



Does Size Matter? Two Subterranean Biodiversity Hotspots in the Lessini Mountains in the Veneto Prealps in Northern Italy
Museo di Storia Naturale of Verona, Lungadige Porta Vittoria, 9, 37129 Verona, Italy
Diversity 2024, 16(1), 25; https://doi.org/10.3390/d16010025
Submission received: 6 October 2023
/
Revised: 14 December 2023
/
Accepted: 26 December 2023
/
Published: 30 December 2023
(This article belongs to the Special Issue Hotspots of Subterranean Biodiversity—2nd Volume)
Abstract
:In the Lessini Mountains, the southernmost prealpine area in the Veneto region, thousands of caves are found, many of which have been extensively studied from the biological point of view. Numerous studies have been carried out on taxonomic and biogeographic aspects over the last hundred years. Two caves, in particular, have been found to be extremely rich in species adapted to life in subterranean environments. These are the Arena Cave in the Monti Lessini Veronesi and the Buso della Rana cave system in the Monti Lessini Vicentini. The two caves have extremely different development: Arena Cave is about 100 meters in length, and the Buso della Rana-Pisatela cave system is more than 37 km in length. Despite this huge difference in size, they both have the highest number of subterranean dwelling species in northern Italy (16 troglobionts and 8 stygobionts in Arena Cave, and 7 troglobionts and 11 stygobionts in the Buso della Rana-Pisatela cave system).
1. Introduction
The Lessini Mountains, located in the western part of the Veneto Prealps (southeastern Italian Alps) [1], extend over a total area of 1403 km2 with a maximum elevation of 1865 m a.s.l. They are bordered by the Adige Valley to the west and the Leogra Valley to the east and northeast. The Val dei Ronchi separates the range from the Pasubio-Carega Group to the northwest. From the geological point of view, the Lessini Mountains are dominated by limestones from the Mesozoic and Cenozoic ages, interspersed by Cenozoic volcanic rocks and Eocenic limestone outcrops [2]. Lessinia includes the western Lessini Mountains (between the Adige and Illasi Valleys) and eastern Lessinia Mountains (between the Illasi and Leogra Valleys). The western Lessini Mountains (Dolomia Principale, Calcari grigi, Rosso Ammonitico, Scaglia Rossa, and Maiolica) mainly consist of carbonate rocks; some areas of the eastern Lessinia Mountains are primarily composed of volcanic rocks developed during the Venetian Tertiary magmatism [2].
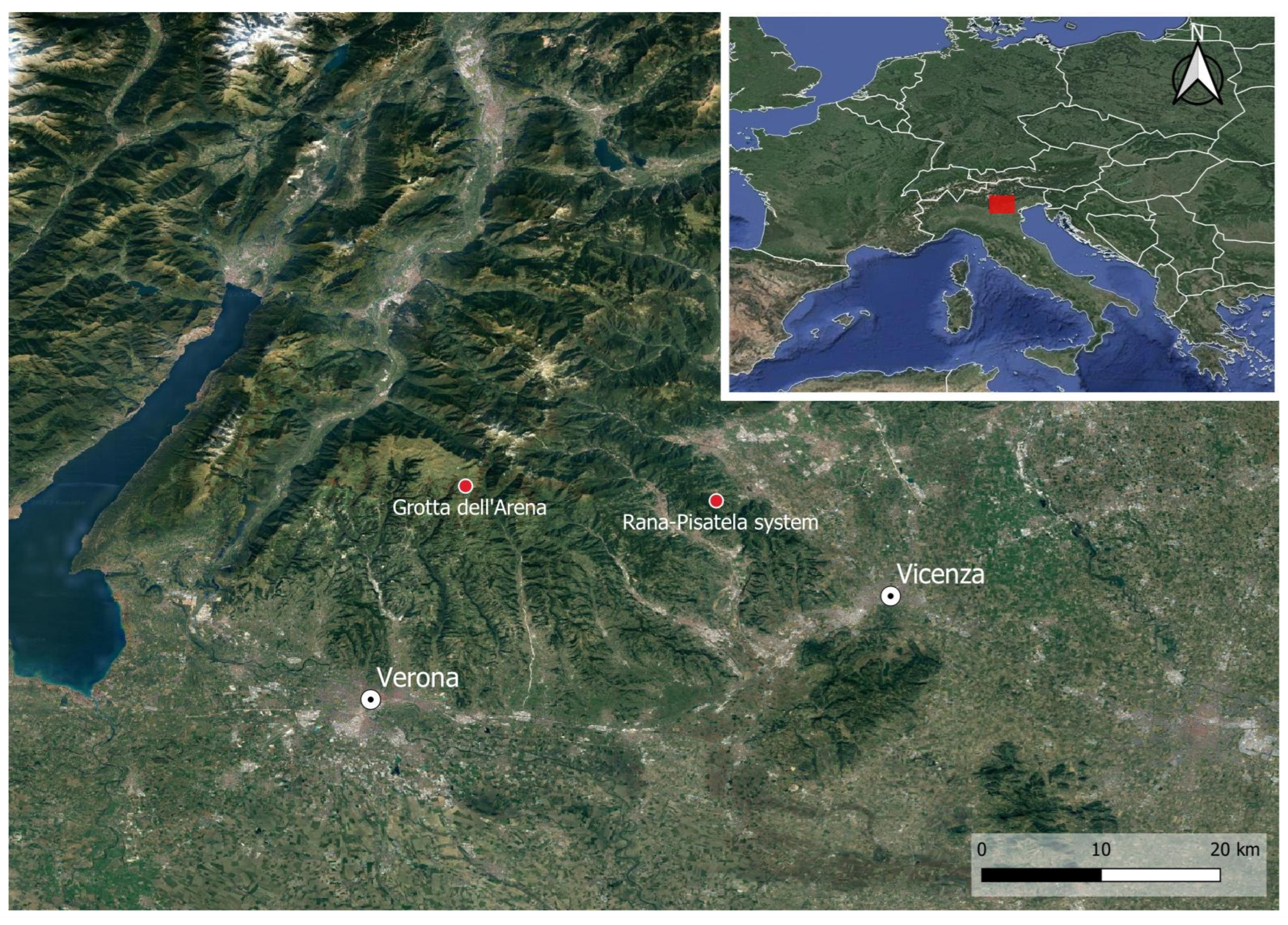
In this mountain range, two caves have a high number of obligate subterranean species: Arena Cave (having 16 troglobionts and 8 stygobionts), which is in the Central-Western Lessini Mountains, and the Buso della Rana-Pisatella cave system (having 7 troglobionts and 11 stygobionts), in the Eastern Lessini Mountains [3,4]. The two caves are located 22 km apart as the crow flies (Figure 1).
Despite the short distance between the two caves, which are situated in the same mountain range, they show enormous differences in development, rock formations, and fauna composition, making the separate analysis and comparison of the two caves interesting. Especially evident is the huge difference in the length of the two caves. This difference in size would suggest that the larger and more diversified one has a greater richness of cave-dwelling species. To verify this hypothesis, we compare here, the number of subterranean species present in each of the two caves. However, the two caves have one important characteristic in common: both can be considered cases of contact karst caves, i.e., karst phenomena and forms influenced by the contact between two or more karstifiable rocks that differ in some characteristics, such as porosity, chemical composition and fracture density, or a karstifiable rock and a non-karstifiable rock [5].
1.1. Arena Cave (476 V/VR)
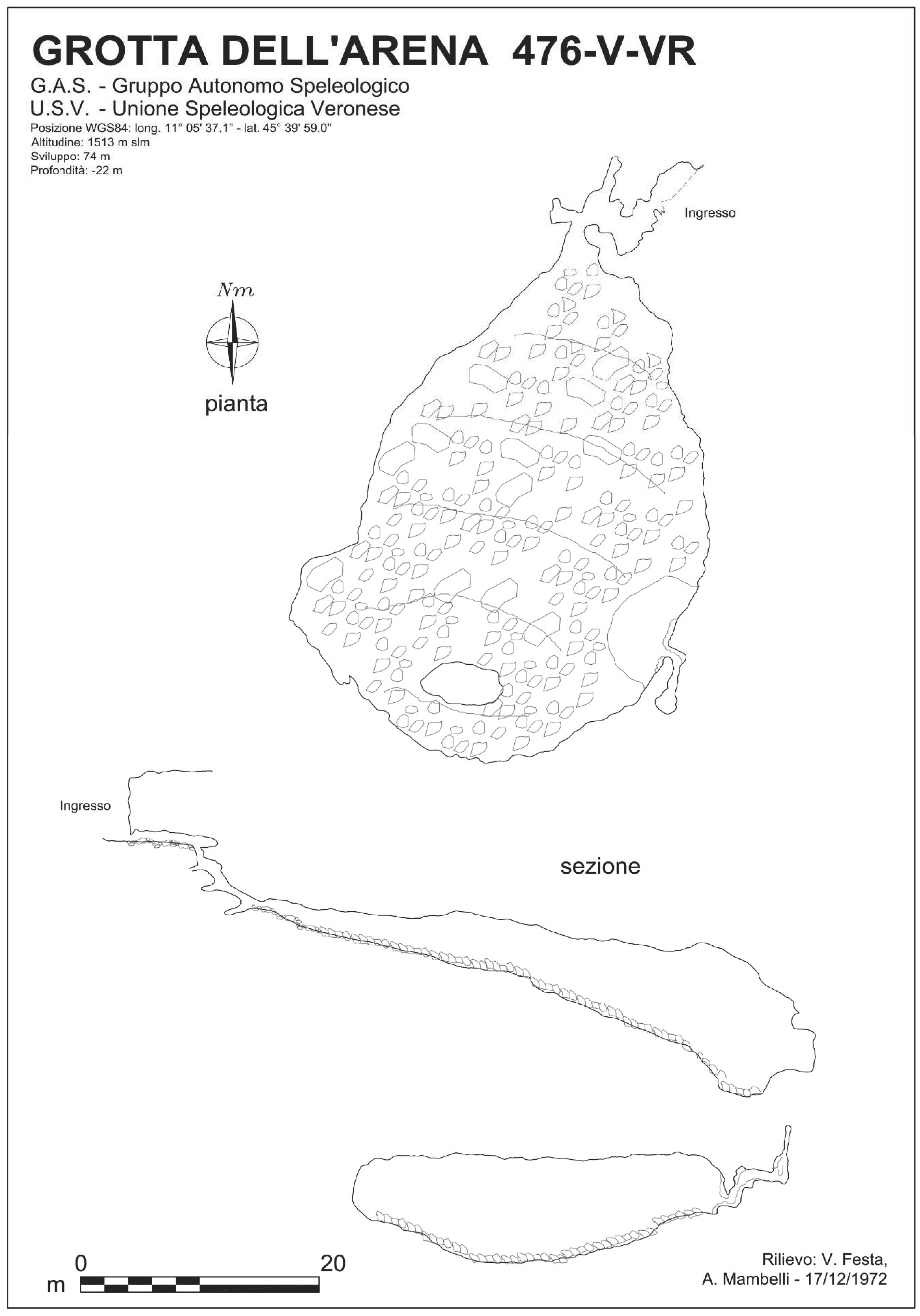
Arena Cave is registered as number 476 in the Cadastre of the Caves of Veneto Region. It is located in the Province of Verona, municipality of Bosco Chiesanuova, in the Malga Bagorno area. Its location, at 11°06′02″ E 45°39′56″ N, has an altitude of 1512 m a.s.l. (Figure 2).
The cave is 74 m long with a difference in elevation of −22 m from the entrance to the bottom. It is formed by a large chamber, roughly elliptical in plane section, with a main diameter of about 50 m. The roof coincides mostly with bedding planes. The southern part of the floor is characterized by a large, asymmetrical, funnel-shaped depression, a type of subterranean doline that developed in the collapse of debris [6] (Figure 3). The chamber is connected to the surface through some narrow passages that start from an open collapse depression located on a slope, which resembles a Roman theatre (i.e., an “Arena”, hence the name of the cave).
From the geological point of view, the cave is an expression of a contact karst, where different limestone types are in contact both stratigraphically and along tectonic structures [5,7,8]. The limestone formations present here are “Calcari del Gruppo di San Vigilio” of lower-middle Jurassic, about 60 m in depth, both pure Oolitic and bio-sparitic/–ruditic, or reef limestones, relatively densely fractured; “Rosso Ammonitico”, a condensed rock unit of middle-upper Jurassic age, about 30 m in depth, made up of nodular micritic limestone that is very resistant to erosion, crossed by widely spaced fractures; and “Biancone”, a chalk-type unit, from the lower and middle Cretaceous, 100–200 m in depth, made up of whitish marly limestone that is closely stratified and densely fractured, and very sensitive to frost and atmospheric agents. Therefore, the cave develops at the stratigraphic contact between the “Calcari del Gruppo di San Vigilio” and the “Rosso Ammonitico” and is close to a fault plane, placing the two above formations in vertical contact with the “Biancone”. The overlying rocks of the cave are formed by the massive beds of lower Rosso Ammonitico, whereas the inner cave is mostly developed inside the Calcari del Gruppo di San Vigilio. At the topographical surface, the line of the normal fault runs along a small valley a few meters to the east of the cave; the displacement of the fault is about 100 m.
From a hydrological point of view, the water circulates diffusely inside the dense network of discontinuities of the Biancone unit; the preferential flow is sub-parallel to the topographical surface and occurs mostly below the dry valley bottoms but is also influenced by the structural setting; vertical losses occur along the fault and fracture zones. In contrast, the water circulation is more concentrated and mostly vertical in the Rosso Ammonitico [6].
1.2. Buso della Rana-Pisatela Cave System (40 V–VI/1707 V–VI)
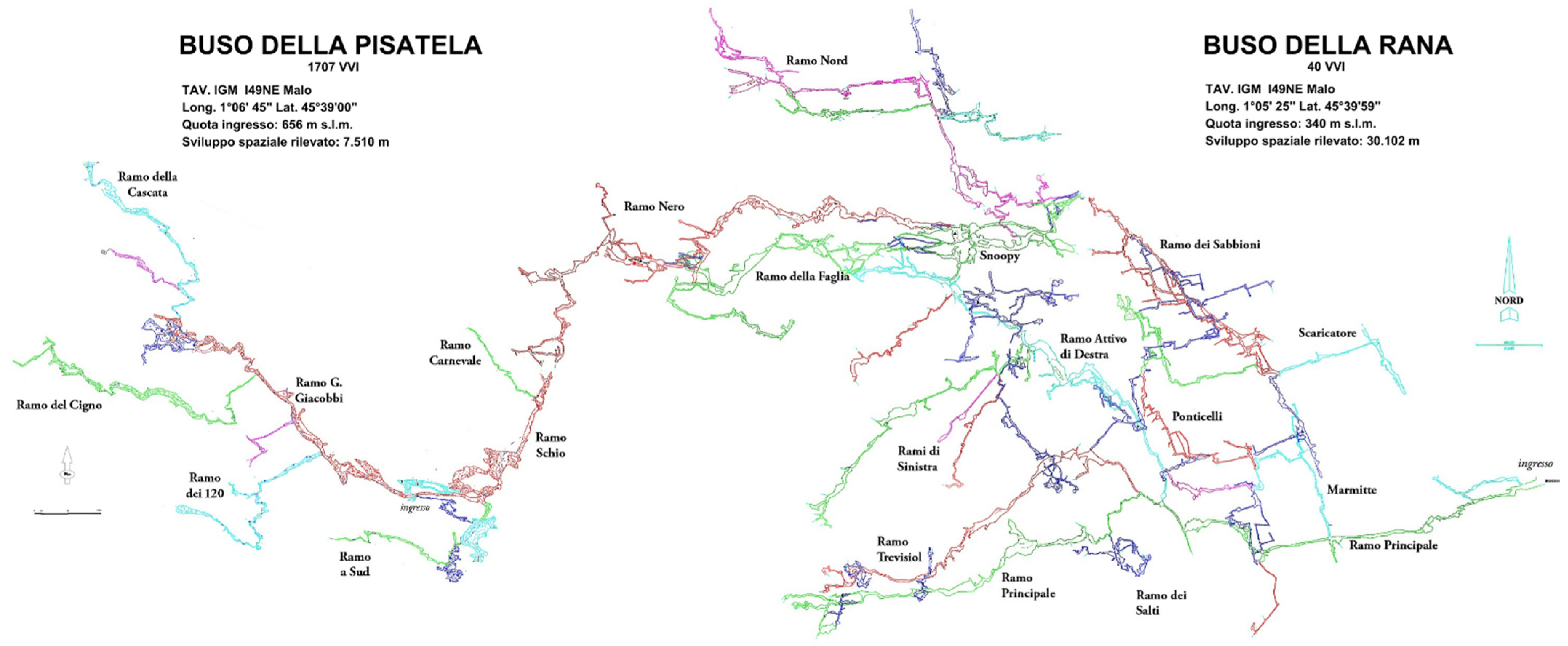
Buso della Rana cave (40 V–VI) opens at an altitude of 340 m a.s.l. in the province of Vicenza, municipality of Monte di Malo. It has a length of 30,102 m and an altitudinal range of 274 m.
In 2012, the cave, which had only one entrance, was connected to the Pisatela Cave (1707 V–VI), a cavity with two entrances (Pater Noster and Pisatela), the highest of which opens at 747 m a.s.l. at a development of 7510 m. The two cavities thus give rise to the Rana-Pisatella cave system, with a development of 37,612 m and an altitude difference of 407 m between the upper (Pisatella) and lower (Rana) entrances (Figure 4).
The cave system is located on the Faedo-Casaron Plateau, which occupies a geographical area of 15 square kilometers in the province of Vicenza.
The area lies between the latitude of 45°36′46″ (Cornedo) to the South and 45°39′35″ (Monte di Malo) to the North and a longitude of 11°19′46″ (Monte Faedo) to the West and 11°22′22″ (Priabona) to the East. Morphologically, it is made up of limestone that gives rise to gentle and rounded surface forms typical of hills, with the valleys oriented according to the main Lessinian tectonic lines.
The Buso della Rana-Pisatela cave system, with its 37,612 m of development, is one of the longest caves in Italy. It developed in the Oligocene limestone through joint networks from the Faedo-Casaron plateau towards the less permeable basalt surface. Here, several independent and differently sized brooks descend along the gradient of the transgressive contact towards the entrance of the Buso della Rana cave, which is the main spring of the karst system forming the Rana River. Pisatela Cave is an inactive sink located inside a doline that reaches the main stream through a series of narrow meanders and shafts [9,10].
The drainage in the cave system is primarily controlled by contact with the less permeable basalt surface, basal conglomerate, and terrigenous marls of the Priabona Formation that rise above the contact in the eastern sector of the system. In some parts of the cave, the conduits evolved entirely in the Boro conglomerate, which is almost 2 m thick in the upstream part of Pisatela Cave. Here, the conglomerate covers the lower basalt contact, while in Buso della Rana, it is often absent, and the calcarenites lie directly on the basalts [9].
Cave morphologies are mostly vadose, with deep canyons and meanders and large collapse rooms at the intersection of different streams and fracture sets. Small paleo-phreatic conduits, evolved entirely in the calcarenite, hanging about 10–15 m above the basalt surface, are related to the phreatic primary drainage and locally ancient epiphreatic conditions. Various vadose narrow shafts reach the contact galleries from the above overlying plateau, showing condensation corrosion morphologies [9].
During periods of heavy rain, the level of the streams rises, often flooding some areas of the cave. In the terminal parts (the last few thousand meters towards the entrance), the river shows a well-structured hyporheic zone (Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9).
Since the Rana-Pisatela system has only recently been joined, much of the research (especially on aquatics) was carried out in the Buso della Rana cave, which will often be cited separately in the text.
2. Materials and Methods
2.1. Sampling and Museum Collections
Both caves have been known since the first half of the last century, and numerous biospeleological investigations have been carried out within them. The results of these surveys have largely been published, but many unstudied specimens are present in the collections of the Museo di Storia Naturale of Verona (Italy).
In recent years, research campaigns aimed to increase the knowledge of the troglobiotic and stygobiotic fauna as a whole were carried out [11,12,13].
Sampling of terrestrial fauna was carried out by means of pitfall traps and direct capture. The pitfall traps consisted of a plastic or glass cup, usually with an opening diameter of 10 cm and a depth of 15 cm. Each was filled with a preserving liquid (NaCl solution) in which was placed a tube containing an attracting bait of blue cheese. The traps were used on a few occasions to integrate hand-collecting and were left in place for about a month each time. In some cases, when we were unsure whether to return to the cave within a short time, the bait was placed without the trap, and fauna were collected at sight.
Aquatic fauna were sampled in both caves by direct capture, hand nets, or using a syringe to collect the water filling the pools. In the Buso della Rana, a drip funnel was placed in the main branch 600 m from the entrance [13]. A drip funnel consists of a funnel supported by a bucket that allows it to direct the dripping water into a plastic container. A 2 cm × 3 cm area on two sides of the square container was cut out and covered with a net (mesh size 60 µm) to retain the animals in the container [14,15].
The specimens sampled in the caves in the province of Verona, both in historical and recent times and not taxonomically identified, have been deposited in the ‘miscellanea Biospeleologica’ collection of the Museum of Verona. In the course of the study for this paper, the material collected in the two caves and not yet identified (i.e., Isopoda, Opilionida, and Collembola) was sent to specialists.
2.2. Bibliographic Research
Many papers on speleological (especially the Buso della Rana cave), geological, and biological aspects have been published from the first half of the last century. As far as biological research is concerned, in addition to the many publications devoted to the description of new species or studies and reviews of certain faunitic groups, there have been few works on the fauna as a whole of the two caves [16,17,18,19].
Therefore, we collected all publications to date and checked and updated the scientific names. This was also important so as to identify and list all the species for which the two caves represent the type locality. More than 60 publications were found in which, in various ways, the presence of animal species in one (or both) of the two (or both) caves under study is mentioned. For reasons of space and usefulness, all of them are obviously not listed in the bibliography.
3. Results
The subterranean fauna of the two caves, as a whole, consists of 46 cave-dwelling species. A total of 35 species are troglobionts or stygobionts, while 12 can be considered eutroglophiles (sensu Ruffo, 1957) [20,21]. In the Arena Cave, 24 obligate subterranean species are known, of which 16 are troglobionts and 8 are stygobionts; for the Rana-Pisatella cave system, 18 species are known, of which 7 are troglobionts and 11 are stygobionts (Table 1).
3.1. Terrestrial Fauna
Mollusca Gastropoda is represented by Zospeum globosum Kušcer, 1928. It is a small mollusc (rarely exceeding 2 mm in height) with a translucent or transparent shell, a body diaphanous, and totally lacking in ocular spots. It is mainly encountered on walls that are damp or wet from dripping water and often covered by silty materials [22].
Among the Opiliones, Ischyropsalis strandi Kratochvil, 1936 is present (Figure 10). This species is endemic to the caves in the Verona Prealps (Monte Baldo and Lessini Mountains). They are present in the two caves studied, and can be found in other caves in the Lessinia Mountains, usually above 600 meters of altitude, being a rather cryophilous species [23]. Within the Rana-Pisatella system, they are more frequently found in the higher elevations of the Grotta della Pisatela.
In regard to Pseudoscorpionida, three troglobiotic species are present in the caves: Chthonius (Chthonius) lessiniensis Schawaller, 1982, with Balkan affinities being a subterranean species. They show a high degree of troglomorphy, ranging from the western Venetian Prealps to the eastern Venetian Prealps [24], and are quite easy to detect under the collapsed stones at the bottom of the hall in the Arena Cave. Neobisium (Blothrus) torrei (E. Simon, 1881) is present in many caves in the Prealps and Alps of Veneto and Friuli Venezia Giulia regions [3]. This species is found in the Arena Cave and is the only troglobiotic pseudoscorpion currently known in the Rana-Pisatella system. Balkanoronchus boldorii (Beier, 1931) is present in some caves of the Prealps of Brescia, in the Monte Baldo and the Lessini Mountains. These specimens were collected both with traps and hand-collecting in the Arena Cave. This species frequents the same habitats as C. lessiniensis.
The Terrestrial Isopoda species is represented by Androniscus (Dentigeroniscus) degener Brian, 1926, and is troglobiont and endemic to the Lessini Mountains in the Verona and Vicenza Provinces. It is quite common in both caves under stones in wetter areas.
To date, troglobiotic millipedes have only been found within Arena Cave; these are the Julidae Troglojulus boldorii Manfredi, 1940n, a species endemic to the caves of the Prealps of Lombardia, Veneto, and Trentino, and the Craspedosomatidae Lessinosoma paoletti Strasser, 1977, which is endemic to the Arena Cave [17].
The Troglobiotic Collembola are also known to date only from Arena Cave. The Onychiuridae species are represented by Onychiurus hauseri Dallai, 1975 and are endemic to the caves in the Veneto and Trentino regions, and the Entomobryidae Pseudosinella concii Gisin, 1950 is a species distributed in different caves in Italy and Switzerland and is quite common in the Arena Cave [3]. Some specimens belonging to the genus Pseudosinella Schaeffer, 1897 are not identifiable on a species level since they are very damaged; these were also sampled in the Arena Cave. However, it is possible to assert that they do not belong to Pseudosinella concii on the basis of the different number of labium setae.
Coleoptera are the most interesting among the terrestrial animals found in the caves considered in this study. Carabidae Trechinae, in particular, shows particularly robust adaptations for life in subterranean environments, and among them is Italaphaenops dimaioi Ghidini, 1964. Endemic to the Lessini Mountains in the Verona area, this species is one of the largest subterranean Trechinae in the world. The colonization of caves by this troglobiont can be traced back to an epoch preceding the Quaternary glaciations [25]. I. dimaioi is known from some caves of the Veronese Lessini, which develop at the contact between different types of karst formations like the Arena Cave. It is not present in the Vicenza Province, so it is not present in the Rana-Pisatella cave system.
Another extremely specialized genus of ancient pre-Quaternary origin of Trechinae is Lessinodytes Vigna Taglianti, 1982. This genus is distributed in the Lombardy and Veneto Prealps with three species, which are all endemic to one or a few caves and is present in the Arena Cave with the rather rare species Lessinodytes pivai Vigna Taglianti & Sciaky, 1988, which is endemic to that cave [26,27].
Orotrechus vicentinus juccii Pomini, 1940, is endemic to the Lessini Mountains in Verona Province and belongs to a group of species distributed in the Venetian Prealps and which, in Arena Cave, co-occurs with Orotrechus pominii Tamanini, 1953. O. pomini is the only known troglobiont Trechinae from the Rana-Pisatella system [28].
With regard to the Leiodidae Cholevinae, Halberria zorzii (Ruffo, 1950) is present in Arena Cave, while in the Buso della Rana, we find Lessiniella trevisioli Pavan, 1941 and Neobathyscia fabianii (Dodero, 1904) (Figure 11). The genus Halberria Conci & Tamanini, 1951 is present, with nine species in the caves in Eastern Veneto and Southern Trentino. In the Western Lessini Mountains, H. zorzi is present only in the caves that open at higher altitudes (above 1400 m a.s.l.), while in the caves at lower altitudes, the species of the genus Neobathyscia show vicariant distributions [17]. It is quite common in the Arena Cave [18].
The genus Lessiniella Pavan, 1941 is phylogenetically close to Halberria [29,30] and consists of two species: Lessiniella trevisoli, of which Buso della Rana is the typical locality where it is found rare in the innermost areas, and Lessiniella berica Piva, 1993, from the nearby Berici Mountains, which are also in the Vicenza Province [29].
To the genus Neobathyscia belongs nine species endemic to the Venetian Prealps, distributed between the Adige and Piave Rivers [30]. N. fabiani is known from several caves in the province of Vicenza that open in localities not far from the Rana-Pisatella cave system [29]. Within the system, it is rather common, especially in the branches of the Buso della Rana cave, while it seems rarer in the Pisatela cave.
3.2. Aquatic Fauna
In Arena Cave, six copepod species were found in a small pool fed by percolating water and a small drain, all stygobiotic. In Buso della Rana, 19 species of copepods are known, and 9 of the copepods are stygobiotic.
The Cyclopidae are represented by Speocyclops infernus (Kiefer, 1930), a stygobiotic species that is widespread over a broader geographical area in the epikarst and vadose zones in the eastern Alpine region and is present in both the caves under study [14,31,32]. First collected in Buso della Rana by Chappuis in the first half of the last century [33,34,35], S. infernus has since been found in many parts of the cave, in both small lakes and pools. It is also found in puddles inside Arena Cave, although the species attribution is not yet certain and is therefore currently reported as S. cf. infernus [3].
Among the Harpacticoida, those of the family Canthocamptidae are the most abundant stygobiotic copepods. The genus Elaphoidella is present in the Arena Cave and Rana-Pisatela system. Elaphoidella is widespread in almost all groundwater habitats in Italy, in both karstic and porous aquifers, as well as the hyporheic zones of rivers, springs, the epikarst, and the saturated karst. Elaphoidella phreatica (Chappuis, 1925) is widely distributed in Italy and across Europe [36]. Elaphoidella ruffoi Chappuis, 1953 is endemic to the Buso della Rana and is rather rare. It was first found during research in 1952 and was not found in the epikarst. Only one species of this genus was found in Arena Cave: Elaphoidella sp. A1 [37]. An Elaphoidella, different from the others, was also found in the Buso della Rana, but the scarce material made it impossible to identify at the species level (Bruno et al. 2018). Ceuthonectes serbicus Chappuis, 1924 was detected in recent research on epikarst fauna in the Buso della Rana cave [13].
The genus Elaphoidella is closely related to the genus Lessinocamptus. Known until a few years ago only from the vadose zone of the Lessinian caves, Lessinocamptus is now also known from a site in Northern Slovenia. In fact, Lessinocamptus pivai Stoch 1997, which was considered endemic to the Buso della Rana, was also found in the Lipnik spring complex in the Julian Alps (NW Slovenia) [38]. Lessinocamptus caoduroi Stoch, 1997, present in the pool in the Arena Cave, was found only in the percolating waters of the vadose zone of caves with an elevation of more than 1000 m a.s.l. in the Lessini Mountains. Lessinocamptus insoletus (Chappuis, 1928) was collected by Chappuis for the first time in the hypogean brook inside Buso della Rana [34]; however, further intensive sampling in the brook did not yield any specimen of L. insoletus and is therefore probable that the vadose zone is the main habitat of the species, from which it can be transported into the brook by percolating water [39]. The Ameiridae are represented by Nitocrella psammophila Chappuis, 1955 and is a stygobiotic species endemic to Italy. It is a widely distributed harpacticoid in the interstitial zone of subterranean streams in caves and the hyporheic in the Po Valley and has been reported in Apennine wells in Central Italy and caves in southern Italy. It is commonly found in the two caves under study. The Parastenocarididae are represented by Parastenocaris ranae Stoch, 2000, which was collected in the Buso della Rana in the large residual pools of the subterranean brook in a dry period [40].
Aquatic isopods have only been found in the Buso della Rana, where Monolistra (Typhlosphaeroma) bericum bericum (Fabiani, 1901) is present. It is a stygobiotic isopod endemic to the Lessini Mountains and Berici Hills (Vicenza Province).
Bathynellacea were found in the Arena Cave. Not yet identified at a species level, Bathynella sp. from Arena Cave was collected in a pond fed by a small water flow.
4. Discussion
The obligate subterranean fauna of Arena Cave and the Rana-Pisatella cave system is exceptionally rich. It comprises 35 troglobionts and stygobionts, representing 74% of the obligate subterranean fauna of the whole caves in the Lessini Mountains (more than 200 caves surveyed).
Despite its small size, Arena Cave is the richest one, with 15 troglobionts and eight stygobionts. The Rana-Pisatella system has a higher number of stygobionts (seven troglobionts and 11 stygobionts).
The geographical proximity between the two caves (22 km) would lead one to suspect a high taxonomic similarity in the fauna inhabiting them. These, on the contrary, have a very different obligate subterranean fauna in terms of terrestrial but especially aquatic species. Only seven species (five troglobionts and two stygobionts) out of thirty-five are in common for the two caves—applying the Jaccard similarity index (and expressed as a percentage similarity), the results in the similarity between the two caves is 21%.
The high richness and differences in faunal composition of the two caves can be explained by the paleogeographical events that occurred in the study area.
During the last glacial period, the Italian Alps were covered by glaciers, except at the top of the highest mountains [41]. In contrast to the Alps, Prealpine areas were only partially covered by glaciers [2,42], and glacial tongues occupied only a few deeper valleys [43]. This favored the colonization of ice-free zones by invertebrates from moist and cold habitats, like forest litter and soil, alpine grasslands, and talus areas. During interglacial periods, as glaciers retreated, populations became isolated in the highest parts of the Prealpine mountains or took refuge in cold, moist interiors of caves [5,6]. Surface populations became extinct or isolated, and there was, therefore, little or no gene flow between cave communities, boosting the evolution of the troglobiont [11]. This is known as the climatic relict hypothesis [44,45,46,47]. The effects of Quaternary glaciations also shaped the stygobiotic species distribution, as the Massif was only marginally covered by ice, and the extensive networks of fractures of the karstic system represented a refuge for stygobionts, boosting isolation and speciation [13]. In fact, the vadose zone and the epikarst of the Lessinian Massif are known to harbor a high diversity of microcrustaceans, including many endemic species due to the ancient geological age of the aquifers, high habitat fragmentation, and isolation of microhabitats, factors of which concurred to promote speciation by vicariance [13,48].
The exceptional subterranean diversity of Arena Cave and the Rana-Pisatella cave system, so different from each other in shape and development (less than 100 m for Arena Cave and about 38 km for the Rana-Pisatella system), can be explained only by their geology, where the two caves developed in different typologies of rocks, namely “contact caves” [5,6].
Contact karst is considered, in a strict sense, a karst phenomenon, where forms are influenced by the contact between a karstifiable rock and a non-karstifiable rock. In a wide sense, the karst phenomena and forms that are influenced by the contact between two karstifiable rocks differ in some of their characteristics, such as chemical composition, porosity, and fracture density [5].
These different rock characteristics create a number of different microhabitats that also influence the life and dispersion of the subterranean animals. Depending on the amount of water retained, humidity, and other factors that are not yet fully known, specimens of different species may prefer one microhabitat over another. This explains the rarity of the findings in Arena Cave of the trechine Lessynodites pivai, a species that most probably do not frequently inhabit the cave proper but rather lives in the wetter interstices of the Rosso Ammonitico formations. The same can be said for Italaphaenops dimaioi; ongoing studies by the lab of the Museum of Verona show that I. dimaioi was sampled almost exclusively in the caves in contact with Rosso Ammonitico rock in the Lessini Mountains.
The same microhabitat characteristics are probably the reason for the abundance of copepods and the presence of Bathynella sp., present in waters that flow from the environments in the small pool inside the cave. It is in similar conditions that, collecting water from the epikarst, we found Ceuthonectes serbicus for the first time in Buso della Rana [13].
The smaller cave is, therefore, the richest in diversity. As is often said, size does not always matter.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
I would like to thank Dave Culver (Washington, DC, USA), Louis Deharveng (Paris, France), Valeria Lencioni (Trento, Italy), and Francesco Sauro (Verona, Italy) for their helpful suggestions in the elaboration of the text; Ivan Petri (Trento, Italy), Barbara Valle (Milan, Italy), Fabio Stoch (Brussels, Belgium) for their taxonomic identification or suggestions; Arianna Spada (Verona, Italy) for the graphic elaboration of some figures; Sandro Sedran (Dolo, Italy) and Filippo Rossetto (Verona, Italy) for allowing the use of photographs of the Buso della Rana and Ischyropsalis strandii. I am also grateful to all the speleologists who shared the cave surveys with me, especially Giorgio Annichini (Verona, Italy), Tarcisio Battagini (Verona, Italy), and Andrea Pasotto (Verona, Italy)—and a special thanks to Dave Culver for revising the English text and a thanks to the four anonymous reviewers.
Conflicts of Interest
The author declares no conflict of interest.
References
- Marazzi, S. Atlante Orografico Delle Alpi—SOIUSA; Priuli and Verlucca: Scarmagno, Italy, 2005. [Google Scholar]
- Sauro, U. Il Paesaggio Degli Alti Lessini: Studio Geomorfologico; Museo Civico di Storia Naturale di Verona: Verona, Italy, 1973; Volume 6, 161p. [Google Scholar]
- Caoduro, G.; Ruffo, S. La Grotta dell’Arena, un biotopo di eccezionale interesse negli alti Lessini. In La Lessinia Ieri Oggi Domani: Quaderno Culturale; Editrice La Grafica: Lavagno, Italy, 1998; pp. 39–44. [Google Scholar]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Sauro, U. Aspects of contact karst in the Venetian Fore-Alps. Acta Carsologica 2001, 30, 89–102. [Google Scholar]
- Latella, L.; Sauro, U. Aspects of the Evolution of an Important Geo-Ecosystem in the Lessinian Mountain (Venetian Prealps, Italy). Acta Carsologica 2007, 36, 69–75. [Google Scholar] [CrossRef]
- Pasa, A. Carsismo ed idrografia carsica del Gruppo del Monte Baldo e dei Lessini Veronesi. CNR Cent. Studi Geogr. Fis. Ric. Sulla Morfol. Idrogr. Carsismo 1954, 5, 1–150. [Google Scholar]
- Sauro, U. Aspetti dell’evoluzione carsica legata a particolari condizioni litologiche e tettoniche negli Alti Lessini. Boll. Soc. Geol. Ital. 1974, 93, 945–969. [Google Scholar]
- Tisato, N.; Sauro, F.; Bernasconi, S.M.; Bruijn, R.H.; De Waele, J. Hypogenic contribution to speleogenesis in a predominant epigenic karst system: A case study from the Venetian Alps, Italy. Geomorphology 2012, 151, 156–163. [Google Scholar] [CrossRef]
- Sistema carsico Rana-Pisatella. Available online: www.busodellarana.it (accessed on 1 August 2023).
- Latella, L.; Verdari, N.; Gobbi, M. Distribution of terrestrial cave-dwelling arthropods in two adjacent Prealpine Italian areas with different glacial histories. Zool. Stud. 2012, 51, 1113–1121. [Google Scholar]
- Avesani, D.; Latella, L. Spatio-temporal distribution of the genus Chionea (Diptera, Limoniidae) in the Buso del Valon ice cave and other caves in the Lessini Mountains (Northern Italy). Boll. Mus. Civ. St. Nat. Verona 2016, 40, 11–16. [Google Scholar]
- Bruno, M.C.; Cottarelli, V.; Grasso, R.; Latella, L.; Zaupa, S.; Spena, M.T. Epikarst crustaceans from some Italian caves: Endemisms and spatial scales. Biogeographia 2018, 33, 1–18. [Google Scholar]
- Pipan, T. Epikarst—A Promising Habitat; ZRC Publishing: Ljubljana, Slovenia, 2005; pp. 1–101. [Google Scholar]
- Pipan, T.; Christman, M.C.; Culver, D.C. Dynamics of epikarst communities: Microgeographic pattern and environmental determinants of epikarst copepods in Organ Cave, West Virginia. Am. Midl. Nat. 2006, 156, 75–87. [Google Scholar] [CrossRef]
- Allegranzi, A.; Bartolomei, G.; Broglio, A.; Pasa, A.; Rigobello, A.; Ruffo, S. Il Buso della Rana (40 V-VI). Ras. Speleol. Ital. 1960, 12, 99–163. [Google Scholar]
- Caoduro, G.; Osella, G.; Ruffo, S. La fauna cavernicola della regione veronese. Memorie del Museo Civico di Storia Naturale di Verona. Sez. Biol. 1994, 6, 1–144. [Google Scholar]
- Latella, L.; Sauro, U. Note di Storia Naturale del sottosuolo dei Monti Lessini e del suo popolamento. Quaderno Culturale Lessinia Ieri Oggi Domani 2006, 34, 57–64. [Google Scholar]
- Latella, L. Il contributo del Museo Civico di Storia Naturale di Verona allo sviluppo della biospeleologia. Studi Trentini Di Sci. Nat. Acta Biol. 2004, 81, 15–22. [Google Scholar]
- Ruffo, S. Le attuali conoscenze sulla fauna cavernicola della Regione Pugliese. Mem. Biogeogr. Adriat. 1957, 3, 1–143. [Google Scholar]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Pezzoli, E. Il genere Zospeum Bourguignat, 1856 in Italia (Gastropoda Polmonata Basommatophora). Censimento delle stazioni ad oggi segnalate. Nat. Brescia. 1992, 27, 123–169. [Google Scholar]
- Petri, I.; Ballarin, F.; Latella, L. Seasonal abundance and spatio-temporal distribution of the troglophylic harvestman Ischyropsalis ravasinii (Arachnida, Opiliones, Ischyropsalididae) in the Buso del Valon ice cave, Eastern Italian Prealps. Subterr. Biol. 2022, 42, 151–164. [Google Scholar] [CrossRef]
- Gardini, G. The species of the Chthonius heterodactylus group (Arachnida, Pseudoscorpiones, Chthoniidae) from the eastern Alps and the Carpathians. Zootaxa 2014, 3887, 101–137. [Google Scholar] [CrossRef]
- Casale, A.; Vigna Taglianti, A. Note su Italaphaenops dimaioi Ghidini (Coleoptera, Carabidae). Boll. del Mus. Civ. Stor. Nat. di Verona 1976, 2, 293–314. [Google Scholar]
- Sciaky, R.; Vigna Taglianti, A. The genus Lessinodytes (Coleoptera, Carabidae, Trechinae). Mém. Biospéol. 1990, 17, 169–173. [Google Scholar]
- Monguzzi, R. Nuovi Dati Per La Conoscenza Del Genere Lessinodytes Vigna Taglianti, 1982 (Coleoptera Carabidae Trechinae). Nat. Brescia. 1993, 29, 179–183. [Google Scholar]
- Faille, A.; Casale, A.; Balke, M.; Ribera, I. A molecular phylogeny of Alpine subterranean Trechini (Coleoptera: Carabidae). BMC Evol. Biol. 2013, 13, 248. [Google Scholar] [CrossRef] [PubMed]
- Latella, L. The Subterranean Cholevinae of Italy. In Cave Biodiversity: Speciation and Diversity of Subterranean Fauna; Johns Hopkins University Press: Baltimore, MD, USA, 2022; p. 164. [Google Scholar]
- Giachino, P.M.; Vailati, D. I Cholevidae delle Alpi e Prealpi italiane: Inventario, analisi faunistica e origine del popolamento del settore compreso fra i corsi dei fiumi Ticino e Tagliamento (Coleoptera). Biogeogr. J. Integr. Biogeogr. 2005, 26, 229–378. [Google Scholar] [CrossRef]
- Deharveng, L.; Stoch, F.; Gibert, J.; Bedos, A.; Galassi, D.; Zagmajster, M.; Brancelj, A.; Camacho, A.; Fiers, F.; Martin, P.; et al. Groundwater biodiversity in Europe. Freshw. Biol. 2009, 54, 709–726. [Google Scholar] [CrossRef]
- Papi, F.; Pipan, T. Ecological studies of an epikarst community in Snežna jama na planini Arto an ice cave in north central Slovenia. Acta Carsologica 2011, 40, 3. [Google Scholar] [CrossRef]
- Chappuis, P.A. Nouveaux Copépodes cavernicoles des genres Cyclops et Canthocamptus (Note préliminaire). Bul. Soc. De Stiint. Din Cluj 1923, 1, 584–590. [Google Scholar]
- Chappuis, P.A. Nouveaux Copépodes cavernicoles. Bul. Soc. De Stiint. Din Cluj 1928, 2, 20–34. [Google Scholar]
- Chappuis, P.A. Biospeologica LIX. Copepodes (premiere serie), avec l’énumeration de tous les copepodes cavernicoles connus en 1931. Arch. De Zool. Expérimentale Et Générale 1933, 76, 1–56. [Google Scholar]
- Galassi, D.M.P. Groundwater copepods: Diversity patterns over ecological and evolutionary scales. Hydrobiologia 2001, 453, 227–253. [Google Scholar] [CrossRef]
- Ruffo, S.; Stoch, F. Checklist and Distribution of the Italian Fauna. Memorie del Museo Civico di Storia Naturale di Verona, II Serie, Sezione Scienze Della Vita 17; Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2006; with CD-ROM. [Google Scholar]
- Mori, N.; Brancelj, A. Differences in aquatic microcrustacean assemblages between temporary and perennial springs of an alpine karstic aquifer. Int. J. Speleol. 2013, 42, 3–9. [Google Scholar] [CrossRef]
- Stoch, F. A new genus and two new species of Canthocamptidae (Copepoda, Harpacticoida) from caves in northern Italy. Hydrobiologia 1997, 350, 49–61. [Google Scholar] [CrossRef]
- Stoch, F. New and little known Parastenocaris (Copepoda, Harpacticoida, Parastenocarididae) from cave waters in Northeastern Italy. Boll. Del Mus. Civ. Di Stor. Nat. Di Verona 2000, 24, 195–206. [Google Scholar]
- Hughes, P.D. Quaternary glacial history of the Mediterranean mountains. Progr. Phys. Geogr. 2006, 30, 334–364. [Google Scholar] [CrossRef]
- Bassetti, M.; Borsato, A. Evoluzione geomorfologica e vegetazionale della bassa valle dell’Adige dall’ultimo massimo glaciale: Sintesi delle conoscenze e riferimenti ad aree limitrofe. Studi Trentini Sci. Nat. Acta Geol. 2007, 82, 31–42. [Google Scholar]
- Sommaruga, M.; Zorzin, R. Evidences of Morphologies and Glacial Deposits into High Valle del Chiampo (Northern Italy, Vicenza Province): First Results. Boll. Del Mus. Civ. Di Stor. Nat. Di Verona Geol. Paleontol. Preist. 2018, 42, 43–71. [Google Scholar]
- Sbordoni, V. Advances in speciation of cave animals. In Mechanisms of Speciation; Barigozzi, C., Ed.; Liss: New York, NY, USA, 1982; pp. 219–224. [Google Scholar]
- Humphreys, W.F. Relict fauna and their derivation. In Ecosystems of the World; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Subterranean Ecosystems; Elsevier: Amsterdam, The Netherlands, 2000; Volume 30, pp. 417–432. [Google Scholar]
- Assmann, T.; Casale, A.; Drees, C.; Habel, J.C.; Matern, A.; Schuldt, A. Review: The dark side of relict species biology: Cave animals as ancient lineages. In Relict Species: Phylogeography and Conservation Biology; Assmann, T., Habel, J.C., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 91–103. [Google Scholar]
- Hampe, A.; Jump, A.S. Climate relicts: Past, present, future. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 313–333. [Google Scholar] [CrossRef]
- Galassi, D.M.; Stoch, F.; Fiasca, B.; Di Lorenzo, T.; Gattone, E. Groundwater biodiversity patterns in the Lessinian Massif of northern Italy. Freshw. Biol. 2009, 54, 830–847. [Google Scholar] [CrossRef]
Figure 1.
Location of the two studied caves in the Lessinia Mountains (the red rectangle in the small photo indicates the position in Italy).
Figure 1.
Location of the two studied caves in the Lessinia Mountains (the red rectangle in the small photo indicates the position in Italy).

Figure 2.
Cave map of Arena Cave. Sezione = cross section; pianta = plan; ingresso = entrance.

Figure 3.
The large chamber of the Arena Cave (Photo: L. Latella).

Figure 4.
Plan of the Rana-Pisatela cave system (scale bar: 100 m).

Figure 5.
“Ramo attivo di destra”, one of the hydrologically active branches in the Buso della Rana (Photo: S. Sedran).
Figure 5.
“Ramo attivo di destra”, one of the hydrologically active branches in the Buso della Rana (Photo: S. Sedran).

Figure 6.
The beautiful coralloid concretions in the “Ramo franchignia, Saletta broccoli” in the Buso della Rana (Photo: S. Sedran).
Figure 6.
The beautiful coralloid concretions in the “Ramo franchignia, Saletta broccoli” in the Buso della Rana (Photo: S. Sedran).

Figure 7.
The left main branch of the Buso della Rana (Photo: S. Sedran).

Figure 8.
The large dimensions of the “Sala dei Massi”. One of the many large halls that characterize the Rana–Pisatella cave system (Photo: S. Sedran).
Figure 8.
The large dimensions of the “Sala dei Massi”. One of the many large halls that characterize the Rana–Pisatella cave system (Photo: S. Sedran).

Figure 9.
The shaft of the “Ramo del Pantano”, a tributary of the Main Branch of the Buso della Rana (Photo: S. Sedran).
Figure 9.
The shaft of the “Ramo del Pantano”, a tributary of the Main Branch of the Buso della Rana (Photo: S. Sedran).

Figure 10.
A specimen of Ischyropsalis strandi: this species is present in both investigated caves (Photo: F. Rossetto).
Figure 10.
A specimen of Ischyropsalis strandi: this species is present in both investigated caves (Photo: F. Rossetto).

Figure 11.
A specimen of Neobathyscia fabianii from the Rana-Pisatela cave system (Photo: L. Latella).
Figure 11.
A specimen of Neobathyscia fabianii from the Rana-Pisatela cave system (Photo: L. Latella).

Figure 12.
Niphargus similis from Arena Cave (Photo: L. Latella).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The list of troglobiotic and stygobiotic species known in the two studied caves. Tb—troglobiont; Stb—stygobiont; 1—present; 0—absent; Tl—Type locality.
Table 1.
The list of troglobiotic and stygobiotic species known in the two studied caves. Tb—troglobiont; Stb—stygobiont; 1—present; 0—absent; Tl—Type locality.
| Class | Order | Family | Genus/Species/Subspecies | Status | Arena | Rana-Pisatella |
|---|---|---|---|---|---|---|
| Gastropoda | Ellobiida | Ellobiidae | Zospeum globosum Kuščer, 1928 | Tb | 1 | 1 |
| Arachnida | Opiliones | Ischyropsalididae | Ischyropsalis strandti Kratochvíl, 1936 | Tb | 1 | 1 |
| Arachnida | Pseudoscorpionida | Neobisiidae | Neobisium (Blothrus) torrei (Simon, 1881) | Tb | 1 | 1 |
| Arachnida | Pseudoscorpionida | Neobisiidae | Balkanoroncus boldorii (Beier, 1931) | Tb | 1 | 0 |
| Arachnida | Pseudoscorpionida | Chthoniidae | Chthonius lessiniensis Schawaller, 1982 | Tb | 1 | 0 |
| Malacostraca | Isopoda | Trichoniscidae | Androniscus (Dentigeroniscus) degener Brian, 1926 | Tb | 1 | 1 |
| Diplopoda | Chordeumatida | Craspedosomatidae | Lessinosoma paolettii Strasser, 1967 | Tb | 1 Tl | 0 |
| Diplopoda | Chordeumatida | Iulidae | Trogloiulus boldorii Manfredi, 1940 | Tb | 1 | 0 |
| Collembola | Poduromorpha | Onychiuridae | Onychiurus hauseri Dallai, 1975 | Tb | 1 | 0 |
| Collembola | Entomobryomorpha | Entomobryidae | Pseudosinella concii Gisin, 1950 | Tb | 1 | 0 |
| Collembola | Entomobryomorpha | Entomobryidae | Pseudosinella sp. | Tb | 1 | 0 |
| Insecta | Coleoptera | Carabidae | Italaphaenops dimaioi Ghidini, 1964 | Tb | 1 | 0 |
| Insecta | Coleoptera | Carabidae | Lessynodytes pivai Vigna Taglianti & Sciaky, 1988 | Tb | 1 Tl | 0 |
| Insecta | Coleoptera | Carabidae | Orotrechus pominii Tamanini, 1953 | Tb | 1 | 1 |
| Insecta | Coleoptera | Carabidae | Orotrechus vicentinus juccii Pomini, 1940 | Tb | 1 | 0 |
| Insecta | Coleoptera | Leiodidae | Halberria zorzii (Ruffo, 1950) | Tb | 1 Tl | 0 |
| Insecta | Coleoptera | Leiodidae | Lessiniella trevisioli Pavan, 1941 | Tb | 0 | 1 Tl |
| Insecta | Coleoptera | Leiodidae | Neobathyscia fabianii (Dodero, 1904) | Tb | 0 | 1 |
| Copepoda | Cyclopoida | Cyclopidae | Speocyclops infernus Kiefer, 1930 | Stb | 1 | 1 |
| Copepoda | Harpactoida | Camptocamptidae | Elaphoidella phreatica (Chappuis, 1925) | Stb | 0 | 1 |
| Copepoda | Harpactoida | Camptocamptidae | Elaphoidella ruffoi Chappuis, 1953 | Stb | 0 | 1 |
| Copepoda | Harpactoida | Camptocamptidae | Elaphoidella sp. A1 | Stb | 1 | 0 |
| Copepoda | Harpactoida | Camptocamptidae | Elaphoidella sp. A | Stb | 0 | 1 |
| Copepoda | Harpactoida | Camptocamptidae | Ceuthonectes serbicus Chappuis, 1924 | Stb | 0 | 1 |
| Copepoda | Harpactoida | Camptocamptidae | Lessinocamptus insoletus (Chappuis, 1928) | Stb | 0 | 1 Tl |
| Copepoda | Harpactoida | Camptocamptidae | Lessinocamptus pivai Stoch, 1997 | Stb | 0 | 1 Tl |
| Copepoda | Harpactoida | Camptocamptidae | Lessinocamptus caoduroi Stoch, 1997 | Stb | 1 Tl | 0 |
| Copepoda | Harpactoida | Camptocamptidae | Bryocamptus (Limocamptus) echinatus (Mrazek, 1893) | Stb | 1 | 0 |
| Copepoda | Harpactoida | Camptocamptidae | Moraria (M.) sp. A1 | Stb | 1 | 0 |
| Copepoda | Harpactoida | Parastenocaridiidae | Parastenocaris ranae Stoch, 2000 | Stb | 0 | 1 Tl |
| Copepoda | Harpactoida | Amaeridae | Nitocrella psammophila Chappuis, 1955 | Stb | 1 | 1 |
| Malacostraca | Amphipoda | Niphargidae | Niphargus costozzae Schellenberg, 1935 | Stb | 0 | 1 |
| Malacostraca | Amphipoda | Niphargidae | Niphargus similis Karaman & Ruffo, 1989 | Stb | 1 | 0 |
| Malacostraca | Isopoda | Sphaeromatidae | Monolistra (Typhlosphaeroma) bericum bericum Fabiani, 1901 | Stb | 0 | 1 |
| Malacostraca | Bathynellacea | Bathynellidae | Bathynella sp. | Stb | 1 | 0 |
| Tot. | 24 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Latella, L. Does Size Matter? Two Subterranean Biodiversity Hotspots in the Lessini Mountains in the Veneto Prealps in Northern Italy. Diversity 2024, 16, 25. https://doi.org/10.3390/d16010025
AMA Style
Latella L. Does Size Matter? Two Subterranean Biodiversity Hotspots in the Lessini Mountains in the Veneto Prealps in Northern Italy. Diversity. 2024; 16(1):25. https://doi.org/10.3390/d16010025
Chicago/Turabian StyleLatella, Leonardo. 2024. "Does Size Matter? Two Subterranean Biodiversity Hotspots in the Lessini Mountains in the Veneto Prealps in Northern Italy" Diversity 16, no. 1: 25. https://doi.org/10.3390/d16010025
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.