

Status of the Italian Freshwater Gastrotricha Biodiversity, with the Creation of an Interactive GIS-Based Web Map


1
Department of Life Sciences, Università di Modena e Reggio Emilia, Via G. Campi, 213/D, 41125 Modena, Italy
2
Department of Earth and Marine Sciences, University of Palermo, Viale delle Science, 90128 Palermo, Italy
3
National Biodiversity Future Center (NBFC), Piazza Marina 61, 90133 Palermo, Italy
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(1), 17; https://doi.org/10.3390/d16010017
Submission received: 23 November 2023
/
Revised: 21 December 2023
/
Accepted: 22 December 2023
/
Published: 25 December 2023
(This article belongs to the Special Issue Diversity in 2023)
Abstract
:Gastrotricha are microscopic benthic animals found in almost all water bodies. To date, over 890 species distributed in 71 genera, 18 families, and two orders are known. There are 376 freshwater species, and several are also found in Italy; however, a compendium of the studies carried out so far, and a georeferenced distribution of the species, still need to be provided. This project summarizes information about the diversity and geographic distribution of Italian gastrotrichs. Diversity data, acquired over 239 years, were reviewed and corrected based on taxonomic and nomenclatural updates and, in addition to distribution information, were organized into a data matrix valuable for statistical analysis. The data were fed into geographic information system software to understand the overall figures more easily. The results indicated that Italy is one of the best-known countries regarding freshwater gastrotrichs. With 92 species in 17 genera and three families from 61 investigated localities, Italy counts 17 type localities and 19 endemic species. Despite this high biodiversity, many Italian regions still need to be investigated, and should, therefore, be the focus of future research. The implementation of a web-mapping plugin enabled the creation of interactive maps for an easy and modern method for sharing the work done, and the information acquired.
1. Introduction
The phylum Gastrotricha (from Greek gaster, stomach, and thrix, hair, referring to the extensive ventral locomotor ciliature), includes microscopic, free-living, aquatic organisms. Marine and brackish-water species lead a wandering life among the interstitial spaces of poorly compacted sandy sediments. Freshwater gastrotrichs, on the other hand, live mainly on the surface of the muddy–sand bottoms of lakes and streams (epibenthic) or among aquatic plants (periphytic). Few species are interstitial, and even fewer live in the water column (planktonic) [1].
The phylogenetic position of the Gastrotricha has been long debated [2]; however, recent robust phylogenomic analyses based on transcriptomic data indicate that these micrometazoans are allied with the Phatyhelminthes, forming a subclade (Rouphozoa) within the spiralian protostomes [3,4,5,6].
Gastrotrichs have a more or less transparent body, spanning 80 µm to 3.5 mm in length, and are convex dorsally, and flattened ventrally [7]. They have a triploblastic, acoelomate anatomical organization with unsegmented tissues and organs. They are characterized by locomotor cilia and a well-developed, bistratified outer cuticle, often forming variously arranged plates, scales, and spines. The cuticle of the gastrotrich is unique in that it enwraps the entire body, including the cilia [8].
Over 890 species of gastrotrichs have been described, divided into two orders, Macrodasyida and Chaetonotida, whose representatives may differ in biology and ecology, and are easily distinguished on a morphological basis (Figure 1).
The order Macrodasyida (about 380 species, divided into 37 genera and ten families, Table 1) includes almost exclusively marine and estuarine gastrotrichs (there are only four freshwater species). They are vermiform in shape, ranging in size from 150 µm to 3.5 mm, and generally have a smooth cuticle, which is sometimes differentiated into scales and/or spines with protective functions. Macrodasyida generally have many adhesive tubes along the body sides, giving the animal a remarkable ability to adhere to the substrate, which is essential for practical life in the interstitial environment. Locomotor cilia may cover the entire ventral surface or be arranged into two longitudinal ciliary bands.
The mouth opening is often very large. The myoepithelial pharynx has a triradiate lumen, typically in the shape of an inverted Y, and possesses two pores in the posterior region that allow for the exit of excess water ingested during feeding. Macrodasyida are usually insufficient hermaphrodites, except for about ten species that are parthenogenetic instead [9].
The order Chaetonotida (about 515 species, divided into 34 genera and eight families; Table 2) includes the gastrotrichs most commonly found in freshwater; however, many species live in marine and estuarine environments, and three families (Muselliferidae, Neodasyidae, and Xenotrichulidae) are exclusively marine [7].
Chaetonotidans are generally smaller than macrodasyidans (the total length is generally between 100 and 300 µm) and their bodies are shaped like tenpins. The caudal region is furcate (named furca, Figure 1), except for species affiliated with the genus Neodasys, which are vermiform in appearance. The cuticle of Chaetonotida is almost always differentiated into plates, scales, and spines of highly variable shapes and sizes. The locomotor cilia are, in general, arranged to form two longitudinal ciliary bands.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
High-ranking classification of Gastrotricha Macrodasyida. The number of marine (M) and freshwater (F) species for each taxon is in brackets.
Table 1.
High-ranking classification of Gastrotricha Macrodasyida. The number of marine (M) and freshwater (F) species for each taxon is in brackets.
| Family CEPHALODASYIDAE Hummon & Todaro, 2010 [34 M spp.] |
| Genus Cephalodasys Remane, 1926 [14 M spp.] |
| Genus Dolichodasys Gagne, 1977 [3 M spp.] |
| Genus Mesodasys Remane, 1951 [8 M spp.] |
| Genus Paradasys Remane, 1934 [7 M spp.] |
| Genus Pleurodasys Remane, 1927 [2 M spp.] |
| Family DACTYLOPODOLIDAE Strand, 1929 [17 M spp.] |
| Genus Dactylopodola Strand, 1929 [11 M spp.] |
| Genus Dendrodasys Wilke, 1954 [6 M spp.] |
| Genus Dendropodola Hummon, Todaro & Tongiorgi, 1993 [1 M sp.] |
| Family HUMMONDASYIDAE Todaro, Leasi & Hochberg, 2014 [1 M sp.] |
| Genus Hummondasys Todaro, Leasi & Hochberg, 2014 [1 M sp.] |
| Family LEPIDODASYIDAE Remane, 1927 [9 M spp.] |
| Genus Lepidodasys Remane, 1926 [9 M spp.] |
| Family MACRODASYIDAE Remane, 1926 [56 M spp.] |
| Genus Kryptodasys Todaro, Dal Zotto, Kånneby, Hochberg, 2019 [7 M spp.] |
| Genus Macrodasys Remane, 1924 [32 M spp.] |
| Genus Thaidasys Todaro, Dal Zotto & Leasi, 2015 [1 M sp.] |
| Genus Urodasys Remane, 1926 [16 M spp.] |
| Family PLANODASYIDAE Rao & Clausen, 1970 [14 M spp.] |
| Genus Crasiella Clausen, 1968 [9 M spp.] |
| Genus Megadasys Schmidt, 1974 [3 M spp.] |
| Genus Planodasys Rao & Clausen, 1970 [2 M spp.] |
| Family REDUDASYIDAE Todaro et al. 2012 [1 M and 3 F spp.] |
| Genus Anandrodasys Todaro et al., 2012 [1 M sp.] |
| Genus Redudasys Kisielewski, 1987 [3 M spp.] |
| Family THAUMASTODERMATIDAE Remane, 1927 [176 M spp.] |
| Subfamily Diplodasinae Ruppert, 1978 [23 M spp.] |
| Genus Acanthodasys Remane, 1927 [13 M spp.] |
| Genus Diplodasys Remane, 1927 [10 M spp.] |
| Subfamily Thaumastodermatinae Remane, 1927 [154 M spp.] |
| Genus Chimaeradasys Kieneke & Todaro, 2020 [2 M spp.] |
| Genus Hemidasys Claparède, 1867 [1 M sp. *] |
| Genus Oregodasys Hummon, 2008 [16 M spp.] |
| Genus Pseudostomella Swedmark, 1956 [17 M spp.] |
| Genus Ptychostomella Remane, 1926 [13 M spp.] |
| Genus Tetranchyroderma Remane, 1926 [86 M spp.] |
| Genus Thaumastoderma Remane, 1926 [18 M spp.] |
| Family TURBANELLIDAE Remane, 1927 [64 M spp.] |
| Genus Desmodasys Clausen, 1965 [3 M spp.] |
| Genus Dinodasys Remane, 1927 [2 M spp.] |
| Genus Paraturbanella Remane, 1927 [24 M spp.] |
| Genus Prostobuccantia Evans & Hummon, 1991 [1 M sp.] |
| Genus Pseudoturbanella d’Hondt, 1968 [1 M sp.] |
| Genus Turbanella Schultze, 1853 [33 M spp.] |
| Family XENODASYIDAE Todaro, Guidi, Leasi & Tongiorgi, 2006 [4 M spp.] |
| Genus Chordodasiopsis Todaro, Guidi, Leasi & Tongiorgi, 2006 [1 M sp.] |
| Genus Xenodasys Swedmark, 1967 [3 M spp.] |
| INCERTAE SEDIS |
| Genus Marinellina Ruttner-Kolisko, 1955 [1 F sp.] |
* Hemidasys agaso Claparède, 1867 has been declared extinct by Hummon and Todaro [10].
Table 2.
High-ranking classification of Gastrotricha Chaetonotida. The number of marine (M) and freshwater (F) species for each taxon is in brackets.
Table 2.
High-ranking classification of Gastrotricha Chaetonotida. The number of marine (M) and freshwater (F) species for each taxon is in brackets.
| Suborder MULTITUBULATINA d’Hondt, 1971 [3 M spp.] |
| Family NEODASYIDAE Remane, 1929 [3 M spp.] |
| Genus Neodasys Remane, 1927 [3 M spp.] |
| Suborder PAUCITUBULATINA d’Hondt, 1971 [145 M * and 372 F spp.] |
| Family CHAETONOTIDAE Gosse, 1864 [112 M * and 320 F spp.] |
| Subfamily Chaetonotinae Kisielewski, 1991 [112 M and 319 F spp.] |
| Genus Arenotus Kisielewski, 1987 [1 F sp.] |
| Genus Aspidiophorus Voigt, 1903 [11 M and 24 F spp.] |
| Genus Bifidochaetus Kolicka & Kisielewski [2 F spp.] |
| Genus Caudichthydium Schwank, 1990 [3 M spp.] |
| Genus Cephalionotus Garraffoni et al., 2017 [1 F sp.] |
| Genus Chaetonotus Ehrenberg, 1830 [47 M ** and 191 F spp.] |
| Genus Fluxiderma d’Hont, 1974 [3 F spp.] |
| Genus Halichaetoderma Rataj Križanová & Vdacný, 2023 [4 F spp.] |
| Genus Halichaetonotus Remane, 1936 [31 M spp.] |
| Genus Heterolepidoderma Remane, 1927 [15 M and 25 F spp.] |
| Genus Ichthydium Ehrenberg, 1830 [3 M and 29 F spp.] |
| Genus Lepidochaetus Kisielewski, 1991 [6 F spp.] |
| Genus Lepidodermella Blake, 1933 [2 M *** and 15 F spp.] |
| Genus Polymerurus Remane, 1926 [16 F spp.] |
| Genus Rhomballichthys Schwank, 1990 [1 F spp.] |
| Subfamily Undulinae Kisielewski, 1991 [1 F sp.] |
| Genus Undula Kisielewski, 1991 [1 F sp.] |
| Family DASYDYTIDAE Daday, 1905 [35 F spp.] |
| Genus Anacanthoderma Marcolongo, 1910 [2 F spp.] |
| Genus Chitonodytes Remane, 1936 [3 F spp.] |
| Genus Dasydytes Gosse, 1851 [9 F spp.] |
| Genus Haltidytes Remane, 1936 [6 F spp.] |
| Genus Ornamentula Kisielewski, 1991 [2 F spp.] |
| Genus Setopus Grünspan, 1908 [9 F spp.] |
| Genus Stylochaeta Hlava, 1904 [4 F spp.] |
| Family DICHAETURIDAE Remane, 1927 [4 F spp.] |
| Genus Dichaetura Lauterborn, 1913 [4 F spp.] |
| Family MUSELLIFERIDAE Leasi & Todaro, 2008 [7 M spp.] |
| Genus Diuronotus Todaro, Kristensen & Balsamo, 2005 [2 M spp.] |
| Genus Musellifer Hummon, 1969 [5 F spp.] |
| Family NEOGOSSEIDAE Remane, 1927 [11 F spp.] |
| Genus Kijanebalola Beauchamp, 1932 [4 F spp.] |
| Genus Neogossea Remane, 1927 [7 F spp.] |
| Family PROICHTHYDIDAE Remane, 1927 [2 F spp.] |
| Genus Proichthydium Cordero, 1918 [1 F spp.] |
| Genus Proichthydioides Sudzuki, 1971 [1 F spp.] |
| Family XENOTRICHULIDAE Remane, 1927 [26 M spp.] |
| Subfamily Draculiciterinae Ruppert, 1979 [1 M sp.] |
| Genus Draculiciteria Hummon, 1974 [1 M sp.] |
| Subfamily Xenotrichulinae Remane, 1927 [25 M spp.] |
| Genus Heteroxenotrichula Wilke, 1954 [9 M spp.] |
| Genus Xenotrichula Remane, 1927 [16 M spp.] |
* includes seven species occasionally found in marine/brackish waters. ** includes six species occasionally found in marine/brackish waters; *** includes one species occasionally found in marine/brackish waters [10].
In almost all species, there are only two adhesive tubes, forming the distal part of the caudal furca (Figure 1). The mouth of Chaetonotida is small and the pharynx has a triradiate, Y-shaped lumen devoid of pores. Reproduction generally occurs by parthenogenesis in freshwater species, while marine forms are usually hermaphroditic, with the exception of members of the family Chaetonotidae [8,11].
The phylum Gastrotricha has a cosmopolitan distribution (i.e., species of the phylum have been found in at least two oceans and/or continents), as are the two orders and almost all the species-rich families and genera. As with other meiobenthic animals, the “meiofauna paradox” [12] also applies to gastrotrichs since many species seem to have a wide distributional range, and some are even considered cosmopolitan.
As for marine species, dispersal by longshore currents over long periods (geologic scale), combined with oceanic dispersal and continental drift, can be invoked to explain the biogeographic distribution patterns of Gastrotricha in many marine systems; however, passive dispersal by phoresy, rafting, and within a ballast medium (water or sand) may also play a relevant role.
For the dispersal of freshwater chaetonotidans, a key role can be attributed to the resting eggs, which are spread accidentally by the wind or by particularly vagile animals (e.g., migratory birds). In suitable conditions, these eggs, coupled with parthenogenetic reproduction, allow for the genesis of new populations in disjunct habitats [13].
The order Macrodasyida includes four freshwater species distributed in two genera whose species have been found either at a single site, such as Marinellina flagellata Ruttner-Kolisko, 1955 in Austria [14] or at a few sites, such as Redudasys fornerise and R. brasiliensis in Brazil [15,16,17], and R. neotemperatus Kånneby & Kirk, 2017 in the USA [18,19]. Resting eggs are not reported for these parthenogenetic species.
Within freshwater Chaetonotida, the families Dasydytidae, Neogosseidae, and especially Chaetonotidae (subfamily Chaetonotinae), are relatively common. They are present on various continents and have many species, several of which have an amphi-atlantic or cosmopolitan distribution [20]. By contrast, Dichaeturidae, Proichthydiidae, and the subfamily Undulinae (Fam. Chaetonotidae) are quite rare. They include 1–4 species whose distribution appears to be confined to single sites/continent. More specifically, Dichaeturidae includes two species found in Europe, one in the USA and one in Japan. Proichthydiidae contains one species found in South America (Uruguay) and another in Asia (Japan). The subfamily Undulinae (Fam. Chaetonotidae) embraces a single species in the Amazon [21].
Knowledge about the diversity, and especially the distribution of the Italian freshwater species, needs to be more detailed than that of marine gastrotrichs [22]. The first finding of a gastrotrich in Italian fresh waters has been attributed to Buonaventura Corti, who writes of a “hairy soft animal”, probably referring to a Chaetonotida [23]. Information of a similar anecdotal nature continued until the early 1900s and concerned a few individuals found in different localities of the northern area of the nation [24,25,26].
The first Italian freshwater gastrotrichs described in some detail are by Marcolongo [27,28], who studied the fauna of the Astroni Lake near Naples (southern Italy). Subsequently, a significant contribution came from studies performed in Sardinia by Mola [29] and especially by Balsamo [30,31,32,33,34], who investigated freshwater biotopes in northeastern and central regions. The most recent taxonomic research involving Italian freshwater gastrotrichs dates back 25 years and deals with interstitial gastrotrichs from lotic Italian waters [35]. More recently, some freshwater species have recently been used in phylogenetic studies based on molecular markers (e.g., Kånneby et al. [36,37]).
Several Italian freshwater gastrotrich species checklists have been proposed over time [38,39]. However, none provide georeferenced information or summaries regarding, for example, regional distributions or the types of localities.
The present work aims to fill these gaps. The study is part of a large Italian national biodiversity project (NBFC-National Biodiversity Future Center) and falls under the mission of Spoke 3, focusing on terrestrial and inland water biodiversity. This prodromal research on gastrotrichs primarily involves acquiring and georeferencing published information on freshwater taxa. Creating interactive maps will allow for an easy and modern worldwide sharing of information to benefit taxonomists, conservation biologists, policymakers, and the wider public [40].
2. Materials and Methods
2.1. Gastrotrichs
Information about the diversity and distribution of Italian freshwater gastrotrichs was derived from the following published papers, listed in ascending temporal order: Corti [23], Maggi [24], Issel [25], Zacharias [26] Grünspan [41], Marcolongo [27,28], Manfredi [42], Mola [29], Balsamo [30,31,32,33], Balsamo and Kisielewski [43], Balsamo and Todaro [44], Bertolani and Balsamo [45], Balsamo [34], Balsamo et al. [46], Balsamo and Fregni [47], Balsamo and Todaro [48], Fregni et al. [35], Leasi et al. [49], Leasi and Todaro [50], and Kånneby et al. [36,37]. Bibliographic data were carefully checked and, if necessary, corrected based on taxonomic and nomenclatural updates that have occurred over the years e.g., Schwank [51] Kisielewski [21], see also WoRMS [52]. As the subgeneric grouping of Chaetonotus and Ichthydium species has been found to be non-monophyletic [37,53], the classification at the subgeneric level was not adopted in the present work.
In many cases, the geographical coordinates of the investigated localities and sites (sites = sampling spots in a given locality/biotope) were not indicated in the consulted papers; consequently, this information was obtained through freely accessible satellite images (Google Earth and ESRI satellite), based on the toponyms and indications reported in the publications. In cases where the geographical coordinates were reported in the original works, satellite maps made it possible to correct errors or improve the precision of the georeference, which in the past was often limited to degrees and minutes only.
Finally, Prof. M. Balsamo kindly provided unpublished information about the sampling localities of some species reported in a previous checklist of Italian gastrotrich species [38] (Supplementary Table S1).
2.2. Data Matrix
The information obtained from the bibliographic consultation was collected using the spreadsheet software MS Excel®. The dataset was used to obtain basic statistics (see below) and to build a database exploitable by the open-access GIS software QGIS (version 3.14) (https://www.qgis.org/en/site/ (accessed on 24 September 2023)) [54]. Therefore, the data were coded in a “columns by rows” matrix, in which the columns represent the variables, and the rows represent the cases. The final dataset comprised 26 columns and 306 rows for a total of 6527 cells. The matrix variables constituted the QGIS attributes (or fields), based on which the program allows for targeted visualizations, searches, and analyses. The attributes we coded fall into three main categories: (i) data relating to the species, such as high-ranking classification, images, Aphia ID, and endemicity; (ii) data relating to the location where the species was found, such as the administrative region, province, locality, type locality, national faunistic provinces, and geographic coordinates, in both DMS (degrees, minutes and seconds) and DD (decimal degrees) systems, the latter of which are used by the program, and (iii) data regarding the researchers involved and the source of the data (i.e., bibliographic reference).
2.3. Statistical Analysis
To investigate the faunal connections between the regions sampled, we analyzed the presence/absence data using the Bray–Curtis algorithm to create a similarity matrix (see Supplementary Table S2). This similarity matrix was then analyzed using multivariate statistical techniques, including cluster analysis (see Figure 2 in the results below) and nonmetric multi-dimensional scaling (nMDS, see Figure 3 in the results below). The creation of the similarity matrix and the statistical analyses were both performed using Past software (version 4.07) (https://www.nhm.uio.no/english/research/resources/past/ (accessed on 22 September 2023)).
2.4. QGIS
2.4.1. Version and Plugins Used and Software Preparation
For the analysis and representation of the data relating to the Italian freshwater gastrotrichs, after a few trials, we selected QGIS 3.14 Hannover version (release 3.16.8) as it proved to be the most stable. Distributional data are georeferenced according to the WGS 84 reference system (encoded in QGIS as EPGS:4326) as it is the most frequently used geolocalization system. Moreover, the QuickMapService plugin was applied to import the reference map (ESRI satellite) and the map BoundariesPlaces (also by ESRI); the qgis2web plugin was used to produce the web map.
2.4.2. Creation of the Data Shapefile Layers
The data matrix in CSV format was imported to QGIS to produce a comprehensive vector shapefile layer containing point-like geometry, with points representing the investigated locations to which all the other attributes were associated (see above). This shapefile (named Italy-All-Species) was subsequently used to create the shapefiles involving: (i) localities; (ii) single species; (iii) type localities; and (iv) endemic species. A further layer, containing the summary information regarding the single Italian administrative regions, was created starting from a pre-existing shapefile downloaded from the Italian National Institute of Statistics (ISTAT) website (https://www.istat.it/it/archivio/222527 (accessed on 29 September 2023)) [55] and modified to suit our purpose. More specifically, the original attributes about the demography and the territorial features were erased, and new ones, including the number of species present in a given region, the number of sites sampled, the number of type localities, and the average number of species at each site, were created.
2.4.3. Web Map Preparation and Exportation
All the produced layers were selected to be part of the final web map created in the Leaflet format. This format was chosen because it is the most widespread open-source JavaScript library for mobile-friendly interactive maps and is easier to use. The reference map and comprehensive layer (Italy-All-Species) were the only ones set to be active at the map’s opening; in contrast, all the other layers were set to be activated by the final users (see results Figure 4). The pop-up and the cluster options were selected for the comprehensive layer, while the pop-up option was selected for all the other points’ vector layers. For clarity, the primary layer’s record points were shown clustered at the default small scale; the single locations making up each given cluster could be visualized at a larger scale by progressive zoom-in actions (see results Figure 5). The pop-up boxes were devised to show significant information about the specific layer (see results Figure 6).
Once all the necessary settings were established, the web map export command was evoked, and the obtained folders and files were uploaded to the server dedicated to Gastrotricha (www.gastrotricha.unimore.it (accessed on 25 September 2023)), which is hosted at the meiofauna laboratory of the Department of Life Sciences, University of Modena, and Reggio Emilia. The map is available at the following ULR: http://gastrotricha-gis-italy.unimore.it/ (accessed on 25 September 2023). A redundant map copy is posted on a commercial server provider at the following URL: www.FreshwaterGastrotrichaIT.altervista.org (accessed on 25 September 2023).
3. Results
3.1. Gastrotrichs
Records of Italian freshwater gastrotrichs apparently cover a 239-year time frame. However, initial data come from simple species lists needing morphometric information on the reported taxa. Consequently, these accounts should be treated with caution. For instance, Marcolongo [28] believed the first recorded Italian freshwater gastrotrich to be the “hairy soft animal” that Bonaventura Corti found in Modena (see Corti, 1774, Tav. II, Figure XI) [23] and that Eherenberg [56] identified with Chaetonotus maximus Eherenberg, 1838. However, the animal depicted by Corti [23] can hardly be identified as a gastrotrich; its elongated body without the trademark caudal furca makes it resemble a microturbellarian instead. A copy of Corti’s paper can be found at the following url: https://archive.org/details/osservazionimic00corti/page/n209/mode/2up/ (accessed on 25 September 2023).
Similarly, we could not confirm the finding of Lepidodermella squamata (Dujardin, 1841) near Trieste (Region Friuli Venezia Giulia) attributed to Grüspan, 1908 [41] by Marcolongo [28] and Mola [29]. On the other hand, based on our bibliographic search, the following historical records are confirmed: the presence of Ichthydium podura (Müller, 1773) and Chaetonotus larus Müller, 1773 in Valcuvia (Lombardia region) Maggi [24]; the occurrence of Heterolepidoderma ocellatum Metschnikoff, 1865 in the Euganean thermal waters (Veneto region) Issel [25]), and of Chaetonotus chuni Voigt, 1901 near Pavia (Lombardia region) Zacharias [26].
The first information on the Italian species, including morphometric data, were recorded in the taxonomic study by the Italian researcher Ines Marcolongo, who investigated the Astroni crater lake near Naples, and reported 17 species, eight of which were described as new to science [27,28]. The most recent published records of Italian freshwater species can be found in two phylogenetic works by Kånneby et al. [36,37], which list Chaetonotus schultzei, Lepidochaetus zelinkai (Grünspan, 1908) and Polymerurus nodicaudus (Voigt, 1901), all found in Lake Pratignano, in the province of Modena [36,37].
Although the period covered by the published literature is relatively broad, the intensity of the studies has not remained constant over time. Relatively long periods of stasis were followed by energetic and productive investigative activity, although this was often restricted to single regions (e.g., Manfredi [42] in Lombardia; Mola [29] in Sardinia). It was only in 1977 that Italian gastrotrichs became the subject of constant and more extensive studies [30,31,32,33,43], and the last decade of the past century proved particularly fruitful in shedding light on the surprising diversity of gastrotrichs in the Italian inland waters. During that period, a series of investigations carried out by researchers of the University of Modena and Reggio Emilia (Italy), headed by professors Maria Balsamo and Paolo Tongiorgi, significantly contributed to the increase in the number of species and Italian regions involved in the survey [34,35,46,47,48].
Our analysis shows that the number of freshwater gastrotrich species currently known in the Italian inland waters amounts to 92 (Table 3 and Table 4), all belonging to the order Chaetonotida. The 92 Italian species, distributed in 17 genera and three families, were found by investigating 61 localities/biotopes and 81 sites (i.e., sampling points). However, of the 20 Italian regions, less than half (nine regions) have been investigated to any extent about the freshwater gastrotrichfauna (Table 3 and Table 4).
The species found in the investigated locations (loc) range between 1 and 33, with an average of 5.33 ± 6.13 spp/loc (Table 5). The location for which the highest number of species has been reported (33 species) is Monte Bondone, in the Trentino Alto Adige region. This total species number was reached by pooling data from samples collected in several circular water bodies locally known as Viotte or Viote. These are craters created by the explosion of artillery shells during World War I, which were then filled with rainwater [48].
Currently, 19 species are considered endemic to Italy (i.e., so far recorded only in this country), of which 12 have their distribution limited to only one of the nine investigated regions. Italy counts 17 type localities concerning freshwater gastrotrich species, some of which are for more than one species (Table 4). Lepidodermella squamata, found in 27 localities, appears to be the most frequently found species in the Italian inland waters (Table 5).
The distribution of the investigated localities, as well as the richness and diversity of the fauna, appears to be uneven. The most studied regions are Emilia Romagna, Sardegna (Sardinia), and Toscana (Tuscany), while others have yet to be investigated (Table 5). The most surveyed region by far is Sardegna, with 21 localities investigated and 34 species found (averaging 1.81 ± 0.93 spp/loc). Most information on the rich gastrotrich fauna from Sardegna dates back more than 90 years, and no additional records have been added since the work by Mola [29], except three species new to Sardinia reported by Fregni et al. [35].
In his seminal work on the Sardinian gastrotrichs, Mola [29] also proposed an interesting new classification for the group, including new subclasses, orders, families, genera etc. However, Mola’s classification was not followed/adopted by succeeding specialists, probably because of the language barrier (the article is written in Italian). Of the taxa proposed by Mola [29], only the name Wolterecka persists in the current classification of the Gastrotricha, designating a subset of the vast genus Chaetonotus [57]. At least part of Mola’s classification could be helpful in light of the recent molecular phylogenetic studies showing the non-monophyletic nature of many current freshwater groups.
The second region based on localities investigated is Emilia Romagna, with 14 biotopes studied and 28 species collected (averaging 4.57 ± 3.08 spp/loc). Data from this region are among the most recent, the latest of which date back to 2013 [37]. Although Tuscany ranks only third in terms of localities investigated (10 locations), it occupies the first position regarding the number of species found (41 spp). The most recent information from Tuscany dates back about 25 years and comes from surveys conducted in lotic waters in the province of Pisa [35]. Despite the low number of sites surveyed, Campania, Trentino Alto Adige, and Lazio (Latium) host a relatively high number of found species and occupy the first, second, and third places, respectively, in terms of species abundance (number of species per locality, Table 5). The Campania records date back to more than a century ago, and the reported simplified anatomy of some of the described species leaves doubts about their reliability, also considering the poor optical instrumentation in use at that time [27,28]. In contrast, the information from Trentino Alto Adige and Lazio is relatively recent and derived from methodologies in line with current taxonomic practices (e.g., differential interference contrast microscopy [48]).
The diversity of the Italian gastrotrich fauna, understood as the number of species, does not seem to be related to the number of localities investigated so much as to the type of biotope studied; eutrophic environments characterized by rich and varied aquatic vegetation seem to offer the best conditions for the development of a highly diversified biocoenosis [31]. However, the peculiarity of fauna associated with oligohaline environments at high altitudes or with biotopes whose waters are characterized by a low pH value, such as pit moss, and even interstitial environments, should be emphasized [33,34]. Plans for future research should take all these considerations into account.
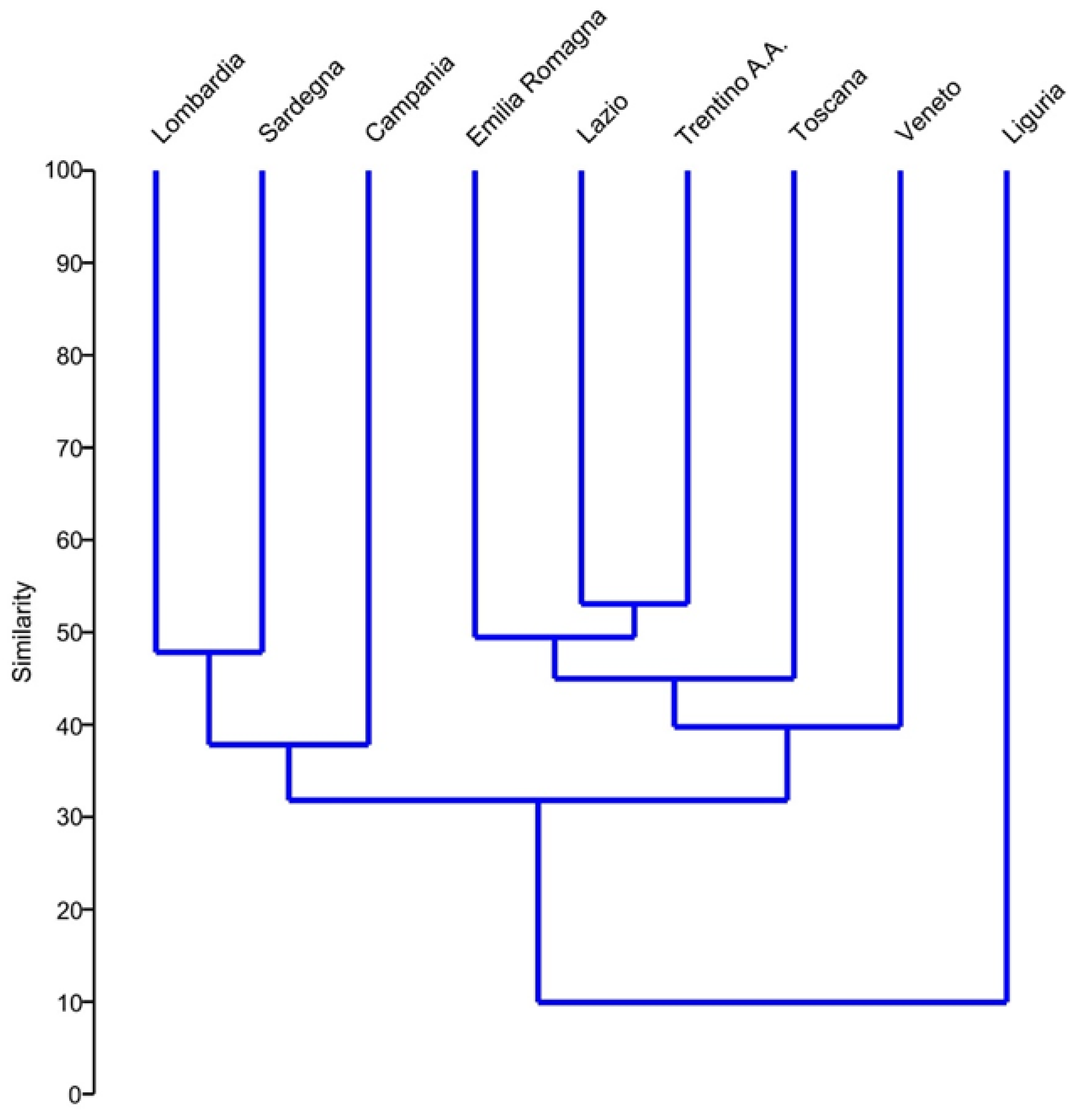
Multivariate statistical analyses based on presence/absence data indicate low faunal similarity between the regions investigated (Figure 2 and Figure 3). The results from the cluster analysis, a classification technique that groups the samples into hierarchical groups based on the similarity matrix (Table S1), show Liguria as separated from the other regions, at a faunal similarity value of around 10% (Figure 2). The species diversity recorded in Liguria is also lower (two species) than in the other regions (Table 4). Among the remaining regions, Lombardia, Sardegna, and Campania are separated from the other five regions at a diversity value of 30%. In the largest cluster, Lazio and Trentino Alto Adige have the highest faunal similarity (about 50%). No prominent hypotheses can be derived from the shared gastrotrichfauna among regions, considering their mutual distance and topographic characteristics.
Results from the Cluster analysis are reflected in the two-dimensional ordering plane obtained from the nMDS analysis (Figure 3). Similar to the outcome of the classification technique, the regions in the two-dimensional space are ordered irrespective of their geographic localization or general topography. Overall, results from our analysis do not point to possible geographic factors (e.g., abiotic) responsible for the distribution of recorded fauna. This result is not surprising considering that current data are affected to a great extent by the different sampling efforts applied to the different regions and by the diverse taxonomic techniques that have been used over the long period between collecting the first data [27] and the most recent data [37].
Figure 2.
Cluster analysis result showing hierarchical clustering of the investigate regions based on Bray–Curtis similarity index on presence/absence of freshwater gastrotrich species.
Figure 2.
Cluster analysis result showing hierarchical clustering of the investigate regions based on Bray–Curtis similarity index on presence/absence of freshwater gastrotrich species.

Figure 3.
Result of the nMDS analysis showing the ordination of the investigated regions based on the Bray–Curtis similarity index on presence/absence of freshwater gastrotrich species.
Figure 3.
Result of the nMDS analysis showing the ordination of the investigated regions based on the Bray–Curtis similarity index on presence/absence of freshwater gastrotrich species.

3.2. QGIS and Web Map
The first screen of the interactive map offered to the user shows a map containing two panels through which specific searches can be made: (i) the panel zoom, through which the user can zoom in and out of areas of interest and (ii) the panel layers, through which the user can query the system in a search for information of interest (Figure 4).
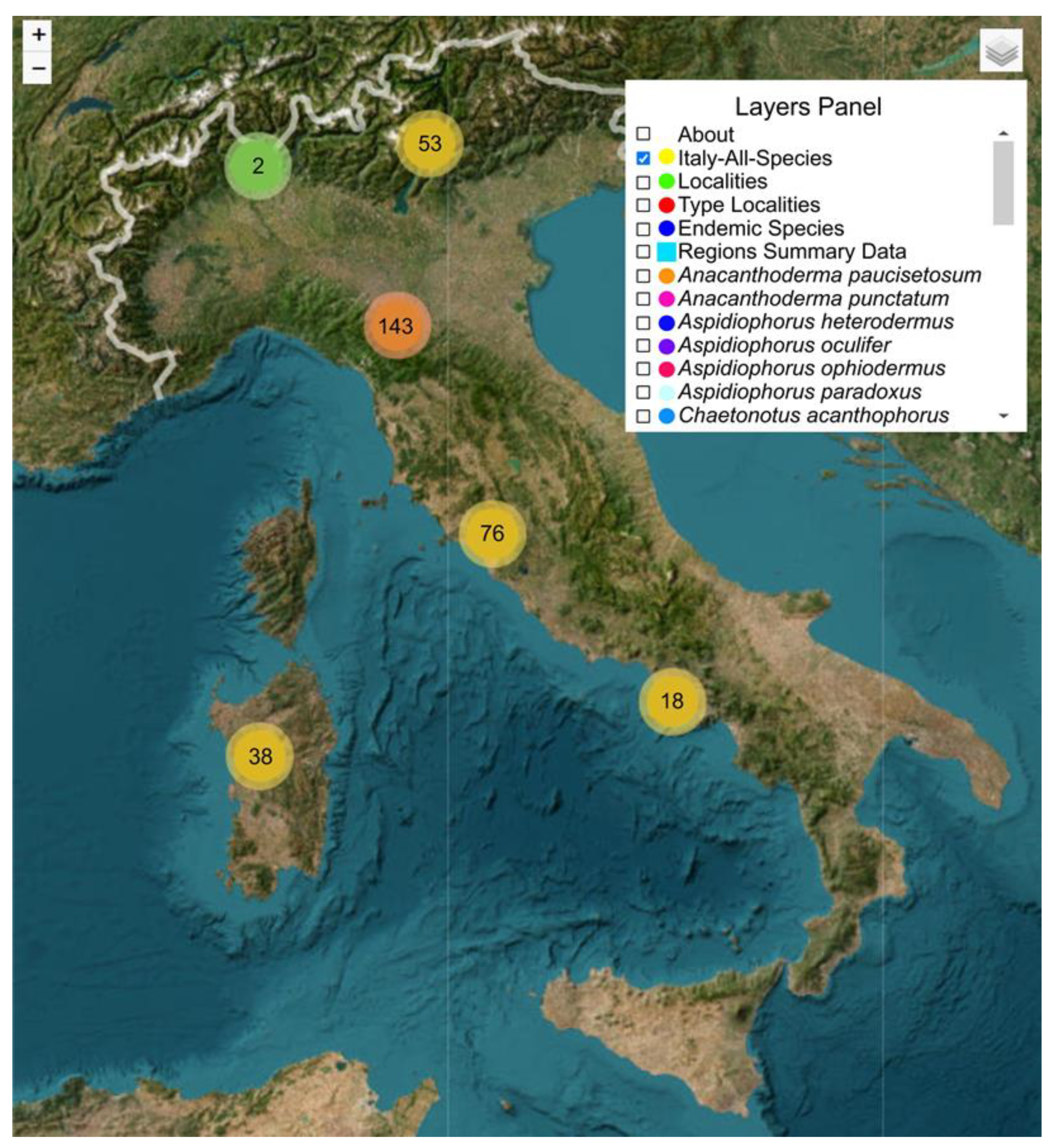
Figure 4.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the initial view of the web map with indications for interactivity: the panel zoom at the top left, and the panel layers at the top right (superimposed is an image showing some of the contained layers). Records appear clustered in six large groups based on reciprocal distance. Numbers at the center of the groups indicate the records associated with each cluster.
Figure 4.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the initial view of the web map with indications for interactivity: the panel zoom at the top left, and the panel layers at the top right (superimposed is an image showing some of the contained layers). Records appear clustered in six large groups based on reciprocal distance. Numbers at the center of the groups indicate the records associated with each cluster.

Regarding interactivity, the most crucial feature is probably the panel layers (Figure 4). A total of 98 layers are featured, one of which represents the reference map (ESRI satellite). All the other layers, except one, can effectively express their contents only if activated in conjunction with the reference map (e.g., Figure 5 and Figure 7). The exception is the layer containing the summary data of each Italian region, which can fully show its information even in the absence of the reference map (see below).
For this research, the comprehensive layer Italy-All-Species assumes particular relevance. Activating this layer, in conjunction with the reference map, allows for the visualization of all the vector points within Italian political boundaries, i.e., it visualizes all the points/locations in which gastrotrich species have been found.
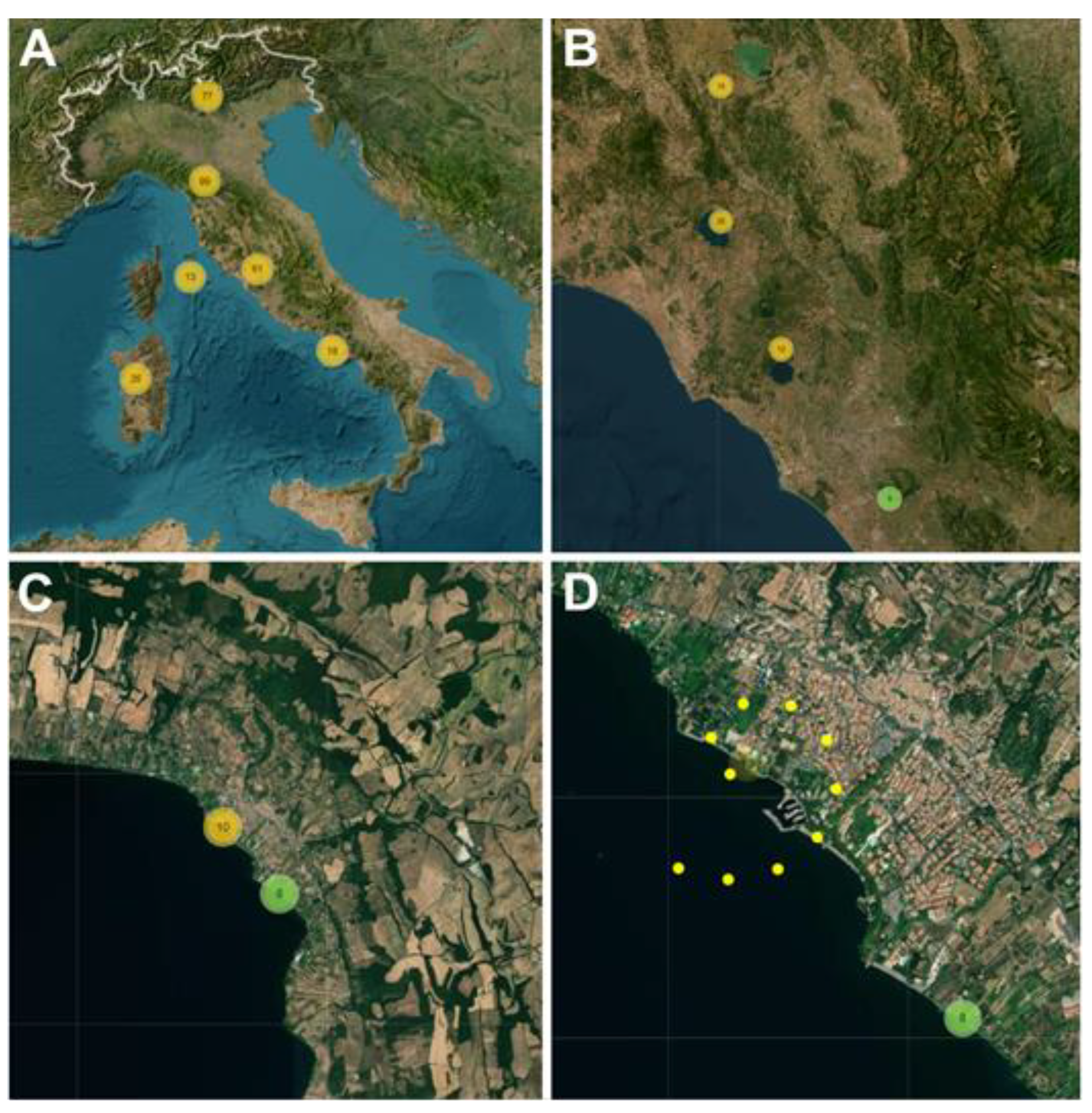
Since it was chosen the points to be shown clustered, the initial screen shows six large clusters in which the vector points are grouped based on their mutual geographical distance (Figure 4). Progressive zooming moves to increasingly smaller clusters until one can visualize the individual sampling points (Figure 5A–C).
Figure 5.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshots of the web map with the layers Italy-All-Species and the reference map active. The images show the vectors grouped into more or less comprehensive clusters depending on the scale (zoom), from the smallest scale (A) up to progressively larger scales (B–D). The numbers at the center of the groups indicate the records associated with the individual clusters. At the largest scale, the point (location) of interest is reached/shown and clicking it will display the associated records surrounding the point itself (D).
Figure 5.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshots of the web map with the layers Italy-All-Species and the reference map active. The images show the vectors grouped into more or less comprehensive clusters depending on the scale (zoom), from the smallest scale (A) up to progressively larger scales (B–D). The numbers at the center of the groups indicate the records associated with the individual clusters. At the largest scale, the point (location) of interest is reached/shown and clicking it will display the associated records surrounding the point itself (D).

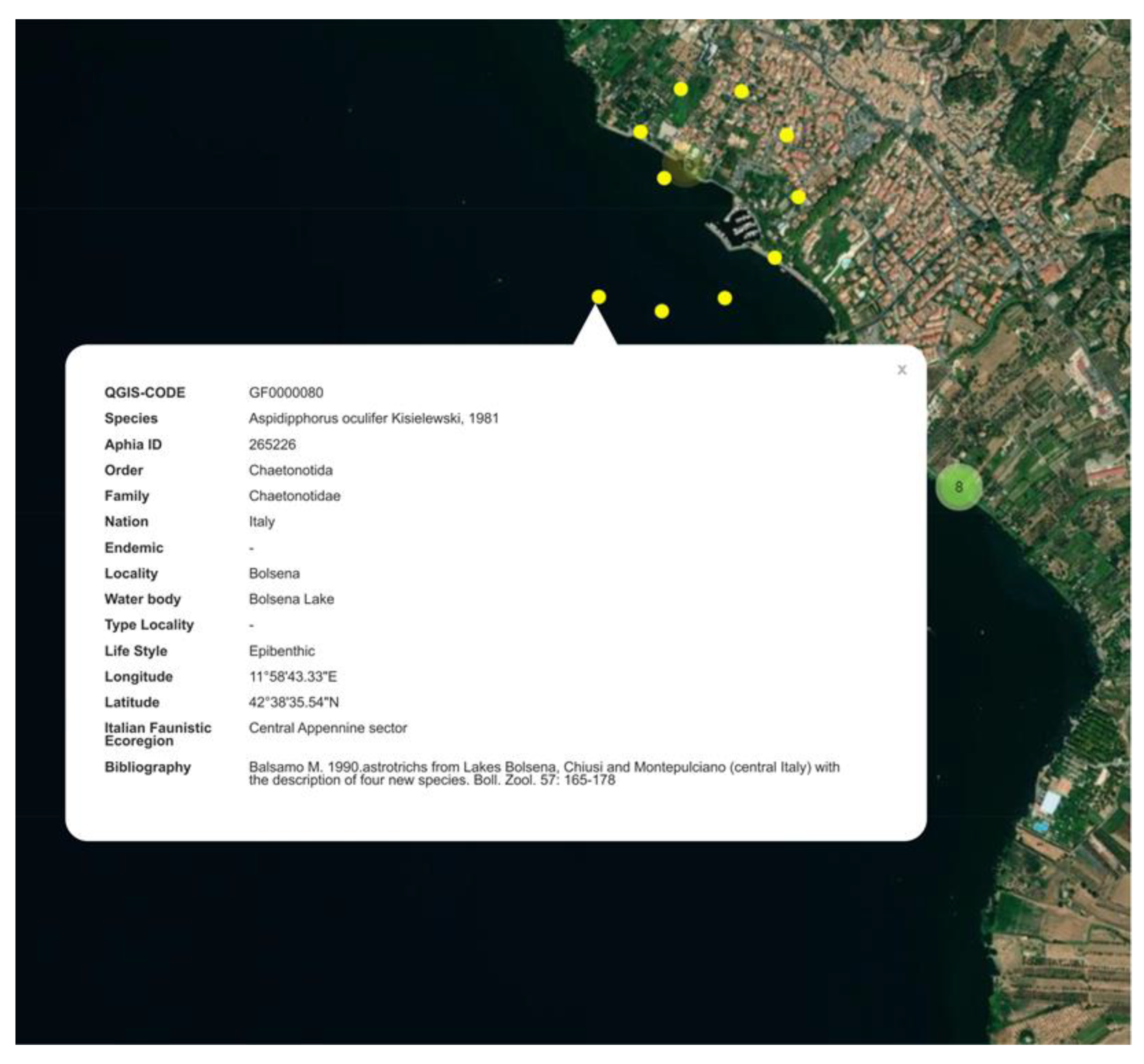
Once the point (location) of interest has been exposed, a number at the disk’s center indicates how many records (i.e., species) are associated with that location. Clicking on the disk makes it possible to “explode” the records, which will be displayed surrounding the point itself (Figure 5D). A click on the single record opens a pop-up box containing all the salient information about that record, e.g., the individual species name, locality, geographical coordinates, bibliographic reference, etc., (Figure 6). In short, the pop-up box reports the 15 attributes chosen to be made visible to the end user, as they are the most valuable. The remaining 11 attributes (see construction of the data matrix above) are visible only to the authors (i.e., us) on the QGIS desktop platform. They are meant to facilitate the housekeeping of the dataset.
Figure 6.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. The same image as Figure 5D above, showing the pop-up box associated with one of the records [34].

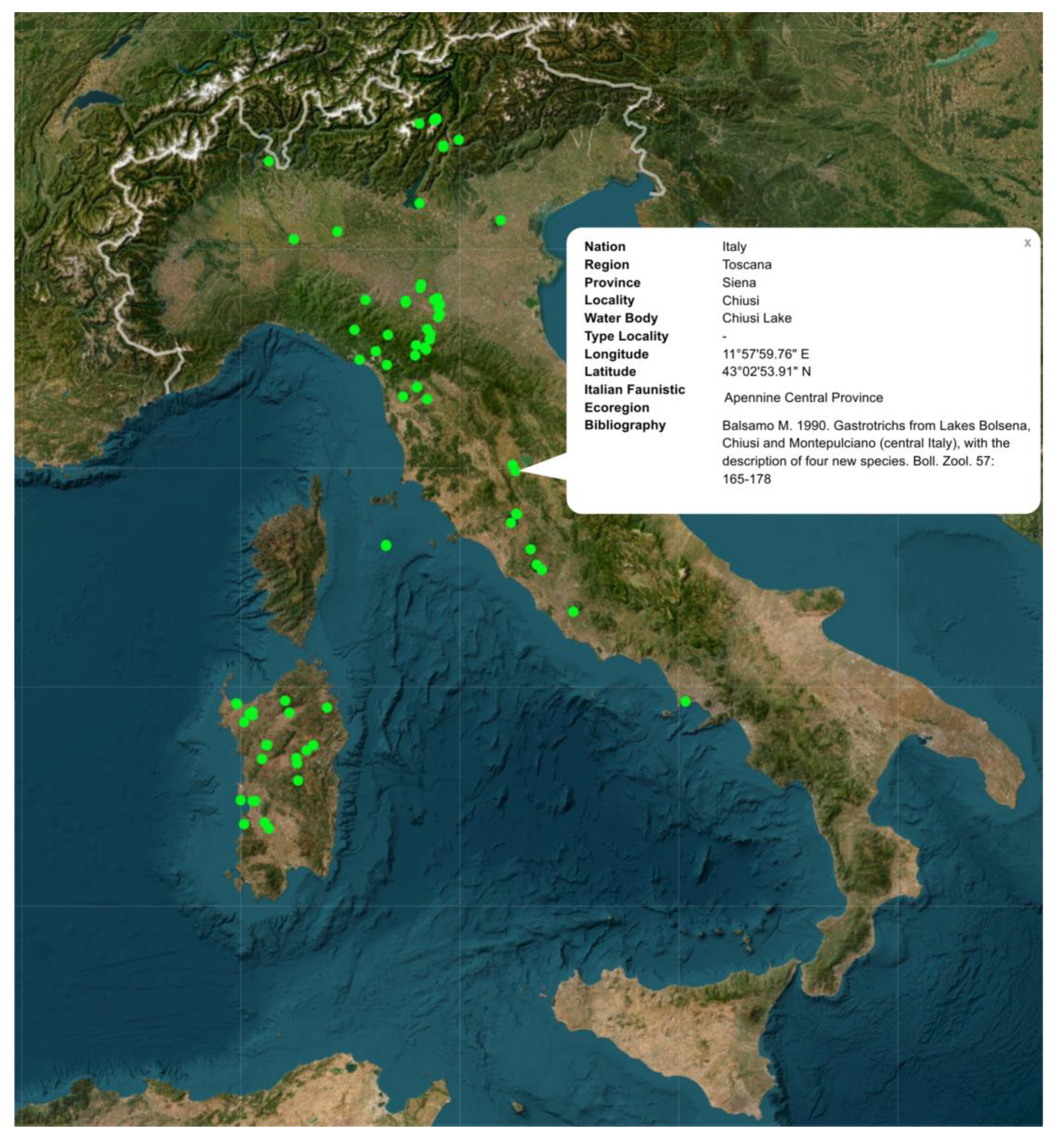
If activated, the layer Localities offer an overall glimpse of the distribution of the investigated localities in Italian territories (Figure 7). Clicking on the individual vector points activates a pop-up card with the relevant location information, including geographical and bibliographic information (Figure 7).
Figure 7.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the interactive map originating from the layer Localities, showing the distribution of the investigated locations along the national boundaries [34].
Figure 7.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the interactive map originating from the layer Localities, showing the distribution of the investigated locations along the national boundaries [34].

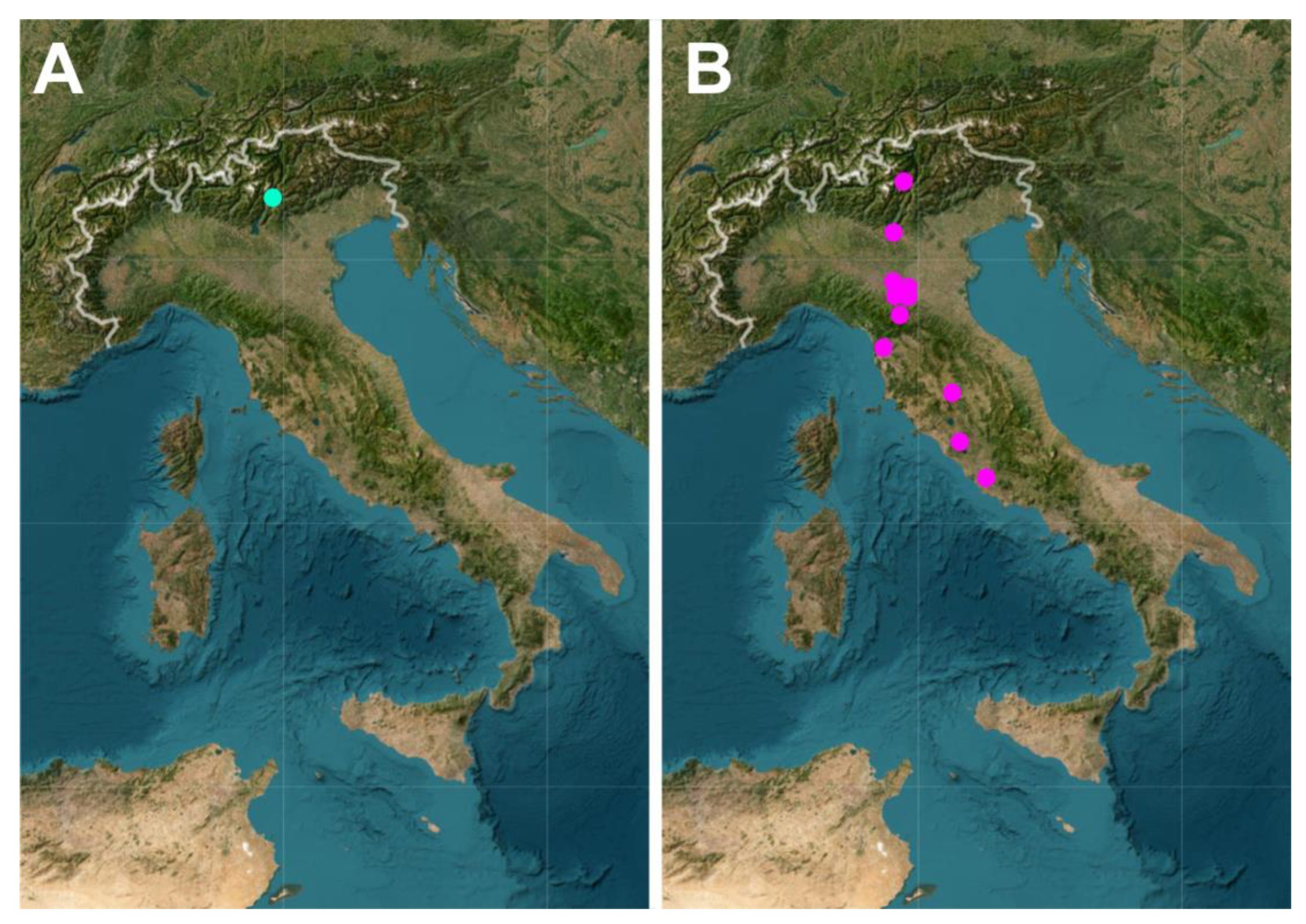
The layers dedicated to the individual species appear to be highly effective in conveying helpful information. By activating them in conjunction with the reference layer, the web map shows the known distribution in the Italian territory of the species of interest; therefore, it is possible to immediately understand whether the species is widespread or confined to a single water body. Paradigmatic examples are shown in Figure 7, where the distribution of a rare (Figure 8A) and of a relatively common species (Figure 8B) are shown.
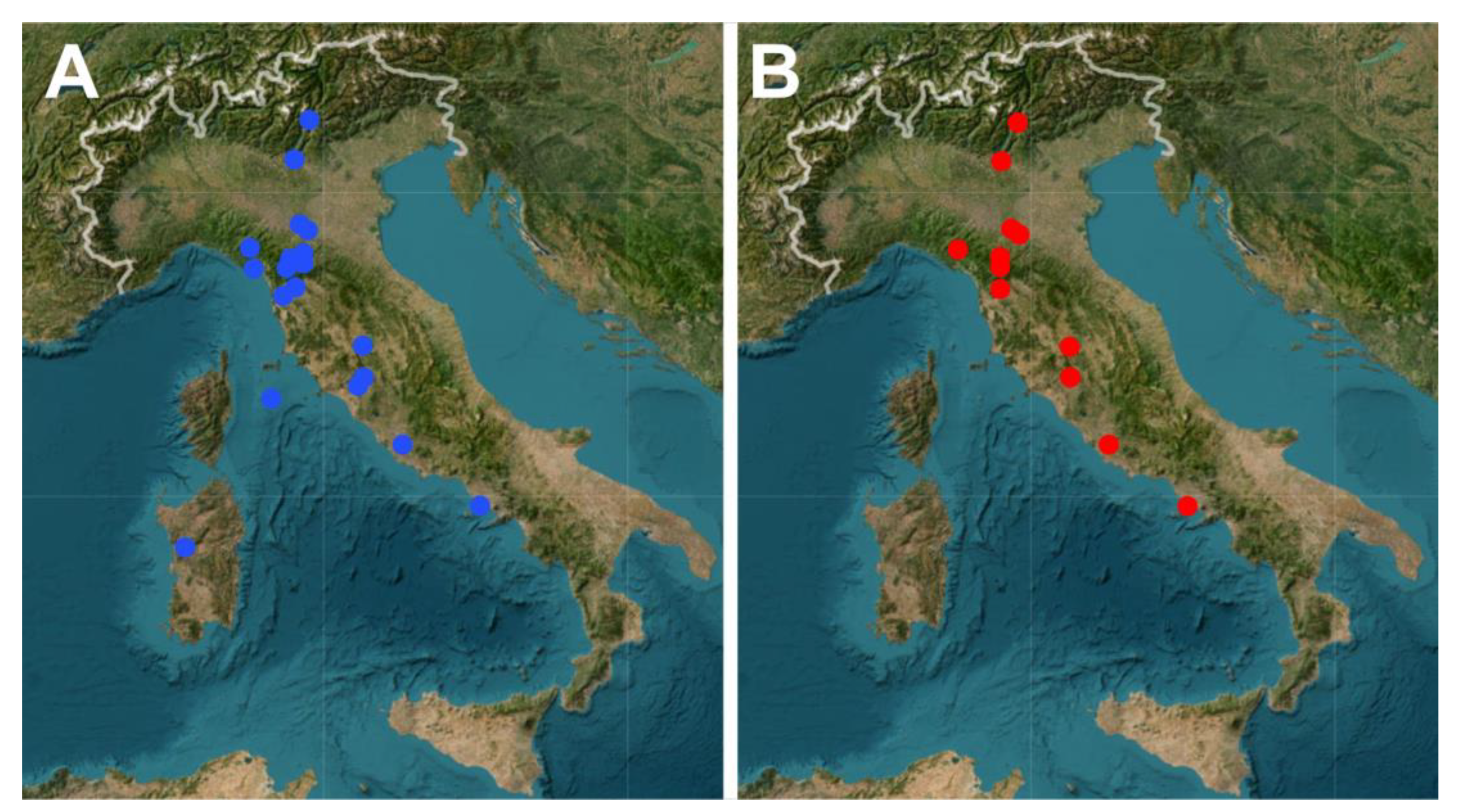
The layer Type Localities is significant because it indicates the locations where one or more species were originally described; therefore, these sites could be the focus of conservation projects (Figure 9A). Similarly, the layer Endemic Species is exceptionally informative because it indicates sites where Italian endemic species have been found; therefore, this layer can also be used to direct possible conservation efforts (Figure 9B).
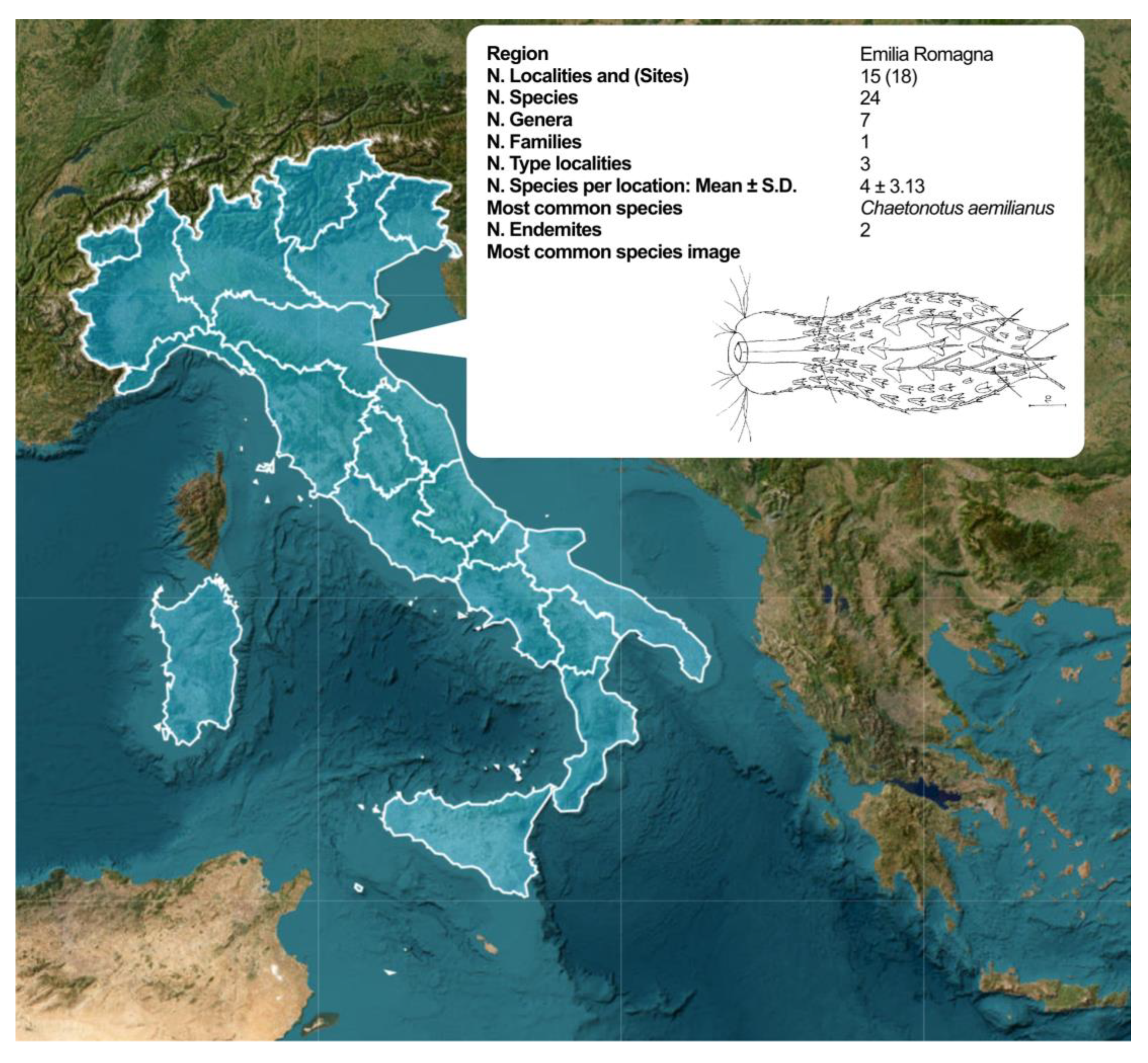
The layer Regions-Summary-Data is also of great interest because it allows for rapid access to summary information on the taxa and the sampling effort regarding the individual Italian regions. The map shows the regional borders, and each region can be activated to show a pop-up box containing information relating to the selected region, including the number of species found, locations investigated, etc. (Figure 10). The interactive map can provide all the valuable information quickly and effectively, like (and more than) a detailed scientific article.
4. Conclusions
4.1. Gastrotrichs
This research has shown that there is a need to further investigate research related to freshwater gastrotrichs in Italy. Over the course of 239 years, 25 studies have been conducted, which have led to the investigation of 61 locations across nine regions. A total of 92 species, belonging to 17 genera and three families, have been found. Out of these, 19 species are unique to Italy, while there are 17 type localities.
After comparing the data with other countries (Table 6), it is evident that Italy has a prominent freshwater gastrotrich fauna, although research has been limited to a few regions. To gain a better understanding of the biodiversity in Italy and the biogeography of its species, future investigations should focus on areas that have been poorly investigated or not investigated at all.
It would also be interesting to screen out genetically disjunct populations of widely distributed species in order to confirm the status of these taxa or, conversely, establish whether they are reproductively isolated but morphologically indistinguishable populations, thus identifying phenomena of cryptic or pseudo-cryptic speciation [58].
Table 6.
Known freshwater gastrotrichfauna in selected nations.
| Nation | N. Species | N. Genera | N. Families | Reference |
|---|---|---|---|---|
| Denmark | 27 | 6 | 2 | [59] |
| France | 36 | 10 | 3 | [60] |
| Germany | 91 | 13 | 3 | [61] |
| Italy | 91 | 17 | 3 | Present study |
| Poland | 107 | 11 | 3 | [62] |
| Romania | 90 | 15 | 3 | [63] |
| Sweden | 54 | 9 | 2 | [64] |
| South Korea | 16 | 6 | 3 | [65] |
| Canada | 39 | 10 | 2 | [66] |
| U.S.A. | 60 | 13 | 4 | [19,67,68] |
| Brazil | 98 | 18 | 4 | [16,17,69,70,71,72,73,74,75,76,77] |
4.2. QGIS and Web Map
QGIS is a compelling informatics platform that offers numerous data management and analysis prospects but requires in-depth knowledge of IT resources. However, this has yet to prevent the purpose of this study from being achieved by offering comprehensive public access, through a web map, to data on the diversity, distribution, and more, of Italian freshwater gastrotrichs. In addition to the effective dissemination of information, maps make it possible to easily highlight gaps in knowledge and offer an immediate indication of areas on the peninsula that future research should focus on, depending on specific goals.
Finally, we would like to emphasize that the map we propose can be continuously improved by adding new layers to make immediately visible information currently distributed in multiple layers (e.g., bibliography) and updated with new faunal information as it becomes available (published). Moreover, we can expand the map to include data from other nations, and even worldwide. We are committed to updating the map of Italian freshwater gastrotrichs regularly as new records are published. Additionally, we will explore possible collaborations with the Global Biodiversity Information Facility (GBIF).
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d16010017/s1, Table S1. Unpublished distributional records of Italian freshwater gastrotrichs provided by Prof. M. Balsamo. Table S2. Similarity matrix based on presence/absence data and the Bray–Curtis algorithm.
Author Contributions
Conceptualization M.A.T.; methodology, resources, data curation and writing F.S. and M.A.T.; funding acquisition M.A.T. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded under the National Recovery and Resilience Plan (NRRP), Mission 4 Component 2 Investment 1.4—Call for tender No. 3138 of 16 December 2021, rectified by Decree n. 3175 of 18 December 2021 of the Italian Ministry of University and Research funded by the European Union—NextGenerationEU. Project Code CN_00000033, Concession Decree No. 1034 of 17 June 2022 adopted by the Italian Ministry of University and Research, CUP E93C22001090001, Project title “National Biodiversity Future Center—NBFC”.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this published paper and supplementary materials.
Acknowledgments
We express our sincere gratitude to M. Balsamo for sharing information with us on localities where she found some of the species reported herein. We thank N. Bekkouche and T. Araújo for their constructive comments on an early draft of the MS.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Balsamo, M.; Grilli, P.; Guidi, L.; d’Hondt, J.L. Gastrotricha—Biology, ecology and systematics. Families Dasydytidae, Dichaeturidae, Neogosseidae, Proichthydiidae. In Identification Guides to the Plankton and Benthos of Inland Waters; Backhuys Publishers: Leiden, The Netherlands, 2014; Volume 24. [Google Scholar]
- Todaro, M.A.; Telford, M.J.; Lockyer, A.E.; Littlewood, D.T.J. Interrelationships of the Gastrotricha and Their Place among the Metazoa Inferred from 18S rRNA Genes. Zool. Scr. 2006, 35, 251–259. [Google Scholar] [CrossRef]
- Struck, T.H.; Wey-Fabrizius, A.R.; Golombek, A.; Hering, L.; Weigert, A.; Bleidorn, C.; Klebow, S.; Iakovenko, N.; Hausdorf, B.; Petersen, M.; et al. Platyzoan Paraphyly Based on Phylogenomic Data Supports a Noncoelomate Ancestry of Spiralia. Mol. Biol. Evol. 2014, 31, 1833–1846. [Google Scholar] [CrossRef] [PubMed]
- Egger, B.; Lapraz, F.; Tomiczek, B.; Müller, S.; Dessimoz, C.; Girstmair, J.; Škunca, N.; Rawlison, K.A.; Cameron, B.C.; Beli, E.; et al. A Transcriptomic-Phylogenomic Analysis of the Evolutionary Relationships of Flatworms. Curr. Biol. 2015, 25, 1347–1353. [Google Scholar] [CrossRef] [PubMed]
- Laumer, C.E.; Bekkouche, N.; Kerbl, A.; Goetz, F.; Neves, R.C.; Sørensen, M.V.; Kristensen, R.M.; Hejnol, A.; Dunn, C.W.; Giribet, G.; et al. Spiralian Phylogeny Informs the Evolution of Microscopic Lineages. Curr. Biol. 2015, 25, 2000–2006. [Google Scholar] [CrossRef]
- Fromm, B.; Tosar, J.P.; Aguilera, F.; Friedländer, M.R.; Bachmann, L.; Hejnol, A. Evolutionary Implications of the microRNA- and piRNA Complement of Lepidodermella Squamata (Gastrotricha). ncRNA 2019, 5, 19. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Kånneby, T.; Hochberg, R. Integrated Data Analysis Allows the Establishment of a New, Cosmopolitan Genus of Marine Macrodasyida (Gastrotricha). Sci. Rep. 2019, 9, 7989. [Google Scholar] [CrossRef]
- Kieneke, A.; Schmidt-Rhaesa, A. Gastrotricha. In Handbook of Zoology; De Gruyter: Berlin, Germany; München, Germany; Boston, MA, USA, 2014; Volume 3, pp. 1–134. [Google Scholar]
- Kieneke, A.; Todaro, M.A. Discovery of Two ‘Chimeric’ Gastrotricha and Their Systematic Placement Based on an Integrative Approach. Zool. J. Linn. Soc. 2021, 192, 710–735. [Google Scholar] [CrossRef]
- Hummon, W.D.; Todaro, M.A. Analytic Taxonomy and Notes on Marine, Brackish-Water and Estuarine Gastrotricha. Zootaxa 2010, 2392, 1. [Google Scholar] [CrossRef]
- Araujo, T.Q.; King-Trudeau, S.; VanDyke, J.; Hochberg, R. First Ultrastructural Description of an Apomictic Opsiblastic Egg in Freshwater Gastrotricha. J. Morphol. 2023, 285, e21659. [Google Scholar] [CrossRef]
- Giere, O. Patterns of Meiofauna Distribution. In Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments; Springer: Berlin/Heidelberg, Germany, 2009; pp. 243–265. [Google Scholar]
- Terhivuo, J.; Saura, A. Dispersal and Clonal Diversity of North-European Parthenogenetic Earthworms. Biol. Invasions 2006, 8, 1205–1218. [Google Scholar] [CrossRef]
- Ruttner-Kolisko, A. Rheomorpha neiswestnovae und Marinellina flagellata, Zwei Phylogenetisch Interessante Wurmtypen Aus Dem Susswasserpsammon. Osterr. Zool. Z. 1955, 6, 33–69. [Google Scholar]
- Kisielewski, J. Two New Interesting Genera of Gastrotricha (Macrodasyida and Chaetonotida) from the Brazilian Freshwater Psammon. Hydrobiologia 1987, 153, 23–30. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Jondelius, U.; Hochberg, R.; Hummon, W.D.; Kånneby, T.; Rocha, C.E.F. Gastrotricha: A Marine Sister for a Freshwater Puzzle. PLoS ONE 2012, 7, e31740. [Google Scholar] [CrossRef] [PubMed]
- Garraffoni, A.R.S.; Araújo, T.Q.; Lourenço, A.P.; Guidi, L.; Balsamo, M. Integrative Taxonomy of a New Redudasys Species (Gastrotricha: Macrodasyida) Sheds Light on the Invasion of Fresh Water Habitats by Macrodasyids. Sci. Rep. 2019, 9, 2067. [Google Scholar] [CrossRef] [PubMed]
- Kånneby, T.; Wicksten, M.K. First Record of the Enigmatic Genus Redudasys Kisielewski, 1987 (Gastrotricha: Macrodasyida) from the Northern Hemisphere. Zoosystema 2014, 36, 723–733. [Google Scholar] [CrossRef]
- Kånneby, T.; Kirk, J.J. A New Species of Redudasys (Gastrotricha: Macrodasyida: Redudasyidae) from the United States. Proc. Biol. Soc. 2017, 130, 128–139. [Google Scholar] [CrossRef]
- Balsamo, M.; d’Hondt, J.-L.; Kisielewski, J.; Pierboni, L. Global diversity of gastrotrichs (gastrotricha) in fresh waters. In Freshwater Animal Diversity Assessment; Balian, E.V., Lévêque, C., Segers, H., Martens, K., Eds.; Developments in Hydrobiology; Springer: Dordrecht, The Netherlands, 2008; Volume 198, pp. 85–91. [Google Scholar]
- Kisielewski, J. Inland-Water Gastrotricha from Brazil. Annal. Zool. 1991, 43, 1–168. [Google Scholar]
- Curini-Galletti, M.; Artois, T.; Delogu, V.; De Smet, W.H.; Fontaneto, D.; Jondelius, U.; Leasi, F.; Martínez, A.; Meyer-Wachsmuth, I.; Nilsson, K.S.; et al. Patterns of Diversity in Soft-Bodied Meiofauna: Dispersal Ability and Body Size Matter. PLoS ONE 2012, 7, e33801. [Google Scholar] [CrossRef]
- Corti, B. Osservazioni Microscopiche Sulla Tremella e Sulla Circolazione Del Fluido in Una Pianta Acquajuola; Rocchi: Lucca, Italy, 1774. [Google Scholar]
- Maggi, L. Primo Elenco Dei Rotiferi o Sistolidi Della Valcuvia. Atti Soc. Ital. Sci. Nat. Pavia 1878, 21, 320. [Google Scholar]
- Issel, R. Saggio Sulla Fauna Termale Italiana. Atti Accad. Sci. Torino 1901, 36, 3–15. [Google Scholar]
- Zacharias, O. Osservazioni Limnologiche Su Alcuni Laghi Italiani. Acquacolt. Lomb. 1908, 8, 5. [Google Scholar]
- Marcolongo, I. Primo Contributo Allo Studio Dei Gastrotrichi Nel Lago-Stagno Craterico Di Astroni. Monit. Zool. Ital. 1910, 21, 315–318. [Google Scholar]
- Marcolongo, I. I Gastrotrichi Del Lago-Stagno Craterico Di Astroni. Atti R. Accad. Sci. Fis. Mat. Napoli Ser. A 1914, 2, 15. [Google Scholar]
- Mola, P. Gastrotricha Delle Acque Dolci Italiane. Internat. Rev. Ges. Hydrobiol. Hydrograph. 1932, 26, 394–423. [Google Scholar] [CrossRef]
- Balsamo, M. Prime Ricerche Sui Gastrotrichi Dulciacquicoli Italiani. Atti Soc. Tosc. Sc. Nat. Mem. 1977, 84, 87–150. [Google Scholar]
- Balsamo, M. Gastrotrichi Della Toscana: Il Lago Di Sibolla. Boll. Mus. Civ. St. Nat. Verona 1980, 7, 547–571. [Google Scholar]
- Balsamo, M. Secondo Contributo Alla Conoscenza Dei Gastrotrichi Delle Acque Dolci Italiane. Atti Soc. Tosc. Sc. Nat. Mem. 1980, 87, 97–126. [Google Scholar]
- Balsamo, M. Three New Gastrotrichs from a Tuscan-Emilian Apennine Lake. Boll. Zool. 1982, 49, 287–295. [Google Scholar] [CrossRef]
- Balsamo, M. Gastrotrichs from Lakes Bolsena, Chiusi and Montepulciano (Central Italy), with the Description of Four New Species. Boll. Zool. 1990, 57, 165–178. [Google Scholar] [CrossRef]
- Fregni, E.; Balsamo, M.; Tongiorgi, P. Interstitial Gastrotrichs from Lotic Italian Fresh Waters. Hydrobiologia 1998, 368, 175–187. [Google Scholar] [CrossRef]
- Kånneby, T.; Todaro, M.A.; Jondelius, U. A Phylogenetic Approach to Species Delimitation in Freshwater Gastrotricha from Sweden. Hydrobiologia 2012, 683, 185–202. [Google Scholar] [CrossRef]
- Kånneby, T.; Todaro, M.A.; Jondelius, U. Phylogeny of Chaetonotidae and Other Paucitubulatina (Gastrotricha: Chaetonotida) and the Colonization of Aquatic Ecosystems. Zool. Scr. 2013, 42, 88–105. [Google Scholar] [CrossRef]
- Balsamo, M.; Tongiorgi, P. Gastrotricha. In Checklist delle Specie della Fauna d’Italia; Calderini: Bologna, Italy, 1995; pp. 1–11. [Google Scholar]
- Todaro, M.A.; Balsamo, M. Gastrotricha. In Checklist of the Italian Fauna; Società Italian di Biologia Marina: Genova, Italy, 2021. [Google Scholar]
- Campos, A.; Garraffoni, A.R.S. A Synopsis of Knowledge, Zoogeography and an Online Interactive Map of Brazilian Marine Gastrotrichs. PeerJ 2019, 7, e7898. [Google Scholar] [CrossRef] [PubMed]
- Grünspan, T. Beiträge Zur Systematik der Gastrotrichen. Zool. Jahrb. Abt. Syst. 1908, 26, 214–256. [Google Scholar]
- Manfredi, P. Brevi Note Intorno Alla Fauna Della Gora Di Bertonico. Ann. Biol. Lacustre 1927, 15, 110–117. [Google Scholar]
- Balsamo, M.; Kisielewski, J. Chaetonotus Fluviatilis, a New Freshwater Gastrotrich. Boll. Zool. 1986, 53, 111–114. [Google Scholar] [CrossRef]
- Balsamo, M.; Todaro, M.A. Life History Traits of Two Chaetonotids (Gastrotricha) Under Different Experimental Conditions. Invertebr. Reprod. Dev. 1988, 14, 161–176. [Google Scholar] [CrossRef]
- Bertolani, R.; Balsamo, M. Tardigradi e Gastrotrichi Del Trentino: Il Lago Di Tovel. Studi Trentini Sci. Nat. 1989, 65, 83–93. [Google Scholar]
- Balsamo, M.; Fregni, E.; Tongiorgi, P. Marine and Freshwater Gastrotricha from the Island of Montecristo (Tuscan Archipelago, Italy), with the Description of New Species. Boll. Zool. 1994, 61, 217–227. [Google Scholar] [CrossRef]
- Balsamo, M.; Fregni, E. Gastrotrichs from Interstitial Fresh Water, with a Description of Four New Species. Hydrobiologia 1995, 302, 163–175. [Google Scholar] [CrossRef]
- Balsamo, M.; Todaro, M.A. Gastrotrichi Del Trentino: Viotte Del Monte Bondone. Studi Trentini Sci. Nat. 1995, 70, 9–22. [Google Scholar]
- Leasi, F.; Rothe, B.H.; Schmidt-Rhaesa, A.; Todaro, M.A. The Musculature of Three Species of Gastrotrichs Surveyed with Confocal Laser Scanning Microscopy (CLSM). Acta Zool. 2006, 87, 171–180. [Google Scholar] [CrossRef]
- Leasi, F.; Todaro, M.A. The Muscular System of Musellifer delamarei (Renaud-Mornant, 1968) and Other Chaetonotidans with Implications for the Phylogeny and Systematization of the Paucitubulatina (Gastrotricha). Biol. J. Linn. Soc. 2008, 94, 379–398. [Google Scholar] [CrossRef]
- Schwank, P. Gastrotricha. In Susswasserfauna von Mitteleuropa; Schwoerbel, J., Zwick, P., Eds.; Gustav Fisher Verlag: Stuttgart, Germany, 1990; p. 252. [Google Scholar]
- WoRMS—World Register of Marine Species. Available online: https://www.marinespecies.org/ (accessed on 14 January 2023).
- Kolicka, M.; Dabert, M.; Olszanowski, Z.; Dabert, J. Sweet or Salty? The Origin of Freshwater Gastrotrichs (Gastrotricha, Chaetonotida) Revealed by Molecular Phylogenetic Analysis. Cladistics 2020, 36, 458–480. [Google Scholar] [CrossRef] [PubMed]
- QGIS.org. QGIS Geographic Information System. QGIS Association. 2023. Available online: http://www.qgis.org (accessed on 24 September 2023).
- ISTAT—Confini delle Unità Amministrative a Fini Statistici al 1° Gennaio. 2023. Available online: https://www.istat.it/it/archivio/222527 (accessed on 14 January 2023).
- Ehrenberg, C.G. Die infusionsthierchen als vollkommene organismen. In Ein Blick in das Tiefere Organische Leben der Natur; L. Voss: Leipzig, Germany, 1838. [Google Scholar]
- Kolicka, M. New Chaetonotus (Wolterecka semovitus sp. nov. (Gastrotricha: Chaetonotida: Chaetonotidae) from a Palm House in Vienna (Austria). Annal. Zool. 2019, 69, 447. [Google Scholar] [CrossRef]
- Rataj Križanová, F.; Vďačný, P. A Huge Undescribed Diversity of the Subgenus Hystricochaetonotus (Gastrotricha, Chaetonotidae, Chaetonotus) in Central Europe. Eur. J. Taxon. 2022, 840, 1–93. [Google Scholar] [CrossRef]
- Grilli, P.; Kristensen, R.M.; Balsamo, M. Contribution to the Knowledge of Freshwater Gastrotricha from Denmark. Steenstrupia 2010, 32, 79–92. [Google Scholar]
- Grilli, P.; d’Hondt, J.L.; Balsamo, M. Contribution à La Connaissance Des Gastrotriches (Gastrotricha) Dulcicoles Des Eaux Françaises. Bull. Soc. Zool. Fr. 2010, 135, 93–107. [Google Scholar]
- Schmidt-Rhaesa, A. German Freshwater Gastrotricha. Gastrotricha World Portal. 2006. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Kolicka, M. Polish Freshwater Gastrotricha. Gastrotricha World Portal. 2015. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Rudescu, L. Gastrotricha; Academiei Republicii Socialiste România: Bucharest, Romania, 1967. [Google Scholar]
- Kånneby, T. Swedish Freshwater Gastrotricha. Gastrotricha World Portal. 2014. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Lee, J.M. Biodiversity of Gastrotricha from Korea. In Proceedings of the Joint Academic Conference of the Korea Association of Marine Science and Technology, Jeju-si, Republic of Korea, 21–23 May 2015. [Google Scholar]
- Kånneby, T. Canadian Freshwater Gastrotricha. Gastrotricha World Portal. 2014. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Kånneby, T.; Weiss, M.J. U.S. Freshwater Gastrotricha. Gastrotricha World Portal. 2016. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Kirk, J.J. Lepidodermella weissi, New Species (Gastrotricha: Chaetonotida: Chaetonotidae) from Northwestern Oregon, U.S.A. Proc. Biol. Soc. Wash. 2021, 134, 116–126. [Google Scholar] [CrossRef]
- Bizzarri, N.; Todaro, M.A. Brazilian Freshwater Gastrotricha. Gastrotricha World Portal. 2010. Available online: http://www.gastrotricha.unimore.it/ (accessed on 5 October 2023).
- Garraffoni, A.R.S.; Araújo, T.Q. Chave de Identificação de Gastrotricha de Águas Continentais e Marinhas Do Brasil. Pap. Avulsos Zool. 2010, 50, 535–552. [Google Scholar] [CrossRef]
- Garraffoni, A.R.S.; Melchior, M.P. New Species and New Records of Freshwater Heterolepidoderma (Gastrotricha: Chaetonotidae) from Brazil with an Identification Key to the Genus. Zootaxa 2015, 4057, 551. [Google Scholar] [CrossRef] [PubMed]
- Minowa, A.K.; Garraffoni, A.R.S. A New Species of Haltidytes Remane, 1936 (Gastrotricha: Chaetonotida: Dasydytidae) from an Urban Lagoon in Brazil with a Phylogenetic Reconstruction of the Genus Based on Morphological Data. Zool. Anz. 2017, 269, 100–109. [Google Scholar] [CrossRef]
- Garraffoni, A.R.S.; Araújo, T.Q.; Lourenço, A.P.; Guidi, L.; Balsamo, M. A New Genus and New Species of Freshwater Chaetonotidae (Gastrotricha: Chaetonotida) from Brazil with Phylogenetic Position Inferred from Nuclear and Mitochondrial DNA Sequences. Syst. Biodivers. 2017, 15, 49–62. [Google Scholar] [CrossRef]
- Minowa, A.K.; Garraffoni, A.R.S. Assessing Biodiversity Shortfalls of Freshwater Meiofauna from the Atlantic Forest: New Species, Distribution Patterns and the First Total-Evidence Phylogeny of Semiplanktonic Gastrotricha. Mol. Phylogenet. Evol. 2020, 152, 106926. [Google Scholar] [CrossRef]
- Guidetti, E.B.; Campos, A.; Batistão, A.R.; Silva, A.T.D.; Bilatto, C.G.; Salgado, K.A.; Araújo, T.Q.; Garraffoni, A.R.S. Gastrotrichs and Tardigrades in a Remnant of Atlantic Forest (Serra Do Japi, SP, Brazil). Biota Neotrop. 2021, 21, e20201165. [Google Scholar] [CrossRef]
- Magpali, L.; Machado, D.R.P.; Araújo, T.Q.; Garraffoni, A.R.S. Long Distance Dispersal and Pseudo-Cryptic Species in Gastrotricha: First Description of a New Species (Chaetonotida, Chaetonotidae, Polymerurus) from an Oceanic Island with Volcanic Rocks. Eur. J. Taxon. 2021, 746, 62–93. [Google Scholar] [CrossRef]
- Minowa, A.; Senna Garraffoni, A.R. Seek and You Shall Find: New Species of the Rare Genus Ornamentula (Gastrotricha: Chaetonotida) and First Record Outside of Type-Locality. Zoologia 2021, 38, 1–9. [Google Scholar] [CrossRef]
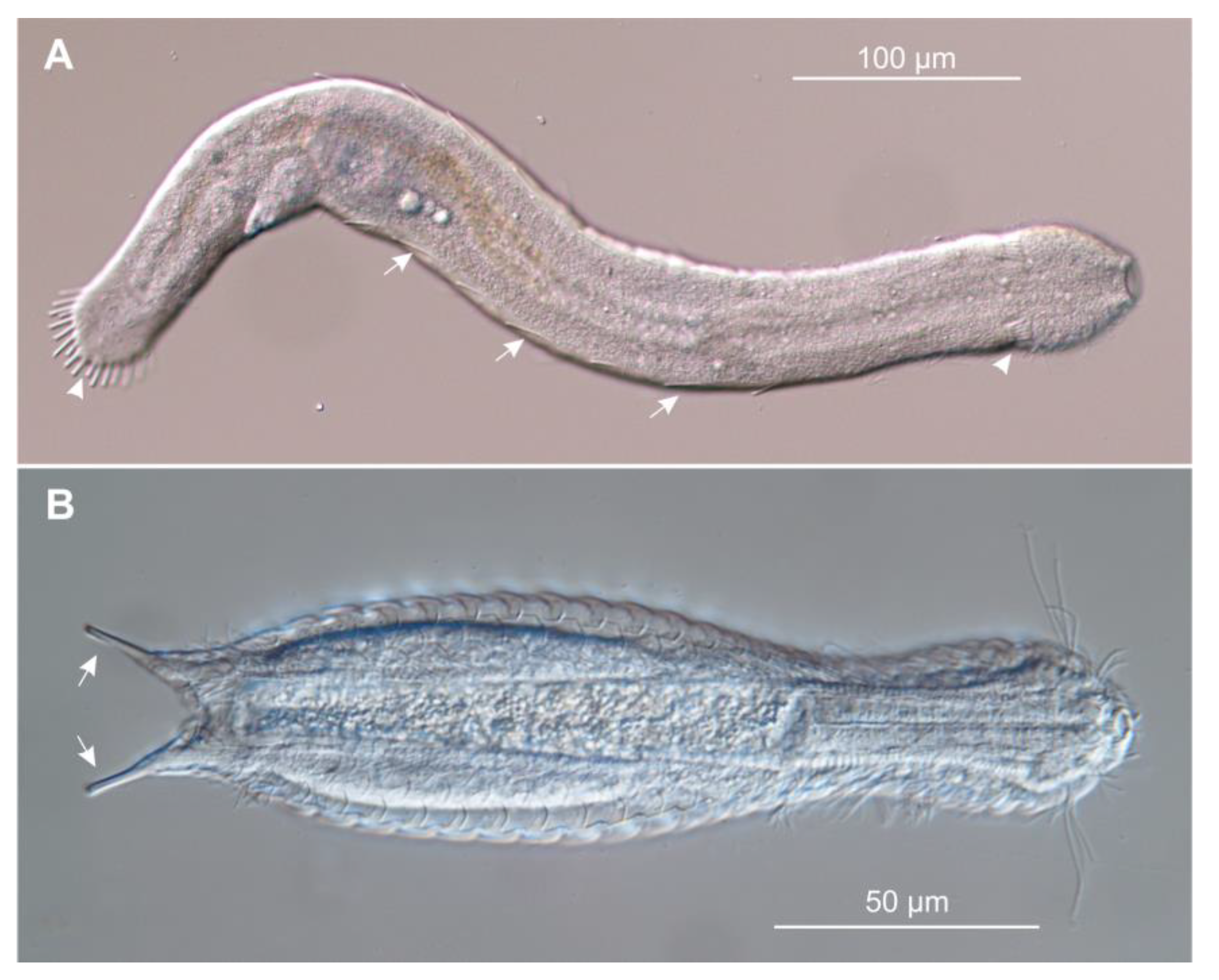
Figure 1.
Photomicrographs of gastrotrichs belonging to the two orders: (A) Cephalodasys sp., order Macrodasyida—notice the elongate body shape with several adhesive tubes along the sides (arrows) and at the anterior and posterior regions (arrowheads); and (B) Lepidodermella sp., order Chaetonotida—notice the peculiar tenpin body shape with the furcate caudum showing a single adhesive tube at the end of each furcal branch (arrows). Differential interference contrast microscopy (DIC).
Figure 1.
Photomicrographs of gastrotrichs belonging to the two orders: (A) Cephalodasys sp., order Macrodasyida—notice the elongate body shape with several adhesive tubes along the sides (arrows) and at the anterior and posterior regions (arrowheads); and (B) Lepidodermella sp., order Chaetonotida—notice the peculiar tenpin body shape with the furcate caudum showing a single adhesive tube at the end of each furcal branch (arrows). Differential interference contrast microscopy (DIC).

Figure 8.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha. Maps showing the distribution of individual species: (A) Chaetonotus arethusae, with distribution limited to a single biotope; and (B) Chaetonotus aemilianus, with a wide distribution.
Figure 8.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha. Maps showing the distribution of individual species: (A) Chaetonotus arethusae, with distribution limited to a single biotope; and (B) Chaetonotus aemilianus, with a wide distribution.

Figure 9.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha: screenshots of the map showing the distribution of type localities (A); and type locality of the endemic species (B).
Figure 9.
Examples of the interactive map dedicated to the Italian freshwater Gastrotricha: screenshots of the map showing the distribution of type localities (A); and type locality of the endemic species (B).

Figure 10.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the interactive map originating from the layer Regions-Summary-Data; by clicking on one of the investigated regions (in the example the Emilia Romagna region), a pop-up box appears with the summary data relating to the investigated locations and the species found, as well as the most common species at the regional level and a sketch.
Figure 10.
Example of the interactive map dedicated to the Italian freshwater Gastrotricha. Screenshot of the interactive map originating from the layer Regions-Summary-Data; by clicking on one of the investigated regions (in the example the Emilia Romagna region), a pop-up box appears with the summary data relating to the investigated locations and the species found, as well as the most common species at the regional level and a sketch.

Table 3.
List of Italian freshwater Gastrotricha.
| Order CHAETONOTIDA Remane, 1925 Rao & Clausen, 1970 |
| Suborder PAUCITUBULATINA d’Hondt, 1971 |
| Family Chaetonotidae Zelinka, 1889 |
| Genus Aspidiophorus Voigt, 1904 |
| Aspidiophorus heterodermus Saito, 1937 |
| Aspidiophorus oculifer Kisielewski, 1981 |
| Aspidiophorus ophiodermus Balsamo, 1982 |
| Aspidiophorus paradoxus (Voigt, 1902) |
| Aspidiophorus squamulosus Roszczak, 1936 |
| Genus Chaetonotus Ehrenberg, 1830 |
| Chaetonotus acanthocephalus Valkanov, 1937 |
| Chaetonotus acanthodes Stokes, 1887 |
| Chaetonotus acanthophorus Stokes, 1887 |
| Chaetonotus aemilianus Balsamo, 1978 |
| Chaetonotus arethusae Balsamo & Todaro, 1995 |
| Chaetonotus balsamoe Kisielewski, 1997 |
| Chaetonotus benacensis Balsamo & Fregni, 1995 |
| Chaetonotus bisacer Greuter, 1917 |
| Chaetonotus brachyurus Balsamo, 1981 |
| Chaetonotus brevispinosus Zelinka, 1889 |
| Chaetonotus cestacanthus Balsamo, 1990 |
| Chaetonotus chuni Voigt, 1901 |
| Chaetonotus daphnes Balsamo & Todaro, 1995 |
| Chaetonotus decemsetosus Marcolongo, 1910 |
| Chaetonotus disjunctus Greuter, 1917 |
| Chaetonotus dracunculus Balsamo, 1990 |
| Chaetonotus enormis Stokes, 1887 |
| Chaetonotus fluviatilis Balsamo & Kisielewski, 1986 |
| Chaetonotus heideri Brehm, 1917 |
| Chaetonotus heterospinosus Balsamo, 1978 |
| Chaetonotus hirsutus Marcolongo, 1910 |
| Chaetonotus hystrix Metschnikoff, 1865 |
| Chaetonotus italicus Balsamo & Todaro, 1995 |
| Chaetonotus laroides Marcolongo, 1910 |
| Chaetonotus larus (Müller, 1773) |
| Chaetonotus longispinosus Stokes, 1887 |
| Chaetonotus lunatospinosus Balsamo, 1981 |
| Chaetonotus macrochaetus Zelinka, 1889 |
| Chaetonotus maximus Ehrenberg, 1830 |
| Chaetonotus microchaetus Preobrajenskaia, 1926 |
| Chaetonotus minimus Marcolongo, 1910 |
| Chaetonotus multispinosus Grünspan, 1908 |
| Chaetonotus mutinensis Balsamo, 1978 |
| Chaetonotus naiadis Balsamo & Todaro, 1995 |
| Chaetonotus octonarius Stokes, 1887 |
| Chaetonotus oculifer Kisielewski, 1981 |
| Chaetonotus oplites Balsamo & Fregni, 1995 |
| Chaetonotus paucisetosus Marcolongo, 1910 |
| Chaetonotus pentacanthus Balsamo, 1981 |
| Chaetonotus persetosus Zelinka, 1889 |
| Chaetonotus polyspinosus Greuter, 1917 |
| Chaetonotus poznaniensis Kisielewski, 1981 |
| Chaetonotus pungens Balsamo, 1990 |
| Chaetonotus robustus Davison, 1938 |
| Chaetonotus schultzei Metchnikoff, 1865 |
| Chaetonotus similis Zelinka, 1889 |
| Chaetonotus simrothi Voigt, 1909 |
| Chaetonotus sphagnophilus Kisielewki, 1981 |
| Chaetonotus spinulosus Stokes, 1887 |
| Chaetonotus succinctus Voigt, 1901 |
| Chaetonotus trispinosus Balsamo, 1990 |
| Chaetonotus uncinus Voigt, 1902 |
| Genus Fluxiderma d’Hondt, 1974 |
| Fluxiderma concinnum (Stokes, 1887) |
| Genus Heterolepidoderma Remane, 1927 |
| Heterolepidoderma brevitubulatum Kisielewski, 1981 |
| Heterolepidoderma gracile Remane, 1927 |
| Heterolepidoderma lamellatum Balsamo & Fregni, 1995 |
| Heterolepidoderma macrops Kisielewski, 1981 |
| Heterolepidoderma majus Remane, 1927 |
| Heterolepidoderma multiseriatum Balsamo, 1978 |
| Heterolepidoderma ocellatum (Metschnikoff, 1865) |
| Heterolepidoderma pineisquamatum Balsamo, 1981 |
| Genus Ichthydium Ehrenberg, 1830 |
| Ichthydium diacanthum Balsamo & Todaro, 1995 |
| Ichthydium forcipatum Voigt, 1901 |
| Ichthydium forficula Remane, 1927 |
| Ichthydium fossae d’Hondt, 1971 |
| Ichthydium plicatum Balsamo & Fregni, 1995 |
| Ichthydium podura (Müller, 1773) |
| Ichthydium squamigerum Balsamo & Fregni, 1995 |
| Ichthydium tanytrichum Balsamo, 1983 |
| Genus Lepidochaetus Kisielewski, 1991 |
| Lepidochaetus zelinkai (Grünspan, 1908) |
| Genus Lepidodermella Blake, 1933 |
| Lepidodermella squamata (Dujardin, 1841) |
| Genus Polymerurus Remane, 1927 |
| Polymerurus nodicaudus (Voigt, 1901) |
| Polymerurus nodifurca (Marcolongo, 1910) |
| Polymerurus rhomboides (Stokes, 1887) |
| Polymerurus serraticaudus (Voigt, 1901) |
| Genus Romballichthys Schwank, 1990 |
| Romballichthys punctatus (Greuter, 1917) |
| Family Dasydytidae Daday, 1905 |
| Genus Anacanthoderma Marcolongo, 1910 |
| Anacanthoderma paucisetosum (Marcolongo, 1910) |
| Anacanthoderma punctatum Marcolongo, 1910 |
| Genus Chitonodytes Remane, 1967 |
| Chitonodytes longisetosus (Metschnikoff, 1865) |
| Genus Dasydytes Gosse, 1851 |
| Dasydytes ornatus Voigt, 1909 |
| Genus Haltidytes Remane, 1936 |
| Haltidytes crassus (Greuter, 1917) |
| Haltidytes saltitans (Stokes, 1887) |
| Genus Setopus Grünspan, 1908 |
| Setopus tongiorgii (Balsamo, 1983) |
| Genus Stylochaeta Hlava, 1904 |
| Stylochaeta fusiformis (Spencer, 1890) |
| Stylochaeta stylifera (Voigt, 1901) |
| Family Dichaeturidae Remane, 1927 |
| Genus Dichaetura Lauterborn, 1913 |
| Dichaetura capricornia (Metschnikoff, 1865) |
| Family Neogosseidae Remane, 1927 |
| Genus Neogossea Remane, 1927 |
| Neogossea antennigera (Gosse, 1851) |
Table 4.
Summary data by Italian regions regarding the freshwater Gastrotricha. Number of investigated localities (and sites), taxa, and type localities.
Table 4.
Summary data by Italian regions regarding the freshwater Gastrotricha. Number of investigated localities (and sites), taxa, and type localities.
| Region | N. Localities and (Sites) | N. Species | N. Genera | N. Families | N. Type Localities |
|---|---|---|---|---|---|
| Abruzzo | - | - | - | - | - |
| Basilicata | - | - | - | - | - |
| Calabria | - | - | - | - | - |
| Campania | 1 (1) | 18 | 4 | 2 | 1 |
| Emilia Romagna | 14 (18) | 28 | 8 | 1 | 3 |
| Friuli Venezia Giulia | - | - | - | - | - |
| Lazio | 5 (7) | 22 | 7 | 1 | 3 |
| Liguria | 1 (1) | 2 | 1 | 1 | 0 |
| Lombardia | 3 (3) | 12 | 5 | 2 | 0 |
| Marche | - | - | - | - | - |
| Molise | - | - | - | - | - |
| Piemonte | - | - | - | - | - |
| Puglia | - | - | - | - | - |
| Sardegna | 21 (24) | 34 | 13 | 3 | 0 |
| Sicilia | - | - | - | - | - |
| Toscana | 10 (12) | 41 | 11 | 2 | 5 |
| Trentino Alto Adige | 4 (14) | 27 | 9 | 2 | 4 |
| Umbria | - | - | - | - | - |
| Valle d’Aosta | - | - | - | - | - |
| Veneto | 2 (2) | 15 | 4 | 1 | 1 |
| Italy | 61 (81) | 92 | 17 | 3 | 17 |
Table 5.
Italian regions with summary data regarding freshwater Gastrotricha. Number of species per locality, most common species, and number of endemites.
Table 5.
Italian regions with summary data regarding freshwater Gastrotricha. Number of species per locality, most common species, and number of endemites.
| Region | N. Specie per Locality (mean ± s.d.) | Most Common Species (% Localities) | N. Endemites |
|---|---|---|---|
| Abruzzo | - | - | - |
| Basilicata | - | - | - |
| Calabria | - | - | - |
| Campania | 18 ± N.A. | N.A. | 4 |
| Emilia Romagna | 4.57 ± 3.08 | C. aemilianus (12.5%) | 3 |
| Friuli Venezia Giulia | - | - | - |
| Lazio | 9.40 ± 5.50 | L. squamata (14.9%) | 1 |
| Liguria | 2 ± N.A. | N.A. | 0 |
| Lombardia | 4.67 ± 5.51 | N.A. | 0 |
| Marche | - | - | - |
| Molise | - | - | - |
| Piemonte | - | - | - |
| Puglia | - | - | - |
| Sardegna | 1.81 ± 0.93 | N.A. | 0 |
| Sicilia | - | - | - |
| Toscana | 7.60 ± 6.24 | N.A. | 3 |
| Trentino Alto Adige | 12.75 ± 15.24 | C. hystrix (13.7%) | 0 |
| Umbria | - | - | - |
| Valle d’Aosta | - | - | - |
| Veneto | 8 ± 9.90 | H. ocellatum (12.5%) | 1 |
| Italy | 5.33 ± 6.13 | L. squamata (8.4%) | 19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Saponi, F.; Todaro, M.A. Status of the Italian Freshwater Gastrotricha Biodiversity, with the Creation of an Interactive GIS-Based Web Map. Diversity 2024, 16, 17. https://doi.org/10.3390/d16010017
AMA Style
Saponi F, Todaro MA. Status of the Italian Freshwater Gastrotricha Biodiversity, with the Creation of an Interactive GIS-Based Web Map. Diversity. 2024; 16(1):17. https://doi.org/10.3390/d16010017
Chicago/Turabian StyleSaponi, Francesco, and M. Antonio Todaro. 2024. "Status of the Italian Freshwater Gastrotricha Biodiversity, with the Creation of an Interactive GIS-Based Web Map" Diversity 16, no. 1: 17. https://doi.org/10.3390/d16010017
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.