
Caenorhabditis elegans as an In Vivo Model for the Discovery and Development of Natural Plant-Based Antimicrobial Compounds

 , , and
, , and
Abstract
:1. Introduction
2. Current Issues with Antimicrobial Drug Discovery
3. Advantages of C. elegans as a Model for the Screening of Novel Compounds
4. Natural Plants as Sources of Novel Antimicrobial Agents
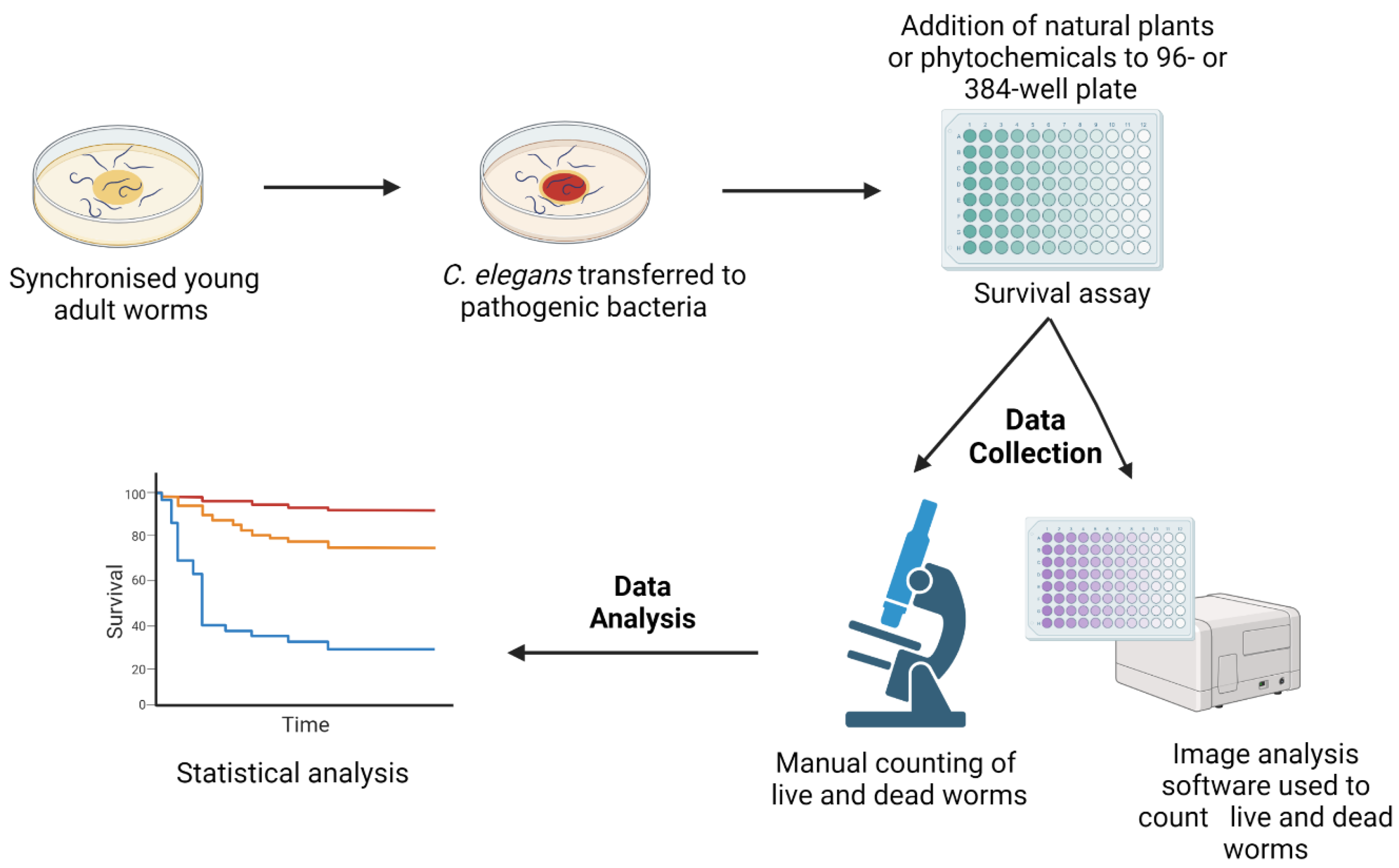
5. Methodology
6. Natural Products Active against Bacterial Infection
7. Natural Products Active against Fungal Infection
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Global Action Plan on Antimicrobial Resistance. Available online: https://www.who.int/publications-detail-redirect/9789241509763 (accessed on 4 April 2023).
- WHO. World Health Statistics. 2022. Available online: https://www.who.int/news/item/20-05-2022-world-health-statistics-2022 (accessed on 4 April 2023).
- Willemsen, A.; Reid, S.; Assefa, Y. A review of national action plans on antimicrobial resistance: Strengths and weaknesses. Antimicrob. Resist. Infect. Control 2022, 11, 90. [Google Scholar] [CrossRef]
- Zinner, S.H. The search for new antimicrobials: Why we need new options. Expert Rev. Anti Infect. Ther. 2005, 3, 907–913. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, D.W.; Gushulak, B.D.; Baine, W.B.; Bala, S.; Gubbins, P.O.; Holtom, P.; Segarra-Newnham, M. Population mobility, globalization, and antimicrobial drug resistance. Emerg. Infect. Dis. 2009, 15, 1727–1732. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Interagency Coordination Group on Antimicrobial Resistance. No time to wait: Securing the future from drug-resistant infections. In Report to the Secretary General of the United Nations; World Health Organisation: Geneva, Switzerland, 2019. [Google Scholar]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, A.Y.; Hooper, D.C. Hospital-acquired infections due to gram-negative bacteria. N. Engl. J. Med. 2010, 362, 1804–1813. [Google Scholar] [CrossRef]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and economic impact of antibiotic resistance in developing countries: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- Navidinia, M. The clinical importance of emerging ESKAPE pathogens in nosocomial infections. Arch. Adv. Biosci. 2016, 7, 43–57. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Link, E.M.; Hardiman, G.; Sluder, A.E.; Johnson, C.D.; Liu, L.X. Therapeutic target discovery using Caenorhabditis elegans. Pharmacogenomics 2000, 1, 203–217. [Google Scholar] [CrossRef]
- Jones, A.K.; Buckingham, S.D.; Sattelle, D.B. Chemistry-to-gene screens in Caenorhabditis elegans. Nat. Rev. Drug Discov. 2005, 4, 321–330. [Google Scholar] [CrossRef]
- Moy, T.I.; Ball, A.R.; Anklesaria, Z.; Casadei, G.; Lewis, K.; Ausubel, F.M. Identification of novel antimicrobials using a live-animal infection model. Proc. Natl. Acad. Sci. USA 2006, 103, 10414–10419. [Google Scholar] [CrossRef]
- Moy, T.I.; Conery, A.L.; Larkins-Ford, J.; Wu, G.; Mazitschek, R.; Casadei, G.; Lewis, K.; Carpenter, A.E.; Ausubel, F.M. High-throughput screen for novel antimicrobials using a whole animal infection model. ACS Chem. Biol. 2009, 4, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Gigante, V.; Sati, H.; Beyer, P. Recent advances and challenges in antibacterial drug development. ADMET DMPK 2022, 10, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.; Hopkins, A. Navigating chemical space for biology and medicine. Nature 2004, 432, 855–861. [Google Scholar] [CrossRef]
- Projan, S.J.; Shlaes, D.M. Antibacterial drug discovery: Is it all downhill from here? Clin. Microbiol. Infect. 2004, 10 (Suppl. S4), 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria. Drugs 2004, 64, 159–204. [Google Scholar] [CrossRef] [PubMed]
- Horrobin, D.F. Modern biomedical research: An internally self-consistent universe with little contact with medical reality? Nat. Rev. Drug Discov. 2003, 2, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Williams, M. A return to the fundamentals of drug discovery? Curr. Opin. Investig. Drugs 2004, 5, 29–33. [Google Scholar] [PubMed]
- Silver, L.L. Challenges of antibacterial discovery. Clin. Microbiol. Rev. 2011, 24, 71–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adonizio, A.; Leal, S.M.; Ausubel, F.M.; Mathee, K. Attenuation of Pseudomonas aeruginosa virulence by medicinal plants in a Caenorhabditis elegans model system. J. Med. Microbiol. 2008, 57, 809–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, A.R.; Kwok, T.C.; Howard, A.; Houston, E.; Johanson, K.; Chan, A.; Cutler, S.R.; McCourt, P.; Roy, P.J. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat. Protoc. 2006, 1, 1906–1914. [Google Scholar] [CrossRef]
- Tampakakis, E.; Okoli, I.; Mylonakis, E. A C. elegans-based, whole animal, in vivo screen for the identification of antifungal compounds. Nat. Protoc. 2008, 3, 1925–1931. [Google Scholar] [CrossRef] [PubMed]
- Gosai, S.J.; Kwak, J.H.; Luke, C.J.; Long, O.S.; King, D.E.; Kovatch, K.J.; Johnston, P.A.; Shun, T.Y.; Lazo, J.S.; Perlmutter, D.H.; et al. Automated high-content live animal drug screening using C. elegans expressing the aggregation prone serpin alpha1-antitrypsin Z. PLoS ONE 2010, 5, e15460. [Google Scholar] [CrossRef] [Green Version]
- Giunti, S.; Andersen, N.; Rayes, D.; De Rosa, M.J. Drug discovery: Insights from the invertebrate Caenorhabditis elegans. Pharmacol. Res. Perspect. 2021, 9, e00721. [Google Scholar] [CrossRef] [PubMed]
- Peterson, N.D.; Pukkila-Worley, R. Caenorhabditis elegans in high-throughput screens for anti-infective compounds. Curr. Opin. Immunol. 2018, 54, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Phillips, P.C. Self-fertilization sweeps up variation in the worm genome. Nat. Genet. 2012, 44, 237–238. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabricant, D.S.; Farnsworth, N.R. The value of plants used in traditional medicine for drug discovery. Environ. Health Perspect. 2001, 109 (Suppl. S1), 69–75. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Garibay, F.; Téllez-Valdez, O.; Moreno-Torres, G.; Calderón, J.S. Flavonoids from Tephrosia major. A New Prenyl-β-hydroxychalcone. Z. Für Naturforschung C 2002, 57, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Bennett, R.N.; Wallsgrove, R.M. Secondary metabolites in plant defence mechanisms. N. Phytol. 1994, 127, 617–633. [Google Scholar] [CrossRef]
- Savoia, D. Plant-derived antimicrobial compounds: Alternatives to antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadi, A.; Nazari, H.; Imani, S.; Amrollahi, H. Antifungal activities and chemical composition of some medicinal plants. J. Mycol. Med. 2014, 24, e1–e8. [Google Scholar] [CrossRef] [PubMed]
- Cappiello, F.; Loffredo, M.R.; Del Plato, C.; Cammarone, S.; Casciaro, B.; Quaglio, D.; Mangoni, M.L.; Botta, B.; Ghirga, F. The Revaluation of Plant-Derived Terpenes to Fight Antibiotic-Resistant Infections. Antibiotics 2020, 9, 325. [Google Scholar] [CrossRef]
- Shin, J.; Prabhakaran, V.S.; Kim, K.S. The multi-faceted potential of plant-derived metabolites as antimicrobial agents against multidrug-resistant pathogens. Microb Pathog. 2018, 116, 209–214. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Lewis, K.; Ausubel, F.M. Prospects for plant-derived antibacterials. Nat. Biotechnol. 2006, 24, 1504–1507. [Google Scholar] [CrossRef]
- Ruddaraju, L.K.; Pammi, S.V.N.; Guntuku, G.S.; Padavala, V.S.; Kolapalli, V.R.M. A review on anti-bacterials to combat resistance: From ancient era of plants and metals to present and future perspectives of green nano technological combinations. Asian J. Pharm. Sci. 2020, 15, 42–59. [Google Scholar] [CrossRef]
- Rodriguez-Yoldi, M.J. Anti-Inflammatory and Antioxidant Properties of Plant Extracts. Antioxidants 2021, 10, 921. [Google Scholar] [CrossRef]
- Cragg, G.; Newman, D. Natural products and drug discovery and development: A history of success and continuing promise for the future. Planta Med. 2014, 80, IL1. [Google Scholar] [CrossRef]
- Kong, C.; Yehye, W.A.; Abd Rahman, N.; Tan, M.W.; Nathan, S. Discovery of potential anti-infectives against Staphylococcus aureus using a Caenorhabditis elegans infection model. BMC Complement Altern. Med. 2014, 14, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Abd-Rashid, B.A.; Ismail, Z.; Ismail, R.; Mak, J.W.; Pook, P.C.; Er, H.M.; Ong, C.E. In vitro effects of active constituents and extracts of Orthosiphon stamineus on the activities of three major human cDNA-expressed cytochrome P450 enzymes. Chem. Biol. Interact. 2011, 190, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yam, M.F.; Mohamed, E.A.; Ang, L.F.; Pei, L.; Darwis, Y.; Mahmud, R.; Asmawi, M.Z.; Basir, R.; Ahmad, M. A simple isocratic HPLC method for the simultaneous determination of sinensetin, eupatorin, and 3’-hydroxy-5,6,7,4’-tetramethoxyflavone in Orthosiphon stamineus extracts. J. Acupunct. Meridian Stud. 2012, 5, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Alexpandi, R.; Prasanth, M.I.; Ravi, A.V.; Balamurugan, K.; Durgadevi, R.; Srinivasan, R.; De Mesquita, J.F.; Pandian, S.K. Protective effect of neglected plant Diplocyclos palmatus on quorum sensing mediated infection of Serratia marcescens and UV-A induced photoaging in model Caenorhabditis elegans. J. Photochem. Photobiol. B 2019, 201, 111637. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, J.W.; Kang, S.G.; Oh, S.; Griffiths, M.W. Bifidobacterium spp. influences the production of autoinducer-2 and biofilm formation by Escherichia coli O157:H7. Anaerobe 2012, 18, 539–545. [Google Scholar] [CrossRef]
- Patel, P.; Joshi, C.; Kothari, V. Anti-Pathogenic Efficacy and Molecular Targets of a Polyherbal Wound- Care Formulation (Herboheal) against Staphylococcus aureus. Infect. Disord. Drug Targets 2019, 19, 193–206. [Google Scholar] [CrossRef]
- Lee, K.M.; Lim, J.; Nam, S.; Yoon, M.Y.; Kwon, Y.K.; Jung, B.Y.; Park, Y.; Park, S.; Yoon, S.S. Inhibitory effects of broccoli extract on Escherichia coli O157:H7 quorum sensing and in vivo virulence. FEMS Microbiol. Lett. 2011, 321, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.J.; Kim, H.I.; Kim, J.A.; Jun, S.Y.; Kang, S.H.; Park, D.J.; Son, S.J.; Kim, Y.; Shin, O.S. The herbal-derived honokiol and magnolol enhances immune response to infection with methicillin-sensitive Staphylococcus aureus (MSSA) and methicillin-resistant S. aureus (MRSA). Appl. Microbiol. Biotechnol. 2015, 99, 4387–4396. [Google Scholar] [CrossRef]
- Zhou, Y.M.; Shao, L.; Li, J.A.; Han, L.Z.; Cai, W.J.; Zhu, C.B.; Chen, D.J. An efficient and novel screening model for assessing the bioactivity of extracts against multidrug-resistant Pseudomonas aeruginosa using Caenorhabditis elegans. Biosci. Biotechnol. Biochem. 2011, 75, 1746–1751. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Mi, D.Y.; Wang, J.; Luo, Y.P.; Yang, X.; Dong, S.; Ma, X.M.; Dong, K.Z. Constituent and effects of polysaccharides isolated from Sophora moorcroftiana seeds on lifespan, reproduction, stress resistance, and antimicrobial capacity in Caenorhabditis elegans. Chin. J. Nat. Med. 2018, 16, 252–260. [Google Scholar] [CrossRef]
- Dharmalingam, K.; Tan, B.K.; Mahmud, M.Z.; Sedek, S.A.; Majid, M.I.; Kuah, M.K.; Sulaiman, S.F.; Ooi, K.L.; Khan, N.A.; Muhammad, T.S.; et al. Swietenia macrophylla extract promotes the ability of Caenorhabditis elegans to survive Pseudomonas aeruginosa infection. J. Ethnopharmacol. 2012, 139, 657–663. [Google Scholar] [CrossRef]
- Evans, E.A.; Kawli, T.; Tan, M.W. Pseudomonas aeruginosa suppresses host immunity by activating the DAF-2 insulin-like signaling pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175. [Google Scholar] [CrossRef] [Green Version]
- Haripriyan, J.; Omanakuttan, A.; Menon, N.D.; Vanuopadath, M.; Nair, S.S.; Corriden, R.; Nair, B.G.; Nizet, V.; Kumar, G.B. Clove Bud Oil Modulates Pathogenicity Phenotypes of the Opportunistic Human Pathogen Pseudomonas aeruginosa. Sci. Rep. 2018, 8, 3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, F.M.; Ahmad, I.; Asif, M.; Tahseen, Q. Influence of clove oil on certain quorum-sensing-regulated functions and biofilm of Pseudomonas aeruginosa and Aeromonas hydrophila. J. Biosci. 2013, 38, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, P.S.; Rai, R.V. Inhibition of quorum-sensing-controlled virulence factors of Pseudomonas aeruginosa by Murraya koenigii essential oil: A study in a Caenorhabditis elegans infectious model. J. Med. Microbiol. 2016, 65, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hafting, J.; Critchley, A.T.; Banskota, A.H.; Prithiviraj, B. Components of the cultivated red seaweed Chondrus crispus enhance the immune response of Caenorhabditis elegans to Pseudomonas aeruginosa through the pmk-1, daf-2/daf-16, and skn-1 pathways. Appl. Environ. Microbiol. 2013, 79, 7343–7350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco(R): A product of Ascophyllum nodosum enhances immune response of Caenorhabditis elegans against Pseudomonas aeruginosa infection. Mar. Drugs 2012, 10, 84–105. [Google Scholar] [CrossRef] [Green Version]
- Patel, H.; Patel, F.; Jani, V.; Jha, N.; Ansari, A.; Paliwal, B.; Rathod, B.; Patel, D.; Patel, P.; Kothari, V. Anti-pathogenic potential of a classical ayurvedic Triphala formulation. F1000Res 2019, 8, 1126. [Google Scholar] [CrossRef] [Green Version]
- Husain, F.M.; Ahmad, I.; Khan, F.I.; Al-Shabib, N.A.; Baig, M.H.; Hussain, A.; Rehman, M.T.; Alajmi, M.F.; Lobb, K.A. Seed Extract of Psoralea corylifolia and Its Constituent Bakuchiol Impairs AHL-Based Quorum Sensing and Biofilm Formation in Food- and Human-Related Pathogens. Front. Cell. Infect. Microbiol. 2018, 8, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, F.M.; Ahmad, I.; Al-Thubiani, A.S.; Abulreesh, H.H.; AlHazza, I.M.; Aqil, F. Leaf Extracts of Mangifera indica L. Inhibit Quorum Sensing—Regulated Production of Virulence Factors and Biofilm in Test Bacteria. Front. Microbiol. 2017, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Nandu, T.G.; Subramenium, G.A.; Shiburaj, S.; Viszwapriya, D.; Iyer, P.M.; Balamurugan, K.; Rameshkumar, K.B.; Karutha Pandian, S. Fukugiside, a biflavonoid from Garcinia travancorica inhibits biofilm formation of Streptococcus pyogenes and its associated virulence factors. J. Med. Microbiol. 2018, 67, 1391–1401. [Google Scholar] [CrossRef]
- Alam, S.T.; Hwang, H.; Son, J.D.; Nguyen, U.T.T.; Park, J.S.; Kwon, H.C.; Kwon, J.; Kang, K. Natural photosensitizers from Tripterygium wilfordii and their antimicrobial photodynamic therapeutic effects in a Caenorhabditis elegans model. J. Photochem. Photobiol. B 2021, 218, 112184. [Google Scholar] [CrossRef]
- Kim, Y.G.; Lee, J.H.; Gwon, G.; Kim, S.I.; Park, J.G.; Lee, J. Essential Oils and Eugenols Inhibit Biofilm Formation and the Virulence of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 36377. [Google Scholar] [CrossRef] [Green Version]
- Ibáñez-Peinado, D.; Pina-Pérez, C.; García-Carrión, G.; Martínez, A.; Rodrigo, D. In vivo Antimicrobial Activity Assessment of a Cauliflower By-Product Extract Against Salmonella Typhimurium. Front. Sustain. Food Syst. 2020, 4, 8. [Google Scholar] [CrossRef]
- Sanz-Puig, M.; Lazaro, E.; Armero, C.; Alvares, D.; Martinez, A.; Rodrigo, D.S. Typhimurium virulence changes caused by exposure to different non-thermal preservation treatments using C. elegans. Int. J. Food Microbiol. 2017, 262, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobo-Salcedo Mdel, R.; Gonzalez-Espindola, L.A.; Alonso-Castro, A.J.; Gonzalez-Martinez Mdel, R.; Dominguez, F.; Garcia-Carranca, A. Antimicrobial activity and cytotoxic effects of Magnolia dealbata and its active compounds. Nat. Prod. Commun. 2011, 6, 1121–1124. [Google Scholar] [CrossRef] [Green Version]
- Eng, S.A.; Nathan, S. Curcumin rescues Caenorhabditis elegans from a Burkholderia pseudomallei infection. Front. Microbiol. 2015, 6, 290. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chen, Z.; Ching, P.; Shi, Q.; Li, X. An integrated microfluidic platform for evaluating in vivo antimicrobial activity of natural compounds using a whole-animal infection model. Lab. Chip. 2013, 13, 3373–3382. [Google Scholar] [CrossRef]
- Kim, H.-I.; Kim, J.-A.; Choi, E.-J.; Harris, J.B.; Jeong, S.-Y.; Son, S.-J.; Kim, Y.; Shin, O.S. In vitro and in vivo antimicrobial efficacy of natural plant-derived compounds against Vibrio cholerae of O1 El Tor Inaba serotype. Biosci. Biotechnol. Biochem. 2015, 79, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Lakhundi, S.; Zhang, K. Methicillin-Resistant Staphylococcus aureus: Molecular Characterization, Evolution, and Epidemiology. Clin. Microbiol. Rev. 2018, 31, 10–1128. [Google Scholar] [CrossRef] [Green Version]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Bag, A.; Bhattacharyya, S.K.; Pal, N.K. Antibacterial potential of hydroalcoholic extracts of triphala components against multidrug-resistant uropathogenic bacteria—A preliminary report. Indian J. Exp. Biol. 2013, 51, 709–714. [Google Scholar]
- Khushboo, P.S.; Jadhav, V.M.; Kadam, V.J.; Sathe, N.S. Psoralea corylifolia Linn.-“Kushtanashini”. Pharmacogn. Rev. 2010, 4, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, S.; Vaitla, P. Streptococcus Pyogenes; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Viszwapriya, D.; Subramenium, G.A.; Prithika, U.; Balamurugan, K.; Pandian, S.K. Betulin inhibits virulence and biofilm of Streptococcus pyogenes by suppressing ropB core regulon, sagA and dltA. Pathog. Dis. 2016, 74, ftw088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef]
- Singulani, J.L.; Scorzoni, L.; Gomes, P.C.; Nazare, A.C.; Polaquini, C.R.; Regasini, L.O.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Activity of gallic acid and its ester derivatives in Caenorhabditis elegans and zebrafish (Danio rerio) models. Future Med. Chem. 2017, 9, 1863–1872. [Google Scholar] [CrossRef]
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 2007, 3, e18. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Liao, K.; Wang, D. Effects of Magnolol and Honokiol on Adhesion, Yeast-Hyphal Transition, and Formation of Biofilm by Candida albicans. PLoS ONE 2015, 10, e0117695. [Google Scholar] [CrossRef] [Green Version]
- Coleman, J.J.; Okoli, I.; Tegos, G.P.; Holson, E.B.; Wagner, F.F.; Hamblin, M.R.; Mylonakis, E. Characterization of plant-derived saponin natural products against Candida albicans. ACS Chem. Biol. 2010, 5, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.X.; Li, D.D.; Hu, D.D.; Hu, G.H.; Yan, L.; Wang, Y.; Jiang, Y.Y. Effect of Tetrandrine against Candida albicans Biofilms. PLoS ONE. 2013, 8, e79671. [Google Scholar] [CrossRef] [Green Version]
- Costa-de-Oliveira, S.; Rodrigues, A.G. Candida albicans Antifungal Resistance and Tolerance in Bloodstream Infections: The Triad Yeast-Host-Antifungal. Microorganisms 2020, 8, 154. [Google Scholar] [CrossRef] [Green Version]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas, E.C.; Correa, L.B.; das Graças Henriques, M. Chapter 28—Antiinflammatory Properties of Schinus terebinthifolius and Its Use in Arthritic Conditions. In Bioactive Food as Dietary Interventions for Arthritis and Related Inflammatory Diseases, 2nd ed.; Watson, R.R., Preedy, V.R., Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 489–505. [Google Scholar]
- De Paula, E.S.A.C.; Costa-Orlandi, C.B.; Gullo, F.P.; Sangalli-Leite, F.; de Oliveira, H.C.; da Silva Jde, F.; Scorzoni, L.; Pitangui Nde, S.; Rossi, S.A.; Benaducci, T.; et al. Antifungal Activity of Decyl Gallate against Several Species of Pathogenic Fungi. Evid. Based Complement. Altern. Med. 2014, 2014, 506273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, C.; Sun, L.; Zhang, W. Thymol has antifungal activity against Candida albicans during infection and maintains the innate immune response required for function of the p38 MAPK signaling pathway in Caenorhabditis elegans. Immunol. Res. 2016, 64, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Murtaza, G.; Karim, S.; Akram, M.R.; Khan, S.A.; Azhar, S.; Mumtaz, A.; Bin Asad, M.H. Caffeic acid phenethyl ester and therapeutic potentials. Biomed. Res. Int. 2014, 2014, 145342. [Google Scholar] [CrossRef] [Green Version]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Ahmad Khan, M.S.; Altaf, M.M.; Sajid, M. Chapter 14—Insights of Phyto-Compounds as Antipathogenic Agents: Controlling Strategies for Inhibiting Biofilms and Quorum Sensing in Candida albicans. In New Look to Phytomedicine; Ahmad Khan, M.S., Ahmad, I., Chattopadhyay, D., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 367–389. [Google Scholar]
- Benson, J.M.; Nahata, M.C. Clinical use of systemic antifungal agents. Clin. Pharm. 1988, 7, 424–438. [Google Scholar] [PubMed]


{kind=link}
| Active Compounds | Plant | Microorganism Tested | In Vitro | In Vivo (C. elegans Infection Model) | MoA * | References |
|---|---|---|---|---|---|---|
| (MIC) | (EC50) | |||||
| Eupatorin, sinensetin, 3′-hydroxy-5, 6, 7,4′-tetramethox-yflavone, and rosmarinic acid | Nypa fruticans Swietenia macrophylla Curcuma longa Eurycoma longifolia Orthosiphon stamineus Silybum eburneum | S. aureus NCTC8325-4 and MRSA ATCC33591 | <100 μg/mL | Reduced bacterial colonisation of the nematode intestines by S. aureus NCTC8325-4 and increased survival by 2.8-fold. Protected worms from MRSA ATCC33591 infection | Immune genes activation and inhibition of bacterial virulence | [46,47,48] |
| No data | ||||||
| <125 μg/mL | ||||||
| N.D. ** | ||||||
| N.D. ** | ||||||
| 100 μg/mL | ||||||
| N.D. ** | Diplocyclos palmatus | S. marcescens PS1 | Inhibition of prodigiosin pigment production in S. marcescens at 2 mg/mL by 85% | Increased survival time by 50% Reduced colonisation of S. marcescens | Inhibition of QS-regulated biofilm formation and virulence factor production in S. marcescens. Upregulation of daf-16 and immune-related genes (clec-87, lys-7, bec-1, and clec-60) | [49] |
| N.D. ** | Bifidobacterium spp. | Enterohemorrhagic Escherichia coli (EHEC) O157:H7 | 98-fold reduction in AI-2 activity in EHEC O157:H7 | Increased survival rate against EHEC O157:H7 | Inhibition of virulence factor production in EHEC O157:H7 | [50] |
| N.D. ** | Azadirachta indica | S. aureus | 0.025–0.5% v/v | Inhibition of bacterial growth and production of staphyloxanthin, a quorum sensing (QS)-regulated pigment, at concentrations ≥0.025% v/v | QS-mediated bacterial virulence and transcription of the genes encoding virulence factors | [51] |
| Acacia nilotica | ||||||
| Ocimum sanctum | ||||||
| Annona squamosa | ||||||
| Curcuma longa | ||||||
| Ricinus communis | ||||||
| N.D. ** | Broccoli extract | Escherichia coli O157:H7 | N.D. ** | Increased survival rate of worms by 28.5% | Inhibition of QS-mediated bacterial virulence genes | [52] |
| Honokiol and magnolol | Magnolia sp. | Methicillin-resistant Staphylococcus aureus (MRSA) | N.D. ** | Protected worms from MRSA infection and increased survival rate | Increased immune-modulatory effects via IFN-β and IFN-λ | [53] |
| N.D. ** | Angelica sinensis | P. aeruginosa | In vitro inhibition of MDRP. aeruginosa | Prolonged C. elegans survival against MDR P. aeruginosa | N.D. ** | [54] |
| Bupleurum chinense | ||||||
| Herba plantaginis | ||||||
| Herba menthae | ||||||
| Stephania japonica | ||||||
| Phoma exigua | ||||||
| Aspergillus sydowii | ||||||
| Herba plantaginis | ||||||
| Plantago depressa | ||||||
| N.D. ** | Sophora moorcroftiana | P. aeruginosa | N.D. ** | Improved C. elegans lifespan and reproductive capacity | N.D. ** | [55] |
| Limonoids | Swietenia macrophylla | P. aeruginosa PA14 | No inhibition | Prolonged C. elegans lifespan by reducing virulence factors and enhancing immune response | Activation of immune modulators gene lys-7, which suppresses QS-mediated mechanisms | [56,57] |
| N.D. ** | Clove bud oil (CBO) | P. aeruginosa | CBO inhibited elastases A and B and regulated P. aeruginosa signalling molecules at concentrations (<3.2%) | Treatment of P. aeruginosa-infected C. elegans with <2% v/v clove oil improved worm survival in vivo | Immune genes activation, QS regulation | [58,59] |
| N.D. ** | Murraya koenigii | P. aeruginosa | N.D. ** | Improved the survival of infected worms | Immune genes activation | [60] |
| N.D. ** | Chondrus crispus | P. aeruginosa | N.D. ** | Improved survival of P. aeruginosa-infected C. elegans at a 500 µg/mL concentration | Immune genes activation (the pmk-1, daf-2/daf-16, and skn-1) | [61] |
| N.D. ** | Ascophyllum nodosum | P. aeruginosa | N.D. ** | Promoted the survival of C. elegans by significantly reducing biofilm formation by P. aeruginosa | Immune genes activation | [62] |
| Triphala (TF) | Phyllanthus emblica | S. aureus, P. aeruginosa, S. marcescens, and C. violaceum | N.D. ** | Reduced virulence (18–45%) towards the nematode host and increase survival of C. elegans by 14–41% | N.D. ** | [63] |
| Terminalia bellerica | ||||||
| Terminalia chebula | ||||||
| N.D. ** | Psoralea corylifolia | P. aeruginosa, | Inhibited in vitro biofilm formation | 1000 µg/mL P. coryliflia seed extract increased infected worms’ survival | Inhibition of QS-mediated virulence and biofilm (LasR and RhlR genes) | [64] |
| A. hydrophila, | ||||||
| C. violaceum, | ||||||
| S. marcescens, and | ||||||
| L. monocytogenes | ||||||
| N.D. ** | Mangifera indica | Chromobacterium violaceum (12472) Pseudamonas aeruginosa PAO1 Aeromonas hydrophila WAF38 | Chromobacterium violaceum (12472) | P. aeruginosa-infected worms treated with Mangifera indica survived longer (72%) compared to untreated nematodes | N.D. ** | [65] |
| = 1000 μg/mL, | ||||||
| P. aeruginosa PAO1 and A. hydrophila WAF38 = 2000 μg/mL | ||||||
| Fukugiside | Garcinia travancorica | Streptococcus pyogenes SF370 | 60, 80, 100, and 120 µg/mL shows 70–90% biofilm inhibition | 80 µg/mL concentration increased survival of C. elegans infected with Streptococcus pyogenes | Inhibition of virulence genes and phagocytic enhancement | [66] |
| N.D. ** | Tripterygium wilfordii | S. pyogenes S. aureus | 70–96% biofilm inhibition at 80 μg/mL | Improved the lifespan of C. elegans infected with S. pyogenes | Photosensitisation | [67] |
| Photosensitise S. pyogenes and S. aureus for the therapy of skin infections | ||||||
| Eugenol | Bay and clove essential oils | Enterohemorrhagic Escherichia coli O157:H7 (EHEC) | 0.0005% inhibited 87% of EHEC biofilm. A 100% inhibition was reported at concentration of 0.005% | 0.005% concentration prolonged C. elegans survival in the presence of EHEC | Biofilm inhibition | [68] |
| N.D. ** | B. oleracea var Botrytis (cauliflower) | Salmonella typhimurium CECT 443 | 3% cauliflower | 3% cauliflower prolonged C. elegans infected with Salmonella typhimurium and | Reduced virulence factors | [69,70] |
| dose-dependently reduced Salmonella typhimurium virulence factors | ||||||
| Honokiol and Magnolol | Magnolia sp. | Vibrio cholerae | N.D. ** | Worm survival significantly prolonged and decreased host inflammatory responses | N.D.** | [71] |
| Curcumin | Curcuma longa | Burkholderia pseudomallei | N.D. ** | Reduced pathogenicity and attenuation of virulence factors in Burkholderia pseudomallei-infected worms | Inhibition of transcription of the genes encoding virulence factors | [72] |
| Aloe-emodin rhein and emodin | Rhubarb | S. aureus ATCC 25923 | Aloe-emodin = 7.5 mg/mL | All compounds rescued C. elegans infected with S. aureus 25,923 at concentration of <60 µg/mL, as detected using microfluidic device survival assay | N.D.** | [73] |
| Rhein = 16.0 mg/mL | ||||||
| Emodin = 6.3 mg/mL | ||||||
| Magnolol | Scutellaria baicalensis, Scutellaria lateriflora, Magnolia officinalis | V. cholerae O1 E1 Tor serotype Inaba strain N16961 | Baicalein = 1.25 µg/mL | 1–10 µg/mL Honokiol and Magnolol prolonged survival of C. elegans infected with V. cholerae O1 | Biofilm inhibition, anti-inflammatory effects | [74] |
| Honokiol | Honokiol = 0.625 µg/mL | |||||
| Quercetin | Magnolol = 5 µg/mL | |||||
| Resveratrol | Quercetin = 0.625 µg/mL | |||||
| Resveratrol = 0.625 µg/mL |
| Active Compounds | Plant | Microorganism Tested | In Vitro | In Vivo (C. elegans Infection Model) | MoA * | References |
|---|---|---|---|---|---|---|
| MIC50 | EC50 | |||||
| Gallic acid and gallates (secondary metabolites) | Derived from various plants | Candida spp. | N.D. ** | Increase nematodes’ lifespans at >1 µg/mL concentrations | N.D. ** | [84] |
| Cryptococcus spp. | ||||||
| Paracoccidioides spp. | ||||||
| Histoplasma capsulatum | ||||||
| Enoxacin | Tabebuia avellaneda | C. albicans | Inhibit C. albicans biofilm formation in vitro | Prolong the survival of C. albicans-infected worms and prevent fungal–hyphal transition | N.D. ** | [85] |
| Caffeic acid phenethyl ester (CAPE) | ||||||
| Lapachol | ||||||
| Magnolol and honokiol | Magnolia officinalis | C. albicans | Significantly suppress in vitro yeast cell adhesion, hyphal growth, and biofilm formation at 4–32 μg/mL concentrations | Prolongs the lifespan of C. albicans-infected worms | Downregulation of virulence genes (Ras1-cAMP-Efg1 pathway) | [86] |
| A2 | Plant-derived natural products compound collection | C. albicans | 27.5 µg/mL | 55.1 µg/mL | Photosensitisation | [87] |
| A8 | 5.8 µg/mL | 23.1 µg/mL | ||||
| A17 | 31.0 µg/mL | 31.0 µg/mL | ||||
| A19 | 26.5.0 µg/mL | 26.5 µg/mL | ||||
| A21 | 16.5 µg/mL | 16.5 µg/mL | ||||
| A24 | 13.3 µg/mL | 13.3 µg/mL | ||||
| A25 (saponins) | 28.7 µg/mL | 28.7 µg/mL | ||||
| All compounds increase C. albicans-infected worms’ lifespans | ||||||
| Tetrandrine | Stephania tetrandra | C. albicans | MIC = 16, 32, 64 mg/L | EC50 = 4, 8 16, 32 mg/L protects worms against C. albicans infections | QS and biofilm inhibition (Ras/cAMP pathway) | [88] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarroug, S.H.O.; Bajaman, J.S.; Hamza, F.N.; Saleem, R.A.; Abdalla, H.K. Caenorhabditis elegans as an In Vivo Model for the Discovery and Development of Natural Plant-Based Antimicrobial Compounds. Pharmaceuticals 2023, 16, 1070. https://doi.org/10.3390/ph16081070
Zarroug SHO, Bajaman JS, Hamza FN, Saleem RA, Abdalla HK. Caenorhabditis elegans as an In Vivo Model for the Discovery and Development of Natural Plant-Based Antimicrobial Compounds. Pharmaceuticals. 2023; 16(8):1070. https://doi.org/10.3390/ph16081070
Chicago/Turabian StyleZarroug, Samah H. O., Juhaina S. Bajaman, Fatheia N. Hamza, Rimah A. Saleem, and Hana K. Abdalla. 2023. "Caenorhabditis elegans as an In Vivo Model for the Discovery and Development of Natural Plant-Based Antimicrobial Compounds" Pharmaceuticals 16, no. 8: 1070. https://doi.org/10.3390/ph16081070