
Ascaridoid Nematodes Infection in Anadromous Fish Coilia nasus from Yangtze River

1
Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China
2
Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, China
*
Authors to whom correspondence should be addressed.
Diversity 2024, 16(3), 167; https://doi.org/10.3390/d16030167
Submission received: 24 January 2024
/
Revised: 25 February 2024
/
Accepted: 2 March 2024
/
Published: 6 March 2024
Abstract
:The longjaw tapertail anchovy Coilia nasus, which migrates from ocean to freshwater for spawning in spring, is an important anadromous fish with ecological and cultural significance. To determine parasite infection in anadromous C. nasus, a total of 103 fish from the Yangtze River were collected and examined in 2021 and 2022. The overall infection prevalence of nematodes in C. nasus was 100%, with a mean intensity of 13.81 ± 16.45. The mean intensity of nematode infections in 2022 was significantly higher than that observed in 2021 across all sampling sites (p < 0.05). Nematodes were widely detected in the mesentery, pyloric cecum, stomach, and liver, among which the mesentery accounted for the highest proportion, reaching up to 53.52%. A total of eight ascaridoid nematodes belonging to the family Anisakidae and Raphidascarididae were identified by using morphological characters and molecular biological techniques, including two species of Anisakis, five species of Hysterothylacium, and one species of Raphidascaris. A. pegreffii was found as the predominant species, accounting for 48.65% of all identified parasitic nematodes in liver, while Raphidascaris sp. was the most common nematode in the mesentery, pyloric cecum, and stomach, reaching up to 39.81%, 36.21%, and 74.36%, respectively. The present study systematically investigated the parasitic status and community structure of the nematode in C. nasus during its migration in the Yangtze River. This research provides a foundation for studying the impact of nematode parasitism on the reproductive migration and population recruitment of C. nasus, and offers valuable insights for biomarker screening and nematode identification in C. nasus.
1. Introduction
Fish parasitic ascaridoid nematodes, such as anisakid and raphidascaridid, are commonly encountered parasites in wild and cultured fish stocks [1,2,3,4]. The life cycle of ascaridoids involves marine mammals, birds, reptiles, amphibians, predatory fish, and elasmobranchs that serve as definitive hosts, and crustaceans, fish, squid, birds, and sea-snakes as intermediate hosts [5,6,7,8]. The growing impact of parasites on fish health and the economic significance of parasites in aquaculture and fisheries have enhanced the need for studies on fish/parasite relationships [9,10]. Meanwhile, the nematodes in the super-family Ascaridoidea have gained global recognition because of their significant impact on human health as the causative agents of anisakidosis, a fish-borne parasitic zoonosis via accidental ingestion of raw or under-cooked fish infected by the family Anisakidae [1,11,12,13].
The longjaw tapertail anchovy Coilia nasus Temminck & Schlegel, 1846 is a small to medium-sized anadromous fish belonging to the order Clupeiformes, and is widely distributed from the coastal waters of China, Korea, and Japan to the interconnected freshwater tributaries [14,15]. The lower and middle reaches of the Yangtze River is the most important channel for the anadromous migration of C. nasus. The migration is a preparatory process for spawning, and wellknown to be a season-dependent and temporally based reproductive activity [16]. Adult C. nasus has been reported to migrate upstream from early February to the end of April, and spawn in the middle and lower reaches of the Yangtze River and connected lakes [14,17,18]. Unfortunately, in recent decades, stocks of C. nasus in the Yangtze River have experienced a significant decline due to anthropogenic influences such as excessive fishing activities, extensive hydraulic construction obstructing migration routes, and degradation of their natural habitats [19,20,21]. Recently, parasite infection in Coilia spp. has attracted much attention [22,23,24,25,26]. Xu et al. [22] found that Anisakis simplex and Epiclavella chinensis only parasitized the anadromous C. nasus in the Yangtze River. Therefore, A. simplex was considered as a “biological indicator” of the anadromous C. nasus in the Yangtze River. Li et al. [23] used the parasites as indicators of fish migratory movements. They pointed out that the monogenean Heteromazocraes lingmueni and the acanthocephalan Acanthosentis cheni are suitable biological tags for the C. nasus population migrating from the middle and lower reaches to the estuary of the Yangtze River, while the nematodes Contracaecum sp. and A. simplex are suitable biological tags for population migrating from the coast and estuary to the Yangtze River. The parasitic infection alters host behavior, damages tissue, affects ingestion, decreases weight, hampers reproduction, and can even cause death in some cases [10,27]. Thus, the parasites seriously threatened the health of C. nasus. In this report, C. nasus caught from the Shanghai and Anqing section of the Yangtze River from 2021 to 2022 were investigated to analyze the parasitic status and community structure of nematodes in C. nasus.
2. Materials and Methods
2.1. Sample Collection
All procedures involved in the handling and treatment conducted as follows were given official approval by the animal ethics committee of the Chinese Academy of Fishery Sciences, and fish collection was approved by the Department of Agriculture and Rural Affairs of Shanghai and Anhui Province with scientific fish collection license. The protocol was approved by the Committee on the Ethics of Animal Experiments of the Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences (Authorization Number: 20221223).
Fish were caught at the Yangtze River estuary (Shanghai section) and the lower reach of Yangtze River (over 580 km to estuary, Anqing section) with gill net between May and July in 2021 and 2022, and immediately transported to laboratory on ice. A total of 103 C. nasus were examined for parasites, and fish body length and sex were recorded. Fish were dissected and the internal organs (abdominal cavity, mesentery, pyloric cecum, stomach, and liver) (Figure 1) were checked under stereomicroscope (Nikon SMZ18). Nematodes isolated from each organ were washed in saline and then fixed with 100% ethanol. In addition, specimens of C. nasus and nematode were all deposited at Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences. Specimen were observed under light microscopy and identified according to the morphological characters of head, tail tip, and excretory and digestive systems.
2.2. Molecular Identification
A total of 250 nematodes isolated from C. nasus at the Shanghai (16 individuals) and Anqing (10 individuals) sections of the Yangtze River, which were sampled in 2022, were all used for molecular identification. Genomic DNA from individuals was extracted using a Column Genomic DNA Isolation Kit (Sangon, Shanghai, China) according to the manufacturer’s instructions. The ribosomal ITS1-5.8S-ITS2 region was amplified using the primers NC2-ITS-F (TTAGTTTCTTTTCCTCCGCT) and NC5-ITS-R (GTAGGTGAACCTGCGGAAGGATCATT) [28] and the predicted PCR production was about 1000 bp. PCR reaction (50 μL) includes 25 μL 2 × Taq Master Mix (Dye Plus) (Vazyme, Nanjing, China), 2 μL of each primer, 4 μL DNA template, and 17 μL ddH2O, and the thermal profile was set as 95 °C, 5 min (initial denaturation) followed by 35 cycles of 95 °C, 30 s (denaturation), 56 °C, 30 s (annealing), 72 °C, 60 s (extension), and a final extension of 72 °C for 10 min. PCR products were purified and sequenced at Sangon Biotech (Shanghai, China). The obtained DNA sequencing chromatograms were checked in Chromas 2.6.6 and assembled with SeqMan (LaserGene package 11.0), aligned in Geneious (5.6) [29]. Sequence similarity was searched against the GenBank database using the Basic Local Alignment Search Tool (BLAST).
Phylogenetic trees were conducted with Bayesian inference (BI) based on the obtained sequences. The best nucleotide substitution model (K2P) was estimated with Modelfinder according to Bayesian information criterion (BIC). BI analyses were conducted using the software MrBayes ver. 3.1.2 with parameter settings nruns = 4, rates = gamma, and ngen = 100,000.
2.3. Data Processing
2.3.1. Terms and Indicators
The terms and indicators were employed here according to Bush et al. [30] and Wu et al. [31], such as Margalef richness index = (N − 1)/ln(N), where N is the number of species;
Aggregate Index: GI = (DI − 1)/(ni − 1), where ni is the total number of worm species i;
Infection Index: Z= ni ×(Pi/N2), where Pi is the number of hosts infected by worm i and N is the total number of worms;
Distribution type and test: DI = variance(S2)/mean value(X). If DI > 1, it is aggregated distribution. If DI = 1, it is random distribution. If DI < 1, it is uniform distribution. Use d = to test the consistency of distribution types. If d < −1.96, accept the uniform distribution. If d > 1.96, accept the aggregated distribution. If d < 1.96, accept the random distribution.
2.3.2. Statistical Analysis
Statistical analyses were performed in SPSS 26 and Excel (Ver. 2401), and the graph was generated using Origin2021. All infection intensity data were expressed as mean ± sd and analyzed using nonparametric test (Mann–Whitney test). Statistical significance was considered at p < 0.05.
3. Results
3.1. Nematode Infection in C. nasus
The body length of longjaw tapertail anchovy C. nasus examined in this study ranged from 23 cm to 35 cm, with an average of 27.71 ± 2.36 cm. Nematode infection in C. nasus samples collected at Shanghai and Anqing in 2021 and 2022 were all 100% (Table 1).
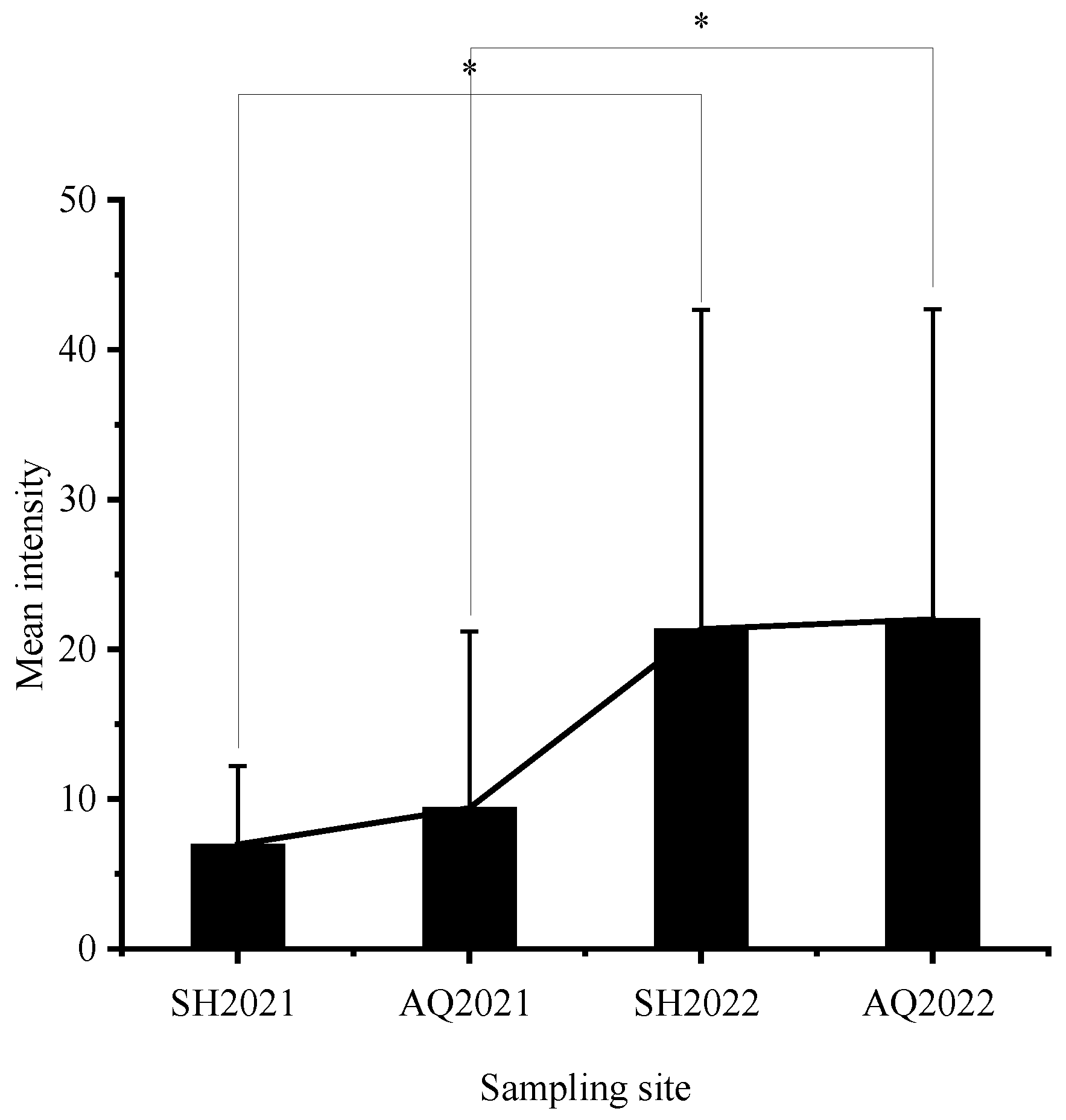
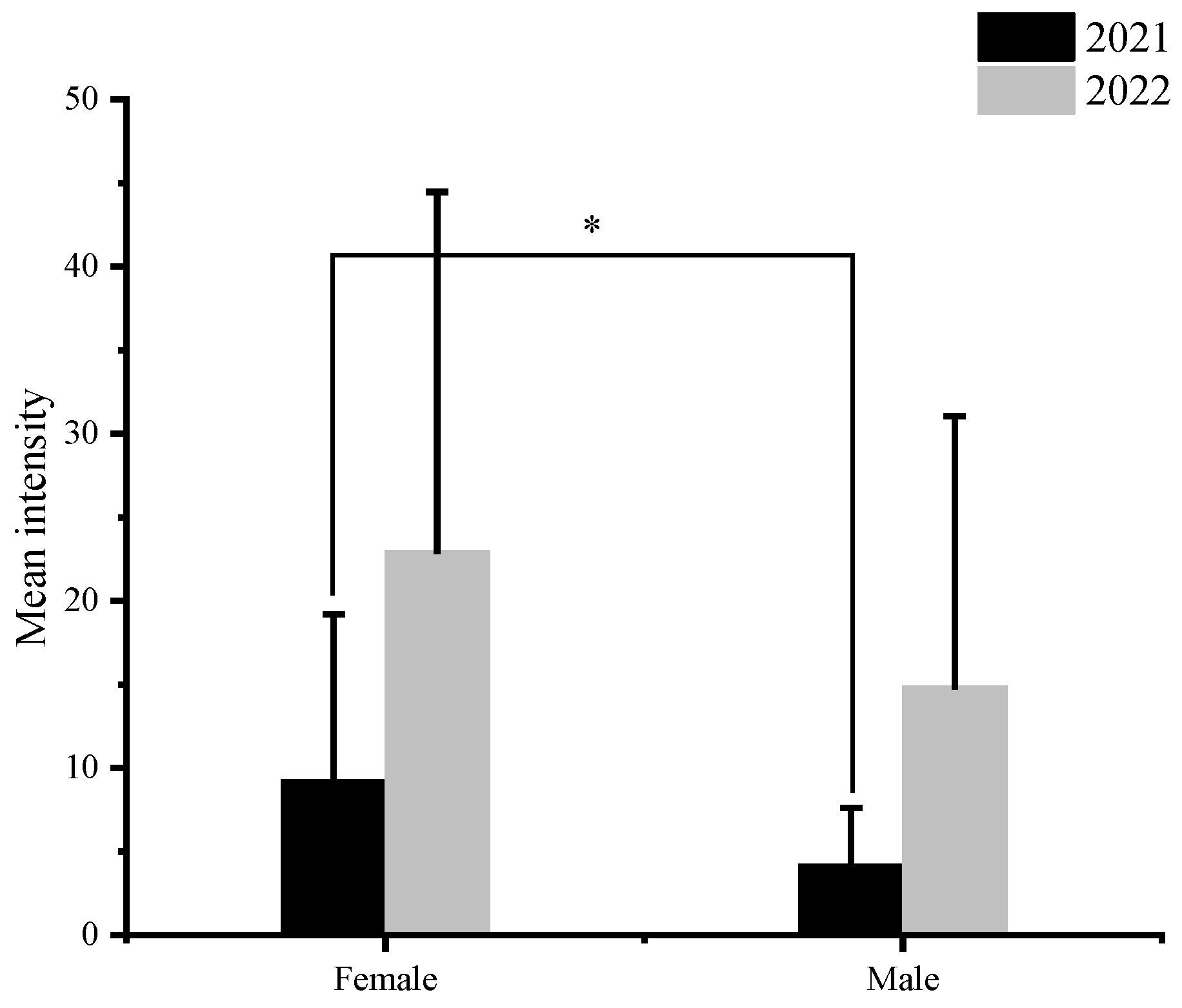
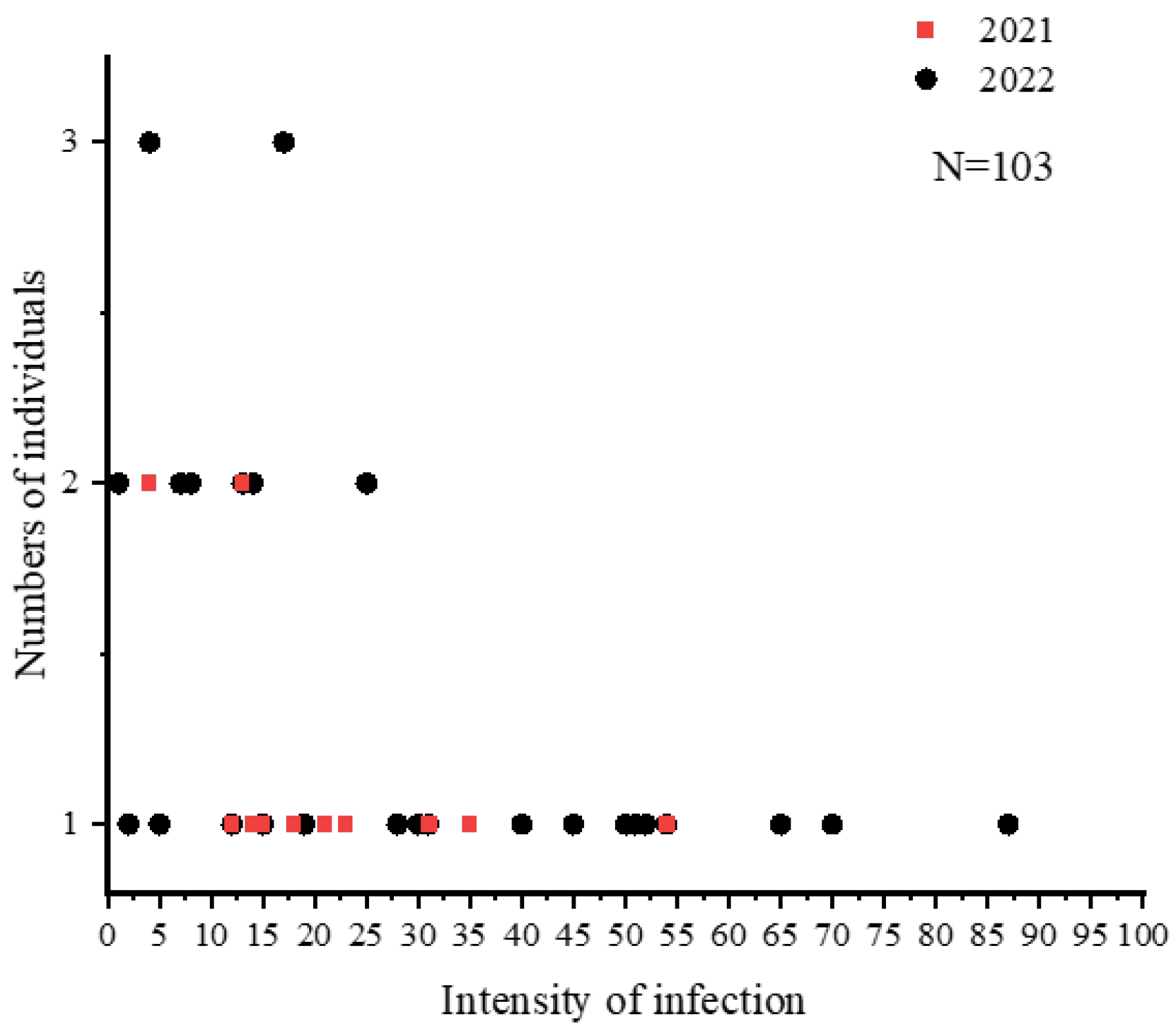
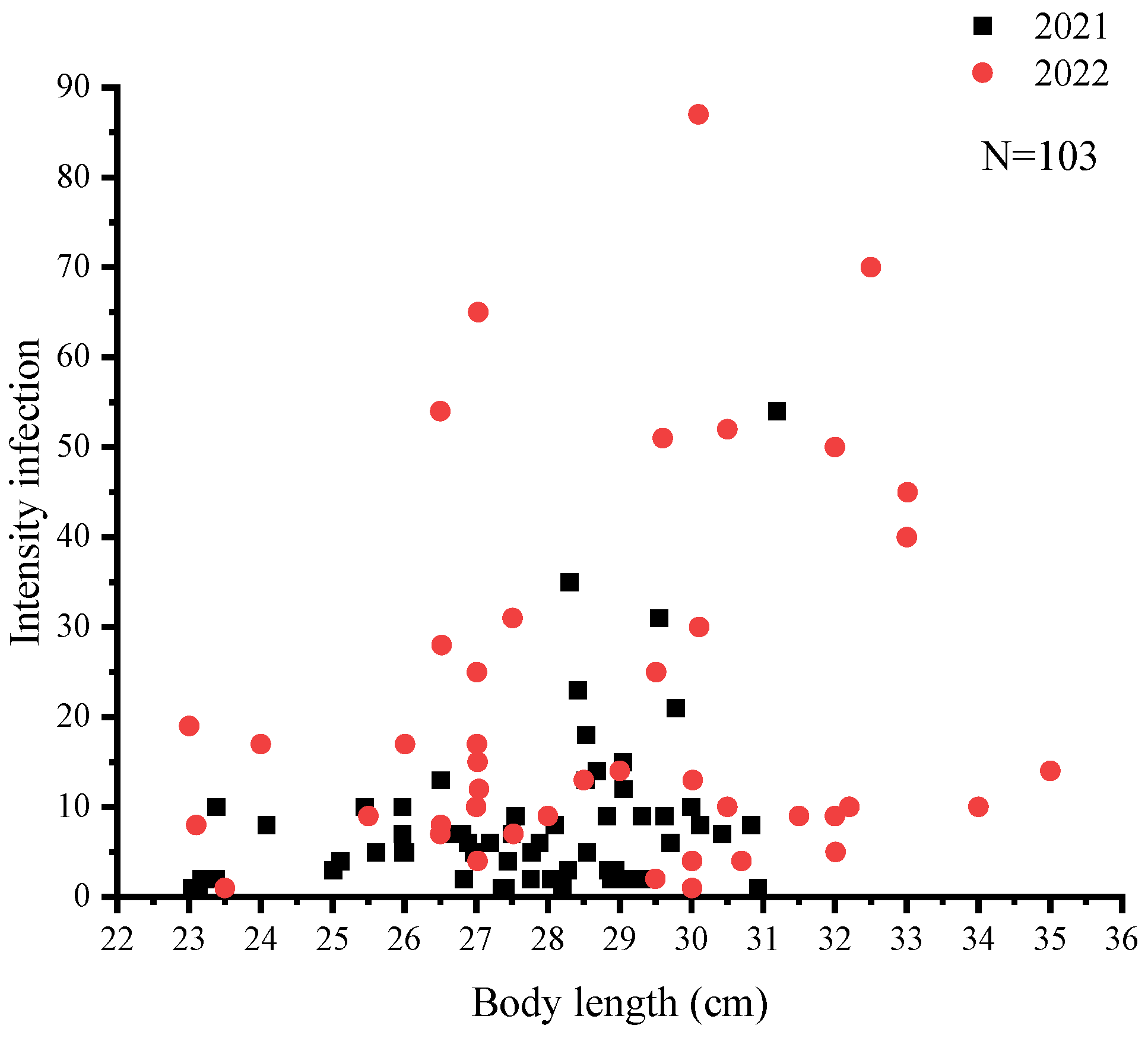
A total of 1422 nematodes were detected in 103 C. nasus, with a mean intensity of 13.81 ± 16.45. The mean intensity of C. nasus that were sampled in 2021 and 2022 was 8.18 ± 9.12 and 21.63 ± 20.80, respectively. The mean intensity was higher at all sampling sites in 2022 than that in 2021, and there was a significant difference (p < 0.05) in mean intensity at the same sampling site in 2022 compared to 2021 (Figure 2). The highest infection intensity was detected in a female C. nasus, which was approximately 30.10 cm in body length, with a total of 87 nematodes. Taking gender differences into account, the results indicated a significant difference in nematode infection intensities between males and females in 2021 (p < 0.05), whereas such sex-related characteristics were not determined in 2022 (Figure 3). In addition, the number of individuals with an intensity of 1–10 was the highest in this study (Figure 4). Finally, a not strict trend of nematodes with an increase in body length in two years was observed (Figure 5).
3.2. Infection Difference among Organs
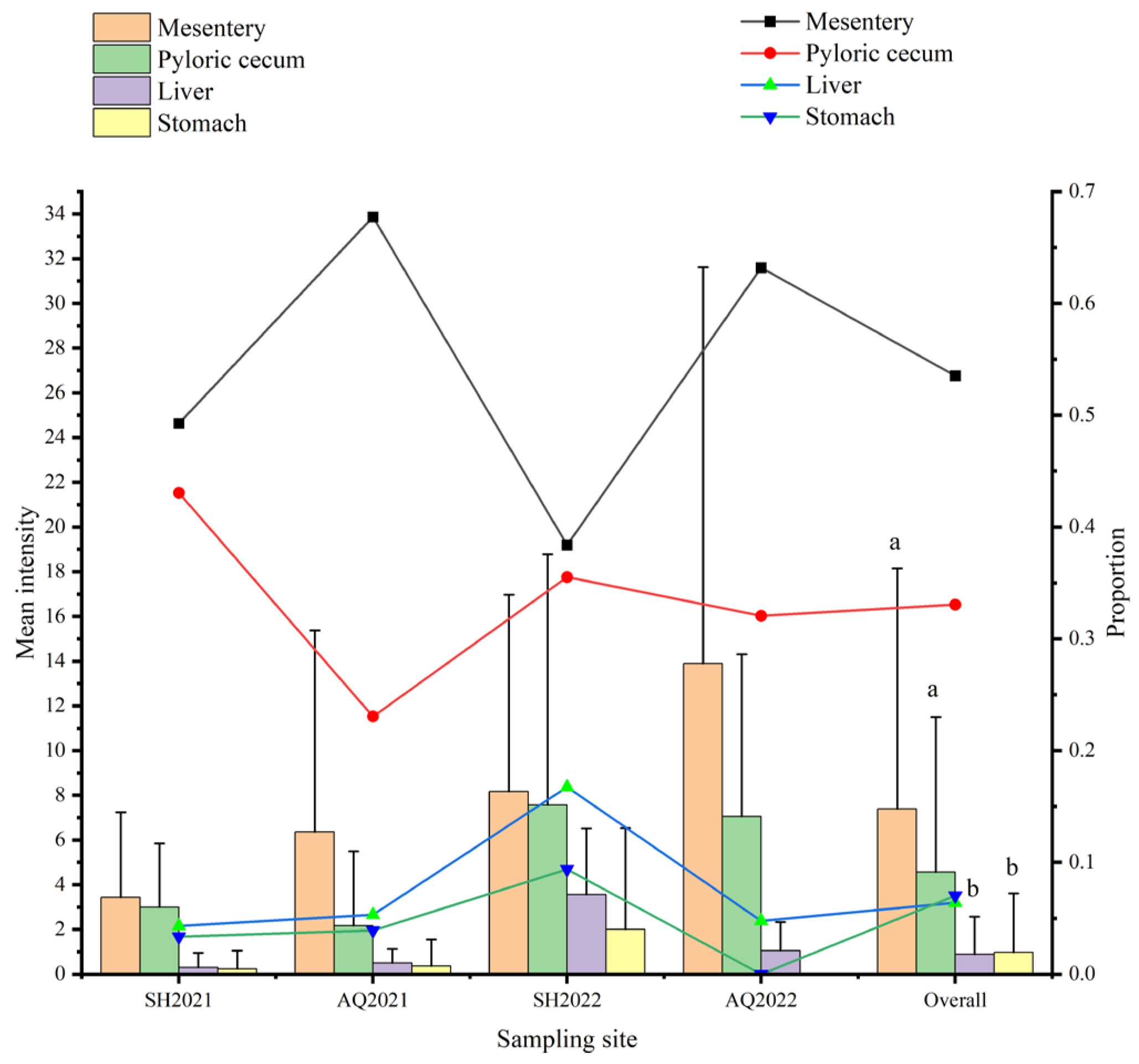
In this study, the mesentery exhibited the highest proportion of nematodes among all tissues of C. nasus, with a total count of 761 nematodes, accounting for 53.52%. There were 470 nematodes in the pyloric cecum, accounting for 33.05%. In addition, the stomach and liver exhibited the lower parasitic level of nematodes, with a count of 100 and 91, accounting for 7.03% and 6.40%, respectively. The mean infection intensity of the mesentery and pyloric cecum was found to be significantly higher (p < 0.05) than that of other organs and tissues (Figure 6). The mesentery exhibited the highest percentage of nematode detection in C. nasus at various sampling locations and times, exceeding or almost 50% in all sampling locations except for the Shanghai section of Yangtze River in 2022. In addition, the stomach consistently displayed the lowest percentage, except for the Shanghai section of Yangtze River in 2022. Interestingly, no nematodes were detected in the stomach of C. nasus at the Anqing section of Yangtze River from 2022. Furthermore, a comparison between mean intensities of nematodes in tissues or organs from different years revealed that the mean intensity was higher in 2022 compared to 2021 (Figure 6).
3.3. Composition of Nematode Species
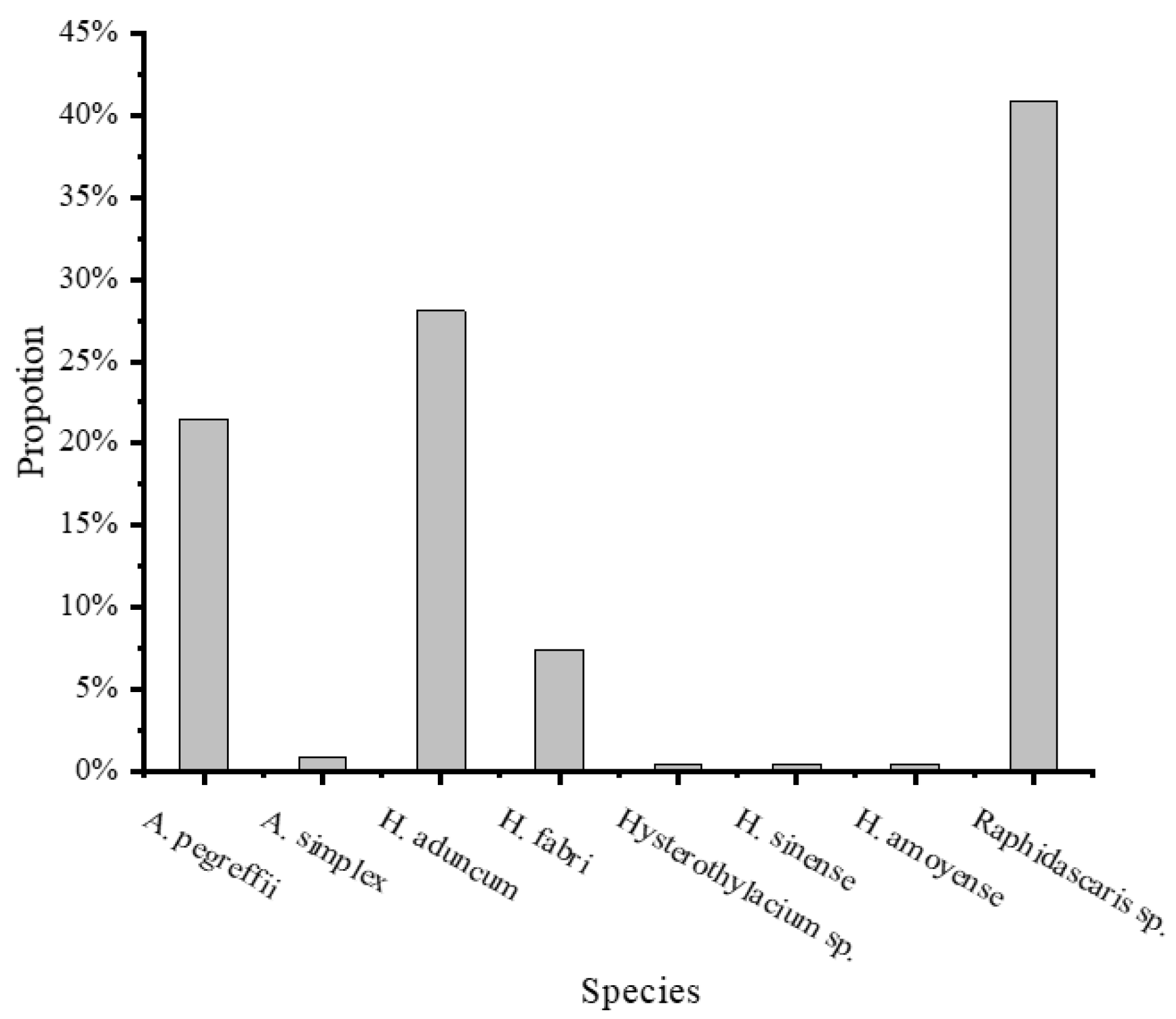
A total of 242 ITS sequences of the 250 nematodes isolated from C. nasus (n = 26) were successfully obtained, accounting for 96.8%. Based on morphological characters and ITS sequence alignment search at the GenBank database, eight nematode species belonging to two families, Anisakidae and Raphidascarididae, were identified, respectively. There were two anisakid nematodes closely related to the following taxa in GenBank: Anisakis pegreffii and Anisakis simplex, and six raphidascaridid nematodes: Hysterothylacium aduncum, Hysterothylacium fabri, Hysterothylacium sinense, Hysterothylacium amoyense, and Hysterothylacium sp., Raphidascaris sp. (Table 2). In the 242 identified nematodes, Raphidascaris sp. was the dominant species, contributing 40.9% of individuals, and it was followed by H. aduncum and A. pegreffii, with 28.1% and 21.5%, respectively. H. fabri was 7.4%, and H. sinense, H. amoyense, and Hysterothylacium sp. accounted for only 0.4%, respectively (Figure 7).
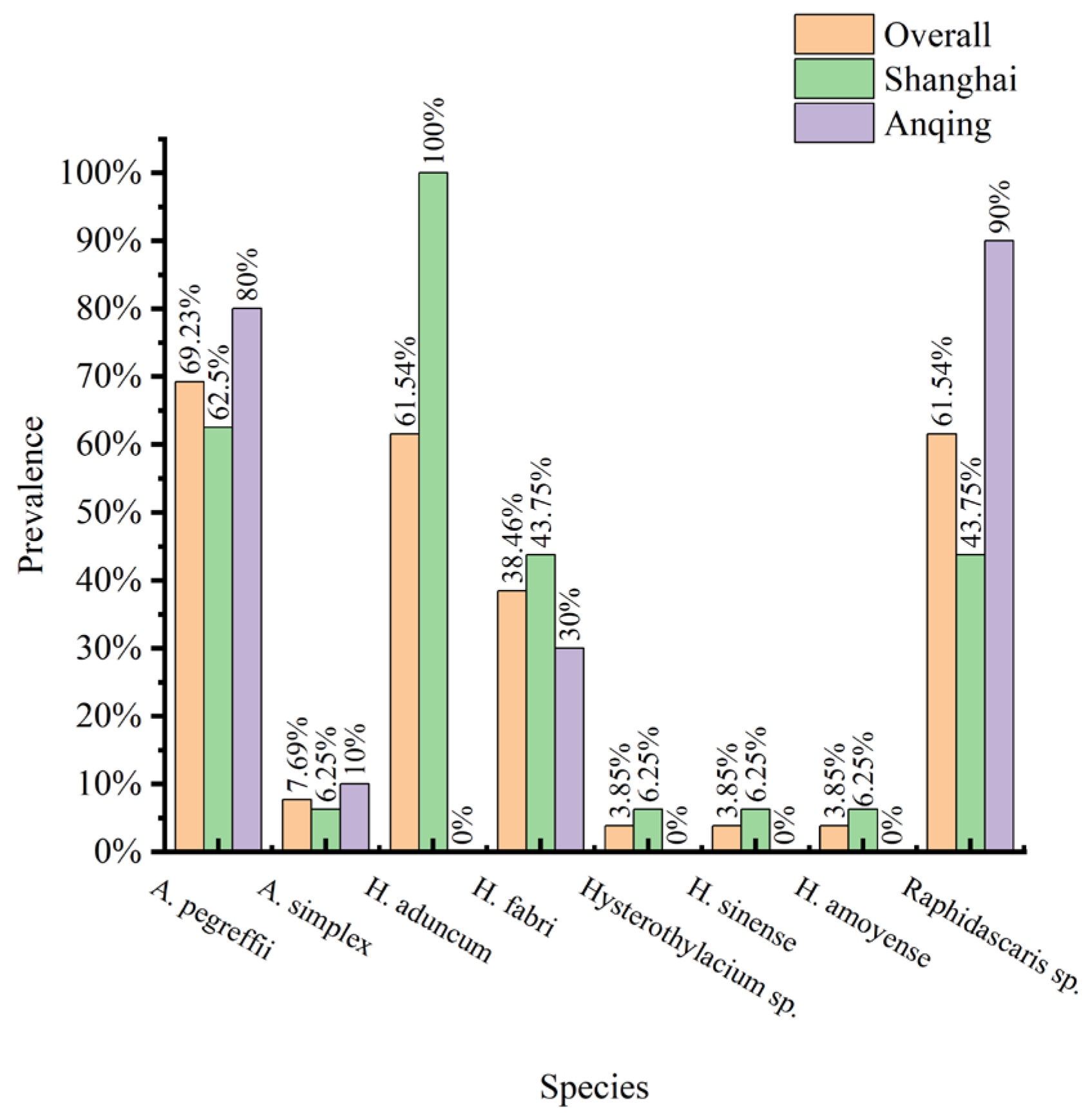
In the 26 C. nasus from which nematodes were isolated and identified by ITS, A. pegreffii had the highest infection prevalence at 69.23%. The infection prevalence of H. aduncum and Raphidascaris sp. was both 61.54%. However, it is worth noting that the infection rate of H. aduncum at the Shanghai section (C. nasus n = 16) was 100%, and it was not detected at the Anqing section (C. nasus n = 10) (Figure 8).
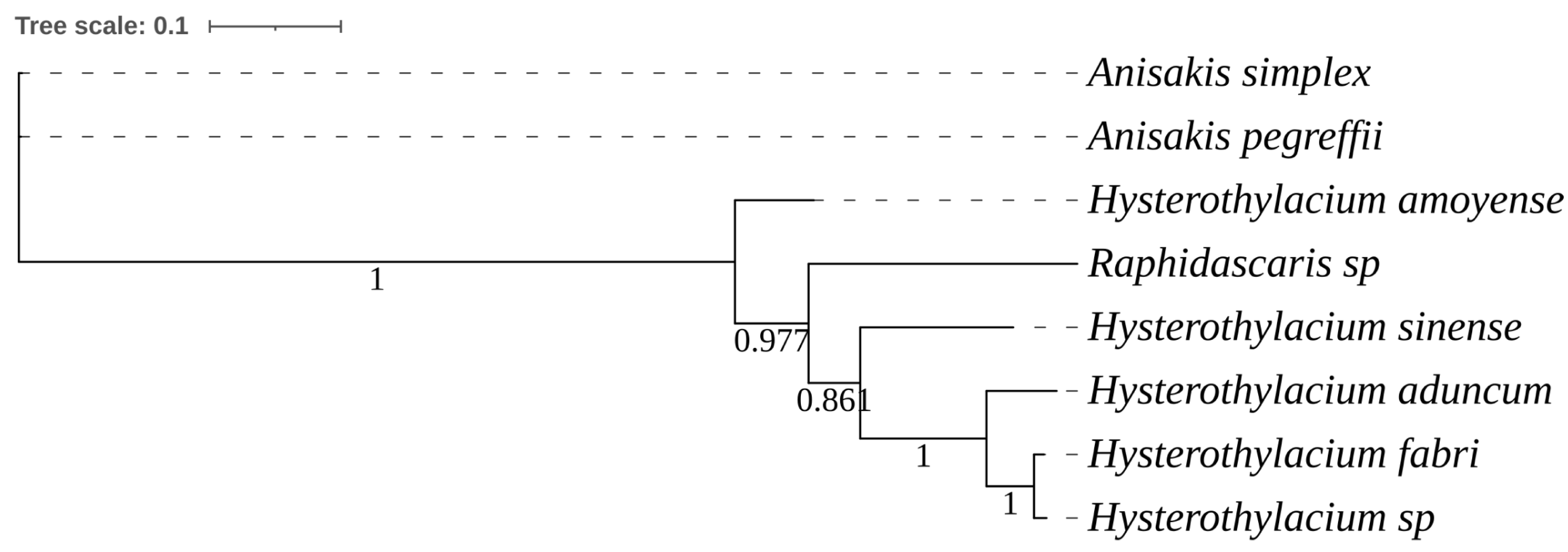
Figure 9 shows a phylogenetic tree constructed from eight ITS sequences of nematode specimens in this study. The nematodes from the family Raphidascarididae formed a separate clade.
3.4. Organ Tropism
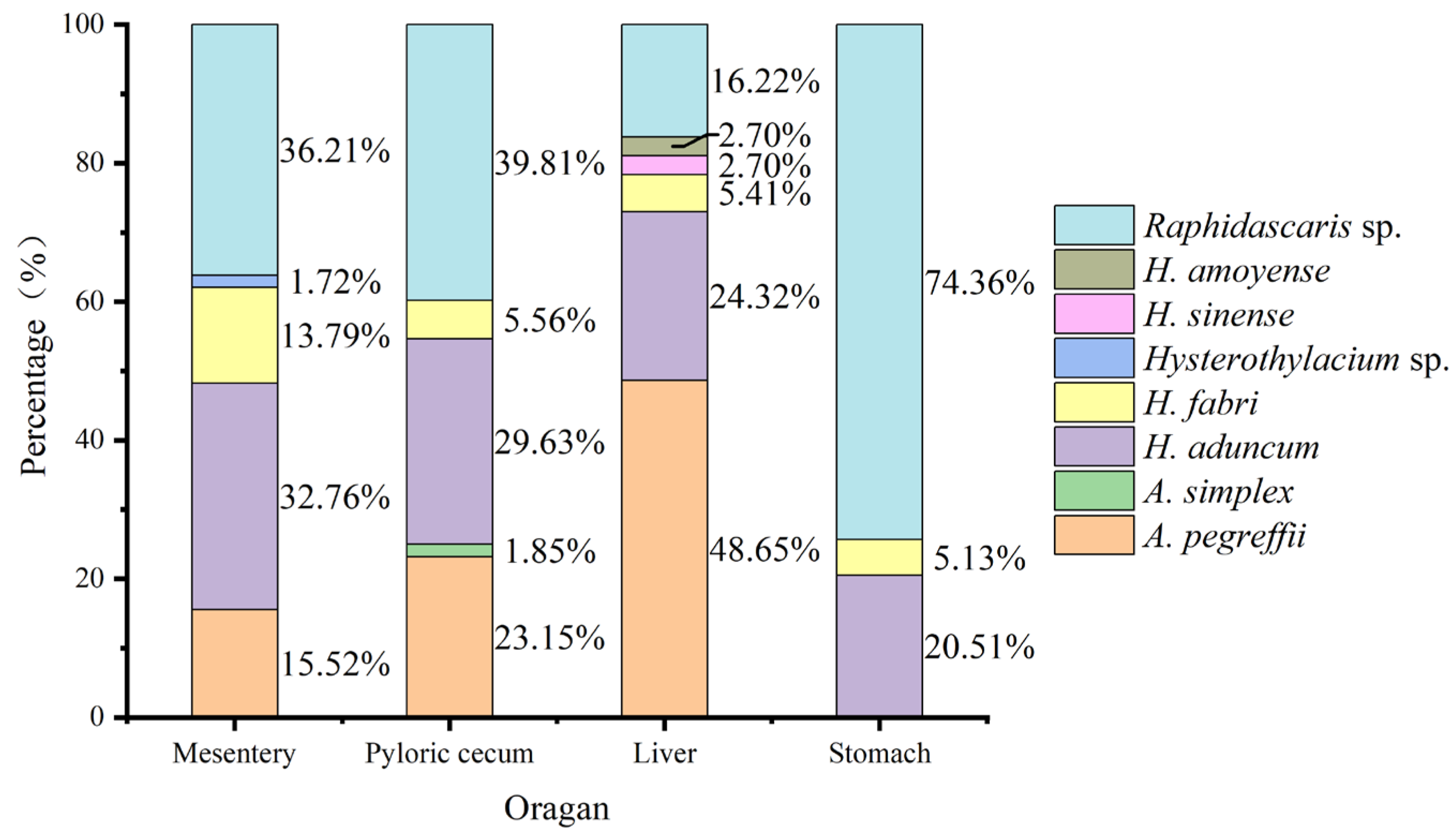
Raphidascaris sp., H. fabri, and H. aduncum did not show strict organ/tissue preference, and were found in the pyloric cecum, stomach, mesentery, and liver of C. nasus. The dominant species Raphidascaris sp. consisted of 74.36%, 39.81%, and 36.21% of individuals in the stomach, mesentery, and cecum, respectively. A. pegreffii occurred in the pyloric cecum, mesentery, and liver, but not in the stomach, and contributed 48.65% of individual numbers in the liver. In the C. nasus examined here, a few A. simplex and H. sinense were detected in the mesentery and liver, respectively (Figure 10).
3.5. Community Structure of Nematodes
The abundance of nematodes in C. nasus was 8, and the average richness was 2.73 ± 1.22. The Margalef richness index was calculated as 1.28, while the Brillouin diversity index was 1.30. The Shannon–Wiener diversity index was 1.35 and the Pielou homogeneity index showed a value of 0.65. The population characteristic values of each nematode in C. nasus were employed to assess the distribution types (Table 3). The results showed that Raphidascaris sp., A. pegreffii, and H. aduncum had an aggregated distribution pattern, whereas H. sinense, H. amoyense, Hysterothylacium sp., and A. simplex displayed random distribution.
Furthermore, community characteristics of nematodes in C. nasus from the Shanghai and Anqing sections of the Yangtze River were assessed, respectively (Table 4 and Table 5). Raphidascaris sp. was found to have an aggregated distribution, while A. pegreffii had an aggregated distribution at the Shanghai section but not in the Anqing section.
4. Discussion
The longjaw tapertail anchovy C. nasus is a commercially valuable fish in China, and was also listed as an endangered (EN) species in a 2018 Red List of Threatened Species report from the International Union for Conservation of Nature (IUCN) (www.iucnredlist.org (accessed on 15 October 2023) [32,33]. In this study, anadromous C. nasus collected from the channel of the Yangtze River at Shanghai and Anqing sections in 2021 and 2022 showed a very high infection rate of nematodes, with 100% prevalence and a mean intensity of 13.81 ± 16.45 (Table 1, Figure 2). In a C. nasus individual, even over 80 nematodes were detected. Eight ascaridoid nematodes belonging to the family Anisakidae and Raphidascarididae were detected and identified in the internal organs, body cavity, mesentery, pyloric cecum, stomach, and liver. The nematode infection posed significant challenges to the health of C. nasus, and the high infection rate of Anisakis spp. was a potential zoonosis risk to the consumer. Additionally, due to the limitation of visual examination employed in this study, some nematode larvae, such as Contracaecum spp. and Spiroxys spp., with a low infection rate in C. nasus may be overlooked [34,35,36].
In the present study, nematode infection in C. nasus was 100%, which is much higher than previous studies [24,25,28,36]. And, it is worth noting that the mean intensity of nematodes in C. nasus between 2021 and 2022 was significantly increased from 8.18 ± 9.12 to 21.63 ± 20.80. The mean intensity in 2022 was the highest recorded, at almost twice the mean intensity of 10.16 ± 9.67 reported by Dai et al. [25] in 2018. This could be attributed to the fact that C. nasus consumed a substantial number of copepods and krill that had been infected by many nematodes, indirectly indicating the severity of offshore organism infections. Meanwhile, according to the results in the present study, the infection rate of nematodes has no significant correlation with the body length. However, if the host diet does not change, the infection rate and intensity of parasites are usually positively correlated with body length [37]. This discrepancy in C. nasus may be attributed to the larger individuals being parasitized with more nematodes, leading to potential damage or mortality during migration processes and consequently resulting in lower nematode abundance in the large C. nasus examined here.
The intensity of nematodes in C. nasus showed no significant variation during the migration at the Shanghai and Anqing sections in the Yangtze River (Figure 2). This could be ascribed to nematodes being in the abdominal cavity and not being susceptible to osmotic pressure generated by migration and changes in the external environment [24]. The nematodes were frequently detected from the mesentery, pyloric cecum, stomach, and liver of C. nasus. Specifically, the highest percentage of infection was found in the mesentery, followed by the pyloric cecum, while the lowest percentage was observed in the stomach or liver. In this study, there were a total of 53.52% nematodes parasitized in the mesentery and 33.05% in the pyloric cecum. The results were consistent with the report of Dai et al. [25]. In addition, nematodes in C. nasus exhibited organ preference, with the highest prevalence observed in the mesentery, constituting over 50% of total nematode occurrences at each site.
Nematode infection caused a certain degree of damage to the liver of C. nasus (Figure 1), and this phenomenon was also reported by Liu et al. [10]. Meanwhile, due to density or developmental requirements, existing nematodes in the host migrated among different organs [38,39]. Balbuena et al. [38] reported that the larvae of H. aduncum could be transferred from the rectum to brain in herring under laboratory rearing conditions. The migration of nematodes would lead to the puncture of the stomach and other organs, which is associated with inflammation and organ damage. Previous studies have demonstrated that ascaridoid infection elicits inflammatory responses and adaptive immunity in C. nasus [28,40,41]. A high nematode infection rate may result in mortality during migration and failure to reach spawning grounds, consequently contributing to a decline in the C. nasus population.
The species composition and abundance of parasites infecting fish are affected by a variety of factors, such as the physicochemical properties of the water, feeding, and the seasons [42]. The anadromous C. nasus exhibits a complex life cycle characterized by feeding offshore and breeding in freshwater (the Yangtze River and its tributaries), followed by a return to diverse habitats encompassing seawater and brackish water after spawning [43]. In this study, eight ascaridoid nematodes were detected in the population of C. nasus migrating from offshore to the spawning grounds. All of these identified nematodes are known as parasites in marine environments [44]. However, nematode Contracaecum spp. and Eustrongylides spp., which were previously reported by Song [36], were not detected in this survey. The difference may be attributed to either the change in nematode community in marine environments or the limitations of the parasite investigation in this study. Moreover, some nematodes had a stronger lethal ability, such as Eustrongylides spp. [45], and result in the death of the infected C. nasus. Meanwhile, no freshwater nematodes were detected in the C. nasus populations as per the previous report [24]. C. nasus refrained from feeding during migration and avoided the infection of freshwater nematodes [46]. Ascaridoid nematodes mainly infect marine fish, and have been detected in fish, krill, and other animals in the East China Sea and the Yellow Sea, exhibiting a broad range of hosts [47,48]. C. nasus serves as a common second intermediate host of ascaridoid nematodes [49]. Krill and copepods, as first intermediate hosts of ascaridoid nematodes, are the main food sources of C. nasus. C. nasus was infected by nematodes mainly through the intaking of krill harboring the larvae of nematodes. Therefore, the variation in nematode species and dominant species observed in this study may be attributed to the changes in natural baits and parasites in coastal waters. In recent reports, a decline in ascaridoids in marine fish has been observed in Japan and Australia [50,51], and this has been ascribed to lower populations of zooplankton and crustaceans [52]. Here, the inconsistent observations were not clear, and further studies are needed to assess the nematode infection in marine fish from China’s offshore water.
Tissue or organ tropism in many parasites is essential for their development and transmission. In nematode genus Contracaecum, Contracaecum radiatum was often found in the gastric wall of the fish host [53]; however, Contracaecum osculatum preferred the liver and body cavity [54]. In arctic ice fish, Ning [55] also discovered that a significant proportion of nematodes parasitized in the liver due to the larger volume and more abundant nutritional resources compared to other tissues and organs. In C. nasus detected here, A. pegreffii, Raphidascaris sp., and H. aduncum were the dominant parasitic nematodes; however, no obvious organ/tissue preference was observed (Figure 10), and they were found in the pyloric cecum, stomach, mesentery, and liver of C. nasus. A. pegreffii accounted for the highest proportion (48.65%) in the liver, and Raphidascaris sp. had the highest proportions in the other tissues: 36.21% in the pyloric cecum, 39.81% in the mesentery, and 74.36% in the stomach. Interestingly, A. simplex and H. sinense were only detected in the mesentery and liver, respectively. However, in this study, only limited nematodes from 26 C. nasus individuals were identified using both morphological and molecular data. Therefore, the tissue or organ distribution of nematodes with a low infection rate may be overlooked.
The application of parasites as biological indicators for the fish host population and environment assessment has recently attracted much attention [56,57]. Xu et al. [22] found that A. simplex only infected the anadromous C. nasus in the Yangtze River, and was not detected in the freshwater residents C. nasus. A. simplex was considered as a “biological indicator” of the anadromous C. nasus in the Yangtze River. Li et al. [23] also reported that parasites infecting C. nasus could be used to indicate the migration between the coast, estuary, and Yangtze River. However, according to the recent parasite investigation on anadromous C. nasus in the Yangtze River, A. simplex had a low infection rate, while A. pegreffii, Raphidascaris sp., and H. aduncum were the common and dominant parasites [25,26].
Author Contributions
B.X. designed the study. Q.Z. wrote the manuscript. Q.Z., L.W. and C.Y. carried out the experiment. B.X. and K.L. provided resources and technical support. All authors reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the earmarked fund for China Agriculture Research System of MOF and MARA (CARS-45) and National Natural Science Foundation of China (32073019).
Institutional Review Board Statement
The animal study protocol was approved by the Committee on the Ethics of Animal Experiments of the Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences (Authorization Number: 20221223).
Data Availability Statement
The authors confirm that the data supporting the findings of this study are available within the manuscript and table.
Conflicts of Interest
The authors declare no conflicts of interests.
References
- Hochberg, N.S.; Hamer, D.H.; Hughes, J.M.; Wilson, M.E. Anisakidosis: Perils of the Deep. Clin. Infect. Dis. 2010, 51, 806–812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, Z.; Chen, H.-X.; Guo, N.; Li, L. Anisakid and Raphidascaridid nematodes (Ascaridoidea) infection in the important marine food-fish Lophius litulon (Jordan) (Lophiiformes: Lophiidae). Int. J. Food Microbiol. 2018, 284, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, S.; Magnabosco, C.; Civettini, M.; Boffo, L.; Mingarelli, G.; Buratti, P.; Giovanardi, O.; Fortuna, C.M.; Arcangeli, G. Survey of Anisakis Sp. and Hysterothylacium Sp. in sardines and anchovies from the North Adriatic Sea. Int. J. Food Microbiol. 2015, 200, 18–21. [Google Scholar] [CrossRef]
- Li, L.; Zhao, J.-Y.; Chen, H.-X.; Ju, H.-D.; An, M.; Xu, Z.; Zhang, L.-P. Survey for the presence of ascaridoid larvae in the cinnamon flounder Pseudorhombus Cinnamoneus (Temminck & Schlegel) (Pleuronectiformes: Paralichthyidae). Int. J. Food Microbiol. 2017, 241, 108–116. [Google Scholar] [CrossRef]
- Klimpel, S.; Palm, H.W.; Rückert, S.; Piatkowski, U. The life cycle of Anisakis simplex in the Norwegian Deep (Northern North Sea). Parasitol. Res. 2004, 94, 1–9. [Google Scholar] [CrossRef]
- Murphy, T.M.; Berzano, M.; O’Keeffe, S.M.; Cotter, D.M.; McEvoy, S.E.; Thomas, K.A.; Maoiléidigh, N.P.; Whelan, K.F. Anisakid larvae in Atlantic Salmon (Salmo salar L.) grilse and post-smolts: Molecular identification and histopathology. J. Parasitol. 2010, 96, 77–82. [Google Scholar] [CrossRef]
- Shamsi, S.; Barton, D.P.; Zhu, X. Description and characterisation of terranova Pectinolabiatan. Sp. (Nematoda: Anisakidae) in great hammerhead shark, Sphyrna mokarran (Rüppell, 1837), in Australia. Parasitol. Res. 2019, 118, 2159–2168. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S.; Briand, M.J.; Justine, J.-L. Occurrence of Anisakis (Nematoda: Anisakidae) larvae in unusual hosts in Southern Hemisphere. Parasitol. Int. 2017, 66, 837–840. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Bosi, G.; DePasquale, J.A.; Manera, M.; Giari, L. Fish innate immunity against intestinal helminths. Fish Shellfish Immunol. 2016, 50, 274–287. [Google Scholar] [CrossRef]
- Liu, K.; Yin, D.; Shu, Y.; Dai, P.; Yang, Y.; Wu, H. Transcriptome and metabolome analyses of Coilia nasus in response to Anisakidae parasite infection. Fish Shellfish Immunol. 2019, 87, 235–242. [Google Scholar] [CrossRef]
- Mattiucci, S.; Nascetti, G. Chapter 2 Advances and trends in the molecular systematics of Anisakid nematodes, with implications for their evolutionary ecology and host—Parasite co-evolutionary processes. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2008; Volume 66, pp. 47–148. [Google Scholar]
- Levsen, A.; Berland, B. Anisakis Species. In Fish Parasites: Pathobiology and Protection; CABI: Wallingford, UK, 2012; pp. 298–309. [Google Scholar] [CrossRef]
- Shamsi, S.; Barton, D.P. A critical review of Anisakidosis cases occurring globally. Parasitol. Res. 2023, 122, 1733–1745. [Google Scholar] [CrossRef]
- Jiang, T.; Yang, J.; Liu, H.; Shen, X. Life History of Coilia nasus from the Yellow Sea inferred from Otolith Sr:Ca Ratios. Environ. Biol. Fishes 2012, 95, 503–508. [Google Scholar] [CrossRef]
- Feng, X.; Yang, X.; Ruan, J.; Wang, Y.; Zhou, Y.; Xu, D.; Fang, D. Molecular cloning and characteristics of DnaJa1and DnaJb1 in Coilia nasus: Possible function involved in oogenesis during spawning migration. BMC Dev. Biol. 2019, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Dou, S.Z.; Yamada, Y.; Okamura, A.; Tanaka, S.; Shinoda, A.; Tsukamoto, K. Observations on the spawning behavior of artificially matured Japanese eels Anguilla japonica in captivity. Aquaculture 2007, 266, 117–129. [Google Scholar] [CrossRef]
- Yuan, C. Spawning migration of Coilia nasus. Bull. Biol. 1987, 12, 1–3. (In Chinese) [Google Scholar]
- Jiang, T.; Yang, J.; Lu, M.J.; Liu, H.B.; Chen, T.T.; Gao, Y.W. Discovery of a spawning area for anadromous Coilia nasus Temminck et Schlegel, 1846 in Poyang Lake, China. J. Appl. Ichthyol. 2017, 33, 189–192. [Google Scholar] [CrossRef]
- Xuan, Z.; Jiang, T.; Liu, H.; Chen, X.; Yang, J. Mitochondrial DNA and microsatellite analyses reveal strong genetic differentiation between two types of estuarine tapertail anchovies (Coilia) in Yangtze River Basin, China. Hydrobiologia 2021, 848, 1409–1431. [Google Scholar] [CrossRef]
- Liang, C.; Pauly, D. Growth and mortality of exploited fishes in China’s coastal seas and their uses for yield-per-recruit analyses. J. Appl. Ichthyol. 2017, 33, 746–756. [Google Scholar] [CrossRef]
- Ma, J.; Li, B.; Zhao, J.; Wang, X.; Hodgdon, C.T.; Tian, S. Environmental influences on the spatio-temporal distribution of Coilia nasus in the Yangtze River Estuary. J. Appl. Ichthyol. 2020, 36, 315–325. [Google Scholar] [CrossRef]
- Xu, H.-N.; Sun, C.-B.; Tong, Y.-R.; Tao, J.-Y. The biological indicator of the spawning migration of anchovy (Coilia ectenes Jordan et Seale) in Yangtze River. J. Nanjing Univ. (Nat. Sci.) 1978, 3, 85–91. (In Chinese) [Google Scholar]
- Li, W.X.; Song, R.; Wu, S.G.; Zou, H.; Nie, P.; Wang, G.T. Seasonal occurrence of helminths in the anadromous fish Coilia nasus (Engraulidae): Parasite indicators of fish migratory movements. J. Parasitol. 2011, 97, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Li, W.X.; Zou, H.; Wu, S.G.; Song, R.; Wang, G.T. Richness and diversity of helminth communities in the Japanese grenadier anchovy, Coilia nasus, during its anadromous migration in the Yangtze River, China. J. Parasitol. 2012, 98, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Guo, Q.; Tian, J.; Ma, F.; Wang, Y.; Liu, K. Nematode infection status and temporal characteristic of estuarine tapertail anchovy Coilia nasus in Anqing section of the Yangtze River. J. Dalian Ocean Univ. 2022, 37, 464–470. (In Chinese) [Google Scholar] [CrossRef]
- Cheng, X.; Yang, Y.; Ying, C.; Ma, F.; Deng, Y.; Liu, K. Investigation on Anisakidae nematode parasitic status of Coilia nasus at the early stages of banning fishing in Yangtze River. Trans. Oceanol. Limnol. 2023, 45, 142–149. (In Chinese) [Google Scholar] [CrossRef]
- Williams, J.D.; Boyko, C.B. The global diversity of parasitic isopods associated with crustacean hosts (Isopoda: Bopyroidea and Cryptoniscoidea). PLoS ONE 2012, 7, e35350. [Google Scholar] [CrossRef] [PubMed]
- Liu, K. Research on Immune Adaptive Response of Coilia nasus Infected with Nematode in Yangtze River Based on Multi-Omics Techniques. Ph.D. Thesis, Anhui Normal University, Wuhu, China, 2019. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology Meets Ecology on Its Own Terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Lü, J.-Y.; Zeng, H.; Yang, D.-W.; Chen, Z.-S. Studies on community ecology of helminthes parasitic in Mugil cephalus in Guangdong Province, China. Acta Ecol. Sin. 2001, 21, 1003–1008. (In Chinese) [Google Scholar]
- Dai, P.; Yan, Y.; Zhu, X.; Tian, J.; Ma, F.; Liu, K. Status of Coilia nasus resources in the National Aquatic Germplasm Resources Conservation Area in the Anqing Section of the Yangtze River. J. Fish. Sci. China 2020, 27, 1267–1276. (In Chinese) [Google Scholar]
- Dai, P.; Ma, F.-J.; Tian, J.-L.; Wang, Y.-P.; Yang, Y.-P.; Liu, K. Community structure and infection characteristics of nematodes in the Coilia nasus in Anqing section of the Yangtze River. Acta Hydrobiol. Sin. 2023, 47, 917–923. (In Chinese) [Google Scholar]
- Shamsi, S.; Turner, A.; Wassens, S. Description and genetic characterization of a new Contracaecum Larval Type (Nematoda: Anisakidae) from Australia. J. Helminthol. 2018, 92, 216–222. [Google Scholar] [CrossRef]
- Shamsi, S.; Pearce, L.; Zhu, X. Characterisation of nematode larvae found in a vulnerable native Australian fish, the Southern Pygmy Perch, Nannoperca Australis Günther. Mar. Freshw. Res. 2023, 74, 1095–1101. [Google Scholar] [CrossRef]
- Song, R. The Ecological Investigation of the Helminths in Coilia nasus. Master’s Thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan, China, 2011. (In Chinese). [Google Scholar]
- Xi, B.-W.; Wang, G.-T.; Wu, S.-G.; Gao, D.; Zou, H.; Yao, W.-J.; Nie, P. Community structure of the intestinal helminths of the Chinese hooksnout carp, Opsariichthys Bidens (g(u)nther), from the Danjangkou reservio. Acta Hydrobiol. Sin. 2009, 33, 177–182. (In Chinese) [Google Scholar] [CrossRef]
- Balbuena, J.A.; Karlsbakk, E.; Kvenseth, A.M.; Saksvik, M.; Nylund, A. Growth and emigration of third-stage larvae of Hysterothylacium aduncum (Nematoda: Anisakidae) in larval herring Clupea harengus. J. Parasitol. 2000, 86, 1271–1275. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.H.; Bresciani, J.; Buchmann, K. Interactions between Ecto- and Endoparasites in Trout Salmo Trutta. Vet. Parasitol. 2002, 103, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-L. Anisakidae Nematodes on Digestive Tract Tissue Damage and the Effect of Immune Response of Coilia nasus. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2022. (In Chinese). [Google Scholar]
- Ying, C.; Fang, X.; Wang, H.; Yang, Y.; Xu, P.; Liu, K.; Yin, G. Anisakidae parasitism activated immune response and induced liver fibrosis in wild anadromous Coilia nasus. J. Fish Biol. 2022, 100, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-X.; Wang, G.-T. Helminth Communities in Coilia nasus from Anadromous, Freshwater and Landlocked Stock. Chin. J. Zool. 2014, 49, 233–243. (In Chinese) [Google Scholar] [CrossRef]
- Li, M.; Jiang, T.; Chen, T.; Liu, H.; Yang, J. Otolith microchemistry of the estuarine tapertail anchovy Coilia nasus from the Anqing section of the Yangtze River and its significance for migration ecology. Acta Ecol. Sin. 2017, 37, 2788–2795. (In Chinese) [Google Scholar]
- Cao, Z.; Liu, J.; He, F.; Lin, R.; Zhu, X. Overview of Anisakis disease. J. Trop. Med. 2004, 4, 494–497. (In Chinese) [Google Scholar]
- Shamsi, S.; Francis, N.; Masiga, J.; Barton, D.P.; Zhu, X.; Pearce, L.; McLellan, M. Occurrence and characterisation of Eustrongylides species in Australian Native birds and fish. Food Waterborne Parasitol. 2023, 30, e00189. [Google Scholar] [CrossRef]
- Li, L. Systematics of the Genus Hysterothylacium from Marine Fishes from Yellow Seea, China (Ascaridoidea: Raphidascaridae). Master’s Thesis, Hebei Normal University, Shijiazhuang, China, 2007. (In Chinese). [Google Scholar]
- Li, X.; Cao, T.; Du, A.; Cao, W. Preliminary investigation on infection of Anisakis larvae in Marine fish in the East China Sea. Chin. J. Vet. Med. 2009, 45, 76–77. (In Chinese) [Google Scholar]
- Xu, Z. Preliminary Studies on Ecology of Parasitic Nematodes from Several Marine Fish from Yellow Sea, China. Master’s Thesis, Hebei Normal University, Shijiazhuang, China, 2007. (In Chinese). [Google Scholar]
- Bouree, P.; Paugam, A.; Petithory, J.-C. Anisakidosis: Report of 25 Cases and Review of the Literature. Comp. Immunol. Microbiol. Infect. Dis. 1995, 18, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Takano, T.; Iwaki, T.; Waki, T.; Murata, R.; Suzuki, J.; Kodo, Y.; Kobayashi, K.; Ogawa, K. Species composition and infection levels of Anisakis (Nematoda: Anisakidae) in the Skipjack Tuna Katsuwonus Pelamis (Linnaeus) in the Northwest Pacific. Parasitol. Res. 2021, 120, 1605–1615. [Google Scholar] [CrossRef] [PubMed]
- Hossen, M.S.; Shamsi, S. Zoonotic nematode parasites infecting selected edible fish in New South Wales, Australia. Int. J. Food Microbiol. 2019, 308, 108306. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S. The occurrence of Anisakis spp. in Australian Waters: Past, present, and future trends. Parasitol. Res. 2021, 120, 3007–3033. [Google Scholar] [CrossRef] [PubMed]
- Palm, H.W.; Reimann, N.; Spindler, M.; Plötz, J. The role of the rock cod Notothenia coriiceps Richardson, 1844 in the life-cycle of Antarctic parasites. Polar Biol. 1998, 19, 399–406. [Google Scholar] [CrossRef]
- Klöser, H.; Plötz, J.; Palm, H.; Bartsch, A.; Hubold, G. Adjustment of anisakid nematode life cycles to the high antarctic food web as shown by Contracaecum radiatum and C. osculatum in the Weddell Sea. Antarct. Sci. 1992, 4, 171–178. [Google Scholar] [CrossRef]
- Ning, J. Host-Parasitic Relationships Between Antarctic Icefish (Channichthyidae) And its Parasites, with Focusing on Nematode and Copepod. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2021. (In Chinese). [Google Scholar]
- Zuo, S.; Kania, P.W.; Mehrdana, F.; Marana, M.H.; Buchmann, K. Contracaecum osculatum and other anisakid nematodes in Grey Seals and Cod in the Baltic Sea: Molecular and ecological links. J. Helminthol. 2018, 92, 81–89. [Google Scholar] [CrossRef]
- Palomba, M.; Marchiori, E.; Tedesco, P.; Fioravanti, M.; Marcer, F.; Gustinelli, A.; Aco-Alburqueque, R.; Belli, B.; Canestrelli, D.; Santoro, M.; et al. An update and ecological perspective on certain sentinel helminth endoparasites within the Mediterranean Sea. Parasitology 2023, 150, 1139–1157. [Google Scholar] [CrossRef]
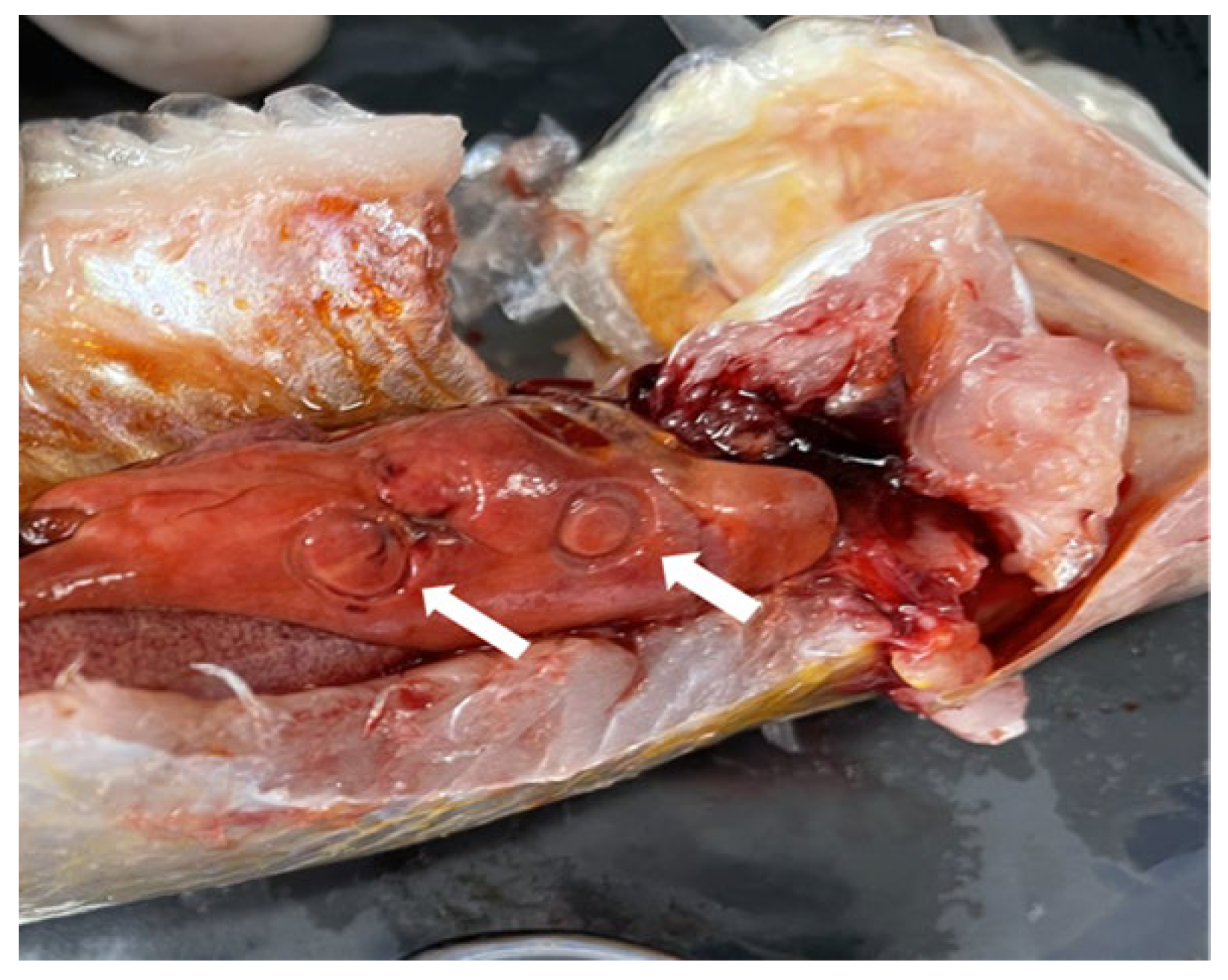
Figure 1.
The nematodes in the liver of C. nasus, indicated with arrows.

Figure 2.
The variations in the mean intensity of nematodes in C. nasus at each sampling site across different years. Abbreviations: SH2021—Shanghai section of the Yangtze River (2021), AQ2021—Anqing section of the Yangtze River (2021), SH2022—Shanghai section of the Yangtze River (2022), AQ2022—Anqing section of the Yangtze River (2022). The numbers enclosed in parentheses indicate sampling date. Bars represent standard deviation. The ‘*’ represents statistically significant disparities (p < 0.05).
Figure 2.
The variations in the mean intensity of nematodes in C. nasus at each sampling site across different years. Abbreviations: SH2021—Shanghai section of the Yangtze River (2021), AQ2021—Anqing section of the Yangtze River (2021), SH2022—Shanghai section of the Yangtze River (2022), AQ2022—Anqing section of the Yangtze River (2022). The numbers enclosed in parentheses indicate sampling date. Bars represent standard deviation. The ‘*’ represents statistically significant disparities (p < 0.05).

Figure 3.
Relationship between the sex of C. nasus and the intensity of infection at various sampling time points. Bars represent standard deviation. The ‘*’ represents statistically significant disparities (p < 0.05).
Figure 3.
Relationship between the sex of C. nasus and the intensity of infection at various sampling time points. Bars represent standard deviation. The ‘*’ represents statistically significant disparities (p < 0.05).

Figure 4.
Number of individuals with different infection intensities of nematodes in C. nasus from two sampling years.
Figure 4.
Number of individuals with different infection intensities of nematodes in C. nasus from two sampling years.

Figure 5.
Relationship between the body length of C. nasus and the intensity of infection in two sampling years.
Figure 5.
Relationship between the body length of C. nasus and the intensity of infection in two sampling years.

Figure 6.
The distribution and mean infection intensity of parasitic nematodes in different tissues and organs of C. nasus at each sampling time and site. Bars represent standard deviation. The presence of distinct letters signifies statistically significant disparities (p < 0.05).
Figure 6.
The distribution and mean infection intensity of parasitic nematodes in different tissues and organs of C. nasus at each sampling time and site. Bars represent standard deviation. The presence of distinct letters signifies statistically significant disparities (p < 0.05).

Figure 7.
The species composition of nematodes isolated from C. nasus (n = 26).

Figure 8.
The species composition of nematodes isolated from C. nasus at each sampling site in 2022.
Figure 8.
The species composition of nematodes isolated from C. nasus at each sampling site in 2022.

Figure 9.
Bayesian inference phylogenetic tree based on the obtained sequences of ITS1-5.8s-ITS2.

Figure 10.
The proportion of parasitic nematode species identified in each organ and tissue of C. nasus in 2022.
Figure 10.
The proportion of parasitic nematode species identified in each organ and tissue of C. nasus in 2022.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biological characteristics of anadromous Coilia nasus migrating to the Yangtze River.
| Site | Year | N | Body Length (cm) | Female to Male Ratio | Prevalence (%) | Mean Abundance |
|---|---|---|---|---|---|---|
| Shanghai section | 2021 | 30 | 26.93 ± 2.26 | 4.00:1 | 100 | 6.97 ± 5.26 |
| Anqing section | 2021 | 30 | 27.40 ± 2.28 | 3.29:1 | 100 | 9.40 ± 11.78 |
| Shanghai section | 2022 | 23 | 28.16 ± 3.12 | 4.75:1 | 100 | 21.35 ± 21.29 |
| Anqing section | 2022 | 20 | 29.50 ± 2.64 | 5.67:1 | 100 | 22.00 ± 20.72 |
Table 2.
Nematodes collected from Coilia nasus (n = 26) and identified by ITS DNA sequence.
| Nematode Species | No. of Individuals Sequenced | Sequence Identity (%) | GenBank Acc. No. | Closely Related Nematode in GenBank | |
|---|---|---|---|---|---|
| Species (Accession) | Sequence Similarity (%) | ||||
| Hysterothylacium aduncum | 68 | 99.4–100 | PP029287 | Hysterothylacium aduncum (MH211517) | 98.3–100 |
| Hysterothylacium fabri | 18 | 99.4–100 | PP029274 | Hysterothylacium fabri (MH211492) | 99.6–100 |
| Hysterothylacium amoyense | 1 | - | PP029291 | Hysterothylacium amoyense (MT269312) | 99.89 |
| Hysterothylacium sinense | 1 | - | PP029292 | Hysterothylacium sinense (MH211574) | 100 |
| Hysterothylacium sp. | 1 | - | PP034301 | Hysterothylacium sp. (MF061683) | 100 |
| Anisakis simplex | 2 | 100 | PP029277 | Anisakis simplex (MT355320) | 100 |
| Anisakis pegreffii | 52 | 99.9–100 | PP029289 | Anisakis pegreffii (MH211473) | 99.6–100 |
| Raphidascaris sp. | 99 | 99.0–100 | PP034302 | Raphidascaris sp. (MW370774) | 99.5–100 |
Table 3.
The population characteristics of nematodes in C. nasus.
| Nematode Species | Mean Value | Variance | Distribution Index | d | Aggregate Index | Infection Index |
|---|---|---|---|---|---|---|
| Anisakis pegreffii | 2.000 | 4.560 | 2.280 | 3.677 | 0.025 | 1.385 |
| Anisakis simplex | 0.077 | 0.074 | 0.960 | −0.072 | −0.040 | 0.006 |
| Hysterothylacium aduncum | 2.615 | 7.206 | 2.755 | 4.737 | 0.026 | 1.609 |
| Hysterothylacium fabri | 0.692 | 1.022 | 1.476 | 1.589 | 0.028 | 0.266 |
| Hysterothylacium sp. | 0.038 | 0.038 | 1.000 | 0.071 | - | 0.001 |
| Hysterothylacium sinense | 0.038 | 0.038 | 1.000 | 0.071 | - | 0.001 |
| Hysterothylacium amoyense | 0.038 | 0.038 | 1.000 | 0.071 | - | 0.001 |
| Raphidascaris sp. | 3.808 | 32.882 | 8.636 | 13.779 | 0.078 | 2.343 |
Table 4.
The population characteristics of nematodes in C. nasus at Shanghai section of the Yangtze River.
Table 4.
The population characteristics of nematodes in C. nasus at Shanghai section of the Yangtze River.
| Nematode Species | Mean Value | Variance | Distribution Index | d | Aggregate Index | Infection Index |
|---|---|---|---|---|---|---|
| Anisakis pegreffii | 1.625 | 4.783 | 2.944 | 4.012 | 0.078 | 1.016 |
| Anisakis simplex | 0.063 | 0.063 | 1.000 | 0.092 | - | 0.004 |
| Hysterothylacium aduncum | 4.250 | 4.600 | 1.082 | 0.313 | 0.001 | 4.250 |
| Hysterothylacium fabri | 0.750 | 1.000 | 1.333 | 0.939 | 0.030 | 0.328 |
| Hysterothylacium sp. | 0.063 | 0.063 | 1.000 | 0.092 | - | 0.004 |
| Hysterothylacium sinense | 0.063 | 0.063 | 1.000 | 0.092 | - | 0.004 |
| Hysterothylacium amoyense | 0.063 | 0.063 | 1.000 | 0.092 | - | 0.004 |
| Raphidascaris sp. | 3.938 | 44.863 | 11.394 | 13.103 | 0.168 | 1.723 |
Table 5.
The population characteristics of nematodes in C. nasus at Anqing section of the Yangtze River.
Table 5.
The population characteristics of nematodes in C. nasus at Anqing section of the Yangtze River.
| Nematode Species | Mean Value | Variance | Distribution Index | d | Aggregate Index | Infection Index |
|---|---|---|---|---|---|---|
| Anisakis pegreffii | 2.600 | 4.044 | 1.556 | 1.168 | 0.022 | 2.080 |
| Anisakis simplex | 0.100 | 0.100 | 1.000 | 0.120 | - | 0.010 |
| Hysterothylacium fabri | 0.600 | 1.156 | 1.926 | 1.765 | 0.185 | 0.180 |
| Raphidascaris sp. | 3.600 | 16.489 | 4.580 | 4.957 | 0.102 | 3.240 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhou, Q.; Wang, L.; Xi, B.; Ying, C.; Liu, K. Ascaridoid Nematodes Infection in Anadromous Fish Coilia nasus from Yangtze River. Diversity 2024, 16, 167. https://doi.org/10.3390/d16030167
AMA Style
Zhou Q, Wang L, Xi B, Ying C, Liu K. Ascaridoid Nematodes Infection in Anadromous Fish Coilia nasus from Yangtze River. Diversity. 2024; 16(3):167. https://doi.org/10.3390/d16030167
Chicago/Turabian StyleZhou, Qingjie, Lijun Wang, Bingwen Xi, Congping Ying, and Kai Liu. 2024. "Ascaridoid Nematodes Infection in Anadromous Fish Coilia nasus from Yangtze River" Diversity 16, no. 3: 167. https://doi.org/10.3390/d16030167
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.