon, G.R. Development and evaluation of a DNA-barcoding approach for the rapid identification of nematodes. Mar. Ecol. Prog. Ser. 2006, 320, 1–9. [Google Scholar] [CrossRef]
- Carta, L.K.; Li, S. Improved 18S small subunit rDNA primers for problematic nematode amplification. J. Nematol. 2018, 50, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Carta, L.K.; Li, S. PCR amplification of a long rDNA segment with one primer pair in agriculturally important nematodes. J. Nematol. 2019, 51, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ley, P.; De Ley, I.T.; Morris, K.; Abebe, E.; Mundo-Ocampo, M.; Yoder, M.; Heras, J.; Waumann, D.; Rocha-Olivares, A.; Jay Burr, A.H.; et al. An integrated approach to fast and informative morphological vouchering of nematodes for applications in molecular barcoding. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1945–1958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowles, J.; Blair, D.; Mcmanus, D.P. Genetic-Variants within the Genus Echinococcus Identified by Mitochondrial-DNA Sequencing. Mol. Biochem. Parasit. 1992, 54, 165–174. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. Clustal-W—Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide-Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Tchesunov, A.V. Biology of Marine Nematodes; KMK Scientific Press: Moscow, Russia, 2006; p. 367. [Google Scholar]
- Lorenzen, S. New and known gonadal characters in free-living nematodes and the phylogenetic implications. Z. Syst. Zool. Evol. 1978, 16, 108–115. [Google Scholar] [CrossRef]
- Filipjev, I.N. The classification of the free-living nematodes and their relation to the parasitic nematodes. Smithson. Misc. Collect. 1934, 89, 1–63. [Google Scholar]
- Khan, S.H.; Coomans, A. Observations on the juvenile stages of Miconchus studeri (Nematoda: Mononchida). Biol. Jaarb. 1980, 48, 111–118. [Google Scholar]
- Vincx, M.; Sharma, J.; Smol, N. On the identity and nomenclature of Paracanthonchus caecus (Bastian, 1865), with a redefinition of the genus Paracanthonchus Micoletzky (Nematoda, Cyatholaimidae). Zool. Scr. 1982, 11, 243–263. [Google Scholar] [CrossRef]
- Coomans, A.; Jacobs, L.J. Halalaimus algeriensis n. sp. (Nematoda) from the Sahara. Hydrobiologia 1983, 102, 39–44. [Google Scholar] [CrossRef]
- Vincx, M. Redescription and ontogenetic of Desmodora schulzi Gerlach 1950. Biol. Jaarb. 1983, 51, 171–179. [Google Scholar]
- Coomans, A.; Heyns, J. Oncholaimus-Jessicae N-Sp (Nematoda, Oncholaimidae) from Fresh-Water in the Transvaal. S. Afr. J. Zool. 1986, 21, 197–201. [Google Scholar]
- Tchesunov, A.V.; Sturhan, D. Studies on Domorganus macronephriticus Goodey, 1947 (Nematoda: Ohridiidae). J. Nematode Morphol. Syst. 2004, 6, 139–150. [Google Scholar]
- Jensen, P. Free-living marine nematodes from a sublittoral station in the North Sea off the Belgian coast. Biol. Jaarb. 1976, 44, 231–255. [Google Scholar]
- Holovachov, O.; de Ley, P. Order Plectida. In Freshwater Nematodes: Ecology and Taxonomy; CABI Publishing: Wallingford, UK, 2006; pp. 611–647. [Google Scholar]
- Martinez-Arce, A.; De Jesus-Nayarrete, A.; Leasi, F. DNA Barcoding for Delimitation of Putative Mexican Marine Nematodes Species. Diversity 2020, 12, 107. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Manunema cf. proboscidis, entire: (A) female; (B) male. The red arrow points to a site of connection of the spermaduct (vas deferens) to a site where anterior and posterior gonads diverge. Scale bar: 100 μm.
Figure 1.
Manunema cf. proboscidis, entire: (A) female; (B) male. The red arrow points to a site of connection of the spermaduct (vas deferens) to a site where anterior and posterior gonads diverge. Scale bar: 100 μm.

Figure 2.
Manunema cf. proboscidis, details of a male: (A) anterior body; (B) posterior body. There are clots of slime emitted from the tips of the cephalic setae and the amphideal fovea. Scale bar: 10 μm.
Figure 2.
Manunema cf. proboscidis, details of a male: (A) anterior body; (B) posterior body. There are clots of slime emitted from the tips of the cephalic setae and the amphideal fovea. Scale bar: 10 μm.

Figure 3.
Manunema cf. proboscidis, female reproductive systems: (A) mature fertilized female, 533 μm long; antidromously reflected ovaries, vagina straight, vulva bulged, spermathecas; (B–D) immature females; outstretched ovaries, vagina bent, vulva not bulged beyond body surface; (B) body, 354 μm long; (C) body, 379 μm long; (D) body, 358 μm long. Scale bars: 20 μm.
Figure 3.
Manunema cf. proboscidis, female reproductive systems: (A) mature fertilized female, 533 μm long; antidromously reflected ovaries, vagina straight, vulva bulged, spermathecas; (B–D) immature females; outstretched ovaries, vagina bent, vulva not bulged beyond body surface; (B) body, 354 μm long; (C) body, 379 μm long; (D) body, 358 μm long. Scale bars: 20 μm.

Figure 4.
Manunema cf. proboscidis, SEM-pictures: (A) female, entire view; (B) somatic setae on sockets; (C) anterior end, amphideal fovea protruded; (D) anterior end, cephalic setae with apical pores; (E) vulva. Scale bars: (A) 20 μm, (B,D) 2 μm, (C) 3 μm, (E) 5 μm.
Figure 4.
Manunema cf. proboscidis, SEM-pictures: (A) female, entire view; (B) somatic setae on sockets; (C) anterior end, amphideal fovea protruded; (D) anterior end, cephalic setae with apical pores; (E) vulva. Scale bars: (A) 20 μm, (B,D) 2 μm, (C) 3 μm, (E) 5 μm.

Figure 5.
Bayesian inference phylogenetic tree of the 18S rRNA gene with Enoplus sp. set as an outgroup, with the posterior probability indicated as a percentage shown at each node.
Figure 5.
Bayesian inference phylogenetic tree of the 18S rRNA gene with Enoplus sp. set as an outgroup, with the posterior probability indicated as a percentage shown at each node.

Figure 6.
Rooted maximum likelihood phylogenetic tree of the 18S rRNA gene based on the GTR + F + I + G4 model, with UFboot/SH-aLRT support displayed at each node.
Figure 6.
Rooted maximum likelihood phylogenetic tree of the 18S rRNA gene based on the GTR + F + I + G4 model, with UFboot/SH-aLRT support displayed at each node.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer related information used in this study.
| Marker | Primer (Direction) | Sequence 5′-3′ | Sequence Length (bp) | Reference |
|---|---|---|---|---|
| mtCOI | JB3 (f) | TTTTTTGGGCATCCTGAGGTTTAT | 350–395 | [23,28] |
| JB5 (r) | AGCACCTAAACTTAAAACATAATGAAAATG | |||
| 18S | MN18F (f) | CGCGAATRGCTCATTACAACAGC | 336–349 | [24] |
| 22R (r) | GCCTGCTGCCTTCCTTGGA | |||
| 18S-CL-F (f) | TCAAAGATTAAGCCATGCAT | 983–1723 | [25,26] | |
| 18S-CL-R2 (r) | GTTGAGTCAAATTAAGCCGCA | |||
| 1912R (r) | TTTACGGTCAGAACTAGGG | |||
| 550F (f) | GGCAAGTCTGGTGCCAGCAGCC | |||
| 530R (r) | GCGGCTGCTGGCACCACACTT | |||
| 18S-CL-F2 (f) | CTGTGATGCCCTTAGATGTCC | |||
| 18S-CL-R5 (r) | GCGGTGTGTACAAAGGGCAGGGAC | |||
| 18S-CL-R7 (r) | ACCTTGTTACGACTTTTGCCCGGTTCA | |||
| 28S | D2A (f) | ACAAGTACCGTGAGGGAAAGTTG | 780 bp | [27] |
| D3B (r) | TCGGAAGGAACCAGCTACTA |
Table 2.
GenBank accession number of sequences obtained from this study.
| Specimen | Species Name | GenBank Accession Number | ||||
|---|---|---|---|---|---|---|
| mtCOI | 18S | 28S | ||||
| JB3 | 18S-CL-F | 18S-CL-F | MN18F | D2A | ||
| /JB5 | /18S-CL-R7 | /1912R | /22R | /D3B | ||
| (~390 bp) | (~1700 bp) | (~1100 bp) | (~300 bp) | (~730 bp) | ||
| 1 | Manunema cf. proboscidis | OR068235 | OR069708 | – | OR069703 | OR069692 |
| 2 | Manunema cf. proboscidis | OR068236 | – | OR069709 | OR069704 | OR069693 |
| 3 | Manunema cf. proboscidis | OR068237 | – | – | OR069705 | OR069694 |
| 4 | Manunema cf. proboscidis | OR068238 | – | OR069710 | OR069706 | OR069695 |
| 5 | Manunema cf. proboscidis | OR068239 | – | OR069711 | OR069707 | – |
Table 3.
Morphometrics of Manunema cf. proboscidis in males and females. All the measurements except ratios are given in μm.
Table 3.
Morphometrics of Manunema cf. proboscidis in males and females. All the measurements except ratios are given in μm.
| Character | Male (from Original Description of Gerlach, 1957) | Males (n = 9) mean ± sd (Range) | Females (n = 15) Mean ± sd (Range) |
|---|---|---|---|
| Body length | 353 | 354.4 ± 54.2 (301.0–477.0) | 415.5 ± 66.0 (330.0–533.0) |
| Pharynx length | 62 | 59.9 ± 5.6 (52.0–69.0) | 63.9 ± 3.9 (58.0–71.8) |
| Tail length | 58 | 63.5 ± 8.1 (52.2–76.0) | 65.2 ± 10.3 (50.0–85.5) |
| a | 16.1 | 17.4 ± 1.6 (14.3–19.4) | 14.6 ± 2.6 (12.2–18.7) |
| b | 5.7 | 5.7 ± 0.8 (5.0–7.6) | 6.5 ± 1.0 (5.4–8.3) |
| c | 6.1 | 5.6 ± 0.6 (4.5–6.6) | 6.2 ± 0.6 (4.7–6.9) |
| c′ | 2.9 | 3.9 ± 0.6 (3.0–4.7) | 4.1 ± 0.5 (3.2–4.6) |
| V | – | – | 49.7 ± 2.6 (43.0–53.3) |
| Body diameter at level of cephalic setae | 5 | 4.3 ± 0.4 (3.8–4.8) | 4.3 ± 0.7 (3.1–5.3) |
| Body diameter at level of amphid | – | 7.5 ± 0.4 (6.9–8.1) | 8.0 ± 0.9 (6.4–8.9) |
| Body diameter at level of nerve ring | – | 24.3 ± 1.8 (22.7–27.5) | 26.8 ± 1.7 (24.6–30.3) |
| Body diameter at level of cardia | – | 21.4 ± 2.8 (16.3–23.6) | 22.8 ± 4.5 (16.0–28.0) |
| Body diameter at level of midbody | 25 | 20.4 ± 2.2 (18.0–24.8) | 28.1 ± 1.5 (25.6–31.0) |
| Body diameter at level of cloaca/anus | 20 | 15.7 ± 1.4 (14.1–17.7) | 15.7 ± 1.4 (13.8–18.6) |
| Cephalic setae length | 5 | 6.7 ± 1.2 (4.7–8.0) | 6.4 ± 0.9 (4.8–8.0) |
| Amphid width | – | 2.7 ± 0.5 (2.0–3.4) | 2.9 ± 1.0 (1.5–4.0) |
| Distance head apex—amphid | – | 18.2 ± 1.3 (17.0–19.7) | 17.6 ± 1.7 (15.0–19.0) |
| Length of pharynx posterior muscular part | – | 30.4 ± 3.3 (24.0–34.2) | 32.9 ± 3.2 (28.2–37.9) |
| Pharynx diameter of preneural muscular swelling | 5–6 | 8.9 ± 0.5 (8.3–9.7) | 2.6 ± 0.5 (1.8–3.6) |
| Pharynx diameter at nerve ring | – | 7.0 ± 0.8 (5.2–7.8) | 8.1 ± 0.8 (6.9–9.3) |
| Pharynx diameter of postneural muscular swelling | 13 | 10.8 ± 1.8 (9.3–15.0) | 13.2 ± 2.1 (10.7–17.3) |
| Length of spicules along arc | 26 | 31.2 ± 4.6 (25.9–40.6) | – |
| Length of gubernaculum | 16 | 11.2 ± 2.3 (7.9–13.2) | – |
| Length of anterior supplement | 15 | 13.9 ± 4.1 (10.4–23.0) | – |
| Length of posterior supplement | 15 | 14.0 ± 3.4 (10.0–19.6) | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jeong, R.; Tchesunov, A.V. Phylogenetic Position of the Genus Manunema (Nematoda, Plectida, Leptolaimidae). Diversity 2023, 15, 914. https://doi.org/10.3390/d15080914
AMA Style
Jeong R, Tchesunov AV. Phylogenetic Position of the Genus Manunema (Nematoda, Plectida, Leptolaimidae). Diversity. 2023; 15(8):914. https://doi.org/10.3390/d15080914
Chicago/Turabian StyleJeong, Raehyuk, and Alexei V. Tchesunov. 2023. "Phylogenetic Position of the Genus Manunema (Nematoda, Plectida, Leptolaimidae)" Diversity 15, no. 8: 914. https://doi.org/10.3390/d15080914
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.